
Development of colour-producing b-keratin nanostructures in avian feather barbs
Upload
independentCategory
view
0download
0
Rapid report
Boreal feather mosses secrete chemical signals to gain nitrogen
Author for correspondence:Ulla RasmussenTel: +46 8 163779
Email: [email protected]
Received: 16 April 2013Accepted: 6 June 2013
Guillaume Bay1, Nurun Nahar2, Matthieu Oubre2, Martin J. Whitehouse3,
David A. Wardle1, Olle Zackrisson4, Marie-Charlotte Nilsson1 and
Ulla Rasmussen2
1Department of Forest Ecology and Management, Swedish University of Agricultural Sciences, SE-901 83 Ume�a, Sweden;
2Department of Ecology, Environment and Plant Sciences, Stockholm University, SE-106 91 Stockholm, Sweden; 3Department of
Geosciences, Swedish Museum of Natural History, Box 50007 Stockholm, Sweden; 4The Institute for Subarctic Landscape Research,
SE-930 90 Arjeplog, Sweden
New Phytologist (2013) 200: 54–60doi: 10.1111/nph.12403
Key words: chemo-attractance,cyanobacteria, feather mosses, hormogonia,nitrogen transfer, symbiosis.
Summary
� The mechanistic basis of feather moss–cyanobacteria associations, a main driver of nitrogen
(N) input into boreal forests, remains unknown. Here, we studied colonization byNostoc sp. on
two feather mosses that form these associations (Pleurozium schreberi and Hylocomium
splendens) and two acrocarpous mosses that do not (Dicranum polysetum and Polytrichum
commune). We also determined how N availability and moss reproductive stage affects
colonization, and measured N transfer from cyanobacteria to mosses.� The ability of mosses to induce differentiation of cyanobacterial hormogonia, and of
hormogonia to then colonize mosses and re-establish a functional symbiosis was determined
throughmicrocosm experiments, microscopy and acetylene reduction assays. Nitrogen transfer
between cyanobacteria and Pleurozium schreberi was monitored by secondary ion mass
spectrometry (SIMS).� All mosses induced hormogonia differentiation but only feather mosses were subsequently
colonized. Colonization on Pleurozium schreberi was enhanced during the moss reproductive
phase but impaired by elevated N. Transfer of N from cyanobacteria to their host moss was
observed.� Our results reveal that feather mosses likely secrete species-specific chemo-attractants when
N-limited, which guide cyanobacteria towards them and fromwhich they gain N.We conclude
that this signalling is regulated by N demands of mosses, and serves as a control of N input into
boreal forests.
Introduction
Pleurocarpous feather mosses have a wide global distribution andfrequently dominate the ground layer in the boreal forest, which isthe Earth’s largest forested biome. They can reach a biomass of upto 3 t ha�1 and can account for up to a third of total forest primaryproductivity (Wardle et al., 2012). They also commonly hostdinitrogen (N2)-fixing cyanobacteria that are epiphytic (i.e. livingon the surface of moss leaves) and which serve as the main agent ofbiological nitrogen (N) input into mature forests (DeLuca et al.,2002). As such, there is emerging recognition of the important roleof feathermosses in the functioning of boreal forests, for example ingoverning nutrient fluxes, carbon sequestration and the establish-ment and growth of vascular plants (Turetsky et al., 2012).However, despite the ecological importance ofmoss–cyanobacteria
associations, little is known about the intrinsic factors regulatingthe capacity ofmosses to host cyanobacteria, or the potential benefitthat theymay obtain from the association. An improved knowledgeof the mechanisms involved will help us better understand themeans by which N enters into extensive areas worldwide in whichcryptogamic species are major contributors (Elbert et al., 2012).
In recent work, we have shown that strains of the N2-fixingcyanobacterial genusNostocwere highly host specific, with differentstrains associating with each of the two dominant and mostwidespread boreal feather mosses, that is Pleurozium schreberi andHylocomium splendens (Ininbergs et al., 2011). This result suggeststhat feather mosses are highly selective towards the cyanobacterialstrains that they host. Consequently, these results raise thepossibility that mosses may secrete compounds that serve aschemo-attractants for cyanobacteria, and that these cyanobacteria
54 New Phytologist (2013) 200: 54–60 � 2013 The Authors
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research

could in turn contribute to moss nutrition by providing mosseswith fixed N, in the same manner as has been shown for vascularplant–cyanobacterial symbioses (Adams & Duggan, 2012).
First, we hypothesized that colonization of feathermosses byN2-fixing cyanobacteria occurs as a result of the secretion of specificchemical signals (a hormogonia inducing factor (HIF) and achemo-attractant) that induce hormogonia formation (i.e. tran-sient motile filaments serving as infection units) and guide themtowards the host moss. Second, we hypothesized that the secretionof the chemo-attractant is maximized when the host moss isdepleted in N. Third, we also hypothesized that the N2 fixed by thecyanobacteria is transferred to their host moss. Testing thesehypotheses would inform on whether feather mosses secretechemicals to attract cyanobacteria from which they then gain anutritional benefit.
Materials and Methods
Sampling site, moss species and cyanobacterial strain
All mosses used in this study (Hylocomium splendens (Hedw.)W.P.Schimp, Pleurozium schreberi (Brid.) Mitt., Dicranum polysetumSw. and Polytrichum commune Hedw.) were collected in northernSweden (65°80′N; 19°10′E) in June 2010. Hylocomium splendensand P. schreberi were used as host species (Ininbergs et al., 2011),while D. polysetum and P. commune were used as nonhost speciesbecause no or few cyanobacteria occur on them (Scheirer &Dolan,1983). A monoculture ofNostoc sp. isolated fromH. splendens wasused for all the experiments.
Hormogonia induction assessment
Gametophytes of all moss species were surface-sterilized to removeall epiphytic cyanobacteria using 0.15% sodium hypochlorite for1 min followed by seven rinseswith sterilewater to remove any traceof sodium hypochlorite (Misaghi & Donndelinger, 1990). Toverify that all cyanobacteria had been removed, the gametophyteswere observed under an epifluorescence microscope equipped witha green excitation filter (510–560 nm). One millilitre of theNostocsp. culture (100 mg ml�1 in BG-110 medium; Rippka et al., 1979)was added to each well of two 12-well BD Falcon plates. For eachmoss species, four replicates were prepared, where each gameto-phyte (each c. 10 mg DW) was placed in a fitting cell culture insert(1 lmmembrane pore size). Only the upper 2 cm green parts of thegametophyte were used as this is where most of the cyanobacterialN2-fixation activity occurs (Solheim et al., 2004). Each insert wasplaced in the cyanobacteria-filled wells of the plates. With this set-up, the gametophytes were in contact with the Nostoc culture butphysically separated from it because the diameter of the Nostocfilaments is 1.5–2 lm, allowing no filaments to pass through themembrane. Nostoc cultures without bryophytes present in the cellinserts were used as negative controls. The plates were placed in agrowth chamber for 72 h at 24°C under constant illumination of18 lmol photons m�2 s�1. Hormogonia formation in the culturemedia wasmeasured every 24 h using a hemocytometer. The pHofthe culture media was determined before each counting.
Assessment of chemo-attraction
The ability of mosses to attract cyanobacteria from the culturemedium was assessed as mentioned earlier, except that an 8 lmpore size membrane was used allowing the movements ofhormogonia through the membrane, and that nine gametophytereplicates of each species were used. From these nine replicates, fourwere incubated in darkness at 24°C to exclude the possibility thatcyanobacterial colonization was due to light attraction. Cyanobac-terial colonizationwas quantified after 8 d as the proportion ratio ofthe total surface area occupied by cyanobacteria, using ImageJsoftware v.1.45s (Rasband, 1997–2012).
To assess whether chemo-attraction of cyanobacteria by mossesdepends on the N demand of the host, we established two1 m9 1 m field plots dominated by P. schreberi in northernSweden (see earlier) in June 2007.One of the plots was fertilized byN, added as ammonium nitrate (NH4NO3) dissolved in water, at5 kg N ha�1 yr�1, and the other was an unfertilized controlamended with equivalent amounts of water. Nitrogen and waterwere distributed at seven application times from June to Septemberbetween 2007 and 2010. This N addition rate is consistent withrates used in previous experiments in boreal forests to study theimpairment of N2-fixation by N fertilization (Gundale et al.,2011). In September 2010, five P. schreberi gametophytes wererandomly collected in each of three 0.29 0.2 m sub-plots. Thegametophytes were exposed to cyanobacteria as described earlier,and cyanobacterial colonization was assessed after 8 d of incubation.
To test if chemo-attraction of cyanobacteria by P. schreberi isaffected by the reproductive costs of producing sporophytes, wecollected 24 P. schreberi gametophytes with well-developed sporo-phytes and12 gametophyteswithout sporophytes from thefield sitein September 2012. For 12 of the 24 gametophytes withsporophytes, the sporophytes were manually removed to reducethe reproductive costs to the gametophytes. All gametophytes werearranged in 12 replicate blocks; each block consisted of all threegametophyte types (i.e. gametophytes with sporophytes, those withsporophytes removed, and gametophytes with no sporophytes).The mosses were left in a growth chamber for 10 d to acclimatizeuntil chemo-attraction was assessed. All gametophytes weresurface-sterilized before experimentation, and assessment ofchemo-attraction and N2-fixation rates was performed 8 d afterexperimental set-up.
N2-fixation measurements
Nitrogen-fixation activity of the gametophytes used during thedifferent experiments was measured using the acetylene reductionassay (ARA; Hardy et al., 1968). The gametophytes were placedindividually in 16-ml glass tubes fitted with rubber septa. Ten percent of the air volumewas removed from the tube and replacedwithacetylene. The tubes were incubated in a growth chamber at 24°Cunder constant illumination of 18 lmol photons m�2 s�1 for 4 h,and measurements of ethylene levels were performed as describedby Ininbergs et al. (2011). A ratio of 3 moles of ethylene per molof N was used to convert acetylene reduction rates tong N2 fixed g moss�1 d�1 (DeLuca et al., 2002).
� 2013 The Authors
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013) 200: 54–60
www.newphytologist.com
NewPhytologist Rapid report Research 55

Nitrogen transfer between cyanobacteria andPleurozium schreberi gametophytes
Cyanobacteria-hosting P. schreberi gametophytes were incubatedwith 15N2 gas to obtain
15N-enriched samples. Sixty gametophyteswere placed in 12 serum bottles (five in each) sealed with rubbercaps. Gametophytes in six of these bottles were exposed to 2% 15N2
gas while those in the other six were exposed to air; these were thenincubated in a growth chamber under a light regime of167 lmol photons m�2 s�1 for 12 h d�1 at 24°C. Half the bottles(i.e. three exposed to 15N2 gas and three exposed to air) wereincubated for 1 wk, and the other half were incubated for 4 wk.The incubations were stopped by adding 2% freshly preparedparaformaldehyde solution to each bottle, and the bottles werefurther incubated for 16 h at 4°C. The gametophytes were washedtwice with phosphate-buffered saline (pH 7.4) and stored at 4°Cuntil further processed for secondary ion mass spectrometry(SIMS) analysis (for embedding and sectioning of samples, seeSupporting Information Methods S1). This analysis is both aqualitative and quantitative technique that was used in thisexperiment to determine the 15N2 isotopic composition on thesurface of moss cross-sections. Before SIMS analysis (for both15N2-exposed and non 15N2-exposed gametophytes), leaf areaswith cyanobacteria were identified by microscopy, and coated with5 nm gold (Au) to provide a conducting surface. Scanning ionimaging was performed using a large geometry CAMECAims1280 SIMS instrument (Cameca SAS, Gennevilliers, France).A 133Cs+ primary beam with 20 kV impact energy was criticallyfocused to produce a < 1 lm analytical spot on the sample, with abeam current of c. 100 pA which was rastered over an area of809 80 lm. Following the removal of the Au-coat from the areato be analysed by using a larger rastered primary beam, thesecondary ion species of 12C14N and 13C15N were sequentiallymeasured in an ion counting electron multiplier at a massresolution (M/DM) of 5000. The dynamic transfer optical systemwas utilized to enable reconstruction of the spatial distribution ofthe measured ion species within the rastered sampling area. Eachspot analysed consisted of 120 cycles through the masses 12C14N�,12C15N� and 13C14N� (1, 5 and 2 s integration time, respec-tively). Image processing of the analysed spots was performed usingCAMECAWinimage software to generate accumulated ion mapsof 15N and 14N. Calculation of the 15N/14N isotopic ratio of mossand cyanobacterial cells was performed by using high resolutionspectra from the control (non 15N2-exposed) mosses and the 15N2-exposedmosses at mass peaks 26 and 27, respectively, to determinethe major mass of 12C14N� and 12C15N� of the chosen area. Themean 15N/14N ratios of non 15N2-exposed samples were used tocorrect the 15N/14N values of 15N2-exposed samples frombackground noise. Due to the heterogeneity of resin-embeddedsamples, the total number of areas (n) analysed for 15N/14N ratioamong moss and cyanobacterial tissues is not equal.
Statistical analysis
The ability of the bryophytes to induce hormogonia formationwas tested in a linear mixed effects model using a Gaussian error,
where the fixed effect was the moss treatment (i.e. each of thefour moss species and the moss-free control) and the randomeffect was the time of exposure. The effect of time onhormogonia formation was tested by analysis of variance(ANOVA) for each time point, on Box-Cox transformed data.The effects of moss species, N amendment and sporophytepresence on cyanobacterial colonization and N2-fixation wereeach assessed by ANOVA, on Box-Cox transformed data.Tukey’s HSD (honestly significant difference) post hoc tests weresubsequently performed to investigate significant differencesbetween moss species, and between treatments (i.e. N amend-ment and sporophyte presence) at P = 0.05. The effect of 15N2
exposure on 15N2 enrichment in gametophytes was assessed by anonparametric Wilcoxon test. All statistical analyses were carriedout using R software v.2.13.1 (R Foundation for StatisticalComputing, Vienna, Austria).
Results
Hormogonia induction assessment
All four bryophyte species induced hormogonia differentiation inNostoc sp. cultures after 24 h of incubation (Fig. 1). After 48 h,more than half of the cyanobacterial filaments in cultures hadformed hormogonia when exposed to mosses, whereas only 9% ofthe filaments formed hormogonia in the absence of mosses.Depending on the moss species, hormogonia differentiationdecreased or levelled out when cyanobacteria were exposed tomosses for longer than 48 h (Fig. 1). Measurements of the pHwithin the cell culture medium revealed a slight acidification of themedium over time regardless of the moss species (Fig. 1).
6.7
6.8
6.9
7
7.1
7.2
7.3
0
10
20
30
40
50
60
70
0 24 48 72
pH
Hor
mog
onia
indu
ctio
n (%
)
Time of incubation with the mosses (h)
P. schreberiH. splendens
P. communeD. polysetum
ControlpH
a
cde
f
e
cb
a a a
Fig. 1 Hormogonial formation of Nostoc sp. culture (percentage of totalnumber of filaments) exposed to Pleurozium schreberi, Hylocomium
splendens, Polytrichum commune and Dicranum polysetum over 72 h(mean� SEM; data points accompanied by different letters are statisticallysignificant at P = 0.05 according to a linearmixed effectsmodel combined toANOVAs for each timepoint, followedbyTukey’s test;n = 4eachdatapoint,df = 4 for eachANOVA), andpHvariation in the cell culture inserts during theexperiment (mean� SEM;n = 4 each data point).Nostoc sp. culturewithoutmoss was used as the control.
New Phytologist (2013) 200: 54–60 � 2013 The Authors
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research Rapid reportNewPhytologist56
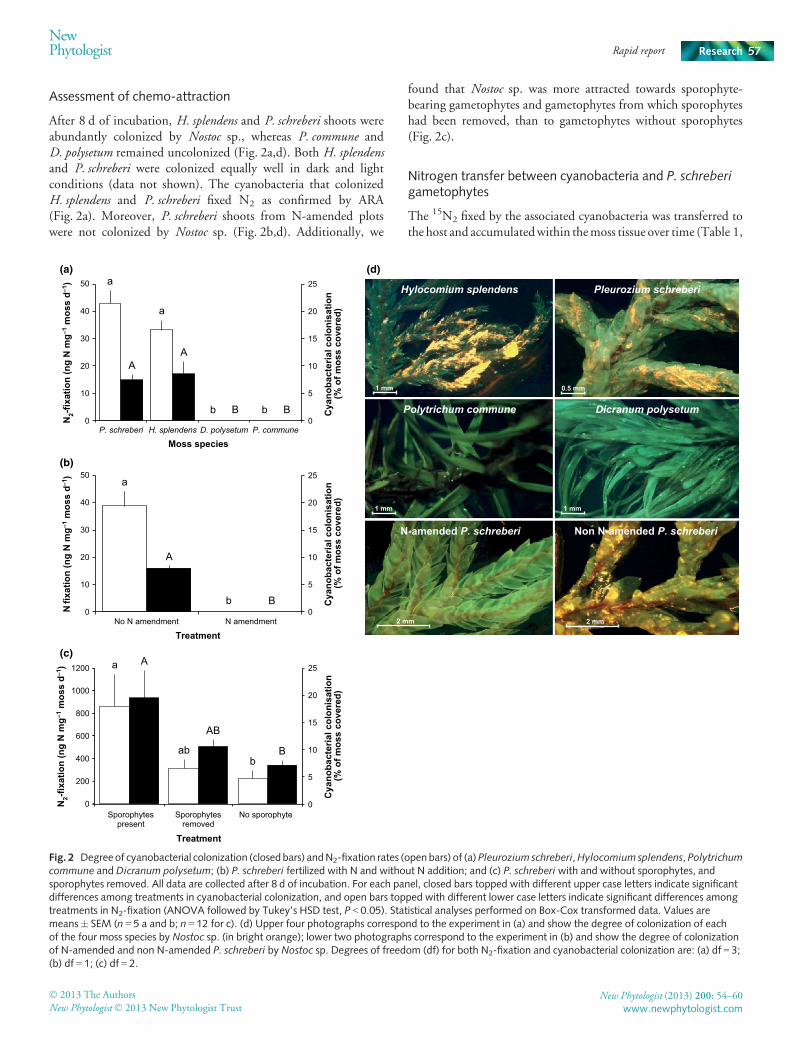
Assessment of chemo-attraction
After 8 d of incubation, H. splendens and P. schreberi shoots wereabundantly colonized by Nostoc sp., whereas P. commune andD. polysetum remained uncolonized (Fig. 2a,d). Both H. splendensand P. schreberi were colonized equally well in dark and lightconditions (data not shown). The cyanobacteria that colonizedH. splendens and P. schreberi fixed N2 as confirmed by ARA(Fig. 2a). Moreover, P. schreberi shoots from N-amended plotswere not colonized by Nostoc sp. (Fig. 2b,d). Additionally, we
found that Nostoc sp. was more attracted towards sporophyte-bearing gametophytes and gametophytes from which sporophyteshad been removed, than to gametophytes without sporophytes(Fig. 2c).
Nitrogen transfer between cyanobacteria and P. schreberigametophytes
The 15N2 fixed by the associated cyanobacteria was transferred tothe host and accumulatedwithin themoss tissue over time (Table 1,
(b)
0
5
10
15
20
25
0
10
20
30
40
50
No N amendment N amendment
Cya
noba
cter
ial c
olon
isat
ion
(% o
f mos
s co
vere
d)
Treatment 2 mm
1 mm
1 mm
0.5 mm
1 mm
Hylocomium splendens
2 mm
Pleurozium schreberi
Polytrichum commune Dicranum polysetum
N-amended P. schreberi Non N-amended P. schreberi
(d)
a
A
B b
a
ab b
A
AB
B
0
5
10
15
20
25
0
10
20
30
40
50
P. schreberi H. splendens D. polysetum P. commune
Cya
noba
cter
ial c
olon
isat
ion
(% o
f mos
s co
vere
d)
N2-
fixat
ion
(ng
N m
g–1 m
oss
d–1)
N2-
fixat
ion
(ng
N m
g–1 m
oss
d–1)
Moss species
a
a
A A
B b B b
(a)
(c)
Sporophytes present
Sporophytes removed
No sporophyte
Treatment
0
5
10
15
20
25
Cya
noba
cter
ial c
olon
isat
ion
(% o
f mos
s co
vere
d)
0
200
400
600
800
1000
1200
N fi
xatio
n (n
g N
mg–1
mos
s d–1
)
Fig. 2 Degree of cyanobacterial colonization (closedbars) andN2-fixation rates (open bars) of (a)Pleurozium schreberi,Hylocomiumsplendens, Polytrichumcommune and Dicranum polysetum; (b) P. schreberi fertilized with N and without N addition; and (c) P. schreberi with and without sporophytes, andsporophytes removed. All data are collected after 8 d of incubation. For each panel, closed bars topped with different upper case letters indicate significantdifferences among treatments in cyanobacterial colonization, and open bars topped with different lower case letters indicate significant differences amongtreatments in N2-fixation (ANOVA followed by Tukey’s HSD test, P < 0.05). Statistical analyses performed on Box-Cox transformed data. Values aremeans� SEM (n = 5 a and b; n = 12 for c). (d) Upper four photographs correspond to the experiment in (a) and show the degree of colonization of eachof the four moss species by Nostoc sp. (in bright orange); lower two photographs correspond to the experiment in (b) and show the degree of colonizationof N-amended and non N-amended P. schreberi by Nostoc sp. Degrees of freedom (df) for both N2-fixation and cyanobacterial colonization are: (a) df = 3;(b) df = 1; (c) df = 2.
� 2013 The Authors
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013) 200: 54–60
www.newphytologist.com
NewPhytologist Rapid report Research 57

see Supporting InformationTable S1 for values of d15N& of 15N2-exposed and non 15N2-exposed cyanobacteria associated withP. schreberi). Even 1 wk after exposure, a significantly higher15N/14N ratio was observed in the host tissue of 15N2-exposedgametophytes than in nonexposed gametophytes. This ratiocorresponds to a 4.1% 15N enrichment when compared withnatural abundance measured in nonexposed P. schreberi gameto-phyte tissues. After 4 wk of exposure, the enrichment in the mosstissue increased to 34.4% (Table 1). The cyanobacteria associatedwith P. schreberi also showed an enrichment of 15N when exposedto 15N2 gas (Table 1, Fig. 3). Contrary to the moss tissue, theenrichment of 15N in cyanobacterial cells did not significantlyincrease with time of exposure.
Discussion
Our aim was to shed light on the underlying mechanisms of Ninput into boreal ecosystems; more specifically, on which factorsdetermine if amoss is a suitable host for cyanobacteria andwhat thefate is of N fixed by the associative cyanobacteria. Our resultsshowed that all four mosses promoted hormogonia differentiationof Nostoc sp. after 24 h of incubation (Fig. 1), which supports ourfirst hypothesis. Hormogonia are induced by a variety of environ-mental stimuli, including chemical signals (HIFs) released by hosts(Campbell&Meeks, 1989; Rasmussen et al., 1994). Therefore, weconclude that these mosses secrete a HIF in a similar manner toGunnera or Anthoceros (Campbell & Meeks, 1989; Rasmussenet al., 1994). Moreover, the observed decrease or plateau after 72 hof incubation (Fig. 1) is in accordance with previous findings byMeeks & Elhai (2002), which showed that Nostoc hormogoniacease to move and revert to vegetative filaments after 48 h.Additionally, the acidification of the medium during the experi-ment did not appear to have any effect on hormogonia differen-tiation and suggests that the HIF secreted by each of the moss
species is strong enough to override the possible negative effects ofthe pH on hormogonia differentiation (Hirose, 1987; Rasmussenet al., 1994).
Our results also showed that H. splendens and P. schreberi werethe only mosses colonized by Nostoc sp. (Fig. 2a,d). This demon-strates that hormogonia are exclusively attracted towards thesefeather mosses due to the probable production of host-specificchemical compounds that would induce chemotactic responses incyanobacteria, and therefore also provides support for our firsthypothesis. Additionally, the cyanobacteria which colonizedP. schreberi and H. splendens fixed N2 (Fig. 2a), suggesting that afunctional symbiosis was formed for both of these mosses aftercolonization. While this study was not aimed at identifying the
Table 1 15N/14N isotopic ratios (mean� SEM) of 15N2-exposed and control(non 15N2-exposed) Pleurozium schreberi gametophyte cells and theirassociated cyanobacteria
Exposuretime Exposed to 15N2 Control P-Value
P. schreberi cells1 wk 3.85e�03� 3.77e�03
(n = 13)3.65e�03� 3.02e�05
(n = 13)1.019e�03
4 wk 4.97e�03� 2.16e�04 *(n = 65)
3.73e�03� 8.05e�05
(n = 31)9.239e�15
Cyanobacterial cells1 wk 7.76e�03� 3.07e�03
(n = 8)3.69e�03� 1.04e�04
(n = 60)5.145e�6
4 wk 7.47e�03� 2.65e�03
(n = 185)3.73e�03� 8.05e�05
(n = 21)2.2e�16
Data are obtained from secondary ion mass spectrometry (SIMS) analyses 1and 4wk after initiation of 15N2-exposure.The number of spots (n) analysed with SIMS is given for each sample.P-Values indicate significant differences between cells exposed to 15N2 andcontrol cells, according to a nonparametric Wilcoxon rank sum test.*Significant enrichment of 15N between P. schreberi cells after 1 and 4 wk ofexposure (P = 1.096e�05).
(a)
(b)
(c)
Fig. 3 Cross-sections of a Pleurozium schreberi gametophyte withcyanobacteria after 4 wkof 15N2exposure. (a) Lightmicroscopy imagewithared box indicating the area subsequently analysed by secondary ion massspectrometry (SIMS). (b) Close-up SIMS images ofmoss tissue (11–13, blackarrows) and cyanobacterial cells (4–10 and 14, blue arrows) chosen for15N/14N ratio measurements. The boxes (1–3, white arrows) to the leftdepict background areas. (c) Colour gradient showing the distribution of15N-enrichment within moss and cyanobacterial cells.
New Phytologist (2013) 200: 54–60 � 2013 The Authors
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research Rapid reportNewPhytologist58

precise molecular structures underpinning why H. splendensand P. schreberi but not D. polysetum and P. commune becomecolonized byNostoc sp., our results help to explainwhyD. polysetumandP. commune remain free of cyanobacteria in natural conditions.Thus, we conclude that although hormogonia differentiation is acrucial step in the cyanobacterial colonization of host plants, forsuccessful colonization to occur the plant must also attractand guide the hormogonia towards suitable colonization sites(i.e. leaves).
In support of our second hypothesis, we found that P. schreberifrom N-amended plots remained uncolonized by Nostoc sp.,indicating that the moss ceases to secrete chemo-attractants underelevated N conditions (Fig. 2b,d). Furthermore, we found that thereproductive status of the moss positively impacted cyanobacterialcolonization and plausible chemo-attractant secretion (Fig. 2c).This result also is consistent with sporophyte formation being aN-demanding process (Rydgren & Økland, 2002) and withsporophytes serving asN sinks (Renault et al., 1989).Moreover, theremoval of sporophytes from the gametophytes probably inducedchanges in the source-sink relationship between these two compo-nents, creating a reduced N demand, and explains why gameto-phytes with sporophytes removed had intermediate colonizationrates (Fig. 2c). Thus, P. schreberi is likely to allocate resources to theproduction of chemo-attractants only under N-limited conditionsor when the moss has a greater need for N (e.g. during sexualreproduction). Source–sink relationships between N2-fixation andflowering have been widely reported for legumes (Sprent, 2009),and our results suggest that a similar phenomenon involving activeproduction of chemo-attractantsmay apply for boreal forest feathermosses. Sporophyte development is seasonal (Longton & Greene,1969) and these results may also help to explain the previouslyobserved seasonality of feather mosses N2-fixation patterns(DeLuca et al., 2002).
Finally, a transfer of N was observed between cyanobacteria andP. schreberi when gametophytes were exposed to 15N2 gas (Fig. 3).Thus, feather mosses acquire N from their associated cyanobac-teria, which eventually accumulates in moss tissues over time(Table 1), and therefore provides support for our third hypothesis.Although the levels of 15N within the cyanobacteria cells alsoincreased throughout the experiment, no accumulation wasobserved with time of exposure (Table 1), most likely because theN fixed by the cyanobacteria is instead transferred to their hostmoss.
Moss–cyanobacteria associations are widespread in boreal forest,tundra and sub-Arctic ecosystems (Turetsky, 2003; Lindo &Gonzalez, 2010), and frequently represent the main form ofbiological N input into N-limited ecosystems. However, we havelimited understanding of how N2-fixing cyanobacteria colonizemosses, andwhether this fixedNbecomes available to themoss andultimately to the rest of the ecosystem (Gavazov et al., 2010). It hasbeen suggested that cyanobacterial colonization of host mosses is arandom phenomenon due to mechanical means, for example wind(Marshall & Chalmers, 1997). Our findings show that theinitiation of associations between boreal feather mosses andhormogonia-forming cyanobacteria is principally directed by themosses, and that the mosses regulate the degree of cyanobacterial
colonization based on their need for N. Our findings furthersuggest that the ability of feather mosses to occupy N-limitedecosystems has evolved through chemical communication withcyanobacteria, and that cyanobacteria fulfil important functions forthem through the provision of fixed N2. Ultimately, these findingsreveal that the feather moss–cyanobacteria association operates as afunctional symbiotic relationship driven by chemical signallingthat supports the N economy of feather mosses, and refines ourunderstanding of how feather mosses govern N input into borealforests.
Acknowledgements
Funding was provided by the Carl Trygger Foundation forScientific Research and Stiftelsen Oscar och Lili Lamms Minne.The authors are grateful to H. Ploug and S. Lindwall for technicalassistance. TheNordsims facility is financed and operated under anagreement between the Nordic countries; this is Nordsimscontribution no. 343.
References
Adams DG, Duggan PS. 2012. Signalling in cyanobacteria–plant symbioses. In:
Perotto S, Balu�ska F, eds. Signalling and communication in plant symbiosis. Berlin,Germany: Springer-Verlag, 93–121.
Campbell EL, Meeks JC. 1989. Characteristics of hormogonia formation by
symbiotic Nostoc spp. in response to the presence of Anthoceros punctatus orits extracellular products. Applied and Environmental Microbiology 55: 125–131.
DeLuca TH, Zackrisson O, Nilsson M-C, Sellstedt A. 2002.Quantifying
nitrogen-fixation in feather moss carpets of boreal forests.Nature 419: 917–920.Elbert W, Weber B, Burrows S, Steinkamp J, Budel B, Andreae MO, Poschl U.
2012. Contribution of cryptogamic covers to the global cycles of carbon and
nitrogen. Nature Geoscience 5: 459–462.Gavazov KS, Soudzilovskaia NA, van Logtestijn RSP, Braster M, Cornelissen
JHC. 2010. Isotopic analysis of cyanobacterial nitrogen fixation associated with
subarctic lichen and bryophyte species. Plant and Soil 333: 507–517.Gundale MJ, DeLuca TH, Nordin A. 2011. Bryophytes attenuate anthropogenic
nitrogen inputs in boreal forests. Global Change Biology 17: 2743–2753.Hardy RWF, Holsten RD, Jackson EK, Burns RC. 1968. The acetylene-ethylene
assay for N2 fixation: laboratory and field evaluation. Plant Physiology 43:1185–1207.
HiroseM. 1987. Energy supply system for the glidingmovement of hormogonia of
the cyanobacterium Nostoc cycadae. Plant and Cell Physiology 28: 587–597.Ininbergs K, BayG, RasmussenU,WardleDA,NilssonM-C. 2011.Composition
and diversity of nifH genes of nitrogen-fixing cyanobacteria associatedwith boreal
forest feather mosses. New Phytologist 192: 507–517.Lindo Z, Gonzalez A. 2010.The bryosphere: an integral and influential component
of the Earth’s biosphere. Ecosystems 13: 612–627.Longton RE, Greene SW. 1969. Growth and reproductive cycle of Pleuroziumschreberi (Brid.) Mitt. Annals of Botany 33: 83–105.
MarshallWA, ChalmersMO. 1997.Airborne dispersal of antarctic terrestrial algae
and cyanobacteria. Ecography 20: 585–594.Meeks JC, Elhai J. 2002. Regulation of cellular differentiation in filamentous
cyanobacteria in free-living and plant-associated symbiotic growth states.
Microbiology and Molecular Biology Reviews 66: 94–121.Misaghi IJ, Donndelinger CR. 1990. Endophytic bacteria in symptom-free cotton
plants. Phytopathology 80: 808–811.RasbandWS. 1997–2012. ImageJ. Bethesda, MD, USA: US National Institutes of
Health. http://imagej.nih.gov/ij
Rasmussen U, Johansson C, Bergman B. 1994. Early communication in the
Gunnera–Nostoc symbiosis: plant-induced cell differentiation and protein synthesis
in the cyanobacterium. Molecular Plant-Microbe Interactions 7: 696–702.
� 2013 The Authors
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013) 200: 54–60
www.newphytologist.com
NewPhytologist Rapid report Research 59

Renault S, Despeghel-Caussin C, Bonnemain J-L, Delrot S. 1989. The proton
electrochemical transmembrane gradients generated by the transfer cells of the
haustorium of Polytrichum formosum and their use in the uptake of amino acids.
Plant Physiology 90: 913–920.Rippka R, Deruelles J, Waterbury JB, Herdman M, Stanier RY. 1979. Generic
assignments, strain histories and properties of pure cultures of cyanobacteria.
Journal of General Microbiology 111: 1–61.Rydgren K, Økland RH. 2002. Ultimate costs of sporophyte production in the
clonal moss Hylocomium splendens. Ecology 83: 1573–1579.Scheirer DC,DolanHA. 1983.Bryophyte leaf epiflora: an SEM and TEM study of
Polytrichum communeHedw. American Journal of Botany 70: 712–718.Solheim B, Wiggen H, Røberg S, Spaink HP. 2004. Associations between arctic
cyanobacteria and mosses. Symbiosis 37: 169–187.Sprent JI. 2009. Legume nodulation: a global perspective. Oxford, UK:
Wiley-Blackwell.
Turetsky MR. 2003. The role of bryophytes in carbon and nitrogen cycling.
Bryologist 106: 395–409.Turetsky MR, Bond-Lamberty B, Euskirchen E, Talbot J, Frolking S, McGuire
AD, Tuittila E-S. 2012. The resilience and functional role of moss in boreal and
arctic ecosystems. New Phytologist 196: 49–67.WardleDA, JonssonM, Bansal S, Bardgett RD,GundaleMJ,MetcalfeDB. 2012.
Linking vegetation change, carbon sequestration and biodiversity: insights from
island ecosystems in a long-term natural experiment. Journal of Ecology 100:16–30.
Supporting Information
Additional supporting information may be found in the onlineversion of this article.
Table S1 d15N& of 15N2-exposed and non 15N2-exposedcyanobacteria associated with Pleurozium schreberi, analysed byelemental analysis isotope ratio mass spectrometry (EA-IRMS)
Methods S1 Embedding and sectioning of gametophytes forsecondary ionmass spectrometry (SIMS) analysis, and 15N contentanalysis of 15N-exposed and non-exposed Pleurozium schreberisamples by elemental analysis isotope ratiomass spectrometry (EA-IRMS).
Please note: Wiley Blackwell are not responsible for the content orfunctionality of any supporting information supplied by theauthors. Any queries (other than missing material) should bedirected to the New Phytologist Central Office.
New Phytologist is an electronic (online-only) journal owned by the New Phytologist Trust, a not-for-profit organization dedicatedto the promotion of plant science, facilitating projects from symposia to free access for our Tansley reviews.
Regular papers, Letters, Research reviews, Rapid reports and both Modelling/Theory and Methods papers are encouraged. We are committed to rapid processing, from online submission through to publication ‘as ready’ via Early View – our average timeto decision is <25 days. There are no page or colour charges and a PDF version will be provided for each article.
The journal is available online at Wiley Online Library. Visit www.newphytologist.com to search the articles and register for tableof contents email alerts.
If you have any questions, do get in touch with Central Office ([email protected]) or, if it is more convenient,our USA Office ([email protected])
For submission instructions, subscription and all the latest information visit www.newphytologist.com
New Phytologist (2013) 200: 54–60 � 2013 The Authors
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research Rapid reportNewPhytologist60
Copyright © 2022 FDOKUMEN




















