
Translation Series No. 3558 - Fisheries and Oceans Canada ...
269
AR C HIVES FISHERIES AND MARINE SERVICE Translation Series No. 3558 The maturation and spawning of fish (Fundamentals and applications) Takashi Eibiya, et al. (Editors) Eleven papers by various authors from a symposium of the Japanese Society of Scientific Fisheries, April 5, 1974 Original title: Gyorui no seijuku to sanrar. From: Nippon Suisan Gakkaishi suisangaku shirizu 6 Koseisha Koseikaku, Tokyo, pp. 1 - 130, October 15, 1974 Translated by the Translation Bureau(JWC) Multilingual Services Division Department of the Secre_ary of State of Canada Department of the Environment Fisheries and Marine Service Vancouver Laboratory Vancouver, B.C. 1975 263 pageu typescript
-
Upload
khangminh22 -
Category
Documents
-
view
5 -
download
0
Transcript of Translation Series No. 3558 - Fisheries and Oceans Canada ...

ARCHIVES
FISHERIES AND MARINE SERVICE
Translation Series No. 3558
The maturation and spawning of fish (Fundamentals and applications)
Takashi Eibiya, et al. (Editors) Eleven papers by various authors from a symposium of the Japanese Society of Scientific Fisheries, April 5, 1974
Original title: Gyorui no seijuku to sanrar.
From: Nippon Suisan Gakkaishi suisangaku shirizu 6 Koseisha Koseikaku, Tokyo, pp. 1 - 130, October 15, 1974
Translated by the Translation Bureau(JWC) Multilingual Services Division
Department of the Secre_ary of State of Canada
Department of the Environment Fisheries and Marine Service
Vancouver Laboratory Vancouver, B.C.
1975
263 pageu typescript

DEPARTMENT OF THE SECRETARY OF STATE
TRANSLATION BUREAU
MULTILINGUAL SERVICES
e«Ï CANADA
DIVISION MULTILINGUES
SECRÉTARIAT D'ÉTAT
BUREAU DES TRADUCTIONS
DIVISION DES SERVICES
TRANSLATED FROM — TRADUCTION DE INTO — EN
Japanese EnD:lish
PLACE OF PUBLICATION LIEU DE PUBLICATION
Tokyo
PUBLISHER— ÉDITEUR
Koseisha Koseikaku
t 974 ctober
•DATE OF REQUEST DATE DE LA DEMANDE 01-04-75
-
•.:Y1 7/)
AUTFIOR — AUTEUR Nihon suisan gakkai (hen)
Japanese Society of Scientific Fisheries (edited). TITLE IN ENGLISH — TITRE ANGLAIS
The maturation and spawning of fish.
TITLE IN FOREIGN LANGUAGE (TRANSLITERATE FOREIGN CHARACTERS) TITRE EN LANGUE ÉTRANGÉRE (TRANSCRIRE EN CARACT1RES ROMAINS)
Gyarui no seijuku to sanran.
REFERENCE IN FOREIGN LANGUAGE (NAME OF BOOK OR PUBLICATION) IN FULL. TRANSLITERATE FOREIGN CHARACTERS. RÉFÉRENCE EN LANGUE ÉTRANGÉRE (NOM DU LIVRE OU PUBLIl ATION), AU COMPLET, TRANSCRIRE EN CARACTaRES ROMAINS.
Nihon suisan gakkai - suisangaku shirizu 6. • REFERENCE IN ENGLISH — RÉFÉRENCE EN ANGLAIS
Japanese Society of Scientific Fisheries, Fisheries Series 6.
YEAR ANNÉE
15 0
DATE OF PUBLICATION DATE DE PUBLICATION
VOLUME ISSUE NO.
NUMÉRO
PAGE NUMBERS IN ORIGINAL NUMÉROS DES PAGES DANS
L'ORIGINAL
1 7 130 NUMBER OF TYPED PAGES
NOMBRE DE PAGES DACTYLOGRAPHIÉES
26 3 (ici. p
REQUESTING DEPARTMENT MI NISTÈR E-C LI ENT Environment TRANSLATION BUREAU NO. 1101002
NOTRE DOSSIER N 0
TRANSLATOR (INITIALS) JWC DIRECTION OU DIVISION TRADUCTEUR (INITIALES) BRANCH OR DIVISION Fisheries & Marine _
OCT 2 1 1975 PERSON REQUESTING Allan T. Reid DEMANDÈ PAR
YOUR NUMBER VOTRE DOSSIER N 0
UNEDITED TRANSLATION For information only
TRADUCTION NON REVISEE. 'Information seulement
SOS•200.10.6 (RE V. 2/øI) 7630.21-029.5333

DEPARTMENT OF THE SECRETARY OF STATE
TRANSLATION BUREAU
SECRÉTARIAT D'ÉTAT
BUREAU DES TRADUCTIONS
DIVISION DES SERVICES
MULTILINGUES
mULTILINGUAL SERVICES
DiVISION
III
'22)
, I • ! ; s •
DERARTaiNT . DIVISION/GRANCH CITY
I MINISTRE DIVISION/DIRECTION . VILLE
Environment Fisheries & i.arine Ottawa, Ont.
LANGUAGE TRANSLATOR (INITIALS) LANGUE TRADUCTEUR (INITIALES)
.O Jn ,.)rese JWC
CT 2 1 19 75
Scientific Fisheries, No. 6.
THE MATURATION AND SPAWNING OF FISH.
(Fundamentals and applications).
The Jah -mose Society of Scientific Fisheries.
Published by Koseisha Koseikaku.
UNEDITED TRANSLATION For information only
TRADUCTION NON REVISEE Information seulement
101 .-200..1 0..31

Yutaka MURAKAMI.
Reijiro HIRANO.
Kazunori TAKANO.
Sadaichi KATO.
Fumio-YAMASAKI.
Hiroshi YOSHIOKA.
Teruo HARADA.
Quality of the Y. The assessment of the spawn.
10. Freshwater fish.
11. Marine fish.
Kiyoshi SAKAI.
Michiyasu KIYONO.
2 ••
Contents and Authors.
Foreword , Takashi HIBIYA, Minoru NOMURA, Yutaka MURAKAMI
and Reijiro HIRANO.
I. Present condition and uncertainties.
1. Freshwater fish.
2. Marine fish.
II. Parent fish for egg collecting.
3. The process of maturation of the gonads.
4 • Detailed characteristics of reproduction.
5. Internal secretions, egg maturation and ovulation.
III. Environment, maturation, and spawning.
6. Freshwater fish.
7. Marine fish.
IV. Maturation and metabolism.
8. Maturation and fat metabolism. Fumio TAKASHIMA.
9. The accumulation of yolk protein. Katsumi AIDA.
General discussion.
Takashi HIBIYA, Minoru NOMURA, Reijiro HIRANO.

3
Foreword.
An increased rate of reproduction of fish is of
fundamental importance in the maintenance of continued
stability of the fishing industry, and particularly important
in the planning of any increase of production. Formally, of
course, this is a matter of pisciculture, and active efforts
have recently come to be concentrated on the production of
fry for the cultivation and stocking of the offshore fish
resources.
There are many questions concerned with the production
of fry, such as the food required in the early stages, the
control of the environment and of disease, but the very first
step is the procurement of good quality eggs from superior
parent fish and the preservation of these eggs in a suitable
hatching place to produce the required number of healthy fry.
There have been very few studies of the culture of the parent
fish from the point of view of breeding, or of the feeding of
the parent fish from which eggs are to be collected. The
methods of collecting eggs have a history reaching back to
the last century, and a great deal of basic research has been
accumulated in relation to the maturation of the gonads, the
acceleration of maturation, and the induction of spawning.
There has also been a very large amount of research on the
life history of the fish from the formation of the eggs to
the hatching of the fry.

However the present discussions show that the results
so far obtained by basic research are by no means sufficient
for practical application in the collection of eggs. This is
because basic research is apt to be limited to a narrow range
of phenomena of an essentially scientific ichracter, and it
is considered that there has not been sufficient recognition
of the mutual stimulation which can accompany the expansion
of both basic research and applied technology.
The Japanese Society for Scientific Fisheries therefore
arr.Lnged a symposium, held on 5 April 1974, entitled "Questions
relating to the collecting of eggs and to the parent fish
used for the production of fry". Discussion during this
symposium centred on the female fish and dealt with the
relation between basic knowledge and practical technique in
the collection of eggs.
The present publication is the record of this
symposium, and is the joint work of many contributors.
Nothing can give more pleasure to all those concerned than
to see it contribute to both scientific and technical
progress in this field.
Takashi HIBIYA (Department of Agriculture, Tokyo University)
Minoru NOMURA (Tokyo Maritime University)
Yutaka MURAKAMI (Department of Aquatic Animal Husbandry,
University of Hiroshima)
Reijiro TAUANO (Department of Agriculture,
Tokyo University).
•

The maturation and siawnincr of fish.
(Fundamentals and applications).
Index.
Foreword. Takashi HIBIYA, Minoru SATOMURA, Yutaka MRAKAMI
and Reijiro HIRANO.
I. Present conditions and uncertainties,
1, Freshwater fish
2. Marine fish.
Page
Yukata MURAKAMI 9
Reijiro HIRANO 20
II. Parent fish for eg collecting.
3. The process of maturation of the
gonads. Kazunori TAKANO 32
1. The structure of the ovary and the oviduct.
2 . Genital cavity fluids. 36
3. The process of ripening of ovarian ova. 39
4 . Ovulation.
5. The functions of ovary hormones. 47
4. Detailed characteristics of
reproduction. Sadaichi KATO 58
1. The ripe eggs. 58
2, The gonad index of mature individuals
and the size distribution of eggs
in the ovary. 59
3. The ripe eggs. 62
4. The weight of an egg. 64
5. The spawning season. 69
33

• 6
5. Internal secretions, egq maturation and ovulation. Fumio YAIUSAKI 76
1. The maturation of the egg. 76
2. Ovulation. 85
3. Piscine GTH. 90
Environment, maturation, and spawning.
6. Freshwater fish. Hiroshi YOSHIOKA 105
1, Illumination. 105
2, Temperature. 117
7. Marine fish. Teruo HARADA 128
1. Water temperature, maturation
and spawning. 129
2. Illumination and spawning. 134
3. Other environmental factors,
maturation and spawning. 135
4. The culture of the parent fish
for egg collecting. 136
5. The stimulation and inhibition
of maturation and spaWning by
environmental control.
6. Future prospects. 142
IV. Maturation and metabolism.
8. Maturation and fat metabolism. Fumio TAKASHIMA 152
1. The chemical properties of
fish egg fats. 153 .
2. Changes in body fat content
during maturation of the
ovary. 156
138

•
•
208
211
212
2 15
3. Endocrine control of changes in
fat metabolism during Maturation
of the ovary. 159
4. Fats in the diet of the parent fish. 163
9. The accumulation of yolk
protein. Katsumi AIDA 176
The appearance and endocrine •
control of female specific
proteins during maturation.
2. The connection between FSSP and
yolk protein. 181
3. The site of synthesis of FSSP. 183
4. The introduction of FSSP into
the maturiag egg. 191
•V. The evaluation of egg quality in spawn.
10. Freshwater fish. Kiyoshi SAKAI 200
1. The Quality of the spawn and
its internal and external
morphological characteristics. 202
2, Change of egg quality with time. 208
3. Hormone administration and the
process of maturation ofthe
ovarian egg.
4. Assessment of the Quality of eggs
spawned near the upper limit of
spawning temperature.
5. Egg quality and egg fractionation
by centrifuging.
6. Egg quality and other criteria.
1.
17 8

• 8
Michiyasu KIYONC 225
1. The characteristics used in
determining egg quality. 226
2. Discussion of methods of
appraisal of egg quality. 232
Questions.
General discussion. 246
•
11. Marine fish.
238
•

• 9 Present condition and uncertainties.
1. Freshwater fish. Yutaka MURAKAMI.*
In discussions of pisciculture and the stocking of
lakes, marshes, and rivers, it is usual to start with the
preservation of the fry. Ideally, the parent fish are
cultured in a pond, the eggs are artificially collected and
fertilized and healthy fry are reared to the required size
so that the production of the quantity needed to meet the
demt.nd can be planned. However methods which will come close
to guaranteeing fry production in this way are known for only
a few species. They are limited to rainbow trout Salmo
gairdneri irideus, Salmo macrostoma, Oncbrhynchus rhodeus,
the carp Cyprinus carpi°, the crucian carp and the goldfish
Carassius auratus.
For the cultivation of such fish as the eel Anguilla
japonica, the sweet smelt Plecoglossus altivelis, and of
Ctenopharyncodon idellus and Hypophthalmichthys molitrix,
it is normal to rely mostly, or even completely, on young
fish or eggs which have been naturally produced. It has
recently become difficult to obtain sufficient quantities of
these fish to meet the demand because the number of natural
fry is diminishing, and there has been a great deal of
experimental study of fry production through artificial egg
collection. In one such experimental study, made with
P. altivelis, it was possible to produce the large quantity
of approximately 10 million fry for stocking.
(Tokyo University of Fisheries).

1 0
In present day production of freshwater fish fry both
cultivated and wild-caught parent fish are used. In some
species, hormone administration is needed for artificial egg
collection. There are two methods of artificial egg collection.
In one method, which is used with carp, goldfish and bluegills,
mature male and female parent fish are kept in the same pond
and they spawn naturally in nests in a man-made spawning bed.
In the second or "stripping" method, the parent fish are
inspected and when it is judged that they have ovulated the
eggs and mut are taken from the body either by pressure on
the abdomen (the squeezing out method) or by cutting (the
incision method). We will here refer to the first method as
the "carp" method of collection, and to the latter as the
"rainbow trout" method.
Table 1.1 shows the methods of egg collection from the
parent fish normally used for the production of fry of the
species most often stocked or cultivated. The table also shows
those which are in the experimental stage. It shows that
various experimental studies are in progress with species for
which the production of fry by artificial egg collection has
not yet been commercially established. The following summary
of present conditions and uncertainties about the production
of fry from parent fish is limited to the topics of parent
fish and egg collection. •

Table 1.1.
Parent fish and methods used for the collection of eggs of freshwater fish.
Parent fish used Cultivated Wild Eggs or fry ob-tained
Hormone use Not used Used Not used Used from the . wild
Method of collection* Rainbow Carp Rainbow Carp Rainbow Carp Rainbow Carp trout type trout type trout type trout type type type type type
Salmo gairdneri X
Salmo macrostoma X Oncorhynchus rhodurus -
Cyprinus carpio X
Carassius carassius X
Carassius auratus X
Lepomis macrochirus X
- Basilichthys bonariensis
Hi lmesus transpacificus X. 1 i..
li Y Y X

. , • Hypomesus transpacificus X . • . ; '
; Plecoglossus altivelis Y . Y Y X
L ,
: Misgurnus anguillicaudatus Y Y , y Y
Anguilla japonica v y X .
Gnathopogon elongatus I Y ' y .
,
Tribolodon hakonensis Y Y Y i X
Ctenopharyncodon and Y . y Y Y X Hypophtalmichthys
Parasilurus azotus Y Y
Zacco platypus Y
Black bass
X: Methods . most usually employed commercially.
Y: Methods in experimental use or in process of transfer to commercial use.
See page 10 of the present article. 1-■ • b-‘
}o^,r

12
In species in which the collection of eggs from
cultured fish is difficult, use is made of artificial
collection from mature parent fish which have assembled
in their r.latural spawning places, and of immature fish which
are approaching the age of natural maturation. Eggs can be
collected from some fish immediately after capture, and others
can be cultured and brought to complete maturation by the
administration of hormones, after which the eggs can be
collected. At the present time the list of such fish, taking
account of those in the experimental stage, includes the
eel Anmilin_inmni.22, the sweet smelt Plecoglossus altivelis,
the loach Misgurnus anguillicaudatus, the roach Gnathopogon
elongatus caerulescens, the dace Tribolodon hakonensis
hakonensis, the Chinese carps Ctenopharyncodon idellus and
Hypophtalmichthys molitrix, the pond smelt Hypomesus
transpacificus nipponensis, the freshwater catfish Parasilurus
azotus and Zacco platypus. When fish captured in the wild
are used, and more particularly if they are captured already
mature at the spawning places, it is known that the results
of egg collection may be greatly influenced by the method of
transportation from the place of capture to the place of
collection, and by the length of the period of cultivation
between capture and collection. For example, with
Ctenopharyncodon and Hypophthalmichthys the results of egg
collection are increased if the fish are anaesthetized during

• 13
transport. Good eggs can be obtained from Misçurnus if
hormones are administered immediately after capture, but
the yield from egg collection is lowered if there is a long
interval.. Good results can be obtained with Tribolodon if
the eggs are collected immediately after capture, but if they
are kept for as long as 24 hours, the eggs become difficult
to extract, and the yield is lowered.
If the required quantity of parent fish can easily be
caught and the price is not high, there is little difficulty
in the use of wild fish. However when, as has recently become
the case with the production of fry of P. altivelis, the
demand becomes large it becomes difficult to maintain a
sufficient quantity of parent fish to meet the demand for this
large production. The price then rapidly rises and it is
better to rely on artifial collection of eggs from parent
fish cultivated in ponds.
With present day techniques of pond cultivation, the
parent fish can be divided into three groups according to the
relative difficulty of attaining maturation and egg collection.
(1) Species which can easily be brought to the size and
age for maturation, and can be matured and made to produce
eggs by the normal methods of pond feeding and rearing.
Examples are Salmo gairdnerii, Salmo macrostoma, Oncorhynchus
rhodurus, Cyprinius _carpi°, Goldfish, Carassius auratus, the
Leyomis macrochirus and the pejerry
(Odontesthes) bonariensis.

• 14
(2) Species in which parent fish can be reared to become
mature even without hormone treatment, although in many cases
hormone treatment is necessary at the time of egg collection,
and the results of egg collection are inferior to those
obtained from wild fish. Examples are Plecoglossus altivelis,
Misgurnus anguillicaudatus, and Tribolodon hakonensis hakonensis.
(3) Species which in cultivation can reach the age and pi°
size of maturation of wild fish, and which can be made to
mature and to produce eggs by the administration of hormones,
but with which this is very difficult. Examples are Anguilla
jamoniu, Ctenopharyncodon idellus and Hypophthalmichthys
molitrix.
Species of the first type •are by no means lacking in
problems with regard to rearing and egg collection, but on the
whole it is those of types 2 and 3 Which present the most
difficulties.
Among the problems which these species have in common,
though to different extents, are
(a) The proportion of the cultured fish from wIlich eggs
can be collected is low.
(h) Only a fraction of the ripe eggs can be collected,
or if all the eggs can be collected, the fraction which
develops is low.
(c) The condition of the eggs ovulated or spawned may
vary from one parent fish to ;another, or even between eggs from
the same parent, so that the results of collection are poor.

15
(d) The fraction which die during development or hatching
is high, or the fraction which are abnormal or feeble when
hatched is high, so that it is difficult to get healthy fry.
The conditions in which the parent fish are raised and
the methods of egg collection are factors in these difficulties.
The environment in which the parent fish are raised
influences maturation and ovulation, and the principal
environmental factors normally considered are illumination and
temperature. The relations between these factors and the
maturation and ovulation of freshwater fish have recently been
clarified, and techniques have been developed whereby fish
maturation and spawning can be controlled by me=s of
artificial variations of temperature and illumination. Using
these methods with goldfish, S. gairdnerii, O. rhodurus,
S. macrostoma, and P. altivelis, it has become possible to
make them spawn outside the natural spawning season, and this
can be of practical utility in the production of fry. When
illumination and temperature are within an approximately
limited range, normal maturation and spawning occur, but when
they are close to the limits, maturation and ovulation may be
greatly hindered. Temperature differs from illumination in
that there may be great differences between different places
of rearing, and it is particularly necessary to take note of
this. The upper limits of temperature at which there is some
hindrance of maturation and ovulation in S.J2,-airdnerii,

• 16
•
•
Cyprinius carpio and P. altivelis have recently been
experimentally determined, and it is considered to be important
to establish such temperature limits for the rearing of parent
fish of many species.
The nutrition of the parent fish (the quality and
quantity of feed) not only influences the maturation of the
fish with regard to the amount of spawn and the quality of
the eggs, but also controls the growth of the fish, and
there is a close relation between growth and the maturation
of the gonads. It is known that egg collection is easy and pll
that good quality eggs can be obtained from P. altivelis if
it is reared in uncrowded conditions. The good results of
egg collection are due to the fact that they ingest not only
the artificial food, but also large quantities of diatoms
from the edges and the bottom of the pond. • Experimental
studies have recently been made of the nutrition of several
types of freshwater fish such as S. gairdnerii and P. altivelis,
but many of them have not obtained any definite results. One
reason for this which can be offered is as follows. The
assessment of the quality of the feed is made by means of many
criteria such as the fraction of the experimental fish from
which eggs can be collected, the quantity of eggs collected,
the fraction of eggs laid which are fertile, the fraction
which hatch, the health of the fry after hatching, and the
fraction of the parent fish which survive spawning. These

•
•
• 17
criteria can be influenced by many factors other than the
quality of the feed occurring during the process of rearing.
These may include individual differences in the reproductive
ability Of the experimental fish, and variations in the
techniques of egg collection, of fertilization and of
incubation, and these may introduce such a range of error
that the experiments are difficult. It will be necessary to
establish satisfactory methods of assessing the quality of
the eggs after laying, of measuring the reproductive quality
of the parent fish, and of standardizing the methods of
egg collection.
There have been many experimental studies of the
artificial collection of eggs by means of the administration
of hormones, and general techniques are being established by
experience. However with some species or in some states of
maturation the reproductive efficiency may be low, and there
are many circumstances in which the administration of hormones
may be considered to be a factor in the death or damaged
health of the parent fish.
In some fish, eggs are to be collected by the "rainbow
trout" method and it may be found that ovulation occurs a
number of times. Since the hormone is thought to have a bad
influence on ovulation and on the health of the parent fish,
it is difficult to determine the appropriate time for egg
collection. In many species, satisfactory results are obtained

• 18
by the collection and fertilization of the eggs immediately
after ovulation. In S. gairdnerii a satisfactory development
ratio of more than 80% is found up to 10 days after ovulation,
but with species in which the temperature best suited to
rearing is high, the fertilizability of the eggs drops off
shortly after ovulation. In such species, the "carp" method
of collection gives better results than the "rainbow trout"
method. In the planned production of fry, the "rainbow trout"
method is superior to the "carp" method. However, in general,
the "carp" method gives satisfactory results with many of the
fish species under discussion. This is because there is
gl› • little increase in artificial handling in the 'carp" method,
and ovulation and spawning are allowed to occur in the natural
way. It is known that the water flow, the water temperature,
the physical and chemical properties of the bottom, and the
mutual stimulation between the sexes all take part in ovulation
and spawning, and all of these are used in the "carp" method.
However it is considered necessary that the relations between
these external stimuli and ovulation and spawning should be
better known, so that they can be applied positively to the
production of fry.
• We have here mentioned a number of the problems
involved in the production of freshwater fish fry, particularly
those concerned with the rearing of the parent fish and the
techniques of egg collection. In order to solve these problems,
p12

•• • 19
it will be of importance to study the process of maturation
of the eggs, the mechanism of ovulation (and its induction),
the connection between nutrition and metabolism of the parent
fish, and the relation of these to the environment in which the
fish are reared. It is necessary to plan for many stages of
study from basic research to practical application so
that the results may be used as a foundation for the rearing
of parent fish and for egg collection. It is also considered
that increasing importance should be given in future to studies
of genetic breeding in order to utilize parent fish of valuable
reproductive quality.
•

• 20
•
2. Marine fish. Reijiro HIRANO*. p13
Studies of the production of marine fish fry have
gradually increased since the last half of the 1950s. At
first, they relied on the collection of eggs from fish caught
in the wild. This method of egg collection involved very
strenuous efforts, and it was very soon recognized that it
was necessary that marine parent fish should be cultured.
Since the beginning of the 1960's progress has been made in
the techniques of culture of marine fish, and as the culture
industry has spread it has become possible to collect eggs
from a number of species. This is at present being done with
the species shown in ?able 2.1. However there are still many
basic questions to be answered about the effects of feed and
environmental conditions on the spaWning of marine fish, and
it is to be hoped that there will in the future be further
fundamental research.
However it will continue to be convenient with some
species or in some regions to keep wild-caught fish for a p14
short period before collecting the eggs, and it is important
that progress should be made in studies in this area. At the
present time, two methods are used for collecting eggs from
marine fish. They are.the so-called "artificial collection"
method, using pressure on the abdomen, and the "natural spawning
collection" method. The "artificial" method may be used to
* Department of Agriculture Science, University of Tokyo.

•
•
21
Table 2.1.
Examples of the collection of eggs from cultured parent fish.
Species Research agency ' Culture facility
Chrysophrys Hiroshima maritime Shore pond (75t) major experimental station Small
Oita maritime Small experimental station
Kanagawa maritime Embankment, 8400m3 experimental station
Tokushima maritime Aquarium (100t) experimental station
Kumamoto maritime Small experimenta7. station
Ishikawa Small
Mie, Owase maritime Shore pond 12.5mi expelimental station
_ . Kinki University Small
. Tokyo University • Shore pond, culture
' pond
Mylio macro- Hiroshima maritime Shore pond (75t) cephalus experimental station
Aichi maritime Shore pond (75t) experimental station
Nagasaki nui7ritional Small laboratory
Kinki University Small
Tokyo University Shore pond, culture pond
Rhabdosaurus Shizuoka maritime Small sarba experimental station
•

• 22
•
Table 2.1 (continued)
Oplegnathus Kumamoto maritime Small fasciatus experimental station
Nagasaki maritime Small . experimental station
Wakayama maritime Small ._ _ _ _ _ experimental - station , _ _
Kinki University Small
Erynnis Kinki University Small . -j,aPanica
Oplegnathus Kinki University Small _ . _ 'plunctata
'Parapristipoma Oita maritime Small 1•- .trilineaturi - --6kpériderital station
1 . _ , Tokushima maritime -e.. (111ari-um (100t)
- experimental station
lac. rack- Oita maritime Small - -- -Ugh - ' experiMental station ._ , ,
nexagrammas Hyoga maritime • q_P-917P pond otaki experimental station
Seriola Nagasaki maritime Small _ quinquera7 experimental station ._ diata
Kochi maritime Small experimental station
Mie Owase maritime Small experimental station
Kinki University Small
Seriola Kinki University Small aureovattata
Seriola Kinki University Small purpurascens

• 23
•
•
obtain eggs from fish caught in the wild, but the period which
is usable for fertilisation is believed to be short, and many
satisfactory results have been obtained with the "natural"
method in species which lay separately floating eggs1 .
When cultured fish are used as parents, it is usual
to employ the "natural spawning" method of egg collection.
To do this the male and female parent fish are housed in the
same tank, the spawn and the fertilized eggs are collected
and used for rearing. The ratio of males to females for this
purpose is usually about unity, but further investigation on
this point is required for each species. With appropriate
environment and management of the rearing, there will be little
difficulty if the place for rearing and the place for egg
collection are the same, but when the parent fish have to be
moved from the place in which they were reared to some other
place for egg collection, or when fish caught in the wild are
cultured for egg collection, consideration must be given to
the following points.
Careful attention must be given to the method and
season of capture and transport of the fish, and also the size
and construction of the tank used for collection. In general
a round tank gives more satisfactory results than a square
tank of the same surface area. In these circumstances, it is
also often effective to use hormones which stimulate the
gOnads, but when hormones are used it is evidently important

24
to consider the type of hormone to be used and its relation
to the state of maturation of the parent fish, and also the
quantity and time for its administration.
If the fish are to be removed from a roomy place of
rearing to a narrow tank for egg collection, the hormone
must be used as soon as possible after the nove. It has
often occurred - with Mylio macrocephalus that the hormone
became ineffective after the lapse of a week. It is believed p1 .5
that this is strongly connected to stress, and further studies
are considered to be needed. •
The hormone which is at present generally used is '
synaholin, but the studies of the relation betwem the state
of maturation of the fish and the amount of hormone to be
administered to a fish of a given weight are not yet complete.
Many experiments have been made in Which 4o to 120 R.U. of
synholin per kilogram of fish body weight are injected into
the dorsal muscles, but there is scope for further investigation.
It is also to be remembered that when wild-caught
fish are to be used, they may not be sufficiently mature when
caught, and it is often found that the administration of
hormones is then completely ineffective.
Another important point to remember with marine fish
is the so-called "successive spawning". A number of studies 2
have already made it well known that in Chrysophorus major
the same parent fish will spawn a number of times. Hibiya et al3

• 25
Table 2.2.
Collection of eegs from mylio microcephalus.
(Hibiya and Sato, 1973).
Date. 1 1 y 2 July 3 July 3 July 4 July 5 July
'Experiment number
B - 2 4o 30,00 0 60 ___ Few 100,000 units units
H - 2 syna- 500,000 syna- --- Few 400,000 holin holin
3 in- in- •600,000 --- 400,00 0 jected jected
7 300,00 0 --- 700,000
8 100,00 --- Few
Table 2.4
(Hibiya, Sato, 1973). DATE t
TEsT. tio • t i 12. 7 12. 10 12. 11 12. 11 12. 12 12. 13 12. 14
1 • 43g• • • 42g —g —.—g ---1;
2 20 40 -:-- 40 — 43• UNITS 'UNITS - 3 SYNA•• — SYNA -• — 15 26
HOE I A HOL IN 43 .71 . 4 INJECT
65 INJECT
5 En o ■ -7-- ED., _____ 66
29 —
6 • _____ '
_____ 52 31 —
g. This note is nowhere explained. (Translator).
•

• • •
Table 2.3
Number of ova spawned by Sillago sihama. (Sato, Hibiya, Kiyono and Hirano, 1973).
Test fish Date and Expti time of First Second Third Fourth
!Female Male hormone Injection night night night night ! injection 1 1
10 July 72 30 to 40 40 to 50 10 to 20 1 5 (1800) Syn 40 thousand thousand thousand
2 ; 5 i 12 July it 50 to 60 50,000 20,000 i (1800) thousand 60,000
•
3 1 3 2 13 July t, 100,000 10,000 1,000 1 (evening)
3 2 • 4 August ti Small 70 to 80 5,000 2 to 3
(1800) quantity thousand thousand
2 4 8 August vi 100,000 40 to 50 (1600-1700) thousand
i
2 3 " " " Syn 100 40 to 50 10,000 thousand
7 2 3 15 August 10,000 80,000
(1400) Syn 40 , I

2 7
have recently shown that when eggs are collected in a tank,
Mylio màcrocephalus, Sillago sihama, and Kareius bicoloratus
show successive spawning. (Tables 2.2, 2.3, and 2.4). It
will therefore be necessary to make further studies of each
species from which it is proposed to collect eggs by means of
the administration of hormones, in order to determine whether
or not their normal spawning habit includes "successive pU;
•
•
spawning". It has been found that when hormones are administered
to Limanda yokohamae "successive spawning" does not occur.
As is of course to be expected, it is difficult to
obtain a large number of eggs by means of pressure on the
abdomen in species in which successive spawning occurs. It
is with such species that rearing of the parent fish is
particularly important.
It has been shown with freshwater flsh that the
external environment, in particular the illumination and the
temperature, are important in the control of maturation
and spawning.
There has been little study of this in relation to
marine fish, but studies of the early egg collection from
Chrysophorus major through the use of warm water have been
in progress at the Mie-Owase marine experimental station p i7
since 1972. It was found that by keeping the water temperature
at 19° C to 20 °C during the winter of 1972 to 1973, it was
possible to maintain normal mature condition for 65 days

• 59
28
Table 2.S ••
Spawning by Chrysophrys major.
(Owase experimental station 1973)
Test Spawning period Number Total Ova per Ova per group of ova fish fish per
females day
(x104 ) (x104 ) (x104 )
,
,
A : 15 Feb - 12 Apr 6* 1662.5 305 5.6 (warmed
B 31 Mar - 15 Apr 5 254 50.8 3.2 (control)
*. Including one which died on 18 March.
Figure 2.1.
Spawning by Chrysophrys major.
(Owase experimental station 1973)
12 1 2 3 4 Date 1973
›
cd 501
0 4
CL..• . 3
NH 2 Q) ><
e n
10/11 20 inu io 2'0 . io L I:1111 Date.
Figure 2.2.
Water temperature during the period of culture.
Water temperature for group A.
0---• Water temperature in the test station rearing facility.

•
2 9
after 15 February. This was one month sooner than in the
control group, and the number of eggs produced per fish was
3 million, which was 6 times the number from the control group.
These experiments have shown that as with fresh-
water fish, the temperature of the water is a very important
factor of the external environment for marine fish. However
we still have insufficient information about the period when
the temperature should be increased and about the limits of
suitable temperature which will result in early spawning.
Further basic studies of this will be necessary.
There is also an intimate connection between the age
of the parent fish and the results of egg collection, and
from what little is known it is clear that there is much
which awaits further study. Individual future studies which
are considered important include the conditions in which the
parent fish are to be reared, the fodder to be given, the
acceleration or stimulation of maturation by the administration
of sex hormones 3 and the physical and chemical properties of
the eggs, especially those which float.
•

References.
1. Reijiro HIRANO.
Kurodai no chigyo shi iku.
Nichi sui shi .15 567 - 569 (1969).
Rearing of the young of the black porgy,
Mylio microcephalus.
Bulletin of the Japanese Society of Scientific
Fisheries, 15 567 - 569 (1969).
2. Hiroshimaken suisan shiken jo.
Madai no oyauo yosei ni kansuru kenkyu.
Showa 46 nendo shubyo seisan kenkyu hohokusho 1971.
Hiroshima prefectural maritime test station.
A study of the nurture of red sea bream çnEunrima2
• major.
Reports of research on the production of fish fry. 1971.
3. Takashi HIBIYA.
Horumon no seijuku sanran no jinko togyo ni
kansuru kenkyu.
Showa 47 nendo florin suisangyo shiken kenkyu
hihojokin ni yoru kenkyu hohokusho 1973.
Studies related to the artificial control of
maturation and spawning by hormones.
Report for 1972 of research subsidized by the Maritime
Industrial test station of the Ministry of
Agriculture and Forests. (1973).
•

• 3 1
4. Mie ken Owase suisan shiken jo.
Onkaisui shiiku ni yoru madai soki sanran ni tsuite.
Gyorui shubyo seisan kenkyu II. 1 - 6 1973.
Mie prefectural maritime test station, Owase.
On the early spawning of the red sea bream,
plirmonrionnnui2E due to rearing in warm sea water.
Studies of the production of fish fry. II 1-6, 1973.
•

32 •
Parent fish for ege collection.
3. The process of maturation of the gonads..
Kazunori TAKANO.
(Faculty of Fisheries,
University of Hokkaido).
The process of maturation of the gonads extends from
the formation of the gametes to their release, and includes
the morphological and functional changes of the gonads and
of many other related organs which accompany this maturation.
Thus this process of maturation begins as the peritoneal
region is formed during the embryonic developmert of the
original gamete, continues with the formation of the gonadal
primordium, and with the attainment of maturity passes on to
the first arrival at a functionally mature condition. This
takes a long time, up to the whole lifetime of fishes which
spawn only once. However in many fish which are able to
breed two or more times, and which after spawning can once
again pass from a sexually inactive condition to funutional
maturity, the process can be properly considered as that of
the ripening of the gonads. Moreover, since the process of
arriving at maturity for the first time involves morphological
changes of the gonads and of the related organs, it is
necessary to distinguish it from ihe so-called reproductive
cycle of the fully grown fish.

• 33
Since the endocrinological processes which take part
directly or indirectly in the maturation of the gonads are
dealt with under a separate heading, the morphological
development will form the main subject of the present outline
of the development of the gonads of female Teleosts.
1. The structure of the ovary and the oviduct.
The ovaries of many Teleostà develop as a pair to
left and right. However in the viviparous swordtail
Xiphophorus helleri 1 , and in the oviparous killifish Oryzias
latipes 2 a simple Dvary only is present. In these fish a
double gonad primordium is produced, but as the organs develop
the ovaries fuse and become a single organ. Th 3 process of
fusion of the left and right lobes depends on the species,
and in fish such as the surf fish Ditrema temmincki the simple
ovary retains traces of the original bilobate structure 3.
Hoar4 recognizes two types of connection between the
fish ovary and oviduct, the cystovarian type and the
gymnovarian type. The ovaries of teleosts are usually of
the cystovarian type. This type of ovary is almost completely
composed of a large number of thin sheets (lamellae) which
contain the gametes, and the ripe eggs which have matured in
the ovary break away into the ovarian cavity. The location
of the ovarian cavity depends on the species. For example
in Oryzias latipes and in the goldfish Carassius auratus it
extends dorsally from the somatic layers of the multi-lamellate

34
•
structure of the ovarian stroma, whereas in Cottus bairdii
it extends ventrally 5, and in the perch Lateolabrax japonicus 6
it passes through the middle of the ovary. However the
cystovarian type of ovary is in all cases connected at the
tip of the ovarian cavity to a short genital tube (oviduct),
and the ripe eggs ovulated to the ovarian cavity directly from
the ovary are discharged to the outside of the body by way of
this oviduct. In contrast to this, the eggs in the ovaries
of the Chondrostei, and in the Holostei family Amia, are all
exposed in the body cavity, and the ripe eggs are released into
the peritoneal cavity. Such a structure is called an ovary
of the gymnovarian type.
In the eel Anguilla japonica the ovary of the immature
fish develops from a long narrow ridge formed on the dorsal
peritoneal wall. It protrudes into the body cavity, and during
maturation swells up into a fimbria. The median side of this
ovary is covered by the ovarian wall, but the body wall side
lacks a covering membrane and the ovarian lamellae are
directly exposed to the body cavity. In salmon and trout the
ovary generally develops a triangular cross-section, and
apart from a portion of each tip the boundaries of the upper
edges on the body wall . lack a covering membrane. Thus in
these fish the ripe eggs formed in the lamellae of the ovary
are all ovulated into the body cavity. In the eel they pass
directly from the body cavity into the genital pore, in salmon

35
•
and trout they pass through an oviduct to the genital papilla - and are discharged to the exterior710 . In these cases in
which there is no genital organ attched to the oviduct, the
body cavity has a temporary role to fulfil, and even though
the ovary is of the cystovarian type in that one part of the
ovarian wall is covered, it would be rational to class it as
of the gymnovarian type.
We turn now to a histological examination of the ovary.
The smooth section of the ovarian wall of the cystovarian type
in Oryzias latipes are well developed and cytoplasmic
protuberances develop into a villus structure only on the
epithelial cells bordering on the ovarian cavity. In the
goldfish, the muscles of the ovarian wall are missing and the
epithelial cells bordering on the ovàrian cavity develop not
only the villi but also a typical 9 plus 2 pattern of ciliae6
According to the descriptions which can be gathered from
published reports, the ovaries of adult teleosts may be
classified on the basis of such structural characteristics
in the following way.
I. Ovary gymnoform, villi developed on epithelial
cells of the body cavity.
Anguilla japonica , Salmo trutta 11 , Oncorhynchus
nerka .
Unpublished.

•
•
• 3, lia. Ovary cystoform, no muscle developed in the
ovarian wall, villi present on the epithelial
cells of the ovarian cavity.
Rhodeus amarus 12 , Rhodeus ocellatus 13, goldfish14 ,
Misgurnus anguillicaudatus .
IIb. Ovary cystoform, muscle developed in the ovarian
wall, villi absent from the epithelial cells of
the ovarian cavity.
Fundulus heteroclitus 15 , Mugil cephalus16 , Pungitius
tymensis 17 , Oryzias latipes 18 and Tilapia
massambica
This classification of the ovaries of adult fish is
closely connected with their organogenesis, and more particularly
with the development of the ovarian cavity. If more of these
observations of the structure of the gonads of adult fish are
obtained, it may become possible to deduce the way in which
the organs are formed and the manner in which fish should be
ree?.red, and to understand the relations of the organs to their
breeding behaviour. This would lead to an increase in utility.
2. Genital cavity fluids.
It is well known that when ova are stripped from
rainbow trout or chum salmon a large amount of liquid is
expelled with the ripe ovà. In species with cystovarian
ovaries such as Oryzia latipes 19 and goldfish these liquids
are secreted by the epithelium of the ovarian cavity wall,

37
and in species such as Oncorhynchus nerka with gymnovarian
ovaries they are secreted from the inner epjAheirld ef the
body cavity and from the mesovarium. In either case, the
epithelium in the sexually immature stage is formed of a
simple flat layer of epithelial cells, but at about the time
that the ova begin to accumulate yolk, a morphological
specialization into a ciliate form begins. As maturity is
approached these cells develop either a cubic or a cylindrical p21
shape. At the time of ovulation the vertices show very
remarkable morphological changes and secrete liquids from an
excretory apparatus 20 . The Golgi bodies of the epithelial
cells have an importan:; role in the detailed structure of
these liquid producers, and water soluble substances are
accumulated in the interior of the cells, mostly by a process
of absorption. The liquids obtained in this way at the time
of o\-ulation have been called "coelomic fluids" or "peritoneal
fluids" according to the structure of the genital organs of
the fish in which they are produced but Ginzburg21 advocates
the use of the generalized name "cavity fluids". Even though
there are variations in the structure of the genital organs,
the place in which the ovulated ova are temporarily accumulated
can be thought of as a "genital cavity", and since the
excreted fluids have similar origins a distinctive term for
general use will be "genital cavity fluids". •

• 38
•
•
The composition, properties and functions of these
genital cavity fluids are not yet sufficiently well known.
It is known that the liquid obtained during the stripping of
ova from trout and salmon is normall y. a rather viscous serum-
like substance. Its properties have been fairly well
investigated in salmonids. The pH is 7.9 to 8.65 21
is relatively rich in Na, K, and Cl, it contains carbohydrates
and proteins23 , and the depression of the freezing point
shows that it is somewhat hypotonie to the contents of the
ripe ova and to the blood serum 21 It is not difficult to
imagine that the liquid has functions related to the retention
of the ripe ova after Dvulation, and to their lubrication and
protection during transport. It is believed that in the • stickleback lumitimensi.s17 this secretion takes part
in the formation of a jelly-like substance in which the ova
are wrapped. It has recently been found that this secretion
in the rainbow trout contains substances which enhance the
motility of the spermatozoa and prolong their life 23 , and it
has been suggested that they may possibly contain an essential
participant in the interaction of the sperm and the ova of
Oncorhynchus keta24 . It has also been found that mammalian
gonad stimulating hormones and fish pituitary extracts could
cause an admittedly incomplete ovulation of in vitro cultures
of rainbow trout, and it is suggested that the genital cavity
fluid may contain substances which control ovulation25 . It
22 . it ,

•
39
is therefore possible that the genital cavity fluids may not
only have physical functions at the time of discharge of the
ripe ova, but may also have physiologically important roles
in the whole process from ovulation to fertilization.
3. The process of ripening of ovarian ova.
The most important role of the ovary is the formation
of functional female gametes. The process of oogenesis can
be divided into a period of proliferation, a period of growth
and a period of maturation.
The period of proliferation is one of repeated mitodis
in the oogonium. In adult fish the period during which this
proliferation occurs depends on the species. In Gasterosteus
aculeatus26 , Fundulus heteroclitus 15 , Phoxinus laevis 27 and
Pleuronectes platessa 28 it is limited to a definite period
after spawning. However in Salmo gairdnerii 29 and in Rhodeus
ocellatus 3° oogonium division can occur throughout the year
and in the guppy Lebistes reticulatus (Poecilia reticulata)31
it occurs in a cycle of ovogenesis seasons. In the interphase
oogonium, a thin cytoplasm surrounds a round nucleus with
uniformly distributed, clear chromonemata and from one to
several karyosomes.
When the period of proliferation by division is
finished the germ cells enter the growth phase. The germ
cells of this phase are called oocytes. The growth phase can
further be divided into primary and secondary phases. The

• 40
•
primary phase starts with nucleolation of the prophases
of maturation division and with chromosome synapsis, and is
characterized by the formation of plasmosomes and an increase
of the egg cytoplasm. This phase can be called the karyosome
or the peri-nucleolus stage, and the oocyte, being without
yolk is in no way affected even by removal of the pituitary.
It next enters the secondary phase. The distinctive character
of the oocyte which is seen in this phase is the accumulation
of yolk materials in the cytoplasm. The yolk materials of
fish eggs are made up of three kinds, yolk vesicles, yolk
globules and fatty droplets 32 The yolk vesicles normally
appear around the periphery of the cytoplasm, and in the next
stage they increase in number and size, but their formation
ceases relatively early, and in the mature egg they are lined
up inside the cortical protoplasm. These yolk vesicles are
homologous to the cortical alveoli, and when the eggs are
fertilized they play an important part role in the formation
of the perivitelline space. It is known that these yolk
vesicles are formed of glycoproteins and contain mucopoly-
saccharides 32 . Thus although the yolk vesicles and the fatty
droplets are simultaneously present in fish ova, they can
both be easily identified by using histochemical methods to
test for polysaccharides.
The yolk globules form an important part of the yolk
substances in most fish ova, and are an important source of
energy during the development of the embryo. In all species

•• • of fish, the yolk globules are at first deeply stained with
haematoxylin and appear as minute granules in the cytoplasm
between the yolk vesicles, and as their number increases
they gather together centripetally. The completed yolk
globules are composed of a central portion wrapped in a
surrounding membrane and a peripheral layer, and it has been
found by means of the electron microscope that the central
part has a crystalline structure'. In the herring
Clutea pallasi135 , the goldfish14 and the bitterling Rhodeus
ocellatus 13 the individual yolk glr)bules do not fuse together
even in the ripe ova. However in the flounder Liopsetta
obscura36 and in the rainbow trout29 , the yolk globules
rapidly fuse together after the germinal cell (nucleus) is
displaced by the progress of vitellogenesis, and in the ripe
egg they form a single yolk mass. In Oryzias latipes37 and
Lebistes reticulatus 38 the period during which this yolk mass
begins to form is quite early, and consequently the nucleus
is also displaced early. Many histochemical studies have
shown that the yolk globules are complex, and consist mostly
of lipoproteins with an admixture of many polysaccharides and
other carbohydrates 32 . In addition to these yolk vesicles and
yolk globules there are also fatty droplets which take
individual shapes which depend on the species. The fatty
droplets principally contain neutral fats, in particular 32 glycerides . The time at which the fatty droplets appear

•• • 42
in the oocytes depends on the species, and in Lebistes
reticulatus 38 , Lateolabrax japonica 6 , and Anguilla japonica39
they appear earlier than the other yolk substances. However
in Oryzias latipes4o and rainbow trout 29 they appear
simultaneously with or somewhat later than the formation of
the yolk vesicles and in Hypomesus àaponicus41 they appear
quite late, after the formation of the yolk globules.
The secondary growth phase can be divided into several
phases like the primary phase. Thus the period during which
the yolk vesicles appear and are formed can be taken as the
yolk vesicle period and divided into first, second and third
stages of the accumulation of yolk globules. Or the process
of oogenesis may be divided into phases characterized by
stages of formation of the fatty droplets. Since the process
of oogenesis is dynamic, these distinctions may be made in
any convenient way, and will depend on the species and on the
objectives of the study. Various methods of listing the
stages and the degree of maturation of fish ova have in the
past been proposed, but the most accurate is the histological
method. When the maturation of the ovarian egg is expressed
in terms of its cytological properties the details are made
as comprehensible as is possible, and comparative studies can
be made more useful by the use of a common terminology for
ail species.

• 43
When the accumulation of yolk is completed the egg
enters a stage of maturation and Cleavage. In many species
other than Oryzias latipes and Lebistes reticulatus the
nucleus, which up to now has been centrally established,
migrates to the animal pole in the direction of a previously
formed micropyle. This stage is called the stage of migration
of the blastula. The mechanism by which the nucleus is
displaced is as yet unknown, and the role played by the cells
of the micropyle presents a problem for the future. Soon
after the nucleus reaches thu surface layer of cytoplasm in
the neighbourhood of the micropyle, its profile becomes
unclear, and a stage of dissolution of the blastula beginsc
During this prematuration stage thread-shaped nucleolus
material, deeply coloured with haemotoxylin appears in the
region surrounding the chromosomes. When this once more
disappears, the chromosomes become clearly distinguishable 39 '
Next comes the first maturation cleavage in which the first
polar body is discharged from the chorion, and then after
the second stage of maturat'..on cleavage is reached ovulation
occurs8 . This phase is known as the phase of maturation.
In Figure 3.1 four species of teleosts are used as
examples to show how the changes in the degree of maturation
of the ovary can be based on the development of the egg,
right up to the first time that functional sexual maturity is
reached. Immediately after birth, the viviparous guppy
42.

- iiV3A ZI II 01 6 j 11 01 6 g L 9 S InJ 1 u cn 6 8 L 9 I H1NOGI
7
(S961
(1961 niaup.tp23 mains iv la Van s')
vy.t au grupu8t1ioDu0
11;0111.11134 . 1113e0,1
3srls sr. lo nonwt u 3d
3D Y1S 3N YUSW3W )410A
—peq. -em-p_se . auTT paq.q.op , poTaed Ge2JeAy 'CD
tenxas unj eiTu -ree eaojact uoTq.uanm /Ç.JUAO UT saetreqo
ave aanzu
35 %US 3ingo1a ' xlcu
3SY1 $ Si S3N3b01,111A
39Y St S3N3SC;13111A
3UYIS SI S3N 30011314
NO I irunivw IsAoolsvm
35 us 3nin›.3idd
35 VIC 3nsv47
3S Y1S
N0 11Yell VW AW 'MO
•

45
Poecilia reticulata contains oocytes from •the karyosome stage
to the peri-nucleolus stage, and within 70 to 120 days after
birth they pass beyond the third yolk droplet stage. Early
individuals . may contain fertilized eggs after 80 days. In
contrast, the ova of the rainbow trout Salmo gairdneri reach
the first yolk stage only in July after one or two years of
maturation, and after this stage egg development proceeds
rapidly, the ripe stage being reached in December29 . The eggs
in the ovary of the sockeye salmon Oncorhvnchus nerka
similarly wait till the year of maturation, and pass from the
'fatty droplet stage to the first yolk droplet phase during the p25
period from mid-April to mid-May, and then grow rapidly to
become ripe in the middle of 0ctober43 . When the eel Anguilla
japonica is artificially brought to maturity, ripe ova are
obtained in less than three months from the first accumulation
of yolk. Thus the length of time after birth needed for the
attainment of functional maturity depends on the species, but
in all cases the second phase of growth, the period needed
for the accumulation of yolk is several months. That is to
say that fish which require a long time for maturation remain
in an infantile condition of sexual immaturity for most of
the time, but once the "trigger is pulled" they reach the
functionally mature condition in a short time. The period
of transformation from the sexually immature state to the
mature state will be characterized by the commencement of
secretion of gonad stimulating hormones, but the physiological
mechanism by means of which the action is triggered is not

• 46
•
•
yet known. There are extra problems in those fish which need
an extremely long time to become mature, and in the many
species which divide into groups, one of which reaches
maturity within a year while the other remains immature.
4. • Ovulation.
Ovulation is the phenomenon of the discharge of the
mature oocyte from the ovarian lamellae into the ovarian
cavity or the body cavity. Before ovulation the oocyte is
wrapped in two layers of theca cells from the interior of the
gra-ulosa membrane cells (the epithelial cells of the
follicles). There is a chorion between the oocyte and the
granulosa cells. The chorion can usually be distinguished at
about the time of the peri-nucleolus stage, and as the oocyte
enters the second stage of growth it can be seen to be
composed of an internal and an external layer44 . The light
microscope shows that the zona radiata is pierced by many
pore canals, through which villi protrude in both directions
from the oocyte and the granulosa cells. These villi are
believed to have an important role in the interchange of
metabolic materials.
As the maturing oocyte approaches ovulation, the villi
are cut off from the pore canals of the chorion and the
adhesion between the chorion and the granulosa cells, or
between the inner and outer thecal cells is weakened. The
granulosa cells take part in producing a fluid filling the
gap which forms44,45 . Soon afterwards the cell layers

•• •
•
•
surrounding the oocyte rupture and the oocyte separates from
the ovarian lamellae. In the chum salmon and the rainbow
trout it is ovulated from a previously cracked portion of
the internal thecal cells 8 , but this does not happen in
goldfish, in which the oocyte continues to be protected by
the upper part of the lamellae.
The morphological changes which accompany ovulation
are preceded by a remarkable absorption of water into the
ovarian egg, and it is suggested that this is one of the
factors which cause ovulation46 ' 47
It is very important to be able to estimate precisely
the degree of maturation of the fish, especiall7 if ovulation
is to be artificially controlled. Methods which are being
tried out on an experimental scale include the insertion of
a polythene tube into the ovary in circler to suck out some of the
ova48 , and the control of the external environment so as to
cause all fish to reach the reproductive phase together49 .
Desirable conditions for such methods include simplicity of
operation, rapidity of determination, and little damage to
the body of the fish. This question of the determination of
the degree of maturation remains one of the principal problems
for the future.
5. The functions of ovarian hormones.
In addition to producing germ cells, the ovary also
has the important function of producing and secreting ovarian
50 hormones. The sex hormones estrone, estradio1-17, and
p26

•• • 11-ketotesto sterone 51 are known to be present in teleost ovaries.
Reactions have been observed in the eranulosa cells52 ' 53 , in
the thecal cells 54 and between them53 ' 55 , and also in the
cells between the granulosa membrane and the follicle56. It
has been suggested that the sex hormones contribute to
maintaining the development of the shape and secreting ability
of the epithelial cells of the ovarian cavity wall", and of
the production of ova58 , and that they indirectly participate,
through the liver cells, in the production and supply of yolk
material 59,60 . Recently 11-deoxycorticosteroids61 have been
extracted from teleost ovaries, and they are supposed to be
concerned in the maturation and ovulation of the egg in the
ovary. In general, if the ovary is surgically removed from
a teleost, the source of the hormone and the tissue on which
it acts are lost at the same time, so that it is difficult to
elucidate the action of the ovarian hormones by physological
means62 . This leaves many problems of very great interest
for the future.
In order to locate the presence and activity of the hormone
producing cells in the ovary, enzyme histochemical tests have
been made with 1.3 -HSD, (z \:5-3p -hydroxysteroid dehydrogenase). 1

References.
1)
8)
10)
J. M. ESSENDERG : Sex-differentiation
in the viviparous teleost Xiphophorus
helleri HECKEL. Biol. Bull., 45, 46
--970923).
2) K.ONITAxE :Morphological studies of
normal sex-differentiation and indu-
ced sex-reversal process of gonads in
the medaka, Oryzias latipes. Annot.
Zool. Japon. 45, 159.-169 (1972).
3) SE E PAGE 53
4) W. S. Ho.ta : The gonads and repro-
duction, in "The Physiology of Fish-
es" (M. E. Buown ed.), Vol. 1, 287—
321, Academic Press, New Yorl • C1957).
5) H. W, HANN : The history of the germ
cells of Cottus bairdii GIRARD. J.
Morph., 43, 427-497 (1927).
6) I. HAYASHI : On the ovarian matura-
tion of the Japanese sea bass, Lateo-
labrax japonicus. Japan. J. Ich-
thyol., 19, 243---254 (1972).
7) D, HEY : The fertility of brown trout
eggs at the Jonkershock Inland Fish
Hatchery. Trans. Am. Fish. Soc.,
77, 65-40 (1949).
SE E PAGE 53
9) V. D. VLADYKOY : Fecundity of wild
speckled trout (Salvelinus fontinalis)
isi Quebec Lakes. J. Fish. Res. Ba.
Canada, 13, 799--841.
SEE PAGE 53
11) K. R. Asumr : The effect of steroid
hormones on the brown trout (Salmo
trutta L.) during the period Of
gonadal differentiation. J. Embryo!.
Exp. Morph., 5, 225-249 (1957).
12) L.H. BRETSCHNEIDER and J.J.Durvstriz
DE WIT : Sexual Endocrinology of
Non-mammalian Vertebrates, 146 pp.,
Elservier Publ. Co., Baltimore (1947).
1.1) K. Sarast : Correlation between the
growth of the ovipositor and ovarian
conditions in the bitterling, Rhodeus
ocellatus. Bull. Fac. Fish. Hokkaido
Univ., 13, 137-151 (1962).
14) K. YAMAMOTO and F. YAILAZAM
' Rhythm of development in the oo-
cytes of the goldfish, Carassius
auratus.. Bull. Fac. Fish. Hokkaido
Univ.,12, 93-410 (1961). • .
15) S. A. MATTHEWS : The seasonal cycle
in the gonads of Fundulue. Biol.
Bull., 75, 66.--.74 (1938).
16) A. H. STENGER A study of the
structure and development of certain
reproductive tissues of Mugil ceph-
alus LINNAEUS. ZOO/OgiCG, 44, 53--70
(1959),
17) T. S. Ysussioro : Eggs and ovaries
of the stickleback, Pungitius tymen-
sis, with a note on the formation
of a jelly-like substance surrounding
the egg. J. Fac. Sci. Hokkaido Univ.
VI, 15, 190,...,201 (1963).
18) K. YAMAMOTO : Cyclical changes in

•
• 5 0
the wall of the ovarian lumen in the medaka, Oryzias latipes. Annot.
Zool. Japon., 36, 179-186 (1963).
19) K. TAEANO : Fine structure of the wall of the ovarian lumen in the teleost, Oryzias latipes. Bull. Fac.
Fish. Hokkaido Univ.,19,76-82(1968).
SE E PAGE 54
21) A.S. Grxzentos : Fertilization in Fishes and the Problem of Poly-
sPerinY. 36 51V., Izdatel'stvo "Nauka", Moskva (1968).
22) SEE PAGE 54
23) T. YOSHIDA and M. Nounne. : A substance enhancing sperm motility in the ovarian fluid of rainbow trout. Bull. Jap. Soc. Sci. Fish., 38, 1073 (1972).
24) SE E PAGE 55
25) B. JALABERT, B. BRETON and C. BRY
Maturation et ovulation in vitro des ovocytes de l'a Truite Arc-en-Ciel Salmo gairdnerii. C. R. Acad. Sci. D, 275, 1139-4142 (1972).
26) A. CRAIO•BENNETT : The reproductive cycle of the t h ree- spi ned stickleback, Gasterosteus aculeatus LINN. Phil.
Trans. roy. Soc. London, 13, 219, 197--279 (1930).
27) W. S. BI:mu:won : Gametogenesis and some endocrine factors affecting it in the adult minnow (Phoxinus laevis
L.).J. Endocrinol., 3, 211---219 (1942).
28) W. A. BARR : The endocrine control of the sexual cycle in the plaice, Pleuronectes platessa (L). I. Cyclical changes in the normal ovary. Gen.
Comp. Endocrinol., 3, 197-204 (1963).
29) SEE PAGE 56
30) K. 7'Am/dim.° and K. SHIRAI : Origin of the yearly crop of eggs in the bitterling, Rhodeus ocellatus. Annot.
Zool. Japon., 35, 218-222 (1962). 31) K. TAXA» : Origin of the oocytes in
the adult guppy, Lebistes reticulatus.
Bull. Fac. Fish. Hokkaido Univ.,18,
137,-142 (1965).
32) S' EL PAGE 56
33) K. YAIIAMOTO and I. 00Tà. : Fine structure of yolk globules in the oocyte of the zebrafish, Brachydanio
rcrio. Annot. Zool. Japon., 40, 20---27 (1967).
34): N. N. GUPTA and K. YAMAMOTO:
Electron microscope study on; the fine structural changes in the oocytes
- of goldfish. Carassius auratus, during
yolk formation stage. Bull Fac. Fish.
Hokkaido Univ., 22, 187-205 (1971).
35) K. YAMAMOTO : Studies on. the formation of fish eggs. VII. The fate of the yolk vesicle in the oocytes of the herring, Clupea
20)

SEE PAGE 56
SEE PAGE 57
• 51
•
during vitellogenesis. Annot. Zool.
Japon., 29, 91 ,-96 (1956).
36) . K. YAMAMOTO : Ditto. XI. The
formation of a continuous mass of
yolk and the chemical nature of lipids
contained in it in the oocyte of the
flounder, Liopsetta obscura. J. Fac.
Sci. Hokkaido Univ., VI, 13, 344-351
(1957). 37) K. YAMAMOTO and H. Yosutou. :
Rhythm of development in the oocyte
of the medaka, Oryzias latipes. Bull.
Fat. Fish. Hokkaido Univ.,15, 5-49
(1964). 38) K. Tars»: On the egg formation
and the follicular changes in Lebist es
reticulatus. Bull. Fac. Fish. Hokkaido
Univ., 15, 147-155 (1964).
39)
40)
41) K. YAMAMOTO : Studies on the for-
:nation of fish eggs. VIII. The fate of the yolk vesicle in the oocyte of
smelt,../iypomesus japonicus, during
vitellogenesis. Embryologic, 3, 131-- 138 (1956).
42) K. YAMAMOTO : Ditto. II. Changes in
the nucleus of the oocyte of Liopsetta
obscura, with special reference to the
activity of the nucleolus. J. Fac. Sci.
Hokkaido Univ., VI, 12, 375-499 (1956).
43) R. IsuinA, K. TArlot and S. ARITA :
Criteria for the differentiation of
mature and immature forms of chum and sockeye salmon in northern seas.
In!. North. Pat. Fish. Corn. Bull., 5, 27-47 (1961).
44) K. HiaosE : The ultrastructure of
the ovarian follicle of medaka, Oryzias latipes. Z. Zellforich., 123.:
316-.329(1972). • 45) K. YememoTo and F. YAmAzearr
Hormonal control of ovulation and spermiation in goldfish. Gunmcr
SymPosia on Endocrinol., 4, 131.-145 . (1967).
SEE P A GE 57
47) K. HIROSE, T. HIRANu and R. !man :- Effects of salmon gonadotropin in
the ayu, Plecoglossus altivelis, with
special reference to water balance. Conde. Biochem. Physiol., 47A, 283 .--,289 (1974).
48) Z. H. SLIEHADEll, C. M. Coo and K.
K. MILISEN: Validation of in vivo
method for monitoring ovarian de-
velopment in the grey mullet (Mugit
cephalus L.). J. Fish. Biol., 5, 469 —496(1973).
49) SEE PAGE 57
50) Y. KATZ, 13. ECKSTEIN, R. 'tux and
R.GermEn: Estrone and estradiol-
na in the ovaries of Tilapia aurez
(TELEOSTEI, CICHLIDAE). Camp. Bi--
ochem.Physiol., 40B,1005--1009(1971).
51) U. EYLATU and B. ECKSTEIN :Isolation
of 11-ketotestosterone and dehydro-epiandrosterone from ovaries of the
common mullet, Mugit capito. Gen.
Camp. Endocrinol.,12, 58-62 (1969).
46)
•

• 52
•
52) J. G..D. LAMBE R T: The ovary of
guppy Poecilia reticulata. The
granulosa cells as sites of steroid
biosynthesis. Ibid., 15, 464-476(1970).
53) Z. YARON : Observations on the
granulosa cells of Acanthobrama
terraesanctae and Tilapta nilotica
(TELEoszst). Ibid., 17, 247-252(1971).
K. YexAmoTo and H. ONOZATO :
Steroid-producing cells in the ovary
of the zebrafish. Brachydanio rerio.
Annot. Zool. Japon., 41, 119-128 (19
68).
55) G. BAHL : Histochemical localization
of ' 413-31?-hydroxysteroid dehydro-
genase in the ovaries of a teleost
fish, Scornber scomber L. Gen. Comp.
Endocrinol., 5, 284.--296 (1965).
56) Y. Iv/mum : Histochemical detection
of 45-3a-hydroxysteriod dehydro-
genase in the ovary of medaka,
Oryzias • latipes, during annual «
reproductive cycle. Bull. Fac. Fish.
Hokkaido Univ., 23, 177-484(1973).
57) H. TAKAHASHI and K. Dace» : Sex
hormone-induced precocious hyper-
trophy and ciliation of epithelial cells
in the ovarian lumen of the goldfish.
Annot. Zool. Japon., 44, 32-41(1971).
58) N. E. STACEY and N. R. Loam :
Regulation of spawning behaviour
in the female goldfish. Nature, 247,
71--72 (1974).
59) K. AIDA, PHAN-VAN-N(3ex and T.
HIBIYA : Physiological studies on
gonadal maturation of fishes-I Sexual
difference in composition of plasma
protein of Ayu in relation to gonadal
maturation. Bull . JaP. Foe. Sci. Fish.,
39, 1091-1106 (1973).
60) K. AIDA, K. HIROSE, M. YOKOTE and
T. HISIYA : Ditto. II Histological
changes in the live:. cells of Ayu
following gonadal maturation and
estrogen administration. Ibid., 39,
1107-1115 (1973). •
61) L. COLOMBO, H. A. BERN, J. PIEPRZYI
and P. W. JOHNSON : Biosynthesis
of 11-deoxycorticosteroids by teleost
ovaries and discussion of their
possible role in oocyte maturation
and ovulation. Gen. Comp. Endo-
crinol., 21, 168-478 (1973).
62) R. REINBOTH : Hormonal control of
. the teleost ovary. Am. Zoologist, 12,
307-424 (1972).
54)
•

• 53
References.
3. MIZUNOE Kazuhiro.
Umitanago no kenkyu III.
Umitanago no ranso no seijuku narabi ni kisetsuteki
junkan ni kansuru kenkyu.
Nagasaki dai suisan ken ho 11 1 -17 (1961)
K. Mizunoe.
Studies of Ditremma temmincki III.
Studies of the maturation of the ovary of, Ditremma
temmincki and of its seasonal cycle.
Reports of the Fisheries research laboratory,
Nagasaki University, 11 1-17 (1961).
8. YAMAMOTO Tadashi.
Nishin, nijimasu oyobi yatsume no hairan katei.
• Gyo zatsu • 182-192 (1955).
Ti Yamamoto.
The process of ovulation in Clupea pallasii, Salmo
gairdnerii and Entosphema japonicus.
Japanese Journal of Ichthyology. 182-192 (1955).
10. NOMURA Minoru.
Nijimasu no jinko sairan ni kansuru kiso kenkyu I.
Seijuku ni tomonau seishokuso no keitai henka to
hairan katei.
Nichi sui shi 28 409-416 (1962).

• ••
•
10. M.Nomura.
Basic research related to the artificial collection
of eggs from Salmo gairdnerii.I.
Morphological changes accompanying maturation of
ovaries and the process of ovulation.
Bulletin of the Japanese Society of Scientific
Fisheries, 28 409-416 (1962).
20. KUROZUMI Kazumasa.
Bunpitsu no keitaigaku ni kansuru denshi
kenbikyot3ki kenkyu.
Denkengakkai ho 14 12-26 (1965).
K. Kurozumi.
Electron microscope studies of the morphology of
internal secretion.
Journal of the electron microscope society,
14 12-26 (1965).
22. NOMURA Minoru.
Nijimasu no jinko sairan ni kansuru kiso kenkyu VI.
Tansui, tochoeki, taikoeki, nyo no kishaku ni yoru
seishi no katsudosei to seieki no chozo ni tsuite.
Nichi sui shi 723-733 (1964).

55
22. M. Nomura.
Basic research on the artificial collection of
eggs from rainbow trout. VI.
The effect on the storage of mut and the activity
of sperm produced by dilution with fresh water,
isotonic fluid, coelomic fluid or urine.
Bulletin of the Japanese Society of Scientific
Fisheries .3..2 723-733 (1964).
24. TAKANO Kazunori, HIROI Osamu, YASUKAWA Masao,
SUETAKE Toshio.
Sake, masu rui no tamago oyobi seishi no hozon ni
kansuru kenky,. I.
Sake (Oncorhynchus keta) mi jusei tamago no hozon
ni tsuite.
Sake, masu fuka basho ho .2.Z 31-37 (1973).
K, Takano, O. Hiroi, M. Yasukawa, T. Suetake.
Studies of the preservation of the roe or mut
of salmonids I.
Preservation of unrertilized eggs of. Oncorhynchus keta.
Salmon and trout hatchëry reports,
• ?I 31-37 (1973).

56
•
29. YAMAMOTO Kiichiro, OOTA Isao, TAKANO Kasunori,
ISHIKAWA Tetsuji.
Nijimasu no seijuku ni kansuru kenkyu I.
Ichinengyo no ranso no hattatsu ni tsuite.
Nichi sui shi 11 123-132 (1965).
K. Yamamoto, I. Oota, K. Takano, T. Ishikawa.
Studies of maturation of the rainbow trout I.
On the development of the ovary in first-year fish.
Bulletin of the Japanese Society of Scientific
Fisheries. 21 123-132 (1965).
32. YAMAMOTO, Kiichiro.
Gyoran ni oke:Ju rano keisei.
Saibo kagaku shinposhiumu 8 119-134 (1958).
K. Yamamoto.
The formation of yolk in fish eggs.
.Cytochemical symposia, 8 119-134 (1958).
39. YAMAMOTO Kiichiro, 6MORI Masaaki, YAMAUCHI Kohei.
Nihonsan unagi (Anguilla japonica) no rankeisei
ni tsuite.
Nichi sui shi 40 9-15 (1974).
K. Yamamoto, M. Omori, K. Yamauchi.
On the development of the ova of the Japanese
eel (Anguilla japonica).
Bulletin of the Japanese Society of Scientific
Fisheries 40 9-15 (1974).

• 57
40. YAMAMOTO Tadashi.
Medaka no ranshi keisei, toku ni sono saibo
kagaku teki kenkyu.
Gyo zatsu 4 170 - 181 (1955).
T. Yamamoto.
Study of the development of the ovum in Oryzias
latipes, with special reference to its cytochemistry.
Journal of Ichthyology 4 170 - 181 (1955).
46. HIROSE Keichi.
Gyorui no hairan no naibunpitsu
To kai sui ken ho /Lk 67 - 81 (1973).
K. Hirose.
Endocrine control of ovulation in fish.
Bulletin of the Tokai regional fisheries
research laboratory .72± 67 - 81 (1973).
49. TAKANO Kazunori, KASUGA Seiichi, SATO Shigeru.
Jinko koshuki shita ni okeru medaka no seishoku
nichishuki.
Hokudai suisan iho 24 91 - 99 (1974).
K. Takano, S. Kasuea, S. Shigeru.
Diurnal• reproduction cycle in Oryzias latipes
under artificial control of the light cycle.
Bulletin of the Faculty of Fisheries,
Hokkaido University, 24 91 - 99 (1974).

• 58
4. Detailed characteristics of reproduction.
Sadaichi KATO.
(Freshwater fisheries research laboratory, Nikko).
ln order to produce fry efficiently in restricted
conditions, it is important to have a sufficient understanding
of the many characteristics peculiar to the reproduction of
the parent fish. By means of changing the methods and
conditions of rearing and the ways of breeding in accordance
with this understanding, it will be possible to make improvements
and to select parent fish which have desirable characteristics.
With this in mind, the Nikko division has investigated the
variability of many aspects of the growth of the rainbow trout
Salmo gairdnerli. Detailed results have been reported about
the relation of the age and the growth of the parent fish to **
the weight of the ripe eggs , the number of ripe eggs ' the ***
weight of an egg , and the season of spawning.
1. The ripe eggs.
The weight of the ripe eggs is determined by the
number of eggs and the weight of each egg, but since it was
found that the weight of the ripe eggs was the quantity which
among these three was most closely correlated to the body
* The gross weight of the completely mature eggs. ** The number of completely mature eggs. **** The average Weight of a completely mature egg.
P 31
•

59
weight of the fish at the time when the eggs were collected,
it was thought that the ratio of the body weight to the
weight of the gonads (which in this case is the weight of
the ripe eggs ) would be a most important reproductive index.
It is found that there is a linear relation between
the weight of ripe eggs and the body weight which is
independent of the age of the fish, and an increase of body
weight is accompanied by an increase in the weight of the
ripe eggs. The weight of ripe eggs shows the same trend when
the fish are large after being raised with a plentiful food
supply. Since the weight of one egg:hardly increases at all
as body weight increases, the increase in the weight of ripe
eggs must necessarily be due to an increase in the number of
eggs. The chance of survival of a single egg is therefore
not normally affected by rearing in conditions where there is
an extremely large food supply.
Since the gonad index is inversely related to the
body weight, its value is lowered in large fish.
2. The gonad index of mature individuals and the Pize
distribution of eggs in the ovary.
Many aspects of the degree of maturation can be
expressed in terms of the gonad index, but its value is low
in the early stages of maturation and it cannot be used to
In this paper the gonad index is _yr .ielt_g gonads x 100. Body weight

•• • 60
•
discriminate between individuals which will mature this year
and individuals which will not. These two classes can be
distinguished by the temporal change of the distribution of
egg diameters in the ovary. Figure 4.1 shows the relation
at various times between the gonad index and the diameter of
the eggs in the ovary .
As the spawning season approaches, two groups can be
distinguished. One group contains individuals in which the
average egg diameter and the gonad index is increasing, the
other contains individuals in which the average egg dnmeter
is less than one millimetre and the gonad index is still low.
As reported by Yamamoto et al 1 , it is supposed that the latter
group contains individuals which will not mature this year.
Since the two groups remain clearly separated after
July, it is believed that no individUals pass from the group
which will not mature into the group which will mature. Thus,
although it is not possible to distinguish positively between
the two groups in May, the individualsin which the egg diameter
exceeds one millimetre are believed to be those whicl-. will
mature this year.
The seasonal changes of the distribution of egg
diameters in the ovaries of individuals which will mature this
year are shown in Figure 4.2. In April and June, the gonad
indices are low, and it is difficult to determine whether an
* The average value for the group of eggs which is in an advanced stage of development.
p33

5
ro
ed 10
0 t..5 •
April GoNA0 INDEX (%).
0.7 .
. 1 . ,
: June . .
,r/\, . . 0.5
, August: , -
1.4 ..
. , September
. 5.2
_
. n October 6.1
,-...... i ....s. 4
10
0
e•—■
• o ,
P 0
0
p1 1
10
O
• 61
0
- May
•
d■ t I • 1
- Jùly
W. • . ,•• *, , , 'AuguSt
. te se, 1
• t
- :0Otober .: ••
>i • ,...... •-• ee
. s.. . - ..e
. .
te, , • , 0 2 4.
Mean diameter of ovarian eggs (mm).
Diameter of ovarian eggs (mm).
Figure 4.1.
Mean diameter of
ovarian es and
gonad index.
Figure 4,2,
Variation with time of
the distribution of diameters
of ovarian eggs in mature
individuals.
•

• 62
•
•
individual will or will not mature this year. However the
distribution of egg diameters in the ovaries shows two peaks,
one containing eggs which are in the course of maturation and are
advanced and one containing eggs which are not maturing and
it is to be supposed that the eggs in the former group, in
which the average egg diameter exceeds one millimetre, are
those which will mature this year. In August, the group in
which the egg diameter is less than one millimetre and the
group in which maturation is proceeding are completely
separated. In September and October the two groups are
similarly distinguishable, and as spawning approaches the
small-diameter eggs r.main unchanged and only the eggs of the
group which is maturing increase in diameter. It is
particularly remarkable that after the maturing and non-
maturing groups have separated, the maturing eggs are not
supplemented from the small egg group. This type of egg
diameter distribution is found to be quite normal in healthy
individuals, but in diseased individuals or in those which
have been kept in conditions of starvation eggs of size
intermediate between those of the maturing group and the
small egg group have been found. Even in these cases no
peaks other than the two already described have been found.
3. The ripe eggs.
It appears that after maturation has begun, none of
the eggs in the group which did not start to mature transfer
p34

• 63
1 0.743
0.838
0.707
0.745
0.582
0.740
0.617
0.686
2
0.587
0.785
0.634
0.635 4
Group No.
Group of fibh spawning for the first time,
Body weight in May and number of ripe eggs*.
Body weight in August and num-ber of ripe eggs*.
Body weight when mature and number of ripe eggs.
to the other group, and if this is so, the number of mature
es •will not exceed the number which were present when the
two peaks separated at the start of maturation. Those shown
in April . in Figure 4.2 can be said to determine an upper limit
to the number of eggs which can be spawned this year.
It is true that the size of the fish has a definite
influence on the number of ripe eggs, but it appears that the
number of ripe eggs will not be increased by delaying the
maturation season in order that the parent fish may be larger.
Table 4.1.
Comparison of correlation coefficients.
* Number of ripe eggs when fully mature.

• 64
•
•
Four groups of fish spawning for the first time were
investigated. The body weight of each individual was measured
in May, in August, and at the time of spawning in November to
January, in order to discover the season at which the body
weight was best correlated with the number of ripe eggs.
The results are shown in Table 4.1, and the highest degree of
correlation was that between the number of ripe eggs and the
body weight in August. As has already been stated, it is
believed that the number of eggs in an individual fish which
will mature during the current year does not increase after
May. Thus if there is any change at all in the number of
• eggs after May, this change can only be a decrease in the
number of eggs which may mature. Indeed, the fact that the ■
number of ripe eggs is highly correlated with the body weight
in August, shows that the number of ripe eggs has in many
individuals already been decided at some time in August. It
therefore appears that if it is desired to increase the
number of ripe eggs in the parent fish, the fish should be
chosen to be large by April at the latest, and from then
until August attention should be transferred to the nutrition,
especially the quantity and type of food, given to the
parent fish.
4. The weight of an egg.
There have been many studies of the relation between
the size of the fish and the weight of the egg, and it is

• 65
P35
•
•
reported that an increase in body weight is accompanied by an
increase in egg weight, but it is not yet known what relation
there may be between the weight of the egg and the size of
the fish in different year groups.
Figure 4.3 shows the relation between body weight and
egg weight according to age. It can be seen that on the
whole an increase of body weight is accompanied by an increase
in egg weight, but there is no evident relation between body
weight and egg weight when fish of the same age are compared.
In the case of the 3-year old fish shown in Figure 4.3, those
of body weight 1000g have eggs weighing 57mg and those of body
weight 2000g have eggs weighing 62mg, so that doubling the
body weight results only in an 8% increase of egg weight.
The relation between body weight and egg weight was also
investigated in the four groups of fish spawning for the
first time, and there was no correlation between the two
quantities in three of the groups. It appears from this that p36
factors other than the size of the fish contribute to the
size of the eggs.
An interesting question is whether individual fish
which have produced large eggs in one year will produce large
eggs in the next year, that is whether the size of the eggs
is a genetic characteristic. For this purpose individuals
were tagged and investigated,-and correlation coefficients
were obtained between the size of the eggs spawned in the

• 66
100
tx0
60 - • 4->
• • •
sa) 3 '
4.0 e) r
j4 t 6
cd
. .
• ..
• • a • .. -
• • • • •
o 0 0 .7
o 0 • •0 o•- • c o ° co! co
o oo. o o o • • • 6 o.
0 0 0 ' Jo. ... :
° • of ° . °
. • ° eeoe°
• • • : 0 0 . 0 .
• 000. _DO O. 0 " ° se.
e0 o •°C..e:e .° -. . •
0.00 .- -.0..0 . '.-.. ... • ....1!,...- a..
. 0 000 "6 . • 0 0 . • ••
• . ibe . 0 • • • %. .0 0 .0 ' O • 0 0
• . • • , .
° • • .. , •• ° : 0 • r.• •
-
•
1000 . 2P09 Body Weiiht (g).
Figure 4.3.
Relation between body wefght, age, and egg weigh.t.
2-year-Old. 0 3-year-old. 0 4-year-old.
3009
•

•
67
second year and the size of those spawned in the third year,
and also between those which spawned in the third and fourth
years. It was found that there was a great probability that
those individuals which produced large (or small) eggs in one
year would produce large (or small) eggs in the next year.
The distribution of weights of the eggs produced by
the fish spawning for the first time was investigated. The
individuals which produced large eggs could be distinguished
from those which produced small eggs, and it was found that
there was a clear difference between the two cases. Five
fish producing large eggs and five fish producing small eggs
were selected from thL group spawning for the first time,
and their individual egg weight distributions are shown in
Figure 4.4. The large eggs were relatively uniform in weight
with little difference, but the small eggs show relatively
large variations in weight. A similar tendency was found in
other first-time spawning fish. It is suggested that this
relation between the size of the eggs and the width of the
variations depends not only on the state of health of the
fish but also on genetic factors related to the accumulation
of yolk during development of the egg. This is thought to be
a very interesting quetion.

• 68
Egg weight (mg).
Figure 4.4.
The difference in e.g weiFht distributions in
individuals which.are producing large eggs (black)
and those producing small eggs (white).
•

• 69
•
•
5. The spawninu season.
The age at maturation and the date of spawning show
very large variations even between individuals of the same
group. In the rainbow trout, it is quite common to find
examples of differences of more than two years in the age of
first spawning, and even among individuals which spawn at the
same age the interval between an early spawning season and a
late spawning season is several months. These differences
lead to many difficulties in egg collection and in the
man -tgement of breeding. Fish which were believed to be
genetically similar were therefore investigated with the idea
of bringing their spawning dates closer together, and the
dates on which they first spawned were recorded. Figure 4.5
shows the length of the first spawning season of fish raised
from five female parents A - E. Shown for comparison is the
length of the first spawning season for fish raised from a
large number of parent fish from which eggs were collected
on the same day. The length of the spawning season was 86
days, which is not significantly different from the average
spawning season length of 82 days obtained from 54 years of
measurements made from 1911 to 1964 in the same place by the
former Nikko fish hatchery. However the length of the first
spawning seasons of the fish from selected parents were 39
to 60 days, so that these spawning seasons were considerably
shorter.

20
0 •
.--.. 20 •
0 •1-1 2
o 0
2
2
-- Oct Nov
• 70
•
Same day collection
2OF Fe,r0Up..
ne,-- 277
Oct NOV Dec Jan Feb
A n=118
Et it=62
ù .. 11=108
D n=49
. ,
. ,
Dec Jan Feb
Figure 4.5.
The length of the spawning season in the groups
hatched from eggs collected on the same day
(upper diagram) and in sibling fry (A - E).
* .The eggs were all collected on the same day, but from a large number of parent fish.

•
•
71
The variability of the day of the first spawning of
the fish from selected parents was small, whereas the
variability of this date in fish from eggs collected on the
same day was large. It may be inferred that eggs collected
onthe same day came from many parent fish whose spawning
periods were centered around different dates.
The next question was the variation of the date of
first spawning with age. This is shown in Figure 4.6 in
relation to the spawning date of the parents. It was found
that the date doe3 vary with the spawning age. The spawning
date of 3-year old fish was considerably earlier than the
spawning date of the parents and the date for 4-year old
fish was only slightly earlier. In 5-year old fish it was
much later. Thus from 3 years old onwards the date of first
spawning gets latser with increasing age. Individuals which
first spawn at two years do so later than the parental
spawning day, but thereafter the trend is the same as those
which first spawn at three years.
From these facts it would appear that if th::
attention were given to concentrating together those which
first spawn on the same date, and to the variation of the
spawning date with age, it might be possible in the management
of breeding not only to shorten the spawning season but also
deliberately to produce a series of spawning periods during
the spawning season.

• 72
-e -20
Days before parental spawnine date.
m 40. Days after parental spawning date.
-- -0 -
Figure 4.6.
Change of spawning season with age.
Changes of median spawning season of siblings.
Mean change with age.

• 73
The relation between size and spawning date in fish
which first spawned at three years of age was also investigated.
It was found that on the whole the large individuals tend to
spawn at an early period (Figure 4.7). However, considering
the results already mentioned that increase of age tends to
be accompanied by a later spawning date, it is thought the
growth is not the only factor that influences the spawning
season, and this question must be left for future clarification.
•
•

?OW
• • • •
• • • • • •
-P 1000 '
0.) ;e. •
•
• •
•
• •
•
• • • • • •
Nov Dec 1.1ari
• 74
Figure 4.7.
Relation between sibling size
and spawning season.

•
75
References.
1. YAMAMOTO Kiichiro, DOPA Isao, TAKANO Kazunori,
ISHIKAWA Tetsuji.
Nijimasu no siejuku ni kansuru kenkyu I.
Ichinen go no ranso no hattatsu ni tsuite.
Nichi sui shi 11 123-132 (1966).
K. Yamamoto, I. Oota, K. Takano, T. Ishikawa.
Studies of maturation of the rainbow trout I.
On the development of the ovary in first-year fish.
Bulletin of the Japanese Society of Scientific
Fisheries. 11 123-132 (1965).
2. KATO Sadaichi.
Nijimasu no seicho heni ni kansuru kenkyu III.
Mesu no sanranki no hendo oyobi sanranki kan
tanshutsu no tame no hitotsu no hobo.
Tan sui ken to 22 1, 41-51 (1973).
S. Kato.
Studies of changes during growth of rainbow trout III.
Variations of the period of spawning in the female and
a method of shortening the period of spawning.
Research reports of the Freshwater Fisheries
Research Laboratories. L2 1, 41-51 (1973).

• 76
•
5. Internal secretions, egg maturation and ovulation.
Fumio YAMAZAKI.
(Faculty of Fisheries, Hokkaido University).
The expression "egg maturation" may in principle be
taken to include both the growth of the egg and ovulation,
but since growth and ovulation differ considerably and there
is a clear endocrinological distinction, they should be
discussed separately. Reviews of knowledge about the
reproduction of fish have been published in Japan by Hibiya 1
and by Yamamoto2,3 but our present knowledge of the
endocrinology of maturation and ovulation remains fragmented.
At the present time t...ere is much which is difficult to under-
stand about the function of internal secretions in the
maturation and ovulation of eggs, and an outline will be given
in this paper of some new information about the secretions
from the pituitary and the ovary.
1. The maturation of the egg.
After finishing division in the ovary the oogonium
enters the growth phase as a primary oocyte (also called an
ovarian egg). In a discussion in outline of this growth
phase on the basis of maturation, it can be divided by the
accumulation of yolk into a primary and a secondary stage
of growth.
•

•• • 77
1.1 The primary stage of the growth phase of the oocyte.
This stage corresponds to the prophase of maturation
cleavage before vitellogenesis, and the nucleus having passed
through the stage of homologous pairs, the leptotene stage,
the zyeotene stage, the bouquet stage, and the pachytene
stage, enters the diplotene stage. The ovoplasm increases
and the oocyte enters the yolkless stage. The size of the
egg at this stage depends on the species and ranges from 13
to 250/).... The influence on the ovary of removal of the
pituitary has been investigated in many species, and it is
known4 that at this stage the egg does not disintegrate but
remains inside the ovary and grows slowly. As observation
of the recently discovered pituitary-deficient "cobalt" p42
rainbow trout has shown, the ovarian egg is in this stage
able to grow independently of the pituitary 5 .
It has been established with certainty that sex hormones
in particular female sex hormones, are secreted by the ovary,
but it is not known whether sex hormones are secreted from
the ovary in the immature stage in which only the yo7kless
primary stage has been reached. However 50 to 100 units of
estradio1-1 were found in ten extractions from the
pituitary of immature goldfish, and the oral administration
of 5?-g of the synthetic sex hormones diethylstilbestrol or
methyltestosterone in each uram of feed for three weeks
clearly caused an acceleration of growth of primary stage

•
•
• 78
ovarian eggs in trout, so that it cannot be denied that sex
hormones may participate in the primary growth stage. Twenty
days of oral administration of 100eg of methyltesterone per
gram of feed to Oryzias latines resulted in a very great
increase in the weight of the ovaries, and it is reported that
the ovaries became active 6 . It may therefore be possible that
ovarian activity may be significantly accelerated by means of
sex hormones, and this may be an important technique for the
control of maturation.
1.2 The secondary stage of the growth phase of the oocyte.
This stage is characterized by vitellogenesis, and is
the most important period during the maturation of the egg.
During this time the oocyte grows rapidly, and the diameter
of the egg, which in the avitelline stage was 100cto 200e,
increases to about 1mm in the carp and in some salmonids and
trout may reach more than 5mm.
1.2.1 Vitellogenesis and the pituitary.
When the fish pituitary is excised the eggs in the
secondary stage of growth disintegrate in a characteristic
way. The most easily affected eggs are those which are in
the process of accumulating yolk. In the goldfish they are
those which are surrounded by the most hypertrophic follicle
cells and, with diameters of 350/tuto 750?.., are in the first
or second stages of vitellogenesis. Those which have formed
yolk vesicles and yolk droplets and are in the third stage of

p4.3
pLA,
79
vitellogenesis remain in the ovary for a relatively long
time, but they are slow to develop and eventually disintegrate,
leaving yolkless eggs only in the ovary. The rapidity of
disintegration depends on the temperature. At 15o C it may
take several months, but above 20 °C it is extremely fast. For
example, at 25 ° C all the eggs containing yolk had disintegrated
in about two weeks4
This egg disintegration after hypophysectomy has been
found in the goby Gobius pagmel1us 7 , the plaice Pleuronectes
platessa8 , the catfish Heteropneustes fossilis 9 and several
other species. On the other hand, reimplantation of the
pituitary or injection of a fluid suspension or extract very
clearly accelerates the secondary stage of growth, and causes
vitellogenesis to be resumed. This fact alone shows that a
hormone which promotes vitellogenesis is present in the
piscine pituitary. This hormone is called gonadotropin (GTH)
and by promoting vitellogenesis and the secretion of sex
hormones it plays an important part in egg maturation.
1.2.2 Sex hormones and vitellogenesis.
GTH is not the only substance which affects
vitellogenesis. The sex hormones also participate in a
complex manner. The connection between the formation of
yolk materials and the female sex hormone has been clearly
shown in the recent work of Aida et al10,11 , and will be
outlined in the present symposium. The female sex hormones

80
••■■
■%
PREOPTIUUS
SUCLEUses\\
CÏÏERIOR\
PROTUBERANCE;
ECONUARY
SEX SYMPTOMS
Aer
.a
.0
o
w 0 m ic 0 r r a IC a a =
.....
Figure 5.1.
Endocrinoloeical processes related to maturation of fish.
• GROW-T-1.1
■ • THYROID. ■ # • GLAND
› •t-- ,
■%
. FEMALE sex NORMONES
1 4
YOL K MATERIAL' _ .
G - T
1H
\isî STADE 9* 2
e
. NDSTAGE
1\ )_ OROWTO ..-Yoik xceuek
WejmuLATIomeirAli
OVIDUCT
E G G

81
act on the liver, the yolk materials are secreted from the
liver and accumulated by the oocyte. The second stage of
growth has been shown to be stimulated in this way in many
species of fish12 .
There are many reports of studies of the sex hormones
secreted by teleost gonads. The female sex hormones most
often extracted from or detected in the ovaries of fish are
estradio1-1773 and estrone 13,14 . The male sex hormones
11-ketotestosterone and dehydroepiandrosterone have also been
isolated from the ovaries of Tilapia aureus and Mueil capito 15 ' 16 .
There are at present no reports in which these sex hormones
have been shown to sti -. -ulate maturity by direct action on the
ovary, but when Takano and Kasuga 6 administered 100/u.g/g of
methyltestosterone or ethinylestradrol orally to Oryzias
latipes for 20 days, they found an enormous increase of the
number of eggs in the yolk vesicle stage. The increase in the
ovarian index was much greater when chum salmon GTH and
diethylstilbestrol were simultaneously administered to rainbow
trout than when they were auministered separately. However
in neither of these experiments was there any great accumulation
of yolk, and the way in which vitellogenesis is related to the
action of sex hormones remains as an important subject for
future research.
•

82
1.2.3 The effects of administration of sex hormones.
The effects of administration of sex hormones depend
on the species. Even in the same species, they depend on the
condition of development of the gonads, the degree of
maturation, and the internal environment of the individual.
Even with the same internal environment, different results
are obtained with different types of hormones or with the use
of different concentrations. For example when 50),,tg/g of
methyltesterone was given to newly hatched rainbow trout for
five months, the development of the gonads was obstructed,
there was a complete absence of gametes and a sterile condition
was reached. However with 1 frg/g growth was stimulated, and
although the development of the ovary was inhibited in the
females, the males were clearly stimulated to mature. When
fry of the pink salmon Oncorhynchus gorbuscha were similarly
given 50 to 100 /kg/g in the feed the ovaries degenerated and
after two weeks the eggs which had been accumulating yolk had p45
completely disintegrated17
, but in Oryzias lijILing. the ovaries
were made to become active by a concentration of 100/g/g6
.
Kasuga18 suggests that at this concentration the cells in
Oryzias latipes which produce GTH were stimulated. Aida et al
injected 2 to 200 tAg/g per fish of estradio1-17p into the
abdominal cavities of Plecoglossus altivelis and found that
the amount of yolk material which appeared in the blood serum
increased in proportion to the amount of estradio1-17p
10

•
83
injected. However when more than an optimum concentration
of the female hormone was injected, maturation was inhibited.
This inhibition of maturation by a sex hormone can be
supposed to be the indirect result of the action of the
hormone in causing the sex control centre to inhibit secretion
of GTH from the pituitary. Such indirect influences on the
ovary will form subjects for future research.
The female sex hormones participate not only in
vitellogenesis but also in the thickening of the epithelium
of the oviduct. The male sex hormones participate in changes
in the pancreas and the digestive organs, and in salmonids
they also participate in changes of the skin thickness and
mucous secretion, in changes of body shape, in the relation
between muscle, blood serum and ovaries, and in the
carotenoid pigmentation of the skin. There are of course
queutions of the inherent natural balance between the two
types of hormones.
Hormones can be administered by injection or by being
included in the feed. Injection can easily damage the fish,
and is not easy to arrange when a large number of fish is to
be treated, so that if there is no need to determine the
exact dose administered to each fish, oral administration,
which is easy with large numbers, is much better than injection.
•

• 84
•
•
1.2.4 The sex control centre.
The sex control centre in fish is the diencephalic
hypothalamus. In particular the two important neurosecretional
nuclei are the nucleus preopticus in the anterior part of the
optic chiasma and the nucleus lateralis tuberis in the
protuberance of the hypothalamus close to the stalk of the
pituitary.
The GTH which is essential for the maturation of the
egg is secreted from the nucleous lateralis tuberis, and it
is believed to be under the control of a GTH releasing hormone
(GTH-RH) 19 . It is known that when the connections between the
pituitary and the hypo -:;halamus are severed, the secretion of
GTH is inhibited20
• 1.2.5 The thyroid hormone.
Changes in the thyroid gland parallel to the course
of maturation have been found in the goldfish and in other
fish. Thyroid activity is increased by the administration of
sex hormones, and the development of the gonads is inhibited
by excision of the thyroid and by the administration of anti-
thyroid materials. Maturation is reported21 to occur sooner
if thyroxin is administered, so it may be supposed that there
is a close connection between maturation and the thyroid
gland. The hormone which inhibits secretion of the thyroid
stimulating hormone (TSH-IH) and (GTH-Rh) are both secreted from
the nucleus lateralis tuberis 19 . They are similar in structure,

85
both TSH and GTH being glycoprotein hormones with molecular
weights of about 30,000, and though they differ in their
action on the control centre they are believed to have
common features, It is however doubtful whether the thyroid
hormone directly stimulates maturation by its action on the
ovarian tract, and it is thought that its action is a
secondary result of the stimulation of bodily growth. There
must be further investigation of the question.
2. Ovulation.
In ovulation the eggs which have completed
vitellogenesis are detached from the follicle and are
discharged into the ovarian cavity or into the body cavity.
Changes also occur in the egg itself, and with the maturation
division and the discarding of the polar bodies the diameter
of the egg is rapidly increased by the absorption of water22,23 .
In connection with ovulation the egg first becomes capable of
fertilization. Many studies in which the pituitary is
excised have shown that ovulation is caused by the secretion
from the pituitary of GTH, and that ovulation can be produced
in hypophysectomized fish by the administration of GTH. The
GTH producing cells in goldfish show extreme morphological
changes at the time of ovulation. For example, 10 hours
after goldfish which had matured in a pond at 13 ° to 14°C

86 •
•
were transferred to a tank at 20 ° C, the GTH producing cells
showed hypertrophy and vacuoles were formed in the cytoulasm,
and ovulation began to occur after 30 to 40 hours 24 . Although
there are few actual known facts about ovulation, it is
thought that ovulation follows the sudden secretion, about
20 hours earlier, of large amounts of GTH.
2.1 The amount of GTH in the blood at the time of ovulation.
It has recently become possible to measure the amount
of GTH in fish blood by means of radio-immuno-assay (RIA).
Breton et al25 used this method to measure the GTH in the
blood of goldfish and found the amount to be greatest during
the day, with 5.75 ng/ml at 8 a.m., 9.55 at 11 a.m. but
3.34 ng/ml during the night. They reported that the quantity
of GTH in the blood on the day of ovulation reached five times
that on an average day, and their results suggest that a large
amount of GTH is secreted by the pituitary at the time of
ovulation.
2.2 The mechanism of ovulation.
In considerations of the mechanism of ovulation it is
important to consider two sequences of events, those that
precede the secretion of GTH from the pituitary, and those
which accompany ovulation and are produced by the action of
the secreted GTH on the ovary.
External environmental factors which are of importance
to ovulation include the water temperature, the illumination,
the rainfall, the diurnal variation of illumination, and the

X/ERNAL
STIMULI. 7 Nuct.Eus _
reEOPTICUS..
HYF °THALAMUS
P.ROTUBER A.NCES
• • . • •• ADRENAL CORTEX
— —
• .. .
desoxycorticosterone
hydrocortisone •
•: .'0VAR IA N OR
ROOT CAVITY • •
:
% On- DUCT
Vital
WATER ADSORPTION AND
INCREASE OF DIAMETER
• 8 7
Figure 5.2.
DiaFram of the process of ovulation in fish.
•

• 88
•
weather conditions. It is believed that these stimuli affect
the sex control centre which by secreting GTH-RH causes the
secretion of a large amount of GTH from the pituitary.
Mammalian FSH releasing hormone (FSH-RH) and LH
releasing hormone (LH-RH) have been extracted from the
hypothalamus and purified. Their structural formulae have
been ascertained and it has become possible to synthesize
them26 . Studies by Hirose and Ishida27 of the effect on
ovulation of Plaecoglossus altivelis of mammalian LH-RH have
clearly shown the similarities between the mammalian and the
piscine sex control centres in their mode of action and in
the relation of LH to GTH p and this raises some very interesting
questions. It is important that the effectiveness of Li-RH
should be investigated in a number of species of fish. At the
same time GTH-RH should be extracted from these fish and p48
purified, so that synthesis can be used for comparative
chemical investigations.
Sundaraj et al28 have made a series of studies of the
way in which the action of GTH on the ovary gives rise to
ovulation in the catfish Heteropneustes fossilis. They find
that GTH does not act directly on the ovary, but that GTH
acts on the adrenal to stimulate the secretion of cortisol
and deoxycorticosterone, and these adrenocortical hormones
act on the ovary to cause ovulation. They claim that the
action goes from the pituitary to the adrenal and thence to

• 8 9
•
the ovary. The experiments of Hirose 29 with Oryzias latipes
in vitro suggest that there is a transfer from pituitary to
adrenal to ovary similar to that in the catfish, in addition
to a direct transfer from pituitary to ovary. We must wait
for future research to establish whether or not the hormones
of the adrenocortex participate in the ovulation in all
species of fish.
Jalabert et al30 made in vitro experiments with
rainbow troUt and found that both progesterone and fish GTH
affected ovulation, but that adrenccortical hormones, male
sex hormones, and female sex hormones had no effect. They
considered that GTH indirectly induced ovulatica either through
progesterone or by stimulating the secretion in the ovary of
some unknown substance. Hirose 29 observed the damage to the
follicular structure during ovulation, and suggested that
some substance contained in the follicles was the primary
cause of ovulation. There is no present certainty about this
substance, but when one takes into account the gaps which
appear between the follicle cells during ovulation, The
fluids which appear in these gaps, the loosening of the
adhesion between the follicle cells and the egg, and the
absorption of water by the eggs, it is true that the damage
to the follicle cells is very large. It has recently been

•
•
90 ••
suggested 31 that the GTH acts on the genes in the egg itself,
and that functional maturity of the egg is brought about by
the development of new proteins through a typical procedure
from DNA to RNA to protein.
3. Piscine GTH.
3.1 Types of GTH.
As has been shown above, GTH secreted by the pituitary
is essential to the maturation and ovulation of the egg, but
it is not yet clear whether the GTH which participates in
maturation is the same substance as the GTH which participates
in ovulation. In other words it is not clear whether piscine
GTH exists in one or in two varieties.
The following facts can be used to support the
existence of two types of GTH.
1. The maturation of the egg (vitellogenesis) and ovulation
differ completely in detail.
2. Mammalian LH, pituitary extract, and chorionic
gonadotropin (HCG) are effective in causing ovulation
in many species of fish but they are not found to have
any noticeable effect on maturation .
3. It has been stated that two types of GTH producing
cells are present in the ee 1 32 .
1)49
•

91
4. Even though the GTH producing cells in the goldfish are
of only one type, two very different types of granules
are present in the cytoplasm, and they are observed to
have different behaviours during maturation and ovulation.
5. It is believed that gametogenesis and sex hormone
secretion in Xiphophorus maculatus are under the
control of different pituitary hormones 32 .
6. Maturation and ovulation in mammalian ovarian follicles
are under the control of two types of hormones, FSH and
LH. This may be thought to support the idea that there
should be two types of GTH in the piscine pituitary,
with individually separate functions in maturation and
ovulation.
The following facts can be used to support the contrary
view, that only one type of piscine GTH exists.
1. The GTH extracted from the pituitaries of carp and chum
salmon are biochemically similar and are of the same
type of glycoprotein34 .
2. Purified chum salmon GTH is effective in producing
maturation of the egg and ovulation in the female fish,
and in producing spermatogenesis and spermiation in
the male 35 .
3. There is only one type of GTH producing cell in the
pituitary of Gasterosteus aculeatus, goldfish, Oryzias
• latipes, Oncorhvnchus keta and Oncorhynchus nerka32,36

••
92
•
4, The changes seen in the GTH producing cells in the
goldfish after ovariotomy and the changes seen at the
time of ovulation have in common the appearance of
vacuoles due to the enlargement of the endoplasmic
reticulum, and both the large and the small types of
granules are observed to participate in the formation
of these vacuoles 36 . This appears to support the
idea that maturation of the egg and ovulation are
controlled by one type of GTH.
Burzawa-Gerard and Fontaine37 propose that there is
only one type of piscine GTH, but that the whole of the
reproductive phenomena are controlled by the existence of
two types of receptors in the gonads. Hyder 38 suggests that
there is only one type of GTH but that there may be two
active centres in the GTH molecule. Whatever the truth about
the types of GTH may be, it is important for the artificial
control of reproduction that it should be found out rapidly
by means of further studies.
3.2 The effectiveness of piscine GTH.
Up to the present time mammalian hormones have been
used to stimulate maturation and ovulation. Examples are
gonadotropin, synaholin, puberogen and gonadoplex. They
affect ovulation in goldfish4 , carp39 , Misgurnus
anguillicaudatus 40 , Plecoglossus altivelis 41 , Heteropneustes •

• 93
fossilis28 , Mylio macrocephalus42 , Mugil cephalus 22 and
Seriola quinqueradiata43 , and also affect maturation in
Mylio macrocephalus. However in salmonids, sturgeon,
Chenopharyncodon idellus, Hypophthalmichthys molitrix and
Anguilla japonica it is difficult to observe any particular
effect of these hormones on either maturation or ovulation.
Yamamoto et a144 have recently achieved maturation and
ovulation in the Japanese eel Anguilla japonica by using salmon
pituitary, and young eels were born from artificial
fertilization for the first time in history. It is supposed
that all types of pituitary hormones were present in the
pituitary fluid suspension used, but it must be considered
that there is at least a suggestion that piscine pituitary or
its extracts can be effective in the maturation and ovulation
of species which are difficult to breed. .
3.3 The purification and chemical properties of GTH.
GTH has recently been extracted and purified from
carp37 , chinook salmon, chum salmon, and Puntius .0 .onionotus46
an :. its chemical properties are gradually being found. The
extract ha s been fractionated with water or dilute ethyl
alcohol (40 - 57%) using Simplex G - 75 or G - 100 and
DEAE cellulose 34 ' 36 ' 37 ' 45
Carp GTH is a glycoprotein containing hexose 8.6%,
hexosamine 4.9% and sialic acid 0.35%, and this amount of
•

• 94
•
sialic acid differs from that contained in sheep FSH or LH34 .
The molecular weight in carp is 27,000 to 31,000, and in
chinook salmon is 28,5000 to 29,400 34 . It has been inferred
that carp or chinook salmon GTH, like mammalian GTH, is
formed from two units 34 . The purified GTH is reported to be
effective both in egg maturation and in ovulation.
The hormones known to be of greatest importance in
egg maturation and ovulation are the pituitary GTH and the P51
sex hormones secreted by the ovary and the adrenal cortex.
The individual and joint actions of these hormones are
essential in the artificlal control of maturation and
ovulation for the production of fry of useful species of fish.
Up to the present commercially available mammalian hormones
have principally been used to stimulate maturation and
ovulation in fish. However there are natural limits to the
results obtainable in fish in which the pattern of reproduction
is different from that in mammals. There is a requirement
for further studies of piscine GTH and for a coordinated
series of investigations of the effective individual actions
and joint actions of the male and female sex hormones, and for
simultaneous practical applications of the results.
•

• 95
References.
1)
SEE PAGE 99
SEL PAGE 99
3) Su PAGE 69
4) F. YAMAZAXI: Endocrinological stud-
ies on the reproduction of the fe
male goldfish. Carassius auratus L., with special reference to the func-tion of the pituitary gland. Mem.
Fac. Fish.Hokkaido Univ. 13, 1-44
(1965).
5)
SEE r AGE 100
SEE PAGE 100
chez les poissons sélaciens et
téléostéens. Bull. Biol. France et Belgique 75, 257--309 (1941).
8) W. A. BARR: The endocrine control of the sexual cycle in the plaice. Pleuronectes platessa (L),II. The
endocrine control of oogenesis. Gen. ComP. endocrinol. 3, 205-.215 (1963).
9) B. L SUNDARARAJ and S. V. Coswenr:
Effeet of short-and long-terrn hypo-physectomy on the ovary and inter-renal of catfish, Heteropneustes
jos•ilis (Bloch). J. Exp. . Zoo!., 168.
58-104 (1968).
10)
SE.E PAGE 100
II) SEE PAGE 1 00
7) J. H. VtviEN: Contribution a l'etude
de la physiologie hypophysaire dans
ses relations avec l'appareil genital, la thyroide et les corps suprarenaux
12) B. I. SUNDARARÀJ and T. C. ANA»:
Effects of piscine and mammalian

gonadotropins on gametogenesis in the catfish, Heteroneustes fossilis
(Bloch). Gen. Comp. Endocrinol.
Supple 3, 688--702 (1972).
13) • W. S. Hoes : Reproduction. in "Fish
Physiolgy" (W. S. Hou & D. J. RANDALL ed) Vol. III. 1--72, Acad. Press, New York (1969).
14) R. REINBOTR: Hormonal control of the teleost ovary. Amer. zool. 12, 307 --324 (1972). B. ECKSTEIN: Metabolic pathways of steroid biosynthesis in o‘arian tis-sue of a teleost, Tilapia aurea. Gen.
Comp. Endocrinol. 14, 303--312(1970).
16) U. Eviieru and B. ECKSTEIN : Isola-tion of 11-ketotestosterone and dehy-droepiandrosterone from ovaries of the common mullet, Mugil capito
Gen.. Comp. Endocrinol. 12, 58-62
(1969).
17) F. YANAZAKI: Effects of methyltes-tosterone on the skin and the gonad
of salmonids. Gen.ComP. Endocrinol.
Supple. 3, 741--750 (1972) 1
SEE PAGE I 01
19) R. E. «PETER: Hypothalamic control • of thyroid gland activity and
gonadal activity in the goldfish. Gen.
Comp. Endocrinol. 11, 334 (1970).
20) .1. N. BALL, M. OLIvEREAu, A. M.
SLICIIER and K. D. KALLMAN: Func- tional capacity of ectopic pituitary transplants in the teleost Poecilia
fornwsa with a comparative discus-sion on the transplanted pituitary,
Phil. Trans. Roy.Soc. Lond. Ser. B..
249, 69--99 (1965).
21) M. SAGE :The evolution of thyroidal
function in fishes. Amer. Zool. 13,
899-905 (1973).
22) Z. H. SUET:LADEN and K. S. NORRIS:
The grey mullet (11fugil ceplzalus L.): induced breeding and larval rearing. Oceanic Inst, Hawaii RePt. No. 01-
72-764, 1-402 (1972). K. HIROSE, T. HIRANO and R. Dunne: Effects of salmon gonadotropin on. ovulPtion in the ayu, Plecoglossur
altivelis, with special reference-to water balance. Comp. Biochem.
Physiol. 97A 283--289 (1973).
24) K.YAMAMOTO and F. Yitueza :Hor-monal control of ovulation and sper-miation in goldfish. Gunma Symposia
on Endocrinol, 4, 131---.145 (1967). B. BRETON, R. BILLARD, B. JALABERV
et G. 'KANN: Dosage radioirnmunolo-
gigue des gonadotropines plasmati-
qus chez Carassius auratus, au cours-
du nycthémère et pendant l'ovulati- on. Gen. Comp. Endocrinol. 18, 461
-.-468 (1972).
26) SEC PAGE 101
SEE PAGE 102
28) B. SlINDARARAJ • and S. Goswiiii:
Role of interrenal in luteinizing
hormone-induced ovulation and spa-wning in the catfish Heteropneuster
fossilis (Bloch). Gen. Comp. Endo-
crinol. Supple. 2, 374-384 (1969).
96
15)
18)
23)
25)
27)

• 97
29) .
SEE PAGE 102 30) B. JALABERT, B. Bavrou and C. BRY:
Maturation et ovulation in vitro
des oocytes de la Truite Arcen-Ciel Salmo gairdncrii. C. R. Hebd.
Seances Acad. Sci Ser. D.275, 1139—
1142 (1972).
31) Effect of actinomycin D, mitornycin,
C, puromycin, and cydoheximide on desoxycorticosterone-induced in vit-
ro maturation in oocytes of the catfish, Hetcropneustes fossilis (Bl-
och). J. Exp. Zoo!. 185, 327-332 (19
73). 1421. P. SCIIREIBMAN, J. F. LEATFIERL-
*ND, and B. A. Mc KEowm Functional
morphology of the teleost pitui,dry
gland.Amer. Zoo1.13, 719-742. (1973)
33) Y. Ne.c.inemA: Histo-physiological
studies on the pituitary gland of some teleost fishes, with special
reference to the classification of hormone-producing cells in the-
adenohypophysis. Mem. Fac. Fish.
Hokkaido Univ. 21, (1973).
34) E. M. DONALDSON: Reproductive'
endocrinology of 'fishes. Amer. Zool.
13, 903-927 (1973). F. ?AMAZAXI and E. M. DONALDSON
: The effects of partially purified
salmon pituitary gonadotropin on spermatogenesis, vitellogenesis, and ovulation in hypophysectomized goldfish (Carassius auratus). Gen.
Comp. Endocrinol. 11, 292-299 (1968).
38) SEE PAGE 102 •
37) E. HURZAWA-GERARD and Y. A. FON-
TAINE: The gonadotropins of lower
vertebrates. Gen. Corn p, Endocrinol. Supple. 3, 715-728 (1972).
38) M. HYDER: Endocrine regulation of reproduction in Tilapia. Gen. ComP.
Endocrinol. Supple. 3, 729 ,-740 (1972).
39)
SEE PAGE 103
40) SEE PAGE 103
SEE PAGE 103
42) SEE PAGE 104
43) SEE PAGE 104
-44) SEE PAGE 104
45) E. M. DONALDSON, F. ?AlIAZAXI, H.
M. Dis and W. W. Putwto: Preparation of gonadotropin from
salmon(Oncorhynchus tschauelscha)
pituitary glands. Gen. ComP.
Endocrinol. 18, 469-481 (1972). 46) B. I. SUNDARARAJ, T. C ANIND and
V. R. P. Smut: Effects of Carp
S.V.Gosweur and B. I. SDNDARARAJ:
32)
35)
41)
•

• 98
pituitary fractions on vitellogensis
ovarian maintenance, and ovulation in hypophysectomized catfish, liete-ropneustes fossilis (Bloch). J. En-door, 54,87-98 (1972).
47) B. I. SUNDARARAJ, T. C ANAND and E. M DONALDSON:. Effects of partia-
Ily purified salmon pituitary gonado-
tropin on ovarian maintenance, ovulation and vitellogenesis
in hypophysectomized catfish,
11 et eropn eust es fossilis (Bloch). Gen Comp. Endocrinol ; 18, 102-.114
(1972).
•

99
•
1. HIBIYA Takashi.
Horumon ni yoru gyorui no seijuku, sanran no kontororu.
Suisan zoshoku 12 239 - 259 (1965).
T.
Hormone control of maturation and spawning in fish.
Fish breeding 12 239 - 259 (1965).
2. YAMAMOTO Kiichiro.
Seishoku, gyorui sein.
KAWAMOTO Nobuyuki hen. 232 - 271.
Koseisha Koseikaku Tokyo 1970.
K. Yamamoto.
Reproduction and fish physiology. (Edited by N. Kawamoto) pp 232 - 271.
Published by Koseisha Koseikaku. Tokyo 1970.
3. YAMAMOTO Kiichiro.
Gyorui jinko zoshoku ni kanren suru naibunpitsu
gakuteki kenkyu, dai ikkai taheiyo no suisan
zoshoku ni kansuru Nisso godo shinpojiumu.
Ronbun shu 13 - 39 Tokai dal 1973.
K. Yamamoto.
Endocrinological studies related to artificial
fish breeding.
Collected papers from the first joint Japanese-
Soviet Pacific symposium on fish breeding.
13 - 39, Tokai University, 1973.

100
5. YAMAZAKI Fumio.
Nijimasu no kikeigyo no iwayuru "kobaruto" ni tsuite
Nichi sui shi ko 17 - 25 (1974).
F. Yamazaki.
On the so-called "cobalt" deformed type of rainbow trout.
Bulletin of the Japanese Society of Scientific
Fisheries, ko 17 - 25 (1974).
6. TAKANO Kazunori, KASUGA Seiichi.
Medaka no ranso ni oyobosu sel suteroido toyo no eikyo.
Do zatsu 82 263 (1973).
K. Takano, S. Kasuga.
The influence on the ovary of the administration of
sex steroids to Oryzias latipes.
Japanese Journal of Zoology, 82 263 (1973).
10. AIDA Katsumi, Fan ban gan, HIBIYA Takashi. • Gyorui no seishoku sen seijuku ni kansuru seini
gakuteki kenkyu I. Sei shokusen seijuku ni tomonau
ayu no kessho tanpaku sosei no shiyusa.
Nichi sui shi 1091 - 1106 (1973).
K. Aida, Phan Van NEan, T. Hibiya.
Physiological studies on gonadal maturation of fishes.
I. Sexual differences in composition of plasma protein
in relation to gonadal maturation.
Bulletin of the Japanese Society of Scientific
Fisheries 22 1091 - 1106 (1973).
•

•
•
101
11 , AIDA Katsumi, HIROSE Keiji, YOKOTE Motoyashi and
HIBIYA Takahashi.
Gyorui no seishoku sen seijuku ni kansuru seini gakuteki kenkyu II. Seijuku oyobi esutorojen shori
ni tomonau ayu kanzo no soshiki gakuteki henka.
Nichi sui shi 12 1107 - 1115 (1973).
K. Aida, K. Hirose, M. Yokota and T. Hibiya.
Physiological studies on the gonadal maturation of
fishes II. Histological changes in the liver cells
of Plecoglossus altivelis following gonadal maturation and estrogen administration.
Bulletin of the Japanese Society of Scientific
Fisheries ,3.2. 1107 - 1115 (1973).
18. KASUGA Seiichi.
Sei suteroido toyo ni yoru medaka no nokasuitai seishokusen shigeki horumon sansei saibo no henka.
Do satsu 82 263 (1973).
S. Kasuga.
Changes in the cells which produce the pituitary
gonadotropin in Oryzias latipes due to the
administration of sex steroids.
.Japanese Journal of Zoology 82 263 (1973).
26. IGARASHI Masao.
FRF to LRH.
Taisha 8 33 - 40 (1971). -
•M. Igarashi.
FRH and LRH.
Metabolism 8 33 - 40 (1971).

• 102
27. HIROSE Keiji, ISHIDA Rikizo,
LH - RH ni yoru ayu no hairan sokushin ni tsuite.
Nichi sui eakkai 49 nen koen hoshi shu (1974).
K. Hirose, R. Ishida.
On stimulation of ovulation in Oryzias . latipes by means
of LH - RH.
Japanese Society of Scientific Fisheries.
Collected Abstracts of Papers, 51 (1974).
29. HIROSE Keiji.
Gyorui no hairan no naibunpitsu shikai.
Tokai sui ken ho 21 67 - 81 (1973).
K. Hirose.
Endocrine control of ovulation in fish.
Bulletin of the Tokai Fisheries Research Laboratory,
21 67 - 81 (1973).
36. YAMAZAKI Fumio.
Gyorui no seishokusen shigeki horumon.
Nichi sui shi 695 - 709 (1969).
F. Yamazaki.
Gonadotropin hormone in fish.
Bulletin of the Japanese Society of Scientific
Fisheries, .25. 695-709 (1969).

•
•
103
39. KAWAJIRI Minoru ., SHIMADACHI Magoi, kOYAMA Haiime,
MIYAJIMA Chojiro.
Horumon ni yoru koi no sanran sokushin shiken (dal
ichi ho).
Nichi sui shi 14 13 - 16 (1948).
M. Kawajiri, M. Shimadachi, H. Koyama and C. Miyajima.
Experiments on the stimulation of spawning in carp
by means of hormones (Part 1).
Bulletin of the Japanese Society of Scientific
Fisheries 14 13 - 16 (1948).
40. MATSUURA Yasuasa.
Dojo no jinko saibyo shiken.
Sui san zo shoku 65 - 73 (1967).
Y. Matsuura.
Experimental production of young of Misgurnus
anguillicaudatus.
Fish breeding 65 - 73 (1967).
41. ISHIDA Rikizo.
Seishokusen shigeki horumon toyo ni yoru gyorui no
seijuku oyobi hairan no sokushin ni kansuru kenkyu II.
Ayu ni okeru taiju 1g atari no tekisei toyorà . o to
seijuku ni tomonau sokushin koka no henka.
Tansuiken ken ho 22 49 - 58 (1972).
R. Ishida.
Studies of the stimulation of maturation and ovulation
in fish by the administration of gonadotropic hormones.
IL Variation of the effectiveness of stimulation of
maturation with the quantity per gram of body weight
administered to Plecoglossus altivelis.
Research reports of the Freshwater Fisheries Research
Laboratory 22 49 - 58 (1972).

•• • 104
42, KASAHARA Shogoro, HIBIYA Takashi.
Kurodai no ryubyo seisan ni kansuru kisoteki kenkyu I. Seishoku sen shigeki horumon toyo ni yoru seijuku
oyobi sanran no sokushin ni tsuite.
Hirodai sui chiku sangaku bu kiyu 105 - 111 (1967).
S. Kasahara, T. Hibiya.
Basic research on the production of fry of Mylio
macrocephalus I. Stimulation of maturation and
spawning by means of administration of gonad
stimulating hormones.
Bulletin of the Department of Aquaculture, Hiroshima
University, 2. 105 - 111 (1967).
43. UMEDA Shin, HIROZAWA Kuniaki, OCHIAI Akira.
Kochi ken Komame gyojo ni raiyu suru buri sanran gun
to shinahorin ni yoru seijuku sokushin ni tsuite.
Nichi sui shi 15. 446 - 450 (1969).
S. Umeda, K. Hirozawa, A. Ochiai.
Stimulation of maturation by means of synaholin in the
yellow tail schools which migrate to Komane (Kochi
prefecture) to spawn.
Bulletin of the Japanese Society of Scientific
Fisheries, 3_5. 446 - 450 (1969).
44 • YAMAMOTO Kiichiro, YAMAUCHI Kohei, KASUGA Seiichi.
Unagi no shoki hassei ni tsuite.
Nichi sui gakkai 49 nen koen yoshi shu 102 (1974).
K. Yamamoto, K. Yamauchi, S. Kasuga.
On the early development of the eel.
Japanese Society of Scientific Fisheries, Collected
Abstracts of Papers, 102, 1974.

• 105
•
•
Environment, maturation, and spawning.
6. Freshwater fish.
Hiroshi YOSHIOKA.
(Hakodate Branch, Hokkaido University of Education).
The physical and chemical characteristics of the
environments in which freshwater fish and marine fish live
are different, and it is therefore to be expected that the
influence of environmental factors on maturation and spawning
will differ.
The principal factors affecting the maturation and
spawning of freshwater fish are illumination and temperature,
and this paper will discuss the relation of these two factors
to maturation and spawning.
1. Illumination.
When considering the relationships between an animal
and such factors of its environment as the illumination,
water quality and water temperature, it is important to
remember that they have . been established during a very long
period extending over many thousands of generations. For
this reason a given quality of illumination may exert a
great influence on one animal, no influence at all on another,

106
•
and even a contrary influence on a third. For example,
animals which normally live in bright daylight become
deficient in vitamin D if the illumination is insufficient,
but fish which live deep in the sea where the illumination
is normally insufficient contain large amounts of vitamin D.
Another important matter when deciding whether or not an
animal is being affected by the illumination, is that it is
often impossible to base a decision on the reaction during a
short period. Experiments concerned with the effect of
illumination on gonadal maturation in both birds and fish
must be continued for at least a month, and in some species
a period of not less than several months is necJIssary.
Studies of the effect of illumination on plants began
long ago, and many phenomena have been described. Basic
studies with vertebrates other than fish, such as birds and
mammals, have been completed and have reached the stage of
practical application. In contrast to this, apart from a few
early studies in Canada and America by Hazzard and Eddy1 and P56
by Hoover and Hubbard2 on the acceleration of spawning in
salmon and trout by artificial light cycles, regular research
on fish has only been carried on since the 1960's. An animal
experiences many variations of light during its life, from
those of a short period of one day to those of long period of
one year. Not all of these are necessarily influential, but
the length of the diurnal cycle of illumination is often

• 107
important. In the reproduction of the higher vertebrates,
this influence is exerted through the annual recurrence at a
fixed season of a diurnal cycle with a set length of
illumination. For this reason, the most frequent means by
which illumination influences maturation and spawning is
through changes in the diurnal cycle.
Garner and Allard divided plants according to the
relation between flowering and length of day into long-day
type plants and short-day type plants, and Yoshioka3 has
divided fish according to the relation between maturation and
length of day into long-day type fish and short-day type fish.
Fish which are categorzed as belonging to the first type are
those which spawn during the long-day season from spring to
summer, and include the medaka Orysiàs latipes, the honmoroko
Gnathopogon caerulescens, the shiner and the marsh killifish.
In these species maturation and spawning can be caused to
occur early by means of illuminated periods longer than those
which would normally occur at an early season (Yoshioka4-7 ,
Hibiya et al8,9 , Harrington10,11 ).
Species of the second type, principally salmonids,
which spawn in the autumn when the diurnal illumination is
short, behave in the opposite way, and can be made to spawn
earlier than in natural conditions by shorter periods of
illumination (Corson12 , Nomura13 , Henderson14 ).

• 108
For example Hazzard and Eddy1 took brook trout
Salvelinus fontinalis which had finished spawning in October
and November and exposed them from the middle of January to
the end of April to periods of illumination longer than
those which would occur in nature. Then the natural light
was blocked off and the period of illumination was made
shorter than in nature, and a species which spawns in November
was made to spawn in July. Shiraishi and Takeda have found
the same behaviour in ayu, Plecoglossus altivelis15,16
It has thus been shown that illumination influences
the maturation of fish, and there are two possibilites.
Either the total quantity of light or the increase or
decrease of the illuminated period may be effective.
Yoshioka4 therefore made observations on medaka Oryzias
latipes. He found that increase or decrease of the illuminated
period did not control maturation, but that there was a P57
critical length, between 12 and 13 hours, for the illuminated
period. Maturation was observed only when the illuminated
period was longer than this critical length. Similar
critical lengths have been observed with other species of
fish, and for Plecoglossus altivelis the period is required
to be less than 12 hour s. for maturation. In natural
conditions, of course, the period of illumination is not
normally constant, and there is some danger that the results
of increase or decrease of the length of the day may be
overlooked. This increase or decrease should be considered

1 09
to have a secondary role, supplementary to other influences.
An interesting observation with Oryzias latines is that if
the fish which were spawning were subjected to an illuminated
period less than the critical length, the large oocytes of
diameter more than 0.4mm completely disintegrated, but if
they were then returned to an illuminated period longer than
critical the oocytes resumed growth and spawning occurred.
Changes of illumination can thus stimulate, inhibit
or retard maturation, but there is a problem of when, during
the process of maturation, the influence is exerted.
Yoshioka4 found that apart from the extreme condition of
extremely long illumination and short dark period, the
length of the illuminated period had no effect on ovulation
or spawning of Oryzias latipes and its food gathering
activities were not affected, but he confirmed histologically
that there was a remarkable influence on the development of
the oocytes (Figure 6.1). Changes in diameter have been used
to observe the influence of illumination on the development
on #-he ovarian egg in the willow shiner Gnathopogon elongatus
caerulescens, the bridled shiner Notropis bifrenatus, the
rainbow trout Salmo gairdnerii, the brook trout Salvellinus
fontinalis and the ayu Plecoglossus altivelis.
Oryzias latipes was used in a further investigation
of the result of making the illumination cycle longer or
shorter than 24 hours, and the effect on the critical length,

•
• 1 10
Figure 6,1,
The influence of the photoperiod on the development
of the ovary in medaka (Oryzias latipes).
•
1. The medaka ovary during oogenesis when exposed to a short illuminated period, 14 days after the start of the experiment. Large oocytes are degenerating.
2. The ovary under a short illuminated period (at a time of no oogenesis). Large cells have almost disintegrated and been absorbed. The ovary is occupied by small oocytes of diameter less than 0.4mm.
3. The medaka ovary when oogenesis has been suppressed by a short illuminated period. 21 days after the start of the experiment.
4. The ovary of (3) after 21 days exposure to a long • illuminated period.

Figure 6.1 continued.
r ---- 7 ..ore.,-7.1M77--;:- . . • ...
,„........_ ,..„,...,...„......,...„,..,,...,,.....,... •
, ..‹,,..„.....,,,..„.,..„ ,, . ,
,,.„....,....„_.;_„,...,.„,....,.,•.._ „.„.„,..., ..,,......„:„_.....,_,,„.......,7„..,.. ..,-_, ,.. e',..-:,,,,, Yuy 4-. .. ' dr';'H..-_;;',.. ..,.
, .', - - .= • ‘.' ...:? .: -. ;?9 1 i'r'I,t-V- ,,J.,.....' ..K.,.
,,':(::-....;.:,..e; ff-, l'..,..';'‘.>;f
"•-r--". 0, - -.v. :1,-,--4,-i'.‘ii'i ,4 ' -'.'''-'-- ,L..e., , _ ... • —
01
111
• ;
*-
4.
••• , (.1
bz.4 e\?1' '
.
-""Vr,,•p-,e?,1.-s . -- • .- - ;N-3' • • - --7e'e
,,à2.,•.?,'4, q---%_: -,... --'," ; i, 4 , .-.3.-. ?.....,C). ■ \I ej''''''Y e.‹. )• •••::. • i. t 1
t>e•Cic.e..:,•ei:,;.is2).,.Z.V:.,;.:,:i4.. ;keip:* „ .. «''''''-'0 et• ie ..,..!,..:y4 :.-='; -:,'-?-'".-)' 7 -
•
. . --"•-., - :•:,.t.f.m..-.-",.1,'-'"--- a
,--e...-1..,- ‘ e'. ...e% .K.f.:‘ pWr-,:b.:‘,4,,,,,te -,46.1c4::::,.'liu:'T.,.1
. . '",z7 ;15,ie-,...e.c,te,i, 1 .:-... 'f'■:e. `'. bi.:1-:-.-
..., ,r\:. ., . •. p .
. ‘ -,C:• ■ ,- . e.f,ocyi i:: Wi...
Ç. C::-.«:•"110:'1 'tic'. -.•-:-..--
I .. 1.,.:.-?' 'Zi..,-4 ■ *:ieft&o .'"?-1/j
5,6. The medaka ovary February, reared
in a period of no spawning in under a long illuminated period.
5. At 14 days.
6. At 21 days.
7,8. The medaka ovary in the period of no spawning in January, reared under natural photoperiod conditions.
5. At 14 days.
6. At 21 days.

112
••
when the cycle was lengthened so that the associated dark
period was even longer, maturation was not stimulated.
Conversely, when the dark period was longer than critical,
but the associated light period was even longer, maturation
was stimulated 7. Thus maturation can be stimulated or
retarded by changing the length of the cycle and by changing
the lit or dark periods. Hibiya et al8 have found
corresponding but inverse influences in Gnathopogon.
Furthermore Hibiya et al8,9 observed that there was
a range of periods in which the influence of illumination in
stimulating or inhibiting maturation was particularly strong.
This has not been observed in other species and is to be
awaited in further studies.
The next subject is the influence on maturation of
the strength of illumination. There exist only a few results
by Shiraishi with Plecoglossus 16 and by Yoshioka with Oryzia.
When Plecoglossus was illuminated by fluorescent light with a
16 hour illuminated period, maturation was inhibited in
females at 0.2 lux and in males at 0.11 lux. In Oryzia the
gonads were completely undeveloped with illumination less
than 5 lux, and with a low degree of illumination in the
range of 10 to 50 lux there was a slow and retarded
development of the gonads. The results were the same as in
natural illumination only with more than 160 lux, and there
•

• 113
was then no change in the rate of development of the gonads.
Up to the present there have been no observations of any
influence on maturation of illumination greater than the
critical amount, and the study of other species is awaited.
The effective critical illumination at which maturation is
inhibited may be thought to be rather low, but the sensitivity
to net changes, and the threshold value may be considerably
affected by variations of the environment in which the fish lives.
Light greatly influences spawning as well as maturation.
Oryzias normally spawns a little before dawn, and this shows
an influence of illumination on spawning. When day and night
were artificially internhanged, spawning occurred at the P59
beginning of the artificially lit period after a few days18 .
In order to find out the lowest illumination needed for
spawning, Yoshioka 19 caused Oryzias to stop spawning by
keeping them in darkness and then provided them with various
amounts of artificial light. He found that in illumination
greater than 5 lux all of the fish which were ovulating spawned,
but in illumination less than one lux some spawned and some
did not. Spawning was also induced by suddenly subjecting
individuals which were already well lit to a very strong light
(more than 5000 lux), and all ovulating individuals spawned.
This confirms that the occasional spawning of Oryzias during
the day or in the evening is the result of changes of illumination.
•

• 114
There are very few studies of the effect of wave-
length. Shiraishi 17 found that yellow and orange light was
more effective in controlling maturation than the short
wave-length blue and green, but these results were not
satisfactory because the filters used had very poor transmission
coefficients. Yoshioka19 used standardized liquid filters and
monochromatic filters, and tried the effect of selected wave-
lengths on the maturation of Oryzias. He found that wave-
lengths shorter than 5000 R had no effect on maturation, but
that visible light longer thal: 5000 R did have an effect. It
was interesting that visible light of wave-length 5800 R to
6700 R was particularly effective, and that nearby but shorter
wave-lengths had only a weak effect. The effects of infrared
and ultraviolet were tested with long periods of illumination,
but no effects were found. However the amount of light
passing the filters was said to be insufficient, and so the
experiments were not able to establish with certainty the
presence or absence of any effect. However, since the amount
of ultraviolet light at the çarth's surface is barely 1% of
the total, and since it is absorbed in water, it would be
very difficult for fish to utilize the ultraviolet in the
environment. In animals, other than fish, such the duck, the
development of the gonads is affected by wavelengths from
6640 R to 740 0 R. Conversely, it is known that the shorter
wavelengths in the blue and green are effective in insects.

115
It is supposed that these differences in the sensitivity of
animals are closely connected with the environment in which
they live. For example, the large amount of green and blue
light reflected from the leaves of the trees surrounding the
insects is in accord with the effect of these short wave-
lengths on their maturation. There are differences in fish
between the colour responses of the light sensitive cells in
freshwater fish and marine fish (the marine fish having
principally rhodopsin and the freshwater fish principally
polyrhodopsin), and it is known that the predominant
wavelength of the light depends on the depth in the water.
In deep places the light is generally blue or green,
(wavelength about 4700 R), in shallow places offshore there
is relatively a larger amount of long wave light. As a
result the deep sea fish are more sensitive to short
wavelengths than the shallow sea fish. Thus it is reasonable
to suppose that the pigmentation is appropriate to the light
conditions in the environment in which the fish live.
It is known that there are periods during which there
is no reaction to light, even though the illumination is
greater than the threshold for the species and is made up of
wavelengths which are believed to be effective. Harrington
found that in the bridled shiner Notropis bifrenatus the
spawning season could be made earlier by increasing the
length of the da y between mid-November and mid-,Tuly, but that

116
at the remaining season, from August to October, there was
no reaction to light. The period during which there is no
reaction is called the refractory period. Observation of
the refractory period in Oryzias showed that it is difficult
to get a' reaction to light for about one month after spawning.
However control of the length of the spawning season by means
of control of the illuminated period before the refractory
period begins makes it possible to some extent to shorten or
lengthen the refractory period. (Yoshioka 5 ). In both the
shiner and Oryzias the refractory period begins.immediately
after the spawning period. The metabolism of the fish is
very much reduced after a long spawning period of several
months, and it is thought that the refractory period is a
rest period of preparation for the following spawning period.
It is by no means clear how the external stimulus of
light can reach the gonads. It is supposed that the normal
channel for the effect will be from the light receptive organs
to the brain, from there to the hypothalamus, and then to the
pituitary and finally to the gonads, and it is believed that
it is the gonadotropin secreted by the pituitary which causes
maturation of the gonads. It is found that the gonads of
hypophysectomized Oryzias remain completely immature, even
if subjected to long days with long illuminated period, and
almost all individuals die within 30 days of the operation.
This shows that the pituitary has an important role and forms
part of the channel through which light affects the maturation
of the gonads.

•
•
1 17
2. Temperature.
Temperature is also an important factor which
influences the maturation and spawning of fish, and it has
been frequently studied. Since the living environment of
fish is in water, the environmental temperature range in which
life is possible is limited and is restricted to -2 ° to 35°C.
Since the range of temperature suitable for reproduction is
even narrower, temperature is frequently the factor which
sets a limit on reproduction. As an example, Oryzias latipes
will not mature at a temperature below 10 00 however fr2.vorable
the illumination conditions may be6 , and even in good
lighting conditions the minnow will not spawn below 7 °C
(Bullough20 ). The goldfish will not spawn at any season
unless the temperature is above 14 °C. In a detailed study of
the critical limits of temperature for Gnathopogon in which
the fish were cultured under a light cycle with an 18 hour
illuminated period and kept at various temperatures from 10 00
to 22 00 it was found that maturation occurred in all cases
except at 10 00. The greatest effect of temperature on
maturation occurred in the range from 16 00 to 1900 and
temperatures above or below this range resulted in a reduced
rate of maturation. In contrast, brook trout Salvelinus
fontinalis raised under natural illumination showed no
influence of temperature on maturation even with a temperature
difference from 8.5 00 to 16 00, and maturation was affected

• 118 ••
only when there were artificial changes between long-day and
short-day periods of illumination. Under short-day
conditions of illumination, maturation arrived sooner at 8.5 °C
than at 16 ° C, but the oocytes developed less rapidly than at
16 ° C. However, since growth continued longer than at 16°C
the eggs which had developed in the ovary were larger than
at 160 0. There are other examples of species in which the
effect of temperature changes when the light cycle is changed.
The minnow requires both a high temperature and a long
illuminated period for maturation, but when the light - cycle
is changed to a short day, the effect of a high temperature
is to inhibit development of the eggs. The sanie thing occurs
with the European bitterling and stickleback.
In the above examples both light and temperature are
limiting factors in maturation and *spawning, but there are
known to be species in which temperature alone is the main
factor in inducing maturation, anà illumination has no role
of importance. The development of the seminal glands in the
killifish Fundulus heteroclitus is not related to tl-„e light
cycle, and spermatogenesis proceeds at temperatures above 10 0 0
but is inhibited at 5.5 ° C. High temperatures from 16 ° C to
21 00 encourage proliferation of spermatogonia and spermiation
in the chub Couesius plumbeus, whereas low temperatures of 5 ° C
to 12 00 encourage the formation of spermatocytes and cause an

• 1 19
enlargement of the seminal glands 21 . It is very interesting
that such changes of temperature should have differing
effects on the process of spermatogenesis.
The relative importance of temperature and illumination
in regulating maturation and spawning of fish depends on the
species, and even in one species depends on the season of
reproduction. It may in general be said that there is a
narrow critical range for each species, and that circumstances
which make it easy to get outside this may be restrictive
factors. Temperature, as shown above, is the important factor
in the maturation of the goldfish, the killifish and the chub,
and illumination appea , 's to have almost no influence, but in
such cases it is to be thought that illumination is effective
over a wide range, and that it has vèry little opportunity to
operate as a limiting factor. For example Kazanskii22 found
that although illumination had been shown not to be important
to the killifish, the ovary did not develop under the most
extreme conditions of short illuminated period (one half hour
of light per day), even at the optimum temperature.
An important influence of temperature on maturation
and spawning is its contribution to the rate of oogenesis or
spermatogenesis. Yamazaki23 measured the changes of weight
of the ovaries of goldfish kept for three weeks from the end
of January into February in temperatures of 10 ° C, 15 °C, 20 ° C
and 25 °C. He found that the higher water temperatures within

• 120
•
•
his experimental range produced remarkable increases in ovary
weight. From a weight at the start of the experiment of
0.57g, the average increase at 10 °C was 0.4g, at 15 ° C 0.75g,
at 20 o C 12.0g and at 25 oC 13.5g . Yoshioka also found
greater increases in ovary weight in Oryzias with the higher
temperatures in the range of 10 °C to 16°C. Egami et al4
measured the rate of spermatogenesis in Oryzias by means of
thymidine labelled with H3 , and found that the interval
required for the cells synthesizing DNA (before meiosis) to
form sperm was about 20 days at 15° C and 12 days at 25° C,
showing that it was strongly influenced by temperature. In
general, oogenesis and spermatogenesis are accelerated by
high water temperature. However, according to Nomura, trout
which live in cold water show the inverse effect, and the
spawning season is earlier at low temperatures. However
according to Henderson's work on brook trout, the rate of
oogenesis is reduced by lowering the temperature, and since
the time taken for maturation becomes longer, cold water
water temperatures act agafaist maturation and spawning. This
must await further clarification in the future.
A further important action of temperature is its
effect on ovulation. K. Yamamoto et al25 found that
temperature plays an important part in the ovulation of the
* Sic. I have not seen the original paper, but could these be 1.20g and 1.35g? Translator.

121
goldfish, and that ovulation did not occur at temperatures
below 14 ° C however favorable the other conditions might be.
In fact, even though vitellogenesis proceeds to completion
in the oocytes in temperatures of 13 ° C to 14°C ovulation
does not occur, and even when males are present there is no
spawning. If these females are transferred to a tank at
about 20 ° C in the company of males, ovulation and spawning
occur in two days. According to Nagahama and Yamamoto26
great changes of the basophils in the adenohypophysis
precede ovulation, and secretory granules disappear. It is
clear from this that the influence of temperature on
ovulation is exerted through the pituitary, and results
from its effects on the secretion of the pituitary hormones.

References.
122
1) T. P. HAzzAan and P. E. EDDY:
Modification on the sexual cycle in,
brook trout, Salvelinus fontinalis by
control of light. Trans. Amer. Fish.
Soc., 80, 158-162 (1951). 2) E. E. Hoovait, and H. E. HUBBARD:
Modification of the sexual cycle in
trout by control of light. CoPeia, 4, 206-410 (1937)..
3) H. YOSHIOKA: On the effects of
environmental factors upon the
reproduction of fishes. 1. The effects
of day-length on the reproduction
of the Japanese killifish, Oryzias
latipes. Bull. Fac. Fish. Hokkaido
Univ., 13, 123-436 (1962). 4) H. YosinosA : Ditto. 2. Effects of
short and long day-lengths on Ory-
zias Wipes during spawnig season.
Ibid., 14, 137,-151 (1963).
5) H. Yostuose : Ditto. 3. The occurr-
ence and regulation of refractory
period in the photoperiodic response
of medaka, Oryzias latipes. J. Hok-
kaido Univ. Education, Ser. 2B 17,
23-43 (1966).
6) H. YosnioxA : Ditto. 4. Effects of
long photoperiod on the development
of ovaries of adult medaka, Oryzias
latiPes, at low temperatures. Ibid.,
21, 14- 20 (1970),
7) H. YOSHIOKA : Ditto. 5. The signifi-
cance of combinations of light and
dark periods in photoperiodic respo-
nse of the ovaries of medaka, Oryzias
latipes, in out-of-breeding seasons.
Seibutu Kyozai, 8, 76-82 (1971). 8)
SEE PAGE 124
9) SEE PAGE 124
10) R. W. HARRINGTON: SeXHal photo-
periodicity of the cyprinid fish, Noto-
pis bifrenatus(C,ope), in reation to
the phases of its annual reproductive
cycle. J. Exp. Zool., 135, 529-556 (19
57). 11) R. W. HARRINGTON : PreSeaSOnal
breeding by I he bridled shiner,
Notropis bifrenatus, induced uncle:
light-temperature control. Copeia, 304-311 (1950).
12) B. W. CORSON : Four year progress
in the use of artificially controled light to induce early spawning of
brook trout. Prog. Fish- Cult., 17, 99---102 (1955).
13)' SEE PAGE 125
14) N. E. HENDERSON. : Influence of light
•

25) 19) SEE PAGE 127 SEE PAGE 127
• 1 2 3
•
and temperature on the reproductive
cycle of the Eastern brook trout
Salvelinus fontinalis (Mitchill). J.
Fisheries Res. Bd. Can., 20, 859-897
_ (1963).
15), SEE P&GE 125
SEE PAGE 126
17)
SEE PAGE 126
18) N. EGAM1 : Effect of artificial photo-
periodicity on time of oviposition in
the fish, Oryzias • Wipes. Annot.
Zoo! , Jap., 27. 57-62 (1954).
21) S. N. ARSAN : Effects of temperature
and light on the cyclical changes
in the spermatogenetic activity of
the lake chub, Couesius plutnbeus
(Aoessiz). Can. J. Zool., 44, 161 ,-171
(1966).
22) B. N. 1{Kie.t;sxll : Experimental an-
alysis of the growth of oocytes in
fish. Dokl. Acad. Nauk. U. S. S. R.,
80, 277-280 (1951).
23) F. YAMAZAFI Endocrinolgical stud-
ies on the reproduction of the fe-
male goldfish, Carassius auratus L.,
with special reference to the funct-
ion of the pituitary gland. Mem. Fac-
Fish. Hokkaido Univ., 13, 1-64(1:45).
24)
SEE PAGE 127
16)
20) W. S. Bum.ouon: A study of the
reproductive cycle of thc minnow
in relation to the environment.
Proc. Zool. Soc. London, 109, 79-102
1939).
26) Y. NAGALTAMA and K. YAMAMOTO:
Basophils in the adenohypophysis
of the goldfish (Caraisius auratus).
Gunma Symposia on Endocrinol., 6,
39--59 (1969).

• 124
8 , HIBIYA Takashi ra.
Honmoroko no seijuku ni oyobosu suion narabi
ni hikari no eikyo.
Showa 47 nendo suisan gakkai shunki daikai koen happy°.
T. Hibiya et ai.
The influence of temperature and illumination on
the maturation of gnathougon elongatus caerulescens.
Papers presented to the Spring Meeting of the
Japanese Society of Scientific Fisheries (1972).
9. HIBIYA Takashi ra.
Honmoroko, Gnathopogon elongatus caerulescens (SAUVAGE).
no seijuku ni oyobosu suion narabi ni hikari
no eikyo III.
Koshu kanjusei no jikan ni yoru sai.
Showa 48 nen do nihon gakkai shunki daikai
koen happyo (1973).
T. Hibiya et al.
The influence of temperature and illumination
on the maturation of Gnathopogon elongatus
caerulescens (SAUVAGE).
Periodic changes in sensitivity to light.
Papers presented to the Spring meeting of the
Japanese Society of Scientific Fisheries (1973).

• 125
13. NOMURA Minoru.
Nijimasu no jinko sairan ni kansuru kiso kenkyu III.
Ko shiki no henka ni yoru sairan no sokika.
Nichi sui shi 28 1070 - 1076 (1962).
M. Nomura.
Basic research on the artificial collection of
eggs from Rainbow trout. III.
Changes of the illumination cycle and artificial
egg collection at an early 'period.
Bulletin of the Japanese. Society of Scientific
Fisheries. 28 1070 - 1076 (1962).
15. SHIRAISHI TAKEDA Tatsugo.
Ayu no seijuku ni oyobosu koshiki ni eikyo.
Tansui ku sui ken hohoku 11 69 - 81, (1961).
Y. Shiraishi, T. Takeda.
The influence of the illumination cycle on
maturation of ayu (Plecoglossus altivelis).
Reports of research of the Freshwater Fisheries
Research Laboratory. 11 69 - 81, (1961).
•

• 126
16. SHIRAISHI Yoshikazu.
Ayu no seijuku ni oyobosu koshiki no eikyo.
Dai 3 ho, shoshako no genkai shodo ni tsuite.
Tansui ku sui ken hohoku 11 69 - 76, (1965).
Y. Shiraishi.
The influence of the illumination cycle on
maturation of ayu (Plecoglossus altivelis).
Third report. On a critical quantity of illumination.
Reports of research of the Freshwater Fisheries
Research Laboratory 11 69 -76, (1965).
17. SHIRAISHI Yoshikazu.
Ayu no seijuku ni oyobosu koshiki no eikyo.
Dai 4 ho, shoshako no hacho to temmetsu shosha
no koka.
Tansui ku sui ken hohoku 11 69 - 76 (1965).
Y. Shiraishi.
The influence of the illumination cycle on
maturation of ayu (Plecoglossus altivelis).
Fourth Report. The effects of switching and of
wavelength of illumination.
Reports of research of the Freshwater Fisheries
Research Laboratory 11 69 - 76 (1965).

• 127
•
19 , YOSHIOKA Yasuko.
Kankyo to seijuku, sanran, tansuigyo.
Showa 49 nendo nihon suisan gakkai shunki dal kai.
Shinpojiumu koen happyo 1974.
Y. Yoshioka.
Environment, maturation and spawning. Freshwater fish.
Collected symposium papers, Spring Meeting of the
Japanese Society of Scientific Fisheries 1974.
24. EGAMI Nobuo, TAGUCHI Yasuko.
Medaka noseishi keisei sokudo ni oyobosu suion
no eikyo (yoho).
Shiken keitai gaku 21 500 (1968).
N. Egami, Y. Taguchi.
The predicted effect of temperature on
spermatogenesis in Oryzias latipes.
Experimental Morphology, 21 500, (1968).
25. YAMAMOTO Kiichiro, NAGAHAMA Yoshitaka, YAMAZAKI Fumio.
Kingyo no shunen sairan ni tsuite.
Nichi sui shi 12 977 - 983 (1966).
K. Yamamoto, Y. Nagahama, F. Yamazaki.
On a method for year-round collection of
goldfish eggs.
Bulletin of the Japanese Society of Scientific
Fisheries 32 977 - 983, (1966).
•

128
7. Marine Fish.
Tertio HARADA.
(Faculty of Agriculture, Kinki University).
It can be imagined that there is a close connection
between the maturation and spawning of marine fish and the
environment in which they live, but there is no detailed
account of the factors which are of importance for individual
species. Investigation of the environment is more difficult
than in the case of freshwater fish, and culture is not easy,
but the main reason for this lack of knowledge is that there
have been very few reports about maturation and spawning.
The intent of this paper is to describe experimental
work on maturation, spawning and egg collection which has
been undertaken by the author for more than ten years in the
Kinki University Fisheries Research Laboratory at Shirahama
in Wakayama prefecture. These studies have been based on the
black porgy (kurodai) Mylio macrocephalus, the porgy (kijinu)
latus, the red sea bream (madai) Chrysophrys major, the
porgy (hedai) Rhabdosargus sarba, the parrot bass (ishidai)
Oplegnathus fasciatus, the parrot bass (ishigakidai) Oplegnathus
punctatus, the yellow tail (buri) Seriola quinqueradiata, the
amberjack (kanpachi) Seriola purpurascens, the amberjack
(hiramasa) Seriola aureovattata, the jack (shimaaji) Caranx
delicatissimus, the sea perch (hirasuzuki) Lateolabrax latus,
and the Pacific fluke (hirame) Paralichthys olivaceus.

• 129
••
1. Water temperature, maturation, and spawning.
It is believed that temperature is the environmental
factor most closely connected to maturation. As is well
known, in natural conditions the gonads mature as the spawning
season approaches, and spawning soon begins. When the water
temperature during the spawning season is measured, it is
found that while there is vigorous activity the temperature is
well within the upper and lower limits of its range.
The natural spawning of yellowtail in the ocean has
been investigated by Uchida et al1 , by Mitani et a1 2 , by
experiments made by Harada et al3,4 on egg collection during
the 10 years from 1.960 to 1969 in the regions surrounding the
Orne islands and the Goto islands in Nagasaki prefecture, and
by Umeda et al5 on the spawning of yellowtail migrating to
the Komame fishing ground in Kochi prefecture. Most of the
spawning was from 17 ° C to 22 °C and the most suitable
temperatures appear to be from 18 °C to 20 0 0 . According to p68
Harada et al 6 , eggs can be collected from cultured yellowtail
by means of the injection of hormones when the water temperature
is 17 ° C to 21 0 0 . These authors cultured zero-year fish as
parent fish, rearing part of them in temperature-controlled
tanks, and part in netting fishpens offshore from Shirahama,
exposed to the natural temperature changes. Eggs were collected
by means of the administration of hormones and vitamin E, and •
samples of the 1970 experiments are shown in Table 7.1.

10 te t1
•
•
1 30
Table 7.1.
Records of the eggs obtained in 1970 by individual collection after the injection of hormones into yellowtail. 15 females and 5 males were used. When several fish spawned on the same day, 2 or 3 selected representative individuals are listed. The column headed "Number of floating egg..s" shows the number collected from a single parent yellowtail.
ATE. PLACE OF TEMPERATURE NUMBER OF RATE OF FER- NUMBER OF F ISH
REARING. AT TIME OF FLOATING TILIZATION FROM WHICH E3G
EGG COLLEC- EGGS. (%)* WERE TAKEN FOR
TIOW. FERTILIZATION. . -
LAND - ' - - -
W'12 17.3 193, 160 56.4 TANK 1
16.7 79,000 0 14 nm o
16.7 19,730 0 .
16 111.1
18.3 14,720 16.0 1
. NET FISH 18.0 199,260 9.3,
21 PEN.
18.0 345,600 -- 7.7 , 7
. 18.0 216,450 20.0 ,
Mu 18.1 93.100 54.2 22 11 .
18.1 40,000 25.0 . 18.0 147,400 81.5
M" 24 MA 201,040 76.7 .
18.0 26,200 84.7
tin 18.4 98,000 52.4 • 26 8
.18.4 155,000 35.2
,flt 18.4 10,920 84.4 28 8
' 18.4 144,000 . 66.7
tin
18.6 tWO 87.8 . 29 •
. 18.6 1,381 52.9 4
,10 1 19.0 39,900 28.3 ' ' 30
19.0 8,170 46.0 6
t”, 18.5 212,900 76.7 ,
V • 2 . • 18.5 209,000 55.3 4
• 18.5 187,350 36.0
18.3
18.3
20.3
20.3 wn
14 20.9
189,000
07,800
309,480
335,300
o
27.9
14.6 4
57.5
46.3
, 0 o
4

• 131
As can be seen from the Table, fertilized eggs were
not obtained below 17 ° C, and they were difficult to obtain
when the temperature was close to 21 00. Eggs could be
obtained from a large number of parent fish when the .water
temperature was between 18 ° C and 20 °C, and at these
temperatures the proportion fertilized was relatively large.
It was possible on rare occasions to obtain eggs from cultured
yellowtail even without giving injections of hormones or
vitamin E, but on such occasions the temperature was between
18 ° C and 20 °C. At temperatures below 16 ° C or above 22 00 no
eggs could be collected even with the use of hormones and
vitamins. Gathering together all these facts, it is evident
that temperatures suited to the spawning of yellowtail are
from 18 °C to 20 00. •
Experiments by Harada et al7,8with cultured parent
fish have shown that the temperature for active spawning of
Chrysophrys major, Mylio macrocephalus, and Rhabdosargus sarba
is almost exactly the same, from 17 °C to 21 °C. Accordinsx to
Fujita the puffer (torafugu) Spheroides rubripes spawns at
exactly the same time, and the author has found the same
temperature limits for the collection of eggs from cultured
karasu (gatora) . According to experiments with cultured parent
fish by Harada et al 10,11 , the temperatures required for active
* Karasu (gatora). I have not been able to identify this fish. "Karasu" is normally Rheinhardtius hippoglossoides, a coldwater fish. There is also the dogfish karasuzame, Etmopterus frontimaculatus, but some puffer species seems to be intended here, or perhaps a misprint for karasumibora, Mugil japonicus. Translator.

13 2
maturation and spawning of Oplegnathus fasciatus and . - Oplegnathus punctatus are 21 o C to 26 ° C, higher than those
for Chrysophrys and Spheroides.
Harada et al12,13 have also experimented with cultured
parent fish of Seriola purpurascens and Seriola aureovittata
and found that water temperatures for maturation and active
spawning were 20 °C to 25 °C.
Harada et al 14 collected eggs from fish in the open
ocean. Temperatures for the bonito (hagatsuo) Sarda orientalis
migrating to the Kii peninsula were 18 °C to 24°C. In further
experiments 15,16 the temperatures for the freight mackerels
(marusoda) Auxis than,rd and (marusoda)Auxis tapeinosoma were
18 °C to 26 00, and for the yellowfin tuna-(kihada) Neothunnus
macropterus the temperature was 26.2 0 0, so that the tuna was
by a little the warmest.
The spawning season for all these species is during
the warming up time from spring to summer, but some species
spawn during the cooling down period from autumn to winter.
The porgy (kijinu) Mylio latus spawns at just the same
temperatures as Mylio macrocephalus and Chrysophrys major, but
in the autumn. The jack(shimaaji) Caranx delicatissimus 18
matures at Shirahama in early winter from December to January,
and the Pacific fluke (hirame) Paralichthys olivaceus 19 spawns
from late winter to early spring in February and March. The
author's results from the spawning seasons and water
'temperatures of useful marine fish are listed in Table 7.2.

• 133
Parent fish culture.
Completely experimental.
tI
11 It
It
lu
11
It
It It
It
tl
Not completely.
Completely experimental.
It
ft
Table 7.2.
Water temperature and spawning seasons for useful marine fish.
Species
Paralichthys olivaceus
Chrysophrys major
Mylio macrocephalus
Rhabdosargus sarba
Seriola quinqueradiata
Sphreoides rubripes
Karasu
Oplegnathus fasciatus
Oplegnathus punctatus
Seriola purpurascens
Seriola aureovittata
Sarda orientalis
Auxis thazard
Auxis tapeinosoma
Neothunnus macropterus
Mylio latus
Caranx delicatissimus
Spawning season.
Feb -May
Mar-June
Mar-June
It fir
Apr -May
It It
May -July
11
May-June
II It
May-JuIy
May-Aug
11
June-Aug
Oct-Nov
Dec-Jan
Spawning temperature
1 4- 17 ° C
17-21 °C
17-21 ° C
▪ 0 0
• It 0
17-20 °C
ft
21-26°C
22 -25 °C
It It • It
18-24°C
18-26°C
II II II
24-28 °C
21-17° C
15-19°C
* See footnote, page 131. •

134
•
The temperatures in which marine fish are reared are
closely connected with their maturation. In the author's
experiments lasting over ten years on the rearing and maturing
of parent . fish of Seriola quiqueradiata, S. purpurascens,
S. aureovittatus and Chrysophrys major it has been found that
if there is a period of extremely cold temperature or extremely
warm temperature within several months before the expected
spawning season, and if the period during which spawning is
stimulated by change of temperature to the proper value is
short, it is difficult to obtain good quality eggs. In extreme
-cases, absolutely no eggs can be collected. This shows that
disturbance of the environment in which marine fish are reared,
may cause difficulty in maturation.
2. tj_ol i*Illumina- lmonandF_•_pa.ymi__na.
One environmental factor closely connected to the
maturation and spawning of marine fish is the illumination.
There are a good many reports about the influence of
photoperiodicity and day-length on the maturation and spawning
of freshwater fish, and marine fish will probably be similar.
The fact that Chrysophrys major and Mylio macrocephalus mature
and spawn in the spring whereas Mylio latus matures and spawns
at the same temperature in the autumn probably shows the
influence of illumination. In the spring the temperature is
rising and at the same time the illuminated part of the day is
lengthening, but in the autumn the temperature is falling and
the illuminated part of the day is becoming shorter.

• 135
Light is also closely connected to the time of
spawning during the day. Cultured Chrysophrys major, Mylio
macrocephalus, Rhadosargus sarba, Oplegnathus fasciatus,and
Oplegnathus punctatus almost always spawn between 3 pm and
10 pm. There is greatest activity around sunset. The eggs,
when discharged, float nearby on the surface of the sea and
directly after spawning they are crowded together, but then
are scattered by winds and waves and spread out. When eggs
are discharged in the light before sunset small fish such as
jack mackerel (maaji) which are in the vicinity gather
,together and they can be seen to be eating the eggs. As the
eggs are dispersed and become less crowded the amount lost
and eaten by small fish should be small. The frequent
spawning between sunset and nighttime may perhaps be an
instinctive use of the diminution orlight during the long
night for shelter against this loss by predation.
3. Other environmental factors, maturation and spawning.
Other environmental factors related to maturation and
spawning are the quantity of dissolved oxygen, the salinity,
noxious substances, and available space. If eggs of high
quality are to be obtained, the amount of dissolved oxygen
must not only be maintained, but an environment high in oxygen
saturation must be provided. There are no published reports
about the influence of low salinity on the maturation or

•• • 136
spawning of individual species, but in the case of the
yellowtail the buoyant eggs will probably sink if the
salinity is low and the density falls below 1.021, so that it
appears necessary to maintain a suitable salinity for spawning.
The environment will be degraded if dangerous substances,
accumulations of excrement or red tide are allowed to flow in,
so it can easily be supposed that maturation and spawning will
be adversely affected. It has also been demonstrated in
experiments on the culture of parent fish that normal
development is not possible if the rearing place is tifo
confined, and maturation and spawning will be hindered.
In short, a healthy environment must first be provided,
and then the conditions directly related to maturation and
spawning must also be provided. However further studies must
be awaited before a general account . can be given of the
relation of these individual conditions to maturation and spawning,
4. The culture of parent fish for eggcalleninE.
Eggs for the production of marine fish fry m,ly be
collected from wild or cultured parent fish. The disadvantages
of the use of wild fish include the uncertainty of the capture
of the parent fish, the relatively large expenditure needed
and the few opportunities for obtaining good quality eggs.
On the other hand, the use of cultured fish requires a long
time (often up to three years), and research on methods of
P7

• 137
•
maturation and egg collection is required, but it makes
possible a planned production and collection of reliable and
good quality eggs. Harada et al have experimented with the
culture of parent fish of Chrysophrys maior 7, Mylio macrocephalus 7
,
Rhabdosargus sarba8 , and Oplegnathus fasciatus 10 and have
found that eggs of better quality than those obtained from
wild parent fish could be reliably produced. With Seriola
guinqueradiata6, Paralichthys olivaceus 19 and Lateolabrax latus
there was no difference between the natural eggs and those
whose production had been pl,nned. Although it was not possible
to collect eggs from wild Seriola purpurascens 12 , Seriola
aureovittata13 , Caranx delicatissmus 18 , and Oplegnathus
Eunctatus 11 , it was possible to collect them from cultivated
Parent fish. It is considered that .future reliable production
of large quantities of fry will necessitate planning the
reliable supply of good quality eggs from cultured parents.
With the improved experimental netting fish pens which
Harada20,3 , has been using (in small sizes) since 1954 for
rearing quarters, the fish are reared in conditions close to
the natural conditions in the ocean. The culture density is
relatively high, the expense is low, they can easily be set up,
the fish can easily be harvested, and the fish can be moved
around in their normally submerged condition. These pens
have so many advantages that they are considered to be

138
appropriate places for rearing parent fish for egg collection.
Rearing places on land have the disadvantages of high building'
costs and high power costs, but they have advantages in the
gathering of naturally spawned eggs and in environmental control.
5. The stimulation and inhibition of maturation and spawning
by environmental control.
When parent fish are cultured, environmental control
can be used to stimulate or inhibit maturation so as to obtain
the desired quantity of eggs at the desired time. The period
during which fry are produced can be extended and their
production thereby prolonged. Harada et al21 reared Chrysophrms
major, Mylio macrocephalus, Seriola quinqueradiata, Oplegnathus
fasciatus and Oplegnathus punctatus in controlled environments
and found that eggs could be obtained earlier than the natural
season by raising the temperature. In these authors'
experiments on early egg collection from Chrysophrys major
from 1969 to 1974, naturally spawned eggs were obtained in the
middle of February, 1 to 1i months early. Figure 7.1 shows
an example of the relation between water temperature and
natural spawning of Chrysophys major reared in heated tanks at p72
the Shirahama Laboratory of Kinki University. It can be seen
in Figure 7.1 that when the water was gradually warmed,
natural spawning began when the neighbourhood of 17 ° C was
reached. When the temperature was lowered to 16° C spawning

139 •
• 1
■-•
a) e-1
4-)
(1)
a) 1 - 4-)
3 -
NATURAL
SPAWNING•
, NATURAL
SPAWN I NG.
NATURAL
. SPAWNING. %.
• NATuRAL
.4- SpAWNING •
NATuRAL ,"P•
SPAWNING;
NATURAL
SPAwNING,\Z,
,INATURAL7
SPAWNING.. •
Figure 7.1.
Culture water temperature and natural spawning of
Chrysophrys major in a heated tank.
10 15 20 15 20 25 1 - - 5
February March
Date.

140
stopped, and on rewarming to the neighbourhood of 17 ° C spawning
began again. Thus it is evident that spawning can be
controlled by control of the environmental water temperature.
Through the cooperation of the Owase City central Sanda steam
driven electricity generating plant from November 1967 to
April 1968, the authors obtained eggs from Seriola
quingueradiata by means of hormone administration on 4 April,
one half month early. Experiments were also made in the
Shirahama Laboratory heated tank, and an example of the
results is shown in the column for the tank on land in
Table 7.1. This shows that at the temperature of 16.7 °C no
fertilized eggs were obtained, but that thy were obtained
at 17.3 ° C and 18.3 ° C. From this it can be seen that Seriola
quinqueradiata can also be made to spawn early by controlling e 7.5
the environment to the optimum spawning temperature from 17 ° C
to 19 ° C. The authors have obtained similar results and
similar possibilities of early spawning with Mylio macrocephalus,
Oplegnathus fasciatus, and Oplegnathus punctatus.
The authors also found that a rapid increase of water
temperature was an effective method of stimulating spawning
in Chrysophrys major and Opleenathus fasciatus. The
temperature of water in which fish thought to be near spawning
were living was suddenly raised by 2 ° C to 3°C.
•

• 141
•
Tanks on land are considered to be appropriate as
rearing places for parent fish from which eggs are to be
obtained by means of control of the environment. Their
advantages include convenience of environmental factors such
as light, temperature, dissolved oxygen and salinity, the
possibility of preventing the entry of red tide and of
pollutants, and the easy collection of naturally spawned eggs
in nets stretched across the water outlets.
Other important points are:-
1. The exchange flow of sea-water requires power an6 is
expensive. This is particularly true with large tanks and
with those established in high locations.
2. The tanks must be suitable for the species used. In
particular, tanks must be deep and wide when large fish are
to be cultured, and this implies high construction costs.
3. Fish can be damaged as they are moved from the sea to
a tank on land. There is a need for improved ways of
capturing easily damaged species such as tuna, bonitos
and jacks.
4. Fish diseases can easily develop in land tanks. Epidemics
can easily cause great harm. Also, in many species, stress
may impede maturation.
5. Control of the rearing and environment by heating or
Cooling consumes a great amount of energy and is expensive.

• 142
6. Future prospects.
The planned production of large quantities of marine
fish fry requires the culture of parent fish to produce good
quality eggs which can be collected at the desired time. Up
to the present parent fish of more than ten species, such as
Chrysophrys maior, Seriola cuinaueradiata, Oplegnathus
fasciatus, Caranx delicatissimus and Paralichthys olivaceus
have been shown experimentally to be suitable for culture.
The individual environmental factors needed for maturation
and spawning are not known, and there are some species, such
as tuna and bonito, which have not been experimentally shown
to be suitable . In order that the technology needed for
economic culture of parent fish may be developed, more
detailed studies are needed of the relation between maturation
and spawning and the environmental and other factors concerned,

• 143
•
ReferenceS.
1. UCHIDA Keitaro, MICHIZU Yoshie, MITO Toshi, NAKAMURA Kantaro.
Buri no sanran oyobi shoki seikatsu shi.
Kyushu daigaku nogaku bu gakusei zasshi
16 (3) 336 - 337 (1958).
K. Uchida, Y. Michizu, T. Mito, K. Nakamura.
Spawning and early life history of the yellowtail,
Seriola ouinaueradiata.
Science Journal of tue Faculty of Agriculture,
Kyushu University, 16 (3) 336 - 337 (1958).
2. MITANI Fumio.
Buri no gyogyo seibutsu gakuteki kenkyu.
Kinki daigaku nogaku bu kiyo 1 211 , 212 (1960).
F. Mitani.
Biological study of the yellowtail (Seriola
quinqueradiata) fishery.
Bulletin of the Faculty of Agriculture, Kinki
University, 1 211 - 212 (1960).
3. HARADA Teruo.
Buri no zoshoku . ni kansuru kenkyu.
Kinki daigaku nogaku bu kiyo 2 40 - 54 (1965).

144
3. T. Harada.
Study of reproduction in yellowtail (Seriola
quinqueradiata).
Bulletin of the Faculty of Agriculture, Kinki
University. 1 40 - 54 (1965).
4. FUJITA Yaro, MICHIZU Yoshie, HARADA Teruo.
Horumon shigeki ni yoru buri no jinko sairan.
Nihon suisan gakkai showa 40 nendo shuki daikai
koen yoshj 15 (1965).
x._Eainp„, Y. Michizu, T. Harada.
Artificial collection of eggs from yellowtail
(Seriola quinqueradiata) by means of the
administration of hormones.
Abstracts of papers, autumn meeting (1965) of the
Japanese Society of Scientific Fisheries 15 (1965).
5. UMEDA Susumu, HIROZAWA Kuniaki, OCHIAI Akiru.
Kochi ken Komame gyojo ni raiyu suru buri sanran
gun to shinahorin ni yoru seijuku sokushin ni tsuite.
Nichi sui shi .m 446 - 449 (1969).
S. Umeda, K. Hirozame, A. Ochiai.
On the acceleration of maturation by means of synahorin
in the spawning shoals of yellowtail (Seriola
• quinqueradiata) which migrate to the Komame fishing
ground, Kochi prefecture.
Bulletin of the Japanese Society of Scientific Fisheries,
31 446 - 449 (1969).

1 4.5
6. HARADA Teruo, KUUI Hidemi, kIZUNO Kenpachiro,and
MURATA Osamu, NAKAMURA Motoji.
Yosei buri kara no sairan, jinko fuka ni tsuite.
Nihon suisan gakkai showa 42 nendo shuki daikai
koen yoshi 20 (1967).
T.Harada, H.Kumai, K.Mizuno, 0.Murata, M.Nakamura.
Collection and artificial hatching of eggs from
cultured yellowtail (Seriola quinqueradiata).
Abstracts of papers, autumn meeting (1967) of the
Japanese Society of Scientific Fisheries 20, (1967).
7. HARADA Teruo, KUMAI Hidemi, UMEDA Susumu.
Madai oyobi kurodai no shingyo no yosei ni tsuite.
Nihon suisan gakkai showa 39 nendo shuki daikai
koen yoshi 27 (1964). -
T. Harada, H. Kumai, S. Umeda.
Culture of parent fish of Chrysophrys major and
Mylio macrocephalus.
Abstracts of papers, autumn meeting (1964) of the
Japanese Society of Scientific Fisheries 27, (1964).

•
•
146
8. HARADA Teruo, MURATA Osamu, KUMAI Hidemi.
Hedai no shingyo yosei, jinkofuka oyobi shigyo
ikusei ni tsuite.
Nihon suisan gakkai showa 42 nendo shuki daikai
kOen hoshi 20 (1967).
T. Harada, O. Murata, H. Kumai.
On the culture of parent fish, the artificial hatching
and the rearing of fry of Rhabdosargus sarba.
Abstracts of papers, autumn meeting (1967) of the
Japanese Society of Scientific Fisheries 20 (1967).
9. FUJITA Yaro.
Yogyogaku kakuron. 554 - 562.
Koseisha Koseikaku (1967).
Y. Fujita.
Complete pisciculture. 554 - 562.
Published by Koseisha Koseikaku (1967).
10 KUMAI Hidemi, NAKAMURA Motoji, HARADA Teruo.
Ishidai no shingyo yosei, jinko fuka oyobi shigyo
ikusei ni tsuite.
Nihon suisan gakkai showa 42 nendo shuki daikai
koen yoshi 20 (1967).
•

• 1 47
10. H. Kumai, M. Nakamura, T. Harada.
On the culture of parent fish, the artificial hatching
and the rearing of fry of Oplegnathus fasciatus.
Abstracts of papers, autumn meeting (1967) of the
Japanese Society of Scientific Fisheries 20 (1967).
11. HARADA Teruo, KUMAI Hidemi, MIZUNO Kenpachiro,
NAKAMURA Moto.11, MIYASHITA Mori, FURUTANI Hideki.
Ishigakidai no jinko fuka to shigyo no shiiku.
Nihon suisan gakkai showa 45 nendo shuki daikai
koen yoshi 62 - 63 (1970).
T. Harada, H Kumai, K. Mizuno, M. Nakamura,
M. Miyashita and H. Furutani.
Artificial hatching and rearing of fry of
Oplegnathus punctatus.
Abstracts of papers, autumn meeting (1970) of the
Japanese Society of Scientific Fisheries 62-63 (1970).
12. HARADA Teruo, MURATA Osamu, MIZUNO Kenpachiro,
FURUTANI Hideki, KUMAI Hidemi, NAKAMURA Moto;li.
Kanpachi no shingyo yosei, jinko fuka shigyo shiiku.
Nihon suisan gakkai showa 42 nendo shuki daikai koen (1970).
T.Harada, 0.Murata, K.Mizuno, H.Furutani, H.Kumai, M.Nakamura.
Artificial hatching and rearing of fry of
Seriola purpurascens.
Papers of the autumn meeting (1967) of the Japanese
Society of Scientific Fisheries (1970).

•
•
148
13. HARADA Teruo, MURATA Osamu, MIYASHITA Mon.
Hiramasa no shingyo yosei, sairan, jinko fuka,
shigyo shiiku.
Nihon suisan gakkai showa 47 nen shuki daikai
koen yoshi chu 308 (1972).
T. Harada, O. Murata, M. Miyashita.
Culture of parent fish, egg collection, artificial
hatching and rearing of fry of Seriola aureovittata.
Collected abstracts of papers, autumn meeting (1972)
the Japanese Society of Scientific Fisheries 308 (1972).
14. HARADA Teruo, MURATA Osamu, MIYASHITA Mon.
Hagatsuo no jinko fuka to shichigyo no shiiku.
Kinki daigaku nogakubu kiyo 1 - 4 (1974).
T. Harada, O. Murata, M. Miyashita.
On the artificial hatching and rearing of fry of
Sarda orientalis.
Bulletin of the Faculty of Agriculture,
Kinki University, 2. 1 - 4 (1974).
15. HARADA Teruo, MURATA Osamu, MIYASHITA Mûri.
Hirasoda no jinko fuka to shichigyo no shiiku ni tsuite.
Kinki daigaku nogakubu kiyo 6 109 - 112 (1973).
T. Harada, O. Murata, M. Miyashita.
On the artificial hatching and rearing of fry
of Auxis thazard.
Bulletin of the Faculty of Agriculture,
Kinki University, 6 109 - 112 (1973).

• 1 49
16. HARADA Teruo, MURATA Osamu, MIYASHITA Mori.
Marusoda no jinko fuka to shigyo shiiku ni tsuite.
Kinki daigaku nogakubu kiyo 6 113 - 116 (1973).
T. Harada, O. Murata, M. Myashita.
On the artificial hatching and rearing of fry of
Auxis tapeinosoma.
Bulletin of the Faculty of Agriculture,
Kinki University, 6 113 - 116 (1973).
17. HARADA Teruo, MIZUNO Kenpachiro, MURATA Osamu,
MIYASHITA Mon, FURUTANI Hideki.
Kihada no jink‘ , fuka to shigyo shiiku ni tsuite.
Kinki daigaku nogakubu kiyo 4 145-151 (1971).
T. Harada, K. Mizuno, O. Myashita, H. Furutani.
On the artificial hatching and rearing of fry.
of Neothunnus macropterus.
Bulletin of the Faculty of Agriculture,
Kinki University, 4 145 - 151 (1971).
18. HARADA Teruo, MURATA Osamu, MIYASHITA
Shimaaji no shingyo yosei, sairan, fuka, shichigyo
no shiiku.
Nihon suisan gakkai showa 48 nendo shuki daikai
koen yoshi 65 (1973).

• 15 0
•
18. T. Harada, O. Murata, M. Miyashita.
The culture of parent fish, egg collection, hatching
and rearing of fry of Caranx delicatissimus.
Abstracts of papers, autumn meeting (1973) of the
Japanese Society of Scientific Fisheries 65 (1973).
19. HARADA Teruo, MURATA Osamu, MIZUNO Kenpachiro,
NAKAMURA Motoil, MIYASHITA Mon, FURUTANI Hideki.
Jinko fuka hirame no ikusei to seijuku ni tsuite.
Nihon suisan gakkai showa 44 nen shuki daikai
koen yoshi 1 (1964).
T. Harada, 0.Mur, -tta, K.Mizuno, M.Nakamura, M.Miyashita,
H. Furutani.
Rearing and maturation of artificially hatched
Paralichthys olivaceus.
Abstracts of papers, autumn meeting (1969) of the
Japanese Society of Scientific Fisheries (1964) 41*.
20. HARADA Teruo, KUMAI Hidemi.
Gosei seni gyomo shiyo ni yoru burl no ikesu
ami yosei ni tsuite.
Nihon suisan gakkai showa 34 nen shunki daikai
koen yoshi 44 (1959).
* Sic, though at least one of the dates must be in error.
•

•
151
20. T. Harada, H. Kumai.
On the culture of Seriola quinqueradiata in fish
pens made of artificial fibres.
Abstracts of papers, spring meeting (1959) of the
Japanese Society of Scientific Fisheries 44 (1959).
21. HARADA Teruo, KUMAI Hidemi, MIZUNO Kenpachiro,
NAKAMURA Motoji, MIYASHITA Mon, FURUTANI Hideki.
Buri, madai, ishidai, ishigakidai kara no kaon ni
yoru soki sairan.
Nihonsuisan gakkai showa 45 nen shyki daikai
koen hoshi 62 - 63 (1970).
T. Harada, H. Kumai, K. Mizuno, M. Nakamura, M.Mimashita
and H. Furutani.
On early egg collection from Seriola quinqueradiata,
Chrysophrys major, 02.1egnathus fasciatus and
Oplegnathus punctatus by means of warming.
Abstracts of papers, autumn meeting (1970) of the
Japanese Society of Scientific Fisheries
62 - 63, (1970).
•

•
152
IV. Maturation and Metabolism.
8. Maturation and fat metabolism.
Fumio TAKASHIMA.
(Tokyo University of Fisheries).
Many differing metabolic processes occur during the
development of the egg. Two which can be considered as
particularly important are the synthesis of the proteins
which comprise the embryonic tissue and those which generate
energy. In the eggs of the rainbow trout the amount of
acetone-soluble fat rapidly diminishes during development1 ' 2 ,
particularly at the time when movement becomes vigorous after
hatching2 . The fats are also diminished in the eggs of
Oryzias latipes just before hatching 3 , and in salmon eggs at
the time when the eyes deve1op4,5 . In all cases it appears
that fats are concerned in the generation of energy during
development. Fats are however not concerned solely in the
generation of energy, but are also concerned in the development
of the embryo. The phospholipids in the eggs of Oryzias
latipes diminish up to the moment of hatching, but it is
believed that after dissolution they are transferred to the
embryonic tissues 3. In these ways fats play important roles
in the course of development of the egg, and it is necessary
that the deposits of fat in the mother's body during this
development should be both qualitatively and quantitatively

• 153
sufficient. From the viewpoints of fat chemistry and of food
chemistry, much is already known about the fats in fish eggs,
but there is insufficient knowledge of the physiology of fat
accumulations and of the control mechanisms. The present
paper seeks to summarize the facts about fat metabolism in
relation to oogenesis from this physiological point of view.
1. The chemical properties of fish egg fats.
The overall analyses of the mature eggs of several
species of fish are shown in Table 8.1. The proportions of
the various components depend on the species but there is
more protein and less fat than in hen eggs. Each egg of the
Atlantic salmon Salmo salar contains 14.6mg of fatty tissue,
of which 67.9% is acetone-soluble fat, a large amount in
comparison to the 32.1% of acetone - insoluble fat . In chum p77
salmon, the egg contains 29.8mg of fat, of which as much as
75% is acetone-soluble. The acetone-soluble fats in the chum
salmon eggs are mostly glycerides, the acetone insoluble fats
which form 23% are compound fats in which the principal
components are phospholipids. The rainbow trout egg contains
10.7e fat, of which 6.4% is neutral fat and 4.3% is complex
fat. Thus fish eggs fats contain great amounts of glycerides
and complex fats, and the complex fats are mostly phospholipids,
the main comnonent being lecithin11 .

•• • 154
•
Table 8.1.
General analysis of fish eggs
Species Water Protein Fat Carbo- Ash Source hydrate (Reference)
Rainbow trout 59.0 28.8 11.8 --- 1.6 2
Rainbow trout 59.0 29.4 10.7 --- 1.7 6
Rainbow trout 66.2 20.2 7.4 0.2 1.3 7
Carp 66 28 2.5 --- 1.4 8
Chum salmon 54.5 30.1 10.4 --- 1.7 8
Cod 72.1 23.0 1.3 --- 2.1 8
Herring 69.2 26.3 4.2 --- 1)4 8
Sardine 70.7 21.0 7.0 0.3 2.1 9
Domestic hen 48.7 16.6 32.3 1.0 1.1 10
Ando investigated the fatty acid composition
of the fats in rainbow trout eggs2,12 , and found a high
proportion of unsaturated fatty acids, which could reach as
much as 82%. Of this 36% was higher fatty acids and 46% lower
fatty acids. 19.9% of the fatty acid constituents of the
acetone-soluble fats in chum salmon eggs13 were saturated, the
principal component being 016:0 , whereas 80.1% were unsaturated.
It is also reported14 that carp eggs were 28.3% saturated and
71.7% unsaturated. The unsaturated acids in rainbow trout
and chum salmon eggs contained more than 10% of highly
(%).

•
155
unsaturated C 22 acids but carp eggs only contained 1.5%14
It has been suggested 2 that the large amount of highly
unsaturated fatty acids contained in salmonid eggs is related
to their development at low temperatures around 10 0 at which
these fats are in a liquid condition.
The general analysis of the fats has been given above,
but in the living egg most of the fats are present either as
lipoproteins in the yolk or as fat droplets. This lipoprotein P78
is called lipovitellin, and reported fat contents are from
2 16. 22% to 25 16% in rainbow trout eggs and 9% in chum salmon
eggs. Half of the fat is phospholipids, mostly lecithin, and
in addition there are neutral fats, free fatty acids, and
cholesterol. The yolk components containing lipovitellin in
the young oocyte may be formed in tlie multithecal bodies
derived from the mitochondria, but in large oocytes they are
supplied from outside the egg by pinocytosis.
Fats are also present in the egg as fatty droplets.
These are entirely composed of neutral fats, and in rainbow
trout eggs 50% of the fatty acids were C18s 1 , C22:6 and
C16016 . They appear at first as small droplets surrounding
the nucleus, but often, depending on the species, they fuse
together into one or several drops.
•

• 156
2. Changes in body fat content during maturation of the ovary.
It is already known that the quantity of fat in the
body of many fishes shows seasonal changes. For example
Lovern19 reports that the fat in the body, though not in the
viscera, of rainbow trout increases from April to October, and
similar seasonal changes are known to Occur in mackere1 20 ,
sockeye salmon21 , and the Japanese sardine22 . It is often
suggested that these seasonal changes' of fat content are
related to water temperature and diet, but a response to the
reproductive cycle has also been suggested. Wada 22 found that
both the male and female gonads of the Japanese sardine enter
the phase of rapid development froM the end of January into
February, and that the fatty tissues decrease and the
proportion of fat in the muscle and the liver also become
lower (Table 8.2). .Suyama 6 found that the amounts of
phospholipids and aminolipids in the ovary of the rainbow
trout increase during maturation, but that at the same time
the amounts in the muscles decrease (Table 8.3). Thus the
changes of the fat content of the body are evidently connected
to the reproductive cycle, and it may be supposed that
oogenesis is accompanied by a "migration" of fat to the ovary
from all parts of the body.
P79

Table 8.2.
Seasonal variations of the fats in the body .organs
of female sardine ( 'o) .
Date caught Liver Fatty tissue Muscle
157
16 Mar 4.84
9 April 8.89
9 May 21.95 71.49
9 Dec 16.87 74.64
10 Jan 11.78 79.14
8 Feb 3.57 73.52
13 Mar 1.35
4.60
4.90
14.44
11.25
12.04
11.78
1.11
Table 8.3.
Changes of fat in the ovary of rainbow trout
during maturation (%) 22 .
Month Gonad Mean egg Extractable Phosphorus Nitrogen when index weight by ether. content of content of caught (%) mg. * (%) ** fat. ' fat.
(%) *** (%) ***
8 1.47 1.3 10.1 0.131 0.716 0.93 0.697 0.91
9 2.45 2.6 10.1 0.263 0.878 2.28 0.659 1.71
10 5.69 . 6.5 8.8 0.572. 1.220 7.39 0.747 4.86
11 9.06 8.2 7.1 0.582 1.300 10.66 0.794 6.51
12 17.2 16.6 6.4 1.04 1.580 26.3 0.930 15.44
* Assuming the average number of eggs per fish to be 2500. ** Converted to mg per egg. *** Converted to /ug per egg.

•
158
The quantity, the pathways and the conditions of this
fat migration contain many points of physiological interest,
23 but they are little known. Takashima et al made quantitative
measurements on 2-year old female trout obtained in December.
When they had been starved for the same period (85 days) the
amount of fat surrounding the digestive tract in immature fish
was 28.8%, but in mature fish it was 2.3%, and there was little
effect on the development of the ovaries. This showed that
the amount of depot fat diverted to the ovaries during
oogenesis is greater than the amount consumed for energy in
movement. Body fat in the female sockeye salmon also decreases
during migration from the river mouth to the spawning place,
but the 8% which disappears was found in an approximate
calculation to have migrated to the ovary24
Idler and Tsuyuki25 found that there was little
development of the ovary of the female sockeye salmon during
the spawning migration before the river mouth was reached, but
that there was an increase in the weight of the liver.
Nomura26 found that the beginning of the spawning season is
the period of greatest hypertrophy of the ovary, and that at
that time the weight of the liver is also high. The same
phenomena were found in land-locked sockeye salmon23 .
Takashima et al23 found hypertrophic livers of high fat
content in 2-year old rainbow trout caught in December.
During the spawning season the amount of fat in the liver of

•
•
159 ••
the Japanese sardine decreased22 , but in the directly
preceding period the liver was hypertrophied and had a high
fat content. Thus hypertrophy of the liver and an increase
of its fat content are found during the period of active
oogenesis. This shows that the liver is an active part of
the pathway through which fats migrate during oogenesis.
In order to investigate the way in which the depot
fat migrates, Takashima et al 28 measured by means of thin
layer chromatography the fat constituents around the digestive
tract, in the liver, and in the blood plasma of rainbow trout
in various stages of maturation. It was found that as
maturation proceeded the neutral fats and the free fatty acids
in the body fat around the digestive tract decreased. In
contrast, they were found to increase in the liver and in the
blood plasma (Figure 8.1). It is known that in birds and
amphibians 29-31 , the depot fats are transported in the blood
fluids as neutral fats and as free- fatty acids, and are
synthesized to lipovitellin on reaching the liver.
3. Endocrinological control of changes in fat metabolism
during maturation of the ovary.
Maturation of the ovary is principally controlled by
the diencephalon and the pituitary, and their internal
secretions are believed to take part in the changes of
metabolism during maturation. One of the ways in which this

16 0 ••
Figure 8.1.
Differences due to the state of ovary maturation of the
composition of the fats in the blood plasma, around the digestive
tract, and in the liver of two-year old female rainbow trout. In
mature fish there is little depot fat around the digestive tract,
but much in the blood plasma and the liver. There is very little
fat or free fatty acid in the depot fat, but much in the liver
and the blood plasma.
Cholesterol esters Neutra.1 fats Frp.e fa-tty acids: Free p.holesterol Non7-Dol ar fats

161
fat metabolism may be affected is through the action of the
sex hormones. It has been shown that the gonads of teleosts
secrete sex hormones under the influence of gonadotropic
hormones from the pituitary. Shreck32 found a concentration
of about 1.4 to 3.8 mg/mi of estrogen in the blood plasma of
rainbow trout, and Cedar et al 33 found that the blood plasma
of rire Atlantic salmon contained a high level of estrogen
which was reduced after the eggs had been discharged.
Eleftheriou et al 34 found maturation of the ovary of the
catfish to be accompanied by an increase of free estrogen in
the blood. The increase of estrogen during maturation is
thought to influence mctny other metabolic processes, such as
the metabolism of proteins, nucleic acid, and calcium, but fat
metabolism is also affected. Ho and Vanstone 3. 5 injected
estradiol monobenzoate into sockeye salmon, and reported an
increase of the total fats, the neutral fats and the total
cholesterol in the blood serum. Takashima et al 28 made four
injections, on alternate days, of 500t,g per fish of the
synthetic female hormone diethylstilbestrol into the abdominal
cavity of rainbow trout, and found an increase of the fats,
particularly the neutral fats and the free fatty acids, in
the blood plasma (Table 8., Figure 8.2). At the same time
there was an increase of lipoproteins in the blood plasma,
and hypertrophy accompanied by an increase of fat content of
the liver. These phenomena closely resemble the pattern of
p81
p82

• .
DES MT CS ACTE TP
• 162 Figure 8.2.
The influence of diethylstilbestrol (DES), methyltestosterone
adreno-cortico-tropic hormone (ACTH) and thyroid extract (TP)
on the fat in the blood plasma of two-year old female rainbow trout.
The blood plasma fats were increased by DES and reduced by TP. DES
also showed a tendency to increase the neutral fats and the free
fatty acids. CA and CS are controls.
Cholesterol . esters Neutral fats Free fatty aciO.s . Free ,cholesterol.; Mono - and di -glycer5.des Non-polar fats
Table 8.4.
The influence of diethylstilbesterol on the fat in the
plasma and the liver of the rainbow trout28 .
Amount injected (g) 0 5 500
Body weiuht (e) hepatosomatic index (%) Plasma fat (e/100m1) Liver fat (e0)
353 322 330
1.2 1.4 1.8
2.8 2.5 4.0
3.7 4.6 4.9 •

• 163
•
•
changes of fats already described in connection with
maturation. Thus the sex hormones take part in the
displacement of fats into the blood from the places in which
they accumulate. The sex hormones take part in the same way
as in birds36 and amphibians 37 in the displacement into the
blood of the accumulated fats, and it is to be supposed that
they are synthesized to lipoproteins in the liver. It has
been shown that thyroid hormones and adrenocortical hormones
take part in fat metabolism30 , and it is possible that they
may in this manner take part in the process of maturacion.
4. Fats in the diet of the parent fish.
As has already been discussed, the fats deposited in
the egg are derived from fats deposited in various parts of
the body, and these fat deposits are principally derived from
fats which have been taken by mouth. Therefore, if oogenesis
is to proceed satisfactorily, the parent fish must be supplied
with, and must accumulate, fats which are both qualitatively
and quantitatively sufficient. Ishida et al 38 explain the
so-called "shiniko" (literally "dead offspring") which is
found in herring roe as being composed of unripe eggs for
which the parent fish had insufficient nutrition before the
spawning season. In this example it is not only fats but
other nutritional elements whose insufficiently obstructed
the development of the reproductive elements. In salmonids,

• 164
the food intake is reduced during.00genesis, but it is
probably important that the conditions in which the parent
fish are reared before this period should provide for the
accumulation of sufficient fats in the body of the mother.
The eggs of rainbow trout contain many highly unsaturated
fatty acids, and it is supposed that they can be selectively
incorporated during oogenesis2 , but the diet given to parent
fish should be monitored with this in mind.
Lovern39 ' 4° has already shown that the fatty acid
components in the fats in the diet of fish have a considerable
influence on the fats in the body. Lasker et al40 and Ando 2
have demonstrated that the fats in the diet influence not only
the body fat but also the fat in the eggs. This is therefore
to be remembered when deciding the diet for the parent fish.
Furthermore, deficiency of essential fatty acids41 or of
choline42 leads to fatty degeneration of the liver, and the
administration of oxidized fats causes hepatoma43 , so that it
is known that nutritional deficiency can lead to abnormal fat
metabolism. However the investigations on this have all been
made with young fish, and there has been no study of parent
fish. It is therefore important that there should be further
studies of this matter in order to ensure satisfactory
development of the gonads.
•

•
165 ••
The eggs of salmonids exhibit a yellow-orange coloured
zone which is principally due to the presence of a carotenoid 4 pigment. Glover et al 1 observed that astaxanthin migrates
from the egg to the body of the embryo during the development •
of the salmon, and Deufel44 has.reported that an admixture of
canthaxanthin into the diet of rainbow trout improved the
quantity of eggs collected from the group and also the
proportion fertilized. This shows that the carotenoid
pigments take part in the formation of healthy embryos, and
suggests the possibility of using them as indicators of "egg
quality". There are other suggestions of the connection
between "egg quality" and pigments, but egg collection,
fertilization, hatching and fecundity ail vary according to
differences in the management and control of rearing. Very
careful and skilled investigation Of the value of pigments
as quality indicators is required. -
Grzenda et al45 reared goldfish on a diet containing
6.1 nmoles of 140 dieldrin, and found that after 32 days
47 picomoles/g had accumulated in the ovary, but that after
128 days there were 7209 picomoles/g, an increase of 150 times .
During this period increasing amounts accumulated in other
internal organs, but the rate of increase in the fatty tissue
and in the ovary was extremely high. Mayer and Sanders46
* Sic, but Grzenda gives 1836 picomoles in the ovary, and an increase from 649 to 7209 in the testes.
Translator. •

Guppy.
Fry per female 33 29
166
exposed Zebra danios (Brachydanio rerio) and guppies
(Poecilia reticulatus) for 90 days to diets containing 50
or 10g per gram of food, and found that the number of fry
decreased as the concentration increased, and that the
survival rate of the fry was reduced (Table 8.5). This may
be taken as a warning that water soluble or food soluble
substances may be concentrated in the ovary and hinder oogenesis.
Table 8.1.
The influence of di-2 -ethylhexyl Phthalat9 on
the reproduction of zebra danios and guppies.
Species. Dietary concentration Wg) 0 50 100
Zebra danio. Number of spawns 6 8 14 Eggs per spawn 20.3 15.2 10.1
'Percent of fry survival 51.1 31.7 11.5
•
•

16 7 • References.
•
1) M.Sureace and C. dorrio: Changes in
chemical composition during deve-
lopment of rainbow trout eggs.
'B. JAI.. Soc. SCI. FIsK. 23 2' 785 - 788 (1958)
: SEE P. 170
3) K.Y.sxso.tur: Phosphorus metabolism
in fish egg-I. Changes in the contents
of some phosphorus compounds
during early development of Oryzias
laliPes. Sci. Pap. Coll. Gen. Educ.,
Univ. Tokyo, 10, 99-408 (1960).
"4) M.GLovEs, R.A. MouTorr, and D.G..
. Rotors Astaxanthin, cholesterol and
lipins in developing salrnon egg.
Biochem. J., 50, 425-429 (1952).
t • 5) SEE P. $70
6) SEE P. 17$
7) S. Smut In "The physiology of
fishes" (ed. by M.E. Buowri), Vol.',
323-459.â, Academic Press, 1■'. Y.,
(1957).
8)
SEE P. 170
9) R. LAUER Efficiency and rate of
yolk utilization by developing em-
bryos and larvae of the Pacific
sardine, Sardinops caerulea. J. Fish-
Res. Bd. Canada, 19, 867-875(1962).
10)
SEE P. 171 11)
SEE P. 172
•

• 168
12) SEE P. 172
13) SE E P. 172
14) SEE P. 173
15) G.Youtio and L I. PITINNEY : On the
fractionation of the protein of egg
yolk. J. Biol. Chem., 193, 73-80 (19
51).
« 16) D. W. JARED and R. A .WAttAca :
' Comparative chromat,..r .aphy of the
yolk protein of teleosts. Comp.
Biochem. Physiol., 24, 437—.443 (19
68).
N. N. GUPTA and K. YAMAMOTO : Ele-
ctron microscope study on the fine
structural changes in the oocytes of
goldfish, Carassins auratus, during
yolk formation stage. Bull. Fac.
Fish. Hokkaido Univ., 22, 187---205
(1971).
18) $EE P. 173
17)
B. JAP. Soc. Sct. FISH
19 525-529 (1953)
21) D. R. IDLER and I. BITNERS : BiOCheITI•
ical studies on sockeye salmon • during spawning migration-II.
Cholestesol, fat, protein and water
in the flesh of standard fish. Can.
J. Biochem. Physiol., 36, 763-798
(1958).
SEE P. 173
23) F. TAEABHIXA, T. HIBIYA, T.
WATANABE, and T. HARA : Endocri-
nological studies on lipid metabolism
in rainbow trout-I. Differences in
lipid content of plasma, liver and
visceral adipose tissue between
sexually immature and mature
females. B. J. SOC Soi FISH _37 307-31 1 (1 9 7 1 )
24) D. R. InLan and L BITNE -RS: Biochem- i•
cal studies on sockeye salmon during
sp.awning migration-IX. Fat, protein
and water in the Major internal
organs and cholesterol in the liver
. and gonads of the standard fish. J.
Fish. Res. Bd. Canada. 17, 113-122
(1960).
25) D. R. IDLER and H. TSUYUKI : Bio-
chemical studies on sockeye salmon
during spawning migration-I. Physi-
- cal measurements, plasma chole-
- sterol, and electrolyte levels. Can. J. Biochem. Physiol., 36, 783-791
(1958).
Su P. 174
27) SEE P. 174
22)
19) 5. A. Lovaax Fat metabolism in
fishes-XII. Seasonal changes in the
composition of herring fat. Biochem. 26)
J.. 32, 676.-680 (1938).
20) E. NOGUCHI and M. BIT() : On the
seasonal variation of the liver weight
and oil content of the mackerel.

1 69
•
•
28) F. TAIIASIIINA, T. HIBITA, PI-tax-VAN Noss, and K. AIDA: Endocrinological
studies on lipid metabolism in
—rainbow trout-II. Effects of sex teroids, thyroid powder and adreno-
corticotropin on plasma lipid
•content. B. Jr. SOC. SC!. FISH. ' 38 43 - 49 (1 972 )
29) H. A. WALKER, M. W. Teroz,n, and
W. C. Rum.: The level and inter-
' relationship of laying hen. Pou!!.
• Sci., 30, 525530 (1951).
80) W.M. Mc Io: A lipophosphoprotein
• complex in hen plasma associated
• with yolk production. Biochem. J.,
72, 153-459 (1963).
31) P. J. HEALD and K. K. BOOKLEDGE :
Effect of gonadal hormones, gonado-
trophins and thyroxin on plasma free 38)
fatty acids in the dotnestic fowl.
J.Endocrinol., 30, 115--130(1964).
C. B. SCHRECK : Plasma oestrogen
leve,ls in rainbow trout. J. Fish.
Die!., 6, 227.--230 (1973). L. CEDARD, M. FONTAINE, and T.
Nouai: Sur la teneau en oestro-
genes du sang du saumon adulte
(Salmo solar L.), en eau douce. C. r.
•• •Seanc. Acd. Sc j., Paris, 252, 2656--
2657 (1961). 34) B. E. &minimal«, K. W. BOEIILICK,
and O. W. TIEMEIR : Free plasma
\ estrogens in the channel catfish.
• Proc. Soc. Exp. Biol. Med.. 121, 85
-48 (1966). 35) F.C. Ho and W.E. VANSTONE : Effects
of estradiol monobenzoate on some serum constituents of migratory
sockeye salmon (Oncorhynchus
nerka). J. Fish. Res. Bd. Canada,
32)
33)
39) J. A. Lovaaa : Fat metabolism in • fishes-XIII. Factors influencing the
composition of the depot fat of fishes. Biochem. J., 32, 1214-4224 (1938).
40) R. LASKER and G. H. THEILACKER :
The fatty acid composition of the lipids of some Pacific sardine tissues
in relation to ovarian maturation and diet. J. Lipid Res.,- 3, 60-44 (1962).
41) T. WsrANABE, F. TAKASIIIMA, and C.
Oottio : Effect of dietary methyl-
linolate on growth of rainbow trout.
B.JAP. Soc , SCI. FISH 40 181 - 188 (1974)
42) C. °onto, N. UKI, Z. IIDA, T. WATA-
NABE, and K. ANDO : 13 vitamin re-quirements of carp-IV. Requirement for cholin.
6. JAP. Soc. Sci. F :sn 36, 1140 -1146 (1970)
18, 859-.864 (1961). 36) D. A. Sciusins, M. WILKENS, R. G.
Mo LANDLESS, R. MUNN, M. PETERSON,
and E. CARSON : Liver synthesis, plasma transport and structural alterations accompanying passage of yolk protein. Am. Zoologist, 3, 167 ---184 (1963).
37) R. A. WALLACE and D. W. JARED t
Studies on amphibian yolk-V11. The estrogen-induced hepatic synthesis of a serum lipophosphoprotein and its selective uptake by the ovary and
transformation into yolk platelet protein in Xenopus laevis. Develop-
mental Biology, 19, 498-526 (1969).
SEE P. 175

43) D. G. ScAttraw : Ultrastructure and biochemical observations on trout bepatorna. U. S. DePt. Interior,
Bureau of Sport Fish. Wildlife, Res.'
70, 56-71 (1967). 44) J. Daum,: Pigmentierungsversuche
mit Canthaxanthin bel Regenbogen-forellen. Archiv. far Fish., 16. 125-- 132 (1965).
45) A. R. GRZENDA, W. J. TAYLOR, and
D. F. PARIS: The uptake and distri-bution of chlorinated residues by goldfish fed a "C•dieldrin contami-nated diet. Trans. Amer. Fish. Soc., No. 2, 215-223 (1971)•
46) F. L. MAYER JR. and H. O. SANDERS:
Toxicology of phthalic acid esters in aquatic organisms. Environm.
Health Perspectives, 4, 153-457 (19 -72).
170
2. ANDO Kazuo.
Yoshoku gyorui no shishitsu
gakuteki kenkyu.
J. Tokyo Univ. Fish.,
ni kansuru seika
ILI 61 - 98 (1968).
K. Ando.
Study of the fats in cultu'red fish.
Journal of the Tokyo University of Fisheries.
.5./1 61 - 98 (1968).
5. TAKAMA Kozo, ZAMA Koichi, IGARASHI Hisataka.
Sake tamago hassei katei chu ni okeru shishitsu
seibun no henka.
Hoku dai sui san iho 20 118 - 126 (1969).
K. Takama, K. Zama, H. Igarashi.
Changes in the composition of fats during the
development of the eggs
Fisheries Bulletin of the
20 118 - 126 (1969).
of the chum salmon.
University of Hokkaido.

17 1
6. SUYAMA Michizo.
Nijimasu tamago seijuku chu no ippan seibun no henka.
Nichi sui shi 24 656 - 659 (1958).
M. Suyama.
Changes of the normal composition of rainbow trout
fat during egg maturation.
Bulletin of the Japanese Society of Scientific
Fisheries 24 656 - 659 (1958).
8. TSUCHIYA Yasuhiko.
Suisan kagaku (kateibun). 243 peiji.
Koseisha Koseikaku, Tokyo 1965.
Y. Tsuchiya.
Fisheries Chemistry. (Revised edition) p. 243.
Published by Koseisba KoseikakU, Tokyo 1965.
10. FUJIMAKI Masanari.
Shin eiyogaku. 746 peiji.
Asakurà Shoten, Tokyo 1968.
M. Fujimaki.
New dietetics. p. 746.
Published by Asakura Shoten, Tokyo 1968.

•
17 2
11. ZAMA Koichi, KATADA Muneo, IGARASHI Hisataka.
Sake tamago no shishitsu - II. Fukugo shishitsu ni tsuite.
• Nichi sui shi 24 739 - 742 (1959).
K. Zama, M. Katada, H. Iuarashi.
Fats in the eggs of chum salmon II. On complex fats.
'Bulletin of the Japanese Society of Scientific Fisheries,
24 739 - 742 (1959).
12 , ANDO Kazuo.
Nijimasu tamago hassei chu no shishitsu no henka.
• Nichi sui shi 28 340 - 343 • (1962).
K. Ando.
Changes in the fat of rainbow trout during egg maturation.
Bulletin of the Society of Scientific Fisheries.
28 340 - 343 (1962).
13. ZAMA Koichi, KATADA Muneo, IGARASHI Hisataka.
Sake tamago no shishitsu - I. Yushi no seibun ni tsuite.
Nichi sui shi 24 569 - 572 (1958).
K. Zama, M. Katada, H. Igarashi.
Fats in the eggs of chum salmon - I.
On the components of the oils and fats.
Bulletin of the Japanese Society of Scientific
• Fisheries 24 569 - 572 (1958).

• 173 ••
14. IGARASHI Hisataka, ZA1U Koichi, KATADA Muneo.
Koi tamago no shishitsu - I. Yushi no seibun ni tsuite.
Nichi sui shi 26 326 - 329 (1960).
H. Igarashi, K. Zama, M. Katada.
Fats in the eggs of carp - I.
On the components of the oils and fats.
Bulletin of the Japanese Society of Scientific
Fisheries 26 326 - 329 (1960).
18. NAKAGAWA Heisuke, TSUCHIYA Yasuhiko.
Nijimasu tamago tanpaku no kenkyu - VI.
Hassei katei chu ni okeru shishitsu san sosei no henka.
Showa 44 nendo nihon suisan gakkai shunki daikai koen No. 466.
H. Nakagawa, Y. Tsuchiya.
Study of proteins in the eggs of rainbow trout - VI.
Changes in the fatty acid composition during development.
Paper No. 466, Spring meeting 1969 of the Japanese
Society of Scientific Fisheries.
22. WADA Shota.
Iwashi tai ni okeru shishitsu no seika gakuteki kenkyu.
No ka kai shi 22 339 - 475 (1953).
S. Wada.
Biochemical studies of body fats of the sardine
Sardincms melanosticta.
Journal of Agricultural Chemistry 22 339 - 475 (1953).

174 ••
26. NOMURA Minoru.
Nijimasu no jinko sairan ni kansuru kiso kDnkyu - V.
Seishokuso no hattatsu to shozangyo no okisa.
Nichi sui shi 22 976 - 984 (1963).
M. Nomura.
Basic studies of artificial egg collection from
rainbow trout - V. Development of the reproductive
organs and size of newborn fish.
Bulletin of the Japanese Society of Scientific Fisheries
z2 976 - 984 (1963).
27. IMURA Takatsugu, SAITO Tsuneyuki.
Gyorui soshiki kosei seibun no taisha kassei no
jiki teki henka I.
Nijimasu soshiki chu no kakusan ryo no henka.
Hoku dal suisan iho 20 202 - 210 (1969).
T. Imura, T. Saito.
Changes of the composition of fish tissues during
period of active metabolism - I.
Changes in the nucleic acid content of rainbow trout tissues.
Fisheries Bulletin, Hokkaido University,
20 202 - 210 (1969).

17 5
38. ISHIDA Rikikazu, SASAKI Takeo, ARITA Setsuko.
Nijimasu no ransoran ni kansuru so hiki gakuteki
kenkyu.
Iwayuru "shiniko" ni tsuite.
Hoku dai ken ken ho 24 171 - 176 (1962).
R. Ishida, T. Sasaki, S. Arita.
Histological studies of ovarian eggs in rainbow trout.
The so-called "dead egg".
Bulletin of the Hokkaidc, Regional Fisheries Research
Laboratory 24 171 - 176 (1962).
•

• 176
9, The accumulation of yolk protein.
KatsumLAIDA.
(Department of Agriculture, Tokyo University).
The ovaries of fish contain large numbers of oocytes, and
when yolk accumulates in the eggs as part of maturation, the
ovaries become greatly hypertrophie. Consequently the gonadosomal
index reaches 20% to 30% in many fish, and in the eel it may even
reach 60% to 75%. The composition of the mature ovary depends on
the species, but the values generally found are 55% to 75% water,
20% to 33% protein, 1% to 25% fat, and 0.7% to 2.2% ash1 . Thus
protein is the largest component other than water, and it
evidently has an important role in the process of morphogenesis
after fertilization. The protein can be divided into chorionic
protein and internal protein. In the mature egg the protein in
the yolk accounts for most of the internal protein of the egg,
and morphologically it is pricipally present as yolk globules and
yolk vesicles 2 . The protein of the fish egg is not a chemically
simple protein. Its main component is a lipoprotein corresponding
to the lipovitellin of the bird's egg, and it is found that there
are also phosphoproteins of the phosvitin type, and also livetin
proteins 3 8
•

110 1 77
There are three possible modes of formation and
accumulation of yolk proteins in the yolk.
Substances of low molecular weight such as amino
.acids and simple sugars are synthesized in the
oocyte itself.
2. Yolk proteins are synthesized in the follicle
cells and accumulated.
3. Substances synthesized in organs other than the
ovary are taken up and accumulated.
: Nakano et al have reported the occurrence of the
11, first Mechanism, the formation of protein in the Oocyte of
medaka Oryzias latipes during maturation 9 . However they did
not determine the type of protein which was produced. Almost
nothing is known as yet about the possibility of the second p89
type of production, the synthesis of protein in the follicle
cells. However, in the course of investigations of the
pl-ysiological changes in the body of the fish which accompany
maturation, Aida et al found results which lead to the third
possible process of yolk accumulation. These'results are
discussed below.
1.
•

• 178
•
1. The appearance and endocrine control of female
specific proteins during maturation.
It is already known that there are sexual differences
in the composition of the proteins in the blood plasma during
maturation of the gonads10-12 . Aida et al, used Ayu
(Plecoglossus altivelis) as the principal subject in a study
of the closeness of the relationship of these differences to
maturation of the ovary, and in particular, to the
accumalation of yolk.
Ayu from Lake . Biwa were reared under the natural
photoperiod. Yolk began to accumulate in early August and
the ovary began to hypertrophy rapidly. Ey late September
the gonadosomal index became 20% and spawning occurred from
late September to November. During spawning the gonadosomal
index was about 30%. The changes in the plasma proteins
during the process of maturation of the gonads were
investigated by means of starch gel electrophoresis, and
fractions 9 and 11 were found to increase rapidly after
vitellogenesis began in early August. After reaching a
maximum in early September they gradually decreased until
the time of death" (Figure 9.1, A). However no such evident
chancres were found in the males (Figure 9.1, B).
Next in order to investigate in detail the relation
between increases of these fractions in the females and the
state of maturation of the eggs, ayu which had been reared in

F. 9 F.11 0
•••- y • '
A , 'ez.e - • .%(
• • .1
7 .
179
1,
.. .
c
••••.; 071.,••• •
D ' , :.• e
•
• Figure 9.1. •
Starch gel electrophoresis (A - E) and agal" immunoelectrophoresis.
(F - J) of ayu plasma protein..
A, G. : Maturing female ayu.
B, F. : Maturing male ayu.
C, H. : Immature male ayu treated with 2 /mg of estradiol 17p .
D, I. : Immature male ayu treated with 20,ktg of estradiol 17C .
E, J. : Immature male ayu treated with 200/m.e of estradiol 17p ,
a : Maturing female ayu plasma and antibody.
Antibody of "a" absorbed by maturing male ayu plasma.
Origin,

180 ••
a suppressive photoperiod for maturation (16 hours of light
and eight hours of darkness, 16L-8D) were transferred to an
accelerative condition (8L-16D) and plasma samples were taken
on alternate days. The ninth fraction was observed to increase
on the 11th day, and at the same time egas appeared in the
ovary in the first stage of vitellogenesis with small
peripheral yolk droplets. The yolk droplets afterwards
increased with time. This result showed that the increase of
the fraction coincided in time with the appearance of yolk
globules around the eggs 13 .
It was next conformed by agar immunoelectrophoresis
that two types of protein (Fm-a, Fm-b) newly aueared in the
plasma of females which had begun to mature after early
August (Figure 9.1, F, G). The new protein Fm-a, present in
large amounts, was composed of the two fractions 9 and 11
found by starch gel electrophoresis. Aida et al named both
Fm-a (fractions 9 and 11) and Fm-b the "Female specific plasma
proteins" (FSPP). Our laboratory has since shown that the
same proteins appear in mature carp (Cyprinus carpioj, in
goldfish (Carassius suratus), in willow shiner (Gnathopogon
caerulescens), in makogarei (Limanda yokohamae), and in
konoshiro (Clupanodon punctatus).
The next question deals with the factors which
participate in the synthesis of FSPP. Urist and Schjeide14
reported an increase of a distinctive component of the blood

• 181
•
serum following injection of 10mg of estrone into the muscle
of the bass Paralabrax clathratus. Aida et al made
intraperitoneal injections of estradio1-17IS (0.2, 2, 20, 200?-g),
testeronè (10 /«g, lmg), hydrocortisone acetate (100tAg, 10mg)
and progesterone (10 .pg, lmg) into ayu in which maturation had
been suppressed. FSSP was found to develop only in the groups
in which 2g or more of estradiol 17/Shad been injected, and
the amount was shown to depend on the quantity of estrogen used 13 .
Figure 9.1, C, C - E, H - J). Estrogen also caused FSSP to
appear in immature female as well as matured ayu, and in
goldfish from which the seminal glands had been excised.
This suggests that estrogen stimulates the synthesis of FSPP
in some other organ than the gonads, and that it is discharged
from this organ into the blood.
2. The connection between FSPP and yolk protein.
Since it has been shown that the appearance of FSPP is
closely related to vitellogenesis, ovary extract from matured
ayu was used in antibody reaction with mature female blood
plasma. The precipitin bands, (Eg-a, b, c), were formed13
(Figure 9.2, A). Among these, Eg-a was of the same antigen
type as Fm-a (Figure 9.2, B, C) and, like Fm-a, was
fractionated by starch gel electrophoresis into the two
components found in the ovary extract. Fm-a and Eg-a combine
•

• 182
Figure 9.2
The relation between FSSP and yolk protein.
A, B Ovary extract from ayu. C. : Maturing female ayu plasma. D, E, F : Goldfish ovary extract.
a. Antibody and maturing female ayu plasma. 1 Antibody of "a" absorbed by maturing male ayu plasma.
Antibody and maturing female goldfish plasma.
d : Antibody and maturing male goldfish plasma.
Antibody of "a" absorbed by maturing male
goldfish plasma.
•

• 183
•
with fats, polysaccharides and calcium and the compounds were
found to have the same surface behaviour. Eg-b and Eg-c were
shown to be plasma proteins which are normally present in ayu
eggs. Yolk protein antigens to FSPP are also present in large
amounts in ovary extract from goldfish, and other serum
proteins are also normally present (Figure 9.2, D, E, F).
Krauel and Ridgway 15 showed by means of immunoelectrophoresis
the presence of a substance of an antigen type in the female
blood serum of red salmon in which the ovary weighed more than
10g, and reported the presence of a large quantity of the same
substance in the eggs. From these results it appears highly
probable that FSPP is synthesized under the stimulation of
estrogen, and is taken up by the egg from the blood in a
macromolecular condition as a precursor of the yolk protein.
3. The site of synthesis of FSPP.
The next question is the site in the body at which
FSPP is synthesized. Since it is known that this synthesis
occurs in the liver of other oviparous vertebrates, it is
supposed that FSPP will be synthesized in the liver of fish.
It is in fact known that the liver of the female is enlarged
during maturation, relative to that of the male, in the deep
water herring, (gisu) Pterothrissus gissu, the frog flounder
(meitagarei) Pleuroichthys cornutus, the half-beak (sayori)
•

• 184
•
Hemiramphus sajori 16 , the Pacific mackerel (saba) Scomber
japonicus 17 , the loach (dojo) Misgurnus anguillicaudatus 18 ,
the medaka Oryzias latipes 19 , and the three-spined stickleback
(togeuo) Gasterosteus aculeatus 20 . Aida et al made detailed
observations in ayu and in makogarei Limanda yokohamae. There
were sexual differences in the hepatosomatic index of ayu at
the time when the gonads begin to ripen, but after maturation
that of the female was 2 to 3 times that of the male.
Histological examination of the changes of the liver showed
that as tbe ovary commenced to mature the female liver cells,
the nuclei, and the nucleoli began to swell, the cytoplasm
became strongly basoellic because of an increase of RNA, and
fats and glycogens tended to diminish. These phenomena
continued until spawning, and sugget that the cells were
actively synthesizing protein21 . Maturation of males was
accompanied by atrViyof the liver and a lowering of its
activity. The hepatosomatic index of mature female Limanda
yokohamaemas three or more times that of the male, and the
cytoplasm was extremely basophilic. The water content was
high in females, but the fat content was found to be high
in males.
The cytological changes in the liver cells of female
which may accompany maturation are known to be reproduced
when estrogen is administered to males or to ovariotomized
females 18-20 . The administration of estradiol 173to immature

• 1 85
•
or mature male ayu induces changes similar to those in
maturing female ayu. The electron microscope shows that the
use of estrogen results in a considerable growth of the rough
surfaced, endoplasmic reticulum (rER) and of the Golgi
apparatus, and it is supposed that estrogen enhances the
ability of the male liver to synthesize protein.
The changes of the nucleic acid content of the liver
were investigated in order to corroborate the histological
results (Figure 9.3, A). The RNA density of the female liver
began to increase in the early part of August, reached a
maximum at the end of August and maintained until the spawning
season a value 1.5 to 2.5 times that normal to the male liver.
However there was practically no sexual difference in the DNA
density or in the protein content. The total DNA and the
total RNA gradually declined in the male, but in the female
they began to increase in early August and reached mardmum
values in mid September, when the RNA was about six times that
of the male, and the DNA about 3 times. They then,gradually
declined, but remained much greater than the normal male
amounts 23 . As shown in Figure 9.3, B, the relation between the
increase in weight of the ovary (at 10 day intervals) and the
integrated sexual difference in the RNA content of the liver
is linear from early August to mid September. After this time,
the relation breaks down, so it appears that some other
mechanism takes part in the increase of weight of the ovary as

-7-70 7/22 8/i
(Ix 100) •
A
Reln.tive quantity of nucleic acid inm the liver.
300
200
100
, FOU(derisity) _
• DNA(density)
11 30 31 21 10/10 9/10 • 20
• 186
Figure 9.3, A.
Changes in the nucleic acid content of the liver,
accompanying sexual maturation.
(The ratio of the weight of the female liver to
that of the male, which is taken as 100%).
•

• Increase in weight of the ovary.
(g).
• Figure 9.3, B.
The relation between the increase in weight of the ayu
ovary (in 10 days) and the total sexual difference in
the RNA content àf the liver..
1 87
1. 22 July to 1 Aug.
2. 1 Aug to 11 Aug.
3. 11 Aug to 21 Aug.
4 • 21 Aug to 31 Aug.
5. 31 Aug to 10 Sep.
6. 10 Sep to 20 Sep.
7. 20 Sep to 30 Sep.
8. 30 Sep to 10 Oct.
Total sexual difference in the
RNA content of the liver.

188 •
•
•
it becomes ripe. Creelman and Tomlinson have found that
female sockeye salmon at the time of going upstream have
larer amounts and densities of RNA-P in the liver than have
the males 23 , and this is in complete agreement with the
results in ayu.
Four types of steroid hormones were used for these
measurements, but, as in the investigation of FSPP, the
changes of the nucleic acid in ayu occurred only in those to
which 2p.g or more of estradiol had been administered, and
the amount of change depended on the quantity of estr-ugen
used (Figure 9.3, C).
Furthermore, when mature female ayu are reared for
one month in conditions which suppress maturation (161, - 8D),
not only does the development of the ovary cease, •but FSPP
disappears, the liver atrophies, its RNA density, RNA quantity,
and DNA quantity greatly decline, and the gonadotrophic
PAS-positive substances diminish.
Ovariotomized immature females were reared for one
month in conditions which accelerate maturation. A ritrong
development of gonadotroph was observed in both the ovariotomized
and the sham operated groups, but FSPP was not observed in the p94
ovariotomized group. The hepatosomatic index, the RNA density,
the quantity of RNA and the quantity of DNA increased very
greatly in the sham-operated group, but no changes were
observed in the ovariotomized group.
k,nm17n,17.171,,vem.5?!mrym M,".137VF's,.

RNA (toU')
400
Cie)
Relative quantity of nucleic acid in the ! liver.
(Treated group 1 X 00 ) (Control group )i
100
HSI
(total) (total)
RNA (density)
DNA (density)
0.2 2 2'0 200n EsuacIMA7/1
• 189
•
Figure 9.3, C.
The effect of estradiol 17C on the quantity of
nucleic acid in the liver.
(The quantity in the treated group relative to that
in the control group which is taken as 100%).
•

• 190
•
In order to observe the action of estrogen in detail,
300 it,1/4g of estradiol 17(3 was administered to immature ayu, and
the times needed for the appearance of FSPP and of chances in
the composition of the nver were measured. FSPP was found
in the plasma after 48 hours, and the increase of FSPP was
accompanied by an increase of the total blood protein to 2.7
times that in the controls. The hepatosomatic index also
began to increase after 48 hours, and reached a value twice
that of the controls. The water content of the liver also
suddenly increased by about 5% after. 24 hours, but after
120 hours it showed no. difference from the controls. The
density of RNA waJ found to increase after 48 hnurs, and remained
high for 240 hours, being 1.6 times that of the controls. The
DNA density showed a tendency to decline after 72 hours, and
this is thought to be connected witli the decline in water P 9
content. The total quantity of RNA increased after 48 hours,
and after 240 hours was 4 times that of the controls. The
increase of total DNA was at least 24 hours later than the
increase of RNA, showing that RNA synthesis precedes DNA
synthesis. All these results are now considered to conform
with the ideas .that FSPP is synthesized in the liver because
of the increase of the quantity of estrogen which accompanies
maturation, and that it is discharged into the blood. Furthermore,
this estrogen produces development through its action on the
metabolism of the water content, the nucleic acids, the sugars,
the fats, and the proteins.

• 191
•
4. The introduction of FSPP into the maturing egre.
The facts so far described suggest that FSPP is taken
up by the egg as a macromolecular substance, and the following
experiments were made in order to test this hypothesis.
Firstly, male gold fish in which FSPP synthesis had
been stimulated by the use of estradiol 17(3 were treated with
100 .eCi of L-leucine 4.5 - H3 , and plasma from blood taken
48 hours later was fractionated in a Sephadex G-200 column.
An extremely high radioactivity was found in the FSPP fraction,
which confirmed the active s-,:mthesis of new protein25 .
This newly synthesized FSPP was administered to
maturing goldfish and its process of migration was investigated.
As shown in Figure 9.4, A, the protein fraction in the plasma
reached a maximum after 10 hours and then rapidly declined in
the females, whereas a relatively high level of radioactivity
was maintained in the males25 . The radioactivity in the ovary
protein fraction gradually increased, as shown in Figure 9.4, B,
and after 120 hours it was about 10 times that in the blood
plasma fraction. However, the radioactivity of the seminal
glands was only one third that of the blood plasma. Moreover,
more than 50% of the quantity administered accumulated in the
ovary, whereas scarcely 2% was present in the seminal glands.
Male and female plasma and ovary extract were fractionated in
Sephadex G-200 96 hours after treatment, and the site of the
radioactivity was measured. In all three, the peak was found

192
Figure 9.4.
410 24 48 .. .72 96 120 Houftl:
1420 24 - 48 • 72 06 Hourts
__ • •
à 0.6 ro csJ
—OA
D 0.2
4 n
2
10 20 30 40 60
Ffte.C1104._
60 70 80
•
• A, B. Changes with time after the administration of
labelled FSPP of the plasma protein (A) and of
the gonad protein (B).
o Male, o Female, Dpm, Disintegrations per minute.
C. The location, in Sephadex G - 200, of the radioactivity
in the ovary extract 96 hours after administration.
Cpm, Counts per minute.
•

• 193
to be in the same place as FSPP. In particular, no radio-
activity was found to have been transferred to the other
protein fractions or to any low molecular weizht fractions in
the ovary extract, and this strongly suggests that the
labelled FSPP was transported to the egg as a macromolecular
substance 25 . In addition, since micropinocytotic vesicles
have been observed during the process of maturation in some
eggs26,27
the egg by micropinocytosis. Aida et al, have shown by means
of the fluorescent antibody method that FSPP is present in
the yolk globules, and that the normal plasma protein
components are contained in the yolk vesic 1es 25, it is concluded from these results that FSPP is
synthesized by the liver when stimulated by estrogen, and
after being discharged into the blood it is taken into the egg
in the macromolecular condition and becomes a structural
component of the yolk globules. (Figure 9.5).
To sum up, there are three possible mechanisms for
the formation and accumulation of yolk protein, but he
author concludes from the results given here that the third
mechanism must now be considered to be the principal mechanism.
However since the other two mechanisms may operate at the
same time, further detailed studies will be required in order
that the full mechanism may be made plain.
it appears reasonable to suppose that FSPP enters

SE CO4DAi; , SYMPTOMS. ;
LIVER. 00,RY.
I ,POLLICLE
CELL LAYER IVOLK
vES ICLE
GLOBULE
• ••
Figure 9.5.
The mechanism of the formation and accumulation of
the yolk rrotein in.fish.
' MrPoTHiLAmus.
194,
PITuiTARY.
•GONADOTROPH IC HORMONE. •
CELLCaÀVAGE Oftt INUCLEATE CELLS ; • •
Estrogen
RNA
I 1 FRESH PROTE IN , k
. SYNTHESIS.
1
ROUGH-SURFACE AM I .NO . RET ICULUM.
DNA
PLASMA
', ROTE IN,
a
r. ACIDS.
4 Boor
TISSUE.
•

References.
•
•
1) SEE P. 197
2) SEE P. 197
3) E. C. YouNo and J. T. Pruxusr: On
the fraction of the proteins of egg
yolk. J. Biol. Chem., 193, 73-80 (19
: 51).
4) T.. E. BARMAN, NOUYEN-KIM BAI
and Nnurstz-Veli THOAI: Studies on
a herring-egg phosphoprotein. Blac-
kens. J., 90, 555-558(1964).
z,) K. ANDO : Ultracentrifugal analysis
of yolk proteins in rainbov trout egg
and their changes during develop-
• ment. Canadian J. Biochem., 43, 373
.--379(1965). •
Y. ITO, T. FUJI', M. NAKAMORI, T.
! HAT-rout and R. YOSHIOKA: Phosvitin
\ of the trout roe. J. Biochem., 60, 726
- —.428(1966).
7) Y: MANO and F. LIPMAN: Characte-
! ristics of phosphoproteins(phosvitin)
from variety of fish roes. J. Biol.
- ChWit., 241, 3822—.3833(1966).
8) D. W. JARED and R. A. WALLACE :
Comparative chromatography of the
yolk proteins of teleosts. ComP•
Biochem. Physiol., 24, 437-443(19
68).
9) E. NAKANO and M. ISHIDA-YAMAMOTO:
Uptake and incorporation of labeled
amino acid in fish oocytes. Acta.
Embryo'. Morphol. ExPeri., 10, 109
"416(1968).
10) W. E. VANSTONE and Cm:run-Wm
Ho: Plasma proteins of coho-salmon.
Oncorhynchus kisutch, as separated
•by zone electro phoresis J. Fish.
Res. Bd. Canada, 18, 393-399 (1961).
11) A. Datum and J. •U. Pius :
Dimorphisme sexual dans les pro
serique de Salmo salar: Etud-éte
electrophoretique. Compt. Rend.
Soc. Biol. , 157, 11, 1897—.1900(1963)
12) R. V. TnousroN: Electrophoretie
pattern of blood serum proteins
from rainbow trout (Salmo gaird
nerii). J. Fish. Res. Bd: Canada;
24, 10, 2169-2188(1967).
13) K. AIDA Pusx-VAN-Nomr and T1
• Phy.sioloïical studies on
gonadal maturation of fishes-I.
' Sexual difference in composition o
•

Bd.
• nucleic acid. J. Fish. Re:.
' Canada, 16, 421-426(1959).
24) SEE P. 199
SEE P. 197
17) SE ( P. 198
• 196
References. continued.
plasma protein of Ayu in relation
to gonadal maturation. Bull. Japan.
Soc. Sci Fish., 39, 1091-1106(1973).
14) M. R. URIST and A. O. SCHJEIDE :
The partition of calcium and protein
In the blood of oviparous vertebra-
tes during estrus. J. Gen. Physiol.
44, 743-456(1961).
15) K. K. KRAUEL and G. J. RIDGWAY :
Immunoectrophoretic studies of red
' salmon serum. Int. Arch. Allergy,
23, 246-253(1963). .
16)
Histological changes in the liver cells of Ayu following gonadal maturation
and *estrogen administration. Bull.
Japan. Soc. Sci. Fish., 39, 1107-- 1115(1973).
22) SU P. 198
23) V. M. CREELKAN and N. TOMLINSON
Biochemical ' studies on sockeye salmon during spawning migration
VI. Ribonucleic acid and deoxyribo-
18) H. KOBAYASHI Effects of eStrogen . upon the structure, weight and fat
• content.
of the liver in the fish,
• Misgurnus anguillicaudatus. Annot.
Zoo!. Japon., 26, 213-416(1953).
19) N. Diem: Effect of estrogen and
androgen on the weight and struc-
ture of the fish, Oryzias latipes.
. ibid., 28, 79-435(1955).
20) C. 00IIRO: Some observations on the • . effect of estrogen upon the liver oi the three spined stickleback, Gasterosteus aculeatus aculeatus
• L., ibid., 29, 19-23(1956). 21) K. AIDA, K. HIROSE, • M. YOROTE
and T. HIBITA: Physiological studies
' on gonadal maturation of .fishes-II:
25) SEE P. 199
26) K. Y,o/AuciTo and I. OOTA :.* An
electron microscope study of the
formation of the *yolk globule in the
• oocytes of zebra-fish, Brachydanio
rerio. Bull. Fat. Fish. Hokkaido
Gniv., 17, 16i-.:174(1967). 7) N. N. GUPTA and K. YAMAMOTO:
Electron microscope study on the fine structural changes in the oocytes
"of goldfish, Carassius auratus,
'during yolk iormation stage. ibid.,
22, 187-205(1971).

• 197
1. TSUCHIYA Yasuhiko.
Suisan kagaku p234 - 251.
Koseisha Koseikaku (1962).
Tsuchiya.
Fisheries chemistry. p234 - 251.
Published by Koseisha Koseikaku, (1962).
2. YAMAMOTO Kiichiri.
Gyorui sein i (sel Shoku). p233 - 271.
Koseisha Koseikaku (1970).
K. Yamamoto.
Fish physiology (reproduction). p233 271.
Published by Koseisha KoSeikaku, (1970).
16. AMEMiYA Ikusaku, TAMURA Tamotsu.
Gyorui kanzo juryo shiyusa ni tsuite.
Sui san gaku kaiho 10 10 - 13 (1948).
I. Amemiya, T. Tamura.
Sexual differences in the weights of the
iivers of fish.
Reports of the Society of Scientific Fisheries,
10, 10 - 13, (1948).

• 1 98
17. NOGUCHI Elzaburo, BITO Masamichi.
Saba kanzo no juryo oyobi shiboryu no
kisetsuteki henka.
'Mehl sui shi 12 525 - 529, (1953).
E. Noeuchi, M. Bito.
On the seasonal variations of the liver weieht
and oil content of the mackerel.
Bulletin of the Japanese Society of Scientific
Fisheries 12 525 - 529, (1953).
22. AIDA Katsumi, HIBIYA Takashi.
Gyorui no seijuku ni kansuru seirigaku teki kenkyu - II.
Shitoku isei kessho tanpaku to rano tanpaku
oyobi kanzo kino to kankei.
Showa 45 nendo nihon suisangakkai nenkai happyo.
K. Aida, T. Hibiya.
Studies of the physiology of maturation in fish - II.
The connections between female specific plasma
protein, the yolk protein, and the liver processes.
Collected papers of the 1970 annual meeting of the
Japanese Society of Scientific Fisheries.
•

1 99
24. AIDA Katsumi, HIBIYA Takashi.
Gyorui no seijuku ni kansuru seirigaku teki kenkyu IV.
Estrogen no kessho tanpaku sosei oyobi kanzo seibun
ni okeru eikyo.
Showa 46 nendo nihon suisan gakkai nenkai happyo.
K. Aido, T. Hibiya.
Studies of the physiology of maturation in fish - IV.
The influence of estrogen on the protein component
of the blood plasma and on the composition of
the liver.
Collected papers of the 1971 annual meeting of the
Japanese Society of Scientific Fisi:eries.
25. AIDA Katsumi, HIBIYA Takashi.
Gyorui no seijuku ni kansuru seirigaku teki kenkyu - V.
Shitoku isei kessho tanpaku no tamago e no torikomi.
Showa 48 nendo nihon suisan gakkai nenkai happyo.
K. Aida, T. Hibiva.
Studies of the physiology of maturation in fish - V.
The take-up of female specific plasma protein into
the egg.
Collected papers of the 1973 annual meeting of the
Japanese Society of Scientific Fisheries.
•

•
•
200
••
V. The evaluation of ep-,g quality in spawn.
10. Freshwater fish.
Kiyoshi SAKAI.
(Tokyo University of Fisheries).
The management of the hatching and rearing of fry, and
the steady artificial production of fingerlings would be
greatly facilitated if it were possible to develop methods by
which the quality of the eggs could be appraised immediately
after collection, and those eggs which were suitable for
artificial fertilization could be selected.
It is at present believed that the quality of the eggs
is closely related to the age of the parent fish, to their
nutrition, ancestry*, conditions of rearing and spawning
season, but studies of the relationships between these factors
and the proportions of the eggs which are fertilized or which
develop have not yet given concordant results1-4
The colour, transparency and elasticity of the eggs
of freshwater fish are used as criteria for the appraisal of
egg quality, but the relations between these criteria and egg
quality are by no means clear 7-9, 14
* For example, it is said that there are differences between marine, lake and artificially reared Ayu-(Plecoglossus altivelis).

201
Experimental work was therefore undertaken with the
objectives of appraising the satifactoriness of the quality
of the spawn, and of defining this quality in terms of
•
•
satisfactory fertilization and hatching of the eggs when
collected. The results of these experiments are discussed
in this paper.
The experiments were made in the following manner.
The fish used as parents were Rainbow trout (Salmo gairdneri),
Ayu (Plecoglossus altivelis), Sogyo (Ctenopharyncodon idellus)
and Hakuren(Hypophthalmichthys molitrix). These were reared
in field ponds at natural temperatures and in laboratory
aquatrons with the temperature controlled in three steps.
Small quantities of eggs were soueezed out at intervals from
fish which had ovulated naturally or . had been caused to ovulate
by hormone treatment, and these eggs were examined by eye and
by microscope. The external examination of the eggs included
the colour, the transparency and the elasticity, and also the
size and distribution of the fatty droplets. An internal
histological examination of the morphological characteristics
wàs also made. At the same time, measurements were made of
the proportion of the eggs collected which were fertilized
and which developed, and these measurements were compared
with the morphological observations. The pH of the eggs and
the refractive index of the yolk obtained by centrifuging
were also measured.
p101

202
1. The Quality of the spawn and its internal and external
morphological characteristics.
Eggs which,remain in the body after ovulation pass
successively through the four following morphological stages
In rainbow trout and ayu, eggs which have just been
ovulated are semitransparent and small fatty droplets are
uniformly distributed throughout the body of the eggs (Stage 1)
(Figure 10.1, -1, 11).
Next, some of the fatty droplets distributed throughout
the egg increase in size and begin to accumulate at a
particular location near the animal pole (Stage II),
(Figure 10.1, -2, 12). Soon after, all the fatty droplets
gather together, and in rainbow trout they form a single
cluster while the remainder of the egg becoMe's transparent or
cloudy white and translucent. In ayu, several large drops
are formed, and the remainder of the egg continues to be
translucent (Stage III), (Figure 10.1, -3, 13).
In rainbow trout, the egg next contracts and loses
its globular shape and its elasticity. In ayu, the fatty
droplets and the protoplasm are collected into one place, and
the rest becomes completely transparent (Stage IV), Figure 10.1
-13). Rainbow trout eggs in Stage III and ayu eggs in Stage IV
are generally called overripe eggs.
* An introduction is presented in this paper to the four stages which have been used as normal divisions of the process of maturation by Nomura et al. in rainbow trout15, 16, by Sakig e al. in Sogyo and Hakuren 1 7 and by Sakai et al. in Ayu1°,19,

• 203
•
From the histological point of view eggs in Stage II
are distinguished from those in Stage I by the increases in
size and the gathering together of the fatty droplets, and by
the loss . of some of the cortical alveoli from the cortical
cytoplasm, but the yolk and the fatty droplets have not yet
penetrated into the cortical cytoplasm (Figure 10.1 - 4, 5, 7,
8, 15, 16). At the time of ovulation, the yolk of the rainbow
trout appears to be homogeneous, but in the ayu it consists of
innumerable small droplets which coalesce and form large drops
during passage through this stage. In the Stage III rainbow
trout egg, the cortical protoplasm which has hitherto surrounded
the periphery of the egg atrophies and contracts, and the yolk
percolates in between the contracted cortical protoplasm and
the chorion. The fatty droplets and the yolk penetrate the
atrophied cortical protoplasm (Figure 10.1-6, 9). In the ayu
egg, the fatty droplets and the yolk globules become large and
penetrate the contracted cortical protoplasm and the chorion
(Figure 10.1 - 17). In Stage IV the content of the rainbow p106
trout egg becomes disordered (Figure 10.1 - 10). In the ayu
egg, the yolk percolates between the atrophied cortical
protoplasm and the chorion (Figure 10.1 - 18).
The eggs of sogyo and hakuren contain no fatty droplets
immediately after ovulation. They are elastic and translucent
(Stage I) and the yolk consists of innumerable small globules
(Figure 10.1 - 19). After ovulation the elasticity is

204
maintained as time passes and the transparency is increased
(Stage II). In this stage the yolk globules coalesce and
increase in size but do not penetrate the cortical protoplasm
(Figure 10.1 - 21). Next, the egg loses its elasticity
(Stage III), the coalesced yolk globules join together in
large clusters, and some of them penetrate the cortical
protoplasm (Figure 10.1 - 21). Next the protoplasm gathers
together in one place and the rest becomes completely
transparent. The egg swells up again and becomes cloudy
(Stage IV). The yolk percolates between the contracted
cortical protoplasm and the chorion in these eggs in the same
way as in overripe eggs of rainbow trout and ayu (Figure 10.1-22).
When the eggs are divided among stages in this way, it
is found that the quality of rainbow trout and ayu eggs is
related to the stage. About 70% to 80% of Stage I eggs develop,
40% to 50% of Stage II, and none at all of Stage III. In sogyo
and hakuren the proportion of live eggs (defined as the
percentage of the total number of eggs which are alive one day
after being exposed to fertilisation) in Stages I and II
shows great variability from 0% to 97 2 with an average of 36%,
but in and after Stage III it is zero.
This shows that the quality of the eggs decreases
gradually after ovulation.
•

urrit.•- ••••
• • ./..•7!--.- r.! • re ,
j*, Jt!.. • , 7 ,
, • f. • . ' • • .;: . . • •
.7%.4%4"4•7,4 "
• ,; • •
.,e.7"1"-le•-#4
ç-! A • 1;
• •
-.6 I •,.
. •
• • • ; • ,;•-.
JF • • r•••
, . • Veltre
le, • •
!-Ç!
,
T JQ •
! •
_
. . :
i.
1
'0

• 2 05
Figure 10.1.
Part 1. Rainbow trout. p102
1. Stage I. Small fatty droplets distributed through the body of the transparent egg (Live egg).
2. Stage II. Fatty droplets beginning to increase in size and coalesce (Live egg).
3. Stage III. Fatty droplets and protoplasm assemble in one location and the remainder of the egg becomes transparent (Fixed egg).
4. Stage I. There are 1 or 2 rows of small fatty droplets immediately inside the cortical protoplasm, and the yolk is forming apparently ho:aogeneous clusters in the centre of the egg. (Chorion removed).
5. .Stage II. rrhe fatty droplets are increasing in size and beginning to coalesce (Chorion removed).
6. Stage III. The surface protoplasm surrounding the egg has contracted and the yolk has percolated 'between the chorion and the cortical protoplasm.
The vicinity of the animal pole of the egg in Stage I. There is a small thickness of fat in the cortical protoplasm, and many
• cortical alveoli are visible.
8. The vicinity of the animal pole of the egg in Stage II. Some of the cortical alveoli have disappeared, but the fatty droplets and yolk globules have not yet penetrated.
9. The atrophied cortical protoplasm of the egg . in Stage III. The fatty droplets and yolk
globules are seen to be penetrating the contracted cortical protoplasm.
10. The profile becomes oval and the interior contents are disordered.
7.

Figure 10.1..
206
p103 Part 2. Ayu.
11. Stage I. The ege transparency is low, and small fatty droplets are distributed throughout the egg (Live egg).
12. Stage II. The ego: transparency and the size of the fatty droplets have slightly increased, and the fatty droplets are beginning to collect at the animal pole (Live egg).
13. Stage III. The egg transparency is increasing and several large fatty droplets can be seen (Live egg).
14. Stage IV. The protoplasm and the fatty droplets are gathering into one location and the remainder becomes completely transparent.
15. Stage I. The cortical protoplasm rs seen inside the chorion and the adhesive membrane. Small fatty droplets are distributed throughout the egg, and innumerable small yolk globules are present.
16. Stage II. The yolk globules and the'fatty droplets slightly increase in size, and the fatty droplets move toward the vegetative pcle. The yolk and the fatty droplets have not . penetrated the cortical protoplasm.
17. Stage III. The fatty droplets have greatly increased in size, and the cortical protoplasm, being penetrated by the enlarged yolk globules, becomes thin.
18. Stage IV. The cortical protoplasm having atrophied, the yolk percolates between the chorion and the cortical protoplasm.
•

f
. • , • :•":" • . z‘. '; - ' • • • ;
., • , . • , „.1 e . • - e•J • •• • . ' ir • • - ••
• - • . • -• . • e. ; -•
'-'!„- .;., . • . • ...• e. . , . :• • e.-•,- • ''.. :̂-•••••„, .,,, .
.-, • - ..,-. , • . .-..'2 .- ' • :..; 1.. j. 1 '4 ,•-f".e--.4 - \••• ' —
. " .'‘..:f''''L\s.. . .- . - * . .. • e... ". -;,', 1 •• : ' k 4 - - s '. e • .. f -: ',.‘ ' .. ... . .. : ' . . - • ' • • '. . 7 ..... • t:' • ". • •
- ' • ) te4•4,e. • , . ■.'. ,,•%,'‘... • ;•.« ; •••••4‹ te ... • ,•_., _.,._ ., .,
• ; r : . :,:si...„......:.. u2.̀...2*,:......
F---. ••-;3r1.- : ,...,•, • -.....i.,-.:•.---••,7 r ,„7,---.-„..,......„......,..
,..›,_,•,-. e••• .e.:•.,,,,,••••,(-..e,..„••• • .••....,.., •.:•:•. ,..•.... _•••• 7.2. ;..:,..„ ..• . __,,e,...,3,:•..,..____,..,. .•,.. ".:,„ 4 . • • ' `. •e•■■1 ' 7 '. . • • • • , • • ..i •••■ "« • ••• , ,:".• ;;;•-• •,•;. ■ • ,.. 1. 4-, '.•,• .. ", ' t " i'.: ' . • - • ' • -...k ' • • • .,
À1. . !•.:•r..... e7.• • *- . . 1..'. , ,,,;..:, ,. i...".".-,... .. L . ,, • 1;5 - - .....' )4',... • ." .., •,1:•;' , - • .. •••-
1 ,..-":' ..., ‘ ,..1,-“. • •-• - ..' "'. 's.•• ‘.3
i- i, '. ••••••' • ‘ ; • ' --::•• '•'''•••••••t.' -'.„.•:. •• :.._ ...,.,-..,7•.Y .7% . *i•••4,.*-•,•4- i•-,.:.-, ?,,-.. -9 1 ' ze •-z„. •:,;•••-.;,:e . - • .. ••• • -4. :-. 41.--.. - •.:- -,,,...,,..,••••••„:, .. ,,,.., e.....erl,..t- i,i,,;,,t
r. e. ,.....1,....,,, ,,:,......, :......:.....„.::. : .4 e . ; :-...... .. . • ,_,,,- 'ti____ 4 e.,4•,,. ..'..kie-r• •• •••
i - -e•!.. 4.-...s - ..1.„- -. • , • ..;.= , f-' .- %_. -......•:,. :J.. :, --.. ./.% :e•- 4 Vte
4_,. ;,....-. ••:., (,•,......., : : . „.•,....,..›,.:,:-...,,-.. , • 1,--,,, e,t,
.., .., re-44 • ' €- 5-- .--• '-:'• ...- -+ t• •!"•• •- ----s-e- r- • ": ,: -•-:•::',-.••'.. 7:`,4,4-`.: : : .,•:. ii."; ree;,*".)- -7.!-; -‘ %.: `•1:-‘.4-'-'1.. -J.: !' •-•.-....te,... ' : • - . . . 4 ",...' • "::. 7 . :::'''' ''.tfw. ':; ',.i. , k . . fa V ../7,,,, ? 5-1 • I
.1 ' . . • : •••-••;: ...:..:.«;;;•••`-''4q. • . •-• :-e,, "e%i'li_, &i.' ''''-./,'•.:.e-. \
.,:.;.:•••••›-,:-..- ,--- -•.,.. : - • - i.. • .-.. •• -:',.,.. ,•-x-r,..,.,• • .: -„:„._ ,. -,.;-;,,,,r , :. . ,... ei.e,,,,:.••?1, ..„.‹ ..4 Ci 4 *;•‘ '.. .;', - .? .<:. ' ,'...i•,;. ft•-• '. '4 '.'15 :•'' " •••;' ...7.' ■ • • ...;•• • II. • S''''-` 1 ....:4i,
'1 ' c•• •• 7! .-:,=''''‘.*,.. jitrii, *.•• .. ine,/,‘ 4 • ..-.: • -_,•,',, : '€';‘;;!--',•;,33. ••• ... , , • _ . : ! 'ilii‘te0 e i .,. 13 ' .`" - - . ' ""4:•£..- ;k:!•:xc e„:•-i.I.,.,:' 14 te,a,,,:ài,-.: ..".....,-•-, :4-,:'<-e--41,..-2;1341'.' 15 . % : : " . •••/''' ' '.% ---1" - .. 're.......u.ed.g.,,,1'.•-ekt! • , , • • • "--,44 4 ". : - --4.- „
- .. e.,,:rf.,_,..,k‘ , • ,d .1 .../.._ ,. , : ... .: ,.. . . teta-‘,:Ài. •4••• •
•
-••••• ,
_ .
w er:i k. e' ' •!-cs.e 411 Are.- • • -.2 •;;. •
-•• • ,-:"1„.'•:‘ • . : • , • • .
4›"I rlç? • • e•e•-••:-. •re.- • • • • • eaL,- . • • 4 tv;^-4/::
(;•%K..re` ': - .- - - ,, • ,•.,..-= -,Ï,4 i---„;•, .-7,..-.)-...,-•,.-ifey y. . .. v .:,
44> e % _., e- , ,..e-', • .
• 4,1 e...•-•.• _.• -
f ■rc tie.a «42"
1...1 1te ;e•,,
d • ••;:•••=7• •••••
•,.e`` ,- '-'-'7,.'%›Ji5e:7?-:',4>■g;•: rr•• • ;s4
. t . .
•
ft•-: :`e• .4 • • : ,•:"en. zr7•-e: • .
1.: :.\-•: .'41;.- 14;1:31;%•.e4: • - • „Ai-- , .7. •
ale•o»,,.•ezer • ....g.x., % • - - : ...I. *;•- •,:'.‘fi..:. •te: • .: •••• e«.;,...,
« 71
1§ 1.1j
1•É
, • 17 -.
4.•••••:4;et - — 8 •
. --- - e ...i;•,:.• • ..,...-z --••• i ---,,,,..:,.. è 4«... , .: - ‘ x f ' 4.;:elir . i... ,1:')`-,Y,... ..2 .
.f4,„_-S'^.....' - ' • e ...rife: • •
:1,44es. • . •
ear7". • -
•
'r•-•
•t• - • -
A . '', • .--;* .r? • , • r 4
e;■••• ••:k P.:. .:',.e•\• e • ■--e•-•4 re-•;• •:"
••ei- r j'• L , 1! '›e•ît;;( ' ee'.' - .\ie r';''' 4 •,. .;*. ,4--,ve e. ... -•-•.: •• r,--.,...--..
,..,---.;,.,,,...- 2,; .....,;..::„% ... eéi„.„.., », ;2.
' ,
-'.
-1*h.), .e ,,,,) el:.6,,r,•ii4 f.5...,,,,....ii•e,.,. ,

Part 4. Sogvo and hakuren ovarian eggs. •
24. Ovarian egg
Stage II.
25. Ovarian egg
Stage III.
26. Ovarian egg
Stage IV.
20 7
11, Figure 10.1. .
p104 Part 3. Sogyo and'hakuren eggs.
19. Stage I. There is a small thickness of fat near the vegetative pole. The yolk is formed of 'numerous small globules, and no fatty droplets are present.
20. Stage The yolk globules are coalescing and increasing in size, but not penetrating the cortical protoplasm.
21. Stage III. The coalescence of the yolk globules has greatly progressed, and the large clusters of yolk globules are penetrating the cortical protoplasm.
22. Stage IV. The cortical protoplasm is atrophying and gathering • into one place, and the yoU.c is penetrating between the.chorion and the cortical protoplasm.
23. Ovarian egg The yolk globules are small, the Stage I. germinal vescicle (nucleus) is situated
in the centre of the egg.
The germinal vesicle is beginning to migrate to one pole of the egg. No change of the yolk globules is seen.
The germinal vesicle has migrated nearly to the periphery of the egg. A few of the yolk globules surrounding the germinal vesicle are beginning to increase in size.
The germinal vesicle has migrated to the periphery of the egg. The protoplasm is collecting around the germinal vesicle, and an increasing number of the yolk globules are increasing in size.
When the egg reaches Stage V it is ovulated (Stage of the spawned egg, Photograph 19). In this case Stage III and Stage IV of the spawned egg become Stage VI and Stage VII (Photographs 21 and 22).
77,57e1.?e7,'IMMMMe=27e.

■
• ••
21
7e, de`l're, ±
4,•....„..41,*...ze .
)c . .e. •1?, .
• 4.3. •• • -
e‘• ,-s--
■4:%•$
^,e%14.-.,•°4 • -• • "
";••••••-.7!•,7" • • • • •
•••.„• !•• ;••• :.--• -
• , • ! •••• 1
— -•
Pt • • ' - •
!••
Z: e•,?"7":>.
• • ;. • •, e ••-, •
te cuk•-- • • - — -71 • • • ,
• *" .; ■ • • -;'
• -se ,: •
.. • ••"- -u- :t
• ;Ix ;• ' ; - • •-.
• •
• ' 20 , . • J , •
ç,s
•-,
• •t • *,
'29 , • I29- b-
• s
•e‘ . .4( ••• - •
r ,`;-• • - - • --e I
•"j•? -
..!!% e '
•
2311? • •
-
•
.........124 a_ -\ .--:-..: ,..... ,-.,-.,. ,,,-..3-.‘-'.!..-- --.:--_,i • ie •'
..._ . 2— _„•'•*-"•••;:•,v-à•I'..1-;:iési.• .. -_,;e- ' . •
....- -,••• . I. 11.; . .1-i". •, '-',..., .2: • . -.'",- ...;_-, V., •
. • 4 ' e e;• «.%••". et.- S--;>-•, -';',-.4:'-)-•'–% ..• • x, ''' s .,:, ".
• e ..„1....,-.-...,......,......-...s....!..."-c...›;,......,,,......!,_...; .......t.. ., r ..i...:r.!•...-3.- - t----.:!: . --.1-.; -;--!q,..•- ,
:1 • - V' • •;--v"-- - ' / .....e-e-^ • ""` ,.....,•.r.:'.'• "r:- -..•,:tyee;:t.'" .
• ...; . ›-;‘-' --- ••••••;-74., , •-e,..-re....,....,..:4.-;.;—:•;-, • - -P!,--- .
.....,,, .„...;_,--, ,. ,....„-... ...„„ ,.„.;.,..•:- , :- „.......,,,-
li è•...e.e':-■•4>r=:à.--••,:-;-:•=:.,'.,t:..?.-' • • ' , - .1 N, ir-s._;e".X-..--...„-,.‘t::,,,;,.-••„:.-.-,,•;„ .
`i /z. \1/4.. .«,, .r•-,,!....: -Lz-„:.2,..:, ;:•:, — ---- \
1,, set-7">••,e'r ;>i•-•i,--ii -,;.J ..;•. \‘:s?? .:t4 ---.:_;.-.,:.. -.„
...,â • •.k. „.,..,-.•.„,.. . • •„, LI
... 7 nr é. • ....--.......,.. • „,...,
4 U 'Yles:%,
......ide......,::. -, 4
•

• 208
2. Change of egg quality with time.
The proportion of eggs which live or develop changed
with time after ovulation. The initial rate of development
in rainbow trout reared at 10 ° C to 12 oC was 75%. After 3 to
6 days it rose to 86% and then sank during the course of time
to 0% after 27 to 30 days. The initial rate in ayu at 15° C
to 16° C was 70% but was zero after 4 to 5 days, and in sogyo
and hakuren (at 20 °C to 22 ° C) an initial proportion of live
eggo of 34% became zero after 6 hours. .
Thus the quality of unspawned eggs decreases after
ovulation, and this decrease is more rapid in fish which
spawn at higher temperatures. For this reason the proper
moment for the collection of eggs must be precisely grasped
if eggs of good quality are to be obtained from fish which
spawn at high temperatures.
3. Hormone administration and the process of maturation
of the ovarian egg.
In some species spawning does not occur unless
hormone treatment is used. Since the quality of unspawned
eggs decreases after ovulation it is particularly necessary
with these species to grasp the moment of ovulation after
hormone treatment in order that eggs of good quality may
be procured.
p107
•

•• • 2 09
•
The process of maturation of the ovarian eggs of
sogyo and hakuren has been divided into 5 stages according to
the location of the nucleus in the egg and the size of the
yolk globules (Stages I to V) (Figure 10.1 -23, 2 )4V , 25, 26, 19).
After the hormone had been administered, the state of maturation
was measured by the use of a narrow vinyl tube on the nozzle of
a hypodermic syringe which was periodically inserted into the
genital opening in order to extract some of the ovarian eggs.
The hormone chosen was the sogyo or hakuren pituitary, and it
was administered twice, with an interval of 6 hours, to each
fish. The results are shown in Figure 10.2.
The fish (No. 3) treated at stage I and kept in water
at 20 °C to 22 °C reached the ripe stage V in 21 hours, and the
fish (No. 23) which started between stages II and III reached
stage V in 15 hours. At 25° C to 26°C, the fish (No. 24) which
started in Stage II reached stage V in 12 hours. The fish
(No. 28) which started in stage II but was treated with
pituitary only once.had not reached stage IV after 24 hours,
but' after being again treated with pituitary reached stage V
in about 12 hours.
From these preliminary experimental results, it is
considered that the moment of ovulation after hormone
treatment can to some extent be predicted from the stage of
maturation of the parent fish, the temperature of the water
P108

Degree of ; maturation ..tr (stage). •
n _u. Hours.
42 30. 36 48
210
Figure 10.2.
Stage of maturation and time elapsed since
hormone treatment of sogyo and hakuren.
The arrow shows.the time of hormone treatment.
.For stages I to VII see the present paper
and Figure 10.1 21 to 26.

• 211
•
and the quantity of hormone. It also considered:that a
catheter of the type described could be used for the
extraction of some of the ovarian eggs from large fish in
order to determine the degree of maturation of the ovary at
the time of treatment.
4. Assessment of the quality of . eges - spawned near the
It has been stated10-12 that the quality of eggs spawned
by rainbow trout and by ayu near the upper limit of spawning
temperature is unsatisfactory. An investigation was made to
find out whether the a -opraisal of the quality of eggs spawned
by rainbow trout and ayu reared at three different controlled
water temperatures could be expressed in terms of the
morphological stages described above.
. Rainbow trout were reared for 30 days from 3 November
to the following 21 January at temperatures of 10 ° C, 15°C and
18 00. Eggs in stages from I to II were extracted from all
f,ish in the 10 °C group. At 18 ° C eggs almost in stage III
were obtained after 20 or more days from the start, and the
group at 15° C was intermediate between the other two. The
mean proportion of development of the spawned eggs was 45% at
10 ° C, 1.6% at 1500 , and zero at 18 ° C, so that the high
temperature lowered the quality of the spawn.
•

• 212
Next ayu fry from Lake Biwa were reared to full growth
for 115 days from 28 June to 20 October at 15o C, 20 0 C and 25 o C,
and the relationship between the proportion of development and
the temperature was measured in eggs ovulated naturally or by
means of hormone treatment. In spawn ovulated naturally at
15° C and at 20 0 0 there was not much difference in the frequency
of arrival at a particular stage or in the proportion which
developed, but in eggs which were spawned after hormone
treatment there were more in stage III at 20 ° C than at 15o C,
and the average rate of development at 15° C was 25% whereas
at 20 o C it was 2.5%. At 25o C there was some vitellogenesis,
but there was also much degeneration, and there was neither
natural nor hormone induced spawning.
It is thus found that the classification by stages
which is applicable to eggs incubated but not spawned after
ovulation is also applicable near the upper temperature limit
for spawning, and that these stages can in general be used as
criteria on which to base appraisal of egg quality.
5. Egg quality and egg fractionation by centrifuging.
When eggs were homogenized and fractionated by
centrifuging, (3000 rpm for 30 minutes) the contents of the
eggs are separated into several layers in the centrifuge.
The number of layers and the volume of each layer in proportion
p109

• 213
to the total volume in rainbow trout eggs was found to remain
unchanged for 36 hours after ovulation. However sogyo and
hakuren eggs showed the following fivefold pattern of
change (Figure 10.3).
Type 1. Four layers are formed. In order from the centre
they are
A layer of protoplasm
A fluid layer (transparent)
A yolk layer
A layer containing cortex and chorion.
Type 2. A part of the protoplasm sinks into the transparent
fluid layer.
Type 3. The sinking of the protoplasm is pronounced, and it
is difficult to distinguish.it from the fluid layer.
Type 4. The protoplasm and fluid layers can be distinguished
but the fluid layer is turbid.
Type 5. The protoplasm layer sinks through the fluid layer
to the centrifugal side, and the four layers in
order from the centre are:
The fluid layer (turbid).
The protoplasm layer.
The yolk layer.
The layer containing cortex and chorion.
The fluid layer widens and the protoplasm and yolk
layers shrink.

1 1
A
D
A
D
0
2 Time elapsed since 3 ovulation. (Minutes)* 6
•
:11
22
25
Figure 10.1.
The centrifue.al fractionation patterns Of soeyo and
hakuren . eqes, and the change with elapsed time of the
percentaq- e of parent fish which show each pattern.
type
214
16.7 16.7 8.3 33.3
memme Kmemi ...........« .... N..1 EMM \s 'N
100 . \\\.
100, \\
50.0 50.0
. - 100.
100 \\
\ 11 \ \
-.‘ \ 100
\ • . • 100
■ 100 \
The arrow shows the direction of centrifugal force.
A. Protoplasm layer. B. Fluid layer. C. Yolk layer. D. Cortex and chorion layer.
111› * Sic, but the text says hours. Translator.

215 • Directly after ovulation all stages from 1 to 5
were displayed by some parent fish, but as time passed many
parent fish changed type, and after 22 hours all parent
fish showed type 5. There was a connection between the type
of stratification of the spawn and the average proportion of
development. In type 1 it was 51% and decreased as the type
changed, being zero in type 5.
It thus appears that the quality of the eggs of
sogyo and hakuren can be to a certain extent determined by
cer..Jrifuging a portion of the spawn. p110
6. Egg quality and other criteria.
Criteria other than those already described which
were investigated were the colour and pH of the'eggs and
the refractive index of the yolk. However no changes were • p111
found and there was a large individual scatter of the values,
so it is considered that these criteria are not suitable
for the appraisal of egg quality.
Experiments were made with the objective of developing
methods for the appraisal of egg quality. If eggs of good
' quality with satisfactory fertilization and development ratios
can be obtained, the yield of fry could be increased and
stabilized.

• 216
•
•
It was found that the quality of the eggs of rainbow
trout, of ayu (Plecoglossus altivelis), of sogyo (Ctenopharyncodon
idellus), and of hakuren (Hypophthalmichthys molitrix), could
to a certain extent be determined from the transparency of the
spawn, from the size and distribution of fatty droplets and
from the morphological features shown in histological sections
of the cortical protoplasm, the yolk globules and the fatty
droplets. It was also shown that it is possible to determine
to some extent the quality of sogyo and hakuren spawn from the
stratification pattern obtained when a portion of the spawn
was centrifuged.
Since these criteria can be applied not only to eggs
which have not been spawned after ovulation, but also to those
formed at the upper limit of spawning temperatures and to the
quality of abnormal eggs directly after ovulation, it is
possible to use them as general criteria for the appraisal
of egg quality.
Eggs formed near the upper limit of spawning temperatre
were of extremely low quality, and the quality of eggs not
spawned after ovulation also decreased with the passage of
time. Since this decrease of quality was particularly rapid
in species in which the temperature suitable for spawning is
' high, the period for collecting eggs from such species is
short. Consequently it is to be supposed that the moment of
ovulation must be precisely ascertained if eggs of good quality
are to be collected.

• 2 17
References. .
1. ITO Takashi, IWAI Hisao, FURUICHI Tatsuya.-
Ayu shubyo seisan ni kansuru kenkyu - XXXIV.
Kaku shu chiayu no yosei shingyo no jinko juseiran
to fuka shigyo no tokusei.
Kiso mikawa kako shigen chosa hohoku No. 4 521 -542 (1967).
T. Ito, H. Iwai, T. Furuichi.
Studies relating to the production of ayu fry - XXXIV.
Artificial fertilization of culture fish as parents
of various types of young ayu, and the characteristics
of the young,fish.
Reports of studies of the resources of the Kiso Mikawa
estuary, No. 4, 521 - 542 (1967).
2. Hiroshima ken tansuigyo shido jo.
Kurogoi shingyo yosei jiryo to ranshitsu, shigyo
ni hikaku (2).
Showa 47 nendo jigyo jisseki 12 (1972).
Hiroshima prefectural freshwater fish promotional centre.
Comparisons of the diet of cultured black carp with -
quality and fry (2).
Annual reports of progress 12 (1972).

• 218
3 , KONDO Kei.
Kingoi no shubyo seisan ni kansuru kenkyu I.
Sairan, saibyo ni kansuru kiso chosa.
Hiroshima ken tansuigyo shidojo chosa kenkyu hohoku No.12.
Kingoi tokushu 1 - 13 (1973).
K. Kondo.
Studies of the production of brocade carp fry - I.
Basic studies of the collection of eggs and of fry.
Research reports of the Hiroshima prefectural
freshwater fish promotional centre, No. 12.
Brocade carp issue, 1 - 13 (1973).
4 • TACHIKAWA Wataru.
Nijimasu shingyo kairyo shiken - I.
Shingyo jiryo no tekisei tanpaku ryo o motomeru
•
shiken narabi ni botai to ranshitsu ni kansuru kento.
Gifu sui shi ho No. 12, 64 - 88 (1965).
W. Tachikawa.
Experiments on the improvement of parent rainbow
trout - I.
Experiments on the quantity of protein required in
parent fish diet and an investigation of the
relation between egg quality and the maternai body,
Research reports of the Gifu fisheries research
laboratory, No. 12 64 - 88 (1965).

• 2 19
•
S. KOGISO Takuo, ISHII Shigeo.
Ayu no shingyo yosei jiryo narabi ni seijuku
togyo ni kansuru shiken.
Gifu sui shi ho No. 13, 19 - 25 (1966).
T. Togiso, S. Ishii.
The diet of cultured parent ayu and experiments
on the control of maturation.
Research reports of the Gifu fisheries research
laboratory No. 1? 19 - 25 (1966).
6. KONDO Kei, FUSHIMI Toru.
Ayu shigyo yo:,ei to sairan shiken (jiryo no sa ni
yoru sairan oyobi fuka kekka).
Hiroshima ken tansuigyo shidojo chosa kenkyu hohoku
No. 10, 73 - 96 (1971).
K. Kondo, T. Fushimi.
Experiments on diet and egg collection with ayu
parent fish. (Results on the variations produced
by various diets on egg collection and hatching).
Research reports of the Hiroshima prefectural
freshwater fish promotion centre .
No. 10 73 - 96 (1971).
p112
•

220
7. ISEDA Hiroshi, HIRATA Mitsuru, ITASAKI Kiyoshi.
Ayu shingyo yosei shiken - * I.
Kumamoto sui shi jigyo hohoku showa 46 nendo 336-363 (1971).
H. Iseda, M. Hirata, K. Itasaki.
Experiments on the rearing of parent ayu - I.
1971 progress report of the Kumamoto fisheries research
laboratory 336-362 (1971).
8. GOTO Katsuaki, KOGISO Takuo, HONJO Tetsuo.
Yosei ayu no ranshitsuni kansurtukenkyu - I.
Gifu sui dai 9 kai ayu bukai shiryo I (1973).
• K. Goto, T. Togisr, T. Honjo.
Studies of the quality of cultured ayu eggs - I.
The appraisal of egg quality ..
Materials from the ninth meeting of the ayu section,
Gifu fisheries research laboratory, I. (1973).
9. ZHONG Lin, LI You Guang, ZHANG Song Tau, LIU Jia Zhao,
CHEN Fen Chang.
Jia yu di sheng wu he ren _gong fan ji.
Ko xue qu ban she (1965).
CHUNG* Lin, LI Yu Kuang, CHANG Sung T'ao, LIU Chia Chao,
CH'EN Fen Ch'ang.
The artificial breeding of domestic fish.
Science publishers (Peking) 1965.
* This reference is in Chinese. The authors' naines have been transliterated in the modern (pin-yin) romanization, but the translation gives the names in the Wade-Giles romanization more commonly used in English language publications. Translator.

•
221
10. OWATARI Satoshi.
Shingyo yoshoku ni yoru sairan shiken.
Saitama sui shi ho 18 86 - 90 (1963).
S. Owatari.
Experiments on egg collection from cultured parents.
Reports of the Saitama fisheries research laboratory,
18 86 - 90 (1963)
11. OWATARI Satoshi.
Shingyo yoshoku shiken.
Saitama sui shi ho 20 56 - . 69 . (1964).
S. Owatari.
Experiments on cultured parent fish.
Reports of the Saitama fisheries research laboratory,
20 56 - 69 (1964). .
12. KOGISO Takuo, FUNASAKA Yoshiro, ISHII Shigeo.
, Ayu no shingyo jiryo narabi ni seijuku togyo ni
kansuru shiken.
Gifu sui shi ho, No. 11 74 -83 (1965).
T. Kogiso; Y. Funasaka, S. IShii.
The diet of cultured parent ayu, and experiments on
the control of maturation.
Research reports of Gifu fisheries research laboratory.
No. 11 74 - 83 (1965).
•

• 222
•
13. OWATARI Satoshi.
Nijimasu shingyo shiiku ni .okeru suion sza sairan
seiseki ni oyobosu eikyo ni tsuite.
Saitama sui shi ho 28 58 - 78 (1969).
S. Owatari.
On the influence of the culture temperature on the results
of egg collection from parent rainbow trout.
Reports of the Saitama fisheries research laboratoty,
28 58 - 78 (1969).
14. ITO Takashi.
Ayu shubyo no jinko seisan ni kansuru kenkyu LXX.
Ayu no jinko saibyo gijutsu no genjo to mondaiten.
Ayu no jinko hoshoku kenkyu.. 1 1 - 7.
Mie ken tate dai suisan gaku bu tansurzoshoku
gaku kenkyushitsu.
•• T. Ito.
Studies of artificial production of ayu fry. LXX.
The present state and problems of the technique of
artificially obtaining ayu fry.
Studies of artificial ayu culture 1 1 - 7.
Freshwater propagation laboratory, Faculty of
Fisheries, Mie prefectural University.
•

• 223
•
15. NOMURA Minoru, TAKASHIMA Fumio, OWATARI Satoshi,
UEMATSU Zenjiro.
Nijimasu no kajukuran keisei katei ni tsuite - I.
Kajuku katei ni okeru juseiritsu sono ta no hendo.
Showa 41 nendo suisan gakkai shunki daikai, koen (1966).
M. Nomura, F. Takashima, S. Owatari, Z. Uematsu.
On the process of oogenesis during maturation of
rainbow trout - I.
On fluctuations of the rate of fertilization during
the process of maturation.
Papers of the 1966 spring meeting of the Japanese
Society of Scientific Fisheries (1966).
16. NOMURA Minoru, SAKAI Kiyoshi, OWATARI Satoshi.
Nijimasu no kajuku rankeisei katei ni tsuite - IV.
Hakken ritsu to tamago kussetsu ritsu to no kankei.
• Showa 42 nendo suisan gakkai shuki daikai koen (1967).
M. Nomura, K. Sakai, S. Owatari.
On the process of oogenesis during maturation of
rainbow trout - IV.
The relation between the proportion developing and
the refractive index of the eggs.
Papers of the 1967 autumn meeting of the Japanese
Society of Scientific Fisheries (1967).

•
224
17. SAKAI Kiyoshi, TSUCHIYA Minoru INABA Denzaburo, NOMURA Minoru
Sogyo no horumon shori go no ransoran no jotai to
sairan seiseki.
Showa 42 nendo suisan gakkai shuki daikai koen (1967).
K. Sakai, M. Tsuchiya, D. Inaba, N. Nomura.
The condition of ovarian eggs in sogyo after hormone
treatment, and the results of egg collection.
Papers of the 1967 autumn meeting of the Japanese
Society of Scientific Fisheries (1967).
18. SAKAI Kiyoshi, TACHIKAWA Wataru, NOMURA Minoru.
Ayu tamago no kajuku katei ni tsuite.
Showa 48 nendo suisan gakkai shunki daikai koen (1973).
K. Sakai, W. Tachikawa, N. Minoru.
On the process of maturation of eggs in ayu.
Papers of the 1973 spring meeting of the Japanese
Society of Scientific Fisheries (1973).
19. SAKAI Kiyoshi, NOMURA Minoru, KOGISO Takuo, GOTO Katsuaki.
Ayu tamago no kajuku katei ni tsuite
Yosei shingyo to tennen shingyo ni okeru hikaku.
Showa 49 nendo suisan gakkai shunki daikai koen (1974).
K. Sakai, M. Nomura, T. Kogiso, K. Goto.
On the process of maturation of eggs in ayu II.
Comparison of cultured and natural parent fish.
Papers of the 1974 spring meeting of the Japanese
Society of Scientific Fisheries (1974). •

• 225 ••
11. Marine Fish.
Michiyasu KIYONO.
(Faculty of Agriculture,. Tokyo University)..
Experience in rearing has suggested that the survival
of fish larvae is greatly influenced not only by the
environmental conditions but also by the quality of the eggs1
.
Nevertheless, this has been little studied. It has been
clearly shown2 ' 3 that the fertilization and hatching of the
eggs are greatly influenced by the ripeness of the eggs (the
length of time after ovulation to exposure to the sperm), by
the treatment of the parent fish just before sp , wning, and by
the timing of the hormone administration. Further studies
are needed to determine whether the problems in the rearing
of fry which are due to egg quality arise simply from the
methods used in the collection of the eggs or whether they
arise from earlier factors such as the state of nutrition of
the parents4 , the rearing environment, or the state of
development of the methods used for culturing the parent fish
and of collecting eggs.
The proauction of fry would be helped if ways not yet
established could be found of appraising the quality of the
eggs in the spawn. In this paper good quality eggs are
provisionally defined, on the basis of the rearinp; of larvae,

•
•
226
as eggs from which satisfactory fry are hatched after they
have survived incubation. A number of investigations have
been made in order to test methods of determining the
proportion of good quality eggs in the spawn, and an outline
introduction to these investigations follows.
1. The characteristics used in determining egg oualitv.
Many of the marine fish of importance in the production
of fry produce floating eggs. The fish used in these
investigations all lay floating eggs and include the black
porgy (kurodai) Mvlio macrocephaluF1 BASILEWSKY, the sand borer
(kisu) Sillago sihama FORSKRL, and the stone flounder (ishigarei)
Kareius bicoloratus BASILEWSKY. Eggs naturaily spawned after
hormone treatment were obtained 6 times from 6 parent porgy in
May and June 1971, and 12 times from 7 parent sand borers in
August and September 1973. Eggs were collected by stripping
25 times after hormone treatment of 13 flounder in December
1973 and January 1974.
With the idea of estimating the proportion of good
quality eggs in the spawn, the characteristics and the
relations between the characteristics of the spawn were
investigated. Careful attention was given to changes of the
feed in the rearing environment, which greatly influence the
satisfactoriness of the survival of the larvae. It was
decided to measure the proportion of good quality eggs,
p114

2 27
the number of eggs in the spawn which did not hatch, the
number which died before the yolk was absorbed, and the
number of fry which hatched but were considered to be abnormal
(in the following called the abnormal egg rate*). The
results are shown in Figures 11.1 to 11.3. The individual
properties will be described below.
1.1 Egg density.
The density was determined by observing whether the
eggs floated or sank in seawater of known density kept at the
proper temperature . This temperature was 20 °C to 23 00 for
porgy, 23.5° C to 24.5°C for sand borer, and 11 °C to 13 ° C for
floun der.
Eggs with opaque portions when discharged were all
unfertilized and sank to the bottom of high density seawater
(C1 240/00). Only those which were transparent were therefore
measured. The eggs in a single group were all very similar
in density, the differences being 0.0002 to 0.0005. Groups
of flounder eggs containing a low proportion of abnormal eggs
were found to have a tendency to gather together, but no such
tendency was found within the limits of the experiments in
sand borer eggs. It was also observed that unfertilized eggs
and eggs with abnormal development often tended to sink before
incubation.
* For convenience, this is the ratio to the total number of floating eggs,

• 228
•
Figure 11.1.
Properties of porgy eggs and larvae.
LARVAL PROPORTION
'ARENT IATCHING SURVIVAL OF EGGS EGG FATTY
;II O. RATE RATE (YOLK WITH MULT.. DIAMETER** DROPLET DENSITY
ADSORBTION tPLE FATTY DIAMETER**
STAGE)* DROPLETS*
.----... -
% 0/
.2,, % mm
1. 99 95 2 0.83±E0n92 — 1.0233 • 2 100 90 7 0.91±0.089 0.24±0.050 —
3 • 99 62 9 0.87±0.093 0.23±0.036 —
4 98 50 21 0.85±0.068 0.23±0.037 1.0233
.5 99 98 2 0.85±0.088 . 0.24±0.048 1.0244
92 40 — 0.87±0.075 —
The abnormal egg rate is (100 - percentage of fry
surviving to the yolk absorption stage) %. The egg
diameters are given as mean standard deviation.
Many eggs from No. 3 and No. 6 were in the cloudy
white morula stage when gathered.
Percentage of the floating . eggs gathered.
Observed in the morula stage.
FI ••
* *
•

• 229
•
Figure 11,2.
Properties of sand borer eges and larvae.
FRY
PARENT SURVI • HATCHED LARVAE , ."0
F ISH No. HATCH VAL EN
•—• INC RATE. EGG FATTY • ITY LENGTH YOLK YOLK VOLUME/
COLLECTION SATE (YOLK DIAMETLR DROPLET • * VOLUME LENGTH
No. (,:l ABSORB *4 DIAMETER
TION 4* STAGE)
4
. _ „, " —
'A 0.,
,0 mm mm mm mm 1- 1 99 78 0.67±0.013 — 1.0214 1.46±0.044 — —
1- 2 88 16 0.68±0.007 0.15 ±0.0041.0207 — — —
1- 3 100 70 0.67±0.014 0.13±0.0061.0215 1.45±0.050 0.18±0.0200.124±0.028
2—Ï 95 90 0.65±0.021 — 1.0185 1.43±0.029 —
2-2 99 80 0.69±0.009 — 1.0192 1.52±0.063 0.18±0.0220.118±0.014
3-1 97 90 0.69±0.017 0.16±0.0051.0217 1.501-0.066 0.19±0.025C.127±0.020 .
4-1 91 84 0.68±0.015 0.16±0.0061.0206 1.50±0.047 0.17±0.0250.113±0.014
4-2 88 58 0.70±0.006 0.15±0.0041.0197 1.52±0.053 0.19±0.0320.125±0.022
5- 1 0 — — — 1.0232 — — —
5- 2 98 74 0.66±0.012 0.16±0.0051.0230 1.39±0.038 0.15±0.0170.108±0.010
6- 1 96 37 0.70±0.010 0.15±0.061.0195 1.52±0.054 0.22±0.0040.145±0.033
7- 1 86 7 0.65±0.010 0.15±0.0031.0235 1.40±0.051 0.19±0.0300.136±0.025
Values for egg diameter are + standard deviation.
In 1-2, 5-1, 5-2 and 7-1, many opaque eggs in the morula stage were found at the time of .collection..
Relative to the number floating when collected.
Observed in the morula stage.
***• Yolk volume =axbx c.
* 41.
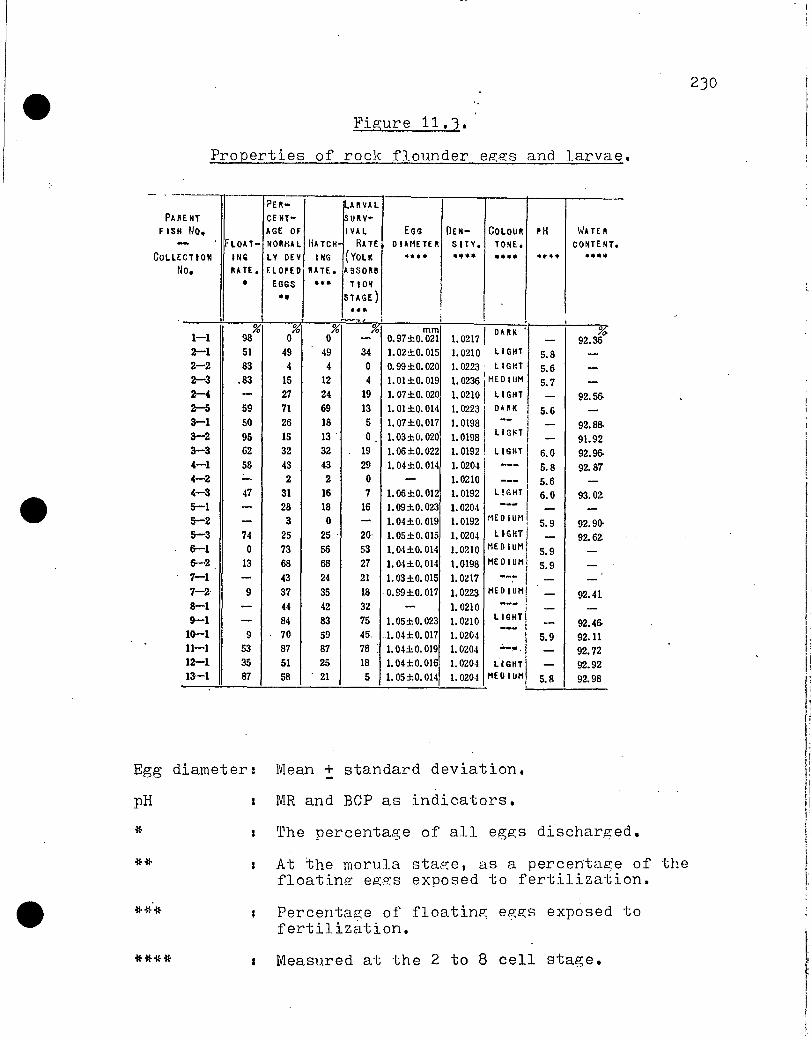
•
FI
COL
•
Figure 11,3:
Properties of rock flounder eggs and larvae.
PER.. AVAL ARENT CENT- SH No. AGE OF IVAL EGG DEN- COLOUR PH WATER --, ' FLOAT... NORMAL HATCH RATE DIAMETER SITY. TONE. CONTENT.
LEcTioN iNG Ly bey 1 NG (You( 4... .*** ...I, 1.4.1. •4, 41.
No, RATE. ELOPED RATE. ADSORB • EGGS '1 ** TION
•* STAGE) ***
■-,-.., % % % % mm I DARK g.
1-1 98 0 0 - 0.97±0.021 10217 - 92.36
2-1 51 49 • 49 34 1.02:10.015 1.0210 L IGHT 5. 5 _
2-2 83 4 4 0 0.99±0.020 1.0223 L IGHT 5.6 -
2-3 .83 15 12 4 1.01±0.019 1.0236 MED I um 5.7 -
2-4 - 27 24 19 1.07±0.020 1.0210 I L iGHT _ 52.56
2-5 59 71 69 13 1.01±0.014 1.0223 DA IIK 5.6 -
3-1 50 26 18 5 1.07±0.017 1.0198 - - 92.88.
3-2 95 15 13 • 0 • 1.03±0.020 1.0198 LIGNT - 91.92
3-3 62 32 32 19 1.06±0.022 1.0192 L 8 GHT 6.0 92.96.
4-1 58 43 43 29 1.04±0.014 1.0204 --- 5.8 92.87
4-2 -. 2 2 0 - 1.0210 --- 5.6 -
4-3 47 31 16 7 1.06±0.012 1.0192 L I GHT 6.0 93.02
5-1 - 28 18 16 1.09 ± 0.023 1.0204 --- - -
5-2 - 3 0 - 1.04±0.019 1.0192 MED i um 5.9 92.90.
5-3 74 25 25 20 1.05 ± 0.015 1.0204 LIGHT - 92.62
6-1 0 73 56 53 1.04±0.014 1.0210 ele "" 5.9 -
6-2 13 68 68 27 1.04±0.014 1.0198 MEDIUM 5.9 -
7-1 - 43 24 21 1.03±0.015 1.0217 --":" - -
7-2 9 37 35 18 0.99±0.017 1.0223 MED tun • - 92.41.
8-1 - 44 42 32 - 1.0210 - - -
9-1 - 84 83 75 1.05±0.023 1.0210 L IGHT- 92.46. ---
10-1 9 70 59 45.1.04±0.017 1.0204 5.9 92.11
11-1 53 87 87 78 1.04±0.019 1.0204 ---.. - 92.72
12-1 35 51 25 18 1.04±0.016 1.0204 L i c 87 ' - 92.92
13-1 87 58 21 5 1.05±0.014 1.0204 MED I UM 5.8 92.98
Egg diameter: Mean + standard deviation.
PH : MR and BOP as indicators.
: The percentage of all eggs discharged.
: At the morula stage, as a percentage of the floating eggs exposed to fertilization.
Percentage of floating eggs exposed to fertilization.
23 0
**
***
** * * Measured at the 2 to 8 cell stage.

23 1
1.2 Egg colour tone.
Some of the floating eggs were transparent when
gathered, but many were yellow. Three shades of yellow could
be distinguished in the flounder eggs. The sample was small,
but within the experimental limits, groups of a dark yellow
colour tended to have a high proportion of abnormal eggs.
p115
1.3 Egg diameter and fatty droplet diameter.
The values for groups of flounder eggs containing
small percentages of abnormal eggs had a slight tendency to
be concentrated. There was also a tendency towards a small
number of abnormal eggs in groups with a large degree of
scatter (as shown by the standard deviation). These tendencies
were especially noticeable in the sand borers.
1.4 Length and yolk volume of newly hatched larvae.
An investigation was made of the sand borer larvae.
Groups in which the average value of the specific yolk volume • pt16
(yolk volume / length), and the standard deviation of the
specific yolk volume were both low, tended to be those in
which the percentage of larvae which died before reaching the
yolk absorption stage was also low.
1.5 The percentage of abnormal eggs.
• The observations made includes-
When the percentage of sinking eggs (the ratio to the
total number of eggs discharged) is high, the percentage which
hatches (the ratio to the number which float) is in general low.

23 2
In the groups in which the proportion which hatch is
low many of the larvae die before reaching the yolk
absorption stage.
If opaque eggs are found in large numbers on the
bottom of the tank when eggs are collected after natural
spawning, many will die before reaching the yolk absorption
stage.
Black porgy eggs normally contain a single fatty
droplet, but if the percentage of eggs which contain multiple
droplets is high, many of the larvae die before the yolk
absorption stage.
Many of the eggs which progress to or beyond the
morula stage will hatch.
The pH of flounder eggs was measured after homogenization.
It was found to be 5.8 to 5.9 in grOups with few abnormal eggs,
and 5.6 to 5.7 in groups in which the percentage of abnormal
eggs was high.
2. Discussion of methods of appraisal of egg quality.
It appears from these results that groups of eggs in
which abnormal eggs were numerous before hatching produced
abnormal larvae. It seems that the proportion of good quality
eggs in the spawn can to some extent be estimated from the
proportion of abnormal eggs at the time of discharge or in the
early stages of development. It also appears that further
criteria, such as the density and the colour, could be obtained
from future trials with separate species.
t

• 233
•
•
The values of pH found for rock flounder are rather
lower than those of Fundulus and medaka (Oryzias latipes) 6 ,
but it can be deduced that the normal and the abnormal eggs
differ in physical composition.
When groups in which the percentage of larvae which
die before reaching the yolk absorption stage is low are
compared with those in which the percentage is high, it is
found that there is relatively little variation in the yolk
volume and that such groups can be considered homogenous.
There was to be founa some relation between the
proportion of abnormal eggs and the degree of scatter of the
egg diameters and of the fatty droplet diameters, However
it was found that in individual sand borers and flounders
which discharge eggs several times, the values, and the
scatter of the values, of the egg and fatty droplet diameters
may vary even between discharges from the same fish. As
silown in Table 11.4 it appeared that there was some connection
between these variations and the administration of the
hormones. In order to investigate the connection between
these values and the quality of the eggs, it will be necessary
to clarify the mechanism which through its action on the eggs,
determines the egg diameter.
Larvae can survive for a long period when reared
without food but they will strongly degenerate unless there
is sufficient food in the surroundings. Some investigations
p118

Parent fish No. -- Collection
Days since EFe. hormone diameter*. treatment
mrn- 2 0.65±0.021
0.69±0.009 2 0.68±0.015 5 0.70±0.006 2 0.67±0.013 5 0.68±0.007 2 (2fiD T Imt) 0.67±0.014
2-1 2-2 4-1 4-2. 1-1 1-2 1-3
• 234
Figure 11.4.
Changes in egF diameter and +he number of days
elapsed after hormone treatment (sand borer eggs).
* Mean + standard deviation.
•
•

• 235
•
have been made to estimate the length of the survival Period.
It is known that there is in herring a normal relationship
between the dry weight of the eggs and the number of days of
survival of the larvae without food ' . In the present
experiments we measured the number of days of survival
(defined as the number of days in which the number of
survivors was reduced to 50%)of each of the groups which had
survived to the yolk absorption stage. This was compared
with the many other characteristics which depend on the
quabtity of yolk* such as the egg diameter, the volume of
yolk in the larvae, and the dry weight of the eggs**, and
also with the density of the eggs and the abnormalities, but
no definite tendencies appeared.
Future investigation of the quality of the eggs must
first of all clarify the problems concerning fry production
which have been discussed in this paper. It will also be
necessary to include the . wayè.ihWhich'quality participates
in the survival and growth of the larvae after the yolk
aborption stage.
The values including the abnormal eggs and abnormal larvae.
** Values calculated from the egg diameter, dehsity, and water content.
•

• 236
•
•
References.
1. KURATA Hiroshi.
Nishin chigyo no'shiiku ni tsuite.
Hoku sui ken ken ho 20 117 - 138 (1959).
H. Kurata.
On the rearing of herring fry.
Bulletin of the Hokkaido Regional Fisheries Research
Laboratory, 20 117 - 138 (1959).
2. HIBIYA Takashi.
Horumon ni yoru gyorui no seijuku, sanran no kontororu.
Sui san zo shoku 12 (4) 239 - 259 (1965).
T. Hibiya.
Control of maturation and spawning 6f fish by
means of hormones.
Fish breeding 12 (4) 239 - 259 (1965).
3. HIRANO Reijiro.
Kurodai no chigyo
Nichi sui shi 11 (6) 567 - 569 (1969).
R. Hirano.
Rearing of black porgy fry.
Bulletin of the Japanese Society of Scientific
Fisheries 31 (6) 567 - 569 (1969).

• 237
•
4. ITO Takahashi, ta.
Ayu shubyo no jinko seisan ni kansuru kenkyu VIII.
Yoshoku ayu no jinko juseiran oyobi fuka shigyo
no tokusei .
Kiso mikawa kako shigen chosa hohoku (2) 825 -381 (1965).
Ito et al.
Studies relating to the artificial production
of ayu fry VIII.
Characteristics of fry hatched from artificially
fertilized eggs of cultured ayu.
Reports of studies of the resources of the Kiso Mikawa
estuary (2) 825 - 881 (1965).
T.
5) R. Cnàbissus :Intracellular hydrogen-
. Ion concentration studies.-V The pHpf
the protoplasm of the Fundulus egg.
J. Cal.. Comp. Physiol., 1. 65-40
(1932).
6) T. Yamaxoyo: Studies on the rhyth-
mical movements of the e3.rly embryo
. of Oryzias -V1 Anaerobic
• movements and oxidation-reduction
potential of the egg limiting the
rhythmical movements. J. Fac. Sci.
Tokyo. Imp. Univ. sec. IV (Zool.), 4.
233--247 (1936).
7) J. l-1. S. &Aetna and G. HE UPEL : The
influence of egg size on herring larvae
(Clupea harengus L.). J. du Cons.. 28, 211-440 (1963).

• 238
QUESTIONS.
I. Present positions and uncertainties.
Chairman: Tsuyoshi HACHIZUKA.
(Kochi University).
Parent fish for eg.9 collection.
Chairman: Kiichiro YAMAMOTO.
(Hokkaido University, Faculty of Fisheries).
For Mr. Takano.
Tamura. (Nagoya University, Agriculture).
To what extent do secretions rather than the villi of
the ovary wall govern or participate in the transport of ova?
Takano. (Hokkaido University, Fisheries).
I believe that the turbulent motion of the secreted
fluids around the egg contributes to local motion in the
reproductive cavity.
Hibiya. (Tokyo University, Agriculture).
Fish begin to accumulate yolk when they reach the
appropriate age. What do you think the trigger mechanism to be?
Takano.
The direct cause of the start of yolk accumulation is
the activation of the productive cells after the gonadotrophic
hormone (GTH) is produced by the pituitary, but the factors
required for this are not known.

•
239
For Mr. Kato.
Hibiya.
How is the weight of the ripe eggs measured? What
is the teMperature used for rearing at the Nikko branch of
the Freshwater Fisheries Research Laboratory?
Kato. (Nikko branch, Freshwater Fisheries Research Laboratory).
The ripe egg weight includes the eggs which have been
squeezed out and the total weight of the eggs which remain in
the body cavity, (partly emptied), but it does not include
the ovary, the ovary fluids or the unripe eggs. The
temperature was 9.5 + 1 00.
For Mr. Yamazaki.
Hibiya.
The cells which produce GTH were stimulated to high
activity by control of the temperature>of the goldfish, but
what happened to the cells which produce thyroid stimulating
hormone (TSH)?
Yamazaki. (Hokkaido University, Fisheries).
Insufficient observation was made of the TSH
producing cells.
•

• 240
Hibiya.
Were any forerunner changes observed in the GTH
producing cells when the temperature was controlled?
Yamazaki:
Some changes are to be expected in the hypothalamus,
but they have not yet been investigated.
111.1 121-Ya.
What do you suppose to be the site which is stimulated
by temperature?
Yamazaki.
The whole body is affected by changes of temperature,
but it is thought that the ovary is made more sensitive by
hormones. Recently there has been discussion of the action
within certain temperature limits of an isohormone.
Hibiya.
Temperature stimulation invigorates the TSH producing
cells, and promotes secretion from the thyroid. Do you think
it possible that this is connected with the secretion of LH
from the pituitary?
Yamazaki.
It has long been known that the thyroid hormone promotes
growth but it is difficult to think of any direct connection
with maturation. Of course one may conjecture that its influence
on the metabolic level could result in promoting maturation.

• 241
Oguri. (Nagoya University, Agriculture).
It is possible that methyltestosterone promotes
maturation of the eggs, but how are we to explain its
physiological action?
Yamazaki.
At the time of maturation the skin of both male and
female salmonids thickens. This has been shown experimentally
to be caused by both male and female hormones. It has recently
been shown that the females also secrete male hormone from the
ovary, and it is supposed that this may have something to do
with the maturation of the eggs, especially in the early stages.
Hirose. (Tokai Regional Fisheries Research Laboratory).
The action of GTH on the interrenal causes the
secretion of corticoids, and these ipromote ovulation, but the
amount of corticoids needed to cause ovulation experimentally
iS large. If instead, corticoids were secreted from the ovary,
could one suppose that ovulation occurs when the local
concentration is high?
Yamazaki.
Corticoids are known to be effective in the ovulation
of catfish and medaka, but I have heard that they are not
effective in salmonids or mullet. Evidently there are
differences between species. Of course it is undeniable
that a condition of strong local concentration would increase
their effectiveness.
p121

242
••
Environment maturation and spawning.
Chairman: Tamotsu TAMURA. (Nagoya University, Agriculture).
For Mr. Yoshioka.
Shiraishi. (Tokyo University, Oceanology).
When the length of the day is limited, what is the
effect of unnatural day lengths?
Yoshioka. (Hokkaido University of Education).
When very short or very long photo periods, such as
8 hours, 48 hours, or 64 hours, which cannot be obtained in
natural conditions, are used, maturation is not so well
encouraged as with a 24 hour period,
For Mr. Harada.
Murakami. (Hiroshima University, Agriculture).
What is the definition of "the proportion which
develop?
Harada. (Kinki University, Agriculture).
This is the percentage of live eggs which progresses
into development, and here it is the percentage which, about
8 hours after fertilization, have reached the 8 to 16 cell stage.

243
Yamato. (Hokkaido University, Fisheries).
If cleavage does not occur after exposure to sperm,
one cannot speak of fertilization. Can one not therefore
say that the proportion which develop is the same as the
proportion fertilized?
Harada.
I would say that that is so immediately after exposure
to sperm. However, in time, development may stop and death
of the egg may be produced, so that the proportion developing
is not in general the sa -, e as the proportion fertilized.
IV. Maturation and metabolism.
Chairman: Michizo SUYAMA. (Tokyo University of Fisheries).
For Mr. Takashima.
Yamamoto.
Has it been reported that changes in the diet produce
quatitative and qualitative changes in the triglycerides in
the fatty droplets?
Takashima. (Tokyo University of Fisheries).
There are very few examples of investigation of such
phenomena in fish eggs, and I know of no such study.

• 244
Tamara.
Has there been any study cif a bad influence on
oogenésis of unnatural circumstances such as those produced
by free fatty acids?
Takashima.
don't believe that this has yet been studied in
fish. However I have heard that growth, survivability, and
oogenesis in rainbow trout were not affected by rearing with
hydrolyzed oils containing C 17 .
For Mr. Aida.
11, Oeuri.
Estradiol 17p was chosen as an estrogen, but were
other estrogens tested?
Aida. (Tokyo University, Agriculture).
Diethylstilbestrol was also used, and produced the
same results.
Oeuri.
Was estradiol 17(3 chosen because it is a fish
estrogen?
Aida.
That is correct.
p122
•

• 245
•
V. The a -Opraisal of egg quality.
Chairman: Hiroshi TSUKAHARA. (Kyushu University, Agriculture).
For Mr. Sakai.
Yamamoto.
How were observations made immediately after
ovulation in rainbow trout? How many specimens were used?
Sakai. (Tokyo University of Fisheries).
Individuals from which eggs could not be collected
on the previous day, but from which they could be collected
the next day were taken to have just ovulated. The number
of specimens was 30.
•

• 246
General Discussion.
p123
Chairmen: Takashi HIBIYA (Tokyo University, Agriculture).
' Minoru NOMURA (Tokyo University of Fisheries).
Reijiro HIRANO (Tokyo University, Agriculture).
Hibiya.
Let us start the discussion with the question of
maturation and external environmental factors.
Oguri.
111, would like to enquire about pathways for the
influence of light in stimulating development of .the gonads.
Yoshiaka.
Up to now it has been supposed that the stimulation
of gonad maturation by light was effected through the
diencephalon pituitary, but there has been no experimental
search throughout the body for pathways for this stimulation.
Medaka mature in long days, but if the pituitary is extirpated
the ovary degenerates. This shows that the pituitary has an
important role in maturation under long daylight. Comparison
of medaka reared under short and long days shows that those
reared under long days contain more aldehyde-fuchsin staining
neurosecretions and more cells containing these secretions in
the diencephalon hypothalamus. This shows that the diencephalon
is concerned in stimulation by light.

24 7
However when the eyeballs are removed from the medaka,
the optic nerves cut, and the pineal body removed at the same
time, the frequency of spawning is reduced, but it is recovered
after 5 weeks. This shows that there is a direct path for
the action of light on the hypothalamus. Since the pigment
in the scales absorbs light whose wavelength is less than
400m/..., it is to be supposed that the light which is effective
has a wavelength longer than 400mp-, It has been experimentally
shown that medaka with the upper part of the eyes removed and
reared in light of wavelength less than 400m/.., do not mature,
but that in longer wavelength light they do mature. Thus there
must be a direct pathway for light to act on th-: hypothalamus
other than that from the eyes to the nerves and thence to the
hypothalamus and the pineal body. However since there is
little pigment which will absorb light-in.:the vicinity of the
hypothalamus, further investigation of the mode of direct
action of light on the hypothalamus is required.
Tamura.
It must be assumed that there is a path for light
stimulation which does not involve the eyes, and this will
probably be a path through the pineal body. The receptor
for the day length is the pineal body, in which melatonin
is produced during dark periods. The melatonin acts on the
pituitary and inhibits the secretion of gonadotrophic hormone.

248
During periods of light, less of this inhibiting substance
is uroduced, and consecuently it is to be supposed that the
secretion of gonadotrophic hormone by the pituitary is
increased. It is also said that melatonin acts directly on
the gonads of the higher animals and inhibits their
development. I feel that this is a field in which there
should be further experimental studies of fish. The fish
pineal emits pulses durilw dark periods which are suppressed
and not emitted during light periods. For this reason
removal of the pineal body is, so far as concerns the pulses,
the same as being in an illuminated place. I think that the
pineal body should be experimentally removed with this point
in mind. The absorption of the pigments suggests that the
wavelength of the most effective light should be 520 to 530y-
for the pineal body and 610mr for the eye. This does not
disagree with the effectiveness of light of wavelength longer
than Lkoome, which has been described by Mr. Yoshioka. However
the shorter wavelengths which are not wholly ineffective must
be included, and the intensity of the light must also be
remembered.
Yamagishi. (Teikyo University).
In natural conditions, fish live in an environment
with seasonally changing temperatures. What is the effect
on spawninn- of rearing them from the fry stage at a fixed
temperature?
p124

• 249
•
•
Yamamoto.
Many fish will form yolk even when reared at temperatures
lower than that required for spawning, but the rate of
vitellogenesis is slow. A definite temperature is needed
for ovulation and spawning. Nevertheless vitelloeenesis often
occurs at temperatures at which spawning is not possible.
Yamagishi.
There is a connection between this and the problem of
rearing parent fish for egg collection. If the fish are
reared at the appropriate temperature for growth and the
temperature is also changed only at the maturation and
spawning season, what is the effect on the maturation and
spawning of these fiSh?
Nomura. (Tokyo University of Fisheries).
In order to obtain good eggs with a development
percentage of not less than 80% from rainbow trout the parent
fish should be raised in temperatures not above 12 °C to 13 °C
and not below 3 °C to 4° C. For maturation and spawning the
needed temperature changes from 13°C to about 18 °C. With ayu
a temperature of 20 °C or higher is good for growth, but for
spawning the temperature is 17 °C to 18 ° C or lower. Mr. Sakai
observed experimentally that oogenesis occurred above 20 0 0,
but that the eggs became abnormal and remained unovulated.
In the culture of rainbow trout, they are kept at a high

• 250
ve
•
in order to increase the number of egg collections, to
obtain eggs from two-year-old fish, and to promote rapid
large growth, and the fish are made to spawn about 2 months
early by being moved into water at the spawning temperature.
In the Kumagaya trout culture experimental ponds of the
Saitama fisheries experimental station, the temperature one
month before the active spawning season is 14.6° C or lower,
and the temperature is then lowered about 1 0 0 every 10 days,
with good results in egg collection .
Hirano. (Tokyo University, Agriculture).
At the Owase station porgy are reared with the water
kept at the high temperature of 20 ° C, and eggs are successfully
obtained in mid-February, 1.5 months earlier than in natural
conditions. Would it be possible to cause them to spawn even
earlier with still higher temperatures? I would like to hear
Mr. Harada's opinion.
Harada.
If porgy spawn 1.5 to 2 months earlier when cultured
than in natural conditions, the fry can be successfully
reared. For early egg collection, porgy are reared at the
spawning temperature or at a slightly lower temperature, and
when the eggs are ripe the temperature is raised by 200 to 3 ° C
* Sic, though it would appear from earlier in this statement that the temperature should be raised. Translator. •

• 251
from just below the spawninP: temperature to just above it.
Ovulation is then efficient. The fish will grow well in
temperatures higher than the spawning temperature, but I do
not think that they will be caused to spawn.
Fushimi. (Hiroshima University Fisheries Research).
The spawning season for cultured red sea bream (madai)
Chrysophrys major in the Inland Sea is from May to July, but
in order to obtain eggs early the fish were reared from the
end of November and through the winter in water in which the
lowest temperature reached was 8 00, 13 00 or 15 0 0. Those for
which the lowest temperature was 15 00 produced mature eggs,
but up to 10 May they had not spawned, and eggs were not
obtained early. Those for which the lowest temperature was
• 13 ° C spawned on 14 April. •
In natural conditions of low winter temperatures they
do not feed and the body weight drops by about 15%, but when
the temperature is increased they start to feed and the
weight increases. The nutritional condition of parent fish
reared at the higher temperatures' remained satisfactory and
it was found experimentally that the amount of spawn and the
proportion hatched were both good.
Hirano.
When porgy are reared in warm water, do you think
that spawning is influenced by the len7th of time before
. spawning at which they are shifted?

• 252
•
Harada.
If there are no large changes in environmental factors
other than temperature, the change of temperature when
shifting will have some effect, but if the change of
temperature is small, shifting the fish will not cause much
of a problem. If the living space after shifting is not
restricted, they will spawn if shifted two months earlier.
I believe that rapid changes of temperature before or after
shifting can cause problems.
Hirano.
I think there are some temperature changes, but at
the time when we move the cultured parent fish in our
experiments into the spawning pond, most of the parent fish
which are treated with hormone and given sufficient food will
spawn. If more than one week elapses between the moving and
the hormone treatment, the ovarian eggs all degenerate and
there is no spawn.
Hibiya.
In general the influence of stress is greater in
marine fish than in freshwater fish. Handling of the fish
may cause polyuria, and there should be a study of the
dehydrated condition. I suppose that this question is
related to the physiology of the interrenal glands, but I
p125
II› would like to hear Mr. Oguri's opinion.

• 2 53
•
•
Oguri.
1 think this is related to the secretion by the
interrenal glands of cortical hormones in order to regulate
the osmotic pressure. For examule, if the interrenal glands
are removed from the eel it becomes unable to live in sea
water, but when cortisol is injected it is found experimentally
to be able to live in sea water. However since the eel is
the only fish from which the interrenal glands can be removed,
and since the method of removal is difficult, there are very
few experiments available, and further inveutigation of methods
of removal is needed. The interrenal glands may thus be
essential to the life of the fish. The data connecting the
interrenal glands with the problems of dehydration are at
present in an unsatisfactory condition.
Yamazaki.
Cortical hormones are secreted in large amounts in
response to stress. According to Fagrlund's experiments with
sockeye salmon the females are more sensitive to stress than
the males. For example, when put into a scoop net in a tank
for 30 minutes, the cortisol in the blood increased five-fold
in the males but ten-fold in the females. Also when chinook
salmon were caught in the wild and reared for one to ten days
in a tank, the cortisol increased by 50 times, and they then
died. Furthermore, when cortisol was injected in Robertson's
experiments, the ovary and other organs deg,enerated. This

254
shows that stress is extremely deleterious to reproduction.
Nevertheless a great deal of cortisol is secreted by salmon
and trout while goinp: up river, but there are few actual
measurements. It appears from the experimental results of
Donald et al that the injection of sex steroids causes a
large discharge of cortisol from the body. Thus, when hormone
is used to cause ovulation in fish which are greatly
influenced by streSs, and sex hormones are administered at
the same time, it is possible that cortisol may bé rapidly
discharged.
Nomura.
In Mr. Kato's paper it was stated that the largest
number of spawnings of rainbow trout were in March to May,
and that the actual number of spawnings was mot closely
correlated with the weight of the parent fish in August. In
other words, eggs which had not yet grown in August were
among those which appeared in March to May. Since it is
important, in the production of fry, to obtain a large number
of eggs spawned from each parent fish, I would like to ask
how this fact can be explained on the 'basis of the
fundamental endocrinology, of the nutritional environment or
of the metabolic process, so as to extract the maximum
possible amount of spawn.
•

• 254a
•
Yama s ak i.
It is the feeding of the body that determines the number of eggs
laid. When the parent fish is poorly fed, few eggs mature and are
released. When it is not fed, regressive eggs grow in number and few
eggs are laid. When the fish is not healthy, the hormone treatment
is ineffective.
Yamamoto
The relationship between growth and multiplication may be
considered as follows. The total amount of energy in the fish is given. If
much energy goes to reproduction it does not go to growth. The opposite
holds too. Generally among cold blood organisms, at a certain time energy
is stored within the body for reproduction and at a certain time it is
used for reproduction. At another time, it is used for growth. I think
that this tendency is the general rule and that fish are no exception.
As we treat scientifically in a concrete fashion of the relationship
between growth and multiplication, I would like to hear the opinion
of a specialist in metabolism.

255 ••
Hibiva.
I would like tc, kncw the opinion rf Mr. Takeuchi.
Takeuchi. (Tokai regional Fisheries Research Laboratory).
As you have suggested, the components of the diet are
applied towards growth and reproduction, and the proportions
allotted to the two functions vary greatly from one stage to
another. The number and quantity of spawnings are important,
and one must also consider the satisfactoriness of the spawn
and the way in which the components of the diet are assembled
into the eggs. It has been found in tuna that some of the
materials taken in during maturation are accumulated in
quantity in the eggs, and some are not. For example, in ayu,
when the fat level in the feed is increased (to the extent of
3% to 20%), the fat content of the body increases, but the fat
content of the ovary does not change. If a large amount of
vitamin E is supplied, much is accumulated in the ovary, but
if only a small amount is supplied there is no spawning even
though the GSI does not change. These migrations of the diet
vary greatly, but may perhaps supply some hint on how to
obtain good eggs.
Hirano.
When one is concerned with the actual production of
fry, egg quality is the most important question. Mr. Kiyono
made his experiments on the assumption that good quality eggs •

• 256
were those with a high larval survivability rate, but I
suggest that egg Quality depends on the process of
maturation and the method of collecting the eggs. What,
from the fundamental point of view, is Mr. Yamamoto's
opinion about this?
Yamamoto.
I originally supported Mr. Sakai's definition of the
satisfactoriness of egg quality in terms of high fertilization
and hatching percentages of the eggs. However I think that
apart from the reduction of fertilization percentage with
lapse of time after ovulation, the essential causes of
reduction of the fertilization and hatching qualities of the
eggs are changes in the metabolism of the parent fish during
the process of oogenesis. For example, when the eggs hatch,
the important factors which influence the metabolism of the
embryo are not only the accumulation of nutrients such as
yolk and fats, but also the substances such as vitamin E
which are present in very small amounts. It is important
that this should receive further study.
Hirose.
I have found that the water content and the ions
which enter the eggs at the time of ovulation are important
to egg quality.

• 257
•
Ogino. (Tokyo University of Fisheries).
The phosphorus content of‘rainbow trout eggs is un-
usually large, and the calcium content unusually low. The
Phosphorus is present in the egg as phospholipids and
phosphoproteins. The phosphorus in the egg cornes from
materials in the mother's body, and probably is derived from
the bones. The relative proportions of phosphorus and
calcium in the mother's body will be very different during
maturation from those at other times. Consequently, simple
reasoning suggests that the supply of phosphates to the
parent fish during maturation of the eggs will be important.
Yamazaki.
It is believed that the yolk c'ontains substances
which promote the development of the embryo. It is not known
when these substances are accumulated, but is it not true
that if they are not properly accumulated the eggs will not
be of good quality even if sufficient yolk has been accumulated?
Hibiya.
There are differences between different species of
fish. For example, even among the same family such as the p127
Salmonidae, the salmon.die after spawning once, but the trout
spawn repeatedly.To add to Mr. Ogino's remarks about phosphorus,
if a fundamental investigation were made of the differences

258
in the state of these species after ovulation, it might be
possible to get some ideas for a technique for their
regeneration. I would like to as Mr. Suyama for his views
on this.
Suyama (Tokyo University of Fisheries).
The composition of the phosphoproteins in fish differs
from that in domestic fowl. For example phosvitin, a protein
containing a large amount of phosphorus, can be isolated from
the fowl egg, but 10 times as much protein of the phosvitin
type is found in rainbow trout eggs. This phosphorus is
present as laro;e quantities of serine esters and small
quantities of threonine esters, but serine is not classed as
a very important amino-acid in fish. If these are decomposed
during development, the phosphorus obtained may be expected
to have an important metabolic action in the formation of
high energy types of ATP and of creatine-phosphoric acid.
For this reason I believe that the phosphoproteins will be
sources of interest in future research.
Murakami.
Supposing that egg quality is so good that the
fertilization and hatching percentages are 100%, is it possible
that 100% fry could be obtained by a perfected technology of
rearing,.? I would like to ask Mr. Hibiya and Dir. Yamamoto
about this.

•
259
Hibiva, Yamamoto. .
Given your conditions, it is possible.
Hibiya.
The information obtained by basic research is detailed,
that obtained by experience is generalized. The two don't
mesh together very well, and this distresses those of us who
participate technically or scientifically in production.
Today we may feel some relief from our distress. Much remains
for the future, such as the most suitable regulation of the
environmental conditions, the development of standard feed
for the parent fish, and the application of genetics to the
breeding of superior parent fish of both sexes. I feel that
this symposium has been successful in fostering debate and
bringing out concrete proposals for future research, and I
may perhaps say that it was been more successful even than
was hoped. All those who were charged with the responsibility
of planning are happy that it was possible to debate so many
important questions during this successful meeting, and I
wish to express our cordial thanks to all the participants
for their cooperation.

Shuzo EGUSA Minoru OKADA Shunji KAWABATA hitoshi KONDO Shoichi TANAKA
Junsaku NONAKA Yoshiro HASHIMOTO Takashi HIBIYA Yoshihiro MACHIDA Motonobu YOKOZEKI
Publishing Committee.
260
Fisheries Science Series (6). 0362-150060-2244.
The Maturation and Spawning of Fish.
Fundamentals and Application.
Price 1100 Yen ,
Published 15 October 1974.
Editors: The Japanese Society of Scientific Fisheries (Incorporated).
c/o Tokyo University of Fisheries Minato minami 4 - 5 - 7 Minato ku Tokyo Japan 108.
Place of publication: Koseisha Koseikaku Ltd.
Saneicho Shinjuku ku .Tokyo.
Gyro .Tokyo 59600
Telephone (359) 7371 - 5.
Copyright: Japanese Society of Scientific *Fisheries, 1974.
Printed by Daishin. Bound by Nakajo.

261
Food Science for Pisciculture.
(Yogyo shiryo gaku)
Edited by Yoshiro HASHIHOTO.
A5, 282 pages, 1600 Yen.
The latest developments in nutritional science are
freely used in a thorough discussion of the theory and
technique of nutrition in pisciculture. In these times of
rapid rises in the price of food in the pisciculture industry,
it is a unique source of reference on the compounding und
distribution of food and on nutritional diseases. It has won.
favorable reports from researchers, from students, and from
those engaged in the culture of fish.
An introduction to the Science of Pisciculture.
(Yogyo gaku so ron).
Edited by p.21.22InIll_xAmL212.
A5, 720 pages, Lp000 Yen.
Concentrating of the fundamentals of pisciculture,
this is a standard reference for the morphology, physiology,
endocrinology, diseases, culture ponds, rearing and
nutritional methods for the cultured species of fish. At
the outset of the ae of cultured fisheries, this book of
detailed explanations by leading researchers has received
favorable criticism.
p130

a 262
Complete Pisciculture.
(Yogyo gaku kakuron).
Edited by Noboyuki KAWAMOTO.
A5, 840 pages, 5000 Yen.
This large volume gives detailed explanations of
all important matters in methods of rearing, the nutrition,
the prevention of disease, the shipping and care of fry of
all the principal species which are cultured in Jauan,
from carp, goldfish, eel, chum salmon, salmon trout, rainbow
trout,yellow tail and porgy to oysters, abalone, laver,
pearls and prawns.
"_Thorough Culture" in shallow seaR.
(Senkai kanzen yoshoku).
Joint Editors:
Takeo IMAI Shun INO Munehisa KUROKI Moritaro YAMAMOTO. Motosaku FUJINAGA
B5, 464 pages, 3500 Yen.
The completely artificial culture of all stages from
the production of fry to the rearing of adults is called
"Thorough Culture". This book discusses in detail biology and
culture techniques of Thorough Culture in Japan of laver,
oysters, abalone, prawns and scallops.
Forthcoming! Diseases of cultured fish. by Shuzo MUSA.
KOSEISHA KOSEIKAKU.




















