
Human platelets synthesize and express functional tissue factor

1993 82: 513-520
J Cermak, NS Key, RR Bach, J Balla, HS Jacob and GM Vercellotti synthesize tissue factorC-reactive protein induces human peripheral blood monocytes to
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
reserved.Copyright 2011 by The American Society of Hematology; all rightsWashington DC 20036.by the American Society of Hematology, 2021 L St, NW, Suite 900, Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly
For personal use only. by guest on July 13, 2011. bloodjournal.hematologylibrary.orgFrom

C-Reactive Protein Induces Human Peripheral Blood Monocytes to Synthesize Tissue Factor
By Jaroslav Cermak, Nigel S . Key, Ronald R. Bach, Jozsef Balla, Harry S . Jacob, and Gregory M. Vercellotti
The acute inflammatory response is frequently accompa- nied by serious thrombotic events. We show that C-reac- tive protein (CRP), an acute-phase reactant that markedly increases its serum concentration in response to inflamma- tory stimuli, induced monocytes to express tissue factor (TF), a potent procoagulant. Purified human CRP in con- centrations commonly achieved in vivo during inflamma- tion (10 to 100 pg/mL) induced a 75-fold increase in TF procoagulant activity (PCA) of human peripheral blood mononuclear cells (PBM), with a parallel increase in TF an- tigen levels. CRP-induced PCA was completely blocked by a monoclonal antibody against human TF but not by irrele- vant murine lgG. Dot blot analysis showed a significant increase of TF mRNA after 4 hours of incubation with CRP. followed by a peak of PCA within 6 and 8 hours. Actinomy- cin D and cycloheximide blocked CRP-stimulated PCA.
HE INFLAMMATORY response includes phagocyte T margination and migration, chemoattractant genera- tion, cytokine formation, and production of a heterogenous group of proteins collectively called acute-phase reactants. The most characteristic human acute-phase protein is C- reactive protein (CRP).’ CRP, consisting of five identical, noncovalently linked subunits of 23,000 molecular weight,2 is synthesized in the liver. Its serum concentration of less than 1 jtg/mL can increase during the first 24 to 48 hours of inflammation or tissue necrosis by several hundred-fold, a response shown primarily to be due to interleukin-6 (IL-6) ti mu la ti on.^.^ CRP is thought to amplify the host defense system by potentiating its recognition capacities, as exem- plified by its activation of the complement cascade and its stimulation of various effector phagocytic cell^.^-^
The linkage between inflammation and coagulation may well involve tissue factor (TF). TF is a membrane-bound glycoprotein that initiates the extrinsic pathway of coagula- tion.’ The TF-factor VIIa (TF-FVIIa) complex activates FX directly or indirectly via FIX activation, leading ultimately to thrombin generation.’ “Resting” endothelial cells and monocytes/macrophages express very low levels of procoag- ulant activity (PCA). However, in both these cell types, an induction of TF expression with associated increased PCA has been shown with exposure to various agents, including inflammatory mediators such as endotoxin (LPS) or cyto- kine^."*^
That CRP might also play a role in upregulating mono- cyte PCA, as suggested by Whisler et al,23 is an attractive possibility, because CRP is known to modulate other mono- cyte function^^^-^^ through a specific re~eptor.*’~~’ Because CRP accumulates at sites of inflammation or tissue damage, we questioned whether CRP might promote localized coagu- lation by stimulating monocytes to produce the specific pro- coagulant, TF. In the following studies, we used highly pun- fied human CRP to assay its effects on peripheral blood monocyte and endothelial cell PCA and to discern mecha- nisms by which CRP promotes TF expression in these cells.
suggesting that de novo TF protein synthesis was re- quired. Endotoxin (LPS) contamination of CRP was ex- cluded as the mediator of TF synthesis because: (1) CRP was Limulus assay negative; (2) induction of TF PCA by CRP was not blocked by Polymyxin B, in contrast to LPS- induced PCA; (3) antihuman CRP IgG inhibited CRP-in- duced PCA, but not LPS-induced PCA; (4) CRP was able to stimulate TF production in LPS-pretreated PBM refractory to additional LPS stimulation; and, (5) unlike LPS. CRP was incapable of inducing TF in human umbilical vein endothe- lial cells. We suggest that CRP-mediated TF production in monocytes may contribute to the development of dissemi- nated intravascular coagulation and thrombosis in inflam- matory states. 0 1993 by The American Society of Hematology.
MATERIALS AND METHODS
Reagents Highly purified (>90%) human CRP was obtained from Sigma
CO (St Louis, MO) (catalogue no. C6 177), as well as endotoxin/lipo- polysaccharide (LPS) (catalogue no. L3 129), lyophilized IgG frac- tion of rabbit serum against human CRP (catalogue no. C3257), and TRIS [tris(hydroxymethyl)aminomethane].
Hanks’ Balanced Salt Solution (HBSS), HEPES buffer solution, and all cell culture media including RPMI 1640 were purchased from GIBCO (Grand Island, NY). Heparin sodium was from Ly- phoMed (Melrose Park, IL); Polymyxin Band recombinant human IL-lp (rIL- 1) were from Upjohn CO (Kalamazoo, MI); CS-depleted serum was from Quidel CO (San Diego, CA); formaldehyde was from Fisher Scientific (Pittsburgh, PA); and cycloheximide was from NBCo (Cleveland, OH). Radiolabeled compounds and nylon membranes were obtained from Amersham (Arlington Heights,
Murine monoclonal antibody (MoAb) against human TF (HTF- 1 ), polyclonal antibody against human TF (anti-HTF pAb), human
IL).
From the Department ofMedicine, University ofMinnesota, Min- neapolis; and the Veterans Administration Medical Center, Minne- apolis, MN.
Submitted November 30, 1992; accepted March 17, 1993. Supported in part by grants f o m the National Institutes of Health
(HL 33793), the Graduate School of the University of Minnesota, and the Department of Veterans’ Affairs. J.C. was supported by the Charles Proshek Fellowship from the Minnesota Medical Founda- tion.
Address reprint requests to Gregory M. Vercellotti, MD, Univer- sity of Minnesota, Department of Medicine/Division of Hematol- ogy, Box 480 UMHC, Harvard St at E River Rd, Minneapolis, MN 55455.
The publication costs of this article were defayed in part by page charge payment. This article must therefore he hereby marked “advertisement” in accordance with I8 U.S.C. section I734 solely to indicate this fact.
0 I993 by The American Society oJHematology. 0006-4971/93/8202-0004$’3.00/0
Blood, VOI 82, NO 2 (July 15). 1993: pp 51 3-520 513
For personal use only. by guest on July 13, 2011. bloodjournal.hematologylibrary.orgFrom

514 CERMAK ET AL
FVIIa and FX, human TF standard, and bovine brain cephalin were prepared as previously de~cribed?~-~’ All other reagents were obtained from Sigma CO, unless otherwise specified.
All reagents, including CRP, were tested for endotoxin contami- nation by the Limulus assay (E-Toxate; Sigma CO) and were nega- tive at levels from 0. I to 0.2 EU/mL (10 to 20 pg/mL).
Peripheral Blood Mononuclear Cells (PBM) and Peripheral Blood Monocyte Isolation
Minimally heparinized (2 U/mL) blood from fasting healthy do- nors (after obtaining their informed consent according to the guide- lines of the Committee on the Use of Human Subjects in Research, University of Minnesota) was layered over Histopaque- 1077 and centrifuged (500g) at 24°C for 30 minutes. The PBM collected from the plasma/Histopaque interface were washed three times with phosphate-buffered saline (PBS) and finally resuspended in RPMI 1640 + 1% fetal calf serum (FCS) at a concentration of 1 X IO6 cells/mL. The PBM contained an average of 80% lymphocytes and 20% monocytes as assessed by light microscopy and nonspecific esterase staining. Contamination by polymorphonuclear leuko- cytes and platelets was less than 1%.
PBM at I X IO6 cells/mL in RPMI 1640 + 1% FCS were incu- bated with various reagents at 37°C in 5% CO, for the appropriate times. After incubation, cells were washed once with fibrometry buffer (0.16 mol/L NaCl + 25 mmol/L HEPES buffer, pH 7.4). The viability ofthe cells as assayed by Trypan blue exclusion was greater than 95% after 6 hours of incubation and 92% after 18 hours of incubation.
To prepare a relatively enriched population of peripheral blood monocytes, PBM at 2 X IO6 cells/mL were plated onto plastic tissue culture wells with RPMI 1640 + 1% FCS. After 2 hours of incuba- tion, nonadherent cells were removed by washing five times with medium. The adherent cells (approximately 75% to 80% mono- cytes and 20% to 25% lymphocytes) were incubated with various reagents under the same conditions as PBM.
Human Umbilical Vein Endothelial Cell (HUVEC) Cultures
HUVEC were isolated and grown as previously described3’ and used from passages 1 through 3 after reaching confluence. HUVEC grown in 2-cm2 tissue culture wells (approximately 200,000 cells/ well) were incubated with various reagents in the same manner as PBM; after the incubation, the cells were washed once with fibrome- try buffer.
TF Assays One-stage clotting assay. PBM cell pellet was resuspended in
0.5 mL fibrometry buffer, frozen at -7O”C, and thawed at 37°C three times and sonicated. Peripheral blood monocytes and HU- VEC were treated the same way. Cells were scraped from the bot- tom of tissue culture wells, frozen, and thawed three times before sonication.
The clotting assay” consisted of 0.1 mL of cell sample and 0. I mL of citrated normal human platelet-poor plasma. The reaction was initiated in a fibrometer cup (Fisher Scientific CO) by the addi- tion of 0.1 mL 25 mmol/L CaCI,. Results were expressed in arbi- trary units per milliliter (U/mL) of T F by comparison of clotting times with a standard curve obtained using a purified human TF standard as previously de~cribed.’~ One unit of TF PCA equals 1 pg of the TF standard. For HUVEC, the results were expressed in U/mg of cell protein assessed by BCA protein assay (Pierce, Rock- ford, IL).
PCA was measured on Coag-a-mate Two-stage clotting assay.
XM (Organon Teknika, Durham, NC). In stage 1 of the assay, 20 pL of sample was incubated with 10 pL of 30 nmol/L human FVIIa and 10 pL of I .5 pmol/L human FX at 37°C for at least 3 minutes. The reaction was initiated by adding 20 pL of 25 mmol/L CaCI,, and the sample was incubated for another 5 minutes. Stage 2 ofthe assay was started by the simultaneous addition of 100 pL substrate plasma (90 pL normal citrated bovine plasma plus 10 pL bovine brain cephalin [6 mg/mL] in TRIS-buffered saline) and 100 pL 25 mmol/L CaCI,. Clotting time was recorded, and TFlevel was calcu- lated from a standard curve obtained using a human brain TF stan- dard.34
T F antigen ELISA was performed as previously described.34 Briefly, 96-well microtiter plates were coated with 100 pL per well of IO pg/mL HTF-I murine MoAb in a bicarbonate buffer (15 mmol/L Na’CO,, 35 mmol/L NaHCO,, pH 9.6) and incubated overnight at 4°C. Wells were then filled with TBS (0.1 mol/L NaC1,0.05 mol/L Tris, pH 7.5) containing 1% bovine serum albumin (BSA) to block nonspecific sites and incubated again at 4°C for at least 4 hours. One hundred microliters of sample for assay or TF standard diluted in TBS containing 0.1% BSA, 0.1% Triton X-100, and 5 mmol/L EDTA were added to triplicate wells. After an overnight incuba- tion, the plates were extensively washed, 100 pL of anti-HTF poly- clonal antibody at a final concentration of 0.2 pg/mL diluted in TBS + I % BSA was added, and samples were incubated for 2 hours at room temperature. After another wash, the samples were incu- bated with 100 pg goat antirabbit horseradish peroxidase-conju- gated IgG (GAR-HRP; Bio-Rad, Richmond, CA) diluted 1 :2,000 in TBS + 1% BSA. After a further wash cycle, a chromogenic detec- tion system was added for 30 minutes. The reaction was stopped with the addition of 25% H2S04, and the sample absorbance was read at 490 nm on a Dynatech MR 600 microplate reader (Dyna- tech Laboratories Inc, Chantilly, VA). TF antigen level was calcu- lated from a standard curve obtained with diluted human T F stan- dard in the range of 0 to 1,000 pg/mL.
TF mRNA analysis. T F mRNA content in PBM was analyzed after treatment with CRP, LPS, serum, or RPMI 1640 + 1% FCS alone. PBM cellular mRNA was isolated by the RNAzol method (Tel-TEST, Inc, Friendswood, TX). Aliquots (4 pg) of total RNA were dissolved in IO pL of H,O and mixed with 20 pL of 100% formamide, 7 pL of 37% formaldehyde, and 2 pL of 20X SSC (3 mol/L NaCl + 0.3 mol/L sodium citrate), applied to slots on a microsample filtration manifold (Minifold I; Schleicher and Schuell, Inc, Keene, NH), and transferred onto nitrocellulose mem- b r a n e ~ ~ ~ using UV cross-linking. The membranes were hybridized at 42°C with a nick-translated ”P-labeled cDNA probe for human TF.36 Autoradiographs were obtained and quantitated by com- puter-assisted videodensitometry.”
TF antigen enzyme-linked immunosorbent assay (ELISA).
Statistical Methods
results are expressed as mean l?r I SD. Statistical analysis was performed using the Student’s t-test. All
RESULTS
Induction of TF Expression on PBM and on Peripheral Blood Monocytes
A significant increase in TF PCA of disrupted PBM was found after 6 hours of incubation with either CRP (100 pg/mL) or LPS (10 pg/mL) (P < .0001 for one-stage clotting assay and P < .O 1 for two-stage clotting assay and TF anti- gen [Table 11). A difference between CRP- and LPS-in- duced TF expression was also significant (P < .O 1 for all 3
For personal use only. by guest on July 13, 2011. bloodjournal.hematologylibrary.orgFrom

CRP INDUCES TISSUE FACTOR
1000 -
800 - 600 -
400 -
200 -
O +
515
I I
- - E
Ns z 3 600-
U. n I-
RPMl U P CRPl LPS LPSl
Table 1. Induction of TF in PBM by Different Stimuli
One-Stage Two-Stage TF Antigen Clotting Assay Clotting Assay ELSA
Reagent ( U / W ( U l W (pg/mL)
Control 13.1 k2.4 8.5 k 0.8 20.3 k 3.5 LPS (10 pg/mL) 415.0 t 46.4 299.6 f 13.0 373.3 t 14.2 CRP (100~~n/mL) 1,044.8 rt 52.8 694.5 f 28.9 726.1 t 17.5
~ _ _ _ ~ ~
PBM at a concentration of 106/mL were incubated for 6 hours with RPMl 1640 + 1 % FCS (control), LPS (10 pg/mL), or CRP (1 00 pg/mL), both in RPMl 1640 + 1 % FCS. The TF PCA was measured by one- or two-stage clotting assay and TF antigen by ELISA. Results represent the mean f 1 SD from five experiments performed in duplicates
assays). In the two-stage clotting assay, PCA was undetect- able when FVIIa was omitted from the assay (data not shown). This result shows that the PCA is indeed TF-me- diated. Most of the TF activity induced by CRP in mono- nuclear cells was encrypted, ie, CRP-treated (100 pg/mL) mononuclear cells added intact to the one-stage clotting as- say had 17% of the procoagulant activity (150 U/mL) com- pared with sonicated, disrupted CRP-treated mononuclear cells (880 U/mL).'
Similarly, under the same conditions, a relatively enriched population of peripheral blood monocytes exhib- ited a significant increase of TF PCA from a control value of 4.2 f 1.1 U/mL to 714.9 k 70.7 U/mL with LPS ( I O pg/ mL) and 1,495.8 f 267.9 U/mL with CRP (100 pg/mL) (measured by the one-stage clotting assay).
To characterize CRP-induced PCA on PBM, the samples previously treated with CRP for 6 hours were incubated with a murine blocking MoAb (HTF-1) for I hour at room temperature. This incubation decreased TF activity in the one-stage clotting assay from 1,044.8 U/mL to 48.8 U/mL (P < .OOOl) (Fig I). Incubation with an irrelevant murine
- E . 2. U. I-
p C 0.0001
1200 1 -
IgG in the same concentration did not significantly affect CRP-induced PCA. This further substantiates that the PCA induced by CRP is TF.
To obtain evidence that CRP is the factor inducing TF PCA, CRP (100 pg/mL) was pretreated with an IgG fraction of rabbit serum against human CRP (200 pg/mL) for 30 minutes at 37°C. Pretreatment of CRP significantly de- creased CRP-induced TF expression by PBM after 6 hours of incubation (P < .OOOl). In contrast, antiserum against human CRP had no effect on LPS induced TF expression (Fig 2).
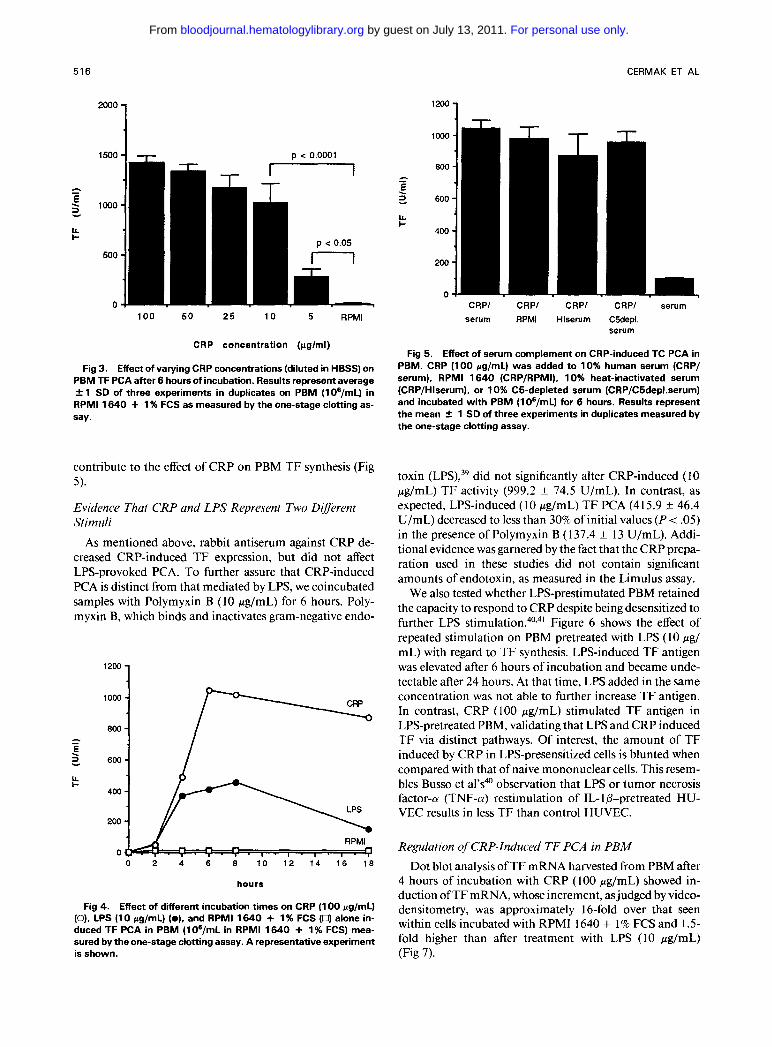
Under normal circumstances, the serum CRP concentra- tion is less than 1 pg/mL, but can increase up to 1,000-fold during acute infections or tissue n e c r o s i ~ . ' ~ , ~ ~ Therefore, we examined a dose response of PBM exposed to varying CRP concentrations for 6 hours; as shown in Fig 3, a concentra- tion of CRP as low as 5 pg/mL significantly induces TF PCA (P < .05), with a further increase in response up to 100 pg/mL. Levels of CRP 5 1 pg/mL did not induce TF PCA (data not shown).
The time course of TF PCA induced in PBM by CRP differed from that noted with LPS induction. As seen in Fig 4, a peak of CRP-stimulated PCA occurred between 6 and 8 hours and remained elevated even after 18 hours. In con- trast, the LPS effect peaked between 4 and 8 hours and tailed off at 18 hours.
Because CRP is able to induce complement activa- t i ~ n , ' * ~ , ~ we tested whether CRP-induced TF PCA in serum was dependent on complement. Using RPMI, C5-depleted human serum, or heat-inactivated human serum (56°C for 30 minutes) as the diluent, we observed only a slight de- crease in the induction of TF PCA by CRP. This result suggests that complement activation does not substantially
p c 0.0001
1200 1 n I 7
RPMl CRP CRP/HTF-1 CRP/IgG CRP Ab CRP Ab
Fig 1. Effect of a blocking murine MoAb (HTF-1) on CRP-in- duced PCA in PBM. After 6 hours of incubation with CRP (100 pg/mL) in RPMll640 + 1 % FCS or RPMll640 + 1 % FCS alone, PBM (1 06/mL) cells were incubated for 1 hour at room temperature either with HTF-1 (200 pg/mL) (CRP/HTF-1) or with irrelevant mu- rine IgG (200 pg/mL) (CRP/IgG). Results represent the mean 2 1 SD of three experiments in duplicates measured by the one-stage clotting assay.
Fig 2. Effect of antihuman CRP antiserum on CRP- and LPS-in- duced TF PCA in PBM. PBM (106/mL in RPMl 1640 + 1% FCS) were incubated for 6 hours either with CRP (1 00 pg/mL) or LPS (1 0 pg/mL), respectively, with CRP or LPS in the same concentration preincubated with 200 pg/mLof purified rabbit antihuman CRP lgG for 30 minutes at 37°C ([CRP/CRPAb], [LPS/CRPAb]). Results rep- resent the mean * 1 SD of three experiments in duplicates mea- sured by the one-stage clotting assay.
For personal use only. by guest on July 13, 2011. bloodjournal.hematologylibrary.orgFrom
516 CERMAK ET AL
1500 I T- D < 0.0001
100 50 25 10 5 RPMl
CRP concentration (pglml)
Fig 3. Effect of varying CRP concentrations (diluted in HBSS) on PBM TF PCA after 6 hours of incubation. Results represent average 2 1 SD of three experiments in duplicates on PBM (lOg/mL) in RPMl 1640 + 1 % FCS as measured by the one-stage clotting as- say.
contribute to the effect of CRP on PBM TF synthesis (Fig 5).
Evidence That CRP and LPS Represent Two Different Stimuli
As mentioned above, rabbit antiserum against CRP de- creased CRP-induced TF expression, but did not affect LPS-provoked PCA. To further assure that CRP-induced PCA is distinct from that mediated by LPS, we coincubated samples with Polymyxin B (10 pg/mL) for 6 hours. Poly- myxin B, which binds and inactivates gram-negative endo-
1200 1
600 i I I& I-
0 2 4 6 8 1.0 1.2 1.4 1 6 1.8
hours
Effect of different incubation times on CRP (1 00 pg/mL) (0). LPS (1 0 pg/mL) (01, and RPMl 1640 + 1 % FCS (0) alone in- duced TF PCA in PBM (10*/mL in RPMl 1640 + 1% FCS) mea- sured by the one-stage clotting assay. A representative experiment is shown.
Fig 4.
CRPI CRPl CRPl CRPI serum serum RPMl Hlserum CWepl.
serum
Fig 5. Effect of serum complement on CRP-induced TC PCA in PBM. CRP (100 pg/mL) was added to 10% human serum (CRP/ serum), RPMl 1640 (CRP/RPMI), 10% heat-inactivated serum (CRP/Hlserum), or 10% C5-depleted serum (CRP/C5depl,serum) and incubated with PBM (1OB/mL) for 6 hours. Results represent the mean f 1 SD of three experiments in duplicates measured by the one-stage clotting assay.
toxin (LPS),39 did not significantly alter CRP-induced (10 pg/mL) TF activity (999.2 f 74.5 U/mL). In contrast, as expected, LPS-induced (10 pg/mL) TF PCA (415.9 f 46.4 U/mL) decreased to less than 30% of initial values (P < .05) in the presence of Polymyxin B ( 1 37.4 f 13 U/mL). Addi- tional evidence was garnered by the fact that the CRP prepa- ration used in these studies did not contain significant amounts of endotoxin, as measured in the Limulus assay.
We also tested whether LPS-prestimulated PBM retained the capacity to respond to CRP despite being desensitized to further LPS s t i m u l a t i ~ n . ~ ~ , ~ ' Figure 6 shows the effect of repeated stimulation on PBM pretreated with LPS ( I O pg/ mL) with regard to TF synthesis. LPS-induced TF antigen was elevated after 6 hours of incubation and became unde- tectable after 24 hours. At that time, LPS added in the same concentration was not able to further increase TF antigen. In contrast, CRP (100 pg/mL) stimulated T F antigen in LPS-pretreated PBM, validating that LPS and CRP induced TF via distinct pathways. Of interest, the amount of TF induced by CRP in LPS-presensitized cells is blunted when compared with that of naive mononuclear cells. This resem- bles Busso et al's4 observation that LPS or tumor necrosis factor-a (TNF-a) restimulation of IL- 1 (3-pretreated HU- VEC results in less TF than control HUVEC.
Regulation of CRP-Induced TF PCA in PBM Dot blot analysis of TF mRNA harvested from PBM after
4 hours of incubation with CRP (100 pg/mL) showed in- duction ofTF mRNA, whose increment, as judged by video- densitometry, was approximately 16-fold over that seen within cells incubated with RPMI 1640 + 1 % FCS and 1.5- fold higher than after treatment with LPS (10 pg/mL) (Fig 7).
For personal use only. by guest on July 13, 2011. bloodjournal.hematologylibrary.orgFrom

CRP INDUCES TISSUE FACTOR 517
.y . , . , . , v, ,LPS
0 6 12 18 2 4 30
Tlmo (hra) 4 CRP 100pg1ml
LPS lOpg/ml
4 TREATMENT: LPS
10pg/ml or
Fig 6. Effect of repeated stimulation of PBM. PBM (1 06/mL in RPMl 1640 + 1 % FCS) were incubated for 24 hours with LPS (1 0 pg/mL). TF antigen expression was measured by ELISA at 0,6, and 24 hours. After 24 hours, the cells were washed, resuspended in RPMl 1640 2 1 % FCS, CRP (1 00 rg/mL) or LPS (1 0 rg/mL) was added, and, after 6 hours of additional incubation, TF antigen was recorded. Results of a representative experiment are shown. Simi- lar results were obtained for these groups when TF PCA was mea- sured by the one-stage clotting assay (data not shown).
To examine the regulatory mechanisms, we also coincu- bated PBM with cycloheximide and actinomycin D. Both cycloheximide and actinomycin D, at concentrations of I O pg/mL, completely blocked CRP (100 pg/mL) induction of T F PCA (one-stage clotting assay) in PBM ( 1 3.4 f 3.2 and 12.2 f 4.0 U/mL, respectively) compared with CRP alone (1,044.8 f 52.8 U/mL), suggesting that de novo protein synthesis was necessary for T F production.
Induction Because both monocytes/macrophages and endothelial
cells are capable of synthesizing T F in response to inflamma- tory mediators such as LPS or cytokines, we tested the abil- ity of CRP to induce TF PCA in endothelial cells. Both LPS ( 1 0 pg/mL) and rIL- 1 p (0. I pg/mL) induced TF expression on HUVEC after 6 hours of incubation (P e .01 when com- pared with RPMI 1640 + 1% FCS). In contrast, CRP (100 pg/mL) failed to increase HUVEC TF expression (Fig 8).
TF Expression in HU VEC
Fig 7. Dot blot analysis of TF mRNA. PBM (in RPMll640 + 1% FCS) were incubated for 4 hours with CRP (100 rg/mL), LPS (10 pg/mL), 10% human serum, or RPMl 1640 + 1% FCS alone. mRNA was isolated from 6 X 10' cells per each group. Intensity of hybridization RE AT1 U E
DISCUSSION
In the genesis of thrombosis, Virchow first stressed the importance of blood flow, the blood's propensity to clot. and the integrity ofthe vessel wall, the last ofwhich could be affected by inflammation.
In the course of inflammation, TF, an initiator of the extrinsic coagulation pathway. may be expressed on cell sur- faces that are not thrombogenic under normal circum- stances, such as monocytes and endothelial cells. Various inflammatory stimuli (eg, LPS. TNF, and IL-1) have previ- ously been shown to increase TF expression on endothelial cell^,'^-^^ whereas in human monocytes/macrophages. en- dotoxin?" activated complement." aggregated IgG,13 and TNFI4 all have been reported to generate procoagulant activ- ity, presumably by inducing T F neosynthesis.' Moreover, an increase in circulating monocyte PCA in various inflam- matory diseases (including Crohn's disease, meningococcal sepsis, lupus erythrematosus, and rheumatic diseases) has been previously Rivers et a146 showed an in- crease in the expression of TF in monocytes from infants with severe infection. We have recently obtained sera con- taining high levels of CRP ( 1 1.6 mg/dL) from a patient with fevers, idiopathic retroperitoneal fibrosis, and deep venous thrombosis that induced T F in normal monocytes. How- ever, our anti-CRP antibody failed to inhibit TF induction by this patient's sera. Defining a singular effect of CRP on mononuclear cell tissue factor induction in sick patients' sera may prove difficult because of concurrent serum eleva- tions of IL-I, TNF, immune complexes, or other factors that themselves can activate TF synthesis. We are planning further TF studies with sera from ill patients who have high serum CRP levels to further delineate CRP's role.
CRP was first reported by Tillet and Francis4' as a serum factor possessing the capacity to precipitate a carbohydrate derived from pneumococcal fraction C. CRP not only binds this pneumococcal polysaccharide but also binds calcium. phosphate monoesters. phosphoryl choline, galactose poly- mers. and p o l y c a t i o n ~ ~ * ~ CRP is also able to bind polymor- phonuclear leukocytes and to modulate neutrophil func- t i o n ~ ~ ~ . ~ ~ and has been found to affect intracellular calcium mobilization, superoxide production, and tumoricidal activ- ity of peripheral monocytes/macrophages2e26 via binding to a specific surface receptor.28 Ballou et aI5' recently showed that CRP can induce TNF-(U, IL-I@. and IL-6 in monocytes
signals was measured by video-
the relative increase in compari- son with RPMl 1640-treated PBM.
densitometry and expressed as I NTE N S ITV- 16x 10 x l x 1 . 5 ~
CRP LPS RPMI Serum
For personal use only. by guest on July 13, 2011. bloodjournal.hematologylibrary.orgFrom

518 CERMAK ET AL
- C .- d 200-
n e p < 0.001 - U
RPMl LPS r lL-1
Fig 8. Effect of various stimuli on TF PCA in HUVEC. Confluent monolayers of HUVEC grown in 2-cmZ wells were incubated for 6 hours with RPMl 1640 + 1 % FCS, CRP (100 pg/mL), LPS (IO pg/ mL), or rlL-I (0.1 pg/mL) (all reagents diluted in RPMl 1640 f 1 % FCS). The results represent the mean 2 1 SD from three experi- ments performed in duplicate and measured by the one-stage clot- ting assay.
in a similar dose and time course as we have shown for TF. Whether these cytokines play a role in the signal transduc- tion for TF synthesis by CRP is now under investigation in our laboratory.
Although induction of monocyte TF PCA in the course of inflammation has previously been ascribed to agents such as LPS, cytokines, or activated complement, the current study shows that CRP may be equally important. The maximal induced increment of TF PCA in PBM was greater than 75-fold over that of the control as measured by one- and two-stage assay and corresponded with a parallel increase in TF antigen. Likewise, Whisler et alz3 previously described increased PCA in PBM after coincubation of LPS or im- mune complexes with CRP and noted that CRP alone also induced some PCA. Our experiments confirm these find- ings and show that the procoagulant activity reflects produc- tion of authentic TF because: ( I ) the activity is inhibited by a blocking MoAb to T F and (2) the procoagulant activity required FVIIa. However, we emphasize that because CRP may increase up to 1,000-fold within 6 to 24 hours in re- sponse to infections or tissue destruction, CRP concentra- tions of 100 fig/mL, used in our experiments, are com- monly achieved during the course of an infectious episode. 1,3,38
Monocytes are considered to be the only circulating leu- kocyte capable of TF synthesis’’; however, CD4+ T-helper lymphocytes may enhance the monocyte TF response to stimuli such as LPS by both direct cell-cell contact as well as via the production of stimulatory lymphokine^.'^,^^ Re- cently, a specific receptor for CRP has been described on monocytes but not on peripheral blood lymphocyte^.'^ We did not observe any significant differences in the results of experiments performed on freshly isolated peripheral blood mononuclear cells (containing approximately 20% to 25% monocytes) or relatively purified (70% to 80%) peripheral
blood monocytes. However, our data do not exclude the possibility that T lymphocytes are critical in mediating the monocyte TF response to CRP.
Although we showed a threefold increase of serum C5a concentration when serum was incubated with CRP (100 pg/mL) in a recent s t ~ d y , ’ ~ CRP-induced monocyte TF ex- pression does not only require complement activation (Fig 5); however, our results do not exclude the possibility that activated complement components may add to the procoag- ulant effect of CRP.
Tebo and Mortensen” described a specific CRP receptor distinct from the IgG receptor on human blood monocytes and the human monocytic cell line U937. Excess of phos- phorylcholine was unable to inhibit CRP binding to mono- cytes; similarly, in our experiments, even a 100-fold molar excess of phosphorylcholine failed to inhibit CRP-induced monocyte PCA (data not shown). The same investigators also recently showed2’ that the CRP/CRP receptor complex was internalized into an endosomal compartment in which CRP was liberated and subsequently degraded. When U937 cells were exposed to CRP, a significant H202 production and tumoricidal activity were detected after 8 hours.
We found that PBM TF mRNA significantly increases after 4 hours of incubation with CRP, which was followed by a peak of TF PCA at 6 to 8 hours. Such findings are comparable to those published by Gregory et a1” who showed induction of TF expression on isolated monocytes exposed to LPS within 4 hours and peak PCA at 6 hours. Similarly, in our studies, coincubation of CRP with both cycloheximide and actinomycin D completely block CRP- induced TF PCA, suggesting that de novo synthesis of TF protein is involved and that CRP induces monocyte TF expression, presumably by acting at the transcriptional level.
We rigorously excluded LPS contamination as a possible artifact in these studies by showing ( 1 ) that all reagents (in- cluding CRP) were negative for significant levels of contami- nating endotoxin as judged by the Limulus amoebocyte as- say; (2) that Polymyxin B inhibited LPS-induced PCA and had no effect on CRP-mediated PCA; (3) that a neutralizing antibody against CRP had a selective blocking effect on CRP-induced PCA; (4) that PBM pretreated with LPS were able to increase TF in response to CRP despite desensitiza- tion to further LPS stimulation; and (5) that in cultured HUVEC, we observed a failure to respond to CRP but not LPS. Thus, we conclude from these data that CRP and LPS represent two distinct stimuli.
It is evident that the vascular endothelium plays an im- portant role in hemostasis and thrombosis, and that inflam- matory agent-stimulated endothelium may affect the acti- vation of the coagulation system. However, we were not able to show a significant increase of HUVEC TF expres- sion after exposure to CRP, in contrast to other inflamma- tory mediators, including LPS, IL-1, and TNF, which in- duce PCA in both monocyte/macrophages and endothelial cells.9. IO. l4,20-22 w e speculate that this intriguing difference might be explained by the absence or considerably reduced expression of the CRP receptor on cultured endothelial cells.
For personal use only. by guest on July 13, 2011. bloodjournal.hematologylibrary.orgFrom

CRP INDUCES TISSUE FACTOR 519
Our results suggest that increased monocyte TF expres- sion during infection or tissue necrosis may be at least par- tially mediated by an increased CRP level. Thus, CRP-me- diated monocyte PCA induction may play an important role in altered microcirculation in inflammatory and ne- crotic tissue as well as contribute to the development of disseminated intravascular coagulation in septic infections. The ability of CRP to reinduce PCA in LPS-stimulated monocytes in a phase in which they are refractory to further LPS stimulation may be an important tool for the mainte- nance of a high monocyte PCA during inflammation. We speculate that this enhanced TF production may better al- low inflammatory cells to ward off bacteria by promoting fibrin formation locally.
ACKNOWLEDGMENT
We thank Dr Henry Gewurz (Rush Presbyterian St Luke’s Medi- cal Center, Chicago, IL) for kind advice; Dong Tuong and Theresa Stella for technical assistance; and Linda Radtke for manuscript preparation.
REFERENCES 1. Gewurz H: Biology of C-reactive protein and the acute phase
response. Hosp Pract 6:67, 1982 2. Osmand AP, Friedenson B, Gewurz H, Painter RH, Hof-
mann T, Shelton E: Characterization of C-reactive protein and the complement subcomponent C 1 t as homologous proteins display- ing cyclic pentameric symmetry (pentraxins). Proc Natl Acad Sci USA 74:739, 1977
3. Pepys MB: C-reactive protein fifty years on. Lancet 1:653, 1981
4. Kilkpatrick JM, Volanakis JE: Molecular genetics, structure and function of C-reactive protein. Immunol Res 10:43, 1991
5 . Kaplan MH, Volanakis JE: Interaction of C-reactive protein complexes with the complement system. I. Consumption of human complement associated with the reaction of C-reactive protein with pneumococcal C-polysaccharide and with the choline phospha- tides, lecithin and sphingomyelin. J Immunol 112:2135, 1974
6. Volanakis JE, Kaplan MH: Interaction of C-reactive protein complexes with the complement system. 11. Consumption of Guinea pig complement by CRP complexes: Requirement for hu- man Clq. J Immunol 113:9, 1974
7. Bach R R Initiation of coagulation by tissue factor. Crit Rev Biochem Mol Biol23:339, 1988
8. Osterud B, Rapaport SI: Activation of factor IX by the reac- tion product of tissue factor and factor VII: Additional pathway for initiating blood coagulation. Proc Natl Acad Sci USA 795260, 1977
9. Rivers RPA, Hathaway WE, Weston WL: The endotoxin-in- duced coagulant activity of human monocytes. Br J Haematol 20:311, 1975
10. Prydz H, Allison AC: Tissue thromboplastin activity of iso- lated human monocytes. Thromb Haemost 39582, 1978
1 1. Muhlfender TW, Niemetz J, Kreutzer D, Beebe D, Ward P, Rosenfeld SI: C5 chemotactic fragment induced leukocyte produc- tion of tissue factor activity. J Clin Invest 63:147, 1979
12. Prydz H, Lyberg T, Deteix P, Allison AC In vitro stimula- tion of tissue thromboplastin (factor 111) activity in human mono- cytes by immune complexes and lectins. Thromb Res 15:465, 1979
13. Rothenberger H, Zimmerman TS, Spielberg HL, Vaughan JH: Leukocyte procoagulant activity. Enhancement of production in vitro by IgG and antigen-antibody complexes. J Clin Invest 59549, 1977
14. Conkling PR, Greenberg CS, Weinberg J B Tumor necrosis factor induces tissue factor-like activity in human leukemia cell line U937 and peripheral monocytes. Blood 72:128, 1988
15. Levy CA, Schwartz BS, Curtiss LK, Edgington TS: Plasma lipoprotein induction and suppression of the generation of cellular procoagulant activity in vitro. Requirements for cellular collabora- tion. J Clin Invest 67:1614, 1981
16. Schwartz BS, Levy CA, Curtiss LK, Edgington TS: Plasma lipoprotein induction and suppression of the generation of cellular procoagulant activity in vitro. Two procoagulant activities are pro- duced by peripheral blood mononuclear cells. J Clin Invest 67:1650, 1981
17. Lorenzet R, Niemetz J, Marcus AJ, Broekman MJ: Enhance- ment of mononuclear procoagulant activity by platelet 12-hy- droxyeicosatetraenoic acid. J Clin Invest 78:418, 1986
18. Rambaldi A, Alessio G, Casali B, Passerini CG, Donati MD, Mantovani A, Semeraro N: Induction of monocyte-macrophage procoagulant activity by transformed cell lines. J Immunol 136:3848, 1986
19. Carlsen E, Stinessen MB, Prydz H: Differential effect of al- pha-interferon on thromboplastin response in monocytes and endo- thelial cells. Clin Exp Immunol 70:47 1, 1987
20. Conway EM, Bach RR, Rosenberg RD, Konigsberg WH: Tumor necrosis factor enhances expression of tissue factor mRNA in endothelial cells. Thromb Res 53:231, 1989
2 I . Scarpati EM, Sadler JE: Regulation ofendothelial cell coagu- lant properties. Modulation of tissue factor, plasminogen activator inhibitors, and thrombomodulin by phorbol 12-myristate 13-ace- tate and tumor necrosis factor. J Biol Chem 26420705, 1989
22. Nawroth PP, Handley DA, Esmon CT, Stern DM: Interleu- kin 1 induces endothelial cell procoagulant while suppressing cell surface anticoagulant activity. Proc Natl Acad Sci USA 83:3460, 1986
23. Whisler RL, Proctor VK, Downs EC, Mortensen RF: Modu- lation of human monocyte chemotaxis and procoagulant activity by human C-reactive protein (CRP). Lymphokine Res 5:223, 1986
24. Zeller JM, Landay AL, Lint TF, Gewurz H: Enhancement of human peripheral blood monocyte respiratory burst activity by ag- gregated C-reactive protein. J Leukoc Biol40:769, 1986
25. Zahedi K, Mortensen R F Macrophage tumoricidal activity induced by human C-reactive protein. Cancer Res 465077, 1986
26. Foldes-Filep E, Filep JG, Sirois P: C-reactive protein inhibits intracellular calcium mobilization and superoxide production by guinea pig alveolar macrophage. J Leukoc Biol 5 I : 13, 1992
27. Tebo JM, Mortensen R F Characterization and isolation ofa C-reactive protein receptor from the human monocyte cell line U-937. J Immunol 144:231, 1990
28. Tebo JM, Mortensen RF: Internalization and degradation of receptor bound C-reactive protein by U-937 cells: Induction of H202 production and tumoricidal activity. Biochim Biophys Acta 1095:210, 1991
29. Carson SD, Ross SE, Bach RR, Guha A: An inhibitory mono- clonal antibody against human tissue factor. Blood 70490, 1987
30. Bach RR, Gentry G, Nemerson Y: Factor VI1 binding to tissue factor in reconstituted phospholipid vesicles: Induction of cooperativity by phosphatidylserine. Biochemistry 25:4007, 1986
3 I . Broze GJ, Majerus PW: Purification and characterization of human coagulation factor VII. J Biol Chem 250:1242, 1980
32. Miletich JP, Broze GJ, Majerus PW: Purification of human coagulation factors 11, IX, and X using sulfated dextran beads. Methods Enzymol80:22 1, 198 1
33. Jaffe EA, Nachman RL, Becker CG, Mimek LR: Culture of human endothelial cells derived from umbilical veins. J Clin Invest 52:2745, 1973
For personal use only. by guest on July 13, 2011. bloodjournal.hematologylibrary.orgFrom

520 CERMAK ET AL
34. Bloem LJ, Chen L, Konigsberg WH, Bach RR: Serum stimu- lation of quiescent human fibroblasts induces the synthesis of tissue factor mRNA followed by the appearance of tissue factor antigen and procoagulant activity. J Cell Physiol 139:4 18, 1989
35. Maniatis T, Fritsch EF, Sambrook J: Molecular Cloning: A Laboratory Manual. Cold Spring Harbor, NY, Cold Spring Harbor Laboratory, 1982
36. Spicer EK, Horton R, Bloem L, Bach R, Williams KR, Guha A, Kraus J, Lin T-C, Nemerson Y, Konigsberg WH: Isolation of cDNA clones coding for human tissue factor: Primary structure of the protein and cDNA. Proc Natl Acad Sci USA 84:5148, 1987
37. Correa-Rotter R, Mariash CN, Rosenberg ME: Loading and transfer control for Northern hybridization. Biotechniques 12: 154, 1992
38. Schultz DR, Arnold DR: Properties of four acute phase pro- teins: C-reactive protein, serum amyloid A protein, a,-acid glyco- protein and fibrinogen. Semin Arthritis Rheum 20:129, 1990
39. Rickles FR, Rick PD: Structural features of Salmonella ty- phimurium lipopolysaccharide required for activation of tissue fac- tor in human mononuclear cells. J Clin Invest 59: 1 188, I977
40. Busso N, Huet S, Nicodeme E, Hiemaux J, Hyafil F Refrac- tory period phenomenon in the induction of tissue factor expres- sion on endothelial cells. Blood 78:2027, 1991
41. Miserez R, Jungl TW: LPS-induced, but not interferon gamma-induced, procoagulant activity ofsuspended human macro- phages is followed by a refractory state of low procoagulant expres- sion. Thromb Res 65:733, 1992
42. Edwards RL, Levine JB, Green R, Duffy M, Mathews E, Brande W, Rickles F R Activation of blood coagulation in Crohn’s disease. Gastroenterology 92:329, 1987
43. Osterud B, Flaegstad T: Increased tissue thromboplastin ac- tivity in monocytes of patients with meningococcal infection: Re- lated to an unfavourable prognosis. Thromb Haemost 495, 1983
44. Cole EH, Schulman J, Urowitz M, Williams C, Levy CA: Monocyte procoagulant activity in glomerulonephritis associated with systemic lupus erythematosus. J Clin Invest 752361, 1985
45. Lyberg T, Prydz H, Baklien K, Hoyeraal HM: Effect of im- mune complex-containing sera from patients with rheumatic dis- ease on thromboplastic activity of monocytes. Thromb Res 25: 193, 1982
46. Rivers RPA, Cattermole HEJ, Wright I: The expression of surface tissue factor apoprotein by blood monocytes in the course of infections in early infancy. Pediatric Res 31:567, 1992
47. Tillet WS, Francis T Jr: Serological reactions in pneumonia with a nonprotein somatic fraction of pneumococcus. J Exp Med 52561, 1930
48. Shephard E, Anderson R, Beer SM, Jansen van Rensburg CE, De Beer FC: Neutrophil lysosomal degradation of human C R P CRP-derived peptides modulate neutrophil function. Clin Exp Im- munol 73: 139, 1988
49. Dobrinich R, Spagnuolo PJ: Binding ofC-reactive protein to human neutrophils. Inhibition of respiratory burst activity. Arthri- tis Rheum 34:1031, 1991
50. Ballou SP, Lozanski G: Induction of inflammatory cytokine release from cultured human monocytes by C-reactive protein. Cy- tokine 4:361, 1992
5 1. Gregory SA, Kornbluth RS, Helin H, Remold HG, Edging- ton TS: Monocyte procoagulant inducing factor: A lymphokine involved in the T cell-instructed monocyte procoagulant response to antigen. J Immunol 137:3231, 1986
52. Edgington TS, Mackman N, Fan S-T, Ruf W: Cellular im- mune and cytokine pathways resulting in tissue factor expression and relevance to septic shock. Nouv Rev Fr Hemato1345 15, 1992
53. Mortensen RF, Kidane A, Femandez MC, Tebo JM: C-reac- tive protein (CRP) receptor-mediated activation of monocytes. J Leukoc Biol20, I99 1 (abstr, suppl2)
54. Wickham NWR, Vercellotti GM, Hammerschmidt DE, Ja- cob HS: Complement activation by C-reactive protein: Enhance- ment by chylomicrons. Clin Res 39:335A, 1991 (abstr)
55. Gregory SA, Morrisey JH, Edgington TS: Regulation of tis- sue factor gene expression in the monocyte procoagulant response to endotoxin. Mol Cell Biol 9:2752, 1989
For personal use only. by guest on July 13, 2011. bloodjournal.hematologylibrary.orgFrom
Copyright © 2022 FDOKUMEN




















