
Homocysteine Induces Expression and Secretion of Monocyte Chemoattractant Protein1 and Interleukin8...
8
Homocysteine Induces Expression and Secretion of Monocyte Chemoattractant Protein-1 and Interleukin-8 in Human Aortic Endothelial Cells Implications for Vascular Disease Ranjana Poddar, PhD; Natarajan Sivasubramanian, PhD; Patricia M. DiBello, MS; Killian Robinson, MD; Donald W. Jacobsen, PhD Background—Proinflammatory cytokines play key roles in atherogenesis and disease progression. Because hyperhomo- cysteinemia is an independent risk factor for cardiovascular disease, we hypothesized that homocysteine could be atherogenic by altering the expression of specific cytokines in vascular endothelial cells. Methods and Results—Northern blot and RNase protection assays showed that DL-homocysteine induced mRNA expression of the proinflammatory cytokines monocyte chemoattractant protein-1 (MCP-1) and interleukin-8 (IL-8) in cultured human aortic endothelial cells (HAECs). Homocysteine had no effect on expression of other cytokines, namely tumor necrosis factor-a, granulocyte-macrophage colony–stimulating factor, interleukin-1b, and transforming growth factor-b. MCP-1 mRNA expression increased 1 hour after homocysteine treatment, reached a maximum within 2 to 4 hours, and declined to basal levels over the next 24 hours. Induction of mRNA expression for both chemokines was observed with as little as 10 mmol/L DL-homocysteine, and maximal expression was achieved with 50 mmol/L DL-homocysteine. Homocysteine also triggered the release of MCP-1 and IL-8 protein from HAECs into the culture medium. The induction was specific for homocysteine, because equimolar concentrations of L-homocystine, L-cysteine, and L-methionine had no effect on mRNA levels and protein release. Furthermore, L-homocysteine induced chemokine expression, but D-homocysteine did not, thus demonstrating enantiomeric specificity. The culture medium from homocysteine-treated HAECs promoted chemotaxis in human peripheral blood monocytes and U937 cells. Anti– human recombinant MCP-1 antibody blocked the migration. Conclusions—Pathophysiological levels of L-homocysteine alter endothelial cell function by upregulating MCP-1 and IL-8 expression and secretion. This suggests that L-homocysteine may contribute to the initiation and progression of vascular disease by promoting leukocyte recruitment. (Circulation. 2001;103:2717-2723.) Key Words: homocysteine n peptides n endothelium n cells n cardiovascular diseases C ytokines play critical roles in atherosclerosis and inflam- matory diseases. Dysregulation of cytokine expression in vascular cells may contribute to the atherosclerotic process, and we now report that homocysteine can enhance the expression of specific cytokines in human aortic endothelial cells (HAECs). Homocysteine, an intermediary metabolite of methionine, is an independent risk factor for cardiovascular disease. 1,2 Whether hyperhomocysteinemia is causal or merely a marker for cardiovascular disease is the subject of active investigation. Previous in vivo and in vitro studies suggest that homo- cysteine changes the endothelium from a nonthrombogenic to a thrombogenic phenotype 3 and limits the bioavailability of nitric oxide. 4 The “response to injury” hypothesis, whereby the vascular endothelium becomes “activated” in response to injurious agents, may apply to homocysteine as well. 5 Acti- vated endothelial cells release proinflammatory cytokines, 5,6 including chemokines, which recruit monocytes, T lympho- cytes, and neutrophils to the site of injury. 6,7 We hypothesize that homocysteine stimulates the production of specific cyto- Received October 6, 2000; revision received February 12, 2001; accepted February 12, 2001. From the Departments of Cell Biology (R.P., P.M.D., D.W.J.) and Molecular Cardiology (N.S.), Lerner Research Institute, and Department of Cardiology, Cleveland Clinic Foundation (K.R.), Cleveland, Ohio. Dr Poddar is now at the Department of Genetics, Yale University School of Medicine, New Haven, Conn; Dr Sivasubramanian is now at the Winters Center for Heart Failure Research, Section of Cardiology, Department of Medicine, Baylor College of Medicine, Veterans Affairs Medical Center, Houston, Tex; and Dr Robinson is now at the Department of Cardiology, Wake Forest University, Baptist Medical Center, Winston Salem, NC. Presented in part at the 70th Scientific Sessions of the American Heart Association, Orlando, Fla, November 9 –12, 1997, and at the Second International Conference on Homocysteine Metabolism, Nijmegen, the Netherlands, April 26 –29, 1998, and published in abstract form (Circulation. 1997;96:I-286) (Neth J Med. 1998;52[suppl]:S1). Correspondence to Donald W. Jacobsen, PhD, Department of Cell Biology, NC10, The Lerner Research Institute, The Cleveland Clinic Foundation, 9500 Euclid Ave, Cleveland, OH 44195. E-mail [email protected] © 2001 American Heart Association, Inc. Circulation is available at http://www.circulationaha.org 2717 Basic Science Reports by guest on November 24, 2015 http://circ.ahajournals.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Homocysteine Induces Expression and Secretion of Monocyte Chemoattractant Protein1 and Interleukin8...

Homocysteine Induces Expression and Secretion ofMonocyte Chemoattractant Protein-1 and Interleukin-8 in
Human Aortic Endothelial CellsImplications for Vascular Disease
Ranjana Poddar, PhD; Natarajan Sivasubramanian, PhD; Patricia M. DiBello, MS;Killian Robinson, MD; Donald W. Jacobsen, PhD
Background—Proinflammatory cytokines play key roles in atherogenesis and disease progression. Because hyperhomo-cysteinemia is an independent risk factor for cardiovascular disease, we hypothesized that homocysteine could beatherogenic by altering the expression of specific cytokines in vascular endothelial cells.
Methods and Results—Northern blot and RNase protection assays showed thatDL-homocysteine induced mRNAexpression of the proinflammatory cytokines monocyte chemoattractant protein-1 (MCP-1) and interleukin-8 (IL-8) incultured human aortic endothelial cells (HAECs). Homocysteine had no effect on expression of other cytokines, namelytumor necrosis factor-a, granulocyte-macrophage colony–stimulating factor, interleukin-1b, and transforming growthfactor-b. MCP-1 mRNA expression increased 1 hour after homocysteine treatment, reached a maximum within 2 to 4hours, and declined to basal levels over the next 24 hours. Induction of mRNA expression for both chemokines wasobserved with as little as 10mmol/L DL-homocysteine, and maximal expression was achieved with 50mmol/LDL-homocysteine. Homocysteine also triggered the release of MCP-1 and IL-8 protein from HAECs into the culturemedium. The induction was specific for homocysteine, because equimolar concentrations ofL-homocystine,L-cysteine,andL-methionine had no effect on mRNA levels and protein release. Furthermore,L-homocysteine induced chemokineexpression, butD-homocysteine did not, thus demonstrating enantiomeric specificity. The culture medium fromhomocysteine-treated HAECs promoted chemotaxis in human peripheral blood monocytes and U937 cells. Anti–humanrecombinant MCP-1 antibody blocked the migration.
Conclusions—Pathophysiological levels ofL-homocysteine alter endothelial cell function by upregulating MCP-1 and IL-8expression and secretion. This suggests thatL-homocysteine may contribute to the initiation and progression of vasculardisease by promoting leukocyte recruitment.(Circulation. 2001;103:2717-2723.)
Key Words: homocysteinen peptidesn endotheliumn cells n cardiovascular diseases
Cytokines play critical roles in atherosclerosis and inflam-matory diseases. Dysregulation of cytokine expression
in vascular cells may contribute to the atherosclerotic process,and we now report that homocysteine can enhance theexpression of specific cytokines in human aortic endothelialcells (HAECs). Homocysteine, an intermediary metabolite ofmethionine, is an independent risk factor for cardiovasculardisease.1,2 Whether hyperhomocysteinemia is causal ormerely a marker for cardiovascular disease is the subject ofactive investigation.
Previous in vivo and in vitro studies suggest that homo-cysteine changes the endothelium from a nonthrombogenic toa thrombogenic phenotype3 and limits the bioavailability ofnitric oxide.4 The “response to injury” hypothesis, wherebythe vascular endothelium becomes “activated” in response toinjurious agents, may apply to homocysteine as well.5 Acti-vated endothelial cells release proinflammatory cytokines,5,6
including chemokines, which recruit monocytes, T lympho-cytes, and neutrophils to the site of injury.6,7 We hypothesizethat homocysteine stimulates the production of specific cyto-
Received October 6, 2000; revision received February 12, 2001; accepted February 12, 2001.From the Departments of Cell Biology (R.P., P.M.D., D.W.J.) and Molecular Cardiology (N.S.), Lerner Research Institute, and Department of
Cardiology, Cleveland Clinic Foundation (K.R.), Cleveland, Ohio. Dr Poddar is now at the Department of Genetics, Yale University School of Medicine,New Haven, Conn; Dr Sivasubramanian is now at the Winters Center for Heart Failure Research, Section of Cardiology, Department of Medicine, BaylorCollege of Medicine, Veterans Affairs Medical Center, Houston, Tex; and Dr Robinson is now at the Department of Cardiology, Wake Forest University,Baptist Medical Center, Winston Salem, NC.
Presented in part at the 70th Scientific Sessions of the American Heart Association, Orlando, Fla, November 9–12, 1997, and at the SecondInternational Conference on Homocysteine Metabolism, Nijmegen, the Netherlands, April 26–29, 1998, and published in abstract form (Circulation.1997;96:I-286) (Neth J Med.1998;52[suppl]:S1).
Correspondence to Donald W. Jacobsen, PhD, Department of Cell Biology, NC10, The Lerner Research Institute, The Cleveland Clinic Foundation,9500 Euclid Ave, Cleveland, OH 44195. E-mail [email protected]
© 2001 American Heart Association, Inc.
Circulation is available at http://www.circulationaha.org
2717
Basic Science Reports
by guest on November 24, 2015http://circ.ahajournals.org/Downloaded from

kines important in recruitment of leukocytes to sites ofvascular injury.
We show that pathophysiological concentrations of homo-cysteine induce mRNA expression and trigger protein releasefor monocyte chemoattractant protein-1 (MCP-1) and inter-leukin (IL)-8 in cultured HAECs. The expression of tumornecrosis factor-a (TNF-a), granulocyte-macrophage colony–stimulating factor (GM-CSF), IL-1b, and transforminggrowth factor-b (TGF-b) was unaffected by homocysteine.Stimulated expression and release of MCP-1, a chemokine formonocytes, and IL-8, a chemokine for T lymphocytes andneutrophils, suggest that homocysteine may alter endothelialcell function and promote atherogenesis.
MethodsCell CulturePrimary cultures of HAECs were established after collagenasetreatment of segments of human thoracic aorta, obtained as discardedtissue from donor hearts used in heart transplant surgery (approvedby the Institutional Review Board of the Cleveland Clinic Founda-tion). Cells were grown in medium 199 (Gibco-BRL) supplementedwith 150mg/mL endothelial cell growth factor,8 90 mg/mL heparin,20% FCS (BioWhittaker), and an antibiotic/antimycotic mixture(Gibco-BRL). For all experiments, confluent HAEC cultures (pas-sages 5 to 7) were incubated with fresh culture medium for 24 hoursand then treated with homocysteine or other compounds. Endotoxinlevels of theDL-homocysteine (ICN Pharmaceuticals) working solu-tions, prepared in complete medium, were,0.012 ng/mL (Limulusamoebocyte lysate assay; Associates of Cape Cod).L-Homocysteineand D-homocysteine were freshly prepared from their respectivethiolactones (Sigma) by a method modified from Hatch et al.9 Finalthiol concentrations were determined spectrophotometrically.10
Northern Hybridization AssaysTotal RNA (10mg), extracted with RNAeasy (Qiagen), was electro-phoresed on agarose gels11 and transferred to nylon membrane(Genescreen, NEN) for Northern blotting. After hybridization11 with32P-labeled human MCP-1 cDNA probes (740-bp insert from ATCC)and autoradiography, relative intensities were quantified by densi-tometry or phosphorimaging and normalized to GAPDH mRNAexpression. Rat GAPDH cDNA (1.3 kb) was obtained from ATCC.
RNase Protection AssaysA 32P-labeled antisense RNA probe cocktail was transcribed from acustom-designed set of cDNA templates (Pharmingen) with an invitro transcription kit (Riboquant, Pharmingen). Each template had aunique length and represented a distinct sequence in human mRNAsfor TNF-a, GM-CSF, IL-1b, MCP-1, IL-8, TGF-b, GAPDH, andL-32 ribosomal RNA. The radiolabeled antisense probes werehybridized to equal amounts of total RNA (2mg) and subjected toRNase digestion with an RNase protection assay kit (Riboquant,Pharmingen). The RNase-protected fragments were purified, re-solved on 4.75% denaturing polyacrylamide gels (National Diagnos-tics), and autoradiographed. The protected fragments, which mi-grated slightly ahead of their respective undigested marker probes,were identified by extent of migration in comparison to the sizes ofundigested marker probes. The relative intensities were quantified bydensitometry or phosphorimaging and normalized to GAPDH andL-32 mRNA. Yeast tRNA served as a negative control.
Measurement of MCP-1 and IL-8 Protein ReleaseFrom HAECsHAEC cultures were treated with or withoutDL-homocysteine orother test compounds for up to 8 hours. MCP-1 and IL-8 proteinconcentrations were determined in the culture supernatants byELISA (Quantikine, R&D Systems) and were normalized to total
cell protein, which was measured with the BCA protein assay kit(Pierce). Basal concentrations of MCP-1 and IL-8 in the mediumfrom untreated cells were subtracted from the concentrations foundin the treated cells at each time point. In some experiments, cellswere pretreated with cycloheximide (10mg/mL) for 30 minutes andthen exposed toDL-homocysteine.
MCP-1 Chemotaxis AssaysChemotaxis in response to MCP-1 was studied both in the humanhistiocytic lymphoma cell line U937 (ATCC CRL 1593) and innormal human peripheral blood monocytic cells (PBMCs). Mono-cytes were isolated from donor blood by Ficoll-Paque (Pharmacia)density gradient centrifugation, followed by adherence to serum-coated culture flasks.12 Adherent cells were detached, washed,resuspended in medium 199, and used in the chemotaxis assays.HAEC cultures were treated with 0, 50, or 500mmol/L DL-homocysteine or 400 U/mL TNF-a (Boehringer Mannheim) for 2 to5 hours. Culture medium was removed and transferred to the lowercompartment of 48-well Boyden chambers (Neuroprobe) (for U937)or 96-well ChemoTx microplates (Neuroprobe) (for monocytes). TheBoyden chamber was assembled with polyvinylpyrrolidone-freepolycarbonate filters (8-mm pore size) and preblocked with FCS.Purified PBMCs or U937 cells (13105 to 2.53105/50 mL) wereadded to the upper compartment and incubated for 90 minutes at37°C. Monocytes adhering to the top surface of the filter werescraped off, and those migrating to the lower chamber and adheringto the underside of the filter were fixed, stained (Hema 3 stain kit,Fisher), and counted (5 fields/well). The U937 cells that hadmigrated to the lower chamber were counted with a hemocytometer.Conditioned medium from untreated HAECs supplemented (1 to 500ng/mL) with recombinant human MCP-1 (rhMCP-1, R&D Systems)and medium from TNF-a–treated HAECs served as positive control.Anti–human MCP-1 polyclonal antibody (R&D Systems) was added(70 mg/mL) to culture medium for 30 minutes to neutralize thesecreted MCP-1 and to assess MCP-1–specific chemotaxis. Normalrabbit IgG was used as negative control. Medium from untreatedcells was used to determine basal migration.
Statistical MethodsResults are expressed as mean6SEM. Probability values werecalculated by 1-way ANOVA with Sigmastat software (Tukey’s testor Fisher’s least significant difference test). In those instances inwhich the data were not normally distributed, the data were subjectedto logarithmic transformation before the parametric analysis. Signif-icant differences were said to exist at a value ofP,0.05. Each datapoint represents the average of 3 or 4 experiments.
ResultsEffect of Homocysteine on CytokinemRNA ExpressionTo determine whether homocysteine modulates the expres-sion of MCP-1 mRNA, HAEC cultures were treated with 500mmol/L DL-homocysteine, and total RNA was isolated atvarious times from 0.5 to 24 hours. Northern blot analysisrevealed that homocysteine induced the expression of MCP-1steady-state mRNA above basal levels within 1 hour, wasmaximal between 2 and 4 hours, and then declined to nearbaseline by 24 hours (Figure 1A). Thus, steady-state MCP-1mRNA levels were significantly increased in homocysteine-treated HAECs compared with untreated cells.
Because endothelial cells express a variety of cytokinesthat could play roles in atherogenesis, we tested the effect ofhomocysteine on expression of TNF-a, GM-CSF, IL-1b,IL-8, and TGF-b mRNAs. HAECs were treated with increas-ing concentrations ofDL-homocysteine (10mmol/L to10 mmol/L) for 3 hours. Total RNA was extracted and
2718 Circulation June 5, 2001
by guest on November 24, 2015http://circ.ahajournals.org/Downloaded from
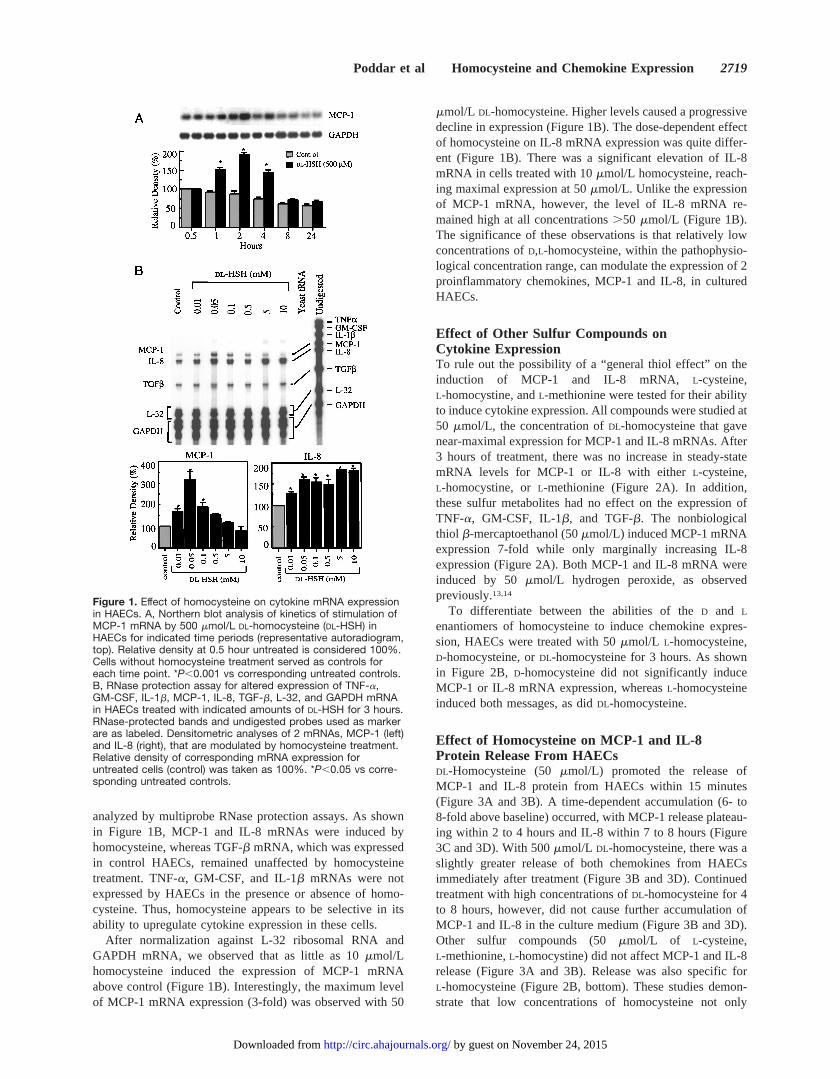
analyzed by multiprobe RNase protection assays. As shownin Figure 1B, MCP-1 and IL-8 mRNAs were induced byhomocysteine, whereas TGF-b mRNA, which was expressedin control HAECs, remained unaffected by homocysteinetreatment. TNF-a, GM-CSF, and IL-1b mRNAs were notexpressed by HAECs in the presence or absence of homo-cysteine. Thus, homocysteine appears to be selective in itsability to upregulate cytokine expression in these cells.
After normalization against L-32 ribosomal RNA andGAPDH mRNA, we observed that as little as 10mmol/Lhomocysteine induced the expression of MCP-1 mRNAabove control (Figure 1B). Interestingly, the maximum levelof MCP-1 mRNA expression (3-fold) was observed with 50
mmol/L DL-homocysteine. Higher levels caused a progressivedecline in expression (Figure 1B). The dose-dependent effectof homocysteine on IL-8 mRNA expression was quite differ-ent (Figure 1B). There was a significant elevation of IL-8mRNA in cells treated with 10mmol/L homocysteine, reach-ing maximal expression at 50mmol/L. Unlike the expressionof MCP-1 mRNA, however, the level of IL-8 mRNA re-mained high at all concentrations.50 mmol/L (Figure 1B).The significance of these observations is that relatively lowconcentrations ofD,L-homocysteine, within the pathophysio-logical concentration range, can modulate the expression of 2proinflammatory chemokines, MCP-1 and IL-8, in culturedHAECs.
Effect of Other Sulfur Compounds onCytokine ExpressionTo rule out the possibility of a “general thiol effect” on theinduction of MCP-1 and IL-8 mRNA, L-cysteine,L-homocystine, andL-methionine were tested for their abilityto induce cytokine expression. All compounds were studied at50 mmol/L, the concentration ofDL-homocysteine that gavenear-maximal expression for MCP-1 and IL-8 mRNAs. After3 hours of treatment, there was no increase in steady-statemRNA levels for MCP-1 or IL-8 with eitherL-cysteine,L-homocystine, orL-methionine (Figure 2A). In addition,these sulfur metabolites had no effect on the expression ofTNF-a, GM-CSF, IL-1b, and TGF-b. The nonbiologicalthiol b-mercaptoethanol (50mmol/L) induced MCP-1 mRNAexpression 7-fold while only marginally increasing IL-8expression (Figure 2A). Both MCP-1 and IL-8 mRNA wereinduced by 50mmol/L hydrogen peroxide, as observedpreviously.13,14
To differentiate between the abilities of theD and L
enantiomers of homocysteine to induce chemokine expres-sion, HAECs were treated with 50mmol/L L-homocysteine,D-homocysteine, orDL-homocysteine for 3 hours. As shownin Figure 2B, D-homocysteine did not significantly induceMCP-1 or IL-8 mRNA expression, whereasL-homocysteineinduced both messages, as didDL-homocysteine.
Effect of Homocysteine on MCP-1 and IL-8Protein Release From HAECsDL-Homocysteine (50mmol/L) promoted the release ofMCP-1 and IL-8 protein from HAECs within 15 minutes(Figure 3A and 3B). A time-dependent accumulation (6- to8-fold above baseline) occurred, with MCP-1 release plateau-ing within 2 to 4 hours and IL-8 within 7 to 8 hours (Figure3C and 3D). With 500mmol/L DL-homocysteine, there was aslightly greater release of both chemokines from HAECsimmediately after treatment (Figure 3B and 3D). Continuedtreatment with high concentrations ofDL-homocysteine for 4to 8 hours, however, did not cause further accumulation ofMCP-1 and IL-8 in the culture medium (Figure 3B and 3D).Other sulfur compounds (50mmol/L of L-cysteine,L-methionine,L-homocystine) did not affect MCP-1 and IL-8release (Figure 3A and 3B). Release was also specific forL-homocysteine (Figure 2B, bottom). These studies demon-strate that low concentrations of homocysteine not only
Figure 1. Effect of homocysteine on cytokine mRNA expressionin HAECs. A, Northern blot analysis of kinetics of stimulation ofMCP-1 mRNA by 500 mmol/L DL-homocysteine (DL-HSH) inHAECs for indicated time periods (representative autoradiogram,top). Relative density at 0.5 hour untreated is considered 100%.Cells without homocysteine treatment served as controls foreach time point. *P,0.001 vs corresponding untreated controls.B, RNase protection assay for altered expression of TNF-a,GM-CSF, IL-1b, MCP-1, IL-8, TGF-b, L-32, and GAPDH mRNAin HAECs treated with indicated amounts of DL-HSH for 3 hours.RNase-protected bands and undigested probes used as markerare as labeled. Densitometric analyses of 2 mRNAs, MCP-1 (left)and IL-8 (right), that are modulated by homocysteine treatment.Relative density of corresponding mRNA expression foruntreated cells (control) was taken as 100%. *P,0.05 vs corre-sponding untreated controls.
Poddar et al Homocysteine and Chemokine Expression 2719
by guest on November 24, 2015http://circ.ahajournals.org/Downloaded from

upregulate steady-state levels of mRNA for MCP-1 and IL-8but also trigger the release of the respective proteins fromHAECs.
To determine whether homocysteine stimulates de novosynthesis of MCP-1 and IL-8, HAECs were incubated withcycloheximide for 30 minutes before treatment with 50mmol/L DL-homocysteine. As shown in Figure 3E, cyclohex-imide treatment alone caused a small increase in MCP-1protein release. The stimulation of MCP-1 protein release byDL-homocysteine, however, was dramatically inhibited withcycloheximide pretreatment. This result suggests that homo-cysteine can enhance the translation of MCP-1 protein inHAECs. A similar inhibition was observed on homocysteine-induced IL-8 protein release from HAECs with cyclohexi-mide pretreatment (data not shown).
Chemotactic Activity of Secreted MCP-1Migration of PBMCs and U937 cells toward media fromDL-homocysteine–treated HAECs (50mmol/L) increased'4.5- and 1.8-fold, respectively, compared with media fromuntreated cells (Figure 4A and 4C). Media from HAECstreated with 500mmol/L DL-homocysteine for 2 hoursshowed a similar increase in migration of U937 cells. Migra-
Figure 2. Specificity of homocysteine action on chemokineexpression. A, Representative autoradiogram (top) of RNaseprotection assay for TNF-a, GM-CSF, IL-1b, MCP-1, IL-8,TGF-b, L-32, and GAPDH mRNA in HAECs treated with 50mmol/L each of DL-homocysteine (DL-HSH), L-cysteine (Cys),L-homocystine (HS-SH), L-methionine (Met), b-mercaptoethanol(BME), or hydrogen peroxide (H2O2). RNase-protected bandsand undigested probes used as marker are as labeled. Bottom,Densitometric analysis of MCP-1 (left) and IL-8 (right) mRNAs.Intensity of corresponding mRNA expression for untreated cells(control) was considered 100%. *P,0.001 vs correspondinguntreated controls. B, Enantiomeric specificity of D- andL-homocysteine action. Top, Representative autoradiogram ofRNase protection assay performed in HAECs treated with 50mmol/L L-homocysteine (L-HSH), D-homocysteine (D-HSH), orDL-homocysteine for 3 hours. Untreated cells served as control.Bottom, ELISA measurements showing MCP-1 protein levelsreleased into medium in response to treatment with homocys-teine enantiomers. *P,0.001 vs corresponding untreatedcontrols.
Figure 3. ELISA measurements showing MCP-1 (A) and IL-8 (B)protein levels released into medium by HAECs treated with 50mmol/L DL-homocysteine (DL-HSH), L-cysteine (Cys),L-methionine (Met), or L-homocystine (HS-SH) for indicated timeperiods. Untreated cells at same time points served as control.*P,0.005 vs corresponding untreated controls. C and D showpattern of MCP-1 and IL-8 protein release after treatment with 0(control), 50, or 500 mmol/L DL-homocysteine over indicatedtime periods. *P,0.05 vs corresponding untreated controls. E,Pretreatment of HAECs with cycloheximide (CHX) inhibitshomocysteine-induced MCP-1 protein release. **P,0.001 forDL-HSH (50 mmol/L) vs CHX1DL-HSH.
2720 Circulation June 5, 2001
by guest on November 24, 2015http://circ.ahajournals.org/Downloaded from

tion was inhibited by preincubation of the medium withanti–rhMCP-1, confirming MCP-1 to be the chemotacticfactor. Migration of PBMCs toward MCP-1 released byTNF-a–treated HAECs and migration of PBMCs and U937cells in response to rhMCP-1 served as positive controls(Figure 4). Anti–rhMCP-1 specifically inhibited the effect,whereas purified rabbit IgG did not. Thus, the MCP-1 proteinsecreted by homocysteine-treated HAECs had biologicalactivity.
DiscussionThere is considerable evidence that atherogenesis is aninflammatory process.5 Cytokines play critical roles as mul-tipotent mediators of inflammation by modulating key func-tions of vascular cells.5,6 In this report, we show thathomocysteine induces both mRNA expression and proteinsecretion of the proinflammatory cytokines MCP-1 and IL-8in cultured HAECs. Other cytokines studied were unaffectedby homocysteine. The effect is specific forL-homocysteine,and most significantly, the secreted MCP-1 is chemotacticallyactive. Because MCP-1 and IL-8 are major chemokines forleukocyte trafficking6 and have already been identified inatheromatous plaques,15,16 our findings suggest a potentialrole for homocysteine in atherogenesis and the progression ofvascular disease. Thus, homocysteine appears to work
through a mechanism involving upregulation of MCP-1 andIL-8 expression and recruitment of leukocytes across aninflammatory site in the vascular endothelium.
The concentrations ofDL-homocysteine (10 to 50mmol/L)used in this study are comparable to pathophysiological levelsobserved in homocystinuric subjects (up to 100mmol/L).17
Recent studies have reported cellular responses that areinduced by clinically relevant levels of homocysteine (up to100mmol/L) but not byL-cysteine.18–20Wang et al18 showedthat homocysteine inhibited endothelial cell proliferation,presumably after its conversion toS-adenosylhomocysteineand subsequent hypomethylation of p21ras. Hajjar’s groupreported that homocysteine, but not cysteine, inhibited thebinding of tissue plasminogen activator to annexin II,19
probably by mechanisms involving thiolation of Cys9.21
Lipton et al20 found that 10 to 100mmol/L DL-homocysteinein the presence of elevated glycine stimulatedN-methyl-D-aspartate receptors in neurons, resulting in neurotoxicity fromexcessive Ca21 influx and generation of reactive oxygenspecies. We also show that clinically relevant concentrationsof DL-homocysteine (10 to 50mmol/L) promote MCP-1 andIL-8 expression and secretion.
The induction of MCP-1 and IL-8 expression appears to bespecific forL-homocysteine, becauseL-homocystine, an oxi-dized form of homocysteine, andL-methionine, the precursorof homocysteine, did not induce chemokine expression.L-Cysteine was also ineffective. Could the atherogenicity ofhomocysteine be related to the high pK of its sulfhydrylgroup? The higher pK of homocysteine relative to cysteinesuggests greater reactivity in nucleophilic exchange reactions.This may explain the tolerance of such high levels of cysteinein vivo. Although b-mercaptoethanol induces expression ofMCP-1 but not IL-8, it should be noted that the thiol pKs ofhomocysteine andb-mercaptoethanol are similar, and thiscould also relate them mechanistically. The fact thatb-mercaptoethanol was a more potent inducer than homocys-teine could be explained by its greater transportability22 thanhomocysteine and its ability to activate nuclear factor-kB.23
Additional studies are needed to address the possibility thathomocysteine-induced chemokine expression, too, may bemediated by activation of specific transcription factors.
We also demonstrate thatL-homocysteine promotes asubstantial release of MCP-1 and IL-8 protein from HAECs.Inhibition of chemokine release by cycloheximide shows thathomocysteine stimulates the translation of MCP-1 and IL-8,whereas the rapidity of the release indicates that homocys-teine may also trigger chemokine secretion from endogenousstores. Cycloheximide treatment for 1 hour reduceshomocysteine-induced MCP-1 release dramatically, to'25%of the value seen with cycloheximide alone. This demon-strates that MCP-1 release requires de novo protein synthesis,although there is likely to be a small contribution fromendogenous stores. Thus, the homocysteine-induced mRNAlevels probably reflect an upregulation of transcription orincreased mRNA stability.
To explain mechanistically the induction of MCP-1 andIL-8 expression by homocysteine in HAECs, 2 possibilitiesshould be considered. First, H2O2 and other reactive oxygenspecies that are generated extracellularly during auto-
Figure 4. Chemotaxis assay. Percentage of migrating PBMCs(A) and U937 cells (C) toward conditioned media from HAECstreated with indicated concentrations of DL-homocysteine (HSH)or TNF-a for 2 to 6 hours relative to that from untreated cells(taken as 100%). Media from untreated cells served as control.*P,0.001 vs corresponding untreated controls. Preincubation ofconditioned media from HSH- or TNF-a–treated cells with neu-tralizing MCP-1 antibody (Ab) resulted in a significant inhibitionof chemotaxis (**P,0.05). B and D, Chemotaxis of PBMCs andU937 cells to purified rhMCP-1. Media that do not contain addi-tional rhMCP-1 serve as untreated control. *P,0.001 vs corre-sponding untreated controls. Preincubation of rhMCP-1–containing medium with neutralizing MCP-1 Ab resulted in a sig-nificant inhibition of chemotaxis (**P,0.05), whereas preincuba-tion with rabbit IgG had no effect.
Poddar et al Homocysteine and Chemokine Expression 2721
by guest on November 24, 2015http://circ.ahajournals.org/Downloaded from

oxidation of homocysteine could mediate the observed effectsthrough oxidative stress. In fact, as we and others havefound,13,14 both MCP-1 and IL-8 mRNA expression can betriggered by reactive oxygen species, including H2O2. Thus,by use of supraphysiological homocysteine concentrations(.1 mmol/L), auto-oxidation could result in oxidative stress,thereby inducing chemokine expression. We found, however,that at high homocysteine concentrations (ie,.50 mmol/L),MCP-1 mRNA expression actually declined.
Furthermore, our observation thatL-homocysteine but notD-homocysteine induces MCP-1 and IL-8 expression suggeststhat the mechanism must involve$1 stereospecific step.Trace-metal–catalyzed auto-oxidation of thiols, includingD-andL-homocysteine, is likely to be nonstereospecific. Thus, ifthe mechanism of chemokine induction involves solely thegeneration of extracellular reactive oxygen species, then bothD- and L-homocysteine should have similar effects. Also, inmild hyperhomocysteinemia, in which concentrations ofplasma total homocysteine usually do not exceed 30mmol/L,the amount of H2O2 generated would probably be of littleconsequence, given the overwhelming antioxidant defensesystem in the blood.24 Although this argues against theoxidative stress hypothesis, additional studies with catalaseand superoxide dismutase are needed.
Alternatively, thiols themselves can act directly within thecell to induce gene expression.23 Two facets of this studysupport the hypothesis that homocysteine is also acting withinthe cell on as yet unidentified molecular targets. First, wewere able to induce chemokine expression at relatively lowhomocysteine concentrations (10 to 50mmol/L), but wefound that equimolar concentrations of cysteine were ineffec-tive. Second, because theD enantiomer of homocysteine didnot induce chemokine expression, stereospecific processessuch as transport and/or enzyme catalysis may be involved inL-homocysteine–induced expression of MCP-1 and IL-8.Homocysteine molecular targeting may occur through alter-ation of the redox status within the cell. A reduction inintracellular glutathione25 and decreased activity and expres-sion of glutathione peroxidase26 after homocysteine treatmentmay lead to the expression of redox-sensitive transcriptionfactors.27 Homocysteine also induces endoplasmic reticulumstress in endothelial cells by a mechanism that may involvealtered redox potential.28 We are currently investigatingaltered redox status in homocysteine-treated endothelial cellsas a mechanism for increased chemokine expression.
Recent evidence suggests that in addition to activation andchemotaxis of neutrophils,6 IL-8 promotes directed migrationof T lymphocytes.29 Both neutrophils and T lymphocytes areimportant players in inflammation and pathogenesis of ath-erosclerosis.5 MCP-1 is known to induce migration of mono-cytes at subnanomolar concentrations30 and to recruit a subsetof T cells.31 Because the gradient for MCP-1 is soluble ratherthan haptotactic,32 it would be difficult to maintain high levelsof the protein on the luminal surface of the endothelial cellsbecause of blood flow in vivo. Therefore, the MCP-1 accu-mulating at the abluminal surface of the endothelium afteractivation establishes an MCP-1 gradient across the endothe-lial monolayer, potentiating transendothelial migration ofmonocytes. This theory has been shown to be true in vitro.32
Accordingly, our finding that the MCP-1 released fromhomocysteine-activated endothelial cells is biologically ac-tive and capable of promoting chemotaxis provides a role forhomocysteine in vascular disease. Because monocyte infiltra-tion is one of the key events in atherogenesis, we propose thathomocysteine induces the accumulation of MCP-1 and IL-8and causes recruitment and directed migration of leukocytesthrough the endothelium. In summary, we have shown thathomocysteine induces the expression of the proinflammatorychemokines MCP-1 and IL-8 in HAECs. Our findings sug-gest a novel role for homocysteine in the pathogenesis ofvascular disease.
AcknowledgmentsThis work was supported by grant RO1-HL-52334 (Dr Jacobsen)from the National Institutes of Health. We thank Dr ChristineMoravec of the Cleveland Clinic Foundation for providing humanaorta segments.
References1. Mayer EL, Jacobsen DW, Robinson K. Homocysteine and coronary
atherosclerosis.J Am Coll Cardiol. 1996;27:517–527.2. Refsum H, Ueland P, Nygård O, et al. Homocysteine and cardiovascular
disease.Annu Rev Med. 1998;49:31–62.3. Lentz SR. Homocysteine and vascular dysfunction.Life Sci. 1997;61:
1205–1215.4. Jacobsen DW. Cellular mechanisms of pathogenesis in atherosclerosis.
In: Carmel R, Jacobsen DW, eds.Homocysteine in Health and Disease.Cambridge UK: Cambridge University Press; 2001.
5. Ross R. The pathogenesis of atherosclerosis: a perspective for the 1990s.Nature. 1993;362:801–809.
6. Rollins BJ. Chemokines.Blood. 1997;90:909–928.7. Dudman NPB, Temple SE, Guo XW, et al. Homocysteine enhances
neutrophil-endothelial interactions in both cultured human cells and ratsin vivo. Circ Res. 1999;84:409–416.
8. Maciag T, Cerondolo J, Ilsley S, et al. Endothelial cell growth factor frombovine hypothalamus: identification and partial characterization.ProcNatl Acad Sci U S A. 1979;76:5674–5678.
9. Hatch FT, Larrabee AR, Cathou RE, et al. Enzymatic synthesis of themethyl group of methionine.J Biol Chem. 1961;236:1095–1101.
10. Ellman GL. Tissue sulfhydryl groups.Arch Biochem Biophys. 1959;82:70–77.
11. Poddar R, Paul S, Chaudhury S, et al. Regulation of tublin and actin geneexpression by thyroid hormone during rat brain development.Mol BrainRes. 1996;35:111–118.
12. Kumagai K, Itoh K, Hinuma S, et al. Pretreatment of plastic Petri disheswith fetal calf serum: a simple method for macrophage isolation.J Immunol Methods. 1979;29:17–25.
13. DeForge LE, Preston AM, Takeuchi E, et al. Regulation of interleukin 8gene expression by oxidant stress.J Biol Chem. 1993;268:25568–25576.
14. Wung BS, Cheng JJ, Hsieh HJ, et al. Cyclic strain-induced monocytechemotactic protein-1 gene expression in endothelial cells involvesreactive oxygen species activation of activator protein 1.Circ Res. 1997;81:1–7.
15. Nelken NA, Coughlin SR, Gordon D, et al. Monocyte chemoattractantprotein-1 in human atheromatous plaques.J Clin Invest. 1991;88:1121–1127.
16. Liu Y, Hulten LM, Wiklund O. Macrophages isolated from human ath-erosclerotic plaques produce IL-8, and oxysterols may have a regulatoryfunction for IL-8 production.Arterioscler Thromb Vasc Biol. 1997;17:317–323.
17. Mansoor MA, Ueland PM, Aarsland A, et al. Redox status and proteinbinding of plasma homocysteine and other aminothiols in patients withhomocystinuria.Metabolism. 1993;42:1481–1485.
18. Wang H, Yoshizumi M, Lai K, et al. Inhibition of growth and p21ras
methylation in vascular endothelial cells by homocysteine but notcysteine.J Biol Chem. 1997;272:25380–25385.
2722 Circulation June 5, 2001
by guest on November 24, 2015http://circ.ahajournals.org/Downloaded from

19. Hajjar KA. Homocysteine-induced modulation of tissue plasminogenactivator binding to its endothelial cell membrane receptor.J Clin Invest.1993;91:2873–2879.
20. Lipton SA, Kim WK, Choi YB, et al. Neurotoxicity associated with dualactions of homocysteine at theN-methyl-D-aspartate receptor.Proc NatlAcad Sci U S A. 1997;94:5923–5928.
21. Hajjar KA, Mauri L, Jacovina AT, et al. Tissue plasminogen activatorbinding to the annexin II tail domain: direct modulation by homocysteine.J Biol Chem. 1998;273:9987–9993.
22. Ishii T, Bannai S, Sugita Y. Mechanism of growth stimulation of L1210cells by 2-mercaptoethanol: role of the mixed disulfide of2-mercaptoethanol and cysteine.J Biol Chem. 1981;256:12387–12392.
23. Das KC, Lewis-Molock Y, White CW. Activation of NF-kappaB andelevation of MnSOD gene expression by thiol reducing agents in lungadenocarcinoma (A549) cells.Am J Physiol. 1995;269:L588–L602.
24. Jacobsen DW. Hyperhomocysteinemia and oxidative stress: time for areality check?Arterioscler Thromb Vasc Biol. 2000;20:1182–1184.
25. Blundell G, Jones BG, Rose FA, et al. Homocysteine mediated endothe-lial cell toxicity and its amelioration.Atherosclerosis. 1996;122:163–172.
26. Upchurch GR Jr, Welch GN, Fabian AJ, et al. Homocyst(e)ine decreasesbioavailable nitric oxide by a mechanism involving glutathione per-oxidase.J Biol Chem. 1997;272:17012–17017.
27. Sen CK, Packer L. Antioxidant and redox regulation of gene tran-scription.FASEB J. 1996;10:709–720.
28. Outinen PA, Sood SK, Pfeifer SI, et al. Homocysteine-induced endo-plasmic reticulum stress and growth arrest leads to specific changes ingene expression in human vascular endothelial cells.Blood. 1999;94:959–967.
29. Balkwill F. The molecular and cellular biology of chemokines.J ViralHepat. 1998;5:1–14.
30. Yoshimura T, Leonard EJ. Human monocyte chemoattractant protein-1:structure and function.Cytokines. 1992;4:131–152.
31. Carr MW, Roth SJ, Luther E, et al. Monocyte chemoattractant protein 1acts as a T-lymphocyte chemoattractant.Proc Natl Acad Sci U S A.1994;91:3652–3655.
32. Randolph GJ, Furie MB. A soluble gradient of endogenous monocytechemoattractant protein-1 promotes the transendothelial migration ofmonocytes in vitro.J Immunol. 1995;155:3610–3618.
Poddar et al Homocysteine and Chemokine Expression 2723
by guest on November 24, 2015http://circ.ahajournals.org/Downloaded from

JacobsenRanjana Poddar, Natarajan Sivasubramanian, Patricia M. DiBello, Killian Robinson and Donald W.
Interleukin-8 in Human Aortic Endothelial Cells: Implications for Vascular DiseaseHomocysteine Induces Expression and Secretion of Monocyte Chemoattractant Protein-1 and
Print ISSN: 0009-7322. Online ISSN: 1524-4539 Copyright © 2001 American Heart Association, Inc. All rights reserved.
is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation doi: 10.1161/01.CIR.103.22.2717
2001;103:2717-2723Circulation.
http://circ.ahajournals.org/content/103/22/2717Wide Web at:
The online version of this article, along with updated information and services, is located on the World
http://circ.ahajournals.org//subscriptions/
is online at: Circulation Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. and Rights Question and Answer
Permissionsthe middle column of the Web page under Services. Further information about this process is available in thethe online version of the published article for which permission is being requested is located, click Request Permissions in
can be obtained via RightsLink, a service of the Copyright Clearance Center, not the Editorial Office. OnceCirculation Requests for permissions to reproduce figures, tables, or portions of articles originally published inPermissions:
by guest on November 24, 2015http://circ.ahajournals.org/Downloaded from




















