
Translation Series No. 3558 - Fisheries and Oceans Canada ...
Upload
independentCategory
view
3download
0
MiniReview
Marine bacteria and biogeochemical cycling of iron in the oceans
Philippe D. Tortell a;*, Maria T. Maldonado b, Julie Granger b, Neil M. Price b
a Department of Ecology and Evolutionary Biology, Princeton University, Princeton, NJ 08544, USAb Department of Biology, McGill University, Montreal, Que. H3A 1B1, Canada
Received 18 June 1998; received in revised form 2 December 1998; accepted 8 December 1998
Abstract
Prokaryotic microbes play a critical role in oceanic Fe cycling. They contain most of the biogenic Fe in offshore waters andare responsible for a large portion of the Fe uptake by the plankton community. In the subarctic North Pacific, surfacepopulations of heterotrophic species assimilate more than 50% of the dissolved Fe and thus compete directly withphytoplankton for this limiting resource. In oligotrophic tropical and subtropical waters, photosynthetic bacteria become moreimportant in Fe cycling as the number of unicellular cyanobacteria increases and the nitrogen-fixing Trichodesmium, whichcontains most of the biogenic Fe in the mixed layer, becomes abundant. Like their terrestrial counterparts, heterotrophic andphototrophic marine bacteria produce Fe-binding siderophores that are involved in Fe acquisition. Evidence exists thatbacteria may modify Fe chemistry in the sea through the production of these ligands and thereby play a significant role inregulating production of eukaryotic phytoplankton. z 1999 Federation of European Microbiological Societies. Publishedby Elsevier Science B.V. All rights reserved.
Keywords: Iron chemistry; Marine bacterium; Iron cycling
1. Introduction
Over the past decade and a half, it has becomeapparent that trace metals can a¡ect oceanic primaryproductivity as limiting nutrients and toxic inhibitors[1]. As a result, the biogeochemical cycling of tracemetals in the oceans has become a subject of greatinterest and research. Of all trace metals, Fe has thusfar received most attention. Shipboard bioassay ex-periments [2^4] as well as two mesoscale in situ Fefertilizations [5,6] have demonstrated that low Feavailability constrains phytoplankton growth in sev-eral large open ocean regions where primary produc-tion is low despite high concentrations of major nu-
trients (nitrate, phosphate, silicate). Iron limitationin these regions decreases the e¤ciency of the `bio-logical carbon pump' through which CO2 is con-sumed in surface waters and transported as sinkingparticulate organic carbon to the deep sea. Given theimportance of this process in controlling atmosphericCO2 [7], Fe biogeochemistry in the oceans has im-portant implications for global carbon cycling andclimate studies.
At present, our understanding of Fe biogeochem-istry is incomplete. Many of the processes that arethought to control dissolved and particulate Fe con-centrations in the oceans are known from laboratorystudies that mimic natural conditions with varyingdegrees of success. One fact that is clear, however,is that biological processes are of great importance.
0168-6496 / 99 / $20.00 ß 1999 Federation of European Microbiological Societies. Published by Elsevier Science B.V. All rights reserved.PII: S 0 1 6 8 - 6 4 9 6 ( 9 8 ) 0 0 1 1 3 - 5
FEMSEC 997 21-4-99
* Corresponding author.
FEMS Microbiology Ecology 29 (1999) 1^11

Most investigations of Fe-biota interactions to datehave focused on eukaryotic phytoplankton, whichaccount for the bulk of CO2 export. Relatively littleattention has been given to the role of phototrophicand heterotrophic bacteria in Fe cycling. This is inmarked contrast to studies of ocean carbon and ni-trogen dynamics where these organisms have beenshown to play a central role [8,9].
In this review, we discuss the biogeochemistry ofFe in the oceans with speci¢c emphasis on the role ofprokaryotes. In particular, we highlight severalmechanisms through which photosynthetic cyano-bacteria and heterotrophic bacteria may a¡ect Febioavailability and cycling. Despite the recently dis-covered abundance of Archaea in the oceans [10,11]we shall not explicitly consider these organisms forlack of relevant physiological data. Given the tre-mendous advances in aquatic microbiology and rap-id development of new molecular techniques, it maysoon be possible to examine bacterial-Fe interactionsacross the broad taxonomic and biochemical diver-sity of marine prokaryotes. Our discussion points toseveral important questions that will need to be ad-dressed by such future research.
2. Biogeochemistry of dissolved iron in the sea
Although Fe is the fourth most abundant elementin the earth's crust, its concentration in most oceanwaters is vanishingly low. Until recently, ubiquitouscontamination of samples during collection andanalysis prevented accurate determination of truedissolved Fe levels. The advent of ultra-trace metalclean techniques [12], however, facilitated accurateand systematic measurements of Fe in the world'soceans [13] and provided new insight into the roleof marine biota in Fe cycling. Below, we brie£y sum-marize the most salient features of dissolved Fe dis-tributions and chemistry, and discuss recent evidencefor bacterial control of Fe speciation and solubility.
Like nearly all trace metals, dissolved Fe concen-trations show strong horizontal and vertical gra-dients, decreasing by as much as several hundred-fold from coastal to o¡shore waters and increasingsigni¢cantly with depth in the upper 500 meters [13].Coastal waters receive large inputs of Fe from riversand anoxic sediments [13] whereas o¡shore regions
rely mainly on atmospheric dust deposition and/orupwelling of deep waters as sources of Fe [14,15].We shall focus our discussion on oceanic (o¡shore)regions that account for s 90% of marine primaryproductivity and are characterized by persistentlylow Fe availability. Such low Fe levels demonstrablylimit primary productivity in high-nutrient oceanicregions [6] and may impose a signi¢cant stress onphytoplankton in oligotrophic subtropical gyres. Re-cent work has demonstrated that coastal upwellingregimes can also be Fe-de¢cient [16].
Dissolved Fe concentrations in o¡shore waters ofthe Paci¢c, Atlantic, and Southern Oceans average0.07 þ 0.04 nmol kg31 at the surface (6 200 m) and0.76 þ 0.25 nmol kg31 at depth (s 500 m) [13]. Suchsurface depletion is typical of `bioactive' micro- andmajor nutrients and results from biological uptake inthe upper water column followed by regenerationduring oxidation of organic detrital matter as it sinksto the sea £oor. Heterotrophic bacteria in surfaceand subsurface layers of the oceans mediate this lat-ter process [9].
FEMSEC 997 21-4-99
Fig. 1. Vertical distribution of dissolved iron and of two classesof iron-binding ligands, L1 and L2, in the central North Paci¢c.The data are replotted from [22].
P.D. Tortell et al. / FEMS Microbiology Ecology 29 (1999) 1^112

Despite its nutrient-type pro¢le, the oceanic distri-bution of Fe di¡ers signi¢cantly from that of otheressential trace metals such as Zn [17]. Most of thesemetals are highly soluble in seawater, whereas Fe ispresent to a large extent in particulate phases (sili-cates, aluminosilicates and oxyhydroxides) (see [18]).The `dissolved' Fe fraction (operationally de¢ned as6 0.4 Wm) consists largely of colloidal hydrolysis spe-cies such as Fe(OH)3 which are rapidly scavengedout of the water column by coagulation and adsorp-tion onto sinking particulate material. As a result,the dissolved Fe pool in deep waters turns over rap-idly with an ocean residence time on the order of 100years [19].
Unlike other particle reactive metals (e.g. Al, Mn,Pb) whose dissolved concentrations decrease withdepth due to removal by particulate adsorption,deep water (s 500 m) dissolved Fe concentrationsappear to be relatively invariant (Fig. 1). This anom-alous behavior is at least partially attributable to thepresence of high a¤nity organic ligands which spe-ci¢cally complex Fe(III) and increase its apparentsolubility by enhancing colloid dissolution and re-ducing scavenging rates [13]. Indeed, UV oxidationof the organic ligands in open ocean water samples,has been shown to decrease the relative proportionof truly dissolved (6 0.025 Wm), i.e. non-colloidal,Fe [20]. Several groups have examined organic Fecomplexation using highly sensitive electrochemicaltechniques [21^23]. Their results suggest that greaterthan 99.9% of dissolved Fe is bound by ligandswhich appear to fall into two classes. The strongerligand class (L1) is present in surface waters at con-centrations of 0.4^1.0 nM and has an inorganic Feconditional stability constant1 of approximately1013 M31 while the L2 class is found throughoutthe water column at approximately 1.5 nM with aninorganic Fe conditional stability constant of about3U1011 M31 (Fig. 1). As a result of this organiccomplexation, inorganic Fe concentrations are lessthan 0.1 pM (i.e. 0.0001 nM) in surface waters and
show more pronounced vertical gradients than totaldissolved Fe [22]. As discussed below, this has im-portant implications for biological Fe acquisition.
The sources and chemical structures of the Fecomplexing agents are presently unknown althoughit is clear that these organic compounds must be ofbiogenic origin. A prominent hypothesis is that theyare bacterial siderophores ^ highly speci¢c Fe-bind-ing compounds utilized for Fe acquisition [24]. Sev-eral lines of indirect evidence are consistent with thishypothesis. Laboratory cultures of oceanic hetero-trophic and phototrophic bacteria produce sidero-phores when grown under low Fe levels typical ofoceanic environments [25,26]. In the ¢eld, evidenceof in situ production of hydroxamate containingsiderophores has been obtained from coastal cyano-bacterial mats [27]. Furthermore, the conditionalstability constant of the strong organic ligand class(L1) in oceanic waters is similar to that of the wellstudied siderophore desferrioxamine B (1016:5 M31)[22]. Although the siderophores of a number of cul-tured marine strains have been isolated and studied(e.g. [28,29]), the low concentration of Fe-bindingligands in seawater precludes the precise structuralcharacterization necessary to trace their biotic ori-gins. Identifying the biological sources and chemicalstructures of organic Fe ligands in seawater will beone of the great challenges facing oceanographersover the next decade.
3. Biological Fe acquisition
If, indeed, the organic Fe chelators in seawater arepredominantly of bacterial origin, prokaryotes maybe largely controlling the availability of dissolved Feto eukaryotic phytoplankton through complexationand assimilation. Laboratory studies indicate thatcertain strains of heterotrophic marine bacteria uti-lize Fe bound to siderophores, including those thatdo not produce them [30]. At least some marine bac-teria may thus rely on Fe-siderophore complexes asthe sole Fe source in situ. Measurements of thechemical speciation of Fe during the second Equato-rial Paci¢c Fe enrichment experiment [6] showed arapid (6 1 day), four-fold increase in L1 concentra-tion that resulted in the complexation of nearly allthe added Fe [31]. As the complexation preceded the
FEMSEC 997 21-4-99
1 The conditional stability constant (KP) is de¢ned as: [FeL]/([FeP]W[LP]) ; where [FeL] is the concentration of organically com-plexed Fe, and [FeP] and [LP] are the concentrations of unbound Feand ligand respectively. KP expresses the relative thermodynamicstability of organic Fe complexes at speci¢ed values of pH andionic strength (8.1 and 0.7 M in seawater).
P.D. Tortell et al. / FEMS Microbiology Ecology 29 (1999) 1^11 3

increase in phytoplankton biomass it may have in-£uenced the community response to Fe enrichmentby a¡ecting Fe bioavailability.
The chemical speciation of Fe has been shown todetermine the extent to which it can be taken up byphytoplankton [32]. While early work suggested thatphytoplankton access only inorganic forms of Fe, wehave recently learned that diatoms are able to utilizeFe bound to a number of organic chelators includingsiderophores via a cell surface reductase mechanism[33]. Evidence of Fe-siderophore use by natural as-semblages of plankton in the subarctic Paci¢c hasalso been obtained [34]. Both autotrophic and heter-otrophic plankton are apparently able to take up Febound to desferrioxamine B and E, and large phyto-plankton reduce the Fe in the chelates extracellu-larly. The ability of indigenous plankton to utilizeFe complexed to siderophores provides indirect evi-dence that these bacterial compounds may be impor-tant in Fe cycling in situ. Very recent work [35] in-dicates that marine photosynthetic £agellates mayacquire Fe by ingesting bacteria which, as discussedbelow, constitute a large pool of particulate Fe inopen ocean waters.
4. Biogenic iron in oceanic surface waters
The majority of Fe in seawater resides in particu-late phases whose biogeochemical cycling is aspoorly understood as that of the dissolved species.Total particulate Fe (s 0.4 Wm) shows high spatialvariability and is only weakly correlated with dis-solved Fe levels [13]. Understanding the dynamicsof the particulate Fe reservoir requires informationon its partitioning between lithogenic (aluminosili-cate), detrital (non-living organic), and biogenic (liv-ing) pools. Recent data indicate that bacteria com-prise a large part of the biogenic Fe in open oceanwaters and play a critical role in Fe cycling.
Quantifying the size of biogenic Fe pools requiresestimates of biomass (mol C) and Fe requirements(commonly referred to as Fe quota, expressed as molFe cell31 or Wmol Fe mol C31) of all producers andconsumers in the ecosystem. While biomass data areavailable from several long-term oceanographic sta-tions, relatively few measurements of Fe:C ratioshave been reported for representative open ocean
plankton groups. Nonetheless, it appears that marineprokaryotes (cyanobacteria and heterotrophic bacte-ria) have a signi¢cantly higher Fe content than eu-karyotic phytoplankton. Under Fe-de¢cient cultureconditions, oceanic diatoms (eukaryotic algae) havean average Fe quota of 3.0 þ 1.5 Wmol Fe mol C31
[36] while open ocean heterotrophic and photosyn-thetic bacteria have Fe:C ratios of approximately7.5 þ 1.7 (¢ve isolates) and 19 (one isolate), respec-tively [37,38]. Some of these laboratory data havebeen substantiated by ¢eld studies at Station Papain the subarctic North Paci¢c Ocean, an o¡shore,Fe-limited region [2]. At this station, Fe:C ratios ofeukaryotic phytoplankton and heterotrophic bacteriaare 3.7 þ 2.3 and 6.1 þ 2.5 Wmol Fe mol C31, respec-tively [34,37], values similar to those measured in Fe-limited cultures. To our knowledge, no ¢eld meas-urements of Fe:C ratios of coccoid cyanobacteriahave yet been reported.
The high Fe quotas of marine bacteria, althoughpossibly surprising to oceanographers, are not unex-pected based on the biochemistry and physiology ofthese organisms. Both phototrophic and heterotro-phic prokaryotes require large quantities of Fe as aredox catalyst in their respective photosynthetic andrespiratory electron transport chains. Indeed, simplecomputation of the Fe content of a heterotrophicbacterium, derived from the concentration of Fe-re-quiring catalysts, indicates that the majority of thecellular Fe is compartmentalized in the respiratoryelectron chain (Table 1). This conclusion is sup-ported by empirical data from Corynebacterium diph-theriae [39]. More data are needed to determinewhether the Fe quotas of photosynthetic marine bac-teria are truly higher than those of their heterotro-phic counterparts.
High bacterial Fe:C ratios change our conceptualmodels of biological Fe cycling in the open ocean.Tortell et al. [37] constructed a preliminary biogenicFe budget for Station Papa by summing the Fe con-tent (biomass multiplied by Fe quota) of eukaryoticphytoplankton, cyanobacteria and heterotrophicbacteria. Their results suggested that the prokaryotesconstitute a striking 80% of the biogenic Fe in thissystem with heterotrophic bacteria alone accountingfor half of this Fe pool. We have recalculated theStation Papa biogenic Fe budget (Table 2) using 3-year average plankton Fe:C ratios recently deter-
FEMSEC 997 21-4-99
P.D. Tortell et al. / FEMS Microbiology Ecology 29 (1999) 1^114

mined for this oceanic region [34]. The result is verysimilar to that obtained previously [37]. The totalbiogenic Fe that we calculated (V17 pM) falls wellbelow the 560 pM particulate Fe previously reportedfor the subarctic Paci¢c Ocean [40] suggesting thatlithogenic and/or detrital Fe pools are large and/orthe biogenic Fe pool is signi¢cantly underestimated.
Fig. 2 illustrates our current understanding of Fecycling among biogenic pools in these waters.
Recently, Price and Morel [18] expanded the Sta-tion Papa biological Fe budget to include metazoanand protozoan grazers using Fe:C ratios [41] andbiomass estimates [42,43]. Addition of the Fe con-tained in these organisms increases the calculated
FEMSEC 997 21-4-99
Table 1Theoretical Fe requirement of Escherichia coli growing aerobically in Fe-replete medium containing yeast extract as a C substrate
Enzyme complex [Complex]a
(Wmol g protein31)Fe contentb
(atom/complex)[Fe]c (molFe/cellU10320)
Fe quotad (WmolFe mol C31)
NADH-Q reductase 0.206 2U2Fe-2S6U4Fe-4S 89.4
Succinate-Q dehydrogenase 0.335 1U2Fe-2S1U3Fe-4S1U4Fe-4S 46.7
Cytochrome b1 0.335 1 5.19Cytochrome oxidase 0.124 2 3.84Aconitase 0.018 1U4Fe-4S 1.15Superoxide dismutase 0.530 1 8.22Catalase 0.011 1 0.17Total ^ ^ 155 43Total measurede 216 61
Note that 94% of the cellular Fe is found in the respiratory chain, associated with NADH-Q reductase, succinate-Q reductase, cytochrome b1,and cytochrome oxidase complexes.aConcentrations Fe containing catalysts were obtained from published values and from measurements of catalytic activity and turnover.NADH-Q reductase was determined from the di¡erence between total £avin (0.541 nmol mg protein31 [63]) minus £avin associated withsuccinate dehydrogenase, where one £avin equivalent exists per NADH-Q or succinate dehydrogenase equivalent. Succinate dehydrogenasewas assumed to be equimolar with cytochrome b1. Cytochrome b1 and cytochrome oxidase were obtained from [63]. Aconitase concentrationwas determined from enzyme activity of E. coli (63.5 nmol cis-aconitate min31 mg protein31 [64]), turnover of puri¢ed enzyme in yeast (50Wmol cis-aconitate min31), and molecular weight (68 500 g mol31 [65]). Fe-SOD concentration was determined from enzyme activity for E.coli (5.6 U mg protein31 [66]), turnover of puri¢ed bovine Cu/Zn-SOD (3300 U mg protein31 [67]), and molecular mass of Fe-SOD (40 000 gmol31) and Cu/Zn-SOD (32 000 g mol31) [68]. Concentration of catalase was obtained from Corynebacterium diphtheridae [39].bIncludes heme and non-heme (Fe-S clusters) Fe.cIron content normalized per cell assuming 1.55U10313 g protein cell31.dIron content normalized per cellular C assuming 0.17 pg C Wm33 [69] and a cell volume of 2.5 Wm3 [70].e[70].
Table 2Iron content and steady-state Fe assimilation rates of autotrophic and heterotrophic plankton at Station Papa in the subarctic Paci¢cOcean (after [37])
Plankton group Biomass(Wmol C l31)
Fe quotaa
(Wmol Fe mol C31)Biogenic Fe(pmol l31)
Turnover rate(day31)
Fe uptake rate(fmol Fe l31 h31)
Eukaryotic phytoplankton 1.41 þ 0.49 3.7 þ 2.3 5.22 0.25 54Cyanobacteria 0.24 þ 0.16 19 4.56 0.2 38Heterotrophic bacteria 1.18 þ 0.37 6.05 þ 2.5 7.13 0.06 18Total 2.83 16.9 110
aEukaryotic phytoplankton and heterotrophic bacteria Fe quotas measured at Station Papa in the subarctic Paci¢c Ocean during threeconsecutive years (average þ S.D. [34]). Cyanobacteria Fe quotas are laboratory measurements [38].
P.D. Tortell et al. / FEMS Microbiology Ecology 29 (1999) 1^11 5
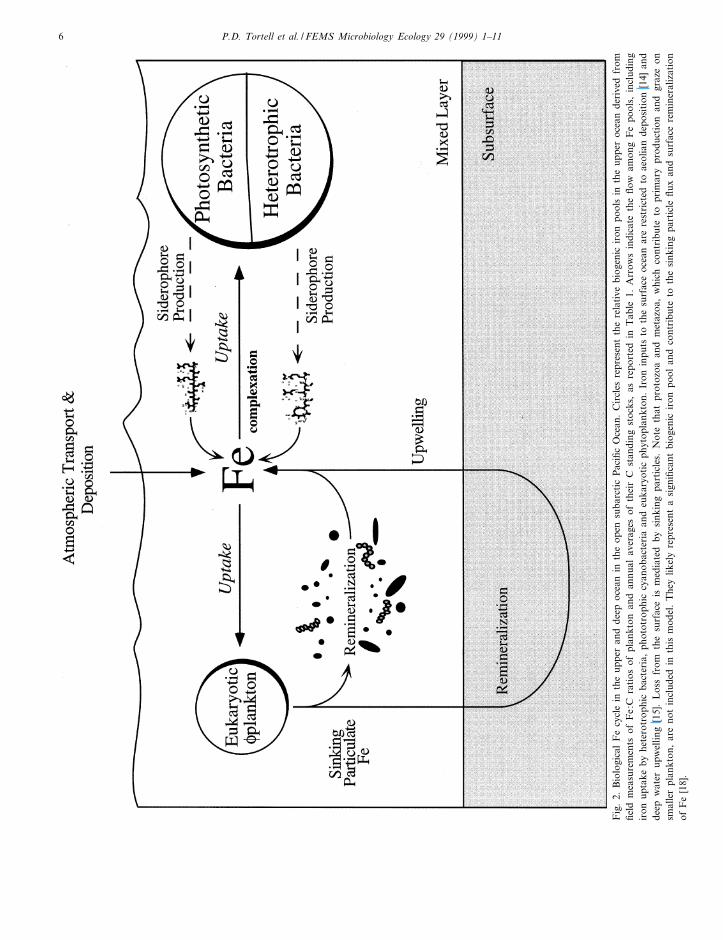
FEMSEC 997 21-4-99
Fig
.2.
Bio
logi
cal
Fe
cycl
ein
the
uppe
ran
dde
epoc
ean
inth
eop
ensu
barc
tic
Pac
i¢c
Oce
an.
Cir
cles
repr
esen
tth
ere
lati
vebi
ogen
icir
onpo
ols
inth
eup
per
ocea
nde
rive
dfr
om¢e
ldm
easu
rem
ents
ofF
e:C
rati
osof
plan
kton
and
annu
alav
erag
esof
thei
rC
stan
ding
stoc
ks,
asre
port
edin
Tab
le1.
Arr
ows
indi
cate
the
£ow
amon
gF
epo
ols,
incl
udin
gir
onup
take
byhe
tero
trop
hic
bact
eria
,ph
otot
roph
iccy
anob
acte
ria
and
euka
ryot
icph
ytop
lank
ton.
Iron
inpu
tsto
the
surf
ace
ocea
nar
ere
stri
cted
toae
olia
nde
posi
tion
[14]
and
deep
wat
erup
wel
ling
[15]
.L
oss
from
the
surf
ace
ism
edia
ted
bysi
nkin
gpa
rtic
les.
Not
eth
atpr
otoz
oaan
dm
etaz
oa,
whi
chco
ntri
bute
topr
imar
ypr
oduc
tion
and
graz
eon
smal
ler
plan
kton
,ar
eno
tin
clud
edin
this
mod
el.
The
ylik
ely
repr
esen
ta
sign
i¢ca
ntbi
ogen
icir
onpo
olan
dco
ntri
bute
toth
esi
nkin
gpa
rtic
le£u
xan
dsu
rfac
ere
min
eral
izat
ion
ofF
e[1
8].
P.D. Tortell et al. / FEMS Microbiology Ecology 29 (1999) 1^116

biogenic Fe to V38 pM with bacteria accounting for40% of the total.
The biogenic Fe budget of the subarctic NorthPaci¢c may not be representative of tropical andsubtropical oceanic ecosystems where photosyntheticprokaryotes dominate phytoplankton biomass [44]and primary productivity [45]. In these waters, bac-teria may be even more important in Fe cycling. Totest this idea, we have constructed a preliminary bio-logical Fe budget for the northern Sargasso Sea (Ta-ble 3) using published biomass data [46,47]. and lab-oratory Fe quota measurements (which include anumber of Sargasso Sea isolates [37]). A salient fea-ture of this open ocean region is the abundance ofProchlorococcus [48,49] that are not found at StationPapa. Because no Fe quotas have been reported forthese organisms and the cyanobacterial estimates arebased on a single Paci¢c Ocean strain we have chos-en upper and lower limits of 19 and 7.5 Wmol Fe molC31 for photosynthetic prokaryotes. (These valuesrepresent Fe:C ratios of cyanobacteria and hetero-trophic bacteria measured in the laboratory.) If weinclude in the budget only unicellular organisms thatassimilate Fe directly from solution, prokaryotes ac-count for 86^91% of the biogenic Fe depending onour choice of Fe:C ratios (Table 3). Consideringprotozoan grazers in the model reduces the contribu-tion of prokaryotes to 62^74% and brings the totalbiological Fe to ca. 20 pM. This value is similar tothe biogenic Fe calculated for the subarctic Paci¢cyet substantially less than the 750 pM particulate Femeasured in the northwest Atlantic [50]. Despite the
higher abundance of photosynthetic prokaryotes inthe Sargasso Sea, heterotrophic bacteria appear tocontribute substantially to the biological Fe poolconstituting between ca. 30^50% of the Fe stocksin producers (Table 3).
The above calculation does not include photosyn-thetic Trichodesmium colonies that are abundant intropical waters [51,52] and have an elevated Fe de-mand associated with N2 ¢xation [53]. Such high Ferequirements may possibly limit the abundance ofthese organisms and the extent of N2 ¢xation inthe sea [53]. As shown in Table 3, it appears thatTrichodesmium could potentially constitute the ma-jority of biological Fe in subtropical gyres containingas much Fe as all other organisms combined. Themeans by which they acquire this Fe is uncertainalthough some novel mechanisms such as utilizationof colloidal Fe have been proposed [53].
In addition to contributing substantially to stand-ing stocks of biogenic Fe, prokaryotes appear toassimilate a large fraction of the dissolved Fe.Steady-state Fe uptake rates calculated from the to-tal Fe in each biological pool and published esti-mates of mean turnover times suggest that heterotro-phic bacteria and cyanobacteria account forapproximately 20 and 30% of total community up-take respectively at Station Papa [37] (Table 2, Fig.2). These calculations are supported by direct meas-urements of size-fractionated 55Fe uptake rateswhich show that 55 þ 20% of Fe assimilation at thisstation is due to prokaryotes (0.2^1.0 Wm) [34]. Nor-malized to carbon biomass, bacterial Fe uptake rates
FEMSEC 997 21-4-99
Table 3Biogenic Fe budget for surface waters of the northern Sargasso Sea (6 200 m)
Plankton group Biomassa (Wmol C l31) Fe quotab (Wmol Fe mol C31) Biogenic Fe (pmol l31)
Eukaryotic phytoplankton 0.47 þ 0.09 3.0 1.41Cyanobacteria 0.16 þ 0.48 7.5^19 1.2^3.04Prochlorococcus 0.39 7.5^19 2.92^7.41Heterotrophic bacteria 0.57 þ 0.12 7.5 4.27Heterotrophic protozoa 0.31 þ 0.01 12 3.72Total 1.90 13.52^19.85Trichodesmium 0.12 þ 0.1 colonies l31 0.23 nmol colony31 27.6Total 41.12^47.45
aData are averages of spring and fall data as reported [46,47]. Biomass of Trichodesmium is reported as abundance in colonies l31 [51].bData for eukaryotic phytoplankton and heterotrophic bacteria are from laboratory measurements as in Tortell et al. [37]. Iron quotas forprotozoa are taken from Chase and Price [41]. For the photosynthetic prokaryotes, we have assumed upper and lower limits for Fe:C ratioscorresponding to laboratory measurements of cyanobacteria and heterotrophic bacteria. Iron content of Trichodesmium is reported per colony[53].
P.D. Tortell et al. / FEMS Microbiology Ecology 29 (1999) 1^11 7

are signi¢cantly faster, by 1.6 times, than those ofeukaryotic phytoplankton, presumably re£ectingtheir high Fe requirements for growth [37].
Spatial and temporal variability in biomass,growth rates, and Fe quotas of various planktongroups will undoubtedly in£uence their contributionsto biogenic Fe pools and assimilation rates. Maldo-nado and Price [34] found that the relative contribu-tion of bacteria and phytoplankton to community Feuptake in the subarctic Paci¢c was signi¢cantly(P6 0.001) correlated with their biomass ratios, butindependent of their production rates. In a multi-year survey of ¢ve stations, they showed that bacte-ria accounted for an average of 58 þ 23% of com-munity Fe uptake in this ocean region. Over largerspatial scales in bodies of water di¡ering greatly introphic status, Fe assimilation by bacteria appears tovary widely and correlate poorly to bacterial produc-tion or biomass [54]. Much more data will be neededto quantify and understand the patterns of biologicalFe uptake.
5. Iron limitation of bacteria in the sea
While a number of studies have examined the ef-fects of Fe de¢ciency on phytoplankton growth incoastal and oceanic waters, very little is knownabout the potential for Fe limitation of bacterialproduction. The high Fe demand of prokaryotesand the observation that Fe:C ratios of Fe-limitedcultures are very similar to those in the ¢eld suggestthat bacteria may su¡er Fe de¢ciency in situ [37].Indeed, Behrenfeld et al. [55]. have shown that theindigenous populations of photoautotrophs com-posed primarily of Synechococcus and Prochlorococ-cus are Fe-stressed in the equatorial Paci¢c Ocean,even though the biomass of these organisms does notappear to increase greatly in response to Fe enrich-ments [6,56]. E¤cient grazing by rapidly growingprotozoans seems to be at least partly responsiblefor this apparent paradox [57]. Despite high grazingpressure, the abundance of heterotrophic bacteriahas been shown to increase in Fe-amended Equato-rial Paci¢c samples [57]. This e¡ect, however, wasattributed to elevated dissolved organic matter(DOM) levels associated with the stimulation oflarge phytoplankton growth rather than Fe limita-
tion per se. By comparison, Fe enrichment of pre-¢ltered Southern Ocean water (free of phytoplanktonand grazers) signi¢cantly stimulated the growth ofheterotrophic bacterial populations suggesting thatthey are truly Fe-de¢cient [58]. It is important tonote that photosynthetic and heterotrophic prokar-yotes have unique requirements for light and DOMrespectively whose availability can partially deter-mine the extent to which they may be Fe-limited(e.g. [59]). Future ¢eld studies will need to considerthese interactions.
Tortell et al. [37]. examined the physiological ef-fects of Fe limitation on open ocean heterotrophicbacteria and the relationship between Fe and C me-tabolism in these organisms. Under Fe-de¢cient cul-ture conditions, respiratory electron transport activ-ity was reduced resulting in a signi¢cant decrease incarbon growth e¤ciencies and an e¡ective co-limita-tion by Fe and C. Recent ¢eld studies examining theresponses of heterotrophic bacteria to Fe and C ad-ditions have provided some preliminary support ofthis co-limitation hypothesis ([54]). It appears, there-fore, that Fe availability may a¡ect the pathways ofcarbon metabolism in heterotrophic bacteria and in-£uence the relative importance of the microbial loopas a `carbon sink' or `carbon link' (sensu Azam et al.[8])
6. Conclusions and future prospects
It is now becoming apparent that marine bacteriaplay a critical role in the biogeochemical cycling ofFe in the oceans. By virtue of their high, Fe-richbiomass in o¡shore waters these organisms take upand sequester large quantities of Fe thereby compet-ing directly with phytoplankton for this potentiallylimiting resource. This competition may be mediatedin part through the production of siderophores bywhich bacteria may be largely controlling Fe speci-ation and solubility in seawater.
Our current understanding of Fe-bacterial interac-tions is based on a very small amount of data. Weneed much more information on the Fe quotas of avariety of bacteria (and phytoplankton) in the labo-ratory and in the ¢eld as well as characterization ofthe Fe-binding ligands. This should provide insightinto how bacteria satisfy their nutritional Fe require-
FEMSEC 997 21-4-99
P.D. Tortell et al. / FEMS Microbiology Ecology 29 (1999) 1^118

ments and elucidate the competitive interactionsamong di¡erent plankton groups. Future studiesneed to examine the extent to which bacteria areFe-limited in situ. The use of physiological ratemeasurements [37] and molecular probes [60] willbe critical in addressing this question.
While we have focused our attention on Fe in thisreview, we believe that bacteria are likely to be im-portant in the oceanic cycling of other trace metalsas well. Most bioactive metals thus far examined arelargely complexed by strong organic ligands of un-known origin [1]. Cyanobacteria have already beenimplicated as producers of Cu-speci¢c ligands [61].Recent work suggests that limitation by trace metalsother than Fe may possibly constrain phytoplanktongrowth in the sea [62]. The extent to which this maybe true of bacteria remains to be discovered.
Acknowledgments
Franc°ois Morel, Phoebe Lam, Klaus Keller andAnne Krapiel provided helpful comments. Fundingfor this work was provided by grants from the Nat-ural Sciences and Engineering Research Council ofCanada.
References
[1] Bruland, K.W., Donat, J.R. and Hutchins, D.A. (1991) Inter-active in£uences of bioactive trace-metals on biological pro-duction in oceanic waters. Limnol. Oceanogr. 36, 1555^1577.
[2] Martin, J.H. and Fitzwater, S.E. (1988) Iron de¢ciency limitsphytoplankton growth in the north-east Paci¢c subarctic. Na-ture 331, 341^343.
[3] deBaar, H.J.W., Buma, A.G.J., Nolting, R.F., Cadee, G.C.,Jacques, G. and Treguer, P.J. (1990) On iron limitation of theSouthern Ocean: experimental observations in the Weddelland Scotia Sea. Mar. Ecol. Prog. Ser. 65, 105^122.
[4] Price, N.M., Andersen, L.F. and Morel, F.M.M. (1991) Ironand nitrogen nutrition of equatorial Paci¢c plankton. DeepSea Res. I 38, 1361^1378.
[5] Martin, J.H. et al. (1994) Testing the iron hypothesis in eco-systems of the Equatorial Paci¢c Ocean. Nature 371, 123^129.
[6] Coale, K.H. et al. (1996) A massive phytoplankton bloominduced by an ecosystem-scale iron fertilization experimentin the equatorial Paci¢c Ocean. Nature 383, 495^501.
[7] Sarmiento, J.L. and LeQueère, C. (1996) Oceanic carbon diox-ide uptake in a model of century-scale global warming. Sci-ence 274, 1346^1350.
[8] Azam, F., Fenchel, T., Field, J.G., Gray, J.S., Meyer-Reil,L.A. and Thingstad, T.F. (1983) The ecological role ofwater-column microbes in the sea. Mar. Ecol. Prog. Ser. 10,257^263.
[9] Cho, B.C. and Azam, F. (1988) Major role of bacteria inbiogeochemical £uxes in the ocean's interior. Nature 332,441^443.
[10] Delong, E.F. (1992) Archaea in coastal marine environments.Proc. Natl. Acad. Sci. USA 89, 5685^5689.
[11] Fuhrman, J.A., McCallum, K. and Davis, A.A. (1992) Novelmajor archaeabacterial group from marine plankton. Nature356, 148^149.
[12] Bruland, K.W., Franks, R.P., Knauer, G.A. and Martin, J.H.(1979) Sampling and analytical methods for the determinationof copper, cadmium, zinc, and nickel at the nanogram per literlevel in seawater. Anal. Chim. Acta 105, 233^245.
[13] Johnson, K.S., Gordon, R.M. and Coale, K.H. (1997) Whatcontrols dissolved iron concentrations in the world ocean?Mar. Chem. 57, 137^161.
[14] Duce, R.A. and Tindale, N.W. (1991) Atmospheric transportof iron and its deposition in the ocean. Limnol. Oceanogr. 36,1715^1726.
[15] Coale, K.H., Fitzwater, S.E., Gordon, R.M., Johnson, K.S.and Barber, R.T. (1997) Control of community growth andexport production by upwelled iron in the equatorial Paci¢cOcean. Nature 379, 621^624.
[16] Hutchins, D.A. and Bruland, K.W. (1998) Iron-limited dia-tom growth and Si:N uptake ratios in a coastal upwellingregime. Nature 393, 561^564.
[17] Bruland, K.W. (1980) Oceanographic distributions of cadmi-um, zinc, nickel, and copper in the North Paci¢c. Earth Plan-et. Sci. Lett. 47, 176^198.
[18] Price, N.M. and Morel, F.M.M. (1998) Biological cycling ofiron in the oceans. In: Metal Ions in Biological Systems (Sigel,A. and Sigel, H., Eds.), Vol. 35, pp. 1^36. Marcel Dekker,New York.
[19] Donat, J.R. and Bruland, K.H. (1994) Trace elements in theoceans. In: Trace Metals in Natural Waters (Salbu, B. andSteinnes, E., Eds.), pp. 247^281. CRC Press, Boca Raton,FL.
[20] Kuma, K., Katsumoto, A., Kawakami, H., Takatori, F. andMatsunaga, K. (1998) Spatial variability of Fe(III) hydroxidesolubility in the water column of the northern North Paci¢c.Deep Sea Res. 45, 91^113.
[21] Gledhill, M. and van den Berg, C.M.G. (1994) Determinationof complexation of iron(III) with natural organic complexingligands in seawater using cathodic stripping voltammetry.Mar. Chem. 47, 41^54.
[22] Rue, E.L. and Bruland, K.W. (1995) Complexation of Fe(III)by natural organic ligands in the central North Paci¢c asdetermined by a new competitive ligand equilibration/absorp-tive cathodic stripping voltammetric method. Mar. Chem. 50,117^138.
[23] Wu, J. and Luther, G.W. III (1995) Evidence for the existenceof Fe(III) organic complexation in the surface water of theNorthwest Atlantic ocean by a competitive ligand equilibriummethod and a kinetic approach. Mar. Chem. 50, 159^177.
FEMSEC 997 21-4-99
P.D. Tortell et al. / FEMS Microbiology Ecology 29 (1999) 1^11 9

[24] Winkelman, G. (Ed.) (1991) Handbook of Microbial IronChelates. CRC Press, Boca Raton, FL.
[25] Trick, C.G. (1989) Hydroxamate-siderophore production andutilization by marine eubacteria. Curr. Microbiol. 18, 375^378.
[26] Wilhelm, S.W. and Trick, C.G. (1994) Iron-limited growth ofcyanobacteria: Multiple siderophore production is a commonresponse. Limnol. Oceanogr. 39, 1979^1984.
[27] Estep, M., Armstrong, J.E. and van Baalen, C. (1975) Evi-dence for the occurrence of speci¢c iron(III)-binding com-pounds in near-shore marine ecosystems. Curr. Appl. Micro-biol. 30, 186^188.
[28] Haygood, M.G., Holt, P.D. and Butler, A. (1993) Aerobactinproduction by planktonic marine Vibrio sp. Limnol. Ocean-ogr. 38, 1091^1097.
[29] Reid, R.T., Live, D.H., Faulkner, D.J. and Butler, A. (1993)A siderophore from a marine bacterium with an exceptionalferric ion a¤nity constant. Nature 366, 455^458.
[30] Granger, J. and Price, N.M. (1999) The importance of side-rophores in iron nutrition of heterotrophic marine bacteria.Limnol. Oceanogr. (in press).
[31] Rue, E.L. and Bruland, K.W. (1997) The role of organic com-plexation on ambient iron chemistry in the equatorial Paci¢cOcean and the response of a mesoscale iron fertilization ex-periment. Limnol. Oceanogr. 42, 901^910.
[32] Hudson, R.J.M. and Morel, F.M.M. (1990) Iron transport inmarine phytoplankton: kinetics of cellular and medium coor-dination reactions. Limnol. Oceanogr. 35, 1002^1020.
[33] Maldonado, M.T. and Price, N.M. (1999) Nitrate regulationof Fe reduction and transport in Fe-limited Thalassiosira oce-anica. Limnol. Oceanogr. (submitted).
[34] Maldonado, M.T. and Price, N.M. (1999) Utilization of ironbound to strong organic ligands by phytoplankton commun-ities in the subarctic Paci¢c Ocean. Deep Sea Res. (in press).
[35] Maranger, R., Bird, D.F. and Price, N.M. (1998) Iron acquis-ition by photosynthetic marine phytoplankton from ingestedbacteria. Nature 396, 248^251.
[36] Maldonado, M.T. and Price, N.M. (1996) In£uence of N sub-strate on Fe requirements of marine centric diatoms. Mar.Ecol. Prog. Ser. 141, 161^172.
[37] Tortell, P.D., Maldonado, M.T. and Price, N.M. (1996) Therole of heterotrophic bacteria in iron-limited ocean ecosys-tems. Nature 383, 330^332.
[38] Brand, L. (1991) Minimum iron requirements of marine phy-toplankton and their implications for the biogeochemical con-trol of new production. Limnol. Oceanogr. 36, 1756^1771.
[39] Righelato, R.C. (1969) The distribution of iron in iron-de¢-cient toxin-synthesizing and in excess-iron non-toxin-synthe-sizing Corynebacterium diphtheriae. J. Gen. Microbiol. 58,411^419.
[40] Martin, J.H., Gordon, R.M., Fitzwater, S.E. and Broenkow,W.W. (1989) VERTEX: phytoplankton/iron studies in theGulf of Alaska. Deep Sea Res. 36, 649^680.
[41] Chase, Z. and Price, N.M. (1998) Metabolic consequences ofiron de¢ciency in heterotrophic marine protozoa. Limnol.Oceanogr. 42, 1673^1684.
[42] Parsons, T.R. and Lalli, C.M. (1988) Comparative oceanic
ecology of the plankton communities of the subarctic Atlanticand Paci¢c oceans. Oceanogr. Mar. Biol. 26, 317^359.
[43] Strom, S.L., Postel, J.R. and Booth, B.C. (1993) Abundance,variability, and potential grazing impact of planktonic ciliatesin the open sub-arctic Paci¢c Ocean. Prog. Oceanogr. 32, 185^203.
[44] Li, W.K.L., Subba Rao, D.V., Harrison, W.G., Smith, J.C.,Cullen, J.J., Irwin, B. and Platt, T. (1983) Autotrophic pico-plankton in the tropical ocean. Science 219, 292^295.
[45] Platt, T., Subba Rao, D.V. and Irwin, B. (1983) Photosyn-thesis of picoplankton in the oligotrophic ocean. Nature 301,702^704.
[46] Li, W.K.W., Dickie, P.M., Irwin, B.D. and Wood, A.M.(1992) Biomass of bacteria, cyanobacteria, prochlorophytesand photosynthetic eucaryotes in the Sargasso Sea. DeepSea Res. 39, 501^519.
[47] Caron, D.A. et al. (1995) The contribution of microorganismsto particulate carbon and nitrogen in surface waters of theSargasso Sea near Bermuda. Deep Sea Res. 42, 943^947.
[48] Chisholm, S.W., Olson, R.J., Zettler, E.R., Goericke, R.,Waterbury, J.B. and Welschmeyer, N.A. (1988) A novelfree-living prochlorophyte abundant in the oceanic euphoticzone. Nature 334, 340^343.
[49] Goericke, R. and Welschmeyer, N.A. (1993) The marine pro-chlorophyte Prochlorococcus contributes signi¢cantly to phy-toplankton biomass and primary production in the SargassoSea. Deep Sea. Res. 40, 2283^2294.
[50] Wu, J. and Luther, G.W. III (1994) Size-fractionated ironconcentrations in the water column of the western North At-lantic Ocean. Limnol. Oceanogr. 39, 1119^1129.
[51] Carpenter, E.J. and Romans, K. (1991) Major role of thecyanobacterium Trichodesmium in nutrient cycling in theNorth Atlantic Ocean. Science 254, 1356^1358.
[52] Capone, D.G., Zehr, J.P., Paerl, H.W., Bergman, B. and Car-penter, E.J. (1997) Trichodesmium, a globally signi¢cant ma-rine cyanobacterium. Science 276, 1221^1229.
[53] Rueter, J.G., Hutchins, D.A., Smith, R.W. and Unsworth,N.L. (1992) Iron nutrition of Trichodesmium. In: Marine Pe-lagic Cyanobacteria: Trichodesmium and other Diazotrophs(Carpenter, E.J., Capone, D.G. and Rueter, J.G., Eds.), pp.289^306. Kluwer Academic, Dordrecht.
[54] Kirchman, D.L. and Hutchins, D.A. (1998) Uptake of iron inbacteria and phytoplankton in the Ross Sea, California up-welling and coastal Delaware waters. Ocean Sciences Meeting,San Diego, CA. EOS Trans. Am. Geophys. Union 79, Suppl.
[55] Behrenfeld, M.S., Bale, A.S., Kolber, Z.S., Aiken, J. and Fal-kowski, P.G. (1996) Con¢rmation of iron limitation of phy-toplankton photosynthesis in the equatorial Paci¢c Ocean.Nature 383, 508^511.
[56] Zettler, E.R., Olson, R.J., Binder, B.J., Chisholm, S.W., Fitz-water, S.E. and Gordon, R.M. (1996) Iron-enrichment bottleexperiments in the equatorial Paci¢c: Responses of individualphytoplankton cells. Deep Sea Res. II 43, 1017^1029.
[57] Price, N.M., Ahner, B.A. and Morel, F.M.M. (1994) TheEquatorial Paci¢c Ocean: Grazer controlled phytoplanktonpopulations in an iron-limited ecosystem. Limnol. Oceanogr.39, 520^534.
FEMSEC 997 21-4-99
P.D. Tortell et al. / FEMS Microbiology Ecology 29 (1999) 1^1110

[58] Pakulski, J.D., Co¤n, R.N., Kelley, C.A., Holder, S.L.,Downer, R., Aas, P., Lyons, M.M. and Je¡rey, W.H. (1996)Iron stimulation of Antarctic bacteria. Nature 383, 133^134.
[59] Sunda, W.G. and Huntsman, S.A. (1997) Interrelated in£u-ence of iron, light and cell size on marine phytoplanktongrowth. Nature 390, 389^392.
[60] LaRoche, J., Boyd, P.W., McKay, R.M.L. and Geider, R.J.(1996) Flavodoxin as an in situ marker for iron stress in phy-toplankton. Nature 382, 802^805.
[61] Mo¡et, J.W., Zika, R.G. and Brand, L.E. (1990) Distributionand potential sources and sinks of copper chelators in theSargasso Sea. Deep Sea Res. 37, 27^36.
[62] Morel, F.M.M., Reinfelder, J.R., Roberts, S.B., Chamberlain,C.P., Lee, J.G. and Yee, D. (1994) Zinc and carbon co-limi-tation of marine phytoplankton. Nature 369, 740^742.
[63] Kim, I.C. and Bragg, P.D. (1971) Properties of nonheme ironin a cell envelope fraction from Escherichia coli. J. Bacteriol.107, 644^670.
[64] Yamamoto, I. and Ishimoto, M. (1975) E¡ect of nitrate re-
duction on the enzyme levels in carbon metabolism in Escher-ichia coli. J. Biochem. 78, 307^315.
[65] Suzuki, T., Yamazaki, O., Nara, K., Akiyama, S., Nakao, Y.and Fukada, H. (1975) The aconitase of yeast. II. Crystalliza-tion and general properties of yeast aconitase. J. Biochem. 77,367^372.
[66] Moody, C.S. and Hassan, H.M. (1984) Anaerobic biosynthe-sis of the manganese-containing superoxide dismutase in Es-cherichia coli. J. Biol. Chem. 259, 12821^12825.
[67] McCord, J.M. and Fridovich, I. (1969) Superoxide dismutase:an enzymic function for erythrocuprein (hemocuprein). J. Biol.Chem. 244, 6049^6055.
[68] Fridovich, I. (1975) Superoxide dismutases. Annu. Rev. Bio-chem. 44, 147^159.
[69] Christian, J.R. and Karl, D.M. (1994) Microbial communitystructure at the U.S.-Joint Global Ocean Flux Study stationALOHA: Inverse method for estimating biochemical indica-tor ratios. J. Geophys. Res. 99, 14269^14276.
[70] Archibald, F.S. (1983) Lactobacillus plantarum, an organismnot requiring iron. FEMS Microbiol. Lett. 19, 29^32.
FEMSEC 997 21-4-99
P.D. Tortell et al. / FEMS Microbiology Ecology 29 (1999) 1^11 11
Copyright © 2022 FDOKUMEN




















