
Pantoea: insights into a highly versatile and diverse genus ...
17
FEMS Microbiology Reviews, fuv027, 39, 2015, 968–984 doi: 10.1093/femsre/fuv027 Advance Access Publication Date: 24 June 2015 Review Article REVIEW ARTICLE Pantoea: insights into a highly versatile and diverse genus within the Enterobacteriaceae Alyssa M. Walterson and John Stavrinides ∗ Department of Biology, University of Regina, 3737 Wascana Parkway, Regina, SK S4S 0A2, Canada ∗ Corresponding author: Department of Biology, University of Regina, 3737 Wascana Parkway, Regina, SK S4S 0A2, Canada. Tel: (306) 337-8478; Fax: (306) 337-2410; E-mail: [email protected] One sentence summary: This review synthesizes what is currently known about the diverse enterobacterial group, Pantoea, which not only exhibits versatility in both host and non-host environments, but also shows biotechnological promise. Editor: Jan Roelof van der Meer ABSTRACT The bacterial genus Pantoea comprises many versatile species that have been isolated from a multitude of environments. Pantoea was delineated as a genus approximately 25 years ago, but since then, approximately 20 species have been identified having a diversity of characteristics. Isolates from water and soil have been harnessed for industrial purposes including bioremediation, and the degradation of herbicides and other toxic products. Other isolates possess nitrogen fixation and plant growth-promoting capabilities, which are currently being explored for agricultural applications. Some isolates are antibiotic producers, and have been developed into biocontrol agents for the management of plant diseases. Pantoea is also known to form host associations with a variety of hosts, including plants, insects and humans. Although often thought of as a plant pathogen, recent evidence suggests that Pantoea is being frequently isolated from the nosocomial environment, with considerable debate as to its role in human disease. This review will explore this highly versatile group and its capabilities, its known associations, and the underlying genetic and genomic determinants that drive its diversity and adaptability. Keywords: Pantoea; enterobacterial plant pathogen; nitrogen fixation; opportunistic pathogen; soil; symbiont; mutualism; epiphyte; antibiotic production; human pathogen; bioremediation; biocontrol; Enterobacter agglomerans; Erwinia herbicola INTRODUCTION The genus Pantoea is a diverse group of yellow-pigmented, rod- shaped Gram-negative bacteria in the Enterobacteriaceae. Some of the first members were recognized as plant pathogens caus- ing galls, wilting, soft rot and necrosis in a variety of agricul- turally relevant plants, but since then, Pantoea strains have been frequently isolated from many aquatic and terrestrial environ- ments, as well as in association with insects, animals and hu- mans (Muraschi, Friend and Bolles 1965; Ewing and Fife 1972; Brady et al. 2008; Volksch et al. 2009; Nadarasah and Stavrinides 2014). Some Pantoea isolates produce antimicrobials, and have been developed into commercial biocontrol products, such as BlightBan C9-1 and Bloomtime Biological, to help control fire blight of apple and pear trees (Johnson et al. 1993, 2000; Johnson and Stockwell 1998), while others have bioremediation poten- tial, with the capacity to degrade herbicides without the gen- eration of toxic by-products (Pileggi et al. 2012). As well, some isolates have been harnessed as immunopotentiators for the de- velopment of supportive drugs for melanoma, infections, aller- gies and the reversal of immunosuppression (Yoshida et al. 2009; Hebishima et al. 2011; Nakata, Inagawa and Soma 2011). The ubiquity, versatility and genetic tractability of Pantoea isolates makes it an ideal group for not only exploring niche-specific adaptation and opportunism, but also for the development of commercially relevant medical, agricultural and environmen- tal products. This review will examine the general biology of Received: 25 August 2014; Accepted: 20 May 2015 C FEMS 2015. All rights reserved. For permissions, please e-mail: [email protected] 968 Downloaded from https://academic.oup.com/femsre/article/39/6/968/547864 by guest on 26 March 2022
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Pantoea: insights into a highly versatile and diverse genus ...

FEMS Microbiology Reviews, fuv027, 39, 2015, 968–984
doi: 10.1093/femsre/fuv027Advance Access Publication Date: 24 June 2015Review Article
REVIEW ARTICLE
Pantoea: insights into a highly versatile and diversegenus within the EnterobacteriaceaeAlyssa M. Walterson and John Stavrinides∗
Department of Biology, University of Regina, 3737 Wascana Parkway, Regina, SK S4S 0A2, Canada∗Corresponding author: Department of Biology, University of Regina, 3737 Wascana Parkway, Regina, SK S4S 0A2, Canada. Tel: (306) 337-8478; Fax: (306)337-2410; E-mail: [email protected] sentence summary: This review synthesizes what is currently known about the diverse enterobacterial group, Pantoea, which not only exhibitsversatility in both host and non-host environments, but also shows biotechnological promise.Editor: Jan Roelof van der Meer
ABSTRACT
The bacterial genus Pantoea comprises many versatile species that have been isolated from a multitude of environments.Pantoea was delineated as a genus approximately 25 years ago, but since then, approximately 20 species have beenidentified having a diversity of characteristics. Isolates from water and soil have been harnessed for industrial purposesincluding bioremediation, and the degradation of herbicides and other toxic products. Other isolates possess nitrogenfixation and plant growth-promoting capabilities, which are currently being explored for agricultural applications. Someisolates are antibiotic producers, and have been developed into biocontrol agents for the management of plant diseases.Pantoea is also known to form host associations with a variety of hosts, including plants, insects and humans. Althoughoften thought of as a plant pathogen, recent evidence suggests that Pantoea is being frequently isolated from thenosocomial environment, with considerable debate as to its role in human disease. This review will explore this highlyversatile group and its capabilities, its known associations, and the underlying genetic and genomic determinants thatdrive its diversity and adaptability.
Keywords: Pantoea; enterobacterial plant pathogen; nitrogen fixation; opportunistic pathogen; soil; symbiont; mutualism;epiphyte; antibiotic production; human pathogen; bioremediation; biocontrol; Enterobacter agglomerans; Erwinia herbicola
INTRODUCTION
The genus Pantoea is a diverse group of yellow-pigmented, rod-shaped Gram-negative bacteria in the Enterobacteriaceae. Someof the first members were recognized as plant pathogens caus-ing galls, wilting, soft rot and necrosis in a variety of agricul-turally relevant plants, but since then, Pantoea strains have beenfrequently isolated from many aquatic and terrestrial environ-ments, as well as in association with insects, animals and hu-mans (Muraschi, Friend and Bolles 1965; Ewing and Fife 1972;Brady et al. 2008; Volksch et al. 2009; Nadarasah and Stavrinides2014). Some Pantoea isolates produce antimicrobials, and havebeen developed into commercial biocontrol products, such asBlightBan C9-1 and Bloomtime Biological, to help control fire
blight of apple and pear trees (Johnson et al. 1993, 2000; Johnsonand Stockwell 1998), while others have bioremediation poten-tial, with the capacity to degrade herbicides without the gen-eration of toxic by-products (Pileggi et al. 2012). As well, someisolates have been harnessed as immunopotentiators for the de-velopment of supportive drugs for melanoma, infections, aller-gies and the reversal of immunosuppression (Yoshida et al. 2009;Hebishima et al. 2011; Nakata, Inagawa and Soma 2011). Theubiquity, versatility and genetic tractability of Pantoea isolatesmakes it an ideal group for not only exploring niche-specificadaptation and opportunism, but also for the development ofcommercially relevant medical, agricultural and environmen-tal products. This review will examine the general biology of
Received: 25 August 2014; Accepted: 20 May 2015C© FEMS 2015. All rights reserved. For permissions, please e-mail: [email protected]
968
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/39/6/968/547864 by guest on 26 March 2022
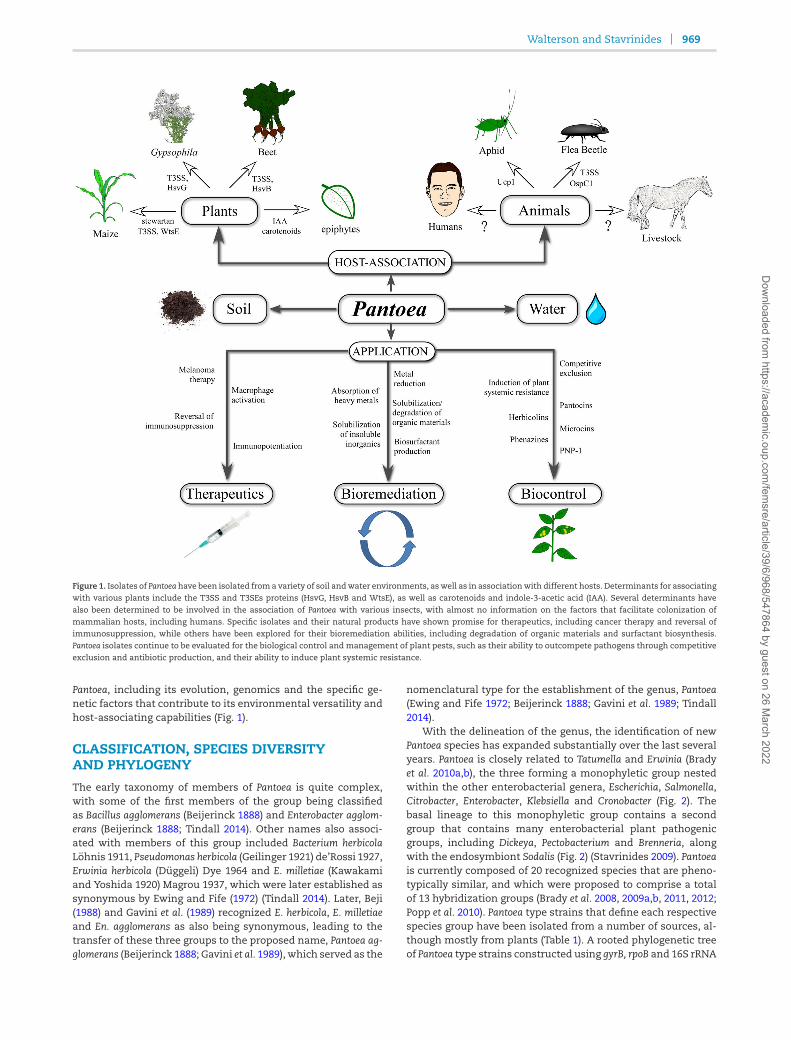
Walterson and Stavrinides 969
Figure 1. Isolates of Pantoea have been isolated from a variety of soil andwater environments, aswell as in associationwith different hosts. Determinants for associatingwith various plants include the T3SS and T3SEs proteins (HsvG, HsvB and WtsE), as well as carotenoids and indole-3-acetic acid (IAA). Several determinants havealso been determined to be involved in the association of Pantoea with various insects, with almost no information on the factors that facilitate colonization of
mammalian hosts, including humans. Specific isolates and their natural products have shown promise for therapeutics, including cancer therapy and reversal ofimmunosuppression, while others have been explored for their bioremediation abilities, including degradation of organic materials and surfactant biosynthesis.Pantoea isolates continue to be evaluated for the biological control and management of plant pests, such as their ability to outcompete pathogens through competitiveexclusion and antibiotic production, and their ability to induce plant systemic resistance.
Pantoea, including its evolution, genomics and the specific ge-netic factors that contribute to its environmental versatility andhost-associating capabilities (Fig. 1).
CLASSIFICATION, SPECIES DIVERSITYAND PHYLOGENY
The early taxonomy of members of Pantoea is quite complex,with some of the first members of the group being classifiedas Bacillus agglomerans (Beijerinck 1888) and Enterobacter agglom-erans (Beijerinck 1888; Tindall 2014). Other names also associ-ated with members of this group included Bacterium herbicolaLohnis 1911, Pseudomonas herbicola (Geilinger 1921) de’Rossi 1927,Erwinia herbicola (Duggeli) Dye 1964 and E. milletiae (Kawakamiand Yoshida 1920) Magrou 1937, which were later established assynonymous by Ewing and Fife (1972) (Tindall 2014). Later, Beji(1988) and Gavini et al. (1989) recognized E. herbicola, E. milletiaeand En. agglomerans as also being synonymous, leading to thetransfer of these three groups to the proposed name, Pantoea ag-glomerans (Beijerinck 1888; Gavini et al. 1989), which served as the
nomenclatural type for the establishment of the genus, Pantoea(Ewing and Fife 1972; Beijerinck 1888; Gavini et al. 1989; Tindall2014).
With the delineation of the genus, the identification of newPantoea species has expanded substantially over the last severalyears. Pantoea is closely related to Tatumella and Erwinia (Bradyet al. 2010a,b), the three forming a monophyletic group nestedwithin the other enterobacterial genera, Escherichia, Salmonella,Citrobacter, Enterobacter, Klebsiella and Cronobacter (Fig. 2). Thebasal lineage to this monophyletic group contains a secondgroup that contains many enterobacterial plant pathogenicgroups, including Dickeya, Pectobacterium and Brenneria, alongwith the endosymbiont Sodalis (Fig. 2) (Stavrinides 2009). Pantoeais currently composed of 20 recognized species that are pheno-typically similar, and which were proposed to comprise a totalof 13 hybridization groups (Brady et al. 2008, 2009a,b, 2011, 2012;Popp et al. 2010). Pantoea type strains that define each respectivespecies group have been isolated from a number of sources, al-though mostly from plants (Table 1). A rooted phylogenetic treeof Pantoea type strains constructed using gyrB, rpoB and 16S rRNA
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/39/6/968/547864 by guest on 26 March 2022

970 FEMS Microbiology Reviews, 2015, Vol. 39, No. 6
Figure 2. Cladogram obtained from the Pathosystems Resource Integration Cen-
ter (Wattam et al. 2014), which was constructed using shared protein homologsderived from whole genomes of entire species groups within each genus in theEnterobacteriaceae. The relative position of Pantoea is indicated with a box. As ge-nomic information for Tatumella (sister taxon to Pantoea) has not yet been in-
corporated into the Pathosystems Resource Integration Center, it is not shown.Pseudomonas (Pseudomonadaceae) was used as the root.
shows the relationships between the established species groups(Fig. 3). Species like P. deleyi, P. anthophila, P. allii, P. cypripedii, P.wallisi, P. rodasii and P. rwandensis have been isolated from onlyplant sources, P. conspicua, P. brenneri, P. septica and P. eucrina from
only clinical sources and P. gavinae from only outdoor environ-mental sources. Isolates of P. calida, P. dispersa and P. gavinaewereidentified from the natural environment or processed products(Table 1). Previously, several other species had been proposed,including P. citrea, P. punctata and P. terrea (Kageyama et al. 1992);however, these were later reclassified into the genus Tatumellausingmultilocus sequence analysis (MLSA)methods (Brady et al.2010a,b).
ADVANCES IN OUR UNDERSTANDING OFPLANT-ASSOCIATED ISOLATES
Most of the early research on Pantoea focused on its parasiticassociation with plant hosts, including maize, cotton, melonand onion (Table 2) (Gitaitis and Gay 1997; Walcott et al. 2002;Medrano and Bell 2007; Kido et al. 2008; Brady et al. 2011).Plant diseases caused by phytopathogenic isolates are diverse(Table 2), and in some cases are shown to be facilitated by a suiteof pathogenicity factors that include the type III secretion sys-tem (T3SS), quorum-sensing systems and exopolysaccharides(EPS) (Roper 2011). The T3SS is responsible for the transport ofvirulence factors, or effector proteins, into host cells, resultingin the subversion of host defenses (Galan and Collmer 1999).For P. stewartii subsp. stewartii, the causative agent of Stewart’swilt disease in corn, the interaction with the host plant is medi-ated by a T3SS, PSI-1 (Correa et al. 2010, 2012). PSI-1 is a 24 kbgenomic island, similar to the Hrc-Hrp1 family of T3SSs thatis also found in P. agglomerans pv. gypsophilae (Frederick et al.2001; Coplin et al. 2002). Regulation of the PSI-1 T3SS involvesthe HrpX/HrpY two-component regulatory system (Merighi et al.2003, 2006). HrpY acts downstream in the activation of hrpS,which subsequently regulates hrpL in response to environmentalstimuli. HrpL, an alternate sigma factor, is then involved in theregulation and expression of effector genes (Merighi, Majerczakand Coplin 2005). HrpX, the other component in this regulatorysystem, is responsible for the expression of hrp genes and is ac-tivated under nutrient-limited conditions (Merighi et al. 2006).
Table 1. Pantoea type strains and source of isolation.
Species Type strain Origin Reference
P. agglomerans LMG 1286 Human, knee laceration Gavini et al. (1989)P. allii LMG 24248 Onion seed Brady et al. (2011)P. ananatis LMG 2665 Pineapple Mergaert, Verdonck and Kersters (1993)P. anthophila LMG 2558 Garden balsam Brady et al. (2009a,b)P. beijingensis LMG 27529 King trumpet mushroom Liu et al. (2013)P. brenneri LMG 5343 Human, urethra Brady et al. (2010a,b)P. calida LMG 25383 Powdered infant formula Popp et al. (2010)P. conspicua LMG 24534 Human, blood Brady et al. (2010a,b)P. cypripedii LMG 2657 Orchid Brady et al. (2010a,b)P. deleyi LMG 24200 Eucalyptus Brady et al. (2009a,b)P. dispersa LMG 2603 Soil Gavini et al. (1989)P. eucalyptii LMG 24197 Eucalyptus Brady et al. (2009a,b)P. eucrina LMG 2781 Human, trachea Brady et al. (2010a,b)P. gavinae LMG 25382 Powdered infant formula Popp et al. (2010)P. rodasii LMG 26273 Eucalyptus Brady et al. (2012)P. rwandensis LMG 26275 Eucalyptus Brady et al. (2012)P. septica LMG 5345 Human, stool Brady et al. (2010a,b)P. stewartii subsp. indologenes LMG 2632 Foxtail millet Mergaert, Verdonck and Kersters (1993)P. stewartii subsp. stewartii LMG 2715 Maize Mergaert, Verdonck and Kersters (1993)P. vagans LMG 24199 Eucalyptus Brady et al. (2009a,b)P. wallisii LMG 26277 Eucalyptus Brady et al. (2012)
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/39/6/968/547864 by guest on 26 March 2022

Walterson and Stavrinides 971
Figure 3.Neighbor-joining phylogeny of Pantoea type strains, based on a concate-nated dataset composed of partial rpoB, gyrB, atpD and 16S rRNA genes using
maximum composite likelihood. Nodes show the result of 1000 bootstrap repli-cates, with values >50% being shown. Symbols adjacent to each representativetype strain indicate whether strains within that species group have been iso-lated from clinical settings (red circle), from plant hosts either as an epiphyte or
a pathogen (green square), or from the natural environment (purple triangle).
Activation of the T3SS results in the injection of type III secretedeffectors (T3SEs) into the host cell, which function to facilitatehost-specific interactions between the bacteriumand host. HsvBand HsvG are T3SEs carried by P. agglomerans pv. betae and P. ag-glomerans pv. gypsophilae, respectively, and have been shown toplay an important role in host-specific colonization (Nissan et al.2006; Weinthal et al. 2011; Kirzinger and Stavrinides 2012). Theintroduction of HsvB into beet and HsvG into gypsophila influ-ences plant transcription and leads to gall formation. When theT3SEswere introduced into the other host (HsvB in gypsophila orHsvG into beet), then no galls would form, suggesting that hostspecificity is determined at the gene level (Nissan et al. 2006).
Quorum sensing is a process whereby bacteria monitor theirpopulation density through the use of small diffusible signalmolecules (Minogue et al. 2005; Koutsoudis et al. 2006), and canalso be involved in regulating gene systems for pathogenesis andhost interactions (Koutsoudis et al. 2006). For P. stewartii subsp.stewartii, quorum sensing influences adhesion, motility and dis-persion, as well as EPS production, and plays a major role in thedevelopment of Stewart’s wilt disease of corn (von Bodman, Ma-jerczak and Coplin 1998; Koutsoudis et al. 2006). Pantoea stewartiicolonizes the xylem of the plant where it multiples and pro-duces EPS, thereby blocking the flow of water and causing theplant to wilt. The production of the EPS is regulated through theEsaI/EsaR quorum-sensing system, which governs the cps genecluster that is responsible for EPS biosynthesis in a cell-density-dependent manner (Koutsoudis et al. 2006). EsaR is thought toact through the repression of the rcsA gene, which in turn acts asa coactivator for RcsA/RcaB-mediated transcriptional activationof the cps gene cluster (Minogue et al. 2002, 2005). EPS not onlyaids in the development of Stewart’s wilt disease symptoms, butis thought to also provide the invading bacteria with the neces-sary protection from plant host defenses (Wehland et al. 1999).The presence of the cps gene cluster is an important geneticcomponent in the overall pathogenicity of P. stewartii, along withthe T3SE, WtsE/AvrE, which results in water soaking, and subse-quent cell death in host plants (Ham et al. 2008).
Epiphytic populations of Pantoea have been identified onmany plants, as well as on vegetables, fruits and grains, sug-gesting they possess specific adaptations that facilitate colo-nization of the phyllosphere (Table 2) (Brandl and Lindow 1998;Beattie and Lindow 1999; Venturini, Oria and Blanco 2002; Lin-dow and Brandl 2003; Oie et al. 2008; Nadarasah and Stavrinides2014). For some isolates, the ability to compete and surviveepiphytically is facilitated by the production of indole-3-aceticacid, which has been shown to contribute to the overall nu-trient leakage of the plant leaves, and overall survival (Brandl,Clark and Lindow 1996; Brandl and Lindow 1998). Likewise, theproduction of carotenoids responsible for the bright yellow pig-ment appears to play a major role in resistance to ultraviolet(UV) radiation, which allows for successful epiphytic coloniza-tion (Mohammadi, Burbank and Roper 2012). Carotenoids, whichare often localized to the hydrophobic regions of themembrane,are able to protect the cell from harmful UV wavelengths (320–400 nm), as well as harmful reactive oxygen species that areactivated in the presence of UV during crucial growing phases(Mohammadi, Burbank and Roper 2012). Mutants that lackedpigmentation had an overall decreased survivorship over wildtype in the presence of H2O2 on nutrient-rich media (Moham-madi, Burbank and Roper 2012).
INSECT ASSOCIATIONS
Muchof the research surrounding the association of Pantoeawithinsects has stemmed from the specific adaptations of P. stew-artii subsp. stewartiiDC283 (DC283) to its flea beetle vector (Roper2011). The bacteria are carried in the insect gut, which are de-posited on the surface of sweet corn via the insect frass (Eskerand Nutter 2003; Menclas et al. 2006; Roper 2011). The bacteriagain entry into the plant via wounds created by the feeding bee-tles, which results in the colonization of either the apoplast orthe xylem vasculature, the production of EPS and the develop-ment of the characteristic wilt commonly attributed to P. stew-artii (Leigh andCoplin 1992). Persistencewithin the corn flea bee-tle gut by DC283 is facilitated by an animal-specific T3SS, PSI-2,which allows for successful colonization of the corn flea beetle,
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/39/6/968/547864 by guest on 26 March 2022
972 FEMS Microbiology Reviews, 2015, Vol. 39, No. 6
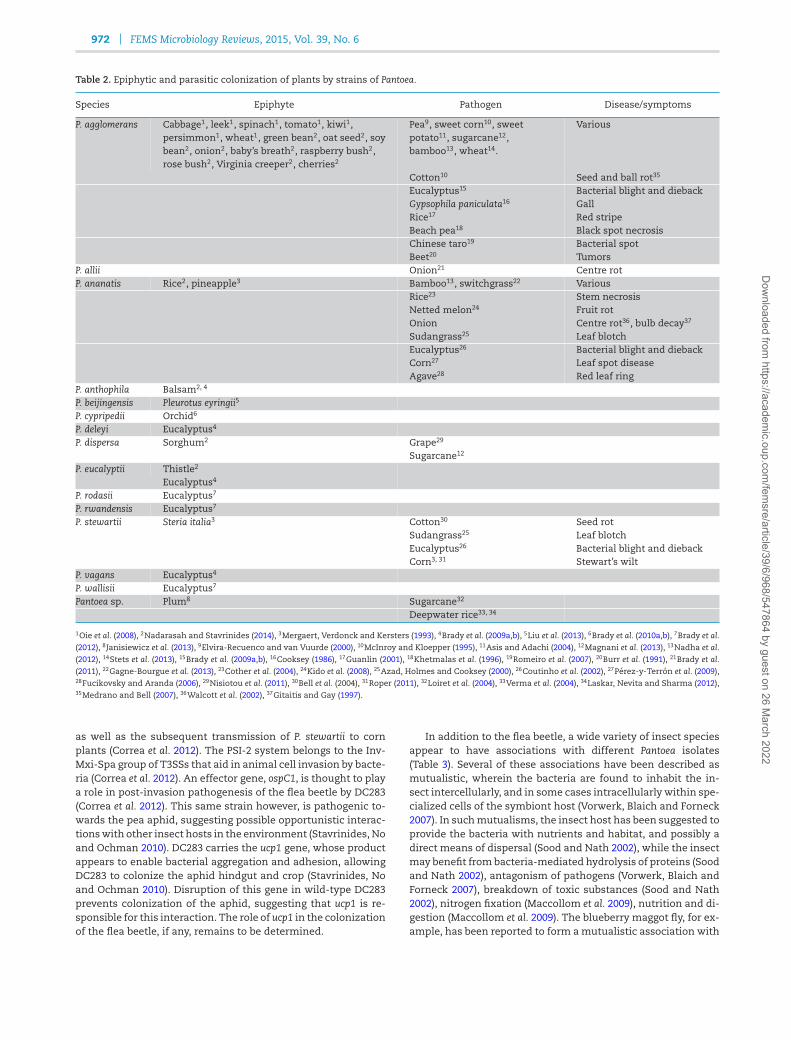
Table 2. Epiphytic and parasitic colonization of plants by strains of Pantoea.
Species Epiphyte Pathogen Disease/symptoms
P. agglomerans Cabbage1, leek1, spinach1, tomato1, kiwi1,persimmon1, wheat1, green bean2, oat seed2, soybean2, onion2, baby’s breath2, raspberry bush2,rose bush2, Virginia creeper2, cherries2
Pea9, sweet corn10, sweetpotato11, sugarcane12,bamboo13, wheat14.
Various
Cotton10 Seed and ball rot35
Eucalyptus15 Bacterial blight and diebackGypsophila paniculata16 GallRice17 Red stripeBeach pea18 Black spot necrosisChinese taro19 Bacterial spotBeet20 Tumors
P. allii Onion21 Centre rotP. ananatis Rice2, pineapple3 Bamboo13, switchgrass22 Various
Rice23 Stem necrosisNetted melon24 Fruit rotOnion Centre rot36, bulb decay37
Sudangrass25 Leaf blotchEucalyptus26 Bacterial blight and diebackCorn27 Leaf spot diseaseAgave28 Red leaf ring
P. anthophila Balsam2, 4
P. beijingensis Pleurotus eyringii5
P. cypripedii Orchid6
P. deleyi Eucalyptus4
P. dispersa Sorghum2 Grape29
Sugarcane12
P. eucalyptii Thistle2
Eucalyptus4
P. rodasii Eucalyptus7
P. rwandensis Eucalyptus7
P. stewartii Steria italia3 Cotton30 Seed rotSudangrass25 Leaf blotchEucalyptus26 Bacterial blight and diebackCorn3, 31 Stewart’s wilt
P. vagans Eucalyptus4
P. wallisii Eucalyptus7
Pantoea sp. Plum8 Sugarcane32
Deepwater rice33, 34
1Oie et al. (2008), 2Nadarasah and Stavrinides (2014), 3Mergaert, Verdonck and Kersters (1993), 4Brady et al. (2009a,b), 5Liu et al. (2013), 6Brady et al. (2010a,b), 7Brady et al.
(2012), 8Janisiewicz et al. (2013), 9Elvira-Recuenco and van Vuurde (2000), 10McInroy and Kloepper (1995), 11Asis and Adachi (2004), 12Magnani et al. (2013), 13Nadha et al.
(2012), 14Stets et al. (2013), 15Brady et al. (2009a,b), 16Cooksey (1986), 17Guanlin (2001), 18Khetmalas et al. (1996), 19Romeiro et al. (2007), 20Burr et al. (1991), 21Brady et al.
(2011), 22Gagne-Bourgue et al. (2013), 23Cother et al. (2004), 24Kido et al. (2008), 25Azad, Holmes and Cooksey (2000), 26Coutinho et al. (2002), 27Perez-y-Terron et al. (2009),28Fucikovsky and Aranda (2006), 29Nisiotou et al. (2011), 30Bell et al. (2004), 31Roper (2011), 32Loiret et al. (2004), 33Verma et al. (2004), 34Laskar, Nevita and Sharma (2012),35Medrano and Bell (2007), 36Walcott et al. (2002), 37Gitaitis and Gay (1997).
as well as the subsequent transmission of P. stewartii to cornplants (Correa et al. 2012). The PSI-2 system belongs to the Inv-Mxi-Spa group of T3SSs that aid in animal cell invasion by bacte-ria (Correa et al. 2012). An effector gene, ospC1, is thought to playa role in post-invasion pathogenesis of the flea beetle by DC283(Correa et al. 2012). This same strain however, is pathogenic to-wards the pea aphid, suggesting possible opportunistic interac-tionswith other insect hosts in the environment (Stavrinides, Noand Ochman 2010). DC283 carries the ucp1 gene, whose productappears to enable bacterial aggregation and adhesion, allowingDC283 to colonize the aphid hindgut and crop (Stavrinides, Noand Ochman 2010). Disruption of this gene in wild-type DC283prevents colonization of the aphid, suggesting that ucp1 is re-sponsible for this interaction. The role of ucp1 in the colonizationof the flea beetle, if any, remains to be determined.
In addition to the flea beetle, a wide variety of insect speciesappear to have associations with different Pantoea isolates(Table 3). Several of these associations have been described asmutualistic, wherein the bacteria are found to inhabit the in-sect intercellularly, and in some cases intracellularly within spe-cialized cells of the symbiont host (Vorwerk, Blaich and Forneck2007). In suchmutualisms, the insect host has been suggested toprovide the bacteria with nutrients and habitat, and possibly adirect means of dispersal (Sood and Nath 2002), while the insectmay benefit frombacteria-mediated hydrolysis of proteins (Soodand Nath 2002), antagonism of pathogens (Vorwerk, Blaich andForneck 2007), breakdown of toxic substances (Sood and Nath2002), nitrogen fixation (Maccollom et al. 2009), nutrition and di-gestion (Maccollom et al. 2009). The blueberry maggot fly, for ex-ample, has been reported to form amutualistic association with
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/39/6/968/547864 by guest on 26 March 2022
Walterson and Stavrinides 973
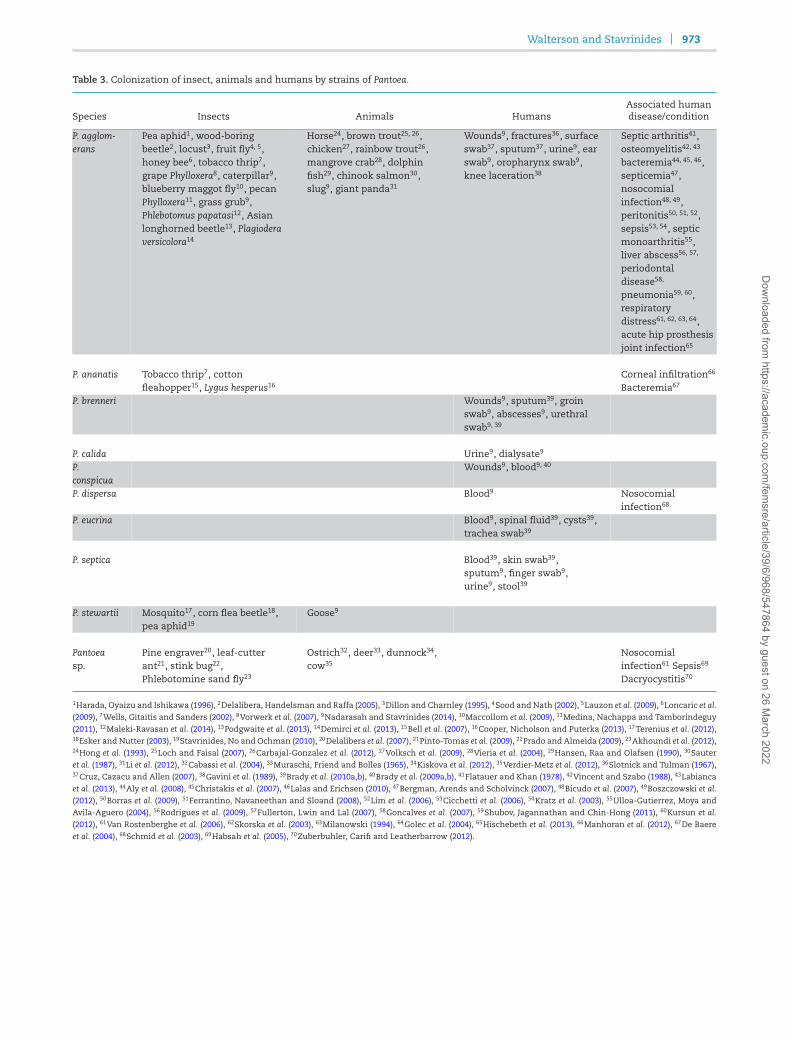
Table 3. Colonization of insect, animals and humans by strains of Pantoea.
Species Insects Animals HumansAssociated humandisease/condition
P. agglom-erans
Pea aphid1, wood-boringbeetle2, locust3, fruit fly4, 5,honey bee6, tobacco thrip7,grape Phylloxera8, caterpillar9,blueberry maggot fly10, pecanPhylloxera11, grass grub9,Phlebotomus papatasi12, Asianlonghorned beetle13, Plagioderaversicolora14
Horse24, brown trout25, 26,chicken27, rainbow trout26,mangrove crab28, dolphinfish29, chinook salmon30,slug9, giant panda31
Wounds9, fractures36, surfaceswab37, sputum37, urine9, earswab9, oropharynx swab9,knee laceration38
Septic arthritis41,osteomyelitis42, 43
bacteremia44, 45, 46,septicemia47,nosocomialinfection48, 49,peritonitis50, 51, 52,sepsis53, 54, septicmonoarthritis55,liver abscess56, 57,
periodontaldisease58,
pneumonia59, 60,respiratorydistress61, 62, 63, 64,acute hip prosthesisjoint infection65
P. ananatis Tobacco thrip7, cotton Corneal infiltration66
fleahopper15, Lygus hesperus16 Bacteremia67
P. brenneri Wounds9, sputum39, groinswab9, abscesses9, urethralswab9, 39
P. calida Urine9, dialysate9
P.conspicua
Wounds9, blood9, 40
P. dispersa Blood9 Nosocomialinfection68
P. eucrina Blood9, spinal fluid39, cysts39,trachea swab39
P. septica Blood39, skin swab39,sputum9, finger swab9,urine9, stool39
P. stewartii Mosquito17, corn flea beetle18,pea aphid19
Goose9
Pantoea Pine engraver20, leaf-cutter Ostrich32, deer33, dunnock34, Nosocomialsp. ant21, stink bug22, cow35 infection61 Sepsis69
Phlebotomine sand fly23 Dacryocystitis70
1Harada, Oyaizu and Ishikawa (1996), 2Delalibera, Handelsman and Raffa (2005), 3Dillon and Charnley (1995), 4Sood and Nath (2002), 5Lauzon et al. (2009), 6Loncaric et al.(2009), 7Wells, Gitaitis and Sanders (2002), 8Vorwerk et al. (2007), 9Nadarasah and Stavrinides (2014), 10Maccollom et al. (2009), 11Medina, Nachappa and Tamborindeguy(2011), 12Maleki-Ravasan et al. (2014), 13Podgwaite et al. (2013), 14Demirci et al. (2013), 15Bell et al. (2007), 16Cooper, Nicholson and Puterka (2013), 17Terenius et al. (2012),18Esker and Nutter (2003), 19Stavrinides, No and Ochman (2010), 20Delalibera et al. (2007), 21Pinto-Tomas et al. (2009), 22Prado and Almeida (2009), 23Akhoundi et al. (2012),24Hong et al. (1993), 25Loch and Faisal (2007), 26Carbajal-Gonzalez et al. (2012), 27Volksch et al. (2009), 28Vieria et al. (2004), 29Hansen, Raa and Olafsen (1990), 30Sauteret al. (1987), 31Li et al. (2012), 32Cabassi et al. (2004), 33Muraschi, Friend and Bolles (1965), 34Kiskova et al. (2012), 35Verdier-Metz et al. (2012), 36Slotnick and Tulman (1967),37Cruz, Cazacu and Allen (2007), 38Gavini et al. (1989), 39Brady et al. (2010a,b), 40Brady et al. (2009a,b), 41Flatauer and Khan (1978), 42Vincent and Szabo (1988), 43Labianca
et al. (2013), 44Aly et al. (2008), 45Christakis et al. (2007), 46Lalas and Erichsen (2010), 47Bergman, Arends and Scholvinck (2007), 48Bicudo et al. (2007), 49Boszczowski et al.(2012), 50Borras et al. (2009), 51Ferrantino, Navaneethan and Sloand (2008), 52Lim et al. (2006), 53Cicchetti et al. (2006), 54Kratz et al. (2003), 55Ulloa-Gutierrez, Moya andAvila-Aguero (2004), 56Rodrigues et al. (2009), 57Fullerton, Lwin and Lal (2007), 58Goncalves et al. (2007), 59Shubov, Jagannathan and Chin-Hong (2011), 60Kursun et al.
(2012), 61Van Rostenberghe et al. (2006), 62Skorska et al. (2003), 63Milanowski (1994), 64Golec et al. (2004), 65Hischebeth et al. (2013), 66Manhoran et al. (2012), 67De Baere
et al. (2004), 68Schmid et al. (2003), 69Habsah et al. (2005), 70Zuberbuhler, Carifi and Leatherbarrow (2012).
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/39/6/968/547864 by guest on 26 March 2022

974 FEMS Microbiology Reviews, 2015, Vol. 39, No. 6
P. agglomerans, whereby the bacterium carries out essential ni-trogen cycling processes in the host gut, while the fly providesnutrition and a suitable habitat (Maccollom et al. 2009). Blueberrymaggot flies are naturally attracted to the degradation productsof uric acid, as well as the production of other volatile chemi-cals (Maccollom et al. 2009). Strains producing uricase are ableto break down uric acid, thus attracting the flies to their foodsource and facilitating the association between the symbionts(Robacker and Lauzon 2002; Maccollom et al. 2009).
Several Pantoea–insect associations identified appear to becommensalistic. The leaf-cutter ant, which cultivates its ownfungus gardens for food is an important player in nutrient cy-cling in forest environments (Wirth et al. 2003; Pinto-Tomaset al. 2009). Pantoea sp. At-9b was consistently isolated as oneof the nitrogen-fixing bacterial groups from the fungus gar-dens, with some isolates carrying a gene cluster that codes forthe molybdenum–iron nitrogen fixation pathway (Pinto-Tomaset al. 2009). The nitrogen fixation provided by Pantoea sp. At-9b and other microbes in these fungus gardens is thought toallow the leaf-cutter ants to use a wider variety of leaf mate-rial that has lower nitrogen concentrations (Pinto-Tomas et al.2009). Whether these bacteria are carried by and inoculated bythe ants, or whether they are naturally present in the soil is cur-rently unknown.
DEADLY PATHOGEN, HARMLESS COMMENSALOR VERSATILE OPPORTUNIST?
There is presently considerable debate as to the animalpathogenic capabilities of different Pantoea species groups.Strains of P. septica, P. calida, P. dispersa, P. ananatis, P. agglom-erans and P. eucalyptii have been isolated routinely from hu-man wounds, fractures, blood and other fluids, skin and sur-face swabs, stool, cysts and abscesses, as well as from swabs ofthe urethra, trachea and oropharynx (Table 3) (Slotnick and Tul-man 1967; Cruz et al. 2007; Coutinho and Venter 2009; Brady et al.2010a,b; Nadarasah and Stavrinides 2014). Pantoea strains havebeen identified from both immunocompetent and immunocom-promised patients, with patient age ranging from premature in-fants to seniors (Flatauer and Khan 1978; Vincent and Szabo1988; Kratz et al. 2003; Schmid et al. 2003; De Baere et al. 2004;Christakis et al. 2007; Aly et al. 2008). Introduction of the bacteriainto or onto the patient has been proposed to be through con-tamination ofmedical instruments and parenteral (intravenous)nutrition, inhalation of organic dust, wounds exposed to organicmaterial or perinatal in nature (Milanowski 1994; De Champset al. 2000; Skorska et al. 2003; Cicchetti et al. 2006; Van Ros-tenberghe et al. 2006; Bicudo et al. 2007; Cruz et al. 2007; Hsiehet al. 2007; Ferrantino, Navaneethan and Sloand 2008; Lee, Chungand Park 2010). A wide range of afflictions have been ascribed toPantoea, including septic arthritis, osteomyelitis, bacteremia andsepticemia, and peritonitis, among many others, although di-rect causation for most of these has not been demonstrated (Ta-ble 3) (Flatauer and Khan 1978; Vincent and Szabo 1988; De Baereet al. 2004; Lim et al. 2006; Bergman, Arends and Scholvinck 2007;Christakis et al. 2007; Aly et al. 2008; Ferrantino, Navaneethanand Sloand 2008; Borras et al. 2009; Lalas and Erichsen 2010; Labi-anca et al. 2013). Several neonatal outbreaks, however, resultedinmultiple deaths from septicemic shock and respiratory failure(Van Rostenberghe et al. 2006; Bergman, Arends and Scholvinck2007). Some work, however, has suggested that these and manyother clinical isolates that have been labeled as a strain of Pan-
toea are, in fact, misidentified (Rezzonico, Smits and Duffy 2012).One study demonstrated that clinical and animal isolates re-ported as members of Pantoea (and most frequently as P. agglom-erans) actually belong to other Pantoea species groups, and evenother genera, like Enterobacter (Rezzonico et al. 2009). In caseswhere clinical isolates are confirmed to be Pantoea, they belongto multiple species groups, including P. septica, P. calida, P. bren-neri, P. eucalyptii and P. agglomerans (Nadarasah and Stavrinides2014).
To try to understand the pathogenic potential of Pantoea iso-lates, and whether there is evidence for the evolution of hostassociation and/or host specialization among lineages, severalMLSA studies have been conducted using collections of vali-dated clinical and environmental Pantoea isolates to evaluatethe extent to which clinical and environmental isolates cluster(Brady et al. 2008, 2009a,b, 2010a,b; Deletoile et al. 2009; Rezzon-ico et al. 2009; Nadarasah and Stavrinides 2014). The separatephylogenetic clustering of clinical and environmental isolates—grouping into pathogenic and non-pathogenic groups—is con-sidered strongly supportive of lineage-specific host adaptation,as seen with other enterics like Escherichia coli (Georgiadesand Raoult 2011). In all phylogenetic studies carried out forPantoea, environmental and clinical isolates of many speciesgroups including P. agglomerans, P. ananatis and P. eucalyptii donot form distinctive clusters within each respective speciesgroup, but rather are intermingled (Brady et al. 2008, 2009a,b,2010a,b; Deletoile et al. 2009; Rezzonico et al. 2009; Nadarasahand Stavrinides 2014). This phylogenetic structure is generallysuggestive of isolates having an unknown capacity for host as-sociation, with clinical isolates possibly having the potential tocolonize plant hosts, and environmental isolates having the po-tential to colonize human hosts (Nadarasah and Stavrinides2014). In addition, a split decomposition analysis of P. agglom-erans has revealed substantive recombination between isolates(Deletoile et al. 2009), demonstrating a capacity for transfer ofgenetic determinants between individual isolates with differentcapabilities.
The evaluation of virulence potential of both plant and clin-ical Pantoea isolates has also been carried out using functionalhost assays. One study, which examined the virulence potentialof multiple species of Pantoea from environmental and clinicalsources using quantitative growth assays in maize, onion andfruit flies, showed that clinical isolates were able to grow in bothplant hosts comparably to environmental isolates (Nadarasahand Stavrinides 2014). There were no apparent growth or hostcolonization patterns that were unique to clinical isolates, andhost growth could not be predicted by phylogeny or source ofisolation (Nadarasah and Stavrinides 2014). A separate studyevaluated five clinical and five plant-associated P. agglomeransstrains quantitatively in both soybean and embryonated heneggs (Volksch et al. 2009). Both clinical and plant strains wereable to establish epiphytically on soybean plants, and there wasno difference in virulence between clinical and plant isolates inthe embryonated egg, suggesting that all P. agglomerans isolatesmight possess equal virulence potential (Volksch et al. 2009).Both of these studies indicate that the host-colonizing capabili-ties of Pantoea isolates remain unpredictable, with an unknownplant or animal host range/host-associating capacity for mostisolates. Notably, Pantoea strains are also found to be associatedwith terrestrial and aquatic animals, including birds, fish, inver-tebrates, bears and ruminants, which could be suggestive of alife history that involves animal hosts (Table 4) (Muraschi, Friendand Bolles 1965; Sauter et al. 1987; Hansen, Raa andOlafsen 1990;
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/39/6/968/547864 by guest on 26 March 2022

Walterson and Stavrinides 975
Table 4. Pantoea isolations from soil and water environments.
Species Soil Source Water Source Location
P. agglomerans Mangrove sludge1, 2 Aquifer Bangladesh10
Pond sediment3 Thermal spring Spain11
Iron-rich soil4 Drinking water South Africa12
High salt and pH soil5 River Chile13
P. ananatis Aquatic environment adjacent to agriculture Brazil14
P. anthophila Drinking water India15
P. dispersa Sub-alpine soil6, soil7
Pantoea sp. Olive mill waste8
Kerosene-rich soil9
1Liu et al. (2012), 2Yeung, Lee and Woodard (1998), 3Francis, Obraztsova and Tebo (2000), 4Sulbaran et al. (2009), 5Son et al. (2006), 6Selvakumar et al. (2008), 7Gavini et al.(1989), 8Pepi et al. (2010), 9Vasileva-Tonkova and Gesheva (2007), 10Sultana et al. (2011), 11Mosso et al. (1994), 12September et al. (2007), 13Escalante et al. (2009), 14Pileggiet al. (2012), 15Pindi, Yadav and Shanker (2013).
Cabassi et al. 2004; Vieria et al. 2004; Loch and Faisal 2007; Volkschet al. 2009; Carbajal-Gonzalez et al. 2012; Kiskova et al. 2012; Liet al. 2012; Verdier-Metz et al. 2012; Nadarasah and Stavrinides2014).
GENETICS AND GENOMICS
There are still considerable gaps in our knowledge of the spe-cific genetic determinants that allow Pantoea isolates to be a suc-cessful colonizer of both host and non-host environments. ForP. stewartii subsp. stewartii and its association with sweet corn,the T3SS (PSI-1) and the EPS, stewartan, appear to be the mainvirulence factors (Roper 2011). Association of P. stewartii subsp.stewartiiwith its flea beetle vector is facilitated by the T3SS, PSI-2. For P. agglomerans phytopathogenicity, the T3SS along withother virulence factors such as T3SEs are required, which arecarried on a plasmid (pPATH) that was suggested to have beenacquired horizontally fairly recently (Clark et al. 1993; Nizanet al. 1997; Valinsky et al. 1998; Ezra et al. 2004; Weinthal et al.2007). Aside from this and a few other select strains, the de-terminants used by most other Pantoea species for establish-ing in both host and non-host environments remains largelyunderexplored.
The availability of sequenced Pantoea genomes has allowedfor the discovery and comparison of genetic factors that maycontribute to the ability of certain isolates to thrive in dif-ferent environments. The genomes of several isolates of Pan-toea, such as P. agglomerans, P. stewartii, P. vagans and P. ana-natis, have been published, revealing genome sizes from 4.5 to6.3 MB and G+C contents of 52–55% (Smits et al. 2010, 2011;Choi et al. 2012; De Maayer et al. 2012b; Hong et al. 2012; Kimet al. 2012; Matsuzawa et al. 2012; Medrano and Bell 2012;Remus-Emsermann et al. 2013; Smith, Kirzinger and Stavrinides2013; Walterson, Smith and Stavrinides 2014). Although therehas been no broad-scale systematic comparative or evolution-ary genomic analysis of Pantoea as of yet, some common ge-nomic features identified inmore specific analyses include acyl-homoserine lactones and other quorum-sensing genes (Smitset al. 2010; Hong et al. 2012), plant growth-promoting genes (Kimet al. 2012), DNA repair genes (Remus-Emsermann et al. 2013),pathogenicity factors (De Maayer et al. 2010, 2012a), as well astype IV and VI secretion systems (Kim et al. 2012; Medrano andBell 2012).
A recent comparative genomic analysis of the type VI se-cretion system (T6SS) was conducted using sequenced Pantoeaand Erwinia genomes, which showed that one T6SS variant was
prevalent in Pantoea isolates from diverse environments (DeMaayer et al. 2011). One locus in particular, the T6SS-1 locus, con-tains two highly conserved core regions that alternate with vari-able regions containing hcp and vgrG, which code for secretedeffector proteins (De Maayer et al. 2011). The hcp and vgrG is-lands contain domains that are conserved across Pantoea strains,and show homology to genes with known roles in antibiosis,fungal cell wall degradation, and animal and plant pathogen-esis (De Maayer et al. 2011). However, the presence of these do-mains in isolates from diverse environments suggests that theremay be an innate capability for genetic versatility and adapt-ability. This adaptability is also reflected in the plasmids of Pan-toea. A comparative genomic study of at least 20 isolates led tothe identification of the Large Pantoea Plasmid family (LPP-1),which ranges from 280 to 789 kb, andwas found to be distributedamong 20 Pantoea isolates representing seven different species,including P. agglomerans, P. vagans, P. eucalyptii, P. anthophila, P.stewartii, P. ananatis and P. cypripedii (De Maayer et al. 2012a).Plasmid-encoded loci are linked to metabolism and transporta-tion of various sugars, carbohydrates, amino acids and organicacids, as well as the assimilation of iron and nitrogen, antibi-otic and heavy metal resistance, host colonization, pathogene-sis and antibiosis (De Maayer et al. 2012a). This illustrates theplasticity of this plasmid family, which may contribute to thediversity of capabilities among the isolates of different speciesgroups.
There are still considerable gaps in our understanding ofthe nature of those determinants that provide Pantoea withenvironmental versatility, and how these determinants maybe regulated by each respective environment. This avenueof research has the potential to advance our understand-ing of host association and the factors that may enable op-portunism or niche expansion in particular species groups.Likewise, genome-level analyses of Pantoea can provide addi-tional information on the biology and evolution of this group,although these have been limited by the availability of fullysequenced genomes spanning the diversity of the genus. Ge-nomic comparisons, while mostly limited to small groups ofisolates, have yielded important insight into the nature of vir-ulence, resistance and metabolic determinants that confer keysurvival and host-association capabilities. The availability of ad-ditional genomes will enable larger comparative genomic anal-yses that can identify the specific evolutionary processes thatare responsible for species-level diversification. Such studieswould also allow for the assessment of the pathogenic po-tential of individual Pantoea isolates given the current use of
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/39/6/968/547864 by guest on 26 March 2022

976 FEMS Microbiology Reviews, 2015, Vol. 39, No. 6
some isolates as biocontrol, bioremediation and therapeuticagents.
PANTOEA IN BIOTECHNOLOGY
Many strains of Pantoea show striking environmental versatil-ity and adaptability, and possess a variety of biosynthetic andbiodegradative capabilities that can be harnessed for potentiallyuseful applications in agricultural, environmental and clinicalsettings.
Biocontrol
Plants are susceptible to colonization by a variety of phy-topathogenic bacteria and fungi, yet epiphytic colonization byPantoea has been shown to decrease the incidence of plant dis-ease (Johnson et al. 1993, 2000; Johnson and Stockwell 1998).Many Pantoea strains have been shown to be strong environmen-tal competitors that produce a variety of natural products withantibiotic activity, such as pantocins, herbicolins, microcins andphenazines (Wright and Beer 1996, 2002; Vanneste et al. 2000,2002; Vanneste, Yu and Reglinski 2002; Giddens, Feng and Ma-hanty 2002; Wright et al. 2006; Vanneste, Yu and Cornish 2008;Smits et al. 2010, 2011), as well as the more recent PNP-1 nat-ural product from P. ananatis BRT175 that has inhibitory activ-ity against E. amylovora, and which is likely similar to FVG, or4-formylaminooxyvinylglycine (Halgren et al. 2013; Trippe et al.2013; Walterson, Smith and Stavrinides 2014). Because of the di-versity of natural products they produce, along with their abilityto persist in the environment, some Pantoea strains have beendeveloped into commercial biocontrol products, such as Blight-Ban C9-1 and Bloomtime Biological, which utilize P. vagans C9-1 and P. agglomerans E325, respectively (Johnson and Stockwell1998; Pusey 2002). These biological control agents are used forfruits, vegetables and other crop plants to control many typesof Gram-negative and Gram-positive bacteria, pathogenic fungiin the Ascomycota and Basidiomycota, pathogenic oomycetesand parasitic nematodes (Table 5). Penicillium digitatum, for ex-ample, is a green mold that is responsible for post-harvest de-cay of citrus fruits (Poppe, Vanhoutte and Hofte 2003). Pantoeaagglomerans CPA-2 when applied during post-harvest has shownto be effective against Penicillium sp. under certain environmen-tal conditions (Canamas et al. 2008; Zamani et al. 2009). Whilethe exact mechanism for the control of Penicillium sp. is stillunclear, it may involve a combination of antifungals and an-tagonism through competitive colonization. Alternatively, Pan-toea may provide protection by inducing plant systemic resis-tance (Munif, Hallmann and Sikora 2001). The production ofcertain compounds, such as 1,2,3-benzothiadiazole carbothioicacid S-methyl ester or harpin, induces plant defenses, as shownthrough the production of β-1,3-glucanases or peroxides (H2O2)(Zhang et al. 1998; Vanneste et al. 2002). Pantoea-induced en-hancement of plant defenses has been used to protect tomatofrom the nematode, Meloidogyne incognita (Munif, Hallmann andSikora 2001), cucumber from the fungus, Colletotrichum orbiculare(Zhang et al. 1998),Arabidopsis from Pseudomonas syringae pv.ma-culicola (Zhang et al. 1998), and kiwifruit and tobacco from Scle-rotinia sclerotiorum (Vanneste et al. 2002). M. incognita is knownto produce root knots on tomato plants (Munif, Hallmann andSikora 2001). Pantoea agglomerans MK-29 was shown to inducesystemic resistance of the tomato plant when used as a soildrench, resulting in a decrease in penetration of the plant rootsby juvenile nematodes, and thereby preventing the developmentof root knots (Munif, Hallmann and Sikora 2001).
Bioremediation
Isolates of Pantoea have been identified in a wide variety of pris-tine and contaminated aquatic and terrestrial environments (Ta-ble 5). In the arsenic-contaminated waters of Bangladesh andChile, Pantoea isolates show arsenic resistance and/or degrada-tion capabilities (Escalante et al. 2009; Sultana et al. 2011). InBrazil, a P. ananatis isolate has shown the ability to break downthe herbicide, mesotrione (Pileggi et al. 2012). Since this environ-ment is adjacent to key agricultural sites, the degradation of her-bicides from agricultural run-off is one important strategy forprotecting aquatic species. Other isolates from a petrochemicalwastewater treatment plant showed the ability to absorb, accu-mulate and tolerate significant levels of copper, chromium andcadmium (Ozdemir et al. 2004). Since the presence of heavymet-als in aquatic environments has been shown to be toxic and car-cinogenic (Ruiz-Manriquez et al. 1998), the use of heavy metal-tolerant Pantoea isolates for the removal of these compoundsfrom wastewaters is of great interest to industry, and of partic-ular environmental significance (Ozdemir et al. 2004).
Pantoea soil isolates have been shown to possess capabil-ities that include metal reduction (Francis, Obraztsova andTebo 2000), solubilization and degradation of organic materi-als (Dastager, Deepa and Pandey 2009; Sulbaran et al. 2009), bio-surfactant production (Vasileva-Tonkova and Gesheva 2007; Ja-cobucci, Oriani and Durrant 2009), iron-based respiration (Wuet al. 2010) and solubilization of insoluble inorganics (Son et al.2006). Petroleum, or crude oil, often poses a significant envi-ronmental danger to marine life and coast-line terrestrial biota,such as those in the Antarctic, because of its ability to persistat toxic levels in the environment (Vasileva-Tonkova and Ge-sheva 2007). An antarctic strain, Pantoea sp. A-13, produces gly-colipid biosurfactants when using the petroleum hydrocarbons,kerosene and n-paraffin as its sole carbon and energy source(Vasileva-Tonkova and Gesheva 2007). These biosurfactants, inturn, result in the emulsification and subsequent biodegrada-tion of the harmful petroleum hydrocarbons (Vasileva-Tonkovaand Gesheva 2007). The use of naturally occurring isolates withthe ability to degrade toxic petroleum hydrocarbons into lessharmful compounds is a promising alternative to currently usedchemical surfactants (Vasileva-Tonkova and Gesheva 2007).
Biosensors and indicators
Some of the more phenotypically distinct isolates of Pantoeaare being explored for their potential as biosensors. One P. ag-glomerans isolate was found to produce a blue pigment in atemperature-dependent manner (Fujikawa and Akimoto 2011).Pigment production was determined to be cell density depen-dent, with cell densities of 106–108 being required for maximalpigment production; however, biosynthesis was also tempera-ture dependent, with bacteria acquiring the blue coloration attemperatures ≥10oC, and retaining their characteristic yellowcoloration at temperatures ≤10oC (Fujikawa and Akimoto 2011).Modeling of blue pigment production yielded a robust descrip-tion of its accumulation and biosynthesis, with the capacity tobe developed into a time-temperature indicator for use in mon-itoring of possible spoilage of food and clinical products (Fu-jikawa and Akimoto 2011).
Therapeutics
Although Pantoea is currently considered problematic in theclinical environment, products derived from Pantoea have been
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/39/6/968/547864 by guest on 26 March 2022
Walterson and Stavrinides 977
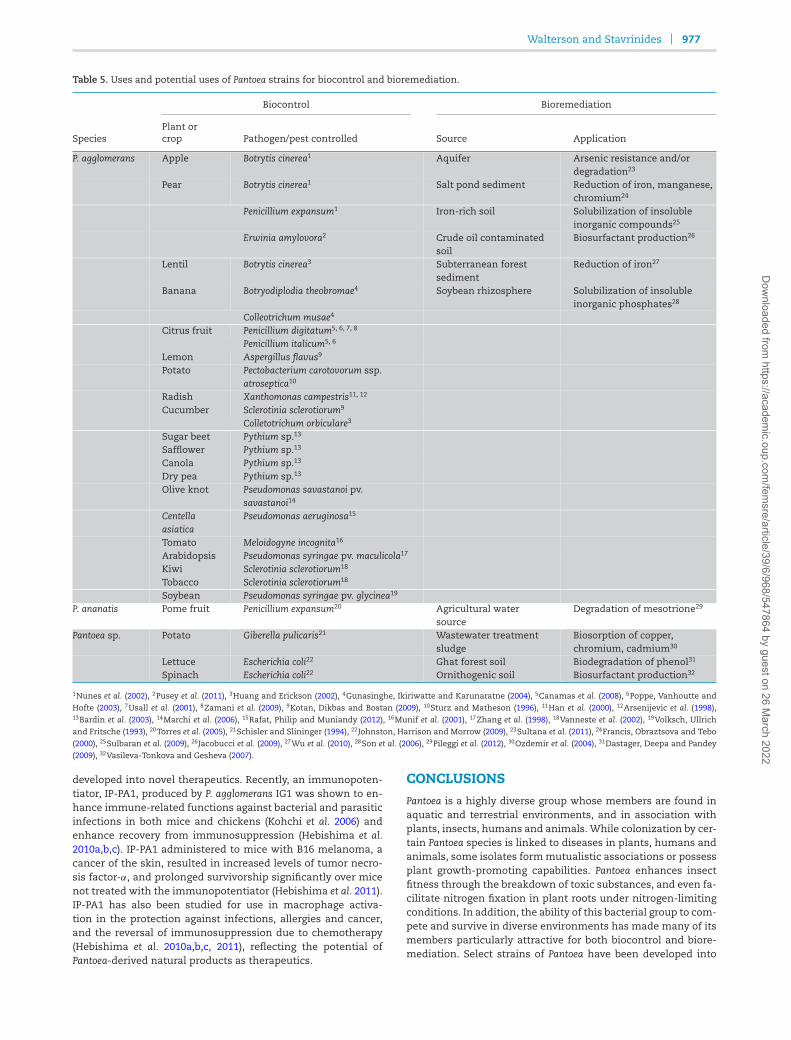
Table 5. Uses and potential uses of Pantoea strains for biocontrol and bioremediation.
Biocontrol Bioremediation
SpeciesPlant orcrop Pathogen/pest controlled Source Application
P. agglomerans Apple Botrytis cinerea1 Aquifer Arsenic resistance and/ordegradation23
Pear Botrytis cinerea1 Salt pond sediment Reduction of iron, manganese,chromium24
Penicillium expansum1 Iron-rich soil Solubilization of insolubleinorganic compounds25
Erwinia amylovora2 Crude oil contaminatedsoil
Biosurfactant production26
Lentil Botrytis cinerea3 Subterranean forestsediment
Reduction of iron27
Banana Botryodiplodia theobromae4 Soybean rhizosphere Solubilization of insolubleinorganic phosphates28
Colleotrichum musae4
Citrus fruit Penicillium digitatum5, 6, 7, 8
Penicillium italicum5, 6
Lemon Aspergillus flavus9
Potato Pectobacterium carotovorum ssp.atroseptica10
Radish Xanthomonas campestris11, 12
Cucumber Sclerotinia sclerotiorum9
Colletotrichum orbiculare3
Sugar beet Pythium sp.13
Safflower Pythium sp.13
Canola Pythium sp.13
Dry pea Pythium sp.13
Olive knot Pseudomonas savastanoi pv.savastanoi14
Centellaasiatica
Pseudomonas aeruginosa15
Tomato Meloidogyne incognita16
Arabidopsis Pseudomonas syringae pv. maculicola17
Kiwi Sclerotinia sclerotiorum18
Tobacco Sclerotinia sclerotiorum18
Soybean Pseudomonas syringae pv. glycinea19
P. ananatis Pome fruit Penicillium expansum20 Agricultural watersource
Degradation of mesotrione29
Pantoea sp. Potato Giberella pulicaris21 Wastewater treatmentsludge
Biosorption of copper,chromium, cadmium30
Lettuce Escherichia coli22 Ghat forest soil Biodegradation of phenol31
Spinach Escherichia coli22 Ornithogenic soil Biosurfactant production32
1Nunes et al. (2002), 2Pusey et al. (2011), 3Huang and Erickson (2002), 4Gunasinghe, Ikiriwatte and Karunaratne (2004), 5Canamas et al. (2008), 6Poppe, Vanhoutte and
Hofte (2003), 7Usall et al. (2001), 8Zamani et al. (2009), 9Kotan, Dikbas and Bostan (2009), 10Sturz and Matheson (1996), 11Han et al. (2000), 12Arsenijevic et al. (1998),13Bardin et al. (2003), 14Marchi et al. (2006), 15Rafat, Philip and Muniandy (2012), 16Munif et al. (2001), 17Zhang et al. (1998), 18Vanneste et al. (2002), 19Volksch, Ullrichand Fritsche (1993), 20Torres et al. (2005), 21Schisler and Slininger (1994), 22Johnston, Harrison and Morrow (2009), 23Sultana et al. (2011), 24Francis, Obraztsova and Tebo(2000), 25Sulbaran et al. (2009), 26Jacobucci et al. (2009), 27Wu et al. (2010), 28Son et al. (2006), 29Pileggi et al. (2012), 30Ozdemir et al. (2004), 31Dastager, Deepa and Pandey
(2009), 32Vasileva-Tonkova and Gesheva (2007).
developed into novel therapeutics. Recently, an immunopoten-tiator, IP-PA1, produced by P. agglomerans IG1 was shown to en-hance immune-related functions against bacterial and parasiticinfections in both mice and chickens (Kohchi et al. 2006) andenhance recovery from immunosuppression (Hebishima et al.2010a,b,c). IP-PA1 administered to mice with B16 melanoma, acancer of the skin, resulted in increased levels of tumor necro-sis factor-α, and prolonged survivorship significantly over micenot treated with the immunopotentiator (Hebishima et al. 2011).IP-PA1 has also been studied for use in macrophage activa-tion in the protection against infections, allergies and cancer,and the reversal of immunosuppression due to chemotherapy(Hebishima et al. 2010a,b,c, 2011), reflecting the potential ofPantoea-derived natural products as therapeutics.
CONCLUSIONS
Pantoea is a highly diverse group whose members are found inaquatic and terrestrial environments, and in association withplants, insects, humans and animals. While colonization by cer-tain Pantoea species is linked to diseases in plants, humans andanimals, some isolates formmutualistic associations or possessplant growth-promoting capabilities. Pantoea enhances insectfitness through the breakdown of toxic substances, and even fa-cilitate nitrogen fixation in plant roots under nitrogen-limitingconditions. In addition, the ability of this bacterial group to com-pete and survive in diverse environments has made many of itsmembers particularly attractive for both biocontrol and biore-mediation. Select strains of Pantoea have been developed into
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/39/6/968/547864 by guest on 26 March 2022

978 FEMS Microbiology Reviews, 2015, Vol. 39, No. 6
biocontrol agents, which prevent plant disease through the an-tagonism of pathogens, and in some cases, through the induc-tion of plant systemic defenses. Additionally, Pantoea has uniquebiodegradative capabilities, including metabolic pathways thatdegrade herbicides and other toxic compounds, providingopportunity for the development and commercialization ofuseful products; however, there is still much uncertainty sur-rounding the host-associating and pathogenic capabilities of in-dividual strains. The development of Panotea as bioagents mustbe considered carefully, given that there are no known pathogenbiomarkers available, and that clinical and environmental iso-lates are phylogenetically indistinguishable in all published phy-logenetic trees (Brady et al. 2008, 2009a,b, 2010a,b; Deletoile et al.2009; Rezzonico et al. 2009; Nadarasah and Stavrinides 2014). Ul-timately, there remains a need to identify those genetic deter-minants that enable niche-specific colonization, including anyfactors that may determine host-colonizing capacity and hostspecificity. Determining the nature of these genetic factors re-mains a promising research direction, andwill undoubtedly helpto unravel the full capabilities of this versatile, broad-niche bac-terial group.
FUNDING
This work was supported by Discovery Grants from the Natu-ral Sciences and Engineering Council (#386654), and a CanadaFoundation for Innovation LOF (#28591).
Conflict of interest. None declared.
REFERENCES
Akhoundi M, Bakhtiari R, Guillard T, et al. Diversity of the bac-terial and fungal microflora from the midgut and cuticleof phlebotomine sand flies collected in North-Western Iran.PLoS One 2012;7:e50259, DOI: 10.1371/journal.pone.0050259.
Aly N, Salmeen H, Lila R, et al. Pantoea agglomerans blood-stream infection in preterm neonates.Med Prin Pract 2008;17:500–3.
Arsenijevic M, Obradovic A, Stevanovic D, et al. Antagonistic ef-fect of some saprophytic bacteria to Pseudomonas syringae pv.phaseolicola and Xanthomonas campestris pv. phaseoli. BullOILB/SROP 1998;21:297–300.
Asis C, Adachi K. Isolation of endophytic diazotroph Pan-toea agglomerans and nondiazotroph Enterobacter asburiaefrom sweetpotato stem in Japan. Lett Appl Microbiol 2004;38:19–23.
Azad H, Holmes G, Cooksey D. A new leaf blotch disease of su-dangrass caused by Pantoea ananas and Pantoea stewartii. PlantDis 2000;84:973–9.
Bardin S, Huang H, Liu L, et al. Control, by microbial seed treat-ment, of damping-off caused by Pythium sp on canola, saf-flower, dry pea, and sugar beet. Can J Plant Pathol 2003;25:268–75.
Beattie GA, Lindow SE. Bacterial colonization of leaves: a spec-trum of strategies. Phytopathology 1999;89:353–9.
Beijerinck MW. Die Bakterien der Papilionaceenknollchen.Botanische Zeitung 1888;46:725–804.
Beji A, Mergaert J, Gavini F, et al. Subjective synonymy of Erwiniaherbicola, Erwinia milletiae, and Enterobacter agglomerans andredefinition of the taxon by genotypic and phenotypic data.Int J Syst Bacteriol 1988;38:77–88.
Bell A, Medrano E, Lopez J, et al. Transmission and importanceof Pantoea ananatis during feeding on cotton buds (Gossyp-
ium hirsutum L.) by cotton fleahoppers (Pseudatomoscelis se-riatus Reuter). Proceedings of World Cotton Research Confer-ence. Lubbock, TX, 2007 (http://wcrc.confex.com/wcrc/2007/techprogram/P1835.htm).
Bergman K, Arends J, Scholvinck E. Pantoea agglomerans sep-ticemia in three newborn infants. Pediatr Infect Dis J2007;26:453–4.
Bicudo E,Macedo V, CarraraM, et al.Nosocomial outbreak of Pan-toea agglomerans in a pediatric urgent care center. Braz J InfectDis 2007;11:281–4.
Borras M, Roig J, Garcia M, et al. Adverse effects of Pan-toea peritonitis on peritoneal transport. Periton Dialysis Int2009;29:234–5.
Boszczowski I, Almeida J, de Miranda E, et al. Nosocomial out-break of Pantoea agglomerans bacteraemia associated withcontaminated anticoagulant citrate dextrose solution: newname, old bug? J Hosp Infect 2012;80:255–8.
Brady C, Cleenwerck I, van derWesthuizen L, et al. Pantoea rodasiisp. nov., Pantoea rwandensis sp. nov. and Pantoea wallisii sp.nov., three novel Pantoea species isolated from Eucalyptus.Int J Syst Evol Micr 2012;62:1457–64.
Brady C, Cleenwerck I, Venter S, et al. Phylogeny and identi-fication of Pantoea species associated with plants, humansand the natural environment based on multilocus sequenceanalysis (MLSA). Syst Appl Microbiol 2008;31:447–60.
Brady C, Cleenwerck I, Venter S, et al. Emended descriptionof the genus Pantoea, description of four species from hu-man clinical samples, Pantoea septica sp. nov., Pantoea euc-rina sp. nov., Pantoea brenneri sp. nov. and Pantoea conspicuasp. nov., and transfer of Pectobacterium cypripedii (Hori 1911)Brenner et al. 1973 emend. Hauben et al. 1998 to the genusas Pantoea cypripedii comb. nov. Int J Syst Evol Micr 2010a;60:2430–40.
Brady C, Goszczynska T, Venter S, et al. Pantoea allii sp. nov.,isolated from onion plants and seed. Int J Syst Evol Micr2011;61:932–7.
Brady C, Venter S, Cleenwerck I, et al. Isolation of Enterobac-ter cowanii from Eucalyptus showing symptoms of bac-terial blight and dieback in Uruguay. Lett Appl Microbiol2009a;49:461–5.
Brady C, Venter S, Cleenwerck I, et al. Pantoea vagans sp nov., Pan-toea eucalypti sp nov., Pantoea deleyi sp nov and Pantoea an-thophila sp nov. Int J Syst Evol Micr 2009b;59:2339–45.
Brady CL, Venter SN, Cleenwerck I, et al. Transfer of Pantoea cit-rea, Pantoea punctata and Pantoea terrea to the genus Tatumellaemend. as Tatumella citrea comb. nov., Tatumella punctatacomb. nov and Tatumella terrea comb. nov and descriptionof Tatumella morbirosei sp nov. Int J Syst Evol Micr 2010b;60:484–94.
Brandl M, Clark EM, Lindow S. Characterization of the indolea-acetic acid (IAA) biosynthetic pathway in an epiphytic strainof Erwinia herbicola and IAA production in vitro. Can J Microbiol1996;42:586–92.
Brandl M, Lindow S. Contribution of indole-3-acetic acid produc-tion to the epiphytic fitness of Erwinia herbicola. Appl EnvironMicrob 1998;64:3256–63.
Burr T, Katz B, Abawi G, et al. Comparison of tumorigenic strainsof Erwinia herbicola isolated from table beet with E. h. gyp-sophilae. Plant Dis 1991;75:855–8.
Cabassi C, Taddei S, Predari G, et al. Bacteriologic findings inostrich (Struthio camelus) eggs from farms with reproductivefailures. Avian Dis 2004;48:716–22.
Canamas T, Vinas I, Usall J, et al. Control of postharvestdiseases on citrus fruit by preharvest application of the
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/39/6/968/547864 by guest on 26 March 2022

Walterson and Stavrinides 979
biocontrol agent Pantoea agglomerans CPA-2 part I. Study ofdifferent formulation strategies to improve survival of cellsin unfavourable environmental conditions. Postharvest BiolTec 2008;49:86–95.
Carbajal-Gonzalez M, Fregeneda-Grandes J, Suarez-Ramos S,et al. Bacterial skin flora variation and in vitro inhibitory activ-ity against Saprolegnia parasitica in brown and rainbow trout.Dis Aquat Organ 2012;96:125–35.
Choi O, Lim J, Seo Y, et al. Complete genome sequence ofthe rice pathogen Pantoea ananatis strain PA13. J Bacteriol2012;194:531.
Christakis G, Perlorentzou S, Aslanidou M, et al. Bacteremiacaused by Pantoea agglomerans and Enterococcus faecalis in apatient with colon cancer. J BUON 2007;12:287–90.
Cicchetti R, Iacobini M, Midulla F, et al. Pantoea agglomerans sep-sis after rotavirus gastroenteritis. Pediatr Infect Dis J 2006;25:280–1.
Clark E, Manulis S, Ophir Y, et al. Cloning and characterizationof iaaM and iaaH from Erwinia herbicola pathovar gypsophilae.Phytopathology 1993;83:234–40.
Cooksey DA. Galls of Gypsophila paniculata caused by Erwinia her-bicola. Plant Dis 1986;70:464–8.
Cooper W, Nicholson S, Puterka G. Salivary proteins of Lygushesperus (Hemiptera: Miridae). Ann Entomol Soc Am 2013;106:86–92.
Coplin D, Majerczak D, Zhang Y, et al. Identification of Pantoeastewartii subsp stewartii by PCR and strain differentiation byPFGE. Plant Dis 2002;86:304–11.
Correa V, Majerczak D, Ammar E, et al. Pantoea stewartii subspstewartii carries two type III secretion systems required foradaptation to insect vector and plant hosts. Phytopathology2010;100:S184.
Correa V, Majerczak D, Ammar E, et al. The bacterium Pantoeastewartii uses two different type III secretion systems to col-onize its plant host and insect vector. Appl Environ Microb2012;78:6327–36.
Cother E, Reinke R, McKenzie C, et al. An unusual stem necro-sis of rice caused by Pantoea ananas and the first recordof this pathogen on rice in Australia. Australas Plant Path2004;33:495–503.
Coutinho T, Preisig O, Mergaert J, et al. Bacterial blight anddieback of Eucalyptus species, hybrids, and clones in SouthAfrica. Plant Dis 2002;86:20–5.
Coutinho TA, Venter SN. Pantoea ananatis: an unconventionalplant pathogen. Mol Plant Pathol 2009;10:325–35.
Cruz A, Cazacu A, Allen C. Pantoea agglomerans, a plantpathogen causing human disease. J Clin Microbiol 2007;45:1989–92.
Dastager S, Deepa C, Pandey A. Isolation and characterization ofhigh-strength phenol-degrading novel bacterium of the Pan-toea genus. Bioremed J 2009;13:171–9.
De Baere T, Verhelst R, Labit C, et al. Bacteremic infection withPantoea ananatis. J Clin Microbiol 2004;42:4393–5.
De Champs C, Le Seaux S, Dubost J, et al. Isolation of Pan-toea agglomerans in two cases of septic monoarthritis afterplant thorn and wood sliver injuries. J Clin Microbiol 2000;38:460–1.
De Maayer P, Chan W, Blom J, et al. The large universal Pantoeaplasmid LPP-1 plays a major role in biological and ecologicaldiversification. BMC Genomics 2012a;13:625.
De Maayer P, Chan W, Rezzonico F, et al. Complete genome se-quence of clinical isolate Pantoea ananatis LMG 5342. J Bacteriol2012b;194:1615–6.
De Maayer P, Chan W, Venter S, et al. Genome sequence of Pan-toea ananatis LMG20103, the causative agent of Eucalyptusblight and dieback. J Bacteriol 2010;192:2936–7.
De Maayer P, Venter S, Kamber T, et al. Comparative genomics ofthe type VI secretion systems of Pantoea and Erwinia speciesreveals the presence of putative effector islands that maybe translocated by the VgrG and Hcp proteins. BMC Genomics2011;12:576.
Delalibera I, Handelsman J, Raffa K. Contrasts in cellulolyticactivities of gut microorganisms between the wood borer,Saperda vestita (Coleoptera : Cerambycidae), and the bark bee-tles, Ips pini andDendroctonus frontalis (Coleoptera : Curculion-idae). Environ Entomol 2005;34:541–7.
Delalibera I, Vasanthakumar A, Burwitz B, et al. Compositionof the bacterial community in the gut of the pine en-graver, Ips pini (Say, Coleoptera) colonizing red pine. Symbiosis2007;43:97–104.
Deletoile A, Decre D, Courant S, et al. Phylogeny and identi-fication of Pantoea species and typing of Pantoea agglomer-ans strains by multilocus gene sequencing. J Clin Microbiol2009;47:300–10.
Demirci M, Sevim E, Demir I, et al. Culturable bacterial micro-biota of Plagiodera versicolora (L., Coleoptera: Chrysomelidae)and virulence of the isolated strains. Folia Microbiol 2013;58:201–10.
Dillon R, Charnley A. Chemical barriers to gut infection in thedesert locust—in vivo production of antimicrobial phenolsassociatedwith the bacterium Pantoea agglomerans. J InvertebrPathol 1995;66:72–5.
Elvira-Recuenco M, van Vuurde J. Natural incidence of endo-phytic bacteria in pea cultivars under field conditions. CanJ Microbiol 2000;46:1036–41.
Escalante G, Campos V, Valenzuela C, et al. Arsenic resistantbacteria isolated from arsenic contaminated river in the At-acama Desert (Chile). Bull Environ Contam Toxicol 2009;83:657–61.
Esker P, Nutter F. Temporal dynamics of corn flea beetle popula-tions infestedwith Pantoea stewartii, causal agent of Stewart’sdisease of corn. Phytopathology 2003;93:210–8.
Ewing W, Fife M. Enterobacter agglomerans (Beijerinck) comb. nov.(the Herbicola-Lathyri Bacteria). Int J Syst Bacteriol 1972;22:4–11.
Ezra D, Barash I, Weinthal D, et al. pthG from Pantoea agglomer-ans pv. gypsophilae encodes an avirulence effector that de-termines incompatibility in multiple beet species. Mol PlantPathol 2004;5:105–13.
Ferrantino M, Navaneethan S, Sloand J. Pantoea agglomer-ans: an unusual inciting agent in peritonitis. Perit Dial Int2008;28:428–30.
Flatauer F, Khan M. Septic arthritis caused by Enterobacter ag-glomerans. Arch Intern Med 1978;138:788.
Francis C, Obraztsova A, Tebo B. Dissimilatory metal reductionby the facultative anaerobe Pantoea agglomerans SP1. Appl En-viron Microb 2000;66:543–8.
Frederick R, Ahmad M, Majerczak D, et al. Genetic organizationof the Pantoea stewartii subsp stewartii hrp gene cluster andsequence analysis of the hrpA, hrpC, hrpN, andwtsE operons.Mol Plant Microbe In 2001;14:1213–22.
Fucikovsky L, Aranda S. Pantoea ananas a new pathogen of agavein Mexico. Phytopathology 2006;96:S37.
Fujikawa H, Akimoto R. New blue pigment produced by Pan-toea agglomerans and its production characteristics at varioustemperatures. Appl Environ Microb 2011;77:172–8.
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/39/6/968/547864 by guest on 26 March 2022

980 FEMS Microbiology Reviews, 2015, Vol. 39, No. 6
Fullerton D, Lwin A, Lal S. Pantoea agglomerans liver abscess pre-senting with a painful thigh. Eur J Gastroen Hepat 2007;19:433–5.
Gagne-Bourgue F, Aliferis K, Seguin P, et al. Isolation and charac-terization of indigenous endophytic bacteria associated withleaves of switchgrass (Panicum virgatum L.) cultivars. J ApplMi-crobiol 2013;114:836–53.
Galan J, Collmer A. Type III secretionmachines: Bacterial devicesfor protein delivery into host cells. Science 1999;284:1322–8.
Gavini F, Mergaert J, Beji A, et al. Transfer of Enterobacter agglomer-ans (Beijerinck 1888) Ewing and Fife 1972 to Pantoea gen-novas Pantoea agglomerans comb nov and description of Pantoeadispersa sp-nov. Int J of Syst Bacteriol 1989;39:337–45.
Georgiades K, Raoult D. Defining pathogenic bacterial species inthe genomic era. Front Microbiol 2011;1:151.
Giddens S, Feng Y, Mahanty H. Characterization of a novelphenazine antibiotic gene cluster in Erwinia herbicola Eh1087.Mol Microbiol 2002;45:769–83.
Gitaitis R, Gay J. First report of a leaf blight, seed stalk rot, andbulb decay of onion by Pantoea ananas in Georgia. Plant Dis1997;81:1096.
Golec M, Skorska C, Mackiewicz B, et al. Immunologic reactivityto work-related airborne allergens in people occupationallyexposed to dust from herbs. Ann Agr Env Med 2004;11:121–7.
Goncalves M, Coutinho W, Pimenta F, et al. Periodontal diseaseas reservoir for multi-resistant and hydrolytic enterobacte-rial species. Lett Appl Microbiol 2007;44:488–94.
Guanlin X. First report of Palea browning in China and charac-terization of the causal organism by phenotypic tests and bi-olog. Int Rice Res Notes 2001;26:25–6.
Gunasinghe R, Ikiriwatte C, Karunaratne A. The use of Pan-toea agglomerans and Flavobacterium sp to control bananapathogens. J Hortic Sci Biotech 2004;79:1002–6.
Habsah H, Zeehaida M, VanRostenberghe H, et al. An outbreakof Pantoea spp. in a neonatal intensive care unit secondaryto contaminated parenteral nutrition. J Hosp Infect 2005;61:213–8.
Halgren A,MaselkoM, AzevedoM, et al.Genetics of germination-arrest factor (GAF) production by Pseudomonas fluorescensWH6: identification of a gene cluster essential for GAFbiosynthesis. Microbiol-SGM 2013;159:36–45.
Ham J, Majerczak D, Ewert S, et al. WtsE, an AvrE-family typeIII effector protein of Pantoea stewartii subsp stewartii, causescell death in non-host plants. Mol Plant Pathol 2008;9:633–43.
Han D, Coplin D, Bauer W, et al. A rapid bioassay for screeningrhizosphere microorganisms for their ability to induce sys-temic resistance. Phytopathology 2000;90:327–32.
Hansen G, Raa J, Olafsen J. Isolation of Enterobacter agglomere-ans fromDolphin Fish, Coryphaena hippurus. J Fish Dis 1990;13:93–6.
Harada H, Oyaizu H, Ishikawa H. A consideration about the ori-gin of aphid intracellular symbiont in connection with gutbacterial flora. J Gen Appl Microbiol 1996;42:17–26.
Hebishima T, Matsumoto Y, Soma G, et al. Immune recov-ery effects of immunopotentiator from Pantoea agglomer-ans 1 (IP-PA1) on low antibody productions in response toSalmonella enteritidis vaccine and sheep red blood cells indexamethasone-treated stressed chicken models. J Vet MedSci 2010a;72:435–42.
Hebishima T, Matsumoto Y, Watanabe G, et al. Protective ef-fects of the immunopotentiator from Pantoea agglomerans 1on chemotherapeutic agent-inducedmacrophage growth in-hibition. Anticancer Res 2010b;30:2033–40.
Hebishima T, Matsumoto Y, Watanabe G, et al. Recovery fromimmunosuppression-related disorders in humans and ani-mals by IP-PA1, an edible lipopolysaccharide. Anticancer Res2010c;30:3113–8.
Hebishima T, Matsumoto Y, Watanabe G, et al. Oral admin-istration of immunopotentiator from Pantoea agglomerans 1(IP-PA1) improves the survival of B16 melanoma-inoculatedmodel mice. Exp Anim 2011;60:101–9.
Hischebeth G, Kohlhof H, Wimmer M, et al. Detection of Pan-toea agglomerans in hip prosthetic infection by sonication ofthe removed prosthesis: the first reported case. Technol HealthCare 2013;21:613–8.
Hong CB, Donahue JM, Giles RC, et al. Etiology and pathology ofequine placentitis. J Vet Diagn Invest 1993;5:56–63.
Hong K, Gan H, Low S, et al. Draft genome sequence of Pantoeasp strain A4, a rafflesia-associated bacterium that producesN-acylhomoserine lactones as quorum-sensing molecules. JBacteriol 2012;194:6610.
Hsieh C, Liang J, Peng S, et al. Seizure associated with total par-enteral nutrition-related hypermanganesemia. Pediatr Neurol2007;36:181–3.
Huang HC, Erickson RS. Biological control of botrytis stem andblossom blight of lentil. Plant Pathol Bull 2002;11:7–14.
Jacobucci D, Oriani M, Durrant L. Reducing COD level on oilyeffluent by utilizing biosurfactant-producing bacteria. BrazArch Biol Techn 2009;52:1037–42.
Janisiewicz WJ, Jurick WM, Vico I, et al. Culturable bacteria fromplum fruit surfaces and their potential for controlling brownrot after harvest. Postharvest Biol Tec 2013;76:145–51.
Johnson KB, Stockwell VO. Management of fire blight: a casestudy in microbial ecology. Annu Rev Phytopathol 1998;36:227–48.
Johnson KB, Stockwell VO, McLaughlin RJ, et al. Effect of antag-onistic bacteria on establishment of honey bee-dispersed Er-winia amylovora in pear blossoms and on fire blight control.Phytopathology 1993;83:995–1002.
Johnson KB, Stockwell VO, Sawyer TL, et al. Assessment of envi-ronmental factors influencing growth and spread of Pantoeaagglomerans on and among blossoms of pear and apple. Phy-topathology 2000;90:1285–94.
Johnston M, Harrison M, Morrow R. Microbial antagonists of Es-cherichia coli O157:H7 on fresh-cut lettuce and spinach. J FoodProt 2009;72:1569–75.
Kageyama B, Nakae M, Yagi S, et al. Pantoea punctata sp nov, Pan-toea citrea sp nov, and Pantoea terrea sp nov isolated from fruitand soil samples. Int J Syst Bacteriol 1992;42:203–10.
Khetmalas M, Bal A, Noble L, et al. Pantoea agglomerans is the eti-ological agent for black spot necrosis on beach peas. Can JMicrobiol 1996;42:1252–7.
Kido K, Adachi R, Hasegawa M, et al. Internal fruit rot of nettedmelon caused by Pantoea ananatis (=Erwinia ananas) in Japan.J Gen Plant Pathol 2008;74:302–12.
Kim H, Lee J, Kang B, et al. Draft genome sequence of Pantoeaananatis B1-9, a nonpathogenic plant growth-promoting bac-terium. J Bacteriol 2012;194:729–9.
Kirzinger MWB, Stavrinides J. Host specificity determinants as agenetic continuum. Trends Microbiol 2012;20:88–93.
Kiskova J, Hrehova Z, Janiga M, et al. Bacterial prevalence inthe Dunnock (Prunella modularis) in sub-alpine habitats ofthe Western Carpathians, Slovak Republic. Ornis Fennica2012;89:34–43.
Kohchi C, Inagawa H, Nishizawa T, et al. Applications oflipopolysaccharide derived from Pantoea agglomerans (IP-PA1)
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/39/6/968/547864 by guest on 26 March 2022

Walterson and Stavrinides 981
for health care based on macrophage network theory. J BiosciBioeng 2006;102:485–96.
Kotan R, Dikbas N, Bostan H. Biological control of post harvestdisease caused by Aspergillus flavus on stored lemon fruits.Afr J Biotechnol 2009;8:209–14.
KoutsoudisM, Tsaltas D, Minogue T, et al.Quorum-sensing regu-lation governs bacterial adhesion, biofilm development, andhost colonization in Pantoea stewartii subspecies stewartii. PNatl Acad Sci USA 2006;103:5983–8.
Kratz A, Greenberg D, Barki Y, et al. Pantoea agglomerans asa cause of septic arthritis after palm tree thorn injury;case report and literature review. Arch Dis Child 2003;88:542–4.
Kursun O, Unal N, Cesur S, et al. A case of ventilator-associatedpneumoniae due to Pantoea agglomerans. Mikrobiyol Bul2012;46:295–8.
Labianca L, Montanaro A, Turturro F, et al. Osteomyelitis causedby Pantoea agglomerans in a closed fracture in a child. Ortho-pedics 2013;36:E252–6.
Lalas K, Erichsen D. Sporadic Pantoea agglomerans bacteremia ina near-term female: case report and review of literature. JpnJ Infect Dis 2010;63:290–1.
Laskar F, Nevita T, Sharma G. Isolation and identification of en-dophytes from different cultivars of rice (Oryza sativa L.) un-der wetland and upland conditions in South Assam. J PureAppl Microbiol 2012;6:357–62.
Lauzon C, McCombs S, Potter S, et al. Establishment and ver-tical passage of Enterobacter (Pantoea) agglomerans and Kleb-siella pneumoniae through all life stages of the MediterraneanFruit Fly (Diptera: Tephritidae). Ann Entomol Soc Am 2009;102:85–95.
Lee N, Chung I, Park J. A case of Pantoea endophthalmitis. KoreanJ Opthalmol 2010;24:318–21.
Leigh JA, Coplin DL. Exopolysaccharides in plant-bacterial inter-actions. Annu Rev Microbiol 1992;46:307–46.
Li D, Chen L, Wang C, et al. Aerobic bacterial flora of nasal cavityof seven giant pandas (Ailuropoda melanoleuca). J Anim Vet Adv2012;11:3008–10.
Lim P, Chen S, Tsai C, et al. Pantoea peritonitis in a patient re-ceiving chronic ambulatory peritoneal dialysis. Nephrology2006;11:97–9.
Lindow SE, Brandl MT. Microbiology of the phyllosphere. ApplEnviron Microb 2003;69:1875–83.
Liu H, Wang G, Zhu D, et al. Improvement of hydrogen produc-tion by transposon-mutagenized strain of Pantoea agglomer-ans BH18. Int J Hydrogen Energ 2012;37:8282–7.
Liu Y, Wang S, Zhang D, et al. Pantoea beijingensis sp nov., iso-lated from the fruiting body of Pleurotus eryngii. Anton Leeuw2013;104:1039–47.
Loch T, Faisal M. Isolation of Pantoea agglomerans from browntrout (Salmo trutta) from Gilchrist Creek, Michigan, USA. BullEur Assn Fish P 2007;27:200–4.
Loiret F, Ortega E, Kleiner D, et al. A putative new endophyticnitrogen-fixing bacterium Pantoea sp from sugarcane. J ApplMicrobiol 2004;97:504–11.
Loncaric I, Heigl H, Licek E, et al. Typing of Pantoea agglom-erans isolated from colonies of honey bees (Apis mellifera)and culturability of selected strains from honey. Apidologie2009;40:40–54.
Maccollom G, Lauzon C, Sjogren R, et al. Association and at-traction of blueberry maggot fly curran (Diptera: Tephri-tidae) to Pantoea (Enterobacter) agglomerans. Environ Entomol2009;38:116–20.
McInroy J, Kloepper J. Survey of indigenous bacterial endophytesfrom cotton and sweet corn. Plant Soil 1995;173:337–42.
Magnani G, Cruz L, Weber H, et al. Culture-independent analysisof endophytic bacterial communities associated with Brazil-ian sugarcane. Genet Mol Res 2013;12:4549–58.
Maleki-Ravasan N, Oshaghi M, Hajikhani S, et al. Aerobic micro-bial community of insectary population of Phlebotomus pap-atasi. J Arthropod Borne Dis 2014;8:69–81.
Manhoran G, Lalitha P, Jeganathan L, et al. Pantoea ananatis as acause of corneal infiltrate after rice husk injury—a case re-port. J Clin Microbiol 2012;50:2163–4.
Marchi G, Sisto A, Cimmino A, et al. Interaction between Pseu-domonas savastanoi pv. savastanoi and Pantoea agglomerans inolive knots. Plant Pathol 2006;55:614–24.
Matsuzawa T, Mori K, Kadowaki T, et al. Genome sequence ofPantoea agglomerans strain IG1. J Bacteriol 2012;194:1258–9.
Medina R, Nachappa P, Tamborindeguy C. Differences in bacte-rial diversity of host-associated populations of Phylloxera no-tabilis Pergande (Hemiptera: Phylloxeridae) in pecan and wa-ter hickory. J Evol Biol 2011;24:761–71.
Medrano E, Bell A. Role of Pantoea agglomerans in opportunis-tic bacterial seed and boll rot of cotton (Gossypium hirsutum)grown in the field. J Appl Microbiol 2007;102:134–43.
Medrano E, Bell A. Genome sequence of Pantoea sp strain Sc1, an opportunistic cotton pathogen. J Bacteriol 2012;194:3019–9.
Menclas B, Block CC, Esker PD, et al. Quantifying the feeding pe-riods required by corn flea beetles to acquire and transmitPantoea stewartii. Plant Dis 2006;90:319–24.
Mergaert J, Verdonck L, Kersters K. Transfer of Erwinia ananas(synonym, Erwinia uredovora) and Erwinia stewartii to thegenus Pantoea emend as Pantoea ananas (Serrano 1928) comb-nov and Pantoea stewartii (Smith 1898) comb-nov, respec-tively, and description of Pantoea stewartii subsp. indologenessubsp. nov. Int J Syst Bacteriol 1993;43:162–73.
Merighi M, Majerczak D, Coplin D. A novel transcriptional au-toregulatory loop enhances expression of the Pantoea stew-artii subsp stewartii Hrp type III secretion system. FEMS Mi-crobiol Lett 2005;243:479–87.
Merighi M, Majerczak D, Stover E, et al. The hrpX/hrpY two-component system activates hrpS expression, the first stepin the regulatory cascade controlling the hrp regulon in Pan-toea stewartii subsp stewartii. Mol Plant Microbe In 2003;16:238–48.
Merighi M, Majerczak D, Zianni M, et al. Molecular characteriza-tion of Pantoea stewartii subsp. stewartii HrpY, a conserved re-sponse regulator of the Hrp type III secretion system, and itsinteraction with the hrpS promoter. J Bacteriol 2006;188:5089–100.
Milanowski J. Effects of Pantoea agglomerans on the respiratorysystem. Part II. Studies in vivo. Ann Agr Env Med 1994;1:52–6.
Minogue T, Carlier A, Koutsoudis M, et al. The cell density-dependent expression of stewartan exopolysaccharide inPantoea stewartii ssp stewartii is a function of EsaR-mediatedrepression of the rcsA gene. Mol Microbiol 2005;56:189–203.
Minogue T, Wehland-von Trebra M, Bernhard F, et al. The au-toregulatory role of EsaR, a quorum-sensing regulator in Pan-toea stewartii ssp stewartii: evidence for a repressor function.Mol Microbiol 2002;44:1625–35.
Mohammadi M, Burbank L, Roper C. The biological role ofpigment production for the bacterial phytopathogen, Pan-toea stewartii subsp. stewartii. Appl Environ Microb 2012;78:6859–65.
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/39/6/968/547864 by guest on 26 March 2022

982 FEMS Microbiology Reviews, 2015, Vol. 39, No. 6
Mosso M, Delarosa M, Vivar C, et al. Heterotrophic bacterial pop-ulations in the mineral waters of thermal springs in Spain. JAppl Bacteriol 1994;77:370–81.
Munif A, Hallmann J, Sikora R. Induced systemic resistance ofselected endophytic bacteria againstMeloidogyne incognita ontomato. Meded Rijksuniv Gent Fak Landbowkd Toegep Biol Wet2001;66:663–9.
Muraschi T, Friend M, Bolles D. Erwinia-like microorganismsisolated from animal and human hosts. Appl Microbiol1965;13:128–31.
Nadarasah G, Stavrinides J. Quantitative evaluation of the host-colonizing capabilities of the enteric bacterium Pantoea usingplant and insect hosts. Microbiology 2014;160:602–15.
Nadha H, Salwan R, Kasana R, et al. Identification and elimina-tion of bacterial contamination during in vitro propagation ofGuadua angustifolia Kunth. Pharmacogn Mag 2012;8:93–7.
Nakata K, Inagawa H, Soma G. Lipopolysaccharide IP-PA1 from Pantoea agglomerans prevents suppression ofmacrophage function in stress-induced diseases. AnticancerRes 2011;31:2437–40.
Nisiotou A, Rantsiou K, Iliopoulos V, et al. Bacterial species asso-ciated with sound and Botrytis-infected grapes from a Greekvineyard. Int J Food Microbiol 2011;145:432–6.
Nissan G, Manulis-Sasson S, Weinthal D, et al. The type III effec-tors HsvG and HsvB of gall-forming Pantoea agglomerans de-termine host specificity and function as transcriptional acti-vators. Mol Microbiol 2006;61:1118–31.
Nizan R, Barash I, Valinsky L, et al. The presence of hrp geneson the pathogenicity-associated plasmid of the tumorigenicbacterium Erwinia herbicola pv gypsophilae. Mol Plant MicrobeIn 1997;10:677–82.
Nunes C, Usall J, Teixido N, et al. Control of Penicillium expan-sum and Botrytis cinerea on apples and pears with the com-bination of Candida sake and Pantoea agglomerans. J Food Prot2002;65:178–84.
Oie S, Kiyonaga H, Matsuzaka Y, et al. Microbial contaminationof fruit and vegetables and their disinfection. Biol Pharm Bull2008;31:1902–5.
Ozdemir G, Ceyhan N, Ozturk T, et al. Biosorption ofchromium(VI), cadmium(II) and copper(II) by Pantoea spTEM18. Chem Eng J 2004;102:249–53.
Pepi M, Lampariello L, Altieri R, et al. Tannic acid degrada-tion by bacterial strains Serratia spp. and Pantoea sp isolatedfrom olive mill waste mixtures. Int Biodeter Biodegr 2010;64:73–80.
Perez-y-Terron R, Villegas M, Cuellar A, et al. Detection of Pan-toea ananatis, causal agent of leaf spot disease of maize, inMexico. Australas Plant Dis Notes 2009;4:96–9.
Pileggi M, Pileggi S, Olchanheski L, et al. Isolation of mesotrione-degrading bacteria from aquatic environments in Brazil.Chemosphere 2012;86:1127–32.
Pindi P, Yadav P, Shanker A. Identification of opportunisticpathogenic bacteria in drinking water samples of differentrural health centers and their clinical impacts on humans.Biomed Res Int 2013;2013:348250.
Pinto-Tomas A, Anderson M, Suen G, et al. Symbiotic nitrogenfixation in the fungus gardens of leaf-cutter ants. Science2009;326:1120–3.
Podgwaite J, D’Amico V, Zerillo R, et al. Bacteria associated withlarvae and adults of the asian longhorned beetle (Coleoptera:Cerambycidae). J Entomol Sci 2013;48:128–38.
Popp A, Cleenwerck I, Iversen C, et al. Pantoea gaviniae sp nov andPantoea calida sp nov., isolated from infant formula and an
infant formula production environment. Int J Syst Evol Micr2010;60:2786–92.
Poppe L, Vanhoutte S, Hofte M. Modes of action of Pantoea ag-glomerans CPA-2, an antagonist of postharvest pathogens onfruits. Eur J Plant Pathol 2003;109:963–73.
Prado S, Almeida R. Phylogenetic placement of pentatomid stinkbug gut symbionts. Curr Microbiol 2009;58:64–9.
Pusey PL. Biological control agents for fire blight of apple com-pared under conditions limiting natural dispersal. Plant Dis2002;86:639–44.
Pusey PL, Stockwell VO, Reardon CL, et al. Antibiosis activ-ity of Pantoea agglomerans biocontrol strain E325 againstErwinia amylovora on apple flower stigmas. Phytopathology2011;101:1234–41.
Rafat A, Philip K, Muniandy S. A novel source of bioactive com-pounds: endophytic bacteria isolated from Centella asiatica. JPure Appl Microbiol 2012;6:11–20.
Remus-Emsermann M, Kim E, Marco M, et al. Draft genome se-quence of the phyllospheremodel bacterium Pantoea agglom-erans 299R. Genome Announc 2013;1:e00036–13.
Rezzonico F, Smits T, Montesinos E, et al. Genotypic comparisonof Pantoea agglomerans plant and clinical strains. BMC Micro-biol 2009;9:204, DOI: 10.1186/1471-2180-9-204.
Rezzonico F, Smits THM, Duffy B. Misidentification slanders Pan-toea agglomerans as a serial killer. J Hosp Infect 2012;81:137–9.
Robacker D, Lauzon C. Purine metabolizing capability of Enter-obacter agglomerans affects volatiles production and attrac-tiveness to Mexican fruit fly. J Chem Ecol 2002;28:1549–63.
Rodrigues A, Lima I, Junior A, et al. Pantoea agglomerans liver ab-scess in a resident of Brazilian Amazonia. Trop Gastroenterol2009;30:154–5.
Romeiro R, Macagnan D, Mendonca H, et al. Bacterial spot of Chi-nese taro (Alocasia cucullata) in Brazil induced by Pantoea ag-glomerans. Plant Pathol 2007;56:1038.
Roper M. Pantoea stewartii subsp. stewartii: lessons learned froma xylem-dwelling pathogen of sweet corn. Mol Plant Pathol2011;12:628–37.
Ruiz-Manriquez A, Magana P, Lopez V, et al. Biosorption of Cu byThiobacillus ferrooxidans. Bioprocess Eng 1998;18:113–8.
Sauter R, Williams C, Meyer E, et al. A study of bacteria presentwithin unfertilized salmon eggs at the time of spawning andtheir possible relation to early lifestage disease. J Fish Dis1987;10:193–203.
Schisler D, Slininger P. Selection and performance of bacterialstrains for biologically controlling Fusarium dry rot of pota-toes incited by Gibberella pulicaris. Plant Dis 1994;78:251–5.
Schmid H, Schubert S, Weber C, et al. Isolation of a Pan-toea dispersa-like strain from a 71-year-old woman withacute myeloid leukemia and multiple myeloma. Infection2003;31:66–7.
Selvakumar G, Kundu S, Joshi P, et al. Characterization of a cold-tolerant plant growth-promoting bacterium Pantoea dispersa1A isolated from a sub-alpine soil in the North Western In-dian Himalayas. World J Microb Biot 2008;24:955–60.
September S, Els F, Venter S, et al. Prevalence of bacterialpathogens in biofilms of drinking water distribution sys-tems. J Water Health 2007;5:219–27.
Shubov A, Jagannathan P, Chin-Hong PV. Pantoea agglomeranspneumonia in a heart-lung transplant recipient: case reportand a review of an emerging pathogen in immunocompro-mised hosts. Transpl Infect Dis 2011;13:536–9.
Skorska C, Mackiewicz B, Gora A, et al. Health effectsof exposure to inhalation of organic dust in hop
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/39/6/968/547864 by guest on 26 March 2022

Walterson and Stavrinides 983
farmers. Ann Univ Mariae Curie Sklodowska Med 2003;58:459–65.
Slotnick I, Tulman L. A human infection caused by an ErwiniaSpecies. Am J Med 1967;43:147–50.
Smith DDN, Kirzinger MWB, Stavrinides J. Draft genome se-quence of the antibiotic-producing cystic fibrosis isolate Pan-toea agglomerans Tx10. Genome Announc 2013a;1:e00904–13.
Smits T, Rezzonico F, Kamber T, et al. Genome sequence ofthe biocontrol agent Pantoea vagans strain C9-1. J Bacteriol2010;192:6486–7.
Smits T, Rezzonico F, Kamber T, et al. Metabolic versatility andantibacterial metabolite biosynthesis are distinguishing ge-nomic features of the fire blight antagonist Pantoea vagansC9-1. PloS One 2011;6:e22247, DOI: 10.1371/journal.pone.0022247.
Son H, Park G, Cha M, et al. Solubilization of insoluble inorganicphosphates by a novel salt- and pH-tolerant Pantoea agglomer-ans R-42 isolated from soybean rhizosphere. Bioresource Tech-nol 2006;97:204–10.
Sood P, Nath A. Bacteria associated with Bactrocera sp.(Diptera:Tephritidae)—isolation and identification. PestManag Econo Zool 2002;10:1–9.
Stavrinides J. Origin and evolution of phytopathogenic bacte-ria. In Jackson RW (ed). Plant Pathogenic Bacteria: Genomics andMolecular Biology. UK: Caister Academic Press, 2009, 1–35.
Stavrinides J, No A, Ochman H. A single genetic locus in thephytopathogen Pantoea stewartii enables gut colonization andpathogenicity in an insect host. Environ Microbiol 2010;12:147–55.
Stets MI, Pinto AS, Jr, Huergo LF, et al. Rapid identification of bac-terial isolates fromwheat roots by high resolution whole cellMALDI-TOF MS analysis. J Biotechnol 2013;165:167–74.
Sturz A, Matheson B. Populations of endophytic bacteria whichinfluence host-resistance to Erwinia-induced bacterial softrot in potato tubers. Plant Soil 1996;184:265–71.
Sulbaran M, Perez E, Ball M, et al. Characterization of the min-eral phosphate-solubilizing activity of Pantoea agglomeransMMB051 isolated from an iron-rich soil in SoutheasternVenezuela (Bolivar State). Curr Microbiol 2009;58:378–83.
SultanaM, Hartig C, Planer-Friedrich B, et al. Bacterial communi-ties in Bangladesh aquifers differing in aqueous arsenic con-centration. Geomicrobiol J 2011;28:198–211.
Terenius O, Lindh J, Eriksson-Gonzales K, et al. Midgut bacterialdynamics inAedes aegypti. FEMSMicrobiol Ecol 2012;80:556–65.
Tindall BJ. The combination Enterobacter agglomerans is to becited as Enterobacter agglomerans (Beijerinck 1888) Ewing andFife 1972 and the combination Pantoea agglomerans is to becited as Pantoea agglomerans (Beijerinck 1888) Gavini et al.1989. Opinion 90. Int J Syst Evol Micr 2014;64:3582–3.
Torres R, Teixido N, Usall I, et al. Post-harvest control of Penicil-lium expansum on pome fruits by the bacterium Pantoea ana-natis CPA-3. J Hortic Sci Biotech 2005;80:75–81.
Trippe K, McPhail K, Armstrong D, et al. Pseudomonas fluorescensSBW25 produces furanomycin, a non-proteinogenic aminoacid with selective antimicrobial properties. BMC Microbiol2013;13:111, DOI: 10.1186/1471-2180-13-111.
Ulloa-Gutierrez R, Moya T, Avila-Aguero M. Pantoea agglomeransand thorn-associated suppurative arthritis. Pediatr Infect DisJ 2004;23:690.
Usall J, Teixido N, Vinas I, et al. Biological control of Peni-cillium digitatum on citrus fruits with the antagonis-tic bacterium Pantoea agglomerans. Acta Hortic 2001;553:377–82.
Valinsky L, Manulis S, Nizan R, et al. A pathogenicity gene iso-lated from the pPATH plasmid of Erwinia herbicola pv. gyp-sophilae determines host specificity. Mol Plant Microbe In1998;11:753–62.
Van Rostenberghe H, Noraida R, WanPauzi W, et al. The clinicalpicture of neonatal infection with Pantoea species. Jpn J InfectDis 2006;59:120–1.
Vanneste J, Cornish D, Yu J, et al. A microcin produced by astrain of Erwinia herbicola is involved in biological controlof fire blight and soft rot caused by Erwinia sp. Acta Hortic2000;513:39–46.
Vanneste J, Cornish D, Yu J, et al. The peptide antibiotic pro-duced by Pantoea agglomerans Eh252 is a microcin. Acta Hortic2002;590:285–90.
Vanneste J, Yu J, Cornish D. Presence of genes homologousto those necessary for synthesis of microcin MccEh252 instrains of Pantoea agglomerans. Acta Hortic 2008;793:391–6.
Vanneste J, Yu J, Reglinski T. Biocontrol agent Pantoea agglomeransstrain NZ501 induces a resistance-like response in kiwifruitand tobacco cells. Acta Hortic 2002;590:279–83.
Vasileva-Tonkova E, Gesheva V. Biosurfactant production byantarctic facultative anaerobe Pantoea sp during growth onhydrocarbons. Curr Microbiol 2007;54:136–41.
VenturiniM, Oria R, BlancoD.Microflora of two varieties of sweetcherries: Burlat and Sweetheart. FoodMicrobiol 2002;19:15–21.
Verdier-Metz I, Gagne G, Bornes S, et al. Cow teat skin, a poten-tial source of diverse microbial populations for cheese pro-duction. Appl Environ Microb 2012;78:326–33.
Verma S, Singh A, Chowdhury S, et al. Endophytic coloniza-tion ability of two deep-water rice endophytes, Pantoea spandOchrobactrum sp using green fluorescent protein reporter.Biotechnol Lett 2004;26:425–29.
Vieria R, de Lima E, Sousa D, et al. Vibrio spp. and Salmonella spp.,presence and susceptibility in crabs Ucides cordatus. Rev InstMed Trop Sao Paulo 2004;46:179–82.
Vincent K, Szabo R. Enterobacter agglomerans - osteomyelitis ofthe hand from a rose thorn - a case report. Orthopedics1988;11:465–7.
Volksch B, Thon S, Jacobsen I, et al. Polyphasic study of plant- andclinic-associated Pantoea agglomerans strains reveals indistin-guishable virulence potential. Infect Genet Evol 2009;9:1381–91.
Volksch B, UllrichM, FritscheW. Occurrence of antimicrobial ac-tivities of bacteria from soybean leaf spots. J Basic Microbiol1993;33:349–55.
von Bodman SB, Majerczak DR, Coplin DL. A negative regula-tor mediates quorum-sensing control of exopolysaccharideproduction in Pantoea stewartii subsp. stewartii. P Natl Acad SciUSA 1998;95:7687–92.
Vorwerk S, Blaich R, Forneck A. Pantoea sp.—an associated bac-teria common in grape Phylloxera (Daktulosphaira vitifoliaeFitch). Acta Hortic 2007;733:47–51.
Walcott R, Gitaitis R, Castro A, et al. Natural infestation of onionseed by Pantoea ananatis, causal agent of center rot. Plant Dis2002;86:106–11.
Walterson AM, Smith DDN, Stavrinides J. Identification ofa Pantoea biosynthetic cluster that directs the synthe-sis of an antimicrobial natural product. PLoS One 2014;9:e96208–8.
Wattam AR, Abraham D, Dalay O, et al. PATRIC, the bacterialbioinformatics database and analysis resource. Nucleic AcidsRes 2014;42:D581–91.
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/39/6/968/547864 by guest on 26 March 2022

984 FEMS Microbiology Reviews, 2015, Vol. 39, No. 6
Wehland M, Kiecker C, Coplin D, et al. Identification ofan RcsA/RcsB recognition motif in the promoters ofexopolysaccharide biosynthetic operons from Erwiniaamylovora and Pantoea stewartii subspecies stewartii. J BiolChem 1999;274:3300–7.
Weinthal D, Barash I, Panijel M, et al.Distribution and replicationof the pathogenicity plasmid pPATH in diverse populations ofthe gall-forming bacterium Pantoea agglomerans. Appl EnvironMicrob 2007;73:7552–61.
Weinthal D, Barash I, Tzfira T, et al. Characterization of nuclearlocalization signals in the type III effectors HsvG and HsvB ofthe gall-forming bacterium Pantoea agglomerans. Microbiology2011;157:1500–8.
Wells M, Gitaitis R, Sanders F. Association of tobacco thrips,Frankliniella fusca (Thysanoptera : Thripidae)with two speciesof bacteria of the genus Pantoea. Ann Entomol Soc Am2002;95:719–23.
Wirth R, Herz H, Ryel J et al.Herbivory of Leaf Cutting Ants: A CaseStudy on Atta Colombica in the Tropical Rain Forest of Panama.Berlin: Springer, 2003.
Wright S, Beer S. The role of antibiotics in biological controlof fire blight by Erwinia herbicola strain EH318. Acta Hortic1996;411:309–11.
Wright S, Beer S. Genes for biosynthesis of pantocin A and B byPantoea agglomerans Eh318. Acta Hortic 2002;590:237–41.
Wright S, JinM, Clardy J, et al. The biosynthetic genes of pantocinA and pantocin B of Pantoea agglomerans Eh318. Acta Hortic2006;704:313–20.
Wu C, Li F, Zhou S, et al. Isolation and characterization of afacultative anaerobe Pantoea agglomerans MFC-3 and its hu-mic substance-and Fe(III)—respiring activity.Huan Jing Ke Xue2010;31:237–42.
Yeung K, Lee K, Woodard R. Isolation and identification of two L-azetidine-2-carboxylic acid-degrading soil microorganisms,Enterobacter agglomerans and Enterobacter amnigenus. J Nat Prod1998;61:207–11.
Yoshida A, Kohchi C, Inagawa H, et al. Improvement of aller-gic dermatitis via regulation of the Th1/Th2 immune sys-tem balance by macrophages activated with lipopolysaccha-ride derived from Pantoea agglomerans (IP-PA1). Anticancer Res2009;29:4867–70.
Zamani M, Tehrani A, Ahmadzadeh M, et al. Control of Penicil-lum digitatum on orange fruit combining Pantoea agglomeranswith hot sodium bicarbonate dipping. J Plant Pathol 2009;91:437–42.
Zhang W, Han D, Dick W, et al. Compost and compost waterextract-induced systemic acquired resistance in cucumberand Arabidopsis. Phytopathology 1998;88:450–5.
Zuberbuhler B, Carifi G, Leatherbarrow B. Acute dacryocystitis ina 2-year old child caused by Pantoea. Orbit 2012;31:13–4.
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/39/6/968/547864 by guest on 26 March 2022




















