
C-reactive protein induces human peripheral blood monocytes to synthesize tissue factor
Upload
independentCategory
view
0download
0
ARTHRITIS & RHEUMATISMVol. 58, No. 11, November 2008, pp 3377–3387DOI 10.1002/art.23966© 2008, American College of Rheumatology
Mature Antigen-Experienced T Helper Cells Synthesize andSecrete the B Cell Chemoattractant CXCL13 in theInflammatory Environment of the Rheumatoid Joint
Antonio Manzo,1 Barbara Vitolo,2 Frances Humby,1 Roberto Caporali,3 David Jarrossay,4
Francesco Dell’Accio,1 Laura Ciardelli,2 Mariagrazia Uguccioni,4
Carlomaurizio Montecucco,3 and Costantino Pitzalis1
Objective. Synovial B cells play a critical role inrheumatoid arthritis (RA), being involved in autoanti-body synthesis, T cell activation, and cytokine produc-tion. CXCL13 is a B cell chemoattractant that is instru-mental in synovial B cell organization; the regulatorydeterminants of CXCL13 in inflammation are poorlycharacterized. This study was undertaken to investigatethe functional involvement of synovial T cells in theectopic expression of CXCL13 in RA.
Methods. CXCL13 production and regulationwere addressed using immunohistochemistry, in situhybridization, quantitative polymerase chain reaction,multicolor flow cytometry, and enzyme-linked immu-
nosorbent assay, by in situ–ex vivo analysis and in vitrofunctional assays with rheumatoid synovial tissue andprimary cells.
Results. CXCL13 messenger RNA and proteinexpression and spontaneous CXCL13 secretion weredetected in RA synovial fluid T cells but were notdetected (or were detected only occasionally) in peri-pheral blood T cells. Analysis of tissue expressionconfirmed cytoplasm localization of CXCL13 in T lym-phocytes infiltrating B cell follicles and small peri-vascular aggregates. Multicolor characterizations insynovial fluid demonstrated CXCL13 expression inantigen-experienced T helper cells, frequently charac-terized by terminal differentiation and the lack of thefollicular helper T cell markers CXCR5 and BCL6protein. In vitro functional assays revealed the enhanc-ing effect of T cell receptor–CD28 engagement onCXCL13 production and secretion in primary cells.
Conclusion. Our findings define a new functionalproperty of synovial T cells, demonstrating their activeinvolvement in the local production of B cell chemoat-tractants, and support a direct contribution of theadaptive immune system and antigen-dependent signalsin the mechanisms of B cell localization in RA.
The recruitment and organization of B cells inthe synovium are common events in rheumatoid arthritis(RA) and are thought to play a critical role in diseasepathogenesis (1,2), by promoting autoantibody produc-tion (3) and the expression of proinflammatory cyto-kines through interaction with resident T cells (4).CXCL13 is a homeostatic B cell chemoattractant (5)that is required for the normal organization of B cellfollicles in several secondary lymphoid organs (6) and issufficient to promote B cell recruitment and ectopic
Dr. Manzo’s work was supported by Guy’s and St Thomas’Charity and the European Union’s Sixth Framework Programme(INNOCHEM, grant LSHB-CT-2005-518167). Ms Vitolo’s and Dr.Montecucco’s work was supported by the IRCCS Policlinico SanMatteo Foundation. Dr. Humby’s work was supported by Guy’s and StThomas’ Charity. Dr. Uguccioni’s work was supported by the HelmutHorten Foundation, the Swiss National Science Foundation, and theEuropean Union’s Sixth Framework Programme (INNOCHEM, grantLSHB-CT-2005-518167). Dr. Pitzalis’ work was supported by theEuropean Union’s Sixth Framework Programme (INNOCHEM, grantLSHB-CT-2005-518167).
1Antonio Manzo, MD, PhD, Frances Humby, MBBS,Francesco Dell’Accio, MD, PhD, Costantino Pitzalis, MD, PhD,FRCP: St. Bartholomew’s and Royal London School of Medicine,London, UK; 2Barbara Vitolo, BSc, Laura Ciardelli, BSc: IRCCSPoliclinico San Matteo, Pavia, Italy; 3Roberto Caporali, MD, Carlo-maurizio Montecucco, MD: IRCCS Policlinico San Matteo, andUniversita’ degli Studi di Pavia, Pavia, Italy; 4David Jarrossay, PhD,Mariagrazia Uguccioni, MD: Institute for Research in Biomedicine,Bellinzona, Switzerland.
Address correspondence and reprint requests to CostantinoPitzalis, MD, PhD, FRCP, Professor of Experimental Medicine andRheumatology, Centre for Experimental Medicine and Rheumatol-ogy, 2nd Floor John Vane Science Centre, William Harvey ResearchInstitute, St. Bartholomew’s and Royal London School of Medicine,Charterhouse Square, London EC1M6BQ, UK. E-mail:[email protected].
Submitted for publication August 23, 2007; accepted inrevised form July 14, 2008.
3377

lymphoid neogenesis when overexpressed in extralym-phoid sites (7).
Accordingly, CXCL13 is induced in the rheuma-toid joint, where it associates with the development of Bcell follicles and germinal center (GC) reactions in thesynovium (8–10) and subchondral bone marrow (11),correlating in terms of plasma levels with disease activity(12). In vivo evidence of the pathogenic relevance of theCXCL13–B cell axis has been recently provided incollagen- and antigen-induced arthritis, in whichCXCL13 blockade or CXCR5 knockout has been shownto interfere with synovial and splenic follicle organiza-tion, thereby improving disease outcome (13,14). Under-standing the mechanisms acting upstream of CXCL13induction in inflammation is critical to define the aber-rant dynamics regulating pathologic B cell interactionsin rheumatoid synovitis.
During embryonic development of murine lym-phoid organs, CXCL13 is physiologically produced bymesenchymal cells upon the action of lymphotoxin �,which is expressed by infiltrating hematopoietic precur-sors (15). Although similar pathways have been con-firmed postnatally (6), recent studies provide evidencethat CXCL13 production can be determined by alterna-tive mechanisms in humans, such as by a direct effect ofimmunologic triggers in cells of the adaptive immunesystem. Indeed, it has been shown that in tonsil GCs,CXCL13 can be directly expressed by T lymphocytes and
induced upon cognate interaction with antigen-presenting cells and T cell receptor (TCR) engagement(16,17). These observations suggest an important func-tion of CXCL13, indicating its capacity to be plasticallymodulated during the immune response, acting as adirect T cell–B cell signal transducer.
Whether this process is physiologically restrictedto tonsils or whether normal T cells can produce B cellchemoattractants in extralymphoid inflammatory reac-tions in humans is, at present, unclear. This issue is ofconsiderable relevance in RA, because synovial T cellsand B cells have been shown to be tightly interconnectedin terms of reciprocal activation (4,18–20), while theprocesses promoting their interaction are poorly charac-terized.
In this report, we challenged the hypothesis thatthis biologic mechanism can be activated in RA synovi-tis, by investigating the capacity of synovial T cells toproduce CXCL13 and analyzing the relationship be-tween TCR–CD28 engagement and CXCL13 produc-tion directly in synovial primary cells.
PATIENTS AND METHODS
Patients and samples. Fifteen synovial tissue speci-mens collected from patients undergoing synoviectomy, arthro-scopic biopsy, or joint replacement were selected for thepresence of mononuclear infiltrates and grade 3 lymphoid
Table 1. Clinical and histologic features of the study population*
Patient/age/sex
Diseaseduration,months IgM-RF
ESR,mm/hour
CRP,mg/liter Erosions† FDCs
DiffuseT cell
infiltration‡
CXCL13�T cell
density§
% CXCL13� T cells
Of subliningT cells¶
Of totalCXCL13� cells
RA1/66/M 360 � NA 3.03 � – 3 1.6 2.50 52RA2/46/F 204 � 88 9.20 � – 3 0.6 0.47 31.71RA3/57/F 120 � 42 0.89 � – 1 0.2 0.88 15.38RA4/44/F 156 � 45 6.50 � � 3 1.2 2.00 28.85RA5/79/F 120 � 85 2.67 � � 1 0.8 0.41 13.85RA6/23/F 408 � 35 NA � � 2 0.4 1.82 22RA7/65/F 144 – 35 0.30 � – 3 1.02 1.20 46.81RA8/75/F 84 – 21 0.30 � – 1 0.15 0.53 9.09RA9/41/M 168 � 38 1.01 � – 3 1.2 2.95 38.30RA10/68/F 12 – 69 5.66 � � 3 2.6 1.55 41.03RA11/61/F 120 � 71 2.72 � – 2 4.2 3.31 29.9RA12/76/F 276 – 65 2.40 � � 2 3 0.69 20.31RA13/66/M 132 � 17 0.30 – – 1 0.11 1.63 7.69RA14/29/F 180 – 120 3.37 � – 3 0.51 1.44 37.5RA15/62/F 2 – 50 5.44 � � 3 1.97 1.40 33.93
* IgM-RF � IgM rheumatoid factor; ESR � erythrocyte sedimentation rate; CRP � C-reactive protein; FDCs � CD21� follicular dendritic cellnetworks; RA � rheumatoid arthritis; NA � not available.† Assessed on standard radiographs of the hands and feet.‡ Assessed by a semiquantitative score based on a 4-point scale, where 0 � occasional diffuse CD3� cells and 3 � infiltration by numerous diffuseCD3� cells.§ Number of cells per microscopic high-power field (see Patients and Methods).¶ See Patients and Methods (Histologic analyses).
3378 MANZO ET AL

aggregates (10) (see Table 1). Ten synovial fluid (SF) samples(�20 ml) obtained by knee arthrocentesis from patients withclinically involved joints (mean age 52 years [range 37–71years], mean disease duration 7 years [range 1–13 years]) werealso included. Peripheral blood (PB) was obtained just beforearthrocentesis. All patients fulfilled the American College ofRheumatology (formerly, the American Rheumatism Associ-ation) 1987 revised criteria for the classification of RA (21)and were treated with steroids and disease-modifying drugs,including hydroxychloroquine, methotrexate, and cyclosporine.Normal PB specimens, tonsils, and normal synovium wereobtained from 4 healthy volunteers, 3 patients undergoingtonsillectomy, and 2 patients undergoing lower limb amputa-tion. Synovial tissue specimens were also obtained from 2patients with osteoarthritis (OA) who were undergoing kneejoint replacement. All samples were collected after obtaininginformed consent and approval by the local ethics committee.
Immunohistologic analysis. The following primary an-tibodies were used for immunohistochemical analysis andimmunofluorescence testing: mouse anti-human CD20 (cloneL26; Dako, Glostrup, Denmark), rabbit anti-human CD3(immunoglobulin; Dako), mouse anti-human CD68 (cloneKP1; Dako), mouse anti-human CD21 (clone 1F8; Dako),mouse anti-human CD8 (clone 1A5; Novocastra, Newcastle,UK), mouse anti-human CD45RO (clone UCHL1; Dako),mouse anti-human CD69 (clone CH11; Novocastra), and goatanti-human CXCL13 (IgG; R&D Systems, Abingdon, UK). Asnegative controls, normal goat IgG (R&D Systems) and irrel-evant isotype-matched antibodies (Dako) were used. All ap-propriate secondary antibodies were obtained from Dako,LiStarFish (Milan, Italy), or Molecular Probes (Leiden, TheNetherlands).
Tissue specimens were formalin-fixed, paraffin-embedded, and cut in consecutive 4-�m sections. The sectionswere then deparaffinized and rehydrated through gradedethanols. Antigen retrieval was performed by heating for 35minutes at 96°C in Dako Target Retrieval Solution or, forCD21, by incubating the sections for 20 minutes at 37°C withprewarmed 0.05% Pronase (Dako). Sections were then washedin Tris buffered saline (TBS) and incubated with Protein Block(Dako). For double immunohistochemistry, the sections wereincubated with the cell marker–specific primary antibodies,with the appropriate peroxidase-conjugated secondary anti-bodies, and then developed with diaminobenzidine (Sigma-Aldrich, Zwijndrecht, The Netherlands). Sections were thensequentially incubated with anti-CXCL13 antibodies, withbiotinylated secondary antibodies and with streptavidin–biotincomplex–alkaline phosphatase (Dako), followed by develop-ment with New Fuchsin Substrate (Dako). For immunofluo-rescence testing, sections were first incubated with the cellmarker–specific primary antibodies, followed by Cy5-conjugated secondary antibodies or biotinylated secondaryantibodies with rhodamine red X–conjugated streptavidin(Jackson ImmunoResearch, Bar Harbor, ME). Sections werethen incubated with goat anti-human CXCL13 antibodies andwith Alexa 488–conjugated secondary antibodies. Immunoflu-orescent stainings were analyzed with a fluorescence micro-scope (Olympus, Lake Success, NY) equipped with an infraredcamera. Primary and secondary antibodies were diluted inDako Antibody Diluent.
Histologic analyses. Synovial aggregates were classi-fied as previously described, by calculating the aggregate radial
cell count (10). The prevalence of CXCL13�CD3� cellsamong sublining CD3� cells was estimated by evaluating ineach sample at least 20 randomly selected high-power fields(hpf) (1,000�) in sublining areas characterized by CD3� cellaccumulation. The prevalence of CXCL13�CD3� cellsamong total CXCL13� cells was calculated by analyzing ineach sample at least 20 CXCL13� hpf (1,000�) in randomlyselected areas of the inflamed sublining. The same evaluationin tonsils was performed by dissecting the whole area of 3secondary follicles per specimen. To calculate the density ofCXCL13� T cells in the inflamed sublining, 30–50 randomlyselected hpf (1,000�) per sample were assessed in areascharacterized by mononuclear infiltrate.
In situ hybridization. 35S-labeled sense and antisenseRNA probes were generated by in vitro transcription (RocheMolecular Biochemicals, Indianapolis, IN). The CXCL13 probecorresponded to positions �24 to 344 of the CXCL13 se-quence. Formalin-fixed paraffin-embedded sections were de-waxed, rehydrated in graded ethanol solutions, and subjectedto in situ hybridization, as previously described (22).
Cell separation. SF mononuclear cells (SFMCs) andPB mononuclear cells (PBMCs) were separated by density-gradient centrifugation with Histopaque-1077 Hybri-Max(Sigma, St. Louis, MO) and stored in liquid nitrogen untilused. SF T cells were isolated from SFMCs by positiveselection (isolated synovial T cells) using anti-CD3 microbeads(Miltenyi Biotec, Sunnyvale, CA), according to the manufac-turer’s instructions.
Quantitative polymerase chain reaction (PCR). TotalRNA was extracted using the RNeasy Total RNA Isolation Kit(Qiagen, Chatsworth, CA) and reverse transcribed into com-plementary DNA (cDNA) with the ThermoScript reversetranscription–PCR system (Invitrogen, San Diego, CA). Quan-titative real-time PCR was performed using Hot Start DNApolymerase (Qiagen) in the presence of 0.1� SYBR Green(Molecular Probes), utilizing an ABI 7900HT detector (Ap-plied Biosystems, Foster City, CA). Serial dilutions of lymphnode cDNA were used to generate a standard curve. Geneexpression was calculated using the standard curve and nor-malized to �-actin values. Primers for human CXCL13 (for-ward 5�-TGTGGACCCTCAAGCTGAAT-3�, reverse 5�-CCATCAGCTTCTGCAAGGTT-3�) and �-actin (forward 5�-CACGGCTGCTTCCAGCTC-3�, reverse 5�-CACAGGACTCCATGCCCAG-3�) were designed using VectorDesigner software (Invitrogen).
Immunofluorescence on cytospin slides. SFMCs andisolated SF T cells were washed in TBS, cytocentrifuged onSuperfrost Plus slides (BDH Chemicals, Poole, UK), andair-dried overnight. Before staining, sections were fixed in2.5% paraformaldehyde solution in phosphate buffered saline(PBS) for 20 minutes at room temperature, washed twice inTBS, and blocked with 20% human serum in TBS for 15minutes at room temperature. The slides were then incubatedwith primary antibodies for the specific cell marker and withthe appropriate fluorescein isothiocyanate–conjugated second-ary antibodies. Sections were then permeabilized with 0.3%saponin solution (Sigma-Aldrich) in TBS, incubated with goatanti-human CXCL13 antibodies (IgG; R&D Systems), andwith biotinylated secondary antibodies, followed by rhodaminered X–conjugated streptavidin. Primary and secondary anti-bodies were diluted in Dako Antibody Diluent before perme-abilization and with 0.3% saponin in TBS thereafter. The
CXCL13 EXPRESSION BY SYNOVIAL T CELLS 3379

following primary antibodies were used: rabbit anti-humanCD3 (Dako), mouse anti-human CD4 (clone 4B12; Novocas-tra), mouse anti-human CD8 (clone 1A5; Novocastra), mouseanti-human CD45RO (clone UCHL1; Dako), mouse anti-
human CD45RA (clone HI100; BD PharMingen, San Diego,CA), mouse anti-human CD27 (clone M-T271; BD PharMin-gen), mouse anti-human BCL6 (clone PG-B6p; Dako), andmouse anti-human CD69 (clone CH/4; Caltag, South SanFrancisco, CA). As negative control for CXCL13 staining,normal goat IgG (R&D Systems) was used. All secondaryantibodies were obtained from Dako or LiStarFish.
Fluorescence-activated cell sorting (FACS) analysis.SFMCs and PBMCs were incubated for 4 hours in completeRPMI medium (RPMI 1640, 2 mM glutamine, 100 units/mlpenicillin plus 100 �g/ml streptomycin, 10% heat-inactivatedfetal calf serum; all from Gibco, Grand Island, NY) supple-mented with 10 �g/ml brefeldin A (Sigma). Brefeldin Atreatment was used as a routine procedure adopted in ourlaboratories for optimal staining of intracellular cytokines.Intracellular CXCL13 was detected after fixation and perme-abilization with Cytofix/Cytoperm (BD PharMingen) followedby incubation of the cells with mouse anti-human CXCL13(clone 53610; R&D Systems) and Alexa 488–labeled goatanti-mouse IgG1 (Molecular Probes). Detection of T cell markerswas performed subsequently, after blocking a specific reaction,with 20% mouse serum in PBS. The following antibodies wereused: phycoerythrin (PE)–labeled mouse anti-human CD3 (cloneUCHT1; Beckman Coulter, Fullerton, CA), PC5-labeledmouse anti-human CD4 (clone 13B8.2; Beckman Coulter),allophycocyanin (APC)–labeled mouse anti-human CD4RO(clone UCHL1; BD PharMingen), and APC-labeled mouseanti-human CD45RA (clone HI100; BD PharMingen).
For surface chemokine receptor staining, cells werefirst incubated with either mouse anti-human CXCR5 (clone51505; R&D Systems) or mouse anti-human CCR7 (clone150503; R&D Systems) antibodies, followed by incubation withPE-labeled secondary antibodies (Southern Biotechnology, Bir-mingham, AL). CXCL13 staining was performed subsequently asdescribed above, followed by incubation with PC5-labeled anti-human CD3 (clone UCHT1; Beckman Coulter). Data wereacquired on a FACSCalibur flow cytometer and analyzed withCellQuest software (Becton Dickinson, Mountain View, CA).
Cell cultures. For in vitro stimulation assays, SFMCswere plated in 6-well plates (Corning, Corning, NY) for 28hours at a seeding density of 2 � 106 ml in complete RPMImedium. Next, the cells were removed, washed, and plated inMaxiSorp 96-well plates (Nunc, Naperville, IL) at a seedingdensity of 5 � 105/ml in complete RPMI medium (200 �l/well)or in one-half autologous SF plus one-half complete RPMImedium (200 �l/well). Precoating of the wells was performedwith 2 �g/ml mouse anti-human CD3 (clone TR66 or cloneOKT3; eBioscience, San Diego, CA) plus 2 �g/ml mouseanti-human CD28 (clone CD28.2; BD PharMingen). Cellswere cultured for the appropriate length of time at 37°C, andbrefeldin A (10 �g/ml) was added for the last 4 hours beforeFACS analysis. For analysis of spontaneous CXCL13 secre-tion, isolated SF T cells were cultured in flat-bottomed 96-wellplates (Corning) in 100 �l/well complete RPMI medium at aseeding density of 106 cells/ml. The CXCL13 concentration inthe supernatants was analyzed by CXCL13 Quantikine ELISAkit (R&D Systems), according to the manufacturer’s protocols.
RESULTS
Synthesis and secretion of CXCL13 by synovialprimary T cells. To assess the capacity of synovialprimary T cells to synthesize CXCL13, we isolated SF T
Figure 1. CXCL13 expression/production in synovial fluid T cells. A,Real-time polymerase chain reaction for CXCL13 mRNA expressionin synovial fluid mononuclear cells (SFMCs) and peripheral bloodmononuclear cells (PBMCs) from patients with rheumatoid arthritis(RA) and healthy donors (n). CXCL13 expression levels are normal-ized to �-actin expression levels. Values are the mean and SEM resultsfrom 3 patients or healthy donors per group. B, CXCL13 mRNArelative quantification (RQ) of the T cell fractions (solid bars) isolatedfrom RA SFMCs, the respective unfractionated SFMCs (shaded bars),and the T cell–depleted fractions (open bars) for patient samples SF5,SF7, and SF8. Values are the mean and SEM results from triplicatereactions. C, Double immunofluorescence staining for CD3 andCXCL13 on cytospin slides of the isolated T cell fractions. CXCL13protein with a Golgi-like staining pattern is visualized in 1 of the 3CD3� cells. D, CXCL13 protein expression analysis in the supernatantof isolated synovial fluid T cell cultures from 3 patients (each linerepresents an individual patient) at 36 hours and 60 hours. Circles anderror bars represent the mean � SEM results for triplicate cultures foreach sample at each time point. Color figure can be viewed in theonline issue, which is available at http://www.arthritisrheum.org.
3380 MANZO ET AL
lymphocytes from 3 patients with RA (�95% purity asdetermined by FACS) and analyzed CXCL13 messengerRNA (mRNA) expression by quantitative PCR.CXCL13 was detected in all unfractionated SFMC sam-ples, with mean gene expression levels 2,124.3-foldhigher and 328.6-fold higher compared with normaldonor and paired RA PBMC samples, respectively (Fig-ure 1A). All CD3� fractions obtained following separa-tion showed high levels of CXCL13 mRNA, with a mean4.3-fold increase compared with the respective unfrac-tionated SFMCs and a 19.1-fold increase compared withthe T cell–depleted fractions (Figure 1B). To evaluatewhether CXCL13 mRNA expression was associated withprotein translation in T lymphocytes, we then performeddouble immunofluorescence staining of the same CD3�preparations adopted for mRNA analysis. In all samples,we detected CXCL13 protein in the cytoplasm of exclu-sively T cells, supporting the PCR data (Figure 1C).Finally, to confirm that intracellularly stored CXCL13could be secreted, we cultured SF CD3� cells undernonstimulatory conditions and analyzed the superna-tants by ELISA. We observed CXCL13 protein in allsupernatants tested, with concentrations at 60 hoursranging from 356 pg/ml to 1,749 pg/ml (Figure 1D).
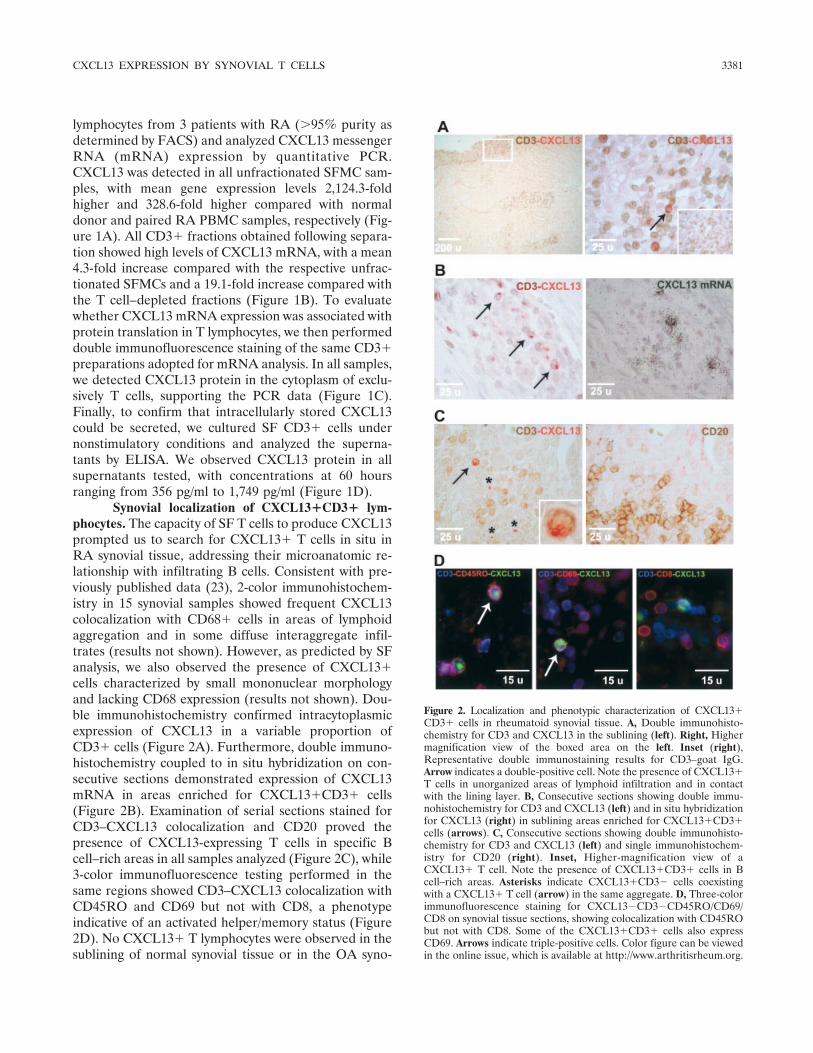
Synovial localization of CXCL13�CD3� lym-phocytes. The capacity of SF T cells to produce CXCL13prompted us to search for CXCL13� T cells in situ inRA synovial tissue, addressing their microanatomic re-lationship with infiltrating B cells. Consistent with pre-viously published data (23), 2-color immunohistochem-istry in 15 synovial samples showed frequent CXCL13colocalization with CD68� cells in areas of lymphoidaggregation and in some diffuse interaggregate infil-trates (results not shown). However, as predicted by SFanalysis, we also observed the presence of CXCL13�cells characterized by small mononuclear morphologyand lacking CD68 expression (results not shown). Dou-ble immunohistochemistry confirmed intracytoplasmicexpression of CXCL13 in a variable proportion ofCD3� cells (Figure 2A). Furthermore, double immuno-histochemistry coupled to in situ hybridization on con-secutive sections demonstrated expression of CXCL13mRNA in areas enriched for CXCL13�CD3� cells(Figure 2B). Examination of serial sections stained forCD3–CXCL13 colocalization and CD20 proved thepresence of CXCL13-expressing T cells in specific Bcell–rich areas in all samples analyzed (Figure 2C), while3-color immunofluorescence testing performed in thesame regions showed CD3–CXCL13 colocalization withCD45RO and CD69 but not with CD8, a phenotypeindicative of an activated helper/memory status (Figure2D). No CXCL13� T lymphocytes were observed in thesublining of normal synovial tissue or in the OA syno-
Figure 2. Localization and phenotypic characterization of CXCL13�CD3� cells in rheumatoid synovial tissue. A, Double immunohisto-chemistry for CD3 and CXCL13 in the sublining (left). Right, Highermagnification view of the boxed area on the left. Inset (right),Representative double immunostaining results for CD3–goat IgG.Arrow indicates a double-positive cell. Note the presence of CXCL13�T cells in unorganized areas of lymphoid infiltration and in contactwith the lining layer. B, Consecutive sections showing double immu-nohistochemistry for CD3 and CXCL13 (left) and in situ hybridizationfor CXCL13 (right) in sublining areas enriched for CXCL13�CD3�cells (arrows). C, Consecutive sections showing double immunohisto-chemistry for CD3 and CXCL13 (left) and single immunohistochem-istry for CD20 (right). Inset, Higher-magnification view of aCXCL13� T cell. Note the presence of CXCL13�CD3� cells in Bcell–rich areas. Asterisks indicate CXCL13�CD3� cells coexistingwith a CXCL13� T cell (arrow) in the same aggregate. D, Three-colorimmunofluorescence staining for CXCL13�CD3�CD45RO/CD69/CD8 on synovial tissue sections, showing colocalization with CD45RObut not with CD8. Some of the CXCL13�CD3� cells also expressCD69. Arrows indicate triple-positive cells. Color figure can be viewedin the online issue, which is available at http://www.arthritisrheum.org.
CXCL13 EXPRESSION BY SYNOVIAL T CELLS 3381

vium characterized by scarce or absent mononuclearinfiltrates, despite the presence of occasional scattered Tcells (results not shown).
CXCL13� T cells were observed in 15 of 15samples analyzed, with a prevalence varying from 0.4%to 3.3% of sublining CD3� cells (mean � SEM 1.5 �0.2%) and ranging from 7.7% to 52% of total CXCL13�cells (mean � SEM 28.5 � 3.5%) (Table 1). Microana-tomic evaluations demonstrated CD3 expression in18.3 � 2.4% of CXCL13-expressing cells in large lym-phoid aggregates (grade 3) (Figures 2B and C), aprevalence similar to that observed in tonsil GCs from 3patients (14.8 � 1.4%) (results not shown). Of note, incontrast to tonsils, where CXCL13� T cells were ob-served only in secondary follicles, synovial CXCL13� Tcells were also detected in some diffusely infiltratedareas and small perivascular aggregates (grade 1), which
are sublining regions devoid of CD21� follicular den-dritic cell networks (10), where they represented 42.9 �5.6% of the total number of CXCL13-expressing cells(Figure 2A). In keeping with these data, the degree ofdiffuse T cell infiltration interdispersed among cellaggregates was significantly correlated with both thepercentage of CXCL13�CD3� cells among totalCXCL13� cells (P � 0.001 by one-way analysis ofvariance) and the overall density of CXCL13�CD3�cells in the inflamed sublining (P � 0.014).
Phenotypic characterization of SF CXCL13� Tcells. These findings induced us to extend the character-ization of the extralymphoid CXCL13� T cells byanalyzing single-cell suspensions from unfractionatedSFMCs. As shown in isolated SF CD3� lymphocytes(see Figure 1C), spontaneous CXCL13 expression wasobserved ex vivo in a subset of CD3� cells from
Figure 3. Quantification/phenotypic characterization of synovial fluid (SF) CXCL13�CD3� cells. SF mononuclear cells (SFMCs) and peripheralblood mononuclear cells (PBMCs) from patients with rheumatoid arthritis (RA) and healthy donors were separated by density-gradientcentrifugation and immunostained as described in Patients and Methods. A, Double immunofluorescence staining for CXCL13 (red) and markersof specific T cell subsets (green) on cytospin preparations of SFMCs. Note the presence of CXCL13 in the cytoplasm of CD3�CD4�CD45RO�cells. Counterstaining of the cell nuclei with 4�,6-diamidino-2-phenylindole is shown in blue. B, Four-color fluorescence-activated cell sorting (FACS)analysis of SFMCs stained for CXCL13, CD3, CD4, and CD45RO. Middle and bottom, Plots representing the profiles of the gate (top), which wasset to select CXCL13� cells. Gates were set based on negative controls with isotype-matched irrelevant antibodies. C, Representative FACS plotsshowing the profile of CXCL13 staining in CD3�CD4�CD45RO� gated cells from normal donor (ND) PBMCs and from paired RA PBMCs andRA SFMCs obtained by synchronized collection. Data from 6 independent normal donors and 6 patients per group are shown in the graph (right).The horizontal line represents the median. FSC � forward scatter.
3382 MANZO ET AL

unfractionated RA SFMCs. Quantitative evaluations byFACS analysis in 6 patients demonstrated CD3 expres-sion in a mean � SEM of 96.2 � 2% of total CXCL13�cells (range 87.5–100%), while CXCL13 expression wasrecognized in a median of 2.7% of the total T cells, witha variable prevalence ranging from 0.8% to 13%. Dou-ble staining on cytospin preparations and 4-color FACSanalysis confirmed the immunohistologic observations,demonstrating CXCL13 expression in the cytoplasmof CD3�CD4�CD45RO� cells (Figures 3A and B)and rare or no colocalization with CD8 or CD45RA.In keeping with the results of mRNA analyses (seeFigure 1A), although a median of 5.6% ofCD3�CD4�CD45RO� T lymphocytes were positivefor CXCL13 in RA SFMCs (interquartile range 4.5–10.8%), negligible CXCL13 expression was detectedin circulating helper/memory T cells from paired RAand normal donor PBMCs (Figure 3C). The frequentactivated phenotype of SF CXCL13� T lymphocyteswas confirmed by CD69 staining, which was detectedin 60–79% of the cells (Figure 4C, and results notshown).
To characterize the differentiation stages ofCXCL13� memory T cells, we analyzed the distributionof CD27 and the chemokine receptor CCR7, which areexpressed in poorly differentiated and central memorycells but are lost in terminally differentiated and effectormemory T lymphocytes (24–26). CCR7 was not homo-geneously distributed in CXCL13�CD3� lymphocytes,being expressed in a mean � SEM of 29.3 � 4.1% of thecells (Figures 4A and C), with a comparable CXCL13median fluorescence intensity (MFI) in theCXCL13�CD3�CCR7� and CCR7� populations(P � 0.2, by paired t-test). Similarly, despite the obser-vation that CD27 appeared variable in its level ofexpression, most CXCL13� T lymphocytes did not showmembrane staining for the antigen (Figures 4A and C),indicating that CXCL13 can be expressed by fully dif-ferentiated helper/memory SF T cells.
Finally, to investigate the possible relationshipbetween synovial CXCL13� T cells with tonsil GCfollicular helper T cells (27,28), the only normal T helpercell subset in which CXCL13 expression has been doc-umented (16,17), we analyzed the distribution ofCXCR5 and BCL6, 2 antigens highly expressed by thesecells at the mRNA and protein levels (17,29). SurfaceCXCR5 was detected in only a very small fraction ofCXCL13� lymphocytes (mean � SEM 5.5 � 0.7%),which did not reveal significant differences in CXCL13MFI if compared with the CXCL13�CD3�CXCR5�subset (P � 0.4, by paired t-test) (Figures 4B and C).Likewise, despite the detection of BCL6 at high levels in
the nucleus of a proportion of CXCL13� cells, themajority of CXCL13� T cells did not express BCL6 orexpressed it at very low levels (Figures 4B and C).
Collectively, these results indicate that although
Figure 4. Differentiation phenotype of SF CXCL13� T cells. SFMCsfrom RA patients with active joint involvement were separated bydensity-gradient centrifugation and immunostained as described inPatients and Methods. A and B, Left, Representative FACS profiles oftriple stainings for colocalization of CXCL13�CD3�CCR7 (A), andCXCL13�CD3�CXCR5 (B). Gates were set based on negativecontrols with isotype-matched irrelevant antibodies. Right, Represen-tative double immunofluorescence staining on cytospin preparationsfor CXCL13 (red) and CD27 (A) or CXCL13 and BCL6 (B). Coun-terstaining of the cell nuclei was performed using 4�,6-diamidino-2-phenylindole. Arrowheads show CXCL13 single-positive cells, whilethe arrows indicate CXCL13�CD27 (A) and CXCL13–BCL6 (B)double-positive cells. C, Percent of each studied marker inCXCL13�CD3� cells. Values are the mean � SEM results obtainedfrom 7 patients (CD69), 4 patients (CCR7 and CXCR5), 9 patients(CD27), and 8 patients (BCL6). See Figure 3 for definitions. Colorfigure can be viewed in the online issue, which is available athttp://www.arthritisrheum.org.
CXCL13 EXPRESSION BY SYNOVIAL T CELLS 3383

CXCR5 and BCL6 are restricted to a small proportionof SF CXCL13� T lymphocytes, CXCL13 can be ex-pressed by activated and by fully differentiated antigen-experienced T helper cells, differing in their phenotypeand microanatomic distribution from tonsil follicular Bhelper T cells.
CXCL13 modulation by immunologic activationof synovial T cells. The finding of spontaneous CXCL13expression in activated T helper lymphocytes ex vivo andin situ prompted us to investigate the functional effect ofTCR stimulation in synovial primary cells on CXCL13production in vitro. Therefore, we separated SFMCs,rested them in culture for 28 hours, and then performedpolyclonal stimulation with anti-CD3/anti-CD28 anti-bodies.
CXCL13 quantitative analysis by ELISA in thesupernatant of cultured SFMCs revealed a potent effectof CD3–CD28 triggering, with mean stimulation indicesof 13.4 and 19.2 after 48 hours and 72 hours, respectively(Figure 5A). The kinetics of CXCL13 mRNA modula-tion, as monitored by quantitative PCR in parallelexperiments, confirmed CXCL13 transcript up-regulation,with a 3 � 1.2–fold increase in stimulated versus restingcells already detectable after 9 hours of culture (Figure5B). In keeping with these data, FACS analysis per-formed at 36 hours demonstrated a significant increasein the percentage of CD3-gated cells expressingCXCL13 after CD3–CD28 stimulation if compared withboth resting populations (7 � 1.2% for stimulated versus1.8 � 0.6% for resting; P � 0.01) and input populations
Figure 5. CXCL13 modulation by immunologic activation of synovial T cells. A, Time course analysis of CXCL13 protein secretion in thesupernatant of synovial fluid mononuclear cell (SFMC) cultures. Circles and error bars represent the mean � SEM results for triplicate cultures of3 samples from 3 different patients. B, Time course analysis of CXCL13 mRNA expression in SFMC cultures. Values are the mean and SEM resultsfor 3 samples from 3 different patients analyzed in triplicate. C, Fluorescence-activated cell sorter (FACS) analysis. After 36 hours of culture withmedium alone (resting), plate-bound CD3–CD28 antibodies, or autologous SF (aSF), SFMCs were collected and analyzed by FACS. Plots showrepresentative profiles of CXCL13 staining in CD3-gated cells. Note that a minor proportion of CD3� cells show a high forward scatter (FSC) signal,probably attributable to residual cell clustering following culture. Gates were set on negative controls with isotype-matched irrelevant antibodies. D,Results of the analysis for all patients. The y-axis indicates the percentage of CXCL13� cells in total CD3-gated cells. Values are the mean and SEMresults for 3 independent samples from 3 different patients. The input column shows the results of the analysis performed in each sample beforeculture. � � P � 0.01 by one-way repeated-measures analysis of variance and by Student-Newman-Keuls test for pairwise multiple comparisons.RQ � relative quantification; RA � rheumatoid arthritis.
3384 MANZO ET AL

(3.3 � 0.5%; P � 0.01) from the respective patients(Figures 5C and D). Similar experiments performed byincubating SFMCs with acellular autologous SF, as acomprehensive source of soluble factors present in situduring synovial inflammation, did not reveal a statisti-cally significant effect (2.7 � 1.9% for SF versus 1.8 �0.6% for resting; P � 0.3) (Figures 5C and D).
DISCUSSION
A large body of evidence indicates that synovial Tcells can establish functional interactions with B cells inRA (18,19). Rheumatoid synovium can indeed hostantigen-dependent differentiation of B cells in ectopicGCs (20), while activation of T cells and production ofproinflammatory cytokines (such as tumor necrosis fac-tor � and interleukin-1) have been shown to be partiallydependent on local B cells (4). However, although themolecular systems adopted by synovial T cells to coop-erate with macrophages and fibroblasts have been ex-tensively investigated (30,31), the mechanisms promot-ing T cell–B cell interaction are less well defined. Here,we present evidence that RA synovial T cells can syn-thesize and secrete the chemokine CXCL13, and thatexpression of this factor is inducible by immunologicactivation of synovial primary cells through the TCR–CD28 pathway. Thus, our data demonstrate a newfunctional property of synovial T lymphocytes, indicat-ing their direct involvement in the regulation of Bcell chemoattractant expression in rheumatoid synovitisand suggest a potential role for CXCL13 as a directsignal transducer involved in local T cell–B cell cooper-ation.
Tissue expression of CXCL13 has been shown tobe critically involved, through the recruitment and orga-nization of B cells, in the process of ectopic lymphoidneogenesis, a phenomenon that is observed in 25–50%of patients with RA, whose developmental mechanismshave not been fully elucidated (7–10). Several studies inrecent years have therefore focused on the identificationof cellular sources and mechanisms of induction ofCXCL13 to obtain insights in the pathologic eventslinking this process with synovial inflammation. Take-mura et al (9) showed that, consistent with the proper-ties of the mesenchymal component of embryonic lymphnodes, CXCL13 can be expressed in RA synovium byfibroblast-like synoviocytes, follicular dendritic cells, andendothelial cells, suggesting a role of the inflamedsynovial stroma in the recruitment and organization ofcirculating B cells (32). Carlsen et al (23), in a subse-quent study, showed that CXCL13 can be expressed by
macrophage/monocytes following lipopolysaccharidestimulation, indicating involvement of the innate im-mune system in ectopic lymphoid neogenesis. Our ob-servations provide evidence of an additional level ofcomplexity demonstrating that CXCL13 can be pro-duced in the synovium by CD4� memory T cells, andthat expression of the chemokine can be modulated insynovial primary cells by TCR–CD28 engagement. Al-though our results do not exclude the possibility thatindirect mechanisms of activation might contribute toCXCL13 expression in synovial cells in vitro, or thatadditional TCR-independent mechanisms could induceCXCL13 expression in synovial T cells in vivo, theydemonstrate that CXCL13 can be produced by cells ofthe adaptive immune system in the joint and modulatedthrough T cell immunologic receptors. Thus, our studyextends current knowledge providing evidence of thepotential contribution of the T cell compartment andantigen-dependent signals in the mechanisms of B celllocalization in RA.
These data represent the first evidence that theCXCL13� phenotype is not restricted to T cells local-ized in tonsil GCs, but that such a functional programcan be activated in normal T cells involved in chronicinflammatory reactions in extralymphoid tissue. We alsodemonstrate that CXCL13 can be expressed by memoryCD4� T cells featuring a highly differentiated pheno-type, as suggested by the frequent absence of CCR7 andCD27 in SF T cells. These findings are consistent withprevious data that showed the potent B helper activity ofsynovial CD45RBdim mature memory T cells and theircapacity to modulate immunoglobulin production (18).
A critical question is whether synovial CXCL13�CD3� cells and tonsil CXCL13�CD3� cells representthe same cell subsets migrating/differentiating in differ-ent organs, or whether different cell types in extralym-phoid tissues can acquire the capacity for CXCL13expression. Indeed, CXCL13 production in tonsil T cellshas been demonstrated in a subset of activated (CD69�)CD4� memory lymphocytes, enriched in the CCR7�fraction and characterized by high efficiency in promot-ing B cell differentiation and immunoglobulin produc-tion (follicular B helper T cells or GC T helper cells)(27,28,33). However, recent studies suggest that, to-gether with their restricted localization in tonsil GCs,follicular B helper T cells and their possible precursorsexpress CXCL13 as a differential feature, because thechemokine is absent or is present at very low levels inother T cell subsets, including naive, central/memory,and Th1- and Th2-polarized lymphocytes (16,17,29).
One of the additional distinctive phenotypic fea-
CXCL13 EXPRESSION BY SYNOVIAL T CELLS 3385

tures of tonsil follicular B helper T cells is the uniformexpression at high levels of the chemokine receptorCXCR5 (17), which is believed to promote their move-ment into the B cell follicle where CXCL13, the CXCR5ligand, is constitutively produced (27). Our data suggestthat this phenotype does not systematically overlap theone displayed by CXCL13� T cells localized in the joint,which we observed were mostly negative for surfaceCXCR5 in all SF samples analyzed. Although the phe-notype in SF cells could differ in part from that inlocalized synovial tissue cells, we observed the absenceof other markers characteristic of tonsil follicular Bhelper T cells, such as the transcriptional repressorBCL6 (17,29), in the majority of SF CXCL13�CD3�cells, and we demonstrated a significant presence ofsynovial tissue–infiltrating CXCL13� T cells outsideectopic GCs.
These findings thus indicate the existence ofregulatory or differentiation discrepancies between syno-vial CXCL13� memory T cells and tonsil follicular BT helper cells, which might reflect differences in thelineage derivation of these populations in physiologicand pathologic conditions. Alternatively, it can be hy-pothesized that despite a common origin from follicularB helper T cell precursors, synovial CXCL13� T cellsmight undergo a process of progressive differentiation inthe synovium, with the modification of their originalphenotype and homing behavior. This could be the casefor CXCR5, which, along with possible ligand-mediatedinternalization, has been shown to be highly unstable inT lymphocytes, being rapidly down-regulated followingTCR stimulation and irreversibly lost in terminally dif-ferentiated cells (24). This hypothesis would be consis-tent with the finding of a small proportion ofCXCL13�CXCR5� and CXCL13�BCL6high T cellsand with the heterogeneous expression of CCR7 andCD27 in SF CXCL13�CD3� cells. Future microarrayanalyses investigating the whole transcriptional programof these cells are thus desirable to address this importantbiologic aspect.
In summary, we provide evidence that normal Tcells infiltrating inflamed extralymphoid tissue can ac-quire interactive properties with local B cells throughthe production of chemoattractants. This study extendsthe known functional properties of synovial T cells andsupports their central role in the cellular network of RAsynovitis (34). The synovial CXCL13� T cell subsetidentified here might turn out to be an attractive targetto manipulate the pathologic interactions established byT cells and B cells in patients with RA.
AUTHOR CONTRIBUTIONS
Dr. Pitzalis had full access to all of the data in the study andtakes responsibility for the integrity of the data and the accuracy of thedata analysis.Study design. Manzo, Dell’Accio, Uguccioni, Montecucco, Pitzalis.Acquisition of data. Manzo, Vitolo, Humby, Caporali, Jarrossay,Ciardelli.Analysis and interpretation of data. Manzo, Vitolo, Jarrossay,Dell’Accio, Ciardelli, Uguccioni, Montecucco, Pitzalis.Manuscript preparation. Manzo, Vitolo, Dell’Accio, Uguccioni, Mon-tecucco, Pitzalis.Statistical analysis. Manzo, Vitolo, Pitzalis.
REFERENCES
1. Edwards JC, Cambridge G, Abrahams VM. Do self-perpetuatingB lymphocytes drive human autoimmune disease [review]? Immu-nology 1999;97:188–96.
2. Dorner T, Burmester GR. The role of B cells in rheumatoidarthritis: mechanisms and therapeutic targets [review]. Curr OpinRheumatol 2003;15:246–52.
3. Moynier M, Abderrazik M, Didry C, Sany J, Brochier J. The B cellrepertoire in rheumatoid arthritis. III. Preferential homing ofrheumatoid factor–producing B cell precursors in the synovialfluid. Arthritis Rheum 1992;35:49–54.
4. Takemura S, Klimiuk PA, Braun A, Goronzy JJ, Weyand CM. Tcell activation in rheumatoid synovium is B cell dependent.J Immunol 2001;167:4710–8.
5. Legler DF, Loetscher M, Roos RS, Clark-Lewis I, Baggiolini M,Moser B. B cell-attracting chemokine 1, a human CXC chemokineexpressed in lymphoid tissues, selectively attracts B lymphocytesvia BLR1/CXCR5. J Exp Med 1998;187:655–60.
6. Ansel KM, Ngo VN, Hyman PL, Luther SA, Forster R, SedgwickJD, et al. A chemokine-driven positive feedback loop organizeslymphoid follicles. Nature 2000;406:309–14.
7. Luther SA, Lopez T, Bai W, Hanahan D, Cyster JG. BLCexpression in pancreatic islets causes B cell recruitment andlymphotoxin-dependent lymphoid neogenesis. Immunity 2000;12:471–81.
8. Shi K, Hayashida K, Kaneko M, Hashimoto J, Tomita T, LipskyPE, et al. Lymphoid chemokine B cell-attracting chemokine-1(CXCL13) is expressed in germinal center of ectopic lymphoidfollicles within the synovium of chronic arthritis patients. J Immu-nol 2001;166:650–5.
9. Takemura S, Braun A, Crowson C, Kurtin PJ, Cofield RH,O’Fallon WM, et al. Lymphoid neogenesis in rheumatoid synovi-tis. J Immunol 2001;167:1072–80.
10. Manzo A, Paoletti S, Carulli M, Blades MC, Barone F, Yanni G,et al. Systematic microanatomical analysis of CXCL13 and CCL21in situ production and progressive lymphoid organization in rheu-matoid synovitis. Eur J Immunol 2005;35:1347–59.
11. Bugatti S, Caporali R, Manzo A, Vitolo B, Pitzalis C, MontecuccoC. Involvement of subchondral bone marrow in rheumatoid ar-thritis: lymphoid neogenesis and in situ relationship to subchon-dral bone marrow osteoclast recruitment. Arthritis Rheum 2005;52:3448–59.
12. Rioja Pastor I, Dickson MC, Binks MH, Lukey PT, Petavy F,McClinton C, et al. B-lymphocyte chemoattractant (BLC/CXCL13): a potential novel disease activity marker of rheumatoidarthritis. Ann Rheum Dis 2007;66 Suppl II:ii452.
13. Zheng B, Ozen Z, Zhang X, De Silva S, Marinova E, Guo L, et al.CXCL13 neutralization reduces the severity of collagen-inducedarthritis. Arthritis Rheum 2005;52:620–6.
14. Wengner AM, Hopken UE, Petrow PK, Hartmann S, Schurigt U,Brauer R, et al. CXCR5- and CCR7-dependent lymphoid neogen-
3386 MANZO ET AL

esis in a murine model of chronic antigen-induced arthritis.Arthritis Rheum 2007;56:3271–83.
15. Honda K, Nakano H, Yoshida H, Nishikawa S, Rennert P, IkutaK, et al. Molecular basis for hematopoietic/mesenchymal interac-tion during initiation of Peyer’s patch organogenesis. J Exp Med2001;19:621–30.
16. Kim CH, Lim HW, Kim JR, Rott L, Hillsamer P, Butcher EC.Unique gene expression program of human germinal center Thelper cells. Blood 2004;104:1952–60.
17. Rasheed AU, Rahn HP, Sallusto F, Lipp M, Muller G. FollicularB helper T cell activity is confined to CXCR5hiICOShi CD4 T cellsand is independent of CD57 expression. Eur J Immunol 2006;36:1892–903.
18. Thomas R, McIlraith M, Davis LS, Lipsky PE. Rheumatoidsynovium is enriched in CD45RBdim mature memory T cells thatare potent helpers for B cell differentiation. Arthritis Rheum1992;35:1455–65.
19. MacDonald KP, Nishioka Y, Lipsky PE, Thomas R. FunctionalCD40 ligand is expressed by T cells in rheumatoid arthritis. J ClinInvest 1997;100:2404–14.
20. Schroder AE, Greiner A, Seyfert C, Berek C. Differentiation of Bcells in the nonlymphoid tissue of the synovial membrane ofpatients with rheumatoid arthritis. Proc Natl Acad Sci U S A1996;93:221–5.
21. Arnett FC, Edworthy SM, Bloch DA, McShane DJ, Fries JF,Cooper NS, et al. The American Rheumatism Association 1987revised criteria for the classification of rheumatoid arthritis.Arthritis Rheum 1988;31:315–24.
22. Mazzucchelli L, Blaser A, Kappeler A, Scharli P, Laissue JA,Baggiolini M, et al. BCA-1 is highly expressed in Helicobacterpylori-induced mucosa-associated lymphoid tissue and gastric lym-phoma. J Clin Invest 1999;104:R49–54.
23. Carlsen HS, Baekkevold ES, Morton HC, Haraldsen G,Brandtzaeg P. Monocyte-like and mature macrophages produceCXCL13 (B-cell-attracting chemokine 1) in inflammatory lesionswith lymphoid neogenesis. Blood 2004;104:3021–7.
24. Sallusto F, Lenig D, Forster R, Lipp M, Lanzavecchia A. Twosubsets of memory T lymphocytes with distinct homing potentialsand effector functions. Nature 1999;401:708–12.
25. Fritsch RD, Shen X, Sims GP, Hathcock KS, Hodes RJ, LipskyPE. Stepwise differentiation of CD4 memory T cells defined byexpression of CCR7 and CD27. J Immunol 2005;175:6489–97.
26. Schaerli P, Willimann K, Lang AB, Lipp M, Loetscher P, Moser B.CXC chemokine receptor 5 expression defines follicular homing Tcells with B cell helper function. J Exp Med 2000;192:1553–62.
27. Breitfeld D, Ohl L, Kremmer E, Ellwart J, Sallusto F, Lipp M, etal. Follicular B helper T cells express CXC chemokine receptor 5,localize to B cell follicles, and support immunoglobulin produc-tion. J Exp Med 2000;192:1545–52.
28. Schaerli P, Loetscher P, Moser B. Cutting edge: induction offollicular homing precedes effector Th cell development. J Immu-nol 2001;167:6082–6.
29. Chtanova T, Tangye SG, Newton R, Frank N, Hodge MR, RolphMS, et al. T follicular helper cells express a distinctive transcrip-tional profile, reflecting their role as non-Th1/Th2 effector cellsthat provide help for B cells. J Immunol 2004;173:68–78.
30. McInnes IB, Leung BP, Liew FY. Cell-cell interactions in synovi-tis: interactions between T lymphocytes and synovial cells. Arthri-tis Res 2000;2:374–8.
31. Brennan F, Foey A. Cytokine regulation in RA synovial tissue:role of T cell/macrophage contact-dependent interactions. Arthri-tis Res 2002;4 Suppl 3:S177–82.
32. Weyand CM, Goronzy JJ. Ectopic germinal center formation inrheumatoid synovitis. Ann N Y Acad Sci 2003;987:140–9.
33. Kim CH, Rott LS, Clark-Lewis I, Campbell DJ, Wu L, ButcherEC. Subspecialization of CXCR5� T cells: B helper activity isfocused in a germinal center-localized subset of CXCR5� T cells.J Exp Med 2001;193:1373–81.
34. Panayi GS, Corrigall VM, Pitzalis C. Pathogenesis of rheumatoidarthritis: the role of T cells and other beasts. Rheum Dis ClinNorth Am 2001;27:317–34.
CXCL13 EXPRESSION BY SYNOVIAL T CELLS 3387
Copyright © 2022 FDOKUMEN




















