
Rhizosphere competent Pantoea agglomerans enhances maize ( Zea mays ) and chickpea ( Cicer arietinum...
9
ORIGINAL PAPER Rhizosphere competent Pantoea agglomerans enhances maize (Zea mays) and chickpea (Cicer arietinum L.) growth, without altering the rhizosphere functional diversity Aradhana Mishra • Puneet Singh Chauhan • Vasvi Chaudhry • Manisha Tripathi • Chandra Shekhar Nautiyal Received: 6 May 2011 / Accepted: 24 May 2011 / Published online: 3 June 2011 Ó Springer Science+Business Media B.V. 2011 Abstract Plant growth promoting Pantoea agglom- erans NBRISRM (NBRISRM) was able to produce 60.4 lg/ml indole acetic acid and solubilize 77.5 lg/ ml tri-calcium phosphate under in vitro conditions. Addition of 2% NaCl (w/v) in the media induced the IAA production and phosphate solubilization by 11% and 7%, respectively. For evaluating the plant growth promotory effect of NBRISRM inoculation a micro plot trial was conducted using maize and chickpea as host plants. The results revealed significant increase in all growth parameters tested in NBRISRM inoc- ulated maize and chickpea plants, which were further confirmed by higher macronutrients (N, P and K) accumulation as compared to un-inoculated controls. Throughout the growing season of maize and chick- pea, rhizosphere population of NBRISRM were in the range 10 7 –10 8 CFU/g soil and competing with 10 7 – 10 9 CFU/g soil with heterogeneous bacterial popula- tion. Functional richness, diversity, and evenness were found significantly higher in maize rhizosphere as compared to chickpea, whereas NBRISRM inoc- ulation were not able to change it, in both crops as compared to their un-inoculated control. To the best of our knowledge this is first report where we demonstrated the effect of P. agglomerans strain for improving maize and chickpea growth without alter- ing the functional diversity. Keywords Chickpea (Cicer arietinum L.) Á Maize (Zea mays) Á Pantoea agglomerans Á Functional diversity Á Nutrient uptake Introduction The use of beneficial soil plant growth promoting rhizobacteria (PGPR) for improving crop production requires the selection of rhizosphere-competent bac- terial strains with multiple plant growth promoting attributes (Nautiyal et al. 2008; Hynes et al. 2008). The group of these beneficial PGPR belongs to various genera e.g. Acetobacter, Acinetobacter, Alca- ligenes, Arthrobacter, Azospirillum, Azotobacter, Bacillus, Burkholderia, Enterobacter, Gluconacetob- acter, Herbaspirillum, Klebsiella, Methylobacterium, Ochrobactrum, Pantoea, Pseudomonas, Rhodococ- cus, Serratia, and Stenotrophomonas (Babalola 2010). Among the above mentioned beneficial PGPR, Pantoea agglomerans is a member of Enterobacteri- aceae found everywhere in nature and inhabiting plants, soil, water, animals and humans (Wright et al. 2001; Chauhan and Nautiyal 2010). Several isolates of P. agglomerans are known to improve the plant growth promotion by various mechanisms which A. Mishra Á P. S. Chauhan Á V. Chaudhry Á M. Tripathi Á C. S. Nautiyal (&) Division of Plant Microbe Interactions, National Botanical Research Institute, Rana Pratap Marg, Lucknow 226001, India e-mail: [email protected] 123 Antonie van Leeuwenhoek (2011) 100:405–413 DOI 10.1007/s10482-011-9596-8
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Rhizosphere competent Pantoea agglomerans enhances maize ( Zea mays ) and chickpea ( Cicer arietinum...

ORIGINAL PAPER
Rhizosphere competent Pantoea agglomerans enhancesmaize (Zea mays) and chickpea (Cicer arietinum L.) growth,without altering the rhizosphere functional diversity
Aradhana Mishra • Puneet Singh Chauhan •
Vasvi Chaudhry • Manisha Tripathi •
Chandra Shekhar Nautiyal
Received: 6 May 2011 / Accepted: 24 May 2011 / Published online: 3 June 2011
� Springer Science+Business Media B.V. 2011
Abstract Plant growth promoting Pantoea agglom-
erans NBRISRM (NBRISRM) was able to produce
60.4 lg/ml indole acetic acid and solubilize 77.5 lg/
ml tri-calcium phosphate under in vitro conditions.
Addition of 2% NaCl (w/v) in the media induced the
IAA production and phosphate solubilization by 11%
and 7%, respectively. For evaluating the plant growth
promotory effect of NBRISRM inoculation a micro
plot trial was conducted using maize and chickpea as
host plants. The results revealed significant increase
in all growth parameters tested in NBRISRM inoc-
ulated maize and chickpea plants, which were further
confirmed by higher macronutrients (N, P and K)
accumulation as compared to un-inoculated controls.
Throughout the growing season of maize and chick-
pea, rhizosphere population of NBRISRM were in the
range 107–108 CFU/g soil and competing with 107–
109 CFU/g soil with heterogeneous bacterial popula-
tion. Functional richness, diversity, and evenness
were found significantly higher in maize rhizosphere
as compared to chickpea, whereas NBRISRM inoc-
ulation were not able to change it, in both crops as
compared to their un-inoculated control. To the best
of our knowledge this is first report where we
demonstrated the effect of P. agglomerans strain for
improving maize and chickpea growth without alter-
ing the functional diversity.
Keywords Chickpea (Cicer arietinum L.) � Maize
(Zea mays) � Pantoea agglomerans � Functional
diversity � Nutrient uptake
Introduction
The use of beneficial soil plant growth promoting
rhizobacteria (PGPR) for improving crop production
requires the selection of rhizosphere-competent bac-
terial strains with multiple plant growth promoting
attributes (Nautiyal et al. 2008; Hynes et al. 2008).
The group of these beneficial PGPR belongs to
various genera e.g. Acetobacter, Acinetobacter, Alca-
ligenes, Arthrobacter, Azospirillum, Azotobacter,
Bacillus, Burkholderia, Enterobacter, Gluconacetob-
acter, Herbaspirillum, Klebsiella, Methylobacterium,
Ochrobactrum, Pantoea, Pseudomonas, Rhodococ-
cus, Serratia, and Stenotrophomonas (Babalola
2010). Among the above mentioned beneficial PGPR,
Pantoea agglomerans is a member of Enterobacteri-
aceae found everywhere in nature and inhabiting
plants, soil, water, animals and humans (Wright et al.
2001; Chauhan and Nautiyal 2010). Several isolates
of P. agglomerans are known to improve the plant
growth promotion by various mechanisms which
A. Mishra � P. S. Chauhan � V. Chaudhry �M. Tripathi � C. S. Nautiyal (&)
Division of Plant Microbe Interactions, National
Botanical Research Institute, Rana Pratap Marg,
Lucknow 226001, India
e-mail: [email protected]
123
Antonie van Leeuwenhoek (2011) 100:405–413
DOI 10.1007/s10482-011-9596-8

mainly includes solubilization of inorganic phosphate
(Son et al. 2006), phytohormone production (Dastag-
er et al. 2009), nitrogen fixation (Loiret et al. 2004),
reduction of ethylene level by the enzyme 1-amino-
cyclopropane-1-carboxylate (ACC) deaminase (Long
et al. 2008) and biological control activity (Braun-
Kiewnick et al. 2000; Canamas et al. 2009) etc.
In many cases PGPR fail to induce the desired
effects when inoculated under the field conditions
because of poor colonization of the rhizosphere of
host plants, which is the most important requirement
for any PGPR for exhibiting all beneficial effects for
crop production (Lugtenberg et al. 2001; Nautiyal
et al. 2008). In fact, any exogenous strain has to
compete with a well-established rhizosphere micro-
flora in order to colonize in the rhizosphere of host
plant effectively. Different biotic and abiotic factors
influence plant development and bacterial diversity
associated with them (Andreote et al. 2010).
The bacterial diversity associated with different
plants in the same soil differ (Salles et al. 2004) and
inoculated efficient PGPR may survive, proliferate
and sometimes even alter the microbial diversity of
the host’s rhizosphere depending on their interactions
with the indigenous rhizosphere microflora (Andreote
et al. 2010; Nautiyal et al. 2010a). However, the
effects of bacterial inoculation on bacterial commu-
nity composition in different host plants are rarely
studied. Functional diversity based on sole carbon
source utilization pattern is a rapid, community level
approach for assessing the changes in the patterns of
host plant rhizosphere microbial diversity due to
inoculation of plant growth promoting microbes
(Nautiyal et al. 2010a, b, c).
In the present study, we aimed to evaluate the
effect of P. agglomerans NBRISRM inoculation in
promoting the chickpea (Cicer arietinum L.) and
maize (Zea mays) plant growth, macronutrient uptake
and followed by evaluating the changes in the
rhizosphere functional diversity of the two crops.
Materials and methods
Bacterial strain and growth conditions
Chickpea rhizosphere-competent Pantoea agglomer-
ans NBRISRM (NBRISRM) was isolated from the
roots of field-grown chickpea (C. arietinum L.) as
described earlier by Nautiyal (1997). Pure culture of
NBRISRM was grown and maintained on nutrient
broth (NB) or nutrient agar (NA) (HI-MEDIA Labo-
ratories, Bombay, India) and in 30% glycerol stocks
stored at -80�C. Identification of P. agglomerans
SRM was based on 95 carbon source utilization
pattern, using Biolog GN2 microtitre panels by fully
automated Biolog Bacteria Identification system (Bio-
log Inc., Hayward, CA, USA) as described earlier by
Nautiyal et al. (2007). Biolog microlog GN release
6.01 Database used for identification of the P. agglom-
erans SRM showed closest homology with P. agglom-
erans with 99% probability. Further identification of
SRM was based on 16S rDNA sequence analysis.
A PCR product of 1465 base pairs 16S rDNA was
sequenced, and data were analysed as described earlier
(Dastager et al. 2009). Sequence data of SRM have
been deposited in the GenBank, nucleotide sequence
database under the accession number GQ225111.
Sequence analysis of the isolate was compared with
16S rRNA sequences using BLAST search in the
NCBI, GenBank database (http://www.ncbi.nlm.
nih.gov). Multiple sequence alignments were per-
formed using CLUSTAL-X (Thompson et al. 1997).
The method of Jukes and Cantor (1969) was used to
calculate evolutionary distances. Phylogenetic den-
drogram was constructed by the neighbour-joining
method (Felsenstein 1985) and tree topologies were
evaluated by performing bootstrap analysis of 1,000
datasets using MEGA 4 (Molecular Evolutionary
Genetic Analysis).
IAA production and phosphate solubilization
by P. agglomerans NBRISRM
IAA production was detected by the modified method
as described by Brick et al. (1991). Bacterial cultures
were grown for 72 h in NB media containing trypto-
phan (100 lg/ml) at 30 ± 2�C. Fully grown cultures
were centrifuged at 3,000 rpm for 30 min. The super-
natant (1 ml) was mixed with 100 ll of orthophos-
phoric acid and 4 ml of the Salkowski reagent (50 ml,
35% of perchloric acid, 1 ml 0.5 M FeCl3 solution).
Development of pink color indicated IAA production.
Optical density was taken at 530 nm using Spectronic
20 D? spectrophotometer (Milton Roy Company,
USA). The concentration of IAA produced by cultures
was compared to standard curves of known IAA
(Hi-media) concentrations from the range of 10–100 lg/
406 Antonie van Leeuwenhoek (2011) 100:405–413
123

ml. National Botanical Research Institute’s phosphate
growth medium (NBRIP) which contained (l-1): glu-
cose, 10 g; Ca3(PO4)2, 5 g; MgCl2�6H2O, 5 g;
MgSO4�7H2O, 0.25 g; KCl, 0.2 g, and (NH4)2SO4,
0.1 g, was used to check the phosphate solubilization
by NBRISRM (Nautiyal 1999). The amount of soluble
phosphate was determined using the Fiske and Subbarow
method (Fiske and Subbarow 1925). Blank samples were
prepared by omitting 1-amino-2-naphthol-4-sulfonic
acid from the assay system as described earlier (Mehta
and Nautiyal 2001). The effects of temperature, salt and
pH were evaluated on the IAA production and phosphate
solubilization ability of NBRISRM in 150 ml Erlen-
meyer flasks containing 50 ml NB with tryptophan
(100 lg/ml) and NBRIP, respectively. The following
conditions were tested: for salt tolerance experiments 2
and 4, % NaCl (w/v), for temperature tolerance flasks
were incubated at 30 and 37�C and for pH tolerance
experiments, pH 9 of medium maintained by adding
NaOH in the media.
Plant growth promotion assay
Micro plot trial was carried out in the field of the
National Botanical Research Institute, Lucknow in a
randomized block design with four replicates using
host plant maize (Zea mays cv. Arkil) and chickpea
(Cicer arietinum cv. Radhey) as described earlier
(Nautiyal et al. 2010a, b). Bacterial inoculum of SRM
for maize and chickpea seeds was prepared by
suspending a 48 h grown culture from NA plates at
28�C in 10 ml of 0.85% saline Milli-Q water (MQW),
containing about 8 log10 CFU/ml. Surface-sterilized
maize and chickpea seeds were soaked in the bacterial
suspension for 4 h at 28�C on a reciprocal shaker at
100 rpm. Control seeds (non-bacterized) were soaked
in 0.85% saline MQW washed from uninoculated NA
plates (Nautiyal et al. 2010a). Each treatment was
raised in eight rows, each of 8 m length 6 m width,
with an intra- and inter-row spacing 10 and 60 cm,
respectively. Harvesting of maize and chickpea was
done at 90 and 105 days after sowing (DAS), respec-
tively and data was recorded as described earlier
(Nautiyal et al. 2010a, b).
Nutrient uptake
Twelve plants chosen at random harvest were rinsed
with Milli-Q water and oven-dried at 75�C for 72 h.
The dried shoot tissues were ground and then
digested using concentrated HNO3 (Page et al.
1982) for the determination of K using an atomic
absorption spectrometer. Total N and P were
extracted by digesting shoot tissue with 3 ml con-
centrated H2SO4 and 1 ml H2O2, respectively, at
360�C, and determined by the Berthelot reaction and
molybdenum blue method, respectively (Page et al.
1982). The amount of NaHCO3-extractable P (avail-
able inorganic P) from the dried shoot tissues was
determined by extracting samples with 0.5 M
NaHCO3 (pH 8.5) at a solution/solid ratio of 20:1
for 30 min (Olsen and Sommers 1982).
Tracking of P. agglomerans NBRISRM
in the rhizosphere of maize and chickpea
In order to monitor the presence of NBRISRM on
plant roots grown in non-sterilized soils, a spontane-
ous rifampicin-resistant (Rifr) strain of NBRISRM
was isolated on NA plates, containing 250 lg rifam-
picin (from Sigma Chemical Co., St. Louis, MO,
USA) as described earlier (Nautiyal 1997). Hetero-
geneous rhizosphere bacterial population was recov-
ered by serial dilution plating of the homogenate on
NA plates and NA plates amended with 50 lg
rifampicin/ml for NBRISRM. Average rhizosphere
colonization of NBRISRM (log10 CFU/g root dry
weight) was determined from four plants at the time
of harvesting each at different time interval as
indicated.
Effect of P. agglomerans NBRISRM inoculation
on functional diversity of maize and chickpea
rhizosphere based on carbon source utilization
pattern
Biolog Eco plates (Biolog, Inc., Hayward, CA, USA)
were used to determine the carbon source utilization
pattern as described earlier Nautiyal et al. (2010a, b).
Four rhizosphere soil samples were collected from
maize and chickpea plants with and without
NBRISRM inoculation. For each soil samples 12
samples were collected and three composite samples
were made by mixing four samples together. Indi-
vidual rhizosphere soil samples (1.0 g) were shaken
in 9.0 ml of sterile saline (0.85% NaCl w/v in Milli-Q
Water) for 60 min and then make up to a final
dilution of 10-3. After incubation, 150 ll of sample
Antonie van Leeuwenhoek (2011) 100:405–413 407
123

was inoculated in each well of Biolog Eco plates and
incubated at 30�C. The rate of utilization is indicated
by the reduction of tetrazolium, a redox indicator dye,
which changes from colorless to purple. Data were
recorded for 7 days at 590 nm and the results
obtained at fourth day were used for statistical
analysis. Microbial activity in each microplate,
expressed as average well color development
(AWCD) was determined as described by Garland
(1996). Diversity, richness and evenness indices were
calculated as described by Nautiyal et al. (2010a, b).
Statistical analyses were performed using SPSS 16.0
and Statistica 7.0.
Results
Isolation and identification of P. agglomerans
NBRISRM
A potential bacterial strain showing IAA production
and phosphate solubilization under in vitro conditions
was isolated from chickpea rhizosphere soil. Identi-
fication of SRM was based on 95 carbon source
utilization pattern using Biolog GN2 microtitre
panels by fully automated Biolog bacteria identifica-
tion system (Biolog Inc., Hayward, CA, USA) as
described earlier by Nautiyal et al. (2007). Further
identification of SRM was based on 16S rRNA gene
sequence analysis. A PCR product of 1,465 base pairs
16S rRNA gene was sequenced, and data were
analyzed as described earlier (Dastager et al. 2009).
The phylogenetic tree was constructed using 16s
rRNA gene sequence showing closest phylogenetic
relationship with P. agglomerans DSM3493T with
99% homology. Sequence data of SRM have been
deposited in the GenBank, nucleotide sequence
database was under the accession number
GQ225111 (Fig. 1).
IAA production and phosphate solubilization
by P. agglomerans NBRISRM
IAA production and phosphate solubilization of
NBRISRM was evaluated under in vitro conditions.
Figure 2 shows that NBRISRM produced 60.4 lg/ml
IAA and solubilized 77.5 lg/ml tri-calcium phosphate
after 72 h incubation under optimum conditions (0%
NaCl, pH-7 and 30�C) in their respective medium. IAA
production and phosphate solubilization of the
Pantoea agglomerans NBRISRM (GQ225111)
Pantoea agglomerans DSM 3493T(AJ233423)
Pantoea conspicua LMG 24534T(EU216737)
Pantoea brenneri LMG 5343T(EU216735)
Pantoea eucalypti LMG 24198T(EF688009)
Pantoea ananatis ATCC 19321T(U80209)
Pantoea vagans LMG 24199T(EF688012)
Pantoea anthophila LMG 2558T(EF688010)
Pantoea deleyi LMG 24200T(EF688011)
Pantoea stewartii LMG 2715T(Z96080)
Pantoea gaviniae A18/07T(GQ367483)
Pantoea calida1400/07T(GQ367478)
Erwinia toletana A64(AF130963)
Erwinia amylovora ATCC 15580T(U80195)
Erwinia tasmaniensis Et1/99T(AM055716)
Erwinia billingiae Eb661(AM055711)
Erwinia persicina ATCC 35998T(U80205)
Erwinia rhapontici ATCC 29283T(U80206)
100
71
53
98
91
90
85
73
83
60
77
71
77
0.005
Fig. 1 Neighbor-joining phylogenetic dendrogram based on
16S rRNA sequences showing relationships between P. ag-glomerans NBRISRM and related taxa. The numbers represent
the confidence levels from 1000 replicate bootstrap sampling.
Only the bootstrap percentages higher than 50% are shown at
branching points. Bar 0.005 substitutions per nucleotide
position
408 Antonie van Leeuwenhoek (2011) 100:405–413
123

NBRISRM was increased by 11 and 7%, respectively,
in treatments containing 2% NaCl (w/v). The addition
of 4% NaCl did not significantly changed IAA
production by NBRISRM. However, significant reduc-
tion by 45% was observed at pH 9, whereas phosphate
solubilization by NBRISRM was significantly reduced
at 4% NaCl (w/v) and pH-9 by 38 and 28%,
respectively.
Colonization and plant growth promotion
of P. agglomerans NBRISRM in maize
and chickpea rhizosphere under micro plot
conditions
Colonization of NBRISRM and heterogeneous bac-
terial population in maize and chickpea rhizospheres
under micro plot conditions were monitored. Popu-
lation of NBRISRM increased in the rhizosphere of
maize from 2.3 9 107 CFU/g root at 7 DAS to
2.22 9 108 CFU/g root on 30 DAS in non-sterilized
soil. Population of NBRISRM in chickpea rhizo-
sphere increased from 2.5 9 107 CFU/g root on 7
DAS to 2.3 9 108 CFU/g root on 30 DAS in non-
sterilized soil. After 30 DAS, colonization of
NBRISRM was 107 CFU/g rhizosphere soil, through-
out the growing season for both crops. Heterogeneous
bacterial populations during the same period were in
the range 107–109 CFU/g root in both maize and
chickpea rhizospheres. Under micro plot conditions,
NBRISRM inoculated maize and chickpea plants
showed significant increase in all plant growth
parameters as compared with un-inoculated control.
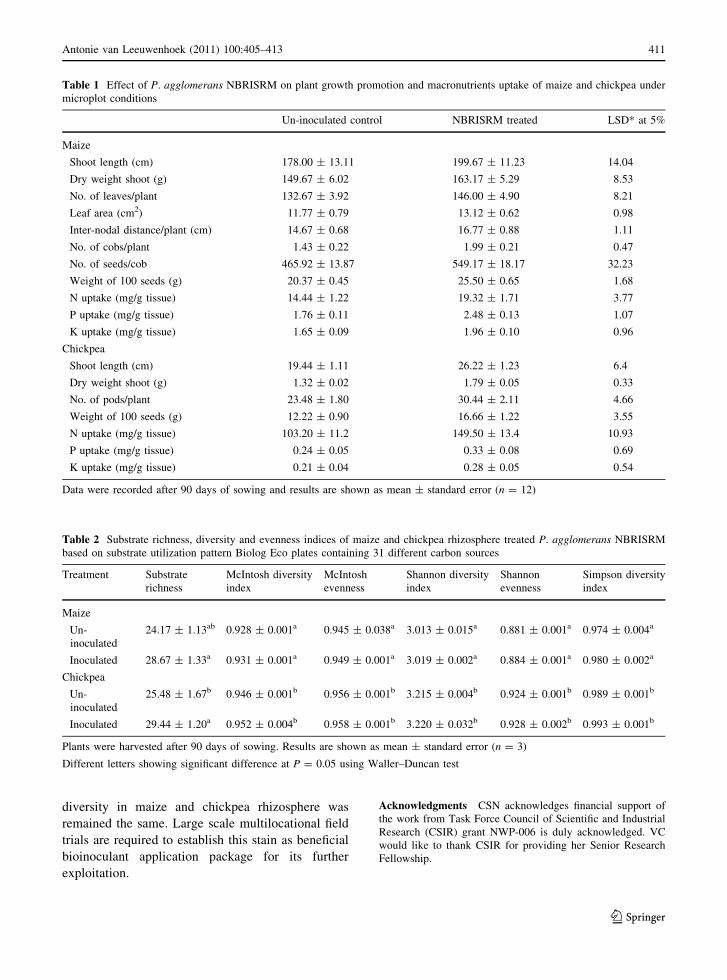
The treatment of maize with NBRISRM resulted in
significant increments by 11.2, 10.9, 11.2, 11.2, 12.1,
13.9, 11.8 and 12.5% in shoot length, shoot dry
weight, number of leaves plant-1, leaf area, inter-
nodal distance plant-1, number of cobs plant-1,
number of seeds cob-1 and weight of 100 seeds,
respectively, as compared to un-inoculated control.
NBRISRM inoculated chickpea plants have signifi-
cantly increase in shoot length, shoot dry weight,
number of pods and weight of 100 seeds by 13.5,
13.5, 12.9, 13.6%, respectively, as compared with un-
inoculated control. The plant-growth promotion abil-
ity of NBRISRM treatment in maize and chickpea
plants was also confirmed by significant increases in
macronutrients uptake. In NBRISRM treated maize
N, P and K accumulation increased by 34, 41, 19%,
respectively, whereas in chickpea by 45, 37, 33%,
respectively, compared to un-inoculated control.
Functional diversity analysis of maize
and chickpea rhizosphere
For assessing the effects of NBRISRM inoculation on
maize and chickpea rhizosphere functional diversity,
carbon source utilization patterns were used to
calculate the richness, diversity and evenness indices.
Overall substrate richness, diversity and evenness
were higher in the chickpea rhizosphere compared to
maize rhizosphere. Substrate richness were found
significantly (P = 0.05) higher in NBRISRM inocu-
lated maize and chickpea rhizosphere as compared to
their respective uninoculated control. Whereas, func-
tional diversity and evenness indices were also found
higher due to inoculation of NBRISRM compared to
uninoculated maize and chickpea rhizosphere, how-
ever, no significant differences (P = 0.05) were
observed using Waller–Duncan test (Fig. 3)
Discussion
Plant growth promoting P. agglomerans NBRISRM
was able to produce IAA and solubilize unavailable
phosphate up to some extent of high temperature,
salinity and alkalinity under in vitro conditions.
Similar to our results, earlier also many PGP strains
from Pantoea sp. were reported for phytohormone
production (Feng et al. 2006; Sergeeva et al. 2007,
Dastager et al. 2009) and phosphate solubilization
IAA
pro
duct
ion
/ P-s
olub
iliza
tion
(µg/
ml)
0
20
40
60
80
100IAA productionP-solbilization
30 0C 2% NaCl pH-937 0C 4% NaCl
Fig. 2 Indole acetic acid (IAA) production and phosphate
solubilization of P. agglomerans NBRISRM under in vitro
conditions. Error bars are the standard error of the means
(n = 3)
Antonie van Leeuwenhoek (2011) 100:405–413 409
123

(Son et al. 2006). Recently, we have reported the
exopolysacharide production and bio-film formation
in the same strain under in vitro conditions (Chauhan
and Nautiyal 2010) which clearly demonstrated that
this PGP strain has potential to perform under
different abiotically stressed environment.
Under micro-plot conditions, NBRISRM colo-
nized the rhizosphere of maize and chickpea plants
with 107–108 (CFU g-1 soil) population and com-
peted with the same titer of native heterotrophic
bacterial population throughout the growing season.
Similar to these results, earlier also we have reported
the colonization ability of P. agglomerans NBRISRM
in chickpea rhizosphere under pot culture conditions
(Chauhan and Nautiyal 2010). Inoculation of
NBRISRM has significantly improved the overall
growth of maize and chickpea. Plant growth promot-
ing abilities of Pantoea sp. has been already reported
for different crops such as rice (Feng et al. 2006;
Verma et al. 2001), wheat (Ruppel et al. 1992;
Amellal et al. 1998; Egamberdiyeva and Hoflich
2001) and cotton (Egamberdiyeva and Hoflich 2001).
Enhanced macronutrients (N, P and K) uptake was
also observed in the NBRISRM treated maize and
chickpea plants as compared to uninoculated plants.
Egamberdiyeva and Hoflich (2001) has reported
earlier that inoculation of Rhizobium trifolii R39
and P. agglomerans PF76/4 isolated from the mod-
erate German climate increased the root and shoot
growth of cotton and wheat in loamy sand soil and
resulted in significantly higher N, P and K contents of
plant (Table 1).
Carbon source utilization pattern-based functional
diversity indices were found lower in maize as
compared to chickpea rhizosphere resident micro-
flora. However, no significant differences (P = 0.5)
were observed in NBRISRM inoculated maize and
chickpea rhizosphere as compared to their respective
uninoculated controls (Table 2). Earlier also Mars-
chner et al. (2001) has reported the bacterial
community composition in the rhizosphere is affected
by a complex interaction between soil type and plant
species. One important factor for PGPR survival is
compatibility between the host plant root exudate
composition and the ability of the PGPR to utilize
those compounds (Strigul and Kravchenko 2006).
Carbon source utilization patterns by environmental
samples using Biolog Eco plate offer a relatively
faster and sensitive means of assessing the changes in
microbial functional diversity as compared to other
regular cultivable methods (Nautiyal 2009). In spite
of the various criticisms, the method remains to be
widely used to a large extent due to the ease and
speed of performance (Winding and Hendriksen
2007, Mishra and Nautiyal 2009; Nautiyal et al.
2010a, b, c). Information based on rhizosphere
microbial communities structure and their diversity
which are related to other essential processes within
the same system such as complexity, natural selec-
tion, symbiosis, parasitism, mutualism, competence,
succession or the effect of disturbances is a better
way to understand the system and for maximum
exploitation of PGPR.
Overall results conclude that the enhancement in
the maize and chickpea growth, nutrient uptake was
recorded under microplot conditions due to inocula-
tion of rhizosphere competent plant growth promot-
ing P. agglomerans NBRISRM. Moreover, functional
4
5
6
7
8
9
10
HeterotrophsNBRISRM
Time (days)0 20 40 60 80 100
Lo
g 1
0 C
FU
/g r
oo
t
4
5
6
7
8
9
A
B
Fig. 3 Survival and competence in the rhizosphere of a Zeamays and b Cicer arietinum L. by P. agglomerans NBRISRM.
Error bars are the standard error of the means (n = 6)
410 Antonie van Leeuwenhoek (2011) 100:405–413
123
diversity in maize and chickpea rhizosphere was
remained the same. Large scale multilocational field
trials are required to establish this stain as beneficial
bioinoculant application package for its further
exploitation.
Acknowledgments CSN acknowledges financial support of
the work from Task Force Council of Scientific and Industrial
Research (CSIR) grant NWP-006 is duly acknowledged. VC
would like to thank CSIR for providing her Senior Research
Fellowship.
Table 1 Effect of P. agglomerans NBRISRM on plant growth promotion and macronutrients uptake of maize and chickpea under
microplot conditions
Un-inoculated control NBRISRM treated LSD* at 5%
Maize
Shoot length (cm) 178.00 ± 13.11 199.67 ± 11.23 14.04
Dry weight shoot (g) 149.67 ± 6.02 163.17 ± 5.29 8.53
No. of leaves/plant 132.67 ± 3.92 146.00 ± 4.90 8.21
Leaf area (cm2) 11.77 ± 0.79 13.12 ± 0.62 0.98
Inter-nodal distance/plant (cm) 14.67 ± 0.68 16.77 ± 0.88 1.11
No. of cobs/plant 1.43 ± 0.22 1.99 ± 0.21 0.47
No. of seeds/cob 465.92 ± 13.87 549.17 ± 18.17 32.23
Weight of 100 seeds (g) 20.37 ± 0.45 25.50 ± 0.65 1.68
N uptake (mg/g tissue) 14.44 ± 1.22 19.32 ± 1.71 3.77
P uptake (mg/g tissue) 1.76 ± 0.11 2.48 ± 0.13 1.07
K uptake (mg/g tissue) 1.65 ± 0.09 1.96 ± 0.10 0.96
Chickpea
Shoot length (cm) 19.44 ± 1.11 26.22 ± 1.23 6.4
Dry weight shoot (g) 1.32 ± 0.02 1.79 ± 0.05 0.33
No. of pods/plant 23.48 ± 1.80 30.44 ± 2.11 4.66
Weight of 100 seeds (g) 12.22 ± 0.90 16.66 ± 1.22 3.55
N uptake (mg/g tissue) 103.20 ± 11.2 149.50 ± 13.4 10.93
P uptake (mg/g tissue) 0.24 ± 0.05 0.33 ± 0.08 0.69
K uptake (mg/g tissue) 0.21 ± 0.04 0.28 ± 0.05 0.54
Data were recorded after 90 days of sowing and results are shown as mean ± standard error (n = 12)
Table 2 Substrate richness, diversity and evenness indices of maize and chickpea rhizosphere treated P. agglomerans NBRISRM
based on substrate utilization pattern Biolog Eco plates containing 31 different carbon sources
Treatment Substrate
richness
McIntosh diversity
index
McIntosh
evenness
Shannon diversity
index
Shannon
evenness
Simpson diversity
index
Maize
Un-
inoculated
24.17 ± 1.13ab 0.928 ± 0.001a 0.945 ± 0.038a 3.013 ± 0.015a 0.881 ± 0.001a 0.974 ± 0.004a
Inoculated 28.67 ± 1.33a 0.931 ± 0.001a 0.949 ± 0.001a 3.019 ± 0.002a 0.884 ± 0.001a 0.980 ± 0.002a
Chickpea
Un-
inoculated
25.48 ± 1.67b 0.946 ± 0.001b 0.956 ± 0.001b 3.215 ± 0.004b 0.924 ± 0.001b 0.989 ± 0.001b
Inoculated 29.44 ± 1.20a 0.952 ± 0.004b 0.958 ± 0.001b 3.220 ± 0.032b 0.928 ± 0.002b 0.993 ± 0.001b
Plants were harvested after 90 days of sowing. Results are shown as mean ± standard error (n = 3)
Different letters showing significant difference at P = 0.05 using Waller–Duncan test
Antonie van Leeuwenhoek (2011) 100:405–413 411
123

Conflict of interest The authors have declared no conflict of
interest.
References
Amellal N, Burtin G, Bartoli F, Heulin T (1998) Colonization
of wheat roots by an exopolysaccharide-producing Pan-toea agglomerans strain and its effect on rhizosphere soil
aggregation. Appl Environ Microbiol 64:3740–3747
Andreote FD, Rocha UN, Araujo WL, Azevedo JL, van
Overbeek LS (2010) Effect of bacterial inoculation, plant
genotype and developmental stage on root-associated and
endophytic bacterial communities in potato (Solanum tu-berosum). Antonie van Leeuwenhoek 97:389–399
Babalola OO (2010) Beneficial bacteria of agricultural
importance. Biotechnol Lett 32:1559–1570
Braun-Kiewnick A, Jacobsen BJ, Sands DC (2000) Biological
control of Pseudomonas syringae pv. syringae, the causal
agent of basal kernel blight of barley, by Antagonistic
Pantoea agglomerans. Phytopathology 90:368–375
Brick M, Bostock RM, Silverstone SE (1991) Rapid in situ
assay for indoleacetic acid production by bacteria immo-
bilized on nitrocellulose membrane. Appl Environ
Microbiol 57:535–538
Canamas TP, Vinas I, Abadias M, Usall J, Torres R, Teixido N
(2009) Acid tolerance response induced in the biocontrol
agent Pantoea agglomerans CPA-2 and effect on its sur-
vival ability in acidic environments. Microbiol Res
164:438–450
Chauhan PS, Nautiyal CS (2010) The purB gene controls
rhizosphere colonization by Pantoea agglomerans. Lett
Appl Microbiol 50:205–210
Dastager SG, Deepa CK, Puneet SC, Nautiyal CS, Pandey A
(2009) Isolation and characterization of plant growth-
promoting strain Pantoea NII-186. From Western Ghat
forest soil, India. Lett Appl Microbiol 49:20–25
Egamberdiyeva D, Hoflich G (2001) Influence of growth pro-
moting bacteria from Uzbekistan and Germany on the
growth and nutrient uptake of cotton and wheat on dif-
ferent soils. In: Horst W, Schenk MK, Burkert A, Claas-
sen N, Flessa H, Frommer WB, Goldbach HE, Olfs HW,
Romheld V, Sattelmacher B, Schmidhalter U, Schubert S,
von Wiren N, Wittenmayer L (eds) Plant nutrition: food
security and sustainability of agro-ecosystems through
basic and applied research (Developments in plant and
soil sciences). Springer, Netherlands, pp 674–675
Felsenstein J (1985) Confidence limits on phylogenies: an
approach using the bootstrap. Evolution 39:783–791
Feng Y, Shen D, Song W (2006) Rice endophyte Pantoeaagglomerans YS19 promotes host plant growth and
affects allocations of host photosynthates. J Appl Micro-
biol 100:938–945
Fiske CH, Subbarow Y (1925) A colorimetric determination of
phosphorus. J Biol Chem 66:375–400
Garland JL (1996) Analytical approaches to the characteriza-
tion of samples of microbial communities using patterns
of potential C source utilization. Soil Biol Biochem
28:213–221
Hynes RK, Leung GCY, Hirkala DLM, Nelson LM (2008)
Isolation, selection, and characterization of beneficial
rhizobacteria from pea, lentil, and chickpea grown in
western Canada. Can J Microbiol 54:248–258
Jukes TH, Cantor CR (1969) Evolution of protein molecules.
In: Munro HN (ed) Mammalian protein 242 metabolism.
Academic Press, New York, pp 21–132
Loiret FG, Ortega E, Kleiner D, Ortega-Rodes P, Rodes R,
Dong Z (2004) A putative new endophytic nitrogen-fixing
bacterium Pantoea sp. from sugarcane. J Appl Microbiol
97:504–511
Long HH, Schmidt DD, Baldwin IT (2008) Native bacterial
endophytes promote host growth in a species-specific
manner; phytohormone manipulations do not result in
common growth responses. PLoS One 3:e2702
Lugtenberg BJJ, Dekkers L, Bloemberg GV (2001) Molecular
determinants of rhizosphere colonization by Pseudomo-nas. Ann Rev Phytopathol 39:461–490
Marschner P, Yang C-H, Lieberei R, Crowley DE (2001) Soil
and plant specific effects on bacterial community com-
position in the rhizosphere. Soil Biol Biochem
33:1437–1445
Mehta S, Nautiyal CS (2001) An efficient method for quali-
tative screening of phosphate-solubilizing bacteria. Curr
Microbiol 43:51–56
Mishra A, Nautiyal CS (2009) Functional diversity of the
microbial community in the rhizosphere of chickpea
grown in diesel fuel spiked soil amended with Tricho-derma ressei using sole-carbon source utilization profiles.
World J Microbiol Biotechnol 25:1175–1180
Nautiyal CS (1997) A method for selection and characteriza-
tion of rhizosphere-competent bacteria of chickpea. Curr
Microbiol 34:12–17
Nautiyal CS (1999) An efficient microbiological growth
medium for screening phosphate solubilizing microor-
ganisms. FEMS Microbiol Lett 170:265–720
Nautiyal CS (2009) Self-purificatory Ganga water facilitates
death of pathogenic Escherichia coli 157:H7. Curr
Microbiol 58:25–29
Nautiyal CS, Chauhan PS, Nene YL (2007) Medicinal smoke
reduces airborne bacteria. J Ethnopharmacol 114:
446–451
Nautiyal CS, Srivastava S, Chauhan PS (2008) Rhizosphere
colonization Molecular determinants from plant-microbe
coexistence perspective. In: Nautiyal CS, Dion P (eds)
Molecular mechanisms of plant, microbe coexistence soil
biology series, vol 15. Springer, Berlin, pp 99–124
Nautiyal CS, Chauhan PS, Bhatia CR (2010a) Changes in soil
physico-chemical properties and microbial functional
diversity due to 14 years of conversion of grassland to
organic agriculture in semi-arid agroecosystem. Soil Till
Res 109:55–60
Nautiyal CS, Chauhan PS, DasGupta SM, Seem K, Varma A,
Staddon WJ (2010b) Tripartite interactions among Pae-nibacillus lentimorbus NRRL B-30488, Piriformosporaindica DSM 11827, and Cicer arietinum L. World J
Microbiol Biotechnol 26:1393–1399
Nautiyal CS, Rehman A, Chauhan PS (2010c) Environmental
Escherichia coli occur as natural plant growth-promoting
soil bacterium. Arch Microbiol 192:185–193
Olsen SR, Sommers LE (1982) Phosphorus. In: Page AL,
Miller RH, Keeney DR (eds) Methods of soil analysis:
part 2. SSSA, Madison, p 403
412 Antonie van Leeuwenhoek (2011) 100:405–413
123

Page AL, Miller RH, Keeney DR (1982) Methods of soil
analysis part 2.1159 chemical and microbiological prop-
erties (2nd ed) agronomy, vol 9. ASA SSSA Publishing,
Madison, p 1159
Ruppel S, Hecht-Buchholz C, Remus R, Ortmann U, Schmel-
zer R (1992) Settlement of the diazotrophic, phytoeffec-
tive bacterial strain Pantoea agglomerans on and within
winter wheat: an investigation using ELISA and trans-
mission electron microscopy. Plant Soil 145:261–273
Salles JF, van Veen JA, van Elsas JD (2004) Multivariate
analyses of Burkholderia species in soil: effect of crop
and land use history. Appl Environ Microbiol 70:
4012–4020
Sergeeva E, Hirkala DLM, Nelson LM (2007) Production of
indole-3-acetic acid, aromatic amino acid aminotransfer-
ase activities and plant growth promotion by Pantoeaagglomerans rhizosphere isolates. Plant Soil 297:1–13
Son HJ, Park GT, Cha MS, Heo MS (2006) Solubilization of
insoluble inorganic phosphates by a novel salt- and pH-
tolerant Pantoea agglomerans R-42 isolated from soybean
rhizosphere. Bioresour Technol 97:204–210
Strigul NS, Kravchenko LV (2006) Mathematical modeling of
PGPR inoculation into the rhizosphere. Environ Mod Soft
21:1158–1171
Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins
DG (1997) The CLUSTAL_X windows interface: flexible
strategies for multiple sequence alignment aided by
quality analysis tools. Nucleic Acids Res 25:4876–4888
Verma SC, Ladha JK, Tripathi AK (2001) Evaluation of plant
growth promoting and colonization ability of endophytic
diazotrophs from deep water rice. J Biotechnol 91:127–141
Winding A, Hendriksen N (2007) Comparison of CLPP and
enzyme activity assay for functional characterization of
bacterial soil communities. J Soils Sed 7:411–417
Wright SAI, Zumoff CH, Schneider L, Beer SV (2001) Pan-toea agglomerans strain EH318 produces two antibiotics
that inhibit Erwinia amylovora in vitro. Appl Environ
Microbiol 67:284–292
Antonie van Leeuwenhoek (2011) 100:405–413 413
123




















