
Potential acidification impacts on zooplankton in CCS leakage scenarios
10
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/authorsrights
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Potential acidification impacts on zooplankton in CCS leakage scenarios

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/authorsrights

Author's personal copy
Potential acidification impacts on zooplankton in CCS leakage scenarios
Claudia Halsband a,⇑, Haruko Kurihara b
a Akvaplan-niva AS, Fram Centre, N-9296 Tromsø, Norwayb Transdisciplinary Research Organization for Subtropical Island Studies, University of Ryukyus, 1 Senbaru, Nishiahara, Okinawa 903-0213, Japan
a r t i c l e i n f o
Keywords:Carbon capture and storageZooplanktonCopepodsMeroplanktonCalcificationReproduction
a b s t r a c t
Carbon capture and storage (CCS) technologies involve localized acidification of significant volumes ofseawater, inhabited mainly by planktonic species. Knowledge on potential impacts of these techniqueson the survival and physiology of zooplankton, and subsequent consequences for ecosystem health in tar-geted areas, is scarce. The recent literature has a focus on anthropogenic greenhouse gas emissions intothe atmosphere, leading to enhanced absorption of CO2 by the oceans and a lowered seawater pH, termedocean acidification. These studies explore the effects of changes in seawater chemistry, as predicted byclimate models for the end of this century, on marine biota. Early studies have used unrealistically severeCO2/pH values in this context, but are relevant for CCS leakage scenarios. Little studied meso- and bathy-pelagic species of the deep sea may be especially vulnerable, as well as vertically migrating zooplankton,which require significant residence times at great depths as part of their life cycle.
� 2013 Elsevier Ltd. All rights reserved.
1. Role of zooplankton in marine ecosystems in the light ofocean acidification and carbon capture and storage activities(CCS)
Attempts to mitigate environmental problems arising fromman-made CO2 emissions include technologies that capture CO2
before its release into the atmosphere and store it undergroundor in the oceans. The options currently discussed and implementedmay pose their own risks. CO2 sequestration in the ocean involveshazards of localized acidification in release plumes at depth or po-tential leakage from storage sites in the seabed (Hawkins, 2004),potentially impacting marine ecosystems.
Zooplankton play a major role in these systems, as they rep-resent a crucial link between primary producers and higher tro-phic levels such as fish, seabirds and marine mammals.Vertically migrating zooplankters are important contributors tothe biological pump. Through their migrations from the surfacewaters to deep ocean layers they vertically ‘pump’ carbon fromthe euphotic to the abyssal ocean, up to 1.1 mmol C m�2 d�1
(Zhang and Dam, 1997; Bradford-Grieve et al., 2001). As such,knowledge of zooplankton responses to stressors from climatechange and pollution is of utmost importance to assess thewider effects of environmental impacts on marine food webs(Hays et al., 2005). Zooplankton include a wide variety of organ-isms, both with (e.g. foraminifera, pteropods, larval stages of cal-cifying benthic animals) and without (e.g. copepods,chaetognaths, cnidarians) carbonate shells that may be impacted
by decreased pH conditions in different ways by (1) climatic in-duced ocean acidification (OA), (2) carbon capture and storage(CCS) leaks and (3) potential synergistic effects of (1) and/or(2) and additional stressors such as warming and pollution.
With their limited ability to move across water masses, zoo-plankton may be one of the most exposed and vulnerable groupsaffected by CCS, whereas phytoplankton live above the depthsaffected by CO2 plumes, and nekton will be able to avoid theplume by swimming (Herzog et al., 1996). Potential avoidancestrategies have not been studied, but it seems that exposurearound CCS release points are inevitable for organisms too smallto swim against currents or across hydrographical boundariesdue to their small Reynolds numbers (e.g. Haury and Weihs,1976). However, the magnitude and severity of the impact willdepend on the type of release technology. Changes in seawaterchemistry resulting from CO2 sequestration could exert negativeeffects on populations living in meso- and bathypelagic habitats,reproduce there, or reside at these depths during diel or ontoge-netic vertical migrations. Acidic conditions could decrease thelongevity or the reproductive output per individual, or beresponsible for increasing maturation time of populations andthus disrupt their seasonal cycles. Such impacts would decreasethe overall fitness of affected populations.
The present paper will explore (1) which CCS scenarios andtechnologies are relevant as potential stressors for marine zoo-plankton, (2) which physiological and life cycle traits are likely tobe affected, (3) review some known physiological impacts studiedin non-planktonic groups that have relevance also for zooplankton,and (4) identify the current knowledge gaps that require furtherstudy to predict the environmental impacts of CCS.
0025-326X/$ - see front matter � 2013 Elsevier Ltd. All rights reserved.http://dx.doi.org/10.1016/j.marpolbul.2013.03.013
⇑ Corresponding author. Tel.: +47 77 75 03 47.E-mail address: [email protected] (C. Halsband).
Marine Pollution Bulletin 73 (2013) 495–503
Contents lists available at SciVerse ScienceDirect
Marine Pollution Bulletin
journal homepage: www.elsevier .com/locate /marpolbul

Author's personal copy
2. CCS scenarios and technologies relevant for zooplanktonimpacts
Stephens and van der Zwaan (2005) estimate that currently25 billion tons of CO2 are released into the atmosphere each year,of which 5–10 billion tons could be stored through CCS technolo-gies. The IPCC (2005) estimates that the economic potential ofCCS could be between 10% and 55% of the total carbon mitigationeffort until the end of this century. Methods suggested includestorage of carbon dioxide in underground geological formationsincluding depleted oil and gas fields, deep aquifers, unminable coalseams and deep-sea sediments (Holloway, 2005; House et al.,2006). Alternatively, CO2 could be stored in the deep sea by inject-ing liquefied CO2 into the water column at great depths.
From the marine CO2 sequestration methods listed by Adamset al. (1997) and Davies et al. (2007), dry ice and confined orunconfined subsurface plumes of liquefied CO2 present the highestprobability of impacting zooplankton (Omori et al., 1998). Impor-tant factors are the resulting pH ranges and spatial dilution pro-cesses, exposure times and the sensitivity of the speciesconcerned. The dry ice release scenario probably invokes negligibleimpacts, while the dense plume scenario has large impacts over alimited area (Herzog et al., 1996). Scenarios that disperse the CO2
(e.g. dry ice and towed pipes) have the least impact, as near-fieldimpacts can be reduced, for example by increasing the number ofrelease points per area or choosing sites with high current speeds(Adams et al., 1997). The longer term effects will depend on thefrequency of injections in the same water body, or the life timeof such CO2 disposal sites, and if the time zooplankton populationsspend in near-field low pH conditions allows them to acclimateand eventually adapt.
3. Zooplankton affected by CCS
Zooplankton most likely affected by CCS impacts are those that(1) reside in meso- to bathypelagic habitats, such as predatorycopepods (e.g. Euchaetidae), some amphipod and chaetognathsspecies and (2) species which undertake extensive diel and/or sea-sonal vertical migrations as part of their daily foraging and ontoge-netic life cycle strategies (e.g. Calanus and Neocalanus copepodspecies and krill). Since deep-sea organisms are adapted to stableenvironmental conditions, they may be vulnerable to high CO2.On the other hand, seawater pCO2 in the deep-sea is naturallyhigher than at the surface (Feely et al., 2009), and hence deep-sea organisms have possibly adapted to high CO2 over evolutionarytime scales. Organisms living in CO2-variable environments, suchas estuaries and deep-sea hydrothermal vents, have a large capac-ity to compensate acid–base balance (Burnett, 1997; Goffredi et al.,1997). However, information on deep-sea zooplankton is scarce.Omori et al. (1998) highlight the unique environmental character-istics of deep sea habitats for plankton and micronekton. These in-clude uniform temperature and salinity, weak but consistentcurrent velocities and extremely stable physical and chemical con-ditions. The enhanced biodiversity and occurrence of rare pelagicspecies at depths around 1000 m and of bentho-pelagic speciesclose to the seafloor may include important functional groups thatare to date little studied. Due to the long-term environmental sta-bility of their habitat, genetic variance in these populations will bereduced, making them less adaptable to chemical perturbations.Consequently, meso- and bathypelagic species may be the leasttolerant to environmental change including CO2 disposal. Naturalvolcanic vents in the Mediterranean provide an example for strongCO2 gradients (Cigliano et al., 2010). Larvae of calcareous forami-niferans, serpulid polychaetes, gastropods and bivalves had signif-icantly reduced recruitment success at high concentrations of CO2
(886–5148 ppm, pH 7.08–7.79). These results show that the settle-ment of planktonic larvae of benthic organisms can be greatly re-duced in areas of increased CO2 levels.
On the other hand, species residing at greater depths may bemore resistant to high pCO2 levels in the first instance, as theycommonly experience higher pCO2 levels in their natural habitatthan surface dwellers. Particularly in subarctic regions with highprimary production and thus high decomposition rates in thewater column, copepods were less affected by high CO2 conditions(Watanabe et al., 2006). In the southern ocean, Antarctic krill (Eup-hausia superba) spawn yolk-rich eggs that sink to 700–1000 m andhatch there (Mauchline and Fisher, 1969), thus spending their earlylife in potentially CCS-affected depths. Antarctic krill embryos (E.superba) reared at 1000 latm CO2 (pH 7.7) showed no impaireddevelopment, however at 2000 latm CO2 (7.4) embryonic develop-ment was completely disrupted (Kawaguchi et al., 2010). BecauseE. superba eggs sink and spend their early life stages in deep water,the species is apparently able to cope with a certain level of CO2.However, the experiment revealed a threshold between 1000 and2000 latm CO2 beyond which the conditions will be detrimental.Thus, added stress from continuing climate-induced OA may createconditions of previously unexperienced low pCO2 and pH also fordeep-living zooplankton. Areas additionally affected by CCS leak-age would further exacerbate adverse effects of low pH on speciesresiding at depth.
Vertically migrating zooplankton include a number of eup-hausiids (Euphausia spp., Thysanoessa spp.) and large copepods(e.g. Calanus spp., Metridia spp., Eucalanus spp. and Neocalanusspp.). These species are important secondary producers and pro-vide huge amounts of biomass for higher trophic level predators.They further contribute significantly to vertical export processesin pelagic systems (Bradford-Grieve et al., 2001). Dormant stagesat depth, such as Calanus copepodites in diapause, might be resis-tant to adverse pCO2 and pH conditions when their metabolism isstrongly reduced. However, species that reproduce at depth suchas the Arctic Calanus hyperboreus (Hirche and Niehoff, 1996) or Par-euchaeta elongata (Ikeda and Hirakawa, 1996) may be vulnerable toCCS leakages in their embryonic and early life stages.
Some taxa, in contrast, may benefit from elevated CO2 condi-tions, if they can tolerate low pH and thus gain a fitness advantageover less resistant competitors. Attrill et al. (2007) related an in-crease of jellyfish in the North Sea with decreasing pH values from8.3 to 8.1 over the past decades. However, a geographically wideranalysis of long-term plankton field observations could not con-firm this conclusion (Richardson and Gibbons, 2008) and thus themechanisms that may potentially favour jellyfish over other taxaare unclear. More research is needed to assess if and which non-calcifying taxa will be winners in high CO2 scenarios.
4. Impacts of CCS on the physiology and life cycle traits ofzooplankton
Recent studies in marine systems look at OA impacts resultingfrom continued human CO2 emissions and focus on both acidificat-ion as a result of increased absorption of CO2 from the atmosphereand subsequent changes in ocean chemistry (Doney et al., 2009;Reid et al., 2009), as well as in relation to CCS scenarios. In orderto assess and predict negative effects of CCS technologies on pela-gic zooplankton, detailed information on the responses of relevantfunctional groups in the pelagic ecosystem is required. Availableliterature of acidification effects on zooplankton is still scarce,and most papers deal with epipelagic and neritic species thatmay not be directly impacted by CCS-induced CO2 pollution. How-ever, a number of experimental studies have been published overthe past decades, and most of the physiological impacts described
496 C. Halsband, H. Kurihara / Marine Pollution Bulletin 73 (2013) 495–503
Author's personal copy
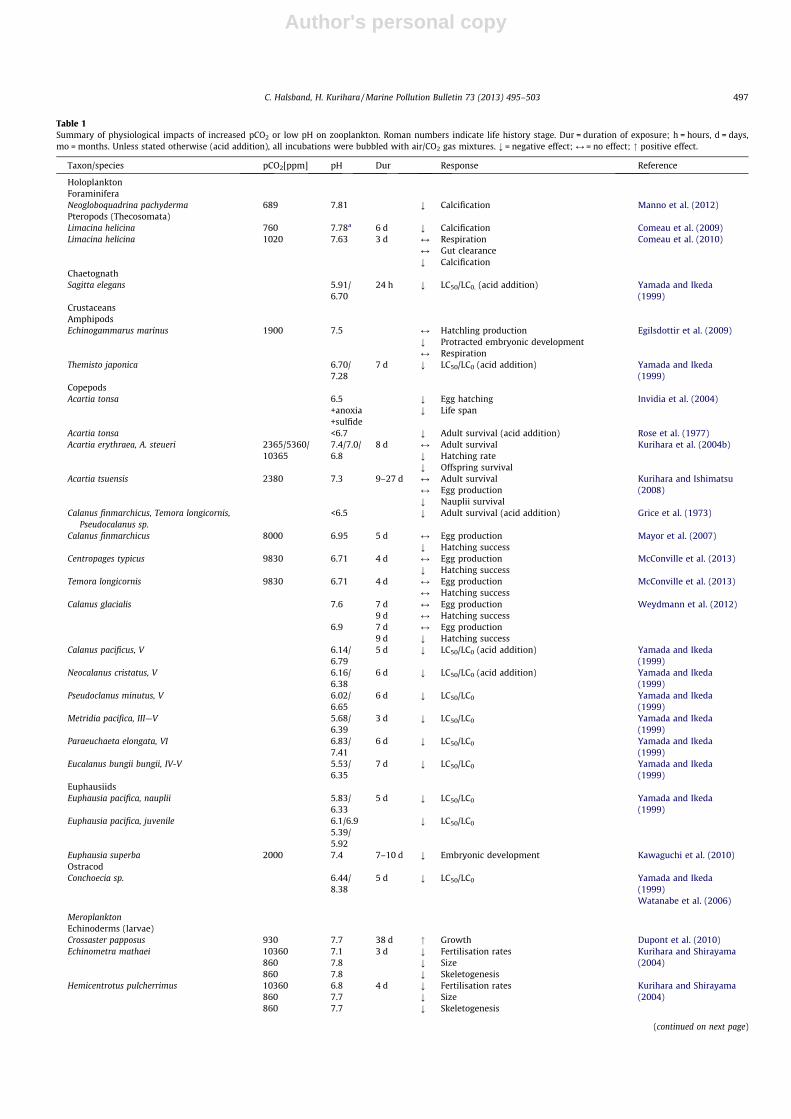
Table 1Summary of physiological impacts of increased pCO2 or low pH on zooplankton. Roman numbers indicate life history stage. Dur = duration of exposure; h = hours, d = days,mo = months. Unless stated otherwise (acid addition), all incubations were bubbled with air/CO2 gas mixtures. ; = negative effect; M = no effect; " positive effect.
Taxon/species pCO2[ppm] pH Dur Response Reference
HoloplanktonForaminiferaNeogloboquadrina pachyderma 689 7.81 ; Calcification Manno et al. (2012)Pteropods (Thecosomata)Limacina helicina 760 7.78a 6 d ; Calcification Comeau et al. (2009)Limacina helicina 1020 7.63 3 d M Respiration Comeau et al. (2010)
M Gut clearance; Calcification
ChaetognathSagitta elegans 5.91/
6.7024 h ; LC50/LC0. (acid addition) Yamada and Ikeda
(1999)CrustaceansAmphipodsEchinogammarus marinus 1900 7.5 M Hatchling production Egilsdottir et al. (2009)
; Protracted embryonic developmentM Respiration
Themisto japonica 6.70/7.28
7 d ; LC50/LC0 (acid addition) Yamada and Ikeda(1999)
CopepodsAcartia tonsa 6.5 ; Egg hatching Invidia et al. (2004)
+anoxia ; Life span+sulfide
Acartia tonsa <6.7 ; Adult survival (acid addition) Rose et al. (1977)Acartia erythraea, A. steueri 2365/5360/
103657.4/7.0/6.8
8 d M Adult survival Kurihara et al. (2004b); Hatching rate; Offspring survival
Acartia tsuensis 2380 7.3 9–27 d M Adult survival Kurihara and Ishimatsu(2008)M Egg production
; Nauplii survivalCalanus finmarchicus, Temora longicornis,
Pseudocalanus sp.<6.5 ; Adult survival (acid addition) Grice et al. (1973)
Calanus finmarchicus 8000 6.95 5 d M Egg production Mayor et al. (2007); Hatching success
Centropages typicus 9830 6.71 4 d M Egg production McConville et al. (2013); Hatching success
Temora longicornis 9830 6.71 4 d M Egg production McConville et al. (2013)M Hatching success
Calanus glacialis 7.6 7 d M Egg production Weydmann et al. (2012)9 d M Hatching success
6.9 7 d M Egg production9 d ; Hatching success
Calanus pacificus, V 6.14/6.79
5 d ; LC50/LC0 (acid addition) Yamada and Ikeda(1999)
Neocalanus cristatus, V 6.16/6.38
6 d ; LC50/LC0 (acid addition) Yamada and Ikeda(1999)
Pseudoclanus minutus, V 6.02/6.65
6 d ; LC50/LC0 Yamada and Ikeda(1999)
Metridia pacifica, III—V 5.68/6.39
3 d ; LC50/LC0 Yamada and Ikeda(1999)
Paraeuchaeta elongata, VI 6.83/7.41
6 d ; LC50/LC0 Yamada and Ikeda(1999)
Eucalanus bungii bungii, IV-V 5.53/6.35
7 d ; LC50/LC0 Yamada and Ikeda(1999)
EuphausiidsEuphausia pacifica, nauplii 5.83/
6.335 d ; LC50/LC0 Yamada and Ikeda
(1999)Euphausia pacifica, juvenile 6.1/6.9 ; LC50/LC0
5.39/5.92
Euphausia superba 2000 7.4 7–10 d ; Embryonic development Kawaguchi et al. (2010)OstracodConchoecia sp. 6.44/
8.385 d ; LC50/LC0 Yamada and Ikeda
(1999)Watanabe et al. (2006)
MeroplanktonEchinoderms (larvae)Crossaster papposus 930 7.7 38 d " Growth Dupont et al. (2010)Echinometra mathaei 10360 7.1 3 d ; Fertilisation rates Kurihara and Shirayama
(2004)860 7.8 ; Size860 7.8 ; Skeletogenesis
Hemicentrotus pulcherrimus 10360 6.8 4 d ; Fertilisation rates Kurihara and Shirayama(2004)860 7.7 ; Size
860 7.7 ; Skeletogenesis
(continued on next page)
C. Halsband, H. Kurihara / Marine Pollution Bulletin 73 (2013) 495–503 497
Author's personal copy
for a variety of taxa will be applicable for zooplankton populationssubject to CCS leaks.
In this chapter we review available information on the physiol-ogy and life cycles of various groups of zooplankton: experimentalstudies focus on single-celled calcite secreting foraminiferans andpteropods (Thecosomata) with aragonitic shells, common copepodspecies and a number of meroplanktonic larvae of benthic organ-isms have been studied (Table 1). We also include studies onnon-planktonic aquatic crustaceans which address processes ortraits potentially relevant for zooplankton.
4.1. Impacts of anthropogenic acid waste
Early work on pH effects focused on the fate of industrial acidwastes in freshwater and marine coastal systems in the 1970s.These investigated lethal effects of extremely low pHs on aquaticorganisms ranging from marine copepods (Grice et al., 1973) tofreshwater gastrotrichs (Faucon and Hummon, 1976), crustaceansand insect larvae (Havas and Hutchinson, 1982), often studied incombination with other toxic stressors such as metals. Short-termincubations over 24–48 h demonstrated that the survival of adultcalanoid copepods of the species Calanus finmarchicus, Temora lon-gicornis and Pseudocalanus sp. is impaired at a pH < 6.5 (Grice et al.,1973). Rose et al. (1977) found a similar increase in mortalityin Acartia tonsa at pH < 6.7. Later, acidification impacts from
anthropogenic sources were mainly a concern for the biodiversityof little buffered freshwater lakes, where acidic conditions elimi-nated sensitive zooplankton species through increased death ratesand delays in reproductive maturity (Walton, 1982; Havens et al.,1993).
4.2. Impacts of CCS approaches
Studies aimed specifically to evaluate the impact of CCS on mar-ine zooplankton have been conducted since the late 1990s. Yamadaand Ikeda (1999) studied ten oceanic zooplankton species in CCSrelevant pH ranges down to pH 4. They tested different functionalgroups including filter feeding herbivores and planktivorous pre-dators: one ostracod, six calanoid copepods, naupliar and juvenilestages of the euphausid Euphausia pacifica and the chaetognathpredator Sagitta elegans. Mortality increased with exposure timeand more rapidly so than for freshwater zooplankton at similarpHs. The sensitivity was species-specific, but no correlations withswimming or feeding bevaviour were detected. The mesopelagiccopepod P. elongata was the most sensitive species, with the high-est LC50 and LC0 values, and may thus be vulnerable to CCS leakageevents.
Ishimaru (2001) determined the 24 h LC50 values of four cope-pod species: Scottocalanus helenae (pH 6.00), Mesocalanus tennui-cornis (pH 6.32), Calanus pacificus (pH 6.58) and Neocalanus
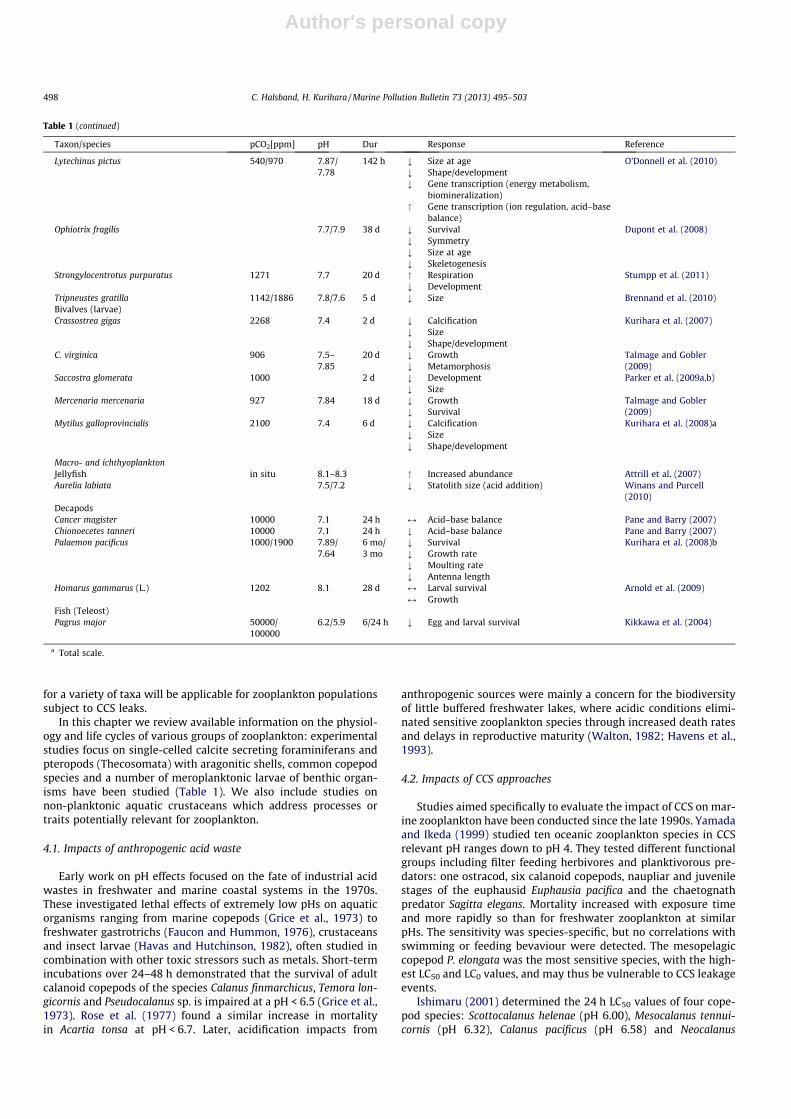
Table 1 (continued)
Taxon/species pCO2[ppm] pH Dur Response Reference
Lytechinus pictus 540/970 7.87/7.78
142 h ; Size at age O’Donnell et al. (2010); Shape/development; Gene transcription (energy metabolism,
biomineralization)" Gene transcription (ion regulation, acid–base
balance)Ophiotrix fragilis 7.7/7.9 38 d ; Survival Dupont et al. (2008)
; Symmetry; Size at age; Skeletogenesis
Strongylocentrotus purpuratus 1271 7.7 20 d " Respiration Stumpp et al. (2011); Development
Tripneustes gratilla 1142/1886 7.8/7.6 5 d ; Size Brennand et al. (2010)Bivalves (larvae)Crassostrea gigas 2268 7.4 2 d ; Calcification Kurihara et al. (2007)
; Size; Shape/development
C. virginica 906 7.5–7.85
20 d ; Growth Talmage and Gobler(2009); Metamorphosis
Saccostra glomerata 1000 2 d ; Development Parker et al. (2009a,b); Size
Mercenaria mercenaria 927 7.84 18 d ; Growth Talmage and Gobler(2009); Survival
Mytilus galloprovincialis 2100 7.4 6 d ; Calcification Kurihara et al. (2008)a; Size; Shape/development
Macro- and ichthyoplanktonJellyfish in situ 8.1–8.3 " Increased abundance Attrill et al. (2007)Aurelia labiata 7.5/7.2 ; Statolith size (acid addition) Winans and Purcell
(2010)DecapodsCancer magister 10000 7.1 24 h M Acid–base balance Pane and Barry (2007)Chionoecetes tanneri 10000 7.1 24 h ; Acid–base balance Pane and Barry (2007)Palaemon pacificus 1000/1900 7.89/
7.646 mo/3 mo
; Survival Kurihara et al. (2008)b; Growth rate; Moulting rate; Antenna length
Homarus gammarus (L.) 1202 8.1 28 d M Larval survival Arnold et al. (2009)M Growth
Fish (Teleost)Pagrus major 50000/
1000006.2/5.9 6/24 h ; Egg and larval survival Kikkawa et al. (2004)
a Total scale.
498 C. Halsband, H. Kurihara / Marine Pollution Bulletin 73 (2013) 495–503

Author's personal copy
cristatus (pH 5.39). The LC50 values increased with exposure time.Consequently, most planktonic copepods will be lethally impactedby leaking CO2 when the pH becomes lower than 6.0.
4.3. Impacts equivalent to CCS long-range and long-term effects
Several studies have evaluated lethal and sub-lethal effect of OAon zooplankton including foraminiferans, pteropods, copepods,shrimps, krill, jellyfish and larval stages of calcifiers in a pH rangeof 8.2–7.6; which are generally higher pHs than those expectedimmediately following CCS leaks. However, these effects are rele-vant in as far as they represent more long-term effects of regularCCS CO2 discharges and possible consequences for zooplanktonin such affected water masses. The data generally suggest that eventhough OA may not have acute lethal effects on zooplankton, sub-lethal effects, including reductions in growth, calcification andmetabolism, and reproductive success are predicted (Kroekeret al., 2010). However, meta-analyses reveal high variability inthe tolerance of different species, where calcification is the mostvulnerable biological process, and crustaceans seem relativelyresistant to high CO2 (Hendriks et al., 2010; Kroeker et al., 2010).
5. Studies on individual taxa and functional groups
5.1. Calcifying zooplankton
The most important calcifying marine zooplankton taxa areforaminifera and pteropods. Negative effects of increased CO2 onshell formation and calcification rates of planktonic clacifiers havebeen reviewed by Fabry et al. (2008 and references therein). Theydescribe the close relationship between aragonite saturation stateof seawater and calcification rates, but point out that only very fewspecies have been studied, while species-specific differences mayexist. They hypothesize that the photosynthetic activity of endo-symboints creates a low pCO2 micro-environment around the shellfavouring shell maintenance and growth. These laboratory studieswere followed by comparisons of present-day foraminifera shellweights with those from deeper sediments, dating back to theHolocene. Recently deposited foraminifera shells were 30–35%lighter than their counterparts from the geological past (Moyet al., 2009), although seasonal patterns such as upwelling alsoplay a role (de Moel et al., 2009). A new study on the Arctic speciesNeogloboquadrina pachyderma highlights the synergistic effects ofdecreasing pH with increasing temperature (Manno et al., 2012):within the optimum temperature range of Neogloboquadrina, high-er temperatures offset a reduction in net calcification. However,Uthicke and Fabricius (2012) show that a profit in endosymbionticphotosynthesis may not compensate for other metabolic costs,such as higher respiration rates, in foraminiferans.
Pteropods are important zooplankton in Polar regions, some ofwhich have calcareous shells and are as such particularly at risk tosuffer from OA effects. The combination of CaCO3 undersaturationand exposure time to undersaturated waters will determine howseverely these animals will be affected (Fabry et al., 2008). A strongemphasis has originally been placed on pteropods in Arctic andAntarctic systems, while less is known about calcareous hetero-pods in the tropics, which possess aragonite shells as veliger larvaeor adults. Clio pyramidata from the subarctic Pacific showedmarked dissolution rates of their shell within 48 h in aragoniteundersaturated water as predicted for 2100 (Orr et al., 2005). Sim-ilar results were found for the Arctic species Limacina helicina(Comeau et al., 2009), where calcification rate decreased by 28%decrease in similar conditions. A decrease in calcium carbonateprecipitation was highly correlated to the aragonite saturationstate and exacerbated by elevated temperature (Comeau et al.,
2010). The relative contribution of pteropods to zooplankton inSouthern ocean is estimated to be around 10%, but sometimescan exceed 85%. Pteropods also perform diel vertical migrationsover 300 m depth. Taking into account the ecological contributionas a food source of higher trophic species and geochemical cyclingand carbon cycle, the impact of CCS on pteropods should beaddressed.
5.2. Epipelagic and neritic species
Some epipelagic and neritic copepod species have been studiedunder high CO2 scenarios equivalent to CCS leak events. Acartiasteueri females were able to survive pH 6.8 over 8 days, but eggproduction rates of both A. steueri and Acartia erythraea decreasedsignificantly (Kurihara et al., 2004a,b). Two subsequent genera-tions of the copepod Acartia tsuensis reared at 2380 ppm CO2 (pH7.3) showed no difference in survival, body size or developmentrates compared with the control at 380 ppm (Kurihara and Ishima-tsu, 2008). Invidia et al. (2004) compared the impact of near anox-ia, combined with high concentrations of sulfide, at pH 8.2 and 6.5and found reduced egg viability and life expectancy after exposuretimes of >15 days. Mayor et al. (2007) studied the effects of oceanicCO2 disposal scenarios on C. finmarchicus hatching success andfound greatly reduced hatching rates after 72 h of incubation atpH 6.95. They concluded that acidification impact studies mustidentify the most sensitive life phases rather than considering onlyadult individuals of a given species. McConville et al. (2013) de-tected significant reductions in reproductive rates of adult femalecopepods and hatching success of their eggs during 4 days of expo-sure to pH 6.8 for Centropages typicus and T. longicornis. Less severeacidification (pH 8.04–7.78), in contrast, showed no effect on anyof the parameters measured, indicating a certain resilience downto a pH of 6.8. The study, however, also showed that the seasonaltiming of experiments was an important factor for the severity ofthe OA impact. This was also pointed out by Mayor et al. (2012),where the temperature effects on hatching success of C. finmarchi-cus were context-dependent and differed between independentbatches of eggs with different maternal history. Thus, long-term ef-fects of acidification on copepod populations could be negative(Fitzer et al., 2012) and adaptation mechanisms need to be studiedover multiple generations.
Differential sensitivity of zooplankton life stages is an impor-tant factor, as it ultimately determines the tolerance of a species,and generally early life stages are predicted to be more vulnerablethan their adult counterparts (Kurihara, 2008). Sensitive stagesrepresent bottlenecks of the population; selection of OA conditionswill primarily act on them. Survival rates of the nauplii stages of A.erythraea were affected at a pH level of 7.0 (Kurihara et al., 2004a).The same conditions had no adverse effect on the survival of adultfemales, but egg production of both A. erythraea and A. steueri wassignificantly suppressed when females were reared at pH 7.0 and6.8 for 6 days, respectively (Kurihara, 2004b). Similarly, neithersurvival nor growth rate, egg production or hatching rate of A. tsu-ensis were affected when reared for two generations under pH 7.4(Kurihara and Ishimatsu, 2008).
5.3. Meroplankton
Impacts on meroplankton, the larval stages of marine benthiccalcifiers, including sea urchins, brittle stars, sea stars and bivalves,are now well documented. However, very few studies exist ondeep sea species. In the deep sea, echinoderms are the most impor-tant epifauna, and in some cases, brittle stars make up 63% of themegafauna (Gage and Tyler, 1991). Although information abouttheir early development is still very scarce, deep-sea species arepredicted to have relatively low fecundity and produce few large
C. Halsband, H. Kurihara / Marine Pollution Bulletin 73 (2013) 495–503 499

Author's personal copy
eggs compared to shallow living species (Tyler, 1986). Any impactsof CCS on the early developmental stages of benthic organismswith such slow reproductive strategies will be detrimental to theirpopulation dynamics.
The larval stages of shallow living calcifiers are very sensitive tohigh CO2. Fertilization rates of sea urchin eggs of the species Hemi-centrotus pulcherrimus and Echinometra mathaei, reared for 15 minunder high CO2, decreased with increasing CO2 concentration, andthe fertilization rate was reduced to 60% and 40% at pH 7.0–7.2,respectively (Kurihara and Shirayama, 2004). The size at age of plu-teus larvae linearly decreased with CO2 rise, and larvae reared un-der pH 6.8 completely lacked skeletogenesis. OA conditions at 0.2pH units below normal caused 100% larval mortality within 8 daysin the brittlestar Ophiotrix fragilis, as well as abnormal develop-ment and decrease in larval body size (Dupont et al., 2008). En-hanced CO2 (pH 7.87–7.78) had significant effects on the shapeand size of echinoplutei larvae, while genes, central to energymetabolism and biomineralization, were down-regulated (O’Don-nell et al., 2010). However, larvae of some species such as larvaeof the sea star Crossaster papposus seemed to benefit from highCO2 conditions (pH 7.7) and responded with increased growthrates to OA conditions (Dupont et al., 2010). Larval stages of marinebivalves such as oysters and mussels are also affected by high CO2
conditions (Kurihara et al., 2007, 2008a; Parker et al., 2009a,b,2010). As already stated for holoplankton, the differential sensitiv-ity of life stages within species is equally important in meroplank-ton such as echinoderm larvae (Dupont et al., 2010). While benthicorganisms may appear to be resistant to ocean acidification asadults (e.g. Wood et al., 2008) and juveniles (e.g. Gooding et al.,2009), some are extremely sensitive as meroplanktonic larvae (Du-pont et al., 2008). The gametes, embryos and meroplanktonic lar-vae of benthic animals may thus be population bottlenecksduring low pH events.
5.4. Macro- and ichthyoplankton
The sensitivity and mortality of macro- and ichthyoplanktonhave also been studied: pH strongly influenced egg weight, but alsoaltered the metal uptake through permeability changes of the eggshell in embryonic cuttlefish Sepia officinale (Lacoue-Labarthe et al.,2009) and in eggs and hatchlings of the squid Loligo vulgaris (La-coue-Labarthe et al., 2011). A long-term study on the marineshrimp Palaemon pacificus over 3 and 6 months revealed that med-ium–high CO2 concentrations (1000–2000 latm CO2, pH 7.72–7.40) reduced their survival, growth rate, moulting rate and anten-na length (Kurihara et al., 2008b). Additionally, the settling size ofjuvenile P. pacificus cultured under 2000 latm CO2 was signifi-cantly smaller compared to the control (Kurihara, 2008). Pro-nounced impacts on early developmental stages are also reportedin marine fishes. Both hatching and survival of larvae of the silverseabream, Pagrus major, declined with sea water pCO2 and themedian lethal CO2 concentration was found to change throughtheir ontogeny. The medium lethal pCO2 was lowest at egg stageand peaked in the preflexion stage (Kikkawa et al., 2004). Becausethe chloride cells of marine teleosts, which are involved in the pHregulation, develop during the embryonic phase (Kaneko et al.,1995; Shiraishi et al., 1997), eggs seem to have less capacity tocompensate a pH decline (Ishimatsu et al., 2004).
5.5. Relevant physiological impacts on non-planktonic groups
Other physiological impacts relevant for zooplankton werefound in benthic groups, such as reduced locomotion and possibledisruption to chemosensory function in hermit crabs (de la Hayeet al., 2011) and other physiological functions reviewed in Widdi-combe and Spicer (2008). Melzner et al. (2009) reviewed the mech-
anisms underlying the differences in CO2 sensitivity betweenorganisms, and pointed out that high metabolic organisms suchas cephalopods and brachyuran crustaceans are more tolerant tohigh CO2 than low metabolic organisms such as echinoderms andbivalves due to their efficient ion-transport systems. Indeed, manyaquatic crustaceans have a higher capacity for extracellular acid–base regulation than other marine invertebrates such as echino-derms and molluscs (Wheatly and Henry, 1992). While the seaurchin Psammechinus miliaris lacks acid–base compensation capac-ity when exposed to pH 7.44 (Miles et al., 2007), the prawns Palae-mon elegans and P. serratus exposed to 3000 latm CO2 completelycompensate their haemolymph pH (Dissanayake et al., 2010). Sim-ilar results were also observed in crabs Necora puber and Canermagister (Pane and Barry, 2007; Spicer et al., 2007). However, thecapacity of extracellular acid–base regulation appears to be spe-cies-specific even within marine crustaceans and their life-stylemay be related. For example, in contrast with shallow-living crabs,the high-CO2 induced acidosis in the haemolymph of the deep-seaTanner crab Chionecetes tanneri was almost uncompensated at10,000 latm CO2 (pH 7.1, Pane and Barry, 2007). In their reviewSeibel and Walsh (2003) suggest that deep-sea fishes, cephalopodsand crustaceans have less pronounced acid–base regulation capac-ity than shallow loving organisms, because they exhibit reducedmetabolic rates.
6. Predicting impacts of CCS on plankton communities
A modelling study quantified the mortality of zooplankton thatpasses through a CCS plume, integrating mortality data with adescription of plume physics (Caulfield et al., 1997). The model de-scribes both the total mortality and the ‘spatial deficit’ of organ-isms (proportion of population lost) for droplet and dense CO2
plumes. High mortalities were caused by pHs lower than 6.3 overlong time scales. To predict effects at ecosystem level, accurateestimates of the organisms’ dose responses are important. How-ever, these are species-specific and thus require sound knowledgeof the ecology, biology and physiological responses to low pH con-ditions of the species concerned. To assess the physiological im-pacts of changes in pH and CO2 chemistry in a comprehensiveway across taxa and functional groups in pelagic systems is diffi-cult, as experimental studies are labour- and infrastructure in-tense, and depend on ‘best guesses’ as to which aspects of aspecies’ physiology – other than calcification – might be affected.So far two meta-analyses have made the attempt of summarisingthese effects (Hendriks et al., 2010, Kroeker, 2010), highlightingthe tremendous variability in experimental results and taxon-dependent differences. A molecular approach used transcriptomicsto examine the variety of OA effects on different aspects of thephysiology in a single species. Sea urchin larvae (Strongylocentrotuspurpuratus) subjected to moderate and high CO2 conditions down-regulated their gene expression in three fundamental processes inaddition to calcification: cellular stress response, metabolism andapoptosis (Todgham and Hofmann, 2009). Such microarray tech-niques will be useful in targeting a variety of different genes andphysiological processes, enabling the detection of unexpected re-sponses. A first study of zooplankton community effects was con-ducted in mesocosms in an Arctic fjord (Niehoff et al., 2012), butthe mechanisms leading to differential changes of developmentalstage ratios under increased pCO2 in some taxa, but not others, re-mained obscure.
Once a clearer picture of the sensitivity or resilience of variousorganisms and functional groups emerges from experiments andmolecular approaches, scaling up from individuals to populationsand communities is required through adequate parameterizationof ecosystem models (Raven et al., 2005; Blackford, 2010).
500 C. Halsband, H. Kurihara / Marine Pollution Bulletin 73 (2013) 495–503

Author's personal copy
7. Knowledge gaps and further research
Although zooplankton are a crucial link between trophic levelsin marine ecosystems, it is a group that has been relatively poorlystudied with respect to OA and CCS impacts. Riebesell & Tortell(2011) attribute this lack of information in part to the greater com-plexity involved in experimental work with these metazoans, ascompared to e.g. single-celled phytoplankton and foraminiferans.Zooplankton have complex life cycles and comparably long gener-ation times, especially at high latitudes. Consequently, realistic ef-fects at population level are difficult to determine and requirelong-term, preferably multi-generation incubations to detect selec-tion pressure of OA on life cycle traits and potential adaptation.
To aid sustainable use of new technology such as CCS, we listthe most urgent requirements for further research:
1. As a pre-requisite to CCS risk assessments, more knowledge isrequired about the basic ecology and life strategies of zooplank-ton, especially for deep sea species and under-studied taxa (i.e.non-copepods), to identify CCS-vulnerable species and lifestages. Considerable progress has been made to determinephysiological impacts at the individual level and more studiesare ongoing to increase our knowledge on the OA responses ofthe varied functional groups among the zooplankton. However,the interactive effects of climate-change stresses, primarily ele-vated CO2 and temperature, have been poorly studied so far,and methodological standards are needed to conduct multiplestressor experiments to identify ontogenetic sensitivities(Byrne, 2012). Davies et al. (2007) raise awareness for the deepsea and urge for the development of regulatory controls toavoid impacts before they occur. From the literature, it isunclear if deep sea zooplankton is more or less sensitive tolow pH, than the better studied surface dwellers. Adams et al.(1997) use the assumption of higher sensitivity in their model,supported by the results of Yamada and Ikeda (1999) on amesopelagic copepod, but on the other hand Watanabe et al.(2006) report enhanced resilience in deep sea zooplankton spe-cies regularly exposed to high CO2 waters. At these depths,where pH is already at 0.5 units less than surface waters, fur-ther decreases in pH may cross a threshold which causes prob-lems for diapausing calanoid copepods. Their overwinteringcopepodite stages (usually CV) may have specific limits withrespect to pCO2 and pH to successfully moult, develop andascent. Reduced survival or delayed development of CVs wouldaffect the recruitment of reproductive adults. More research isneeded on the specific effects of CO2 disposal in deep sea habi-tats, with its unique species assemblage and great hydrostaticpressures. Laboratory results from experiments conducted atsurface pressure may not be transferable to these conditionsand species (Omori et al., 1998).
2. To determine which locations are best suitable for CCS, regionaland latitudinal differences need to be taken into account inorder to minimise impacts on ecosystem function and commu-nity resilience. Information on the ecology and vulnerability oflocal and regional fauna need to be combined with factors suchas latitude, depth, prevailing currents and advection or upwell-ing. This includes possible synergistic impacts of multiplestressors, e.g. pH/alkalinity and elevated temperatures, or pollu-tants and stressors other than CO2. To tackle the complexity ofCO2 effects and responses, new model systems need to bedeveloped (Blackford, 2010).
3. Behavioural responses to CCS conditions need to be studied.They may include impaired mate finding (pheromone recep-tion) and mating behaviour, escape behaviour from predatorsand feeding behaviour (food detection, impact on ‘taste’, i.e.
chemical reception). Different reproductive strategies may alsodetermine the vulnerability of organisms, e.g. differential spermviability in species with external (e.g. sea urchins, fishes) andinternal (spermatophores of copepods) fertilisation.
4. Potential shifts in food web structure need to consider also theresilient species, for which CCS plumes may serve as sanctuariesfrom competition and predation, similar to hypoxic zones (Van-derploeg et al., 2009). Some species seem to emerge as potentialwinners from acidified conditions, including jellyfish, copepodsand sea star larvae (Attrill et al., 2007; Dupont et al., 2010,McConville et al., 2013). Depending on their role within theirrespective food webs, these taxa may exert higher predationcontrol, new indirect pressures on their more vulnerable com-petitors and explore new ecological niches previously occupiedby taxa now struggling to compensate for CCS effects.
5. Finally, where zooplankton are predicted to be significantlyaffected by CCS pollution, the consequences for organismshigher up the food chain need to be addressed, including com-mercial species such as fish, shellfish and crustaceans. A socio-economic assessment of CCS needs to be conducted in the con-text of ecosystem impacts.
Acknowledgements
The authors were supported by the Norwegian Fram CentreFlagship for Ocean Acidification (CH) and by the Japan Scienceand Technology (JST), Rising Star Program (HK).
References
Adams, E.E., Caulfield, J.A., Herzog, H.J., Auerbach, D.I., 1997. Impacts of reduced pHfrom ocean CO2 disposal: sensitivity of zooplankton mortality to modelparameters. Waste Manage. 17, 375–380.
Arnold, K.E., Findlay, H.S., Spicer, J.I., Daniels, C.L., Boothroyd, D., 2009. Effect of CO2-related acidification on aspects of the larval development of the Europeanlobster, Homarus gammarus (L.). Biogeosciences 6 (8), 1747–1754.
Attrill, M.J., Wright, J., Edwards, M., 2007. Climate-related increases in jellyfishfrequency suggest a more gelatinous future for the North Sea. Limnol. Oceanogr.52, 480–485.
Blackford, J.C., 2010. Predicting the impacts of ocean acidification: challenges froman ecosystem perspective. J. Mar. Sys. 81, 12–18.
Bradford-Grieve, J.M., Nodder, S.D., Jillett, J.B., Currie, K., Lassey, K.R., 2001. Potentialcontribution that the copepod Neocalanus tonsus makes to downward carbonflux in the Southern Ocean. J. Plankton Res. 23, 963–975.
Brennand, H.S., Soars, N., Dworjanyn, S.A., Davis, A.R., Byrne, M., 2010. Impact ofocean warming and ocean acidification on larval development and calcificationin the sea urchin Tripneustes gratilla. PLoS One 5 (6), e11372.
Burnett, L.E., 1997. The challenges of living in hypoxic and hypercapnic aquaticenvironments. Am. Zool. 37, 633–640.
Byrne, M., 2012. Global change ecotoxicology: Identification of early life historybottlenecks in marine invertebrates, variable species responses and variableexperimental approaches. Mar. Environ. Res. 76, 3–15.
Caulfield, J.A., Auerbach, D.I., Adams, E.E., Herzog, H.J., 1997. Near field impacts ofreduced pH from ocean CO2 disposal. Energy Conv. Manage. 38 (Supplement),S343–S348.
Cigliano, M., Gambi, M.C., Rodolfo-Metalpa, R., Patti, F.P., Hall-Spencer, J.M., 2010.Effects of ocean acidification on invertebrate settlement at volcanic CO2 vents.Mar. Biol. 157, 2489–2502.
Comeau, S., Gorsky, G., Jeffree, R., Teyssié, J.L., Gattuso, J.P., 2009. Impact of oceanacidification on a key Arctic pelagic mollusc (Limacina helicina). Biogeosciences6, 1877–1882.
Comeau, S., Jeffree, R., Teyssié, J.L., Gattuso, J.P., 2010. Response of the Arcticpteropod Limacina helicina to projected future environmental conditions. PLoSOne 5, e11362.
Davies, A.J., Roberts, J.M., Hall-Spencer, J., 2007. Preserving deep-sea naturalheritage: emerging issues in offshore conservation and management. Biol.Conserv. 138, 299–312.
de la Haye, K.L., Spicer, J.I., Widdicombe, S., Briffa, M., 2011. Reduced sea water pHdisrupts resource assessment and decision making in the hermit crab Pagurusbernhardus. Anim. Behav. 82, 495–501.
de Moel, H., Ganssen, G.M., Peeters, F.J.C., Jung, S.J.A., Brummer, G.J.A., Kroon, D.,Zeebe, R.E., 2009. Planktic foraminiferal shell thinning in the Arabian Sea due toanthropogenic ocean acidification? Biogeosci. Disc. 6, 1811–1835.
C. Halsband, H. Kurihara / Marine Pollution Bulletin 73 (2013) 495–503 501

Author's personal copy
Dissanayake, A., Clough, R., Spicer, J.I., Jones, M.B., 2010. Effects of hypercapnia onacid–base balance and osmo-/ion-regulation in prawns (Decapoda:Palaemonidae). Aquatic Biol. 11, 27–36.
Doney, S.C., Fabry, V.J., Feely, R.A., Kleypas, J.A., 2009. Ocean acidification: the otherCO2 problem. Annu. Rev. Mar. Sci. 1, 169–192.
Dupont, S., Havenhand, J., Thorndyke, W., Peck, L., Thorndyke, M., 2008. Near-futurelevel of CO2-driven ocean acidification radically affects larval survival anddevelopment in the brittlestar Ophiothrix fragilis. Mar. Ecol. Prog. Ser. 373, 285–294.
Dupont, S., Lundve, B., Thorndyke, M., 2010. Near future ocean acidificationincreases growth rate of the lecithotrophic larvae and juvenile of the seastarCrossaster papposus. J. Exp. Zool. 314B, 382–389.
Egilsdottir, H., Spicer, J.I., Rundle, S.D., 2009. The effect of CO2 acidified sea waterand reduced salinity on aspects of the embryonic development of the amphipodEchinogammarus marinus (Leach). Mar. Poll. Bull. 58, 1187–1191.
Fabry, V.J., Seibel, B.A., Feely, R.A., Orr, J.C., 2008. Impacts of ocean acidification onmarine fauna and ecosystem processes. ICES J. Mar. Sci.: J. du Conseil 65, 414–432.
Faucon, A.S., Hummon, W.D., 1976. Effects of mine acid on the longevity andreproductive rate of the gastrotricha Lepidodermella squammata (Dujardin).Hydrobiologia 50, 265–269.
Feely, R.A., Doney, S.C., Cooly, S.R., 2009. Present conditions and future changes in ahigh-CO2 world. Oceanography 22, 36–47.
Fitzer, S.C., Caldwell, G.S., Close, A.J., Clare, A.S., Upstill-Goddard, R.C., Bentley, M.G.,2012. Ocean acidification induces multi-generational decline in copepodnaupliar production with possible conflict for reproductive resourceallocation. J. Exp. Mar. Biol. Ecol. 418, 30–36.
Gage, J.D., Tyler, P.A., 1991. Deep-sea biology: a natural history of organisms of thedeep-sea floor. Cambridge Uni. Press, Cambridge.
Goffredi, S., Childress, J., Desaulniers, N., Lee, R., Lallier, F., Hammond, D., 1997.Inorganic carbon acquisition by the hydrothermal vent tubeworm Riftiapachyptila depends upon high external pCO2 and upon proton-equivalent iontransport by the worm. J. Exp. Biol. 200, 883–896.
Gooding, R.A., Christopher, D.G., Harley, D.G., Tang, E., 2009. Elevated watertemperature and carbon dioxide concentration increase the growth of akeystone echinoderm. PNAS 106, 9316–9321.
Grice, G.D., Wiebe, P.H., Hoagland, E., 1973. Acid-iron waste as a factor affecting thedistribution and abundance of zooplankton in the New York Bight: I. Laboratorystudies on the effects of acid waste on copepods. Estuarine Coastal Mar. Sci. 1,45–50.
Haury, L., Weihs, D., 1976. Energetically efficient swimming behavior of negativelybuoyant zooplankton. Limnol. Oceanogr. 6, 797–803.
Havas, M., Hutchinson, T.C., 1982. Aquatic invertebrates from the Smoking Hills,N.W.T.: effect of pH and metals on mortality. Can. J. Fish. Aquat. Sci. 39, 890–903.
Havens, K.E., Yan, N.D., Keller, W., 1993. Lake acidification: effects on crustaceanzooplankton populations. Environ. Sci. Technol. 27, 1621–1624.
Hawkins, D.G., 2004. No exit: thinking about leakage from geologic carbon storagesites. Energy 29, 1571–1578.
Hays, G.C., Richardson, A.J., Robinson, C., 2005. Climate change and marineplankton. Trends Ecol. Evol. 20.
Hendriks, I.E., Duarte, C.M., Alvarez, M., 2010. Vulnerability of marine biodiversityto ocean acidification: a meta-analysis. Est. Coastal Shelf Sci. 86, 157–164.
Herzog, H.J., Adams, E.E., Auerbach, D., Caulfield, J., 1996. Environmental impacts ofocean disposal of CO2. Energy Conver. Manage. 37, 999–1005.
Hirche, H.-J., Niehoff, B., 1996. Reproduction of the Arctic copepod Calanushyperboreus in the Greenland Sea-field and laboratory observations. PolarBiol. 16, 209–219.
Holloway, S., 2005. Underground sequestration of carbon dioxide – a viablegreenhouse gas mitigation option. Energy 30, 2318–2333.
House, K.Z., Schrag, D.P., Harvey, C.F., Lackner, K.S., 2006. Permanent carbon dioxidestorage in deep-sea sediments. PNAS 103, 12291–12295.
Ikeda, T., Hirakawa, K., 1996. Early development and estimated life cycle of themesopelagic copepod Pareuchaeta elongata in the southern Japan Sea. Mar. Biol.126, 261–270.
Invidia, M., Sei, S., Gorbi, G., 2004. Survival of the copepod Acartia tonsa followingegg exposure to near anoxia and to sulfide at different pH values. Mar. Ecol.Prog. Ser. 276, 187–196.
IPCC, 2005. IPCC special report on carbon dioxide capture and storage. Prepared byworking group III of the Intergovernmental panel on climate change. In: Metz,B., Davidson, O., de Coninck, H. C., Loos, M., Meyer, L.A., (Eds.). CambridgeUniversity Press, Cambridge, United Kingdom and New York, NY, USA, 442 pp.
Ishimaru, T., 2001. Impact of CO2 on marine organisms. Kaiyo Monthly 377, 785–790, in Japanese.
Ishimatsu, A., Kikkawa, T., Hayashi, M., Lee, K.-S., Kita, J., 2004. Effects of CO2 onmarine fish: larvae and adults. J. Oceanogr. 60, 731–741.
Kaneko, T., Hasegawa, S., Uchida, K., Ogasawara, T., Oyagi, A., Hirano, T., 1995.Hypoosmoregulatory ability of eyed-stage embryos of chum salmon. Mar. Biol.122, 165–170.
Kawaguchi, S., Kurihara, H., King, R., Hale, L., Berli, T., Robinson, J.P., Ishida, A.,Wakita, M., Virtue, P., Nicol, S., Ishimatsu, A., 2010. Will krill fare well underSouthern Ocean acidification? Biol. Lett.. http://dx.doi.org/10.1098/rsbl.2010.0777.
Kikkawa, T., Kita, J., Ishimatsu, A., 2004. Comparison of the lethal effect of CO2 andacidification on red sea bream (Pagrus major) during the early developmentalstages. Mar. Pollut. Bull. 48, 108–110.
Kroeker, K.J., Korda, R.L., Crim, R.N., Singh, G.G., 2010. Meta-analysis revealsnegative yet variable effects of ocean acidification on marine organisms. Ecol.Lett. 13, 1419–1434.
Kurihara, H., 2008. Effects of CO2-driven ocean acidification on the earlydevelopmental stages of invertebrates. Mar. Ecol. Prog. Ser. 373, 8–9.
Kurihara, H., Ishimatsu, A., 2008. Effects of high CO2 seawater on the copepod(Acartia tsuensis) through all life stages and subsequent generations. Mar. Pollut.Bull. 56, 1086–1090.
Kurihara, H., Shirayama, Y., 2004. Effects of increased atmospheric CO2 on seaurchin early development. Mar. Ecol. Prog. Ser. 274, 161–169.
Kurihara, H., Shimode, S., Shirayama, Y., 2004a. Sub-lethal effects of elevatedconcentration of CO2 on planktonic copepods and sea urchins. J. Oceanogr. 60,743–750.
Kurihara, H., Shimode, S., Shirayama, Y., 2004b. Effects of raised CO2 concentrationon the egg production rate and early development of two marine copepods(Acartia steueri and Acartia erythraea). Mar. Pollut. Bull. 49, 721–727.
Kurihara, H., Kato, S., Ishimatsu, A., 2007. Effects of increased seawater pCO2 onearly development of the oyster Crassostrea gigas. Aquatic Biol. 1, 91–98.
Kurihara, H., Asai, T., Kato, S., Ishimatsu, A., 2008a. Effects of elevated pCO2 on earlydevelopment in the mussel Mytilus galloprovincialis. Aquatic Biol. 4, 225–233.
Kurihara, H., Matsui, M., Furukawa, H., Hayashi, M., Ishimatsu, A., 2008b. Long-termeffects of predicted future seawater CO2 conditions on the survival and growthof the marine shrimp Palaemon pacificus. J. Exp. Mar. Biol. Ecol. 367, 41–46.
Lacoue-Labarthe, T., Martin, S., Oberhänsli, F., Teyssié, J.-L., Markich, S., Ross, J.,Bustamante, P., 2009. Effects of increased pCO2 on trace element (Ag, Cd, Zn)bioaccumulation in the eggs of the common cuttlefish, Sepia officinalis.Biogeosciences 6, 2561–2573.
Lacoue-Labarthe, T., Réveillac, E., Oberhänsli, F., Teyssié, J.L., Jeffree, R., Gattuso, J.P.,2011. Effects of ocean acidification on trace element accumulation in the early-life stages of squid Loligo vulgaris. Aquatic Toxicol. 105, 166–176.
Manno, C., Morata, N., Bellerby, R., 2012. Effect of ocean acidification andtemperature increase on the planktonic foraminifer Neogloboquadrinapachyderma (sinistral). Polar Biol.. http://dx.doi.org/10.1007/s00300-012-1174-7.
Mauchline, J., Fisher, L.R., 1969. The biology of euphausiids. Adv. Mar. Biol. 7, 1–454.Mayor, D.J., Matthews, C., Cook, K., Zuur, A.F., Hay, S., 2007. CO2-induced
acidification affects hatching success in Calanus finmarchicus. Mar. Ecol. Prog.Ser. 350, 91–97.
Mayor, D.J., Everett, N.R., Cook, K.B., 2012. End of century ocean warming andacidification effects on reproductive success in a temperate marine copepod. J.Plankton Res. 34, 258–262.
McConville, K., Halsband, C., Fileman, E.S., Findlay, H.S., Somerfield, P.J., Spicer, J.I.,2013. Effect of elevated CO2 on the reproduction of two calanoid copepods. Mar.Poll. Bull.
Melzner, F., Gutowska, M.A., Langenbuch, M., Dupont, S., Lucassen, M., Thorndyke,M.C., Bleich, M., Pörtner, H.O., 2009. Physiological basis for high CO2 tolerance inmarine ectothermic animals: pre-adaptation through lifestyle and ontogeny?Biogeosciences 6, 2313–2331.
Miles, H., Widdicombe, S., Spicer, J.I., Hall-Spencer, J., 2007. Effects of anthropogenicseawater acidification on acid-base balance in the sea urchin Psammechinusmiliaris. Mar. Pollut. Bull. 54, 89–96.
Moy, A.D., Howard, W.R., Bray, S.G., Trull, T.W., 2009. Reduced calcification inmodern Southern Ocean planktonic foraminifera. Nat. Geosci. 2, 276–280.
Niehoff, B., Knüppel, N., Daase, M., Czerny, J., Boxhammer, T., 2012.Mesozooplankton community development at elevated CO2 concentrations:results from a mesocosm experiment in an Arctic fjord. Biogeosci. Discuss. 9,11479–11515.
O’Donnell, M.J., Todgham, A.E., Sewell, M.A., Hammond, L.M., Ruggiero, K., Fangue,N.A., Zippay, M.L., Hofmann, G.E., 2010. Ocean acidification alters skeletogenesisand gene expression in larval sea urchins. Mar. Ecol. Prog. Ser. 398.
Omori, M., Norman, C.P., Ikeda, T., 1998. Oceanic disposal of CO2: potential effectson deep-sea plankton and micronekton – a review. Plankton Biol. Ecol. 45, 87–99.
Orr, J.C., Fabry, V.J., Aumont, O., Bopp, L., Doney, S.C., Feely, R.A., Gnanadesikan, A.,Gruber, N., Ishida, A., Joos, F., Key, R.M., Lindsay, K., Maier-Reimer, E., Matear, R.,Monfray, P., Mouchet, A., Najjar, R.G., Plattner, G.K., Rodgers, K.B., Sabine, C.L.,Sarmiento, J.L., Schlitzer, R., Slater, R.D., Totterdell, I.J., Weirig, M.F., Yamanaka,Y., Yool, A., 2005. Anthropogenic ocean acidification over the twenty-firstcentury and its impact on calcifying organisms. Nature 437, 681–686.
Pane, E.F., Barry, J.P., 2007. Extracellular acid-base regulation during short-termhypercapnia is effective in a shallow-water crab, but ineffective in a deep-seacrab. Mar. Ecol. Prog. Ser. 334, 1–9.
Parker, L.M., Ross, P.M., O’Connor, W.A., 2009a. The effect of ocean acidification andtemperature on the fertilization and embryonic development of the Sydneyrock oyster Saccostrea glomerata (Gould 1850). Glob. Change Biol. 15 (9), 2123–2136.
Parker, L.M., Ross, P.M., O’Connor, W.A., 2009b. The effect of ocean acidification andtemperature on the fertilization and embryonic development of the Sydneyrock oyster Saccostrea glomerata (Gould 1850). Glob. Change Biol. 15, 2123–2136.
Parker, L.M., Ross, P.M., O’Connor, W.A., 2010. Populations of the Sydney rock oyster,Saccostrea glomerata, vary in response to ocean acidification. Mar. Biol. 158,689–697.
Raven, J., Caldeira, K., Elderfield, H., Hoegh-Guldberg, O., Liss, P., Riebesell, U.,Shepherd, J., Turley, C., Watson, A., 2005. Ocean acidification due to increasingatmospheric carbon dioxide. Policy Document 12/05. Royal Society, London.
502 C. Halsband, H. Kurihara / Marine Pollution Bulletin 73 (2013) 495–503

Author's personal copy
Reid, P.C., Fischer, A.C., Lewis-Brown, E., Meredith, M.P., Sparrow, M., Andersson,A.J., Antia, A., Bates, N.R., Bathmann, U., Beaugrand, G., Brix, H., Dye, S., Edwards,M., Furevik, T., Gangst, R., Hbtn, H., Hopcroft, R.R., Kendall, M., Kasten, S.,Keeling, R., Le Quéré, C., Mackenzie, F.T., Malin, G., Mauritzen, C., Ëlafsson, Jn,Paull, C., Rignot, E., Shimada, K., Vogt, M., Wallace, C., Wang, Z., Washington, R.,2009. Impacts of the oceans on climate change. In: David, W.S. (Ed.), Advancesin Marine Biology, vol. 56. Academic Press, pp. 1–150, Chapter 1.
Richardson, A.J., Gibbons, M.J., 2008. Are jellyfish increasing in response to oceanacidification? Limnol. Oceanogr. 53, 2035–2040.
Riebesell, U., Tortell, P.D., 2011. Effects of ocean acidification on pelagic organismsand ecosystems. In: Gattuso, J.-P., Hansson, L. (Eds.), Ocean Acidification. OxfordUniversity Press, pp. 99–121.
Rose, C.D., Williams, W.G., Hollister, T.A., Parrish, P.R., 1977. Method fordetermining acute toxicity of an acid waste and limiting permissibleconcentration at boundaries of an oceanic mixing zone. Environ. Sci. Technol.11, 367–371.
Seibel, B.A., Walsh, P.J., 2003. Biological impacts of deep-sea carbon dioxideinjection inferred from indices of physiological performance. J. Exp. Biol. 206,641–650.
Shiraishi, K., Kaneko, T., Hasegawa, S., Hirano, T., 1997. Development ofmulticellular complexes of chloride in the yolk-sac membrane of tilapia(Oreochromis mossambicus) embryos and larvae in seawater. Cell Tiss. Res.288, 583–590.
Spicer, J.I., Raffo, A., Widdicombe, S., 2007. Influence of CO2-related seawateracidification on extracellular acid-base balance in the velvet swmming crabNecore puber. Mar. Biol. 151, 1117–1125.
Stephens, J.C., van der Zwaan, B., 2005. The case of carbon capture and storage. Iss.Sci. Technol. 22, 68–76.
Stumpp, M., Wren, J., Melzner, F., Thorndyke, M.C., Dupont, S.T., 2011. CO2 inducedseawater acidification impacts sea urchin larval development I: elevatedmetabolic rates decrease scope for growth and induce developmental delay.Comp. Biochem. Physiol. – Part A: Mol. Int. Physiol. 160 (3), 331–340.
Talmage, S.C., Gobler, C.J., 2009. The effects of elevated carbon dioxideconcentrations on the metamorphosis, size, and survival of larval hard clams(Mercenaria mercenaria), bay scallops (Argopecten irradians), and Eastern oysters(Crassostrea virginica). Limnol. Oceanogr. 54 (6), 2072–2080.
Todgham, A.E., Hofmann, G.E., 2009. Transcriptome response of sea urchin larvaeStrongylocentrotus purpuratus to CO2-driven seawater acidification. J. Exp. Biol.212, 2579–2594.
Tyler, P.A., 1986. Studies of a benthic time series: reproductive biology of benthicinvertebrates in the Rockall Trough. Proc. R. Soc. Edinb. (Sect. B) 88, 175–190.
Uthicke, S., Fabricius, K.E., 2012. Productivity gains do not compensate for reducedcalcification under near-future ocean acidification in the photosyntheticbenthic foraminifer species Marginopora vertebralis. Glob. Change Biol. 18 (9),2781–2791.
Vanderploeg, H.A., Stuart, A., Ludsina, S.A., Cavaletto, J.F., Höök, T.O., Pothovend, S.A.,Brandt, S.B., Liebig, J.R., Lang, G.A., 2009. Hypoxic zones as habitat forzooplankton in Lake Erie: refuges from predation or exclusion zones? J. Exp.Mar. Biol. Ecol. 381, 108–120.
Walton, W.E., Compton, S.M., Allan, J.D., Daniels, R.E., 1982. The effect of acid stresson survivorship and reproduction of Daphnia pulex (Crustacea: Cladocera). Can.J. Zool. 60, 573–579.
Watanabe, Y., Yamaguchi, A., Ishida, H., Harimoto, T., Suzuki, S., Sekido, Y., Ikeda, T.,Shirayama, Y., Mac Takahashi, M., Ohsumi, T., Ishizaka, J., 2006. Lethality ofincreasing CO2 levels on deep-sea copepods in the western North Pacific. J.Oceanogr. 62, 185–196.
Weydmann, A., Søreide, J.E., Kwasniewski, S., Widdicombe, S., 2012. Influence ofCO2-induced acidification on the reproduction of a key Arctic copepod Calanusglacialis. J. Exp. Mar. Biol. Ecol. 428, 39–42.
Wheatly, M.G., Henry, R.P., 1992. Extracellular and intracellular acid–baseregulation in crustaceans. J. Exp. Zool. 263, 127–142.
Widdicombe, S., Spicer, J.I., 2008. Predicting the impact of ocean acidification onbenthic biodiversity: what can animal physiology tell us? J. Exp. Mar. Biol. Ecol.366, 187–197.
Winans, A.K., Purcell, J.E., 2010. Effects of pH on asexual reproduction and statolithformation of the scyphozoan, Aurelia labiata. Hydrobiologia 645 (1), 39–52.
Wood, H.L., Spicer, J.I., Widdicombe, S., 2008. Ocean acidification may increasecalcification rates, but at a cost. Proc. R. Soc. B. http://dx.doi.org/10.1098/rspb.2008.0343.
Yamada, Y., Ikeda, T., 1999. Acute toxicity of lowered pH to some oceaniczooplankton. Plankton Biol. Ecol. 46, 62–67.
Zhang, X., Dam, H.G., 1997. Downward export of carbon by diel migrantmesozooplankton in the central equatorial Pacific. Deep-Sea Res. Part II:Topical Stud. Oceanogr. 44, 2191–2202.
C. Halsband, H. Kurihara / Marine Pollution Bulletin 73 (2013) 495–503 503




















