
Effects of ocean acidification on marine biodiversity and ecosystem function
Upload
independentCategory
view
0download
0
REGULAR ARTICLE
Positive feedback between acidification and organic phosphatemineralization in the rhizosphere of maize (Zea mays L.)
Xiaodong Ding & Li Fu & Chenjia Liu &
Fanjun Chen & Ellis Hoffland & Jianbo Shen &
Fusuo Zhang & Gu Feng
Received: 22 December 2010 /Accepted: 24 March 2011 /Published online: 7 April 2011# Springer Science+Business Media B.V. 2011
Abstract To test the hypothesis that rhizosphereacidification would enhance the hydrolyzation oforganic phosphates by increasing phosphatase activity.A Petri dish experiment with sterile agar and a potexperiment with a low P soil were used. In the Petri dishexperiment, roots of each plant were cultured in twocompartments, each of which contained agar with oneof three nitrogen combinations: NH4
+/N0 (N0 =nitrogen free), NH4
+/NO3-, and NO3
-/N0. Phytin wassupplied as the sole phosphorus (P) source to allcompartments. In the pot experiment, the soil in eachpot was treated with N0, KNO3, or (NH4)2SO4)together with 0 or 75 mg kg−1 phytin-P. Dry weight,P concentration, and P content of roots were highest inthe NH4
+ compartments in the Petri dish experiment.In the pot experiment, dry weight, P concentration, and
P content of both shoots and roots were higher withNH4
+ than with NO3-. NH4
+ treatments reducedrhizosphere pH, promoted the hydrolization of phytin,enhanced acid phosphatase activity in the rhizosphere,and increased phytin-P utilization relative to N0 andNO3
- treatments. Phosphatase activity was negativelycorrelated with rhizosphere pH but was positivelycorrelated with plant P content in both experiments.Rhizosphere acidification optimized the activity of acidphosphatase excreted by maize roots and promotedphytin mineralization. NH4
+-induced acidification inthe maize rhizosphere improved the growth of maizeroots by improving P uptake from phytin; theimproved growth, in turn, increased NH4
+ uptake andacidification.
Keywords Acidification . Rhizosphere . Organicphosphate . Acid phosphatases . Zea mays L.
Introduction
Plants use various strategies to acquire phosphorus(P) from inorganic or organic phosphate pools in soil.For example, plant roots may excrete carboxylates orprotons to mobilize the nonsoluble inorganic phos-phates (Wang et al. 2008; Lambers et al. 2008; Rengeland Marschner 2005) or excrete acid phosphatase tomineralize organic phosphates (Wang et al. 2008;Turner 2008; Giaveno et al. 2010).
Organic P represents 20–80% of the total P in mostsoils and consequently represents a significant reserve
Plant Soil (2011) 349:13–24DOI 10.1007/s11104-011-0783-1
Responsible Editor: Tim Simon George.
X. Ding : L. Fu : C. Liu : F. Chen : J. Shen : F. Zhang :G. Feng (*)College of Resource and Environmental Science,China Agricultural University, Key Laboratory of PlantNutrition, MOA; Key Laboratory of Plant-Soil Interactions,MOE,Beijing 100193, People’s Republic of Chinae-mail: [email protected]
X. DingSouth Subtropical Crop Research Institute,Chinese Academy of Tropical Agricultural Science,Zhanjiang, Guangdong 524091, People’s Republic of China
E. HofflandDepartment of Soil Quality, Wageningen University,P.O. Box 47, 6700 AA Wageningen, The Netherlands

of potentially available P (Fransson and Jones 2007).Organic P mainly consists of a mixture of phos-phate monoesters or/and phosphate diesters thatmust first be mineralized by phosphomonoesteraseand phosphodiesterase, respectively, to releasephosphates that are available to the plant (Turner2008). Phosphatases with various substrate specific-ities (e.g., phosphomono- and phosphodi-esterases)and pH optima have been characterized for plantroots (Bosse and Köck 1998; Tadano and Sakai1991), bacteria (Tang et al. 2006), and fungi (Bucher2007; Feng et al. 2003; Koide and Kabir 2000). Theactivities of phosphatases are pH dependent. Theyare classified as alkaline or acid phosphatases if theiroptimal pH for catalysis is above or below pH 7.0,respectively (Duff et al. 1994; Vincent et al. 1992).The two types of phosphatase are usually measuredin acid or alkaline pH buffer (George et al. 2002). Itis reported that plant-produced phosphatase is exclu-sively acid phosphatase (Tarafdar and Claassen,1988; Tarafdar and Marschner, 1994; George et al.2002). Alkaline phosphatase does not have a plantorigin and is produced by bacteria, fungi and earth-worms (Hebrien and Neal, 1990).
Rhizosphere soil becomes acidified when plants areP deficient or absorb more cations, such as NH4
+, thananions (Gahoonia et al. 1992; Nye 1981). In calcareousor neutral soils, root proton exudation can reduce therhizosphere pH by more than 1 unit compared to thebulk soil (Li et al. 1991; Tagliavini et al. 1995), andthis has been considered to be an important way bywhich plants solubilise and forage for inorganicphosphates from soil. It is still not known whether achange in rhizosphere pH can alter the activity of acidphosphatases and consequently alter the mineralizationrate of organic phosphates. In the current study, wetested the hypothesis that rhizosphere acidificationinduced by uptake of NH4
+ from a neutral soil willprovide an optimal pH for acid phosphatase activity inthe rhizosphere, and consequently will increase maizeuptake of P from phytin (a common form of organic Pin soil). We also hypothesized that this increasedbioavailability of P from phytin will result inincreased plant growth, which will in turn increasethe uptake of NH4
+ and the release of protons intoand acidification of the rhizosphere. If our hypoth-eses is correct, it would describe a positive feedbackmechanism in the rhizosphere that has not beendemonstrated before.
Materials and methods
The study consisted of two experiments, one in Petridishes under axenic conditions with agar as thesubstrate, and the other in pots containing nonsterilesoil. The P-efficient maize (Zea mays L.) genotype181, which was developed for cultivation on calcar-eous soils in the northeastern China, was used in bothexperiments. The P uptake-related root physiologicaltraits of genotype 181, e.g., proton and acid phospha-tase excretion under P-deficient conditions, have beenpreviously described (Liu et al. 2004).
Petri dish experiment
An axenic cultivation system was used to exclude anyeffect of microbial acid phosphatase. The systemconsisted of a two-compartment Petri dish (150 mmdiameter, 14 mm depth) and a 550-mL plastic bottle(Fig. 1). The two-compartment Petri dish allowedspatial separation of two root compartments (RC1 andRC2). A filter (<0.45 μm pore diameter) was glued onthe top of the bottle allowed gas exchange but preventedcontamination. The root compartments were filled withsterilized nutrient-containing agar (see next paragraph).The merit of this split-root system is that it can be usedto compare the response of the same root system toadditions of NH4
+ (applied to one half) or NO3-
(applied to the other half) at the same shoot P demand.
CC
AMD
RC1 RC2
SC
Shoot
Root
Barrier
Fig. 1 Diagram of the Petri dish system used in the Petri dishexperiment. The two root compartments (RC1 and RC2) containsterilized agar with different N forms. CC, Culture Compartment;SC, Shoot Compartment; AMD, Adhesive Microfiltration Disc
14 Plant Soil (2011) 349:13–24

Seeds of maize were disinfested by placing them in10% hydrogen peroxide (H2O2) for 10 min and then in70% ethanol for 3 min. The seeds were then washed10–12 times in sterile-distilled water to remove alltraces of the sterilizing chemicals. The disinfested seedswere subsequently transferred aseptically to sterile agarplates. The agar powder, which was purchased fromShanghai Genebase Gene-Tech Co., Ltd., China (CAS:9002-19-0; Prod. No. A-2180), was P free. Althoughthis agar is solid and transparent when prepared as a0.6% medium, it becomes liquid again at roomtemperature upon stirring. The seeds were germinatedin a growth chamber at 27°C in the dark. The endospermwas removed before transplanting. The plant at trans-plantation contained 0.15±0.01mg P and 4.56±0.25mgN. The root system of each 3-day-old maize seedlingwas divided into two halves, each of which was placedin a separate compartment of the Petri dish (Fig. 1).
The experiment had a completely randomizeddesign. The main treatments were three combinationsof nitrogen forms referred to as NH4
+/N0, NH4+/
NO3-, and NO3
-/N0. These combinations describe theN supplied to the two halves of the root system, i.e.,the two compartments (RC1 and RC2) of each Petridish. NH4
+/N0 indicates that NH4+ was added to one
half and no nitrogen was added to the other, etc.The basic agar medium contained (in mM): K2SO4
1.5, KCl 0.2, CaCl2 4.0, MgSO4 1.3, MnSO4 1.0×10−4, (NH4)6Mo7O24 5.0×10−6, H3BO3 1×10−3,ZnSO4 1.0×10−4, CuSO4 5×10−4, and Fe-EDTA3.6×10−2. Phytin (0.37 gL−1), which had a P contentof 20.9% and was purchased from Tokyo Kasei Co.Ltd. (Tokyo, Japan), was added to the nutrientsolution before sterilization. The pH was adjusted to6.5 with 1.0 mmol L−1 NaOH or HCl. The solutionwas sterilized (120°C, 30 min), cooled, and pouredinto Petri dishes (40 mL per compartment). NH4
+ andNO3
- were added to the agar as required by thetreatment combination before sterilization as NH4Cland Ca(NO3)2, respectively, and both were at 4 mM.
Six replicate systems were prepared for each treat-ment. The systems were incubated in a growth chamberwith 12 h light, light intensity 500 μmol m−2 s−1, arelative humidity of 70%, and temperature of 25°C. Theseedlings were harvested 21 days after transplanting.
For determination of rhizosphere pH, agar wascollected with a mini-scoop along a 60-mm length(ending at the root tip) of the first order lateral root.Three scoops of agar were collected with an interval
of 20 mm at each side of the root and within 2 mm ofthe root surface. All six scoops of agar collected fromeach root compartment were put together in a 2-mLEppendorf tube and stirred with tweezers until theagar liquefied. The pH of the liquefied agar, i.e., therhizosphere pH, was measured with a microelectrode(Waterproof pHTestr® BNC 10 pH tester, EutechInstruments Pte Ltd., Singapore).
Standard assays of acid phosphatase are usuallyconducted at the optimal pH (pH 6.5) to allowcomparison of potential acid phosphatase activities(Carreira et al. 2000). In the current study, however,we wanted to measure acid phosphatase activity at thepH in situ. In measuring acid phosphatase activity inthe rhizosphere and in root exudates, we thereforeused pH buffers with pH values similar to thosemeasured in the rhizosphere of our experiments asdescribed in the next paragraph.
Maize seedlings were removed from the two-compartment Petri dish. Agar blocks were collectedfrom a zone that was less than 2 mm around andbeneath the root print, and the agar blocks were mixedtogether in a tube. One mL of each liquefied agar samplewas pipetted into a 5-mL Eppendorf tube that contained2 mL of a 50 mM acetate buffer with the meanrhizosphere pH of a similar rhizosphere. Subsequently,0.4 mL of the substrate (150 mM p-nitrophenylphosphate, NPP; Sigma St. Louis, MO, USA) in acetatebuffer was added. Themixture was incubated at 20°C for15 min. The reaction was terminated by adding 2.5 mLof 0.5 M NaOH. After the mixture was centrifuged for5 min at 1,500g, acid phosphatase activity in therhizosphere was quantified by comparing the absorptionat 410 nm to a standard curve of diluted p-nitrophenolphosphate solutions (Carreira et al. 2000).
To assay acid phosphatase activity in root exudates,roots were cut from the stem and gently removed fromthe agar medium. All the roots in each compartmentwere incubated in flasks containing 10 mL of 50 mMacetate buffer for 5 min. Then, 2 mL of the incubationsolution was added to 0.4 mL substrate solution (seeabove). The rest of the procedure was as describedabove. The total length and dry weight of the rootsegments were determined.
For measurement of N and P concentration, roots andshoots were separated and placed in an oven at 105°C for30 min to stop metabolic activity. They were then driedat 65°C for 72 h, weighed, and ground (1mm). The driedand sieved plant material was digested in a H2SO4-H2O2
Plant Soil (2011) 349:13–24 15

mixture at 370°C for 2 h. Nitrogen was determined bythe Kjeldahl method (Shi 1994), and P was determinedby the molybdenum blue method (Allen 1974).
Pot experiment
A moderately acid (pHwater 6.4) clay soil was collectedin Tai’an, Shandong Province, China. It contained7.27 g/kg organic matter, 7.2 mg kg−1 mineral nitrogen(NO3
- and NH4+, analyzed using Continuous Flow
Analysis [TRAACS 2000]), 253 mg kg−1 total organicphosphorus (assessed by Saunders and Williams1955), 3.3 mg kg−1Olsen-P, and 97.6 mg kg−1
NH4Cl-exchangeable K. The soil was air-dried, sieved(2 mm), and sterilized with γ radiation (RadiationDosimetry: 10 kGy 60Co γ-ray, supported by BeijingRadiation Application Research Center) to kill indig-enous microorganisms. The soil (1,800 g) was mixedwith 200 g of fine river sand to reduce compactness ofthe soil and placed in a 19×18×16 cm plastic pot. Soilmoisture was kept at 18–20% (w/w, about 70% ofwater holding capacity) as determined by weighing thepots every 2 days during the experiment. All potsreceived basal mineral nutrients as follows: 50 mg Mgper kg of soil (as MgSO4), 5 mg Cu per kg of soil (asCuSO4), and 5 mg Zn per kg of soil (as ZnSO4). Threeseeds were sown in each pot. Seedlings were thinnedto one per pot after emergence.
The experiment was conducted in a greenhousewith temperatures ranging from 24 to 30°C. Theexperimental design was a randomized block withtwo factors: (1) two organic P levels and (2) three Nforms. The organic P levels were no organic P (P0)and 75 mg kg−1 P supplied as phytin (P75). The formsof N were N0 (no N), and 200 mg kg−1N as KNO3 or200 mg kg−1N as (NH4)2SO4. The nitrificationinhibitor 3, 4-dimethylpyrazole phosphate (DMPP)was added to N0 (no N), KNO3 and (NH4)2SO4 at arate of 1% of nitrogen applied. There were fourreplicate pots for each treatment.
The plants were harvested 6 weeks after emer-gence. Rhizosphere soil was collected by removingthe soil that adhered to the roots after the roots wereshaken; a brush was used to remove the rhizospheresoil. The pH of the rhizosphere soil wasmeasured in a 1:5soil to water (w/v) mixture. Acid phosphatase activities inthe rhizosphere soil were measured using buffers with apH according to the measured rhizosphere pH.
The harvested roots and shoots were dried at 65–70°C for 3 days and weighed. N and P concentrationwere estimated by the same method as in Petri dishexperiment.
Statistics analysis
A test of Normality and Bartlett’s test for homogeneityof variance were conducted by PROC UNIVARIATEusing the SAS™ software (SAS Institute Inc. 1989) toconfirm that all data were normally distributed and thevariances were homogenous. Then the data from thePetri dish experiment were subjected to a one-wayANOVA, and data from the pot experiment weresubjected to a two-way ANOVA. Treatment meanswere compared by Tukey at P=0.05. Correlationanalysis was also performed. SAS™ software wasused for all statistical analyses.
Results
Effects of NH4+ and NO3
- on the growth of maize
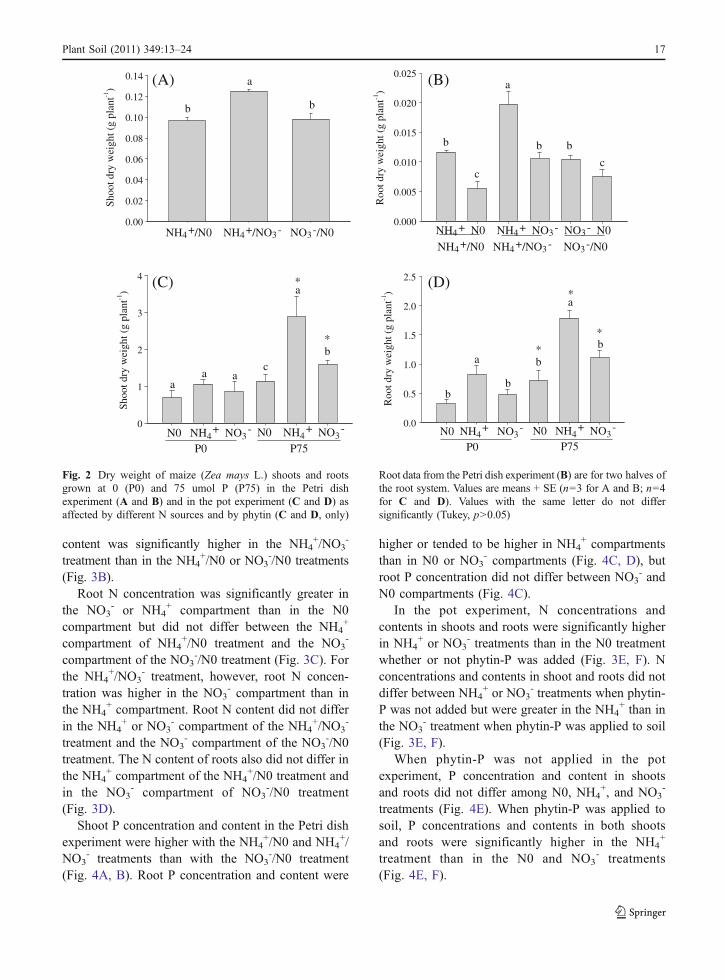
In the Petri dish experiment, the dry weight of shootsdepended on the N combination that was supplied (P=0.002) (Fig. 2A). Shoot dry weight was significantlyhigher in the NH4
+/NO3- treatment than in the NO3
-/N0and NH4
+/N0 treatments, while the difference betweenthe latter two treatments was not significant (Fig. 2A).
Both NH4+ and NO3
- promoted root growth, butroot growth was greater with NH4
+ than with NO3-
(Fig. 2B). Root dry weights were lowest with N0.In the pot experiment, NH4
+ and NO3- increased
the shoot growth only if phytin-P was applied to thesoil; there were no significant differences in shoot dryweights among three nitrogen treatments when no Pwas added (P0) (Fig. 2C). When phytin was added,root dry weight was significantly higher in the NH4
+
treatment than in the NO3- or N0 treatments (Fig. 2D).
The statistical interaction between phytin-P andnitrogen treatments was significant (P=0.000) forboth shoot and root dry weight.
Effects of NH4+ and NO3
- on N and P uptake by plants
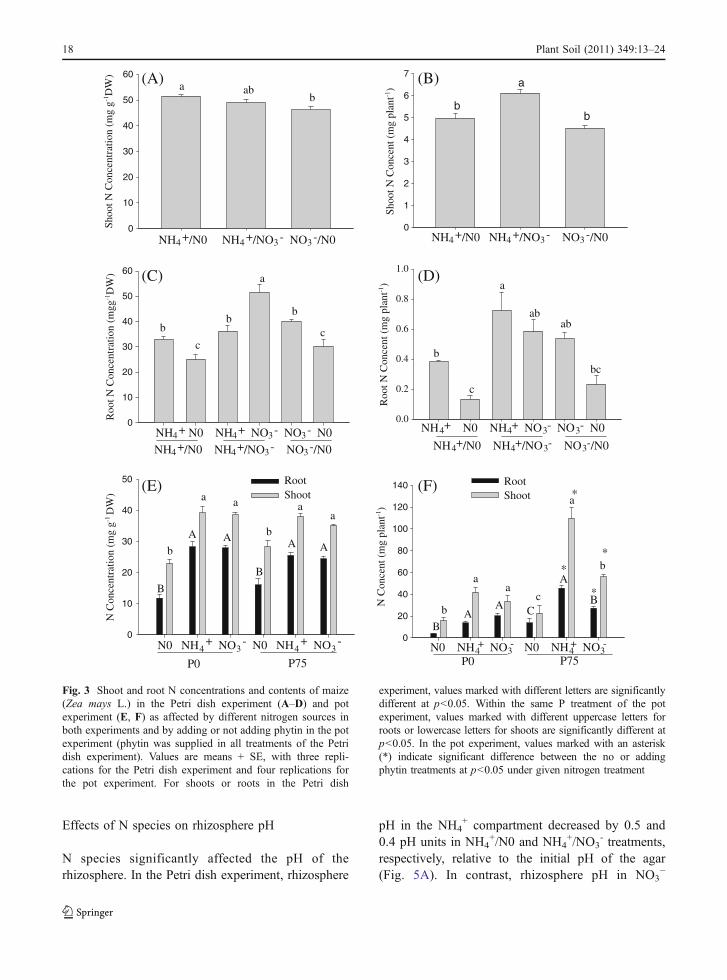
In the Petri dish experiment, shoot N concentration wasnot affected by the treatments (Fig. 3A), but shoot N
16 Plant Soil (2011) 349:13–24
content was significantly higher in the NH4+/NO3
-
treatment than in the NH4+/N0 or NO3
-/N0 treatments(Fig. 3B).
Root N concentration was significantly greater inthe NO3
- or NH4+ compartment than in the N0
compartment but did not differ between the NH4+
compartment of NH4+/N0 treatment and the NO3
-
compartment of the NO3-/N0 treatment (Fig. 3C). For
the NH4+/NO3
- treatment, however, root N concen-tration was higher in the NO3
- compartment than inthe NH4
+ compartment. Root N content did not differin the NH4
+ or NO3- compartment of the NH4
+/NO3-
treatment and the NO3- compartment of the NO3
-/N0treatment. The N content of roots also did not differ inthe NH4
+ compartment of the NH4+/N0 treatment and
in the NO3- compartment of NO3
-/N0 treatment(Fig. 3D).
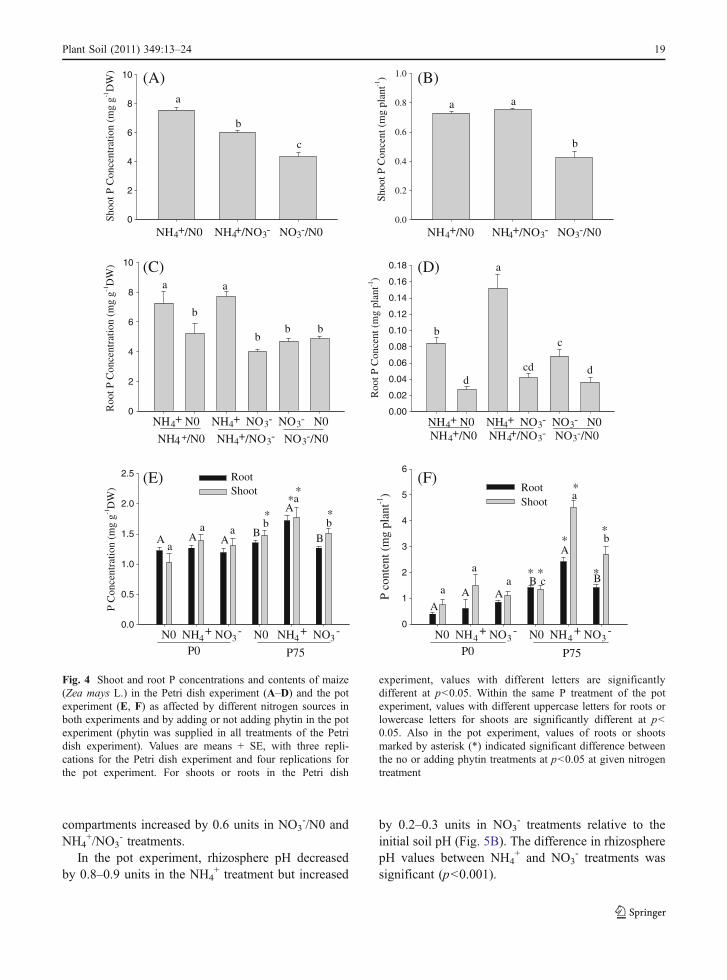
Shoot P concentration and content in the Petri dishexperiment were higher with the NH4
+/N0 and NH4+/
NO3- treatments than with the NO3
-/N0 treatment(Fig. 4A, B). Root P concentration and content were
higher or tended to be higher in NH4+ compartments
than in N0 or NO3- compartments (Fig. 4C, D), but
root P concentration did not differ between NO3- and
N0 compartments (Fig. 4C).In the pot experiment, N concentrations and
contents in shoots and roots were significantly higherin NH4
+ or NO3- treatments than in the N0 treatment
whether or not phytin-P was added (Fig. 3E, F). Nconcentrations and contents in shoot and roots did notdiffer between NH4
+ or NO3- treatments when phytin-
P was not added but were greater in the NH4+ than in
the NO3- treatment when phytin-P was applied to soil
(Fig. 3E, F).When phytin-P was not applied in the pot
experiment, P concentration and content in shootsand roots did not differ among N0, NH4
+, and NO3-
treatments (Fig. 4E). When phytin-P was applied tosoil, P concentrations and contents in both shootsand roots were significantly higher in the NH4
+
treatment than in the N0 and NO3- treatments
(Fig. 4E, F).
0.000
0.005
0.010
0.015
0.020
0.025 a
b b b
cc
NH4+ N0 NH4+ NO3- NO3- N0
NH4+/N0 NH4+/NO3- NO3-/N0
Roo
t dry
wei
ght (
g pl
ant-1
)R
oot d
ry w
eigh
t (g
plan
t-1)
(B)
Shoo
t dry
wei
ght (
g pl
ant-1
)Sh
oot d
ry w
eigh
t (g
plan
t-1)
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0.14
b
a
b
NH4+/N0 NH4+/NO3- NO3-/N0
(A)
0
1
2
3
4
bc
aaa
a
N0 NH4+ NO3
-
P0 P75
*
*
N0 NH4+ NO3
-
(C)
0.0
0.5
1.0
1.5
2.0
2.5
N0 NH4+ NO3
-
P0 P75N0 NH4
+ NO3-
a
ba b
bb
*
*
*
(D)
Fig. 2 Dry weight of maize (Zea mays L.) shoots and rootsgrown at 0 (P0) and 75 umol P (P75) in the Petri dishexperiment (A and B) and in the pot experiment (C and D) asaffected by different N sources and by phytin (C and D, only)
Root data from the Petri dish experiment (B) are for two halves ofthe root system. Values are means + SE (n=3 for A and B; n=4for C and D). Values with the same letter do not differsignificantly (Tukey, p>0.05)
Plant Soil (2011) 349:13–24 17
Effects of N species on rhizosphere pH
N species significantly affected the pH of therhizosphere. In the Petri dish experiment, rhizosphere
pH in the NH4+ compartment decreased by 0.5 and
0.4 pH units in NH4+/N0 and NH4
+/NO3- treatments,
respectively, relative to the initial pH of the agar(Fig. 5A). In contrast, rhizosphere pH in NO3
−
0
10
20
30
40
50
60 a
bb
b cc
NH4+ N0 NH4+ NO3- NO3- N0NH4+/N0 NH4+/NO3- NO3-/N0
(C)
0
20
40
60
80
100
120
140 RootShoot
aa
b AA
B
a
b
c
A
BC
N0 NH4+ NO3
-
P0 P75
*
*
*
*
N0 NH4+ NO3
-
(F)
0
10
20
30
40
50 RootShoot
B
a
b
A A
B
a aa
bA A
N0 NH4+ NO3
-
P0 P75
N0 NH4+ NO3
-
(E)
0.0
0.2
0.4
0.6
0.8
1.0
a
abab
b
bc
c
NH4+ N0 NH4+ NO3- NO3- N0
NH4+/N0 NH4+/NO3- NO3-/N0
(D)
0
1
2
3
4
5
6
7a
bb
NH4 +/N0 NH4 +/NO3 - NO3 -/N0
(B)
Shoo
t N C
once
ntra
tion
(mg
g-1D
W)
N C
once
ntra
tion
(mg
g-1 D
W)
Roo
t N C
once
ntra
tion
(mgg
-1D
W)
Shoo
t N C
once
nt (
mg
plan
t-1)
Roo
t N C
once
nt (
mg
plan
t-1)
N C
once
nt (
mg
plan
t-1)
0
10
20
30
40
50
60a ab
b
NH4 +/N0 NH4 +/NO3 - NO3 -/N0
(A)
Fig. 3 Shoot and root N concentrations and contents of maize(Zea mays L.) in the Petri dish experiment (A–D) and potexperiment (E, F) as affected by different nitrogen sources inboth experiments and by adding or not adding phytin in the potexperiment (phytin was supplied in all treatments of the Petridish experiment). Values are means + SE, with three repli-cations for the Petri dish experiment and four replications forthe pot experiment. For shoots or roots in the Petri dish
experiment, values marked with different letters are significantlydifferent at p<0.05. Within the same P treatment of the potexperiment, values marked with different uppercase letters forroots or lowercase letters for shoots are significantly different atp<0.05. In the pot experiment, values marked with an asterisk(*) indicate significant difference between the no or addingphytin treatments at p<0.05 under given nitrogen treatment
18 Plant Soil (2011) 349:13–24
compartments increased by 0.6 units in NO3-/N0 and
NH4+/NO3
- treatments.In the pot experiment, rhizosphere pH decreased
by 0.8–0.9 units in the NH4+ treatment but increased
by 0.2–0.3 units in NO3- treatments relative to the
initial soil pH (Fig. 5B). The difference in rhizospherepH values between NH4
+ and NO3- treatments was
significant (p<0.001).
Shoo
t P C
once
ntra
tion
(mg
g-1D
W)
Roo
t P C
once
ntra
tion
(mg
g-1D
W)
P C
once
ntra
tion
(mg
g-1D
W)
Shoo
t P C
once
nt (
mg
plan
t-1)
Roo
t P C
once
nt (
mg
plan
t-1)
0
2
4
6
8
10
a
b
c
NH4+/N0 NH4+/NO3- NO3-/N0
(A)
0
2
4
6
8
10
aa
b
NH4+ N0 NH4+ NO3- NO3- N0
NH4+/N0 NH4+/NO3- NO3-/N0
b
b b
(C)
0.0
0.2
0.4
0.6
0.8
1.0
NH4+/N0 NH4+/NO3- NO3-/N0
a a
b
(B)
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0.14
0.16
0.18 a
bc
cd d
d
NH4+ N0 NH4+ NO3- NO3- N0NH4+/N0 NH4+/NO3- NO3-/N0
(D)
0.0
0.5
1.0
1.5
2.0
2.5 RootShoot
a bA
BB
N0 NH4+ NO3
-
P0 P75
a
a
a
bAA A
* *
*
*
(E)
N0 NH4+ NO3
-
P co
nten
t (m
g pl
ant-1
)
0
1
2
3
4
5
6
Root Shoot
a
Ab
c B B a
A
a a
A A
*
*
*
*
* *
(F)
N0 NH4+ NO3
-
P0 P75
N0 NH4+ NO3
-
Fig. 4 Shoot and root P concentrations and contents of maize(Zea mays L.) in the Petri dish experiment (A–D) and the potexperiment (E, F) as affected by different nitrogen sources inboth experiments and by adding or not adding phytin in the potexperiment (phytin was supplied in all treatments of the Petridish experiment). Values are means + SE, with three repli-cations for the Petri dish experiment and four replications forthe pot experiment. For shoots or roots in the Petri dish
experiment, values with different letters are significantlydifferent at p<0.05. Within the same P treatment of the potexperiment, values with different uppercase letters for roots orlowercase letters for shoots are significantly different at p<0.05. Also in the pot experiment, values of roots or shootsmarked by asterisk (*) indicated significant difference betweenthe no or adding phytin treatments at p<0.05 at given nitrogentreatment
Plant Soil (2011) 349:13–24 19
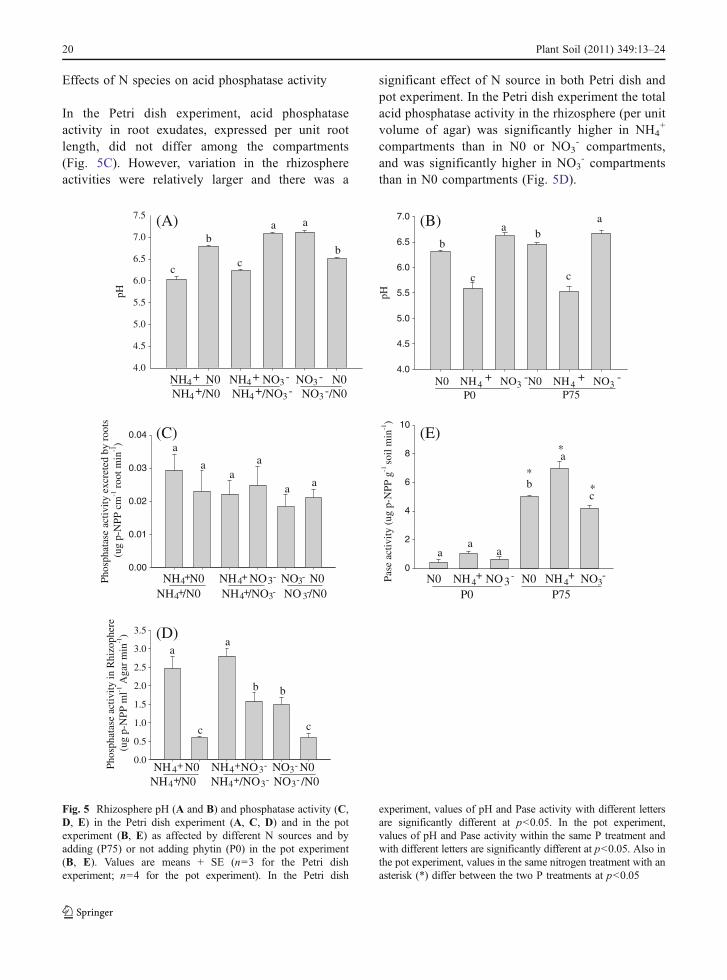
Effects of N species on acid phosphatase activity
In the Petri dish experiment, acid phosphataseactivity in root exudates, expressed per unit rootlength, did not differ among the compartments(Fig. 5C). However, variation in the rhizosphereactivities were relatively larger and there was a
significant effect of N source in both Petri dish andpot experiment. In the Petri dish experiment the totalacid phosphatase activity in the rhizosphere (per unitvolume of agar) was significantly higher in NH4
+
compartments than in N0 or NO3- compartments,
and was significantly higher in NO3- compartments
than in N0 compartments (Fig. 5D).
pH
4.0
4.5
5.0
5.5
6.0
6.5
7.0
N0 NH4+ NO3
-
P0 P75
a b
a
c
b
c
N0 NH4+ NO3
-
(B)
pH
4.0
4.5
5.0
5.5
6.0
6.5
7.0
7.5
NH4+ N0 NH4+ NO3- NO3- N0NH4 +/N0 NH4+/NO3- NO3-/N0
a a
b
cc
b
(A)
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
aa
b b
c c
NH4+ N0 NH4+ NO3- NO3- N0 NH4+/N0 NH4+/NO3- NO3- /N0
(D)
Pase
act
ivity
(ug
p-N
PP g
-1 so
il m
in-1
)
0
2
4
6
8
10
N0 NH4+ NO3
-
P0
a a
a
b
a
c
*
*
*
N0 NH 4+ NO3
-
(E)
Phos
phat
ase
activ
ity e
xcre
ted
by r
oots
(ug
p-N
PP c
m-1
roo
t min
-1)
Phos
phat
ase
activ
ity in
Rhi
zoph
ere
(ug
p-N
PP m
l-1 A
gar
min
-1)
0.00
0.01
0.02
0.03
0.04a
aa
a
aa
NH4+/N0 NH4+/NO3- NO 3-/N0NH4+ N0 NH4+ NO3- NO3- N0
(C)
P75
Fig. 5 Rhizosphere pH (A and B) and phosphatase activity (C,D, E) in the Petri dish experiment (A, C, D) and in the potexperiment (B, E) as affected by different N sources and byadding (P75) or not adding phytin (P0) in the pot experiment(B, E). Values are means + SE (n=3 for the Petri dishexperiment; n=4 for the pot experiment). In the Petri dish
experiment, values of pH and Pase activity with different lettersare significantly different at p<0.05. In the pot experiment,values of pH and Pase activity within the same P treatment andwith different letters are significantly different at p<0.05. Also inthe pot experiment, values in the same nitrogen treatment with anasterisk (*) differ between the two P treatments at p<0.05
20 Plant Soil (2011) 349:13–24
When phytin-P was not supplied in the pot experi-ment, acid phosphatase activity in the rhizosphere wasrelatively low and did not differ among N0, NH4
+, andNO3
- treatments (Fig. 5E); when phytin-P was sup-plied, acid phosphatase activity was significantlyhigher in the NH4
+ treatment than in NO3- and N0
treatments and was greater in the N0 treatment than inthe NO3
- treatment (Fig. 5E).
Correlations between acid phosphatase activityand rhizosphere pH and between P content and acidphosphatase activity
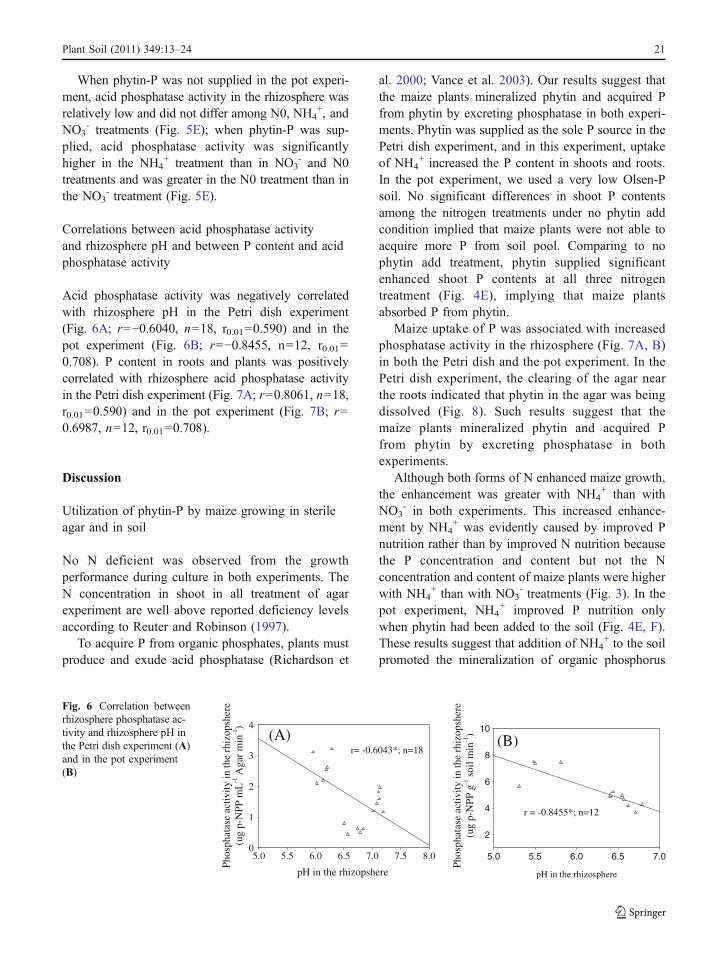
Acid phosphatase activity was negatively correlatedwith rhizosphere pH in the Petri dish experiment(Fig. 6A; r=−0.6040, n=18, r0.01=0.590) and in thepot experiment (Fig. 6B; r=−0.8455, n=12, r0.01=0.708). P content in roots and plants was positivelycorrelated with rhizosphere acid phosphatase activityin the Petri dish experiment (Fig. 7A; r=0.8061, n=18,r0.01=0.590) and in the pot experiment (Fig. 7B; r=0.6987, n=12, r0.01=0.708).
Discussion
Utilization of phytin-P by maize growing in sterileagar and in soil
No N deficient was observed from the growthperformance during culture in both experiments. TheN concentration in shoot in all treatment of agarexperiment are well above reported deficiency levelsaccording to Reuter and Robinson (1997).
To acquire P from organic phosphates, plants mustproduce and exude acid phosphatase (Richardson et
al. 2000; Vance et al. 2003). Our results suggest thatthe maize plants mineralized phytin and acquired Pfrom phytin by excreting phosphatase in both experi-ments. Phytin was supplied as the sole P source in thePetri dish experiment, and in this experiment, uptakeof NH4
+ increased the P content in shoots and roots.In the pot experiment, we used a very low Olsen-Psoil. No significant differences in shoot P contentsamong the nitrogen treatments under no phytin addcondition implied that maize plants were not able toacquire more P from soil pool. Comparing to nophytin add treatment, phytin supplied significantenhanced shoot P contents at all three nitrogentreatment (Fig. 4E), implying that maize plantsabsorbed P from phytin.
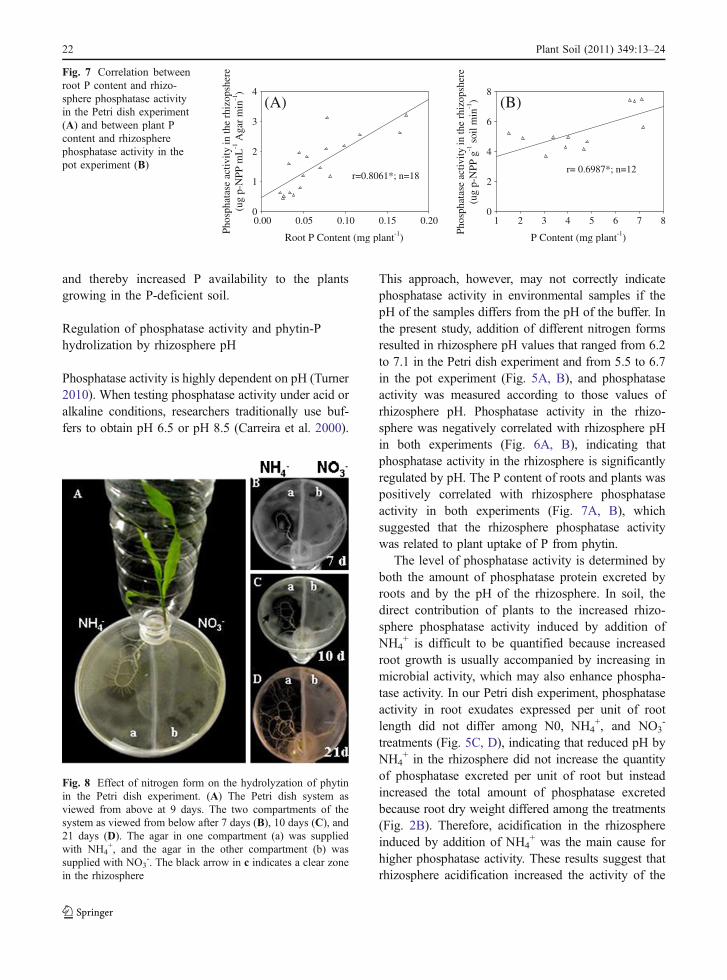
Maize uptake of P was associated with increasedphosphatase activity in the rhizosphere (Fig. 7A, B)in both the Petri dish and the pot experiment. In thePetri dish experiment, the clearing of the agar nearthe roots indicated that phytin in the agar was beingdissolved (Fig. 8). Such results suggest that themaize plants mineralized phytin and acquired Pfrom phytin by excreting phosphatase in bothexperiments.
Although both forms of N enhanced maize growth,the enhancement was greater with NH4
+ than withNO3
- in both experiments. This increased enhance-ment by NH4
+ was evidently caused by improved Pnutrition rather than by improved N nutrition becausethe P concentration and content but not the Nconcentration and content of maize plants were higherwith NH4
+ than with NO3- treatments (Fig. 3). In the
pot experiment, NH4+ improved P nutrition only
when phytin had been added to the soil (Fig. 4E, F).These results suggest that addition of NH4
+ to the soilpromoted the mineralization of organic phosphorus
pH in the rhizopshere
5.0 5.5 6.0 6.5 7.0 7.5 8.0
Phos
phat
ase
activ
ity in
the
rhiz
opsh
ere
(ug
p-N
PP m
L-1
Aga
r m
in-1
)
Phos
phat
ase
activ
ity in
the
rhiz
opsh
ere
(ug
p-N
PP g
-1 s
oil m
in-1
)
0
1
2
3
4
r= -0.6043*; n=18(A)
pH in the rhizosphere
5.0 5.5 6.0 6.5 7.0
2
4
6
8
10
r = -0.8455*; n=12
(B)
Fig. 6 Correlation betweenrhizosphere phosphatase ac-tivity and rhizosphere pH inthe Petri dish experiment (A)and in the pot experiment(B)
Plant Soil (2011) 349:13–24 21
and thereby increased P availability to the plantsgrowing in the P-deficient soil.
Regulation of phosphatase activity and phytin-Phydrolization by rhizosphere pH
Phosphatase activity is highly dependent on pH (Turner2010). When testing phosphatase activity under acid oralkaline conditions, researchers traditionally use buf-fers to obtain pH 6.5 or pH 8.5 (Carreira et al. 2000).
This approach, however, may not correctly indicatephosphatase activity in environmental samples if thepH of the samples differs from the pH of the buffer. Inthe present study, addition of different nitrogen formsresulted in rhizosphere pH values that ranged from 6.2to 7.1 in the Petri dish experiment and from 5.5 to 6.7in the pot experiment (Fig. 5A, B), and phosphataseactivity was measured according to those values ofrhizosphere pH. Phosphatase activity in the rhizo-sphere was negatively correlated with rhizosphere pHin both experiments (Fig. 6A, B), indicating thatphosphatase activity in the rhizosphere is significantlyregulated by pH. The P content of roots and plants waspositively correlated with rhizosphere phosphataseactivity in both experiments (Fig. 7A, B), whichsuggested that the rhizosphere phosphatase activitywas related to plant uptake of P from phytin.
The level of phosphatase activity is determined byboth the amount of phosphatase protein excreted byroots and by the pH of the rhizosphere. In soil, thedirect contribution of plants to the increased rhizo-sphere phosphatase activity induced by addition ofNH4
+ is difficult to be quantified because increasedroot growth is usually accompanied by increasing inmicrobial activity, which may also enhance phospha-tase activity. In our Petri dish experiment, phosphataseactivity in root exudates expressed per unit of rootlength did not differ among N0, NH4
+, and NO3-
treatments (Fig. 5C, D), indicating that reduced pH byNH4
+ in the rhizosphere did not increase the quantityof phosphatase excreted per unit of root but insteadincreased the total amount of phosphatase excretedbecause root dry weight differed among the treatments(Fig. 2B). Therefore, acidification in the rhizosphereinduced by addition of NH4
+ was the main cause forhigher phosphatase activity. These results suggest thatrhizosphere acidification increased the activity of the
Root P Content (mg plant-1) P Content (mg plant-1)
0.00 0.05 0.10 0.15 0.200
1
2
3
4
r=0.8061*; n=18
(A)
1 2 3 4 5 6 7 80
2
4
6
8
r= 0.6987*; n=12
(B)
Phos
phat
ase
activ
ity in
the
rhiz
opsh
ere
(ug
p-N
PP m
L-1
Aga
r m
in-1
)
Phos
phat
ase
activ
ity in
the
rhiz
opsh
ere
(ug
p-N
PP g
-1 s
oil m
in-1
)
Fig. 7 Correlation betweenroot P content and rhizo-sphere phosphatase activityin the Petri dish experiment(A) and between plant Pcontent and rhizospherephosphatase activity in thepot experiment (B)
Fig. 8 Effect of nitrogen form on the hydrolyzation of phytinin the Petri dish experiment. (A) The Petri dish system asviewed from above at 9 days. The two compartments of thesystem as viewed from below after 7 days (B), 10 days (C), and21 days (D). The agar in one compartment (a) was suppliedwith NH4
+, and the agar in the other compartment (b) wassupplied with NO3
-. The black arrow in c indicates a clear zonein the rhizosphere
22 Plant Soil (2011) 349:13–24

excreted phosphatase and thereafter enhanced phytin-Phydrolyzation and improved the P nutrition of maize.The results also indicate that in situ pH should beconsidered when soil phosphatase activity is measured.
Low availability of substrates to phosphatase canlimit the mineralization of organic phosphates becausemost organic phosphates are bound by cations in soil(Giaveno et al. 2010). In our Petri dish experiment, theagar medium to which phytin was added changed fromopaque to transparent when NH4
+ was supplied(Fig. 8), indicating that calcium bound with phytatewas dissolved. This was one way in which addition ofNH4
+ promoted phytin-P mineralization. The situationis true in the pot experiment because maize plant didnot acquire more P from soil pool, but acquiredsignificant more P from the added-phytin when NH4
+
was supplied (Fig 4F).
Feedback between phytin-P mineralizationand rhizosphere acidification
Rhizosphere acidification is a common strategy thatplants use to acquire P from soils (Hinsinger et al.2003; Neumann and Römheld 1999). Plants oftenacidify their rhizospheres when they are P deficient(George et al. 2002) or when they are supplied withNH4
+ fertilizers (Gahoonia et al. 1992; Nye 1981).Previous studies have mainly focused on the effect ofrhizosphere acidification on the mobilization ofinorganic phosphates (Hinsinger et al. 2003; Lamberset al. 2009; Richardson et al. 2009). In the currentstudy, levels of inorganic P were very low, and P wasadded in the organic form of phytin. The acidificationof the rhizosphere following addition of NH4
+
provided an optimal pH for acid phosphatase andmay also have increased the availability of phytate tothe enzyme by dissolving out Ca2+ and Mg2+ fromphytin. The two processes would promote phytinmineralization, which would further improve plant Pnutrition and plant growth. The resulting increase inplant growth would mean that more NH4
+ would betaken up and consequently more protons would beproduced and released into the rhizosphere, whichwould further increase phosphatase activity and accel-erate the mineralization of soil organic phosphates. Thepresent results provide support for a positive feedbackbetween organic phosphate mineralization and rhizo-sphere acidification under P-deficient conditions. Inother words, a drop in rhizosphere pH increases the
activity of phosphatase that mineralize organic P;increased P availability increases plant growth; andincreased plant growth increases the release of protonsand phosphatase into the rhizosphere, resulting infurther increases in P availability. This positivefeedback would clearly be a significant aspect ofplant–soil interactions in many soils and especially inforest soils, where a large fraction of P is in organicforms and where NH4
+ is the dominant N form.
Acknowledgements The study was supported by the NationalNatural Science Foundation of China (the major program of30890132 and the Science Fund for Creative Research Groups of30821003) and Chinese University Scientific Fund (2009TD15).
References
Allen S (1974) Chemical analysis of ecological materials.Halsted Press.
Bosse D, Köck M (1998) Influence of phosphate starvation onphosphohydrolases during development of tomato seed-lings. Plant Cell Environ 21:325–332
Bucher M (2007) Functional biology of plant phosphate uptakeat root and mycorrhiza interfaces. New Phytol 173:11–26
Carreira J, Garcia-Ruiz R, Lietor J et al (2000) Changes in soilphosphatase activity and P transformation rates induced byapplication of N-and S-containing acid-mist to a forestcanopy. Soil Biol Biochem 32:1857–1865
Duff S, Sarath G, Plaxton W (1994) The role of acidphosphatases in plant phosphorus metabolism. PhysiolPlantarum 90:791–800
Feng G, Song Y, Li X et al (2003) Contribution of arbuscularmycorrhizal fungi to utilization of organic sources ofphosphorus by red clover in a calcareous soil. Appl SoilEcol 22:139–148
Fransson A, Jones D (2007) Phosphatase activity does not limitthe microbial use of low molecular weight organic-Psubstrates in soil. Soil Biol Biochem 39:1213–1217
Gahoonia T, Claassen N, Jungk A (1992) Mobilization ofphosphate in different soils by ryegrass supplied withammonium or nitrate. Plant Soil 140:241–248
George T, Gregory P, Robinson J et al (2002) Changes inphosphorus concentrations and pH in the rhizosphere ofsome agroforestry and crop species. Plant Soil 246:65–73
Giaveno C, Celi L, Richardson AE, Simpson RJ et al (2010)Interaction of phytases with minerals and availability ofsubstrate affect the hydrolysis of inositol phosphates. SoilBiology and Biochemistry 42:491–498
Hebrien SA, Neal JL (1990) Soil pH and phosphatase activity.Commun Soil Sci Plant Anal 21:439–456
Hinsinger P, Plassard C, Tang C et al (2003) Origins of root-mediated pH changes in the rhizosphere and their responsesto environmental constraints: a review. Plant Soil 248:43–59
Koide R, Kabir Z (2000) Extraradical hyphae of the mycorrhizalfungus Glomus intraradices can hydrolyse organic phos-phate. New Phytol 148:511–517
Plant Soil (2011) 349:13–24 23

Lambers H, Chapin F, Pons T (2008) Plant physiologicalecology. Springer Verlag
Lambers H, Mougel C, Jaillard B et al (2009) Plant-microbe-soil interactions in the rhizosphere: an evolutionaryperspective. Plant Soil 321:83–115
Li X, George E, Marschner H (1991) Phosphorus depletion andpH decrease at the root-soil and hyphae-soil interfaces ofVA mycorrhizal white clover fertilized with ammonium.New Phytol 119:397–404
Liu Y, Mi G, Chen F et al (2004) Rhizosphere effect and rootgrowth of two maize (Zea mays L.) genotypes withcontrasting P efficiency at low P availability. Plant Sci167:217–223
Neumann G, Römheld V (1999) Root excretion of carboxylicacids and protons in phosphorus-deficient plants. PlantSoil 211:121–130
Nye P (1981) Changes of pH across the rhizosphere induced byroots. Plant Soil 61:7–26
Rengel Z, Marschner P (2005) Nutrient availability andmanagement in the rhizosphere: exploiting genotypicdifferences. New Phytol 168:305–312
Reuter DJ, Robinson BJ (eds) (1997) Plant analysis: Aninterpretation manual. CSIRO, Collingwood
Richardson A, Hadobas P, Hayes J (2000) Acid phosphomono-esterase and phytase activities of wheat (Triticum aestivumL.) roots and utilization of organic phosphorus substratesby seedlings grown in sterile culture. Plant Cell Environ23:397–405
Richardson A, Barea J, McNeill A et al (2009) Acquisition ofphosphorus and nitrogen in the rhizosphere and plantgrowth promotion by microorganisms. Plant Soil321:305–339
SAS Institute (1989) SAS/STAT User’s Guide. Cary. NC: SASInstitute Inc.
Saunders WMH, Williams EG (1955) Observations on thedetermination of total organic phosphorus in soil. J Soil Sci6:254–267
Shi R (1994) Determination of the total nitrogen in plantmaterials. In: Shi R, Bao S, Qin H, An Z (eds) Soilagriculture chemistry analysis method. China Agriculture,Beijing, pp 213–216
Tadano T, Sakai H (1991) Secretion of acid phosphatase by theroots of several crop species under phosphorus-deficientconditions. Soil Sci Plant Nutr 37:129–140
Tagliavini M, Masia A, Quartieri M (1995) Bulk soil pH andrhizosphere pH of peach trees in calcareous and alkaline soilsas affected by the form of nitrogen fertilizers. Plant Soil176:263–271
Tang J, Leung A, Leung C et al (2006) Hydrolysis ofprecipitated phytate by three distinct families of phytases.Soil Biol Biochem 38:1316–1324
Tarafdar JC, Claassen N (1988) Organic phosphorus compoundsas a phosphorus source for higher plants through the activityof phosphatase produced by plant roots and microorganisms.Biol Ferti Soils 5:308–312
Tarafdar JC, Marschner H (1994) Phosphatase activity in therhizosphere and hyphosphere of VA mycorrhizal wheatsupplied with inorganic and organic phosphorus. Soil BiolBiochem 26:387–395
Turner B (2008) Resource partitioning for soil phosphorus: ahypothesis. J Ecol 96:698–702
Turner B (2010) Variation in pH optima of hydrolytic enzymeactivities in tropical rain forest soils. Appl EnvironMicrobiol 76:6485–6493
Vance C, Uhde-Stone C, Allan D (2003) Phosphorus acquisitionand use: critical adaptations by plants for securing anonrenewable resource. New Phytol 157:423–447
Vincent J, Crowder M, Averill B (1992) Hydrolysis ofphosphate monoesters: a biological problem with multiplechemical solutions. Trends Biochemical Sci 17:105–110
Wang X, Tang C, Guppy CN et al (2008) Phosphorus acquisitioncharacteristics of cotton (Gossypium hisutum L.), wheat(Triticum aestivum L.) and white lupin (Lupinus albus L.)under P deficient conditions. Plant Soil 312:117–128
24 Plant Soil (2011) 349:13–24
Copyright © 2022 FDOKUMEN




















