
Early expression of monocyte chemoattractant protein-1 correlates with the onset of...
10
http://jra.sagepub.com/ System Journal of Renin-Angiotensin-Aldosterone http://jra.sagepub.com/content/9/3/154 The online version of this article can be found at: DOI: 10.1177/1470320308096408 2008 9: 154 Journal of Renin-Angiotensin-Aldosterone System Gamaliel E. Ordenes, Raul Vivar Sanchez, Sergio Lavandero and Guillermo Díaz-Araya Miguel Copaja Soto, Rodrigo Valenzuela, Adiela Saldaña, María Paz Ocaranza, Jorge E. Jalil, Carlos Vio, Paul Lijnen, polymorphism isoproterenol-induced cardiac fibrosis in rats with distinct angiotensin-converting enzyme Early expression of monocyte chemoattractant protein-1 correlates with the onset of Published by: http://www.sagepublications.com On behalf of: Frequently Asked Questions can be found at: Journal of Renin-Angiotensin-Aldosterone System Additional services and information for http://jra.sagepub.com/cgi/alerts Email Alerts: http://jra.sagepub.com/subscriptions Subscriptions: http://www.sagepub.com/journalsReprints.nav Reprints: http://www.sagepub.com/journalsPermissions.nav Permissions: http://jra.sagepub.com/content/9/3/154.refs.html Citations: What is This? - Oct 20, 2008 Version of Record >> by guest on March 14, 2014 jra.sagepub.com Downloaded from by guest on March 14, 2014 jra.sagepub.com Downloaded from
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Early expression of monocyte chemoattractant protein-1 correlates with the onset of...

http://jra.sagepub.com/System
Journal of Renin-Angiotensin-Aldosterone
http://jra.sagepub.com/content/9/3/154The online version of this article can be found at:
DOI: 10.1177/1470320308096408
2008 9: 154Journal of Renin-Angiotensin-Aldosterone SystemGamaliel E. Ordenes, Raul Vivar Sanchez, Sergio Lavandero and Guillermo Díaz-Araya
Miguel Copaja Soto, Rodrigo Valenzuela, Adiela Saldaña, María Paz Ocaranza, Jorge E. Jalil, Carlos Vio, Paul Lijnen,polymorphism
isoproterenol-induced cardiac fibrosis in rats with distinct angiotensin-converting enzyme Early expression of monocyte chemoattractant protein-1 correlates with the onset of
Published by:
http://www.sagepublications.com
On behalf of:
Frequently Asked Questions
can be found at:Journal of Renin-Angiotensin-Aldosterone SystemAdditional services and information for
http://jra.sagepub.com/cgi/alertsEmail Alerts:
http://jra.sagepub.com/subscriptionsSubscriptions:
http://www.sagepub.com/journalsReprints.navReprints:
http://www.sagepub.com/journalsPermissions.navPermissions:
http://jra.sagepub.com/content/9/3/154.refs.htmlCitations:
What is This?
- Oct 20, 2008Version of Record >>
by guest on March 14, 2014jra.sagepub.comDownloaded from by guest on March 14, 2014jra.sagepub.comDownloaded from

AbstractIInnttrroodduuccttiioonn.. Isoproterenol treatment of BrownNorway and Lewis rats (high and low plasmaangiotensin-I-converting enzyme activity,respectively) results in similar cardiac hypertrophybut higher cardiac fibrosis in Brown Norway rats.MMaatteerriiaallss aanndd mmeetthhooddss.. Rats were infused in vivowith isoproterenol for two or 10 days. Cardiacfibrosis and inflammation were evaluatedhistochemically. We measured the mRNAs ofpro-fibrotic factors (transforming growth factorβ1, endothelin-1) and pro-inflammatory factors(monocyte chemoattractant protein-1). In studieswith cardiac fibroblasts incubated withisoproterenol in vitro, we measured cellproliferation, angiotensin-I-converting enzymeand matrix metalloprotease 2 activities anddeposition of collagen type I and fibronectin.RReessuullttss.. After treatment with isoproterenol fortwo days, there were large areas of myocardialinjury and numerous inflammatory foci in the leftventricle, these being greater in Brown-Norwaythan in Lewis rats. After treatment withisoproterenol for 10 days, there were large areasof damage with extensive collagen depositiononly in the left ventricle; both strains exhibitedthis damage which was, however, more severe inBrown-Norway than in Lewis rats. Aftertreatment with isoproterenol for two, but not 10,days, greater amounts of monocytechemoattractant protein-1 mRNA were found inBrown Norway than in Lewis rats. Cellproliferation, activities of angiotensin-I-converting enzyme and matrix metalloprotease 2,amounts of collagen type I and fibronectin weresimilar in cardiac fibroblasts from both strains;changes after isoproterenol(10 µM) were also similar in both strains.CCoonncclluussiioonn.. We conclude that the greater cardiacfibrosis in Brown Norway rats treated withisoproterenol correlates with the early andhigher expression of proinflammatory factors.
IntroductionComponents of the renin-angiotensin system(RAS), including angiotensin-converting enzyme(ACE) and angiotensin II (Ang II), have beenimplicated in the development of cardiac fibro-sis.1,2 ACE cleaves angiotensin I to produce AngII, which then binds to its receptors to producepotent vasoconstriction, and, in the heart, mod-ulating hypertrophy and fibrosis.3 The identifica-tion of a microsatellite marker in the rat ACEgene has permitted distinction in ACE allelesbetween Brown-Norway (BB) and Lewis (LL)rats, with high and low plasma ACE activities,respectively.4,5 We have shown that chronicadministration of isoproterenol (ISO) producesmicroinfarct-like lesions in the rat myocardiumand that these are associated with a similardegree of heart hypertrophy but more cardiacfibrosis in BB than in LL rats.6 These results sug-gest that ACE may have a role in the develop-ment of cardiac fibrosis.
Cardiac fibrosis (defined as extracellular matrix[ECM] protein deposition) is mainly regulated bycardiac fibroblasts. These cells are the major sourceof extracellular connective tissue matrix, and therecruitment, accumulation and stimulation of thesecells are thought to play important roles in bothnormal healing and the development of fibrosis.1
These cells have a number of functional receptors,including adrenergic, Ang II, bradykinin andpurinergic receptors, all coupled to G protein-linked signalling pathways, in a pattern that is dis-tinct from those in cardiac myocytes.7 In the heart,β2-adrenergic receptors are mainly expressed in car-diac fibroblasts, indicating that ISO could directlymodulate cardiac fibroblast function.8
Several lines of evidence have shown that tissueAng II is involved in inflammatory and healingresponses, regulating the expression of mediators
Paper
154
Journal ofthe Renin-Angiotensin-AldosteroneSystem(Including otherPeptidergic systems)
September 2008Volume 9Number 3 SAGE Publications 2008 Los Angeles, London, New Delhi and Singapore 10.1177/1470320308096408
* Centro FONDAPEstudios Moleculares dela Célula, Universidad deChile, Santiago, Chile.
† DepartamentoEnfermedadesCardiovasculares,Escuela de Medicina,Pontificia UniversidadCatólica de Chile,Santiago, Chile.
# Facultad de CienciasBiológicas, PontificiaUniversidad Católica deChile, Santiago, Chile.Hypertension andCardiovascularRehabilitation Unit,Katholieke Universiteit vonLeuven, Leuven, Belgium.
§ Facultad de Medicina,Universidad de Chile,Santiago, Chile.
¥ These authors contributedequally to the work.
Correspondence to:Dr Guillermo Díaz-ArayaFacultad CienciasQuímicas yFarmacéuticas,Universidad de Chile,Olivos 1007,Santiago 8380294, Chile.Tel: +56 2 978 2975Fax: +56 2 737 8920Email:[email protected]
Early expression of monocyte chemoattractantprotein-1 correlates with the onset ofisoproterenol-induced cardiac fibrosis inrats with distinct angiotensin-convertingenzyme polymorphism
Miguel Copaja Soto,*,¥ Rodrigo Valenzuela,*,¥ Adiela Saldaña,* María Paz Ocaranza,† Jorge E. Jalil,†
Carlos Vio,# Paul Lijnen, Gamaliel E. Ordenes,§ Raul Vivar Sanchez,* Sergio Lavandero,*,§ GuillermoDíaz-Araya*
Key words:ACE genepolymorphism,cardiac fibrosis,isoproterenol,pro-fibroticfactors, pro-inflammatoryfactors
by guest on March 14, 2014jra.sagepub.comDownloaded from

Paper
155SAGE Publications 2008 Los Angeles, London, New Delhi and Singapore 10.1177/1470320308096408
Journal ofthe Renin-Angiotensin-AldosteroneSystem(Including otherPeptidergic systems)
September 2008Volume 9Number 3
of repair at vascular and non-vascular sites ofcardiac injury.9 Ang II increases the expression ofmonocyte chemoattractant protein-1 (MCP-1), animportant chemokine with strong influence onthe amplification of inflammation processes, bypromoting recruitment of macrophages to inflam-matory sites.9 However, Ang II also regulates theexpression of the pro-fibrotic factors transforminggrowth factor (TGF)-β1 and endothelin (ET)-1, aswell as the expression of ECM proteins.10,11 Therole of these pro-inflammatory/pro-fibrotic fac-tors on cardiac fibrosis induced by ISO in ratswith different ACE polymorphisms is unknown.Elucidation of this role was the aim of this study.
Our results indicate that ISO-induced cardiacfibrosis correlates with increased levels of pro-inflammatory factors in Brown Norway rats only;although cardiac fibroblasts have an importantrole in ECM deposition, cardiac fibroblasts iso-lated from both rat strains responded in vitro in asimilar manner to ISO in proliferation, collagentype I and fibronectin secretion, and matrix met-alloprotease (MMP)-2 and ACE activities.
MethodsExperimental designThe investigation conformed to the Guide for theCare andUse of Laboratory Animals published bythe National Institutes of Health (NIH publicationno. 85-23, revised 1985). We used an experimen-tal ACE genetic model previously described byChallah et al.,4 in which two normotensive strainsof rats, the Brown Norway and Lewis strains, havecontrasting levels of plasma ACE. F0 BrownNorway males (BB ACE genotype) and F0 Lewisfemales (LL ACE genotype) were from CharlesRiver (Wilmington, MA, USA). Rats were mated toproduce F1 hybrids (LB genotype). F1 hybridswere mated to obtain the F2 cohorts, which wereused in this study.6
TreatmentHomozygous F2 male rats (80–100 g) wereinjected subcutaneously with ISO (5 mg·kg–1
day–1; Sigma) or saline (control) for 10 days.Animals were divided into four experimentalgroups: (1) LL-C: LL rats receiving saline injection;(2) BB-C: BB rats receiving saline injection; (3) LL-ISO: LL rats receiving ISO injection; and (4) BB-ISO:BB rats receiving ISO injection. After two or 10days of treatment, animals were weighed and killedby decapitation at day three before the followinginjection or at day 11 (24 h after the last injection).The blood was collected into pre-chilled tubes con-taining heparin for the determination of plasmaACE activity. The heart was rapidly excised, and
the left ventricle (LV) plus septum and right ven-tricle (RV) were weighed (LVW and RVW, respec-tively). The LV was sectioned transversely intotwo slices. One was fixed in 4% formalin for his-tochemical studies, and the other was rapidlyfrozen in liquid N2 and stored at −70°C until usedfor measurement of ACE activity and mRNA(ET-1, TGF-β1, MCP-1, ACE, and glyceraldehyde-3-phosphate dehydrogenase [GAPDH]).
Determination of plasma and cardiacfibroblast ACE activityPlasma and cellular ACE activities were assayed byfollowing the release of His-Leu from the syntheticsubstrate Z-Phe-His-Leu (Bachem Biosciences Inc.,King of Prussia, PA, USA) with or without enalaprilaccording to the fluorometric assay previouslydescribed.5 A sample of plasma or homogenisedcardiac fibroblast membranes was then incubatedat 37°C in a shaking waterbath with Z-Phe-His-Leu for 30 minutes or 2 hours for plasma and car-diac fibroblast membranes, respectively. Thereaction was stopped by addition of 100 µl ofcold 10% trichloracetic acid and 280 mM NaOH.Phthalaldehyde (0.1%) was added to the samples,which were then incubated for 10 minutes at 37°Cbefore reaction was stopped with 3 M HCl. Thefluorescence at 486 nm was measured with anexcitation of 364 nm (Fluorescence Spectro -photometer 650-10, Perkin Elmer, Waltham, MA,USA). Plasma ACE activity was expressed as nmolof His-Leu liberated per minute per ml of plasma(U/ml plasma); cellular ACE activity was expressedas nmol of His-Leu liberated per minute per mgprotein at 37°C (U/mg protein).
Evaluation of cardiac hypertrophyThe degrees of LV hypertrophy (LVH) and RVhypertrophy (RVH) were quantified by the rela-tive LV weight ratio (RLVW, LV weight 100/bodywt) and the relative RV weight ratio (RRVW, RVweight 100/body wt), respectively.
Tissue processing and stainingThe equatorial regions of each LV were routinelyprocessed and paraffin embedded. Sections (5 µm)were stained with hematoxylin and eosin forhistopathological analysis. Sections (5 µm-thick)were also stained with Masson’s trichrome fordetermination of interstitial collagen.
Histopathological analysisSemi-quantification of myocardial injury andinflammatory lesions was performed by a trainedpathologist who had no knowledge of the exper-imental groups during the evaluation of the tis-sues. Two categories were used to score the levelof myocardial injury and inflammatory lesions in
by guest on March 14, 2014jra.sagepub.comDownloaded from

Paper
156SAGE Publications 2008 Los Angeles, London, New Delhi and Singapore 10.1177/1470320308096408
Journal ofthe Renin-Angiotensin-AldosteroneSystem(Including otherPeptidergic systems)
September 2008Volume 9Number 3
several sections from each heart. A score of zerorepresented no damage. A score of one repre-sented clear area of myocardial necrosis andinflammatory infiltrate.
Determination of MCP-1, TGF-ββ1 andET-1 mRNAs by reversetranscription-polymerase chainreaction (RT-PCR)Total RNA was isolated from the LV with Trizol(Invitrogen, Carlsbad, CA, USA). RNA pelletswere suspended in distilled and DEPC water, andtheir concentrations were measured by UV spec-troscopy. RNA integrity was assessed from theintensity of the staining with ethidium bromide of18 and 28S ribosomal RNA after agarose elec-trophoresis. The amounts of tissue TGF-β1, MCP-1,and ET-1 mRNAs were determined by RT-PCR.For reverse transcription, 1 µg RNA was incubatedwith or without reverse transcriptase (GIBCO-BRL, Gaithersburg, MD, USA) in a mixture con-taining random primers, deoxynucleotides, andRNAs in reverse transcription buffer. For amplifi-cation of the resulting cDNA, 1 µM of each specificprimer pair, 0.2 mM deoxynucleotides, 1.5 mMMgCl2, and 1.5 U Taq polymerase (GIBCO-BRL)were added to 3 µl of each RNA sample in a finalvolume of 50 µl. All the sequences of the senseand antisense primers are described in table 1.After PCR, the amplification products were frac-tionated on a 1.5% (w/w) agarose gel and visu-alised by staining with ethidium bromide. Bandintensities were quantified by computerised den-sitometry and normalised with respect toGAPDH mRNA.
Cardiac fibroblast isolation and cultureHearts were rapidly extracted and cardiac fibro -blasts from BB and LL male adult rats (130±20 g)were isolated by cardiac retrograde aortic perfu-sion as described previously,12 with a few modi-fications. Briefly, the heart was digested with asolution containing collagenase-hyaluronidase(1:1); the resulting cells were centrifuged at
1,000 rpm for 5 minutes, and then resuspendedin Dulbecco’s modified Eagle’s medium (DMEM)-F12 containing 10% foetal bovine serum andseeded in non-treated culture dishes for 2 hours.The cells were then washed with phosphate-buffered saline to eliminate debris and non-adherent cells. Passage two fibroblasts werewashed twice with Moscona’s solution and cul-tured in serum-free DMEM-F12 (SFM), supple-mented with ascorbic acid (50 µg/ml) andglutamine (50 µg/ml) for 24 hours. They werethen incubated with or without 10 µM ISO in SFMfor 24–48 hours. Cardiac fibroblasts derived fromeach rat were used in an independent experi-ment. Fibroblasts from both rat strains were mor-phologically characterised by microscopy.
Cardiac fibroblast proliferationCardiac fibroblasts were seeded into 96-wellplates (5,000 cells per well), incubated overnightand growth-arrested using SFM for 24 hours. SFMwas then replaced with 0.2 ml DMEM-F12 con-taining the appropriate stimulus. This time wasdesignated ‘Day 0’. Medium and stimulus werereplaced after 24 hours by DMEM-F12 withoutphenol red and quantified by Cell Titer®(Molecular Probes, Seattle, OR, USA), followingthe manufacturer’s instructions.
Collagen and fibronectin protein levelsSerum-starved cardiac fibroblasts were treatedwith 10 µM ISO or vehicle. At the indicated times,culture medium was collected and concentratedusing Centricon plus-20 tubes (Millipore,Billerica, MA, USA). Non-reducing SDS-PAGEsample buffer was added directly to the concen-trated medium and samples were subjected toelectrophoresis. Proteins prepared using non-reducing sample buffer (25 µg per lane) wereseparated by SDS-PAGE on 12% (w/v) gels andtransferred electrophoretically to nitrocellulose.The blots were incubated with anti-fibronectinantibody (1:10,000; BIODESIGN International,Maine, USA) or anti-collagen type I antibody
Table 1List of sense and antisense primers used in RT-PCR of rats with distinct ACE gene polymorphism.
Primers Sequence (5’ → 3’) Annealing temperature [ºC] Size [bp]
TGF-β1 Sense 5’-AAGCCCTGTATTCCGTCTCC-3’Antisense 5’-CAACGCCATCTATGAGAAAACC-3’ 52 290
ET-1 Sense 5’-CTAGGTCTAAGCGATCCTTG-3’Antisense 5’-TTCTGGTCTCTGTAGAGTTC-3’ 50 319
MCP-1 Sense 5’-CAGGTCTCTGTCACGCTTCT-3’Antisense 5’-GTGCTTCAGGTGGTTGTGG-3’ 53 450
GAPDH Sense 5’-CCATCACCATCTTCCAGGAG-3’Antisense 5’-CCTGCTTCACCACCTTCTTG-3’ 60 541
by guest on March 14, 2014jra.sagepub.comDownloaded from

Paper
157SAGE Publications 2008 Los Angeles, London, New Delhi and Singapore 10.1177/1470320308096408
Journal ofthe Renin-Angiotensin-AldosteroneSystem(Including otherPeptidergic systems)
September 2008Volume 9Number 3
(1:5,000; Chemicon, Tamecula, CA, USA). Afterwashing, blots were incubated with anti-rabbitIgG antibodies linked to horseradish peroxidase(1:10,000). Bands were detected using ECL, andquantified by laser scanning densitometry.Membranes were stained with ponceau red toconfirm equal loading.
MMP activitiesAssessment of MMP-2 activity was performed byzymography. Concentrated culture media (20 µgprotein) were treated with non-reducing samplebuffer and resolved on 12% SDS-PAGE gels con-taining 0.1% gelatin. Gels were washed twicewith 2.5% triton X-100 for 30 minutes at roomtemperature, incubated at 37°C for 18 hours in 50mM HEPES (pH 7.5) containing 0.2 M NaCl, 5mM CaCl2 and 20 µM ZnCl2 and subsequentlystained with coomassie blue R-250. ActivatedMMPs produced clear areas and bands werequantified by densitometry.
Statistical analysisResults were expressed as mean±SEM. Differencesbetween strains were estimated by one-way ANOVAfollowed by Student-Newman-Keuls assessmentfor the number of independent experiments indi-cated (n). The limit of statistical significance wasset at p < 0.05.
ResultsCardiac hypertrophyFigure 1 shows that the relative LV weight ratiodid not change after two days of ISO treatment.The LV ratio was, however, remarkably increasedafter 10 days of ISO treatment when comparedwith that in normal rats.
Myocardial histopathologyHearts from control rats were histologically nor-mal. No myocardial lesions were identified inhearts from rats receiving saline solution after twodays (figure 2A LL-C, BB-C) or 10 days (figure 2BLL-C, BB-C) of treatment. In contrast, myocardialalterations were observed in two-day samples inboth rats (figure 2 LL-ISO, BB-ISO). Myocardialchanges were characterised by degeneration andnecrosis of cardiomyocytes and replacement byinflammatory cells (predominantly histiocytes),distributed mainly in interstitial space. The semi-quantitative scores for myocardial injury in ani-mals treated for two days are shown in table 2.After two days of treatment, the score for myocar-dial alterations was 0 for all LL-C and BB-C rats,while it was 1 for only fifty percent of LL-ISO rats(three rats); in contrast, it was 1 for all BB-ISOrats (five animals).
Strong and markedly different myocardial alter-ations were observed after 10 days between bothrat strains (figure 2 LL-ISO, BB-ISO). The semi-quantitative scores for myocardial injury in ani-mals treated for 10 days are shown in table 2.After 10 days of treatment, the score for myocar-dial damage was 0 for LL-C and BB-C rats; mean-while it was similar between BB-ISO and LL-ISOrats (score 0–1 with similar number of rats).However, in BB-ISO rats the myocardial changeswere characterised by large areas with isolatedcardiomyocytes individually or in groups, sur-rounded by ECM protein and non-myocyte cells,and low presence of inflammatory areas. Thesedamage areas looked as if wound healing wasfinished, as evidenced by lower infiltration ofinflammatory cells, which were replaced byexcessive ECM protein deposition (figure 3 BB-ISO vs. LL-ISO and respective controls). In con-trast, in LL-ISO group, areas with granulationtissue and extensive infiltration of inflammatorycells were observed; however, low collagen dep-osition was observed by low trichrome Massonstain (figure 3 LL-ISO).
Myocardial fibrosisAfter two days there was neither collagen depo-sition nor a difference between both experimen-tal groups (data not shown). The myocardialinterstitial collagen deposition of rats receiving
Figure 1Effect of ISO on LVH in rats with different ACE polymorphisms.LVH at two days (A), and at 10 days (B) of ISO treatmentwas measured as mentioned in methods. Data are mean±SEM;n = 6–7 in duplicate; *p < 0.01 vs. control.
by guest on March 14, 2014jra.sagepub.comDownloaded from

Paper
158SAGE Publications 2008 Los Angeles, London, New Delhi and Singapore 10.1177/1470320308096408
Journal ofthe Renin-Angiotensin-AldosteroneSystem(Including otherPeptidergic systems)
September 2008Volume 9Number 3
vehicle or ISO was, however, clearly differentonly after 10 days of treatment (figure 3). In theBB-ISO group, there was a strong increase ininterstitial collagen with respect to its control, andthe LL-ISO group showed as blue stain withtrichromo Masson. At 10 days, the perivascularfibrosis was similar for both groups receiving ISO(data not shown).
Gene expression of pro-inflammatory andpro-fibrotic mediatorsConsistent with the histological evidence ofinflammatory changes induced by ISO in theheart, the amounts of MCP-1 mRNA were signifi-cantly increased at two days of treatment only inthe BB-ISO group (figure 4). The contents ofTGF-β1 and ET-1 mRNA were also increased aftertwo days of ISO treatment (figure 4); there were,
however, no statistical differences between thetwo rat strains. There were no changes in theamounts of MCP-1, TGF-β1, and ET-1 mRNAs inboth strains at 10 days (figure 4).
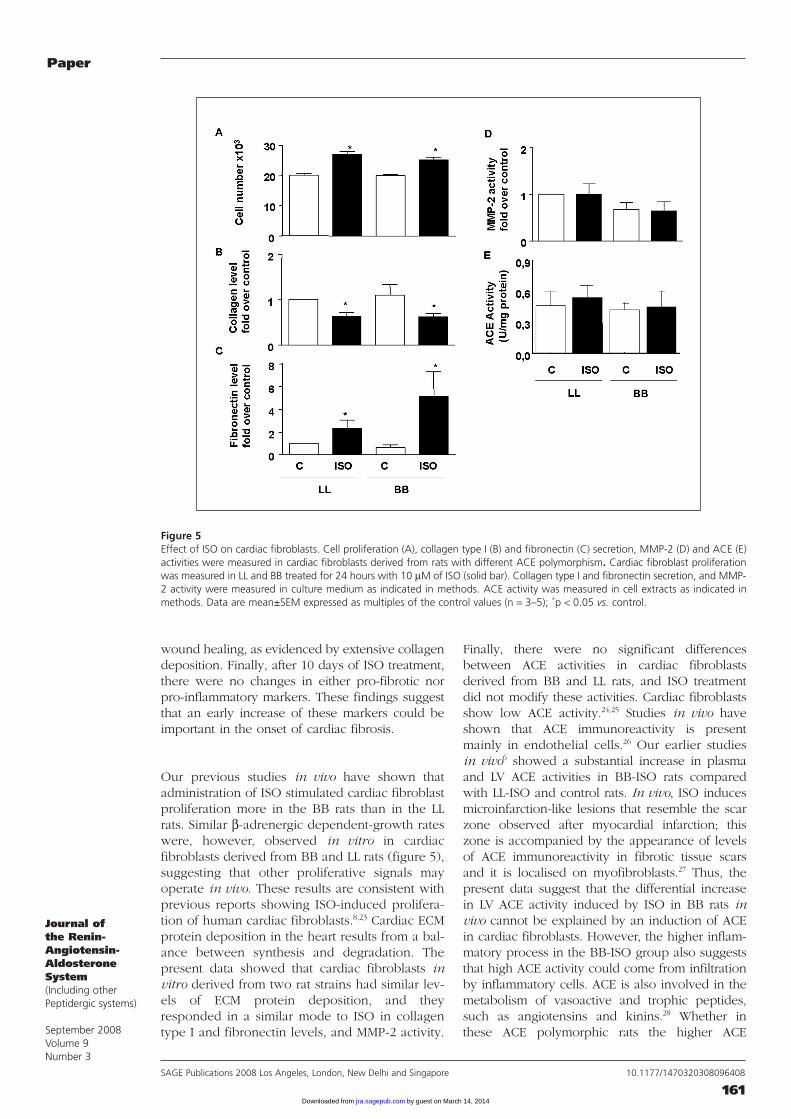
Effect of ISO on proliferation, collagen andfibronectin secretion, and ACE and MMP-2activities of cardiac fibroblasts derived fromrats with distinct ACE polymorphismCardiac fibroblasts cultured in SFM (control) didnot proliferate over a 24 hour period. Incubationwith ISO in vitro increased (p < 0.01) cardiacfibroblast proliferation obtained from both ratstrains (34±7% for LL-ISO and 26±8% for BB-ISO)(figure 5A).
Under basal conditions, the amounts of collagentype I and fibronectin in cardiac fibroblasts fromboth rat strains were similar. However, incubationwith ISO (10 µM) decreased the amount of col-lagen type I, but increased fibronectin in cardiacfibroblasts from both rat strains (figures 5B and5C), although there was no statistical differencebetween the two ISO groups. Finally, in cardiacfibroblasts isolated from both rat strains, MMP-2and ACE activities were unchanged in all condi-tions examined (figures 5D and 5E).
DiscussionThe main findings of the present study were: (1)great and numerous inflammation areas observedearly (two days) in BB-ISO rats; (2) increasedMCP-1 mRNA levels were observed early (twodays) in BB-ISO rats; (3) ISO increased TGF-β1
mRNA levels, but there was no statistical differ-ence between the two rat strains; and (4) cardiacfibroblasts derived from LL or BB rats exhibited asimilar response to ISO in vitro with regard tocell proliferation, ECM protein deposition, andMMP-2 and ACE activities.
In animals, the effects of ISO on hearts are linkedto hypertrophy and cardiac fibrosis.13-16 Thesepathological effects can be divided into early andlate phases, similar to that of the cardiac remodel-ling observed in rats with myocardial infarcts. Ourstudy showed that ISO injection induced bothphases of cardiac remodelling in both rat strains, anearly inflammatory process (figure 2), and latehypertrophy (figure 1) and fibrotic process (figure3). Our histopathology study showed that at twodays of treatment in both rat strains, ISO inducedan acute inflammatory process characterised bymyocardial changes with degeneration and necro-sis of cardiomyocytes and replacement by inflam-matory cells, distributed mainly in interstitial space,classified at score 0–1 for LL-ISO group and only at
Figure 2Effect of ISO on LV histology of ACE polymorphic rats. Lewis(LL) and Brown Norway (BB) rats were injected with saline (C)or ISO (5 mg·kg–1·day–1) for two (upper panel) or 10 (lowerpanel) days. Histological view of LV sections (transversal cross-section) of rat ventricular myocardium, stained with hema-toxylin and eosin, was photographed. Arrows indicate isolatedcardiomyocytes surrounded by infiltrative cells (arrowheads)and connective tissue. At two days in LL-ISO rat, focal and iso-lated lesions were found, meanwhile in BB-ISO rat, multifocaland confluent lesions were found. At 10 days in LL-ISO rat,multifocal and confluent lesions were found, meanwhile in BB-ISO rat, a focal and isolated lesion were found.
by guest on March 14, 2014jra.sagepub.comDownloaded from

Paper
159SAGE Publications 2008 Los Angeles, London, New Delhi and Singapore 10.1177/1470320308096408
Journal ofthe Renin-Angiotensin-AldosteroneSystem(Including otherPeptidergic systems)
September 2008Volume 9Number 3
score 1 for BB-ISO group. In the chronic phase (10days), the damage area looked a finished inflam-matory process in BB-ISO rats with low granulationtissue and extensive ECM protein deposition;meanwhile, in LL-ISO rats, there appeared to be anactive inflammatory process with extensive granu-lation tissue and without excessive deposition ofECM protein. In concordance with these morpho-logical observations, our study showed a strong
increase in MCP-1 mRNA levels only in the BB-ISOgroup in the early phase (two days). Although ourstudy did not determine in which cardiac cell typethe expression was enhanced, MCP-1 expressionhas been localised in cardiac cells.17 Kralisch et al.18
have shown that ISO increases MCP-1 mRNA levelsin fibroblasts. Whether higher ACE activity in BBrats regulates the expression of MCP-1 is unclear;however, others have demonstrated that ACE
Table 2Score of ISO-induced inflammatory and cardiac damage in rats with distinct ACE gene polymorphism.
Two days of treatment
Score LL-C LL-ISO BB-C BB-ISO
0 4 3 4 01 0 3 0 5Total 4 6 4 5
Ten days of treatment
Score LL-C LL-ISO BB-C BB-ISO
0 4 4 7 41 1 4 0 3Total 5 8 7 7
Key: Values are number of rats in this score. Homozygous F2 Lewis (LL) and Brown Norway (BB) rats were injected with saline(C) or ISO (5 mg/kg/day) for two or 10 days. One day after the last injection, LV cross-sections were evaluated for inflammatoryor cardiac damage as described in methods.
Figure 3Effect of ISO on LV interstitial fibrosis of rats with different ACE polymorphism. Lewis (LL) and Brown Norway (BB) rats wereinjected with saline (C) or ISO (5 mg·kg–1·day–1) for 10 days. Histological view of LV sections (transversal cross-section) of rat ven-tricular myocardium, stained with trichrome Masson’s, was photographed. Arrows indicate collagen deposition (blue), and arrow-heads infiltrative inflammatory cell (brown). In LL-ISO rat, low blue stain was found, meanwhile in BB-ISO rat, intense blue stainwas observed.
by guest on March 14, 2014jra.sagepub.comDownloaded from

Paper
160SAGE Publications 2008 Los Angeles, London, New Delhi and Singapore 10.1177/1470320308096408
Journal ofthe Renin-Angiotensin-AldosteroneSystem(Including otherPeptidergic systems)
September 2008Volume 9Number 3
inhibitors prevent the increase in the amounts ofMCP-1 and TGF-β1 mRNAs caused by Ang II.19 Theimportance of the inflammatory cell response inleading to fibrous tissue formation has beenshown before.20,21 A key role is played by MCP-1,which controls not only the accumulation ofmacrophages, but also fibroblast proliferation, theinduction of TGF-β1 and the appearance of fibro-sis.20 Omura et al.22 have shown that ISO induces amarked increase in TGF-β1 in vivo, and this canstimulate the differentiation of fibroblasts to myofi-broblasts. Although pro-fibrotic factors (TGF-β1,ET-1) can stimulate ECM protein deposition, theyare by themselves not sufficient to induce fibrosis.We can, however, suggest that these factorstogether with inflammatory factors are implicatedin the greater extent of cardiac fibrosis in theBB-ISO group compared with the LL-ISO group.
We have previously described that, at two days oftreatment, ISO increases both cell proliferation ofnon-cardiomyocyte cells and MMP-9 activity in BB-ISO rats more than in LL-ISO rats.6 These findingsare coincident with an inflammatory process inwhich MCP-1 seems to have a key role. The higherMMP-9 activity in the BB-ISO group suggests thatthis MMP activity could come from infiltration byinflammatory cells.
Extensive myocardial alterations were observedafter 10 days of ISO injection in both BB and LLrats. In BB-ISO rats, however, the myocardialchanges were characterised by large areas withisolated cardiomyocytes individually or in groupssurrounded by extensive deposition of ECMprotein (mainly collagen) and non-myocyte cells.These damage areas resembled the final stage of
Figure 4Effect of ISO on mRNA levels of MCP-1, TGF-β and ET-1, in LV tissue of rats with different ACE polymorphism. Lewis (LL) andBrown Norway (BB) rats were injected with saline (C) or ISO (5 mg·kg–1·day–1) for two or 10 days. One day after the last injection,mRNA of LV tissue was extracted and analysed by RT-PCR. Data are mean±SEM expressed as multiples of the control value; n = 5–6;*p < 0.05 vs. control, #p < 0.05 vs. genotype.
by guest on March 14, 2014jra.sagepub.comDownloaded from
Paper
161SAGE Publications 2008 Los Angeles, London, New Delhi and Singapore 10.1177/1470320308096408
Journal ofthe Renin-Angiotensin-AldosteroneSystem(Including otherPeptidergic systems)
September 2008Volume 9Number 3
wound healing, as evidenced by extensive collagendeposition. Finally, after 10 days of ISO treatment,there were no changes in either pro-fibrotic norpro-inflammatory markers. These findings suggestthat an early increase of these markers could beimportant in the onset of cardiac fibrosis.
Our previous studies in vivo have shown thatadministration of ISO stimulated cardiac fibr oblastproliferation more in the BB rats than in the LLrats. Similar β-adrenergic dependent-growth rateswere, however, observed in vitro in cardiacfibroblasts derived from BB and LL rats (figure 5),suggesting that other proliferative signals mayoperate in vivo. These results are consistent withprevious reports showing ISO-induced prolifera-tion of human cardiac fibroblasts.8,23 Cardiac ECMprotein deposition in the heart results from a bal-ance between synthesis and degradation. Thepresent data showed that cardiac fibroblasts invitro derived from two rat strains had similar lev-els of ECM protein deposition, and theyresponded in a similar mode to ISO in collagentype I and fibronectin levels, and MMP-2 activity.
Finally, there were no significant differencesbetween ACE activities in cardiac fibroblastsderived from BB and LL rats, and ISO treatmentdid not modify these activities. Cardiac fibroblastsshow low ACE activity.24,25 Studies in vivo haveshown that ACE immunoreactivity is presentmainly in endothelial cells.26 Our earlier studiesin vivo6 showed a substantial increase in plasmaand LV ACE activities in BB-ISO rats comparedwith LL-ISO and control rats. In vivo, ISO inducesmicroinfarction-like lesions that resemble the scarzone observed after myocardial infarction; thiszone is accompanied by the appearance of levelsof ACE immunoreactivity in fibrotic tissue scarsand it is localised on myofibroblasts.27 Thus, thepresent data suggest that the differential increasein LV ACE activity induced by ISO in BB rats invivo cannot be explained by an induction of ACEin cardiac fibroblasts. However, the higher inflam-matory process in the BB-ISO group also suggeststhat high ACE activity could come from infiltrationby inflammatory cells. ACE is also involved in themetabolism of vasoactive and trophic peptides,such as angiotensins and kinins.28 Whether inthese ACE polymorphic rats the higher ACE
Figure 5Effect of ISO on cardiac fibroblasts. Cell proliferation (A), collagen type I (B) and fibronectin (C) secretion, MMP-2 (D) and ACE (E)activities were measured in cardiac fibroblasts derived from rats with different ACE polymorphism. Cardiac fibroblast proliferationwas measured in LL and BB treated for 24 hours with 10 µM of ISO (solid bar). Collagen type I and fibronectin secretion, and MMP-2 activity were measured in culture medium as indicated in methods. ACE activity was measured in cell extracts as indicated inmethods. Data are mean±SEM expressed as multiples of the control values (n = 3–5); *p < 0.05 vs. control.
by guest on March 14, 2014jra.sagepub.comDownloaded from

Paper
162SAGE Publications 2008 Los Angeles, London, New Delhi and Singapore 10.1177/1470320308096408
Journal ofthe Renin-Angiotensin-AldosteroneSystem(Including otherPeptidergic systems)
September 2008Volume 9Number 3
plasma activity induced by ISO influences themetabolism of kinins and its relationship with thedevelopment of LV fibrosis remain unknown.Although an increase in cardiac ACE activity, asshown in BB-ISO rats, could mean a decrease ofkinin levels, it has recently been demonstrated thatendogenous Ang II modulates kinin B1 receptorexpression in hypertensive rats;29 this could havestrong consequences on inflammatory process.Thus, an imbalance between local kinin levels andreceptor number in the heart could have conse-quences in mediating inflammatory responses.Finally, it remains to be confirmed in other cardiaccells expressing high ACE activity, such as cardiacmyofibroblasts and/or pro-inflammatory cells fromthese polymorphic rats, whether ISO induces aselective increase in its activity.
In summary, our study confirms that constantβ-adrenergic stimulation serves as a stimulus totrigger LVH and fibrosis, and that they areincreased in Brown-Norway as compared toLewis rats. Further studies need to be performedto delineate the mechanism whereby ISO inducesthe expression of the genes for pro-inflammatorycytokines in BB-ISO rat hearts.
AcknowledgementsWe thank Dr Christopher Pogson for his help inreviewing this manuscript, and Fidel Albornoz forhis technical support. This work was supportedby FONDECYT Grant 1020587 (GDA) and FON-DAP 1501006 (SL).
References1. Weber KT, Brilla CG. Pathological hypertrophy and car-diac interstitium. Fibrosis and renin-angiotensin-aldosteronesystem. Circulation 1991;38:1849-65.2. Dzau VJ. Tissue renin-angiotensin system in myocardialhypertrophy and failure. Arch Int Med 1993;153:937-42.3. de Gasparo M, Catt KJ, Inagami T, Wright JW, Unger T.International union of pharmacology. XXIII. The angiotensinII receptors. Pharmacol Rev 2000;52:415-72.4. Challah M, Villard E, Philippe M et al. Angiotensin I-converting enzyme genotype influences arterial response toinjury in normotensive rats. Arterioscler Thromb Vasc Biol1998;18:235-43.5. Ocaranza MP, Piddo AM, Faundez P, Lavandero S, JalilJE. Angiotensin I-converting enzyme gene polymorphisminfluences chronic hypertensive response in the rat Goldblattmodel. J Hypertens 2002;20:413-20.6. Ocaranza MP, Diaz-Araya G, Carreno JE et al.Polymorphism in gene coding for ACE determines differentdevelopment of myocardial fibrosis in rats. Am J Physiol2004;286:H498-H506.7. Meszaros JG, Gonzalez AM, Endo-Mochizuki Y, VillegasS, Villarreal F, Brunton LL. Identification of G protein-coupledsignaling pathways in cardiac fibroblasts: cross talk betweenGq and Gs. Am J Physiol 2000;278:C154-C162.8. Turner NA, Porter KE, Smith WH, White HL, Ball SG,Balmforth AJ. Chronic beta2-adrenergic receptor stimulation
increases proliferation of human cardiac fibroblasts via anautocrine mechanism. Cardiovasc Res 2003;57:784-92.9. Sun Y, Zhang J, Lu L, Bedigian MP, Robinson AD, WeberKT. Tissue angiotensin II in the regulation of inflammatoryand fibrogenic components of repair in the rat heart. J LabClin Med 2004;143:41-51.10. Villarreal FJ, Kim NN, Ungab GD, Printz MP, DillmannWH. Identification of functional angiotensin II receptors onrat cardiac fibroblasts. Circulation 1993;88:2849-61.11. Gray MO, Long CS, Kalinyak JE, Li HT, Karliner JS.Angiotensin II stimulates cardiac myocyte hypertrophy viaparacrine release of TGF-β1 and endothelin-1 from fibroblasts.Cardiovasc Res 1998;40:352-63.12. Claycomb WC, Palazzo MC. Culture of the terminallydifferentiated adult cardiac muscle cell: a light and scanningelectron microscope study. Dev Biol 1980;80:466-82.13. Stanton HC, Brenner G, Mayfield ED Jr. Studies on iso-proterenol-induced cardiomegaly in rats. Am Heart J 1969;77:72-80.14. Laks M, Morady MF, Swan HJ. Myocardial hypertrophyproduced by infusion of subhypertensive doses of norephine -phrine in the dog. Chest 1973;64:75-8.15. Boluyt MO, Long X, Eschenhagen T et al. Isoproterenolinfusion induces alterations in expression of hypertrophy-asso-ciated genes in rat heart. Am J Physiol 1995;269:H638-H647.16. Sun Y, Weber KT. Animal models of cardiac fibrosis.Methods Mol Med 2005;117:273-90.17. Hohensinner PJ, Kaun C, Rychli K et al. Monocytechemoattractant protein (MCP-1) is expressed in human car-diac cells and is differentially regulated by inflammatorymediators and hypoxia. FEBS Lett 2006;580:3532-8.18. Kralisch S, Klein J, Lossner U et al. Isoproterenol stim-ulates monocyte chemoattractant protein-1 expression andsecretion in 3T3-L1 adipocytes. Regul Pept 2006;135:12-16.19. Amann B, Tinzmann R, Angelkort B. ACE inhibitorsimprove diabetic nephropathy through suppression of renalMCP-1. Diabetes Care 2003;26:2421-5.20. Kuwahara F, Kai H, Tokuda K et al. Hypertensivemyocardial fibrosis and diastolic dysfunction - another modelof inflammation? Hypertension 2004;43:739-45.21. Weber KT. From inflammation to fibrosis: a stiff stretchof highway. Hypertension 2004;43:716-1922. Omura T, Kim S, Takeuchi K, Iwao H, Takeda T.Transforming growth factor beta 1 and extracellular matrixgene expression in isoprenaline induced cardiac hypertrophy:effects of inhibition of the renin-angiotensin system.Cardiovasc Res 1994;12:1835-42.23. Colombo F, Noel J, Mayers P, Calderone A. Beta adren-ergic stimulation of rat cardiac fibroblast promotes proteinsynthesis via the activation of phosphatidylinositol 3-kinase.J Mol Cell Cardiol 2001;33:1091-106.24. Petrov VV, Fagard RH, Lijnen PJ. Transforming growthfactor-ß1 induces angiotensin-converting enzyme synthesis inrat cardiac fibroblasts during their differentiation to myofi-broblasts. J Renin Angiotensin Aldosterone Syst 2000;1:342-52.25. Lijnen PJ, Petrov VV. Role of intracardiac renin-angiotensin-aldosterone system in extracellular matrix remod-eling. Methods Find Exp Clin Pharmacol 2003;25:541-64.26. Falkenhahn M, Franke F, Bohle RM et al. Cellular distri-bution of angiotensin-converting enzyme after myocardialinfarction. Hypertension 1995;25:219-26.27. Sun Y, Weber KT. Angiotensin converting enzyme andmyofibroblasts during tissue repair in the rat heart. J Mol CellCardiol 1996;28:851-8.28. Tschope C, Schultheiss HP, Walther T. Multiple interac-tions between the renin-angiotensin and the kallikrein-kininsystems: role of ACE inhibition and AT1 receptor blockade.J Cardiovasc Pharmacol 2002;39:478-87.29. Fernandes L, Ceravolo GS, Fortes ZB et al. Modulationof kinin B1 receptor expression by endogenous angiotensin IIin hypertensive rats. Regul Pept 2006;136:92-7.
by guest on March 14, 2014jra.sagepub.comDownloaded from




















