
Bacterial diversity of the rhizosphere of maize (Zea mays) grown in tropical soil studied by...
14
Plant and Soil 232: 167–180, 2001. © 2001 Kluwer Academic Publishers. Printed in the Netherlands. 167 Bacterial diversity of the rhizosphere of maize (Zea mays) grown in tropical soil studied by temperature gradient gel electrophoresis N. C. M. Gomes 2 , H. Heuer 1 , J. Schönfeld 1 , R. Costa 2 , L. Mendonça-Hagler 2 & K. Smalla 1,3 1 Federal Biological Research Centre for Agriculture and Forestry, Messeweg 11/12, 38104 Braunschweig, Ger- many 2 Federal University of Rio de Janeiro, Institute for Microbiology, IMPPG-CCS, CEP 21941-590 Rio de Janeiro, Brazil 3 Corresponding author ∗ Key words: maize rhizosphere, tropical soil, TGGE analysis Abstract The bacterial diversity and population dynamics in the rhizosphere of two maize cultivars (Nitroflint and Nitrodent) grown in tropical soils was studied, by traditional cultivation techniques and 16S rRNA gene-based molecular analysis of DNA directly extracted from soil and rhizosphere samples. Rhizosphere and soil samples were taken at three different plant growth stages. Total aerobic bacterial counts were determined. Fingerprints of the most dominant bacterial population were generated by TGGE separation of 16S rRNA gene fragments amplified from total community DNA using eubacterial specific primers. To reduce the complexity of TGGE fingerprints or to analyse less abundant populations, primers specific for different phylogenetic groups have been used. A comparison of the cfu obtained for rhizosphere of both cultivars indicated significant differences only for rhizosphere and soil samples taken 40 days after sowing. However, a comparison of TGGE patterns indicated that the composition of the bacterial community analysed at different plant growth stages for both cultivars was similar. A comparison of α-, β -proteobacterial and actinomycete TGGE patterns of both cultivars confirmed this observation. The eubacterial TGGE profiles reflected strong seasonal population shifts in the bacterial rhizosphere community of both maize cultivars which could be also observed in the TGGE patterns of α- and β -proteobacteria and to a lesser extent for actinomycetes. The rhizosphere effect was much more pronounced for young roots compared to samples taken from mature maize plants. The rhizosphere fingerprints showed a reduced complexity for young plants with up to five dominating bands while for mature plants patterns similar to those of soil were observed. Sequencing of dominant clones indicated that the dominant population found at all plant growth stages can be assigned to Arthrobacter populations. Introduction The rhizosphere defined as the fraction of soil adher- ing to plant roots is thought to be of great importance to plant health and soil fertility. Root exudates se- lectively influence the growth of bacterial and fungal populations by altering the presence of substrates in soil in the vicinity of roots (Jaeger et al., 1999; Yang and Crowley, 2000). The structural and functional di- versity of rhizosphere populations is supposed to be affected by differences in root exudation and rhizode- ∗ Fax: 49-531-2993013. E-mail: [email protected] position in different root zones and in relation to soil type, plant species, growth stage, cultural practices, such as tillage and crop rotation and other environ- mental factors (De Leij et al., 1994; Latour et al., 1996; Westover et al., 1997; Grayston et al., 1998; Horwath et al., 1998; Lupwayi et al., 1998). Rhizo- sphere microorganisms in turn exert strong effects on plant growth and health by nutrient solubilisation, N 2 fixation, or by the production of plant hormones (Höf- lich et al., 1994; van Loon et al., 1998). Increased plant productivity also results from the suppression of deleterious microorganisms by antagonistic bac- teria while soil-borne pathogens can greatly reduce
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Bacterial diversity of the rhizosphere of maize (Zea mays) grown in tropical soil studied by...

Plant and Soil 232: 167–180, 2001.© 2001 Kluwer Academic Publishers. Printed in the Netherlands.
167
Bacterial diversity of the rhizosphere of maize (Zea mays) grown intropical soil studied by temperature gradient gel electrophoresis
N. C. M. Gomes2, H. Heuer1, J. Schönfeld1, R. Costa2, L. Mendonça-Hagler2 & K. Smalla1,3
1Federal Biological Research Centre for Agriculture and Forestry, Messeweg 11/12, 38104 Braunschweig, Ger-many2Federal University of Rio de Janeiro, Institute for Microbiology, IMPPG-CCS, CEP 21941-590 Rio de Janeiro,Brazil3Corresponding author∗
Key words: maize rhizosphere, tropical soil, TGGE analysis
Abstract
The bacterial diversity and population dynamics in the rhizosphere of two maize cultivars (Nitroflint and Nitrodent)grown in tropical soils was studied, by traditional cultivation techniques and 16S rRNA gene-based molecularanalysis of DNA directly extracted from soil and rhizosphere samples. Rhizosphere and soil samples were takenat three different plant growth stages. Total aerobic bacterial counts were determined. Fingerprints of the mostdominant bacterial population were generated by TGGE separation of 16S rRNA gene fragments amplified fromtotal community DNA using eubacterial specific primers. To reduce the complexity of TGGE fingerprints or toanalyse less abundant populations, primers specific for different phylogenetic groups have been used. A comparisonof the cfu obtained for rhizosphere of both cultivars indicated significant differences only for rhizosphere and soilsamples taken 40 days after sowing. However, a comparison of TGGE patterns indicated that the composition of thebacterial community analysed at different plant growth stages for both cultivars was similar. A comparison of α-,β-proteobacterial and actinomycete TGGE patterns of both cultivars confirmed this observation. The eubacterialTGGE profiles reflected strong seasonal population shifts in the bacterial rhizosphere community of both maizecultivars which could be also observed in the TGGE patterns of α- and β-proteobacteria and to a lesser extent foractinomycetes. The rhizosphere effect was much more pronounced for young roots compared to samples taken frommature maize plants. The rhizosphere fingerprints showed a reduced complexity for young plants with up to fivedominating bands while for mature plants patterns similar to those of soil were observed. Sequencing of dominantclones indicated that the dominant population found at all plant growth stages can be assigned to Arthrobacterpopulations.
Introduction
The rhizosphere defined as the fraction of soil adher-ing to plant roots is thought to be of great importanceto plant health and soil fertility. Root exudates se-lectively influence the growth of bacterial and fungalpopulations by altering the presence of substrates insoil in the vicinity of roots (Jaeger et al., 1999; Yangand Crowley, 2000). The structural and functional di-versity of rhizosphere populations is supposed to beaffected by differences in root exudation and rhizode-
∗ Fax: 49-531-2993013. E-mail: [email protected]
position in different root zones and in relation to soiltype, plant species, growth stage, cultural practices,such as tillage and crop rotation and other environ-mental factors (De Leij et al., 1994; Latour et al.,1996; Westover et al., 1997; Grayston et al., 1998;Horwath et al., 1998; Lupwayi et al., 1998). Rhizo-sphere microorganisms in turn exert strong effects onplant growth and health by nutrient solubilisation, N2fixation, or by the production of plant hormones (Höf-lich et al., 1994; van Loon et al., 1998). Increasedplant productivity also results from the suppressionof deleterious microorganisms by antagonistic bac-teria while soil-borne pathogens can greatly reduce

168
plant growth. Clearly, bacterial and fungal communit-ies can be influenced by the plant as well as viceversa (Watkinson, 1998). Studies of rhizosphere mi-crobial communities performed by traditional cultiva-tion techniques indicated that the presence of differentplant species influences the composition of the mi-crobial community due to the differential response ofbacterial and fungal populations to different root ex-udation patterns (Miller et al., 1989). Understandingand knowledge of the structural composition and dy-namics of rhizosphere microbial communities has longbeen limited because only a minor fraction of micro-bial populations is accessible by traditional cultivationtechniques (Amann et al., 1995). Nucleic acid-basedanalysis of bacterial communities allows us to over-come biases of cultivation-dependent methods knownas the ‘Great plate count phenomenon’ (Staley andKonopka, 1985). To study spatial and temporal vari-ation of rhizosphere and soil bacterial communities,multiple sample analysis is essential (Van Elsas andSmalla, 1996; Muyzer and Smalla, 1998; Gelsominoet al., 1999). For this purpose approaches based oncloning and sequencing of 16S rDNA fragments PCR-amplified from community DNA or on the character-isation of bacterial isolates are too labour-intensiveand time-consuming. Molecular fingerprinting tech-niques based on 16S rDNA fragments amplified fromcommunity DNA have opened a new dimension tostudy bacterial rhizosphere communities.
The maize cultivars Nitroflint and Nitrodent weredeveloped in a breeding programme by EMBRAPA(Brazil) to improve N-utilisation. While Nitroflint hasbeen commercialised a few years ago, Nitrodent is stillbeing evaluated in extensive field tests. Plant breed-ing by traditional techniques or gene technology mightaffect the structural and functional diversity of rhizo-sphere microbial communities, e.g., by an altered rootmorphology and physiology, plant exudation (Engel-hard et al., 2000), and thus affect the balance ofplant-beneficial and deleterious microbes. Potentialeffects of a new cultivar on the bacterial communitystructure and function need to be assessed in the con-text of natural variability, e.g., seasonal shifts. Thetwo different maize cultivars Nitroflint and Nitrodenthave been used as model plants to explore the resolu-tion power of temperature gradient gel electrophoresis(TGGE) of PCR-amplified 16S rDNA fragments toanalyse (i) whether there are soil bacterial populationswhich are enriched in the rhizosphere of maize, (ii)whether the abundance and composition of bacterialpopulations in the rhizosphere of maize changes dur-
ing the vegetation period, and (iii) whether there isan influence of the cultivar on the composition of thebacterial community.
To study the bacterial diversity and populationdynamics in the rhizosphere of two maize cultivarsgrown in tropical soils, rhizosphere and soil sampledat three different time points during the growing sea-son were analysed by traditional cultivation techniquesand 16S rRNA gene-based molecular analysis of DNAdirectly extracted from soil and rhizosphere samples.TGGE analysis of 16S rRNA gene fragments amp-lified from total community DNA using eubacterialspecific primers generates fingerprints of the mostdominant bacterial population. To reduce the complex-ity of TGGE fingerprints or to analyse less abundantpopulations, primers specific for different phylogen-etic groups have been used. PCR products obtainedwith taxon-specific primers without GC clamps servedas template in TGGE-PCR with eubacteria-specificprimers (Heuer and Smalla, 1997; Heuer et al., 1997).In addition, a clone library was generated from 16SrRNA genes amplified from rhizosphere communityDNA. Clones matching dominant bands of the com-munity pattern have been sequenced.
The data provided by the study should be of in-terest as a ‘baseline’ for the successful applicationof biological control agents and for the evaluationof potential effects of transgenic maize plants onrhizosphere microbial communities.
Materials and methods
Field design and sampling
The field test was performed at EMBRAPA (EmpresaBrasileira de Pesquisa Agropecuária), located inSeropédica, Rio de Janeiro state, Brazil. Two cultivarsof maize (Zea mays), Nitroflint and Nitrodent, wereeach planted in three plots (3 × 6 m) in a random-ised block design as part of a larger field test. Twoindependent soil and rhizosphere samples were takenfrom each plot 20, 40 and 106 days after sowing. Eachof the six replicate rhizosphere samples per samplingcomprised total roots from five maize plants. The rootswere shaken vigorously to separate soil not tightlyadhering to the roots. Each of the six replicate soilsamples per sampling consisted of eight cores (15 cmtop soil) which were mixed and sieved. The soil coreswere taken between the maize rows (approx. 50 cmdistance from plants), they were free of roots and

169
homogenous with depth. Samples were collected insterile plastic bags and processed the same day for de-termination of bacterial counts. Subsamples for DNAextraction were kept frozen at −20◦C.
Determination of bacterial counts
Five grams of soil or maize roots with adhering soilwere resuspended in 45 ml of 0.1% sodium pyro-phosphate containing 0.1% Tween 80 in Erlenmeyerflasks with glass beads. After 45 min shaking at 200rpm, serial dilutions were made with sterile saline andplated onto 1/10 TSA (Trypticase soy agar) and King’sB agar. Counts were determined after 48 h incubationat 29◦C.
Total community DNA isolation
Total community DNA was extracted from 1 g offrozen soil or rhizosphere (root with tightly adheringsoil particles) samples using the protocol described bySmalla and van Elsas (1995). This protocol involvesan enzymatic (lysozyme), mechanical (bead beating)and chemical (SDS) treatment of the samples followedby a phenol extraction of the soil slurry. In contrastto other soils, we observed that during extraction thephenol phase was the upper and the aqueous phasethe lower phase. The crude DNA was purified usingCsCl and sodium acetate precipitation and glass milkpurification as described by Smalla et al. (1993).
Genomic DNA isolation from strains
Genomic DNA was isolated from freshly grown colon-ies using the CTAB protocol described by Wilson et al.(1987).
PCR-amplification of 16S rRNA gene fragments forTGGE analysis
PCR was performed with a DNA Thermal Cycler 480(Perkin-Elmer Cetus, Norwalk, USA). For amplifica-tion of 16S rDNA fragments used for TGGE analysis,the reaction mixture was: 1 µl template DNA (ca.20 ng), Stoffel buffer (10 mM KCl, 10 mM Tris–HCl, pH 8.3), 0.2 mM dNTPs, 3.75 mM MgCl2, 4%(w/v) acetamide, 100 nM F984GC, 100 nM R1378,and 2 U/25 µl taq DNA polymerase (Stoffel fragment,Perkin-Elmer Cetus).
All primers which are listed in Table 1 were optim-ised using the Oligotm programme vers. 4.0 (Rychlikand Rhoads, 1989). A GC-rich sequence is attached
to primer F984GC, so the PCR products have a GC-clamp to prevent complete melting during separationin the temperature gradient gel (Sheffield et al., 1989).Acetamide was added to the reaction mixture to facil-itate the denaturation of dsDNA and to circumvent theformation of secondary structures. A 24-µl aliquot ofthe PCR mix was added to a 0.5-ml reaction tube andoverlaid with a drop of mineral oil before adding thetemplate DNA. After 5 min of denaturation at 94◦C,30 thermal cycles of 1 min at 94◦C, 1 min at 53◦C,and 2 min at 72◦C were performed, PCR was finishedby an extension step at 72◦C for 10 min.
Group-specific PCR of 16S rRNA gene fragments
For amplification of α-, β-proteobacterial or actin-omycete 16S rDNA fragments, the reaction mixturewas: 1 µl template DNA (ca. 20 ng), Stoffel buffer(10 mm KCl, 10 mM Tris–HCl, pH 8.3), 0.2 mMdNTPs, 3.75 mM MgCl2, 5% (w/v) DMSO, 100 nMof the forward group-specific primer (see Table 1),100 nM R1494, and 1 U/24 µl taq DNA polymerase(Stoffel fragment, Perkin-Elmer Cetus). Bovine serumalbumin (2.5 µg/25 µl) was added to all group-specificPCR reactions. After initial 5 min of denaturationat 94◦C, 20 thermal cycles of 1 min 94◦C, 1 min56◦C (α-proteobacteria)/61◦C (β-proteobacteria), and2 min 72◦C were performed. PCR was finished by afinal extension step at 72◦C for 10 min. A hot startwas performed for β-proteobacterial and actinomy-cete PCR to improve the specificity. One to 30 dilutedPCR products served as template for a TGGE-PCRperformed with 20 cycles only. PCR-amplification ofActinomycetales populations was done as describedby Heuer et al. (1997).
Temperature gradient gel electrophoresis
For TGGE analysis the Diagen TGGE system (Dia-gen, Düsseldorf, Germany) was used according to themanufacturer’s instructions, and as described in moredetail by Heuer et al. (1997). TGGE with a gradientof 38–53◦C was done for 16 h at 110 V. Acid silverstaining was used for detection of DNA (Riesner etal., 1989).
Cloning and sequencing
PCR products (0.5 µl) amplified from two rhizosphereDNAs (first sampling) with primers (F27, R1494)were cloned using the TOPO-TA pCR 2.1 cloning

170
Table 1. Primer sequences used for PCR-amplification of 16S rDNA fragments
Primer Sequence 5′–3′ (target 16S rDNA) Reference
F984GC AACGCGAAGAACCTTAC (Bacteria) Heuer et al., 1997
GC-clamp CGCCCGGGGCGCGCCCCGGGCGGGGCGGG Nübel et al., 1996
GGCACGGGGGG, 5′ attached to F984GC
F27 AGAGTTTGATC(A/C)TGGCTCAG (Bacteria) Weisburg et al., 1991
F243HGC GGATGAGCCCGCGGCCTA (Actinomycetales) Heuer et al., 1997
F203α CCGCATACGCCCTACGGGGGAAAGATTTAT This study
(α-Proteobacteria)
F948β CGCACAAGCGGTGGATGA (β-Proteobacteria) This study
R1378 CGGTGTGTACAAGGCCCGGGAACG (Bacteria) Heuer et al., 1997
R1494 CTACGG(T/C)TACCTTGTTACGAC (Bacteria) Weisburg et al., 1991
kit according to the instructions of the manufac-turer (Invitrogen, Groningen, The Netherlands). Whitecolonies were picked and screened by TGGE for thepresence of an insert. Clones matching bands of thecommunity pattern were selected and sequenced.
Results
We have applied a combination of traditional cultiva-tion and molecular techniques (TGGE fingerprinting)to study the structural diversity of the bacterial com-munity in the rhizosphere of the two different maizecultivars Nitroflint and Nitrodent grown in tropicalsoil. The approach chosen should allow us to linkbands from community fingerprints to isolates, whichwere obtained from the rhizosphere and soil samplesanalysed, at a later stage.
Determination of bacterial counts
The cfu of maize rhizosphere samples taken at threedifferent time points during plant development weredetermined on TSA (1/10) and ranged from 9 × 107
to 4 × 108 per g of fresh weight. The total cfu de-termined in soil was 6 × 107 to 5 × 108 per gand thus was in the same range as for rhizosphere.All cfu counts (data not shown) were analysed byANOVA. When analysing seasonal shifts of cfucounts, significant differences of cfu counts (0.003)were observed only between rhizosphere samples ofthe first and second sampling. The mean cfu num-bers were higher for rhizosphere sampled 40 days aftersowing compared to samples from young plants. Acomparison of the cfu counts obtained for rhizosphereof both cultivars indicated significant differences only
for rhizosphere (0.02) and soil (0.014) samples taken40 days after sowing. A collection of bacterial isolateswas obtained from TSA and King’s B media to allowa characterisation of dominant isolates and a link withthe TGGE-fingerprints at a later stage.
Total community DNA extraction and TGGE analysis
Total community DNA was extracted after a rigorouslysis from frozen rhizosphere and soil samples. Typic-ally, good yields of high-molecular weight DNA wereobtained. The 16S rDNA fragments could be obtainedfrom all 36 soil and all 36 rhizosphere communityDNA samples. Fingerprints of the most dominantpopulations were obtained after separation of PCRproducts in a temperature gradient from 38 to 53◦C.To reduce the complexity of the patterns and to al-low the analysis of minority populations, a groupspecific PCR enrichment of α- and β-proteobacteriaand Gram-positive high G+C bacteria followed by aTGGE-PCR was done. PCR products obtained witheubacterial primers (F984GC; R1378) or after group-specific PCR were analysed in different combinationson TGGE gels to test the following hypotheses.
Are soil bacterial populations enriched in therhizosphere of maize?
A comparison of the rhizosphere fingerprints of thetwo cultivars Nitroflint and Nitrodent and the corres-ponding bulk soil fingerprints showed an enrichmentof several populations in the rhizosphere. However,the rhizosphere effect was much more pronouncedfor young roots (20 days after sowing) compared tosamples taken from mature maize plants (106 daysafter planting). While at all sampling times the soil

171
Figure 1. TGGE fingerprints of bacterial communities by TGGEseparation of 16S rDNA fragments amplified with primers F984GCand R1378: lanes: (1–6) soil DNA from plots planted with Ni-troflint; (7–12) rhizosphere of Nitroflint (20 days after sowing);(13) bacterial standard. cp, Clostridium pasteurianum DSM 525;pc, Pectobacterium carotovora DSM 30168; at, Agrobacteriumtumefaciens DSM 30205; pf, Pseudomonas fluorescens R2f; pa,Pantoea agglomerans; na, Nocardia asteroides N3; rl, Rhizobiumleguminosarum DSM 30132; av, Actinomadura viridis DSM 43462;ka, Kineosporia aurantiaca JCM 3230; no, Nocardiopsis atraATCC 31511; ap, Actinoplanes philippiensis JCM 3001.
pattern consisted of one intense band and a largenumber of equally abundant faint bands, the rhizo-sphere fingerprints showed a reduced complexity foryoung plants with up to five dominating bands (seeFig. 1). The dominant band [a] of the soil patternsappears also as the dominant population in the rhizo-sphere (Fig. 1). In particular, for rhizosphere samplestaken 40 days after sowing band [a] seems to repres-ent the numerically dominant population. All otherdominant bands observed in the rhizosphere patternof young roots did not represent dominant populationsin bulk soil. Thus for the dominant rhizosphere band[b] (Fig. 1), no corresponding band could be detec-ted in the bulk soil patterns. Some dominant bands inthe rhizosphere patterns do not appear in all sampleswhile others do. The TGGE patterns of the rhizo-
sphere of mature plants look very much like those ofthe bulk soil (no data shown). Although the rhizo-sphere patterns were slightly more intense, both soiland rhizosphere samples typically show a large num-ber of faint bands and a more intense band with theelectrophoretic mobility as band [a] in Fig. 1.
Does the abundance and composition of bacterialpopulations in the rhizosphere of maize changeduring the vegetation period?
The TGGE profiles reflected strong seasonal popu-lation shifts in the bacterial rhizosphere communityof both maize cultivars, as shown for Nitroflint inFig. 2. The profiles from young plants (Fig. 2; lanes2–5) showed a few dominant bands (four intenseand approximately 10 fainter bands), indicating thedominance of a few populations. Due to the strongdominance of only one band, a reduced complexityof the TGGE patterns was observed for samples taken40 days after sowing. For the first and the secondsampling the majority of bands was localised in themiddle part of the gel, and almost no bands werevisible in the lower part of the gel indicating thatpopulations with a high-G+C such as Streptomycetesdid not belong to the numerically dominant popula-tions. In contrast to the first and second sampling,the TGGE patterns of mature plants (106 days afterplanting) showed highly complex banding patterns.More than 50 faint bands were visible indicating thatmany equally abundant populations were present. Astriking diversity was observed, in particular, in thelower part of the TGGE gel for rhizosphere of ma-ture plants. The two most intense bands [a] and [c]were detected in all parallels of young plants whileband [b] was detected only in three of four replicates.The dominant band [a] was, however, observed onlyin the rhizosphere of young plants and disappearedat the later sampling stages. The relative intensityof the dominant band [c] which could be detectedin the TGGE patterns at all plant growth stages in-creased in intensity when rhizosphere patterns of thefirst and second samplings were compared and be-came less dominant for mature plants. Band [c] couldbe also detected as dominant band in the TGGE pat-terns of high G+C Gram-positives in the rhizosphereof the first and second sampling after prior enrichment(Fig. 3). This band could be matched with the most fre-quently recovered 16S rDNA clone. Sequencing of the16S rDNA of six clones of identical electrophoreticalmobility as this band showed that they all belonged

172
Figure 2. Seasonal shifts of bacterial rhizosphere communities: fingerprints by TGGE separation of 16S rDNA fragments amplified withprimers F984GC and R1378 from Nitroflint rhizosphere DNA. Lanes: (2–5) 20 days after sowing; (6–10) 40 days after sowing; (11–15) 106days after sowing, lanes 1 and 16 bacterial standard.
to the Arthrobacter genus, showing high similaritywith the type strains Arthrobacter polychromogenes orArthrobacter nicotinovorans (Table 2). All six clonesdiffered in two or more nucleotides of the TGGE-fragment, and showed substantial sequence variationin the V3-region.
The seasonal shifts of high G+C populations wereless pronounced (Fig. 3) than observed for eubacteria.At all three plant growth stages analysed the patterns
were rather complex. With the exception of a fewbands, a large number of high G+C Gram-positivepopulations seems to be present throughout the season.The relative abundance of band [a] in Fig. 3 (band [c]in Fig. 2), which presumably resembles Arthrobacterpopulations, decreases with increasing plant age. Thepatterns indicate an impressive diversity of high G+CGram-positives in the rhizosphere of maize grown intropical soil.

173
Figure 3. Seasonal shifts of high G+C Gram-positive communities: fingerprints by TGGE separation of 16S rDNA fragments amplified withprimers F984GC and R1378 from amplicons which were obtained by PCR with primers F243HGC and R1494 from Nitroflint rhizosphereDNA. Lanes: (2–6) 20 days after sowing; (7–10) 40 days after sowing; (11–15) 106 days after sowing; (1, 16) bacterial standard.
The seasonal shift of α-proteobacterial populationsshowed a similar trend as seen for the eubacterialpatterns (Fig. 4). The rhizosphere samples taken 20or 40 days after sowing showed one dominant band[a] and approximately 15 fainter bands. The dom-inating band [a] showed an identical electrophoreticmobility as the A. tumefaciens strain of the standardand might indeed represent an A. tumefaciens pop-
ulation, since when a collection of α-proteobacteriawas screened by the same PCR-TGGE approach anidentical electrophoretic mobility was observed onlyfor A. tumefaciens strains (data not shown). The rel-ative abundance of this population is decreasing withincreasing plant age. However, while band [a] wasdetected in the eubacterial patterns only in the rhizo-sphere of young plants (Fig. 2), this population could

174
Table 2. Analysis of abundant 16S rDNA fragments
Clone No. of Type strain with the most similar 16S rRNA Similarity
nucleotides
(bp)
RF5-4 465 Arthrobacter polychromogenes DSM 20136 0.994
RF5-1 1447 Arthrobacter polychromogenes DSM 20136 0.972
RF3-H12 1447 Arthrobacter polychromogenes DSM 20136 0.973
RF5-31 1462 Arthrobacter polychromogenes DSM 20136 0.929
RF5-9 1451 Arthrobacter nicotinovorans DSM 420 0.995
RF5-20 466 Arthrobacter nicotinovorans DSM 420 0.972
be detected in the α-proteobacterial patterns also inthe maize rhizosphere 40 days after sowing (Fig. 4).In comparison with the rhizosphere patterns at earlierplant growth stages the α-proteobacterial patterns ofmature plants showed a higher complexity; however,still less complex than found for the bacterial patterns.Although some variability was observed between theparallels, seven bands could be detected in the TGGEpatterns of all samples from mature plants.
Is there an influence of the cultivar on thecomposition of the microbial community?
TGGE fingerprints of young maize plants were com-pared for both cultivars Nitroflint and Nitrodent. Fivereplicates of each cultivar per sampling time were runnext to each other in a TGGE gel allowing us to as-sess the variability between different replicates andbetween the two cultivars. Although some variabil-ity was observed in the eubacterial TGGE patterns,there was no band which could be observed only inthe pattern of one or the other cultivar. The TGGEpatterns obtained for young plants of both cultivars,Nitroflint (lanes 3–7) and Nitrodent (lanes 8–12), werealso compared after prior enrichment by specific amp-lification of 16S rDNA of β-proteobacteria (Fig. 5A),α-proteobacteria (Fig. 5B), and High G+C Gram-positives (Fig. 5C). The highest variability betweendifferent lanes was observed for β-proteobacterialpopulations. The number of dominant bands (up toeight bands) and their electrophoretic mobility washighly variable. Only two bands were visible in alllanes. However, no band could be identified whichonly occurred in Nitroflint or Nitrodent. None of thebands in the β-proteobacterial patterns matched theelectrophoretic mobility of dominant bands in the eu-bacterial TGGE patterns (lane 2, Nitroflint; lane 3,
Nitrodent), indicating that β-proteobacteria do not be-long to the dominant populations in the rhizosphereof young maize. In contrast to the β-proteobacterialpatterns less variability was observed for the TGGEpatterns of α-proteobacteria (Fig. 5B) or high G+CGram-positives (Fig. 5C). Several dominant bands inthe α-proteobacterial patterns running with the elec-trophoretic mobility of A. tumefaciens and abovematched with the electrophoretic mobility of domin-ant bands in the eubacterial pattern (Fig. 5B; lanes 2and 13). This result suggests that α-proteobacteria be-long to the dominant populations in the rhizosphere ofyoung maize plants. Again, the two most intense bandsof the high G+C patterns (Fig. 5C) can be assignedto dominant bands in the eubacterial community pat-tern. Interestingly, the intensity of both bands in thehigh G+C patterns is similar, the lower band is evenmore intense while in the eubacterial patterns the up-per band is clearly more intense. This might suggestthat another non-high G+C population contributes tothis dominant band. In contrast to the eubacterial com-munity patterns several bands of high G+C popula-tions are visible also in the lower part of the gel. Thesepopulations were obviously not abundant enough to bedirectly amplified with primers F984GC and R1378.Overall, for none of the populations analysed here dif-ferences between the patterns of both cultivars couldbe detected.
Discussion
We have explored the potential of TGGE fingerprintsbased on 16S rDNA fragments amplified by PCR fromcommunity DNA to study the population dynamics inthe rhizosphere of two maize cultivars grown in trop-ical soils over the growing season. The objective was
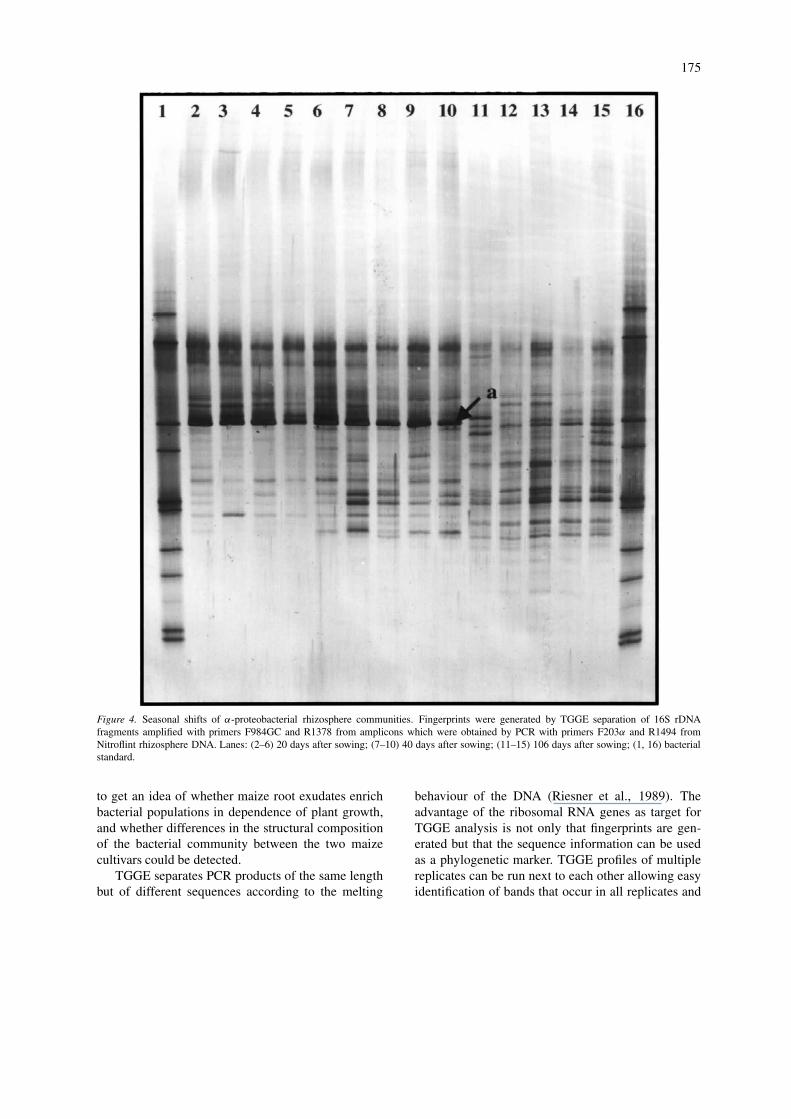
175
Figure 4. Seasonal shifts of α-proteobacterial rhizosphere communities. Fingerprints were generated by TGGE separation of 16S rDNAfragments amplified with primers F984GC and R1378 from amplicons which were obtained by PCR with primers F203α and R1494 fromNitroflint rhizosphere DNA. Lanes: (2–6) 20 days after sowing; (7–10) 40 days after sowing; (11–15) 106 days after sowing; (1, 16) bacterialstandard.
to get an idea of whether maize root exudates enrichbacterial populations in dependence of plant growth,and whether differences in the structural compositionof the bacterial community between the two maizecultivars could be detected.
TGGE separates PCR products of the same lengthbut of different sequences according to the melting
behaviour of the DNA (Riesner et al., 1989). Theadvantage of the ribosomal RNA genes as target forTGGE analysis is not only that fingerprints are gen-erated but that the sequence information can be usedas a phylogenetic marker. TGGE profiles of multiplereplicates can be run next to each other allowing easyidentification of bands that occur in all replicates and
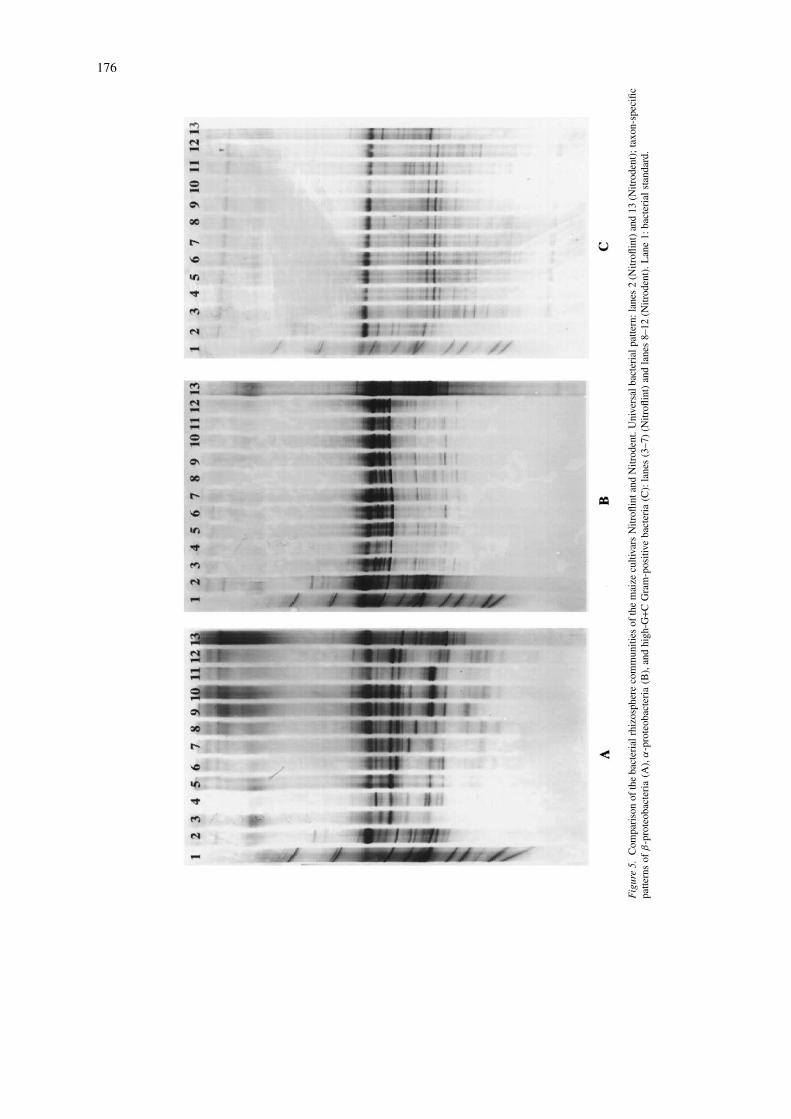
176
Fig
ure
5.C
ompa
riso
nof
the
bact
eria
lrhi
zosp
here
com
mun
ities
ofth
em
aize
culti
vars
Nitr
oflin
tand
Nitr
oden
t.U
nive
rsal
bact
eria
lpat
tern
:lan
es2
(Nitr
oflin
t)an
d13
(Nitr
oden
t);t
axon
-spe
cific
patte
rns
ofβ
-pro
teob
acte
ria
(A),
α-p
rote
obac
teri
a(B
),an
dhi
gh-G
+C
Gra
m-p
ositi
veba
cter
ia(C
):la
nes
(3–7
)(N
itrofl
int)
and
lane
s8–
12(N
itrod
ent)
.Lan
e1:
bact
eria
lst
anda
rd.

177
populations that are not present in all samples. If sucha block of replicates taken at a sampling time is fol-lowed by a block of replicates taken at a later stageof plant development, populations appearing or disap-pearing from this fingerprint can be easily identified byeye. Similarly, as for traditional cultivation approacheswhere colonies are usually picked from plates of di-lutions giving 10–100 colonies for further character-isation, the TGGE patterns of 16S rDNA fragmentsobtained with eubacteria-specific primers are finger-prints of the most dominant populations (Muyzer etal., 1993; Heuer and Smalla, 1997). However, a directcorrelation between the intensity of a band and the cellnumber is not possible because bacteria harbour differ-ent numbers of 16S rDNA operons (Fogel et al., 1999;Klappenbach et al., 2000). The intensity of a TGGEband reflects the abundance of 16S rDNA fragmentssharing an identical melting behaviour. The nestedPCR approach of using taxon-specific primers in afirst PCR followed by the TGGE-PCR with eubacterialprimers not only reduces the complexity of the patternbut also allows analysis of minority populations.
In the present study traditional cultivation tech-niques were combined with TGGE analysis of 16SrDNA fragments amplified from DNA extracted dir-ectly from roots with tightly adhering soil. Due tothe extraction method applied, rhizoplane and endo-phytic bacterial populations could have contributedto the rhizosphere TGGE fingerprints as well. How-ever, the cfu of endophytic bacteria (data not shown)were three orders of magnitude lower than observedrhizosphere bacterial counts, and thus it is unlikelythat endophytic bacterial populations contributed tothe TGGE profiles. The rhizosphere effect, namelythe enrichment of bacterial population possibly dueto root exudates, was found to be most pronouncedfor young roots. A few, intense bands were present intheir TGGE patterns while for soil many faint bandswere observed. This observation indicates that a fewpopulations were numerically dominant in the rhizo-sphere of young maize plants (low evenness) whilein soil and in the rhizosphere of mature maize plantsmany were equally abundant (high diversity) withthe exception of one more abundant population. Thispopulation seems to be dominant in soil and in themaize rhizosphere at different plant growth stages. Se-quencing of clones obtained after amplification of thecomplete 16S rDNA from the rhizosphere of youngplants, that co-migrated with the major band in soiland maize rhizosphere TGGE patterns, showed thatthese populations belonged to the genus Arthrobac-
ter. Furthermore, the sequence revealed that all cloneshad sequence differences mainly in the V3 but alsoin the V6 region, thus the band probably represen-ted a variety of different Arthrobacter strains. Brimet al. (1999) also reported that all Arthrobacter strainsisolated from a Zn-contaminated soil, which showedan identical electrophoretic mobility of the 16S rDNAfragment, displayed a higher degree of diversity basedon their PCR-REP and their substrate utilisation pat-terns. This is an example that a large diversity mightbe hidden behind a TGGE-band. Microbacterium sp.was identified as predominant DNA band in the rhizo-sphere of barley by Yang and Crowley (2000), and alsocomprised 9% of the culturable bacteria in the rhizo-sphere of sugar beet (Lilley et al., 1996). Mahaffee andKloepper (1997) found that Bacillus and Arthrobacterspp. were dominant culturable aerobic bacteria isol-ated from soil, while Gram-negative bacteria tendedto be more abundant in the rhizosphere of field-growncucumber. In addition, Miller et al. (1989) have repor-ted the predominance of Actinomycetes in the maizerhizosphere, based on cultivation. Among the explan-ations suggested for the numerical predominance ofArthrobacter populations in soil is their extreme res-istance to drying and their nutritional versatility (Jonesand Keddie, 1992). However, the role of Arthrobac-ter populations and other coryneform genera in therhizosphere is largely unknown.
For both cultivars, the eubacterial TGGE patternsobserved at different growth stages indicated seasonalshifts of the bacterial community. In general, a higherproportion of equally abundant populations was foundfor mature plants. Seasonal shifts seem to be lesspronounced for high G+C Gram-positives than thoseobserved with eubacterial, α- and β-proteobacterialpatterns. However, we cannot exclude that the kindof roots, in particular the proportion of fine roots,collected in a composite sample might have affectedseasonal shifts observed. Temporal and spatial vari-ation of root exudates has been shown by Jaeger etal. (1999) for the annual grass Avena barbata that re-leased tryptophan from older roots while sucrose wasmost abundant in soil near the root tip. In contrast tothe data reported here, Duineveld et al. (1998) foundthat the DGGE patterns of bacterial populations inthe rhizosphere of the chrysanthemum had very littlevariation over time and between different root partsat different ages. Distinct and very consistent DGGEpatterns in different root zones of barley were repor-ted by Yang and Crowley (2000). In the present studythe TGGE patterns of maize rhizosphere populations

178
were most variable for young roots. Strong bands ap-pearing only in the TGGE patterns for rhizosphere ofyoung maize plants were not always detected in allreplicates. Yang and Crowley (2000) also observed avariability in unique bands and suggested that certaincomponents in the rhizosphere might occur randomly.Furthermore, the authors reported on the appearanceof multiple taxonomically different bacteria at thesame location in the gel.
While the cfu numbers of Nitroflint and Nitrodentrhizosphere samples taken 40 days after sowing weresignificantly different (0.02), no differences were de-tected when the eubacterial and group-specific TGGEpatterns were compared for rhizosphere samples takenafter 20 and 40 days (data not shown). Lottmann et al.(2000) could not detect differences in the rhizospherebacterial community of transgenic T4-lysozyme pro-ducing potatoes and the wild-type based on a com-parison of DGGE fingerprints while the method wassensitive enough to detect seasonal shifts of dominantbacterial populations.
While we have studied the seasonal shifts ofdominant bacteria or of populations of α- and β-proteobacteria or high G+C-bacteria in the maizerhizosphere in a holistic approach, other groups ana-lysed the maize rhizosphere communities at differ-ent growth stages based on bacterial isolates. Thesestudies mainly focussed on plant growth-promotingrhizobacteria isolated from maize such as fluorescentpseudomonads (Picard et al., 2000), Burkholderia ce-pacia (Di Cello et al., 1997, Dalmastri et al., 1999),Paenibacillus polymyxa (Seldin et al., 2000), 2,4-Diacetylphloro-glucinol-producing bacteria (Picard etal., 2000) and maize endophytes (Chelius and Triplett,2000). Although the ‘observation window’ and inparticular the level of resolution between the two ap-proaches is clearly different, many findings observedfor selected isolates are consistent with those ob-tained with the holistic community-level approach.The analysis of RAPD patterns generated from B.cepacia isolates revealed a high level of genetic di-versity in these B. cepacia populations which repres-ented 4% of the total culturable fraction. B. cepaciaseemed to be strictly associated with the rhizosphere(Di Cello et al., 1997). A higher degree of poly-morphism found for B. cepacia isolated during theearly stages of maize growth was also reported byDi Cello et al. (1997). These authors suggested thatthe higher genetic diversity might be due to the factthat the rhizosphere of young plants represents a moreunstable ecosystem than does the rhizosphere of ma-
ture plants. The genetic diversity among B. cepaciaassociated with the root system of maize was ana-lysed by RAPD patterns to assess the influence ofsoil type and maize cultivar on the degree of theirdiversity. The degree of diversity was significantlyhigher between B. cepacia populations recovered frommaize planted in different soils than between B. cepa-cia populations from different cultivars (Dalmastri etal., 1999). Picard et al. (2000) followed the abund-ance and biodiversity of 2,4-Diacetylphloroglucinol(DAPG)-producing bacteria at different stages of plantgrowth. The stage of plant growth seemed to signific-antly affect the abundance of DAPG producers and thelevel of genetic diversity (mostly Pseudomonas fluor-escens). DAPG producers were detected in the maizerhizosphere during all stages of plant developmentbut were present at low density at the first stage ofmaize growth. In comparison to the degree of diversitywhich can be found when PCR-fingerprints of bac-terial isolates or their metabolic genes are compared(e.g., nifH) it is obvious that profiling of rhizospheresbased on PCR-amplified 16S rDNA fragments has amuch lower level of resolution.
Despite several pitfalls of PCR-based rRNA ana-lysis (Heuer and Smalla, 1997; von Wintzingerode etal., 1997; Suzuki et al., 1998), profiling of bacterialrhizosphere communities by denaturing gradient gelsproved to be a powerful method for rapidly analys-ing temporal and spatial variation in the rhizosphere.However, prediction of ecological characteristics ofa bacterial population based on the sequence of a16S rRNA gene fragment used for TGGE is not pos-sible. Either the limited sequence database may lacka well-studied reference strain related closely enoughto have a similar phenotype, or the resolution of the16S rDNA fragment is not high enough to distinguishbetween closely related bacteria differing in the traitof interest, e.g., aggressiveness to infect a plant, mak-ing it necessary to search for actually present strains.Thus, PCR-TGGE analysis should be applied in a mul-tiphasic approach together with traditional cultivationand sequencing of a representative clone library. Astrategy for linking 16S rDNA from community fin-gerprints to pure isolates was recently proposed byHeuer et al. (1999). Although new nucleic acid-basedtechniques have a great potential and allow a new di-mension in studying rhizosphere and soil microbiotathe tools presently available have limitations. In partic-ular, tools to study the functional diversity in situ, e.g.,by mRNA analysis to follow the expression of geneswhich might be of importance for plant health and

179
soil quality are not yet fully available. In comparisonto this our ability to analyse the structural composi-tion of complex bacterial communities is much moreadvanced.
Acknowledgements
This study was financed by a bilateral (Ger-man/Brazilian) cooperation in science and technology(WTZ98/005).The authors are particularly grateful forthe close collaboration with EMBRAPA (EmpresaBrasileira de Pesquisa Agropecuária), Seropédica, Riode Janeiro state, where the field testing of Nitroflintand Nitrodent was performed. Support for consum-ables was provided by MONSANTO to K. Smalla.The development of β-proteobacteria-specific primerswas done in the FATE project (FAIR5-CT97-3632).
References
Amann R I, Ludwig W and Schleifer K-H 1995 Phylogenetic identi-fication and in situ detection of individual microbial cells withoutcultivation. Microbiol. Rev. 59, 143–169.
Brim H, Heuer H, Krögerrecklenfort E, Mergeay M and SmallaK 1999 Characterization of the bacterial community of a zinc-polluted soil. Can. J. Microbiol. 45, 326–338.
Chelius M K and Triplett E W 2000 Immunolocalization of dinitro-genase reductase produced by Klebsiella pneumoniae in associ-ation with Zea mays L. Appl. Environ. Microbiol. 66, 783–787.
Dalmastri C, Chiarini L, Cantale C, Bevivino A and Tabaccioni1999 Soil type and maize cultivar affect the genetic diversityof maize root-associated Burkholderia cepacia populations. Mi-crob. Ecol. 38, 273–284.
De Leij F A A M, Whipps J M and Lynch J M 1994 Theuse of colony development for the characterization of bacterialcommunities in soil and on roots. Microb. Ecol. 27, 81–97.
Di Cello F, Bevivino A, Chiarini L, Fani R, Paffetti D, TabacchioniS and Dalmastri C 1997 Biodiversity of a Burkholderia cepaciapopulation isolated from the maize rhizosphere at different plantgrowth stages. Appl. Environ. Microbiol. 63, 4485–4493.
Duineveld B M, Rosado A S, Van Elsas J D and Van Veen J A1998 Analysis of the dynamics of bacterial communities in therhizosphere of the Chrysanthemum via denaturing gradient gelelectrophoresis and substrate utilization patterns. Appl. Environ.Microbiol. 64, 4950–4957.
Engelhard M, Hurek T and Reinhold-Hurek B 2000 Preferentialoccurrence of diazotrophic endophytes, Azoarcus spp., in wildrice species and land races of Oryza sativa in comparison withmodern races. Environ. Microbiol. 2, 131–141.
Fogel G B, Collins C R, Li J and Brunk C F 1999 Prokaryoticgenome size and SSU rDNA copy number. Microb. Ecol. 38,93–113.
Gelsomino A, Keijzer-Wolters A, Cacco G and van Elsas J D1999 Assessment of bacterial community structure in soil bypolymerase chain reaction and denaturing gradient gel electro-phoresis. J. Microbiol. Methods 38, 1–15.
Grayston S J, Wang S, Campbell C D and Edwards A C 1998 Se-lective influence of plant species on microbial diversity in therhizosphere. Soil Biol. Biochem. 30, 369–378.
Heuer H and Smalla K 1997 Application of denaturing gradientgel electrophoresis (DGGE) and temperature gradient gel elec-trophoresis (TGGE) for studying soil microbial communities. InModern soil microbiology. Eds J D van Elsas, E M H Wellingtonand J T Trevors. pp 353–373. Marcel Dekker, New York.
Heuer H, Krsek M, Baker P, Smalla K and Wellington E M H1997 Analysis of actinomycete communities by specific amp-lification of genes encoding 16S rRNA and gel-electrophoreticseparation in denaturing gradients. Appl. Environ. Microbiol. 63,3233–3241.
Heuer H, Hartung K, Wieland G, Kramer I and Smalla K 1999Polynucleotide probes that target a hypervariable region of 16SrRNA genes to identify bacterial isolates corresponding to bandsof community fingerprints. Appl. Environ. Microbiol. 65, 1045–1049.
Höflich G, Wiehe W and Kühn G 1994 Plant growth stimulation byinoculation with symbiotic and associative rhizosphere microor-ganisms. Experientia 50 (Birkhäuser Verlag, Basel) 897–905.
Horwath W R, Elliott L F and Lynch J M 1998 Influence of soilquality on the function of inhibitory rhizobacteria. Lett. Appl.Microbiol. 26, 87–92.
Jaeger C H III, Lindow S E, Miller W, Clark E and Firestone MK 1999 Mapping of sugar and amino acid availability in soilaround roots with bacterial sensors of sucrose and tryptophan.Appl. Environ. Microbiol. 65, 2685–2690.
Jones D and Keddie R M 1992 The Genus Arthrobacter. In TheProkaryotes, Vol. 2, 2nd edition. Eds A Balows, H G Trüp-per, M Dworekin, W Harder and K-H Schleifer. pp 1283–1299,Springer, New York.
Klappenbach J A, Dunbar J M and Schmidt T M 2000 rRNA op-eron copy number reflects ecological strategies of bacteria. Appl.Environ. Microbiol. 66, 1328–1333.
Latour X, Philippot L, Corberand T and Lemanceau P 1996 The es-tablishment of an introduced community of fluorescent pseudo-monads in the soil and in the rhizosphere is affected by the soiltype. FEMS Microbiol. Ecol. 30, 163–170.
Lilley, A K, Fry, J C, Bailey, M J and Day M J 1996 Comparisonof aerobic heterotrophic taxa isolated from four root domains ofmature sugar beet (Beta vulgaris). FEMS Microbiol. Ecol. 21,231–242.
Lottmann, J, Heuer H, de Vries J, Mahn A, Düring K, WackernagelW, Smalla K and Berg G 2000 Establishment of introduced ant-agonistic bacteria in the rhizosphere of transgenic potatoes andtheir effect on the bacterial community. FEMS Microb. Ecol. 33,41–49.
Lupwayi N Z, Rice W A and Clayton GW 1998 Soil microbial di-versity and community structure under wheat as influenced bytillage and crop rotation. Soil Biol. Biochem. 30, 1733–1741.
Mahaffee W F and Kloepper J W 1997 Temporal changes in the bac-terial communities of soil, rhizosphere, and endorhiza associatedwith field grown cucumber (Cucumis sativus L.). Microb. Ecol.34, 210–223.
Miller H J, Henken G and Van Veen J A 1989 Variation and com-position of bacterial populations in the rhizospheres of maize,wheat, and grass cultivars. Can. J. Microbiol. 35, 656–660.
Muyzer G and Smalla K 1998 Application of denaturing gradientgel electrophoresis (DGGE) and temperature gradient gel elec-trophoresis (TGGE) in microbial ecology. Antonie van Leeuwen-hoek 73, 127–141.
Muyzer G, de Waal E C and Uitterlinden A G 1993 Profilingof complex microbial populations by denaturing gradient gel

180
electrophoresis analysis of polymerase chain reaction-amplifiedgenes encoding for 16S rRNA. Appl. Environ. Microbiol. 59,695–700.
Nübel U, Engelen B, Felske A, Snaidr J, Wiesenhuber A, Amann RI, Ludwig W and Backhaus H 1996 Sequence heterogeneities ofgenes encoding 16S rRNAs in Paenibacillus polymyxa detectedby temperature gradient gel electrophoresis. J. Bacteriol. 178,5636–5643.
Picard C, Di Cello F, Ventura M, Fani R and Guckert A 2000 Fre-quency and biodiversity of 2,4-diacetylphloroglucinol-producingbacteria isolated from the maize rhizosphere at different stages ofplant growth. Appl. Environ. Microbiol. 66, 948–955.
Riesner D, Steger G, Zimmat R, Owens R A, Wagenhöfer M, HillenW, Vollbach S and Henco K 1989 Temperature-gradient gel elec-trophoresis of nucleic acids: analysis of conformational trans-itions, sequence variations, and protein–nucleic acid interactions.Electrophoresis 10, 377–389.
Rychlik W and Rhoads R E 1989 A computer program for choos-ing optimal oligonucleotides for filter hybridization, sequencingand in vitro amplification of DNA. Nucleic Acids Res. 17,8543–8551.
Sambrook J, Fritsch E F and Maniatis T 1989 Molecular Cloning: aLaboratory Manual. Cold Spring Harbor Laboratory Press, ColdSpring Harbor, NY.
Seldin L, Rosado A S, Da Cruz D W, Nobrega A, Van Elsas J D andPaiva E 2000 Comparison of Paenibacillus azotofixans strainsisolated from rhizoplane, rhizosphere, and non-root-associatedsoil from maize planted in two different Brazilian soils. Appl.Environ. Microbiol. 64, 3860–3868.
Sheffield, V C, Cox R D, Lerman L S and Myers R M 1989. At-tachment of a 40-base-pair G+C-rich sequence (GC-clamp) togenomic DNA fragments by the polymerase chain reaction res-ults in improved detection of single-base changes. Proc. Natl.Acad. Sci. USA 86, 232–236.
Smalla K and Van Elsas J D 1995 Application of the PCR for de-tection of antibiotic resistance genes in environmental samples.In Nucleic Acids in the Environment. Eds J T Trevors, J D vanElsas. pp 241–256. Springer, Berlin, Heidelberg.
Smalla K, Cresswell N, Mendonca-Hagler L C, Wolters A andVan Elsas J D 1993 Rapid DNA extraction protocol from soilfor polymerase chain reaction-mediated amplification. J. Appl.Bacteriol. 74, 78–85.
Staley, J T and Konopka A 1985. Measurement of in situ activitiesof nonphotosynthetic microorganisms in aquatic and terrestrialhabitats. Annu. Rev. Microbiol. 39, 321–346.
Suzuki M, Rappé M S and Giovannoni S J 1998 Kinetic bias inestimates of coastal picoplankton community structure obtainedby measurements of small-subunit rRNA gene PCR ampliconlength heterogeneity. Appl. Environ. Microbiol. 64, 4522–4529.
Van Elsas, J D and Smalla K 1996 Methods for sampling soilmicrobes. In Manual of Environmental Microbiology. Eds C JHurst, G R Knudsen, M J McInerney, L D Stetzenbach and M VWalter. pp 383–390. ASM Press, Washington, DC.
Van Loon L C, Bakker P A H M and Pieterse C M J 1998 Sys-temic resistance induced by rhizosphere bacteria. Annu. Rev.Phytopathol. 36, 453–483.
von Wintzingerode F, Göbel U B and Stackebrandt E 1997 De-termination of Microbial Diversity in Environmental Samples:Pitfalls of PCR-based rRNA Analysis. FEMS Microbiol. Rev. 21,213–229.
Watkinson A R 1998 The role of the soil community in plantpopulation dynamics. TREE 13, 171–172.
Weisburg W G, Barns S M, Pelletier D A and Lane D J 1991 16S ri-bosomal DNA amplification for phylogenetic study. J. Bacteriol.173, 697–703.
Westover K M, Kennedy A C and Kelley S E 1997 Patternsof rhizosphere microbial community structure associated withco-occurring plant species. J. Ecol. 85, 863–873.
Wilson K 1987 Preparation of genomic DNA from bacteria. In Cur-rent Protocols in Molecular Biology. Eds J A Ausubel, R Bent, RE Kingston, D D Moore, J A Smith, J G Seidmann and K Struhl.pp 2.10–2.12. Wiley, New York, NY.
Yang C-H and Crowley D E 2000 Rhizosphere microbial com-munity structure in relation to root location and plant ironnutritional status. Appl. Environ. Microbiol. 66, 345–351.




















