
Ultrasound promoted efficient and green protocol for the expeditious
Upload
independentCategory
view
3download
0
The Journal of Immunology
Marginal Zone Precursor B Cells as Cellular Agents forType I IFN–Promoted Antigen Transport in Autoimmunity
John H. Wang,* Jun Li,* Qi Wu,* PingAr Yang,* Rahul D. Pawar,* Shutao Xie,*
Laura Timares,† Chander Raman,* David D. Chaplin,‡ Lu Lu,x John D. Mountz,*,{ and
Hui-Chen Hsu*
The pathogenic connection of type I IFN and its role in regulating the migration response of Ag delivery by B cells into lymphoid
follicles in an autoimmune condition has not been well-identified. Here, we show that there was a significantly larger population of
marginal zone precursor (MZ-P) B cells, defined as being IgMhiCD1dhiCD21hiCD23hi in the spleens of autoimmune BXD2 mice
compared with B6 mice. MZ-P B cells were highly proliferative compared with marginal zone (MZ) and follicular (FO) B cells.
The intrafollicular accumulation of MZ-P B cells in proximity to germinal centers (GCs) in BXD2 mice facilitated rapid Ag
delivery to the GC area, whereas Ag-carrying MZ B cells, residing predominantly in the periphery, had a lower ability to carry Ag
into the GCs. IFN-a, generated by plasmacytoid dendritic cells, induced the expression of CD69 and suppressed the sphingosine-1-
phosphate-induced chemotactic response, promoting FO-oriented Ag transport by MZ-P B cells. Knockout of type I IFN receptor
in BXD2 (BXD2-Ifnar2/2) mice substantially diffused the intrafollicular MZ-P B cell conglomeration and shifted their location to
the FO-MZ border near the marginal sinus, making Ag delivery to the FO interior less efficient. The development of spontaneous
GCs was decreased in BXD2-Ifnar2/2 mice. Together, our results suggest that the MZ-P B cells are major Ag-delivery B cells and
that the FO entry of these B cells is highly regulated by type I IFN–producing plasmacytoid dendritic cells in the marginal sinus in
the spleens of autoimmune BXD2 mice. The Journal of Immunology, 2010, 184: 442–451.
The localization ofBcells in appropriatemicroenvironmentsof secondary lymphoid organs plays an important role indetermining the fate of B cell immunity. Specifically, the B
cellmigrationpattern into thefollicular (FO)regionsof thespleencanalter the tempo and frequency ofT–Bcell contact and, thus, has beenproposed as one mechanism to regulate the development of autor-eactive B cells (1, 2). Early studies by Cyster et al. (3) showed thatunder conditions of clonal selection, anergic B cells failed to enterthe FO areas, instead arresting at the T–B cell boundary in a processknown as FO exclusion. FO exclusion has been proposed as one Bcell checkpoint mechanism to prevent B cell tolerance loss (4).Although chemokines and chemokine receptors have been para-mount in shaping B cell migration patterns, we and other inves-
tigators (5–7) showed that local cytokine production could provideadditional signals to regulate the migration and retention of B cellsin the FOs. In autoimmune murine models, B cells can act as APCsand/or Ag-delivery cells to promote the development of spontane-ous autoreactive germinal centers (GCs) (8–12). Treatment of au-toimmune disease with B cell–depletion therapy depletes B cellsthat mainly serve as APCs or Ag-delivery cells (9, 13–15).The BXD2 mouse strain is one ofz20 BXD recombinant inbred
strains that we analyzed for the development of autoimmune dis-ease, including the development of features resembling humansystemic lupus erythematosus and rheumatoid arthritis (16). Thedevelopment of highly mutated multireactive autoantibodies thatcan induce kidney and joint disease is one of the important patho-genic features of BXD2 mice (16, 17). We found that BXD2 micespontaneously formGCs in the spleens and that the expression of thegene encoding activation-induced cytidine deaminase (Aicda) in theGC B cells can be stimulated by activated CD4+ T cells from BXD2mice (18). There is an abundance of Th-17 cells in the FOs and GCarea. The high levels of IL-17 produced by these cells is associatedwith the upregulation of regulator of G-protein signaling (Rgs)13and Rgs16 and the migration arrest of B cells in response to FO- andGC-oriented chemokines, CXCL12 and CXCL13, enabling them toform stable T–B cell conjugates and facilitate GC formation in thespleens of BXD2 mice (6).The importance of IFN-a in the spontaneous development of
autoantibodies and immune complex deposition in the glomeruliwas demonstratedusingmurinemodels of lupus (19–21).Clues to thepossible role of IFN-a in the production of high-affinity autoanti-bodies can be derived from their normal physiologic mechanisms ofaction. IFN-awas shown to promote Ab class-switch recombinationandplasmacell differentiationunder nonautoimmuneconditions (22,23).Todetermine the reason for the coexistence of high levels of IFN-a and IL-17 in the spleens of BXD2 mice, we observed that therewas a significantly increased IgMhiCD1dhiCD21hiCD23hi B cell
*Department of Medicine, †Department of Dermatology, and ‡Department of Micro-biology, University of Alabama at Birmingham, Birmingham, AL 35294; xDepart-ment of Anatomy and Neurobiology, University of Tennessee Health Science Center,Memphis, TN 38163; and {Birmingham VA Medical Center, Birmingham, AL 35233
Received for publication March 17, 2009. Accepted for publication October 21,2009.
This work was supported by a grant from the American College of RheumatologyResearch and Education Foundation Within Our Reach: Finding a Cure for Rheu-matoid Arthritis campaign, the Alliance for Lupus Research – Target Identification inLupus program, a Department of Veterans Affairs Merit Review Grant(1I01BX000600-01), Daiichi-Sankyo, and National Institutes of Health Grants(1AI 071110-01A1 and ARRA 3RO1AI71110-02S1) (all to J.D.M.), The Universityof Alabama at Birmingham Skin Diseases Research Center (to L.T.), and the ArthritisInvestigator Award supported by the Arthritis Foundation (to H.-C.H).
Address correspondence and reprint requests to Dr. Hui-Chen Hsu, Department ofMedicine, The University of Alabama at Birmingham, 1825 University Boulevard,SHELB 311, Birmingham, AL 35294. E-mail address: [email protected]
The online version of this article contains supplemental material.
Abbreviations used in this paper: DOTAP, N-[1-(2,3-Dioleoyloxy)propyl]-N,N,N tri-methylammonium methylsulfate; FO, follicle or follicular; GC, germinal center; MS,marginal sinus; MZ, marginal zone; MZ-P, marginal zone precursor; pDC, plasma-cytoid dendritic cell; PNA, peanut agglutinin; QRT-PCR, quantitative real-time PCR;S1P, sphingosine-1-phosphate; TNP, (2,4,6-trinitrophenyl)-protein; UAB, Universityof Alabama at Birmingham; WT, wild-type.
www.jimmunol.org/cgi/doi/10.4049/jimmunol.0900870

population in the spleens of BXD2mice. B cells with this phenotypeare referred to asmarginal zone precursor (MZ-P)B cells inmice (24,25). We further identified that the entry of MZ-P B cells into the FOregion is promoted by IFN-a, which is primarily produced by plas-macytoid dendritic cells (pDCs) that are localized in the marginalsinus (MS). Abrogation of IFN-aR in BXD2 mice attenuated thedevelopment of spontaneous GCs and autoimmune disease. Im-portantly, injection of BXD2 and BXD2-Ifnar2/2 mice with 2,4,6-trinitrophenyl-protein (TNP)-Ficoll demonstrated thatMZ-PB cellsserve as the major Ag-transporting B cells that directly deliver TNPinto the GCs. Deficiency of IFN-a abrogated this response. Ourresults suggest that FO entry ofAg-deliveryMZ-PB cells facilitatedby IFN-a may be an important mechanism to promote the autoim-mune response in BXD2 mice.
Materials and MethodsMice
Female homozygous C57BL/6 and BXD2 recombinant inbred mice wereobtained fromThe JacksonLaboratory (BarHarbor,ME); B6-Ifnar2/2micewere obtained from Dr. Jocelyn Demengeot (Instituto Gulbenkian deCiencia, Oeiras, Portugal). B6-Ifnar2/2- micewere backcrossedwith BXD2mice for seven generations using amarker-assisted speed congenic approachwith 146 markers. All mice were housed in the University of Alabama atBirmingham (UAB)Mouse Facility under specific pathogen-free conditionsin a room equipped with an air-filtering system. The cages, bedding, water,and food were sterilized. All mouse procedures were approved by the UABInstitutional Animal Care and Use Committee.
Flow cytometry analysis and cell sorting
Flow cytometry was performed on fluorescently labeled single-cell sus-pensions derived from spleens using the method we described previously (6,18). For the detection of various cell types, including pDCs and B cells,fluorescent-conjugated anti-PDCA1 (eBioscience, San Diego, CA), anti-CD19 (Biolegend, San Diego, CA), anti-IgM (Southern BiotechnologyAssociates, Birmingham, AL), anti-CD21/35 (Biolegend), anti-CD23 (Bi-olegend), and anti-CD93/AA4 Abs were used. For the detection of CD69,CD80, and CD86, fluorescent-conjugated anti-CD69 (Biolegend), anti-CD80 (Biolegend), and anti-CD86 (Biolegend) Abs were used. For thedetection of TNP, biotinylated anti-TNP (BD Biosciences, San Jose, CA)was used followed by fluorochrome-conjugated streptavidin. For the la-beling and isolation of pDCs, all spleens were cut and treated with colla-genase D (2 mg/ml) for 30 min at 37˚C/5%CO2 before obtaining single-cellsuspensions for Ab labeling. For the intracellular detection of IFN-a, cellswere permeabilized (Cytofix/cytoperm, BD Biosciences), washed (Perm/Wash, BD Biosciences), and then incubated with a 1:1 mixture of anti-IFN–a Abs (clones RMMA-1 and [18F]). Cells (100,000/sample) werewashed twice with FACS buffer and fixed in 1% paraformaldehyde/FACSsolution before analysis by flow cytometry using a BD LSR II flow cy-tometer (BD Biosciences). The analysis was performed using FlowJoSoftware (Tree Star, Ashland, OR). Forward-angle light scatter was used toexclude dead and aggregated cells. Some results are presented as fluores-cence histogramswith the relative number of cells on a linear scale versus therelative fluorescence intensity on a log scale. Acquisition and gating of FO,marginal zone (MZ), andMZ-PB cells were carried out based on themethoddescribed by Allman and Pillai (24). FO B cells were defined as CD23hi
sIgMlo+hisIgDhiCD21intCD1dloCD19+ cells; MZ B cells were defined asCD23null+losIgMhisIgDloCD21hiCD1dhiCD19+ cells; MZ-P B cells weredefined as CD23hisIgMhisIgDhiCD21hiCD1dhiCD19+ cells. All gatings wereset up to ensure that the percentage of each subpopulation of B cells obtainedfrom normal B6 mice was equivalent to the previously reported results (24).
For cell sorting to isolate the different B cell populations, anti-CD19microbeads (Miltenyi Biotec, Auburn, CA) were used to isolate whole B cells.Subsequently, the single-splenic B cell preparation was labeled with fluorescentconjugatedAbsandsortedintoFO,MZ,andMZ-PBcellsbasedontheexpressionofCD21,CD23, and IgM,asdescribedabove.SortingwascarriedoutusingaBDFACSAriaCellSorter (BDBiosciences).All sorting for pDCswascompletedbypositive selection using anti-PDCA1 microbeads (Miltenyi Biotec).
Immunofluorescent staining and confocal image analysis
Spleens from mice were collected, embedded in Frozen Tissue Media(Fisher Scientific, Pittsburgh, PA), and snap-frozen in 2-methylbutane.Frozen sections (8 mm thick) were dried in air for 30 min and fixed inacetone for 15 min prior to rehydration with 1% BSA for 10 min. The
sections were blocked with 10% horse serum for 30 min at room tem-perature and then stained for 30 min at room temperature with various Abs.The following primary Abs conjugated to the Alexa Fluor dyes were usedto stain spleen tissues for confocal image analysis: biotin-peanut agglutinin(PNA) (Vector Laboratories, Burlingame, CA) to Alexa 350–streptavidin(Invitrogen, Carlsbad, CA); PNA to Alexa 350; anti-mouse CD35 (clone8C12, BD Bioscience) to Alexa 488; anti-IgM to Alexa 555; anti-CD4(RM4-5, Invitrogen) to Alexa 488 or 647, anti-CD23 (B3B4) to Alex488,and anti-PDCA1 to Alexa 647. For the detection of CD1d Ag, tissue wasincubated with anti-CD1d rat IgG (Biolegend) followed by anti-rat IgGconjugated to Alexa 555 (Invitrogen), after which tissuewas incubatedwith10% normal rat serum at room temperature for 30min. After 10% normal ratserum treatment, the tissueswere stainedwith the otherAbs. For detection ofTNP, anti-TNP hamster IgG (BD Sciences) incubation was followed by bi-otinylated anti-hamster IgG (Biolegend) incubation and fluorochrome con-jugated–streptavidin treatment. The sectionsweremounted in Fluormount G(Southern Biotechnology Associates) and viewed with a Leica DM IRBEinverted Nomarski/epifluorescence microscope outfitted with LeicaTCS NT(Leica Microsystems, Bannockburn, IL) laser confocal optics. In situquantitation of the PNA+ GC response and TNP+ cells was carried out usingOlympus DP2-BSW software (Olympus America, Center Valley, CA) ac-cording to the method we described previously (6).
Cell cultures
Cells, including pDCs and B cell subpopulations, were cultured with RPMI1640 medium (Invitrogen) supplemented with 2 mM L-glutamine, 25 mMHEPES, 100 U/ml penicillin, 100 mg/ml streptomycin, 5.5 3 1025 M2-mercaptoethanol, and 10% FCS for all in vitro experiments.
IFN-a stimulations and measurements
ForCD69 expression on different B cell subpopulations in response to IFN-aunder in vitro conditions, single spleen cells were cultured in 96-well(Costar, Cambridge, MA) tissue-culture plates at 37˚C/5% CO2 in triplicatewells (13 104 cells per well) and stimulated with IFN-a (20 ng/ml) for 12 h.To measure IFN-a levels produced by pDCs, single-cell suspensions wereprepared by collagenase D (2 mg/ml) digestion of spleen tissues for 30 minat 37˚C/5%CO2 before sorting for pDCs. Enriched pDCs were sub-sequently cultured in 96-well (Costar) tissue-culture plates at 37˚C/5%CO2 in triplicate wells (1 3 104 cells per well) and stimulated withmedium only, CpG (3 mM), or CpG (3 mM)-N–[1-(2,3-Dioleoyloxy)propyl]-N,N,N trimethylammonium methylsulfate (DOTAP) complex(F. Hoffman-La Roche, Boulder, CO). After 24 h, culture supernatantswere collected, and IFN-a levels were measured by ELISA (PBLBiomedical, Piscataway, NJ). CpG-DOTAP preparation for in vitroculture was formulated by mixing 19.2 mg (3 nmol) CpG in 120 mlPBS with 30 ml DOTAP, incubating for 15 min, and then diluting with850 ml medium to make 3 mM CpG-DOTAP solution. For in vivomeasurements of IFN-a, 5 mg CpG was administered i.v. into eachmouse tail vein. Five micrograms of CpG was mixed with 50 ml PBS,whereas in another reaction tube, 30 ml DOTAP was mixed with 70 mlPBS. The CpG solution was then mixed with the DOTAP solution andincubated at room temperature for 15 min prior to administration.Serum levels of IFN-a in each mouse were measured 4 h later.
Cell migration assay
Single-cell suspensions of anti-CD19 MACS column (Miltenyi Biotec)-pu-rifiedspleenBcells fromB6,BXD2,orBXD2-Ifnar2/2micewerestimulatedwith medium only control or mouse recombinant IFN-a (20 ng/ml, R&DSystems,Minneapolis,MN) for 6 h. The cell-migration assaywas carried outusing the protocol described by Moratz and Kehrl (26). The stimulated cellswere loaded (2 3 106) into the upper well insert (5-mm pore size) ofa Transwell system (5 mm pore size, Costar), and sphingosine-1-phosphate(S1P) was added to the bottom chamber at a final concentration of 20 nM.After incubation for 2 h at 37˚C in a 5% CO2 incubator, the migrated cellswere harvested and resuspended in 300 ml FACS buffer. The cells that re-mained in the inserts or migrated to the lower chamber were counted usinga flow cytometer, and the separation of the CD19+ B cells into migrated andnonmigrated cell subsets was determined. The chemotaxis index was cal-culated by dividing the number of cells that migrated in response to che-mokine by the number of cells that migrated in the absence of chemokine.
Isolation and quality control of RNA
RNAwas isolatedfrom0.2 to103106cellsusingTRIzol reagent (Invitrogen)orfrom ,0.2 3 106 cells using the PicoPure RNA Isolation Kit (Molecular De-vices, Sunnyvale,CA).Thequalityof the isolatedRNAwasdeterminedusinganExperion Automated Electrophoresis System (Bio-Rad, Hercules, CA). RNAs
The Journal of Immunology 443
that exhibited an 18S/28S ratio .1.8 were converted to cDNA using the FirstStrand cDNA Synthesis Kit (Fermentas, Glen Burnie, MD).
Quantitative real time-PCR
The expression of Aicda, S1p1, and S1p3 was analyzed using a quantitativereal-time PCR (QRT-PCR) method. Briefly, the QRT-PCR mixtures con-tained SYBRGreen PCRMasterMix (Bio-Rad) with the following primers:Aicda forward: GCCACCTTCGCAACAAGTCT; Aicda reverse:CCGGGCACAGTCATAGCAC (137 bp); S1p1 forward: TGCTGTAACT-GAAGGCTCAC; S1p1 reverse: GGATGCTAGTGGACACCATAG (108bp); S1p3 forward: CCGTAGTGATTGTGGTGAGTG; S1p3 reverse:GGACAGCCAGCATGATGAAC; Cdca3 forward: GAGTAGCAGACCC-TCGTTCAC; Cdca3 reverse: TCTCTACCTGAATAGGAGTGCG; Cdca5forward: GGACTTCACTTACTAAGCCTTC; Cdca5 reverse: GA-CATCTGGGACCTCTACTG; Cdca8 forward: ACAAGGAAGAGGCA-GAAG; Cdca8 reverse: CCGTTGATGGAGATGTTG; Gapdh forward:AGGTCGGTGTGAACGGATTTG; Gapdh reverse: TGTAGACCATGTA-GTTGAGGTCA (136 bp). A final volume of 25ml was used forQRT-PCR inan IQ5 thermocycler (Bio-Rad).Amplification conditionswere 95˚C for 3min,followed by 40 cycles of 95˚C for 15 s and annealing for 30 s. The annealingtemperature used was 54˚C for Cdca8; 56˚C for Gapdh; 57˚C for S1p3; 58˚Cfor Aicda, Cdca3, and Cdca5; and 60˚C for S1p1 and S1p3.
QRT-PCR products were normalized relative to that of Gapdh to correctfor potential differences in template input to determine relative mRNAlevels. Results are expressed as fold differences in the expression of theindicated gene relative to the expression of Gapdh. Standard curves weregenerated for every target using six 4-fold serial dilutions.
TNP-Ficoll
To specify cellular agents for Ag transport, 50 mg TNP-Ficoll (BiosearchTechnologies, Novato, CA) was administered i.v. into each mouse, and
mice were sacrificed at the indicated time points. A portion of the spleenwas snap-frozen in 2-methylbutane for histological analysis, as describedabove. Another portion of the spleen was made into a single-cell suspen-sion and labeled for detection of TNP, as described above.
Statistical analysis
All results are shown asmean6 SEM.A two-tailed t test was usedwhen twogroups were compared for statistical differences. The ANOVA test was usedwhen more than two groups were compared for statistical differences. Thedistribution differences of MZ and MZ-P in the anatomical MZ area versusthose in the anatomical follicular area between different strains of miceweretested using the x2 test. p values ,0.05 were considered significant.
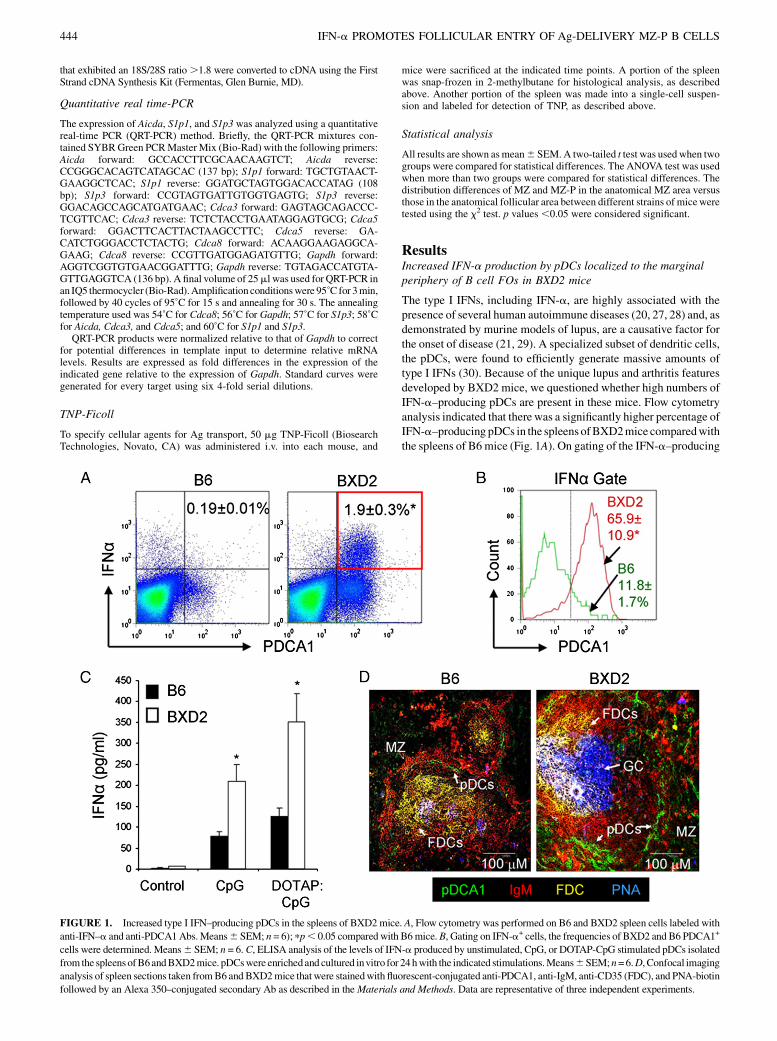
ResultsIncreased IFN-a production by pDCs localized to the marginalperiphery of B cell FOs in BXD2 mice
The type I IFNs, including IFN-a, are highly associated with thepresence of several human autoimmune diseases (20, 27, 28) and, asdemonstrated by murine models of lupus, are a causative factor forthe onset of disease (21, 29). A specialized subset of dendritic cells,the pDCs, were found to efficiently generate massive amounts oftype I IFNs (30). Because of the unique lupus and arthritis featuresdeveloped by BXD2 mice, we questioned whether high numbers ofIFN-a–producing pDCs are present in these mice. Flow cytometryanalysis indicated that there was a significantly higher percentage ofIFN-a–producing pDCs in the spleens ofBXD2mice comparedwiththe spleens of B6mice (Fig. 1A). On gating of the IFN-a–producing
FIGURE 1. Increased type I IFN–producing pDCs in the spleens of BXD2mice. A, Flow cytometry was performed on B6 and BXD2 spleen cells labeled with
anti-IFN–a and anti-PDCA1Abs.Means6SEM; n = 6); pp, 0.05 comparedwith B6mice.B, Gating on IFN-a+ cells, the frequencies of BXD2 and B6 PDCA1+
cells were determined. Means6 SEM; n = 6.C, ELISA analysis of the levels of IFN-a produced by unstimulated, CpG, or DOTAP-CpG stimulated pDCs isolated
from the spleens ofB6andBXD2mice. pDCswere enrichedand cultured invitro for 24hwith the indicated stimulations.Means6SEM;n=6.D, Confocal imaging
analysis of spleen sections taken fromB6 and BXD2mice thatwere stainedwith fluorescent-conjugated anti-PDCA1, anti-IgM, anti-CD35 (FDC), and PNA-biotin
followed by an Alexa 350–conjugated secondary Ab as described in theMaterials and Methods. Data are representative of three independent experiments.
444 IFN-a PROMOTES FOLLICULAR ENTRY OF Ag-DELIVERY MZ-P B CELLS
cells, we found that the pDCs are the primary IFN-a–producing cellsin the spleens of BXD2 mice (Fig. 1B), which is consistent withprevious reports that pDCs are efficient producers of type I IFNs(30–34). Stimulation of pDCs by type A CpG, a Toll-like receptor9 ligand, can elicit the expression and secretion of IFN-a (30, 33).We found that pDCs isolated from the spleens of BXD2 mice weremore responsive to in vitro stimulation with type A CpG or type ACpG-DOTAP and produced significantly greater amounts of IFN-ain response to these agents than equivalent numbers of pDCs iso-lated from the spleens of B6 mice (Fig. 1C). Immunofluorescentimaging of spleen sections of B6 mice showed small numbers ofpDCs (green) that formed a single layer surrounding nonreactiveFOs (Fig. 1D). In the spleens of wild-type (WT) naive BXD2 mice,there were large FOs with active PNA+ GCs (blue). Surroundingthese active GCs were increased numbers of pDCs (green) that werelocated away from the GCs but were primarily in the marginal pe-riphery of the FOs in proximity to the MS (Fig. 1D).
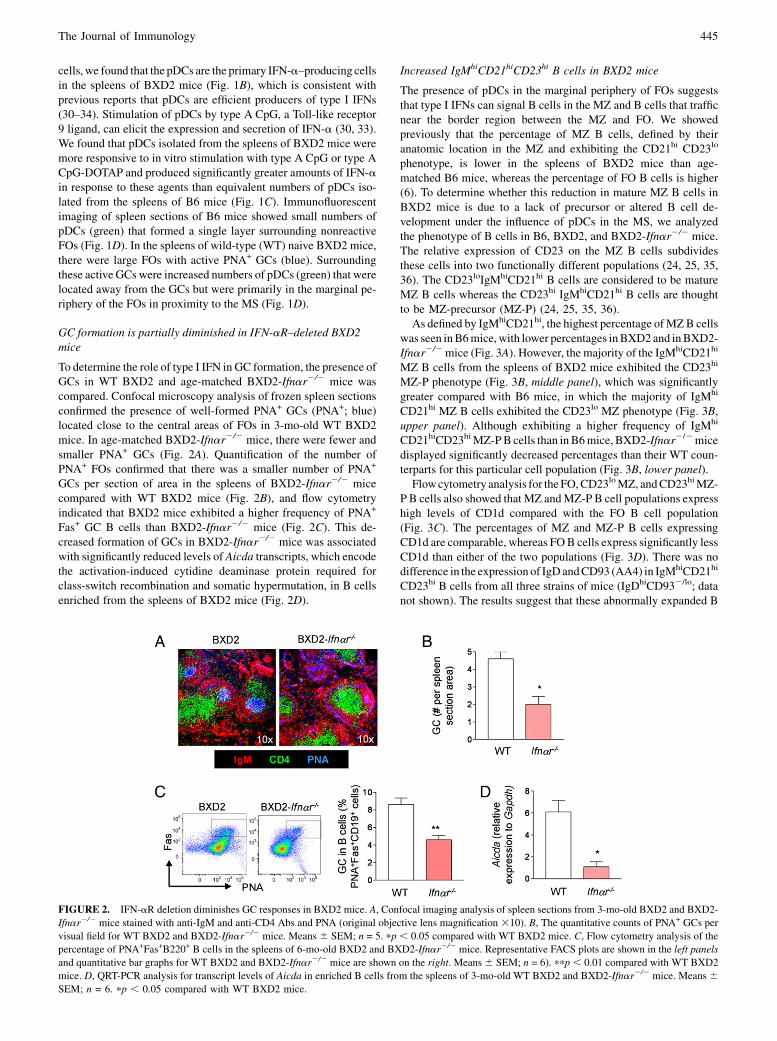
GC formation is partially diminished in IFN-aR–deleted BXD2mice
To determine the role of type I IFN in GC formation, the presence ofGCs in WT BXD2 and age-matched BXD2-Ifnar2/2 mice wascompared. Confocal microscopy analysis of frozen spleen sectionsconfirmed the presence of well-formed PNA+ GCs (PNA+; blue)located close to the central areas of FOs in 3-mo-old WT BXD2mice. In age-matched BXD2-Ifnar2/2 mice, there were fewer andsmaller PNA+ GCs (Fig. 2A). Quantification of the number ofPNA+ FOs confirmed that there was a smaller number of PNA+
GCs per section of area in the spleens of BXD2-Ifnar2/2 micecompared with WT BXD2 mice (Fig. 2B), and flow cytometryindicated that BXD2 mice exhibited a higher frequency of PNA+
Fas+ GC B cells than BXD2-Ifnar2/2 mice (Fig. 2C). This de-creased formation of GCs in BXD2-Ifnar2/2 mice was associatedwith significantly reduced levels of Aicda transcripts, which encodethe activation-induced cytidine deaminase protein required forclass-switch recombination and somatic hypermutation, in B cellsenriched from the spleens of BXD2 mice (Fig. 2D).
Increased IgMhiCD21hiCD23hi B cells in BXD2 mice
The presence of pDCs in the marginal periphery of FOs suggeststhat type I IFNs can signal B cells in the MZ and B cells that trafficnear the border region between the MZ and FO. We showedpreviously that the percentage of MZ B cells, defined by theiranatomic location in the MZ and exhibiting the CD21hi CD23lo
phenotype, is lower in the spleens of BXD2 mice than age-matched B6 mice, whereas the percentage of FO B cells is higher(6). To determine whether this reduction in mature MZ B cells inBXD2 mice is due to a lack of precursor or altered B cell de-velopment under the influence of pDCs in the MS, we analyzedthe phenotype of B cells in B6, BXD2, and BXD2-Ifnar2/2 mice.The relative expression of CD23 on the MZ B cells subdividesthese cells into two functionally different populations (24, 25, 35,36). The CD23loIgMhiCD21hi B cells are considered to be matureMZ B cells whereas the CD23hi IgMhiCD21hi B cells are thoughtto be MZ-precursor (MZ-P) (24, 25, 35, 36).As defined by IgMhiCD21hi, the highest percentage ofMZB cells
was seen inB6mice,with lower percentages inBXD2and inBXD2-Ifnar2/2mice (Fig. 3A). However, the majority of the IgMhiCD21hi
MZ B cells from the spleens of BXD2 mice exhibited the CD23hi
MZ-P phenotype (Fig. 3B, middle panel), which was significantlygreater compared with B6 mice, in which the majority of IgMhi
CD21hi MZ B cells exhibited the CD23lo MZ phenotype (Fig. 3B,upper panel). Although exhibiting a higher frequency of IgMhi
CD21hiCD23hiMZ-PB cells than inB6mice, BXD2-Ifnar2/2micedisplayed significantly decreased percentages than their WT coun-terparts for this particular cell population (Fig. 3B, lower panel).Flowcytometry analysis for theFO,CD23loMZ, andCD23hiMZ-
P B cells also showed that MZ andMZ-P B cell populations expresshigh levels of CD1d compared with the FO B cell population(Fig. 3C). The percentages of MZ and MZ-P B cells expressingCD1d are comparable, whereas FOB cells express significantly lessCD1d than either of the two populations (Fig. 3D). There was nodifference in the expression of IgDandCD93 (AA4) in IgMhiCD21hi
CD23hi B cells from all three strains of mice (IgDhiCD932/lo; datanot shown). The results suggest that these abnormally expanded B
FIGURE 2. IFN-aR deletion diminishes GC responses in BXD2 mice. A, Confocal imaging analysis of spleen sections from 3-mo-old BXD2 and BXD2-
Ifnar2/2 mice stained with anti-IgM and anti-CD4 Abs and PNA (original objective lens magnification 310). B, The quantitative counts of PNA+ GCs per
visual field for WT BXD2 and BXD2-Ifnar2/2 mice. Means 6 SEM; n = 5. pp , 0.05 compared with WT BXD2 mice. C, Flow cytometry analysis of the
percentage of PNA+Fas+B220+ B cells in the spleens of 6-mo-old BXD2 and BXD2-Ifnar2/2 mice. Representative FACS plots are shown in the left panels
and quantitative bar graphs for WT BXD2 and BXD2-Ifnar2/2 mice are shown on the right. Means 6 SEM; n = 6). ppp , 0.01 compared with WT BXD2
mice. D, QRT-PCR analysis for transcript levels of Aicda in enriched B cells from the spleens of 3-mo-old WT BXD2 and BXD2-Ifnar2/2 mice. Means 6SEM; n = 6. pp , 0.05 compared with WT BXD2 mice.
The Journal of Immunology 445
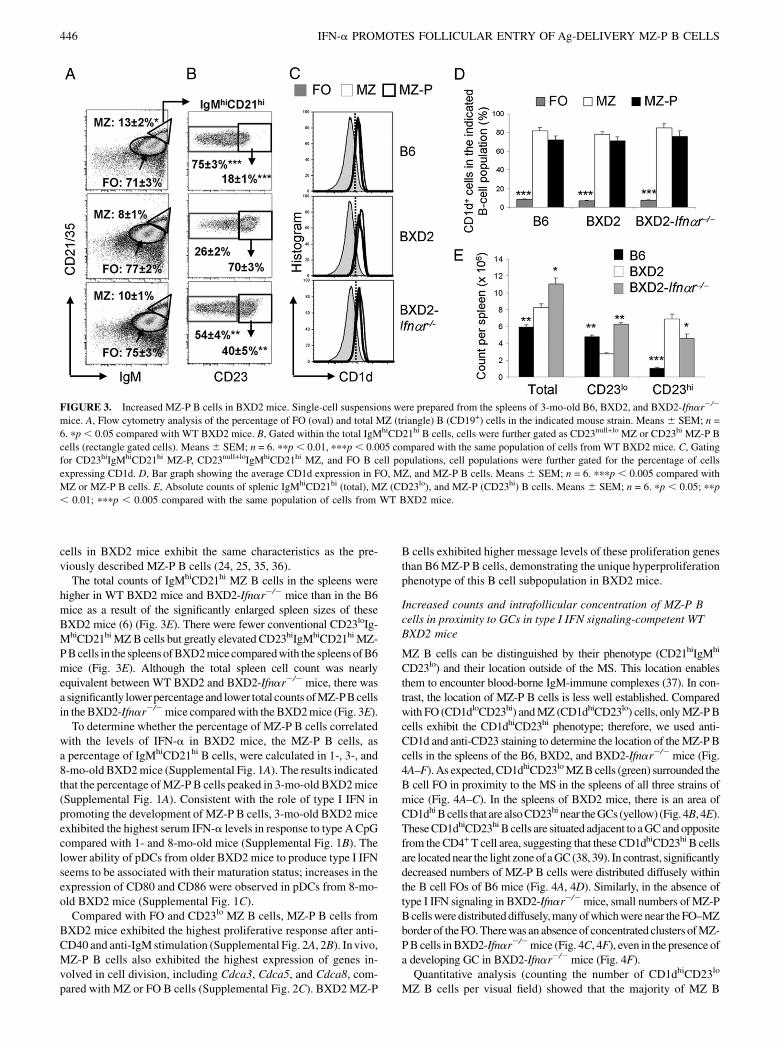
cells in BXD2 mice exhibit the same characteristics as the pre-viously described MZ-P B cells (24, 25, 35, 36).The total counts of IgMhiCD21hi MZ B cells in the spleens were
higher in WT BXD2 mice and BXD2-Ifnar2/2 mice than in the B6mice as a result of the significantly enlarged spleen sizes of theseBXD2 mice (6) (Fig. 3E). There were fewer conventional CD23loIg-MhiCD21hiMZB cells but greatly elevated CD23hiIgMhiCD21hiMZ-PBcells in the spleens ofBXD2micecomparedwith the spleens ofB6mice (Fig. 3E). Although the total spleen cell count was nearlyequivalent between WT BXD2 and BXD2-Ifnar2/2 mice, there wasa significantly lowerpercentage and lower total counts ofMZ-PBcellsin the BXD2-Ifnar2/2mice comparedwith the BXD2mice (Fig. 3E).To determine whether the percentage of MZ-P B cells correlated
with the levels of IFN-a in BXD2 mice, the MZ-P B cells, asa percentage of IgMhiCD21hi B cells, were calculated in 1-, 3-, and8-mo-old BXD2mice (Supplemental Fig. 1A). The results indicatedthat the percentage ofMZ-PB cells peaked in 3-mo-old BXD2mice(Supplemental Fig. 1A). Consistent with the role of type I IFN inpromoting the development of MZ-P B cells, 3-mo-old BXD2miceexhibited the highest serum IFN-a levels in response to type ACpGcompared with 1- and 8-mo-old mice (Supplemental Fig. 1B). Thelower ability of pDCs from older BXD2 mice to produce type I IFNseems to be associated with their maturation status; increases in theexpression of CD80 and CD86 were observed in pDCs from 8-mo-old BXD2 mice (Supplemental Fig. 1C).Compared with FO and CD23lo MZ B cells, MZ-P B cells from
BXD2 mice exhibited the highest proliferative response after anti-CD40 and anti-IgM stimulation (Supplemental Fig. 2A, 2B). Invivo,MZ-P B cells also exhibited the highest expression of genes in-volved in cell division, including Cdca3, Cdca5, and Cdca8, com-pared with MZ or FO B cells (Supplemental Fig. 2C). BXD2MZ-P
B cells exhibited higher message levels of these proliferation genesthan B6MZ-P B cells, demonstrating the unique hyperproliferationphenotype of this B cell subpopulation in BXD2 mice.
Increased counts and intrafollicular concentration of MZ-P Bcells in proximity to GCs in type I IFN signaling-competent WTBXD2 mice
MZ B cells can be distinguished by their phenotype (CD21hiIgMhi
CD23lo) and their location outside of the MS. This location enablesthem to encounter blood-borne IgM-immune complexes (37). In con-trast, the location of MZ-P B cells is less well established. Comparedwith FO (CD1dloCD23hi) andMZ (CD1dhiCD23lo) cells, onlyMZ-PBcells exhibit the CD1dhiCD23hi phenotype; therefore, we used anti-CD1d and anti-CD23 staining to determine the location of theMZ-PBcells in the spleens of the B6, BXD2, and BXD2-Ifnar2/2 mice (Fig.4A–F).As expected,CD1dhiCD23loMZBcells (green) surrounded theB cell FO in proximity to the MS in the spleens of all three strains ofmice (Fig. 4A–C). In the spleens of BXD2 mice, there is an area ofCD1dhiBcells that are alsoCD23hi near theGCs (yellow) (Fig. 4B, 4E).TheseCD1dhiCD23hi B cells are situated adjacent to aGCandoppositefrom theCD4+ T cell area, suggesting that these CD1dhiCD23hi B cellsare locatednear the light zoneof aGC (38, 39). In contrast, significantlydecreased numbers of MZ-P B cells were distributed diffusely withinthe B cell FOs of B6 mice (Fig. 4A, 4D). Similarly, in the absence oftype I IFN signaling in BXD2-Ifnar2/2mice, small numbers of MZ-PBcellsweredistributeddiffusely,manyofwhichwere near theFO–MZborder of theFO.Therewas an absenceof concentrated clusters ofMZ-PBcells inBXD2-Ifnar2/2mice (Fig. 4C, 4F), even in the presence ofa developing GC in BXD2-Ifnar2/2mice (Fig. 4F).Quantitative analysis (counting the number of CD1dhiCD23lo
MZ B cells per visual field) showed that the majority of MZ B
FIGURE 3. Increased MZ-P B cells in BXD2 mice. Single-cell suspensions were prepared from the spleens of 3-mo-old B6, BXD2, and BXD2-Ifnar2/2
mice. A, Flow cytometry analysis of the percentage of FO (oval) and total MZ (triangle) B (CD19+) cells in the indicated mouse strain. Means 6 SEM; n =
6. pp, 0.05 compared with WT BXD2 mice. B, Gated within the total IgMhiCD21hi B cells, cells were further gated as CD23null+lo MZ or CD23hi MZ-P B
cells (rectangle gated cells). Means6 SEM; n = 6. ppp, 0.01, pppp, 0.005 compared with the same population of cells from WT BXD2 mice. C, Gating
for CD23hiIgMhiCD21hi MZ-P, CD23null+loIgMhiCD21hi MZ, and FO B cell populations, cell populations were further gated for the percentage of cells
expressing CD1d. D, Bar graph showing the average CD1d expression in FO, MZ, and MZ-P B cells. Means 6 SEM; n = 6. pppp , 0.005 compared with
MZ or MZ-P B cells. E, Absolute counts of splenic IgMhiCD21hi (total), MZ (CD23lo), and MZ-P (CD23hi) B cells. Means 6 SEM; n = 6. pp , 0.05; ppp
, 0.01; pppp , 0.005 compared with the same population of cells from WT BXD2 mice.
446 IFN-a PROMOTES FOLLICULAR ENTRY OF Ag-DELIVERY MZ-P B CELLS
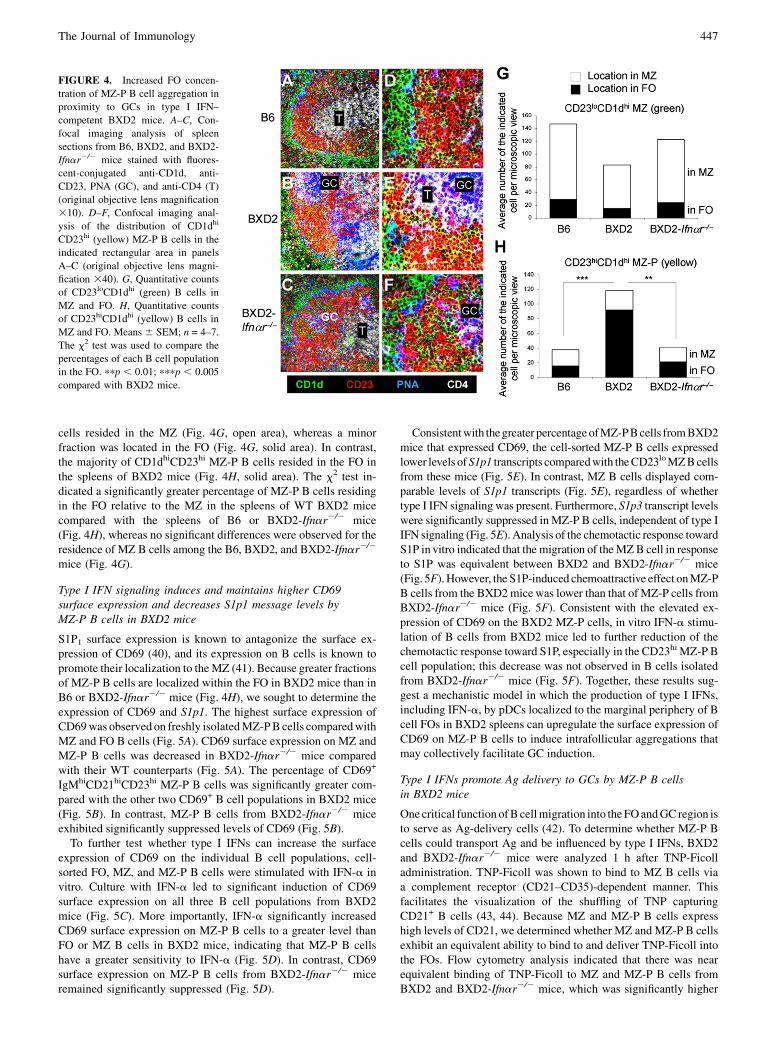
cells resided in the MZ (Fig. 4G, open area), whereas a minorfraction was located in the FO (Fig. 4G, solid area). In contrast,the majority of CD1dhiCD23hi MZ-P B cells resided in the FO inthe spleens of BXD2 mice (Fig. 4H, solid area). The x2 test in-dicated a significantly greater percentage of MZ-P B cells residingin the FO relative to the MZ in the spleens of WT BXD2 micecompared with the spleens of B6 or BXD2-Ifnar2/2 mice(Fig. 4H), whereas no significant differences were observed for theresidence of MZ B cells among the B6, BXD2, and BXD2-Ifnar2/2
mice (Fig. 4G).
Type I IFN signaling induces and maintains higher CD69surface expression and decreases S1p1 message levels byMZ-P B cells in BXD2 mice
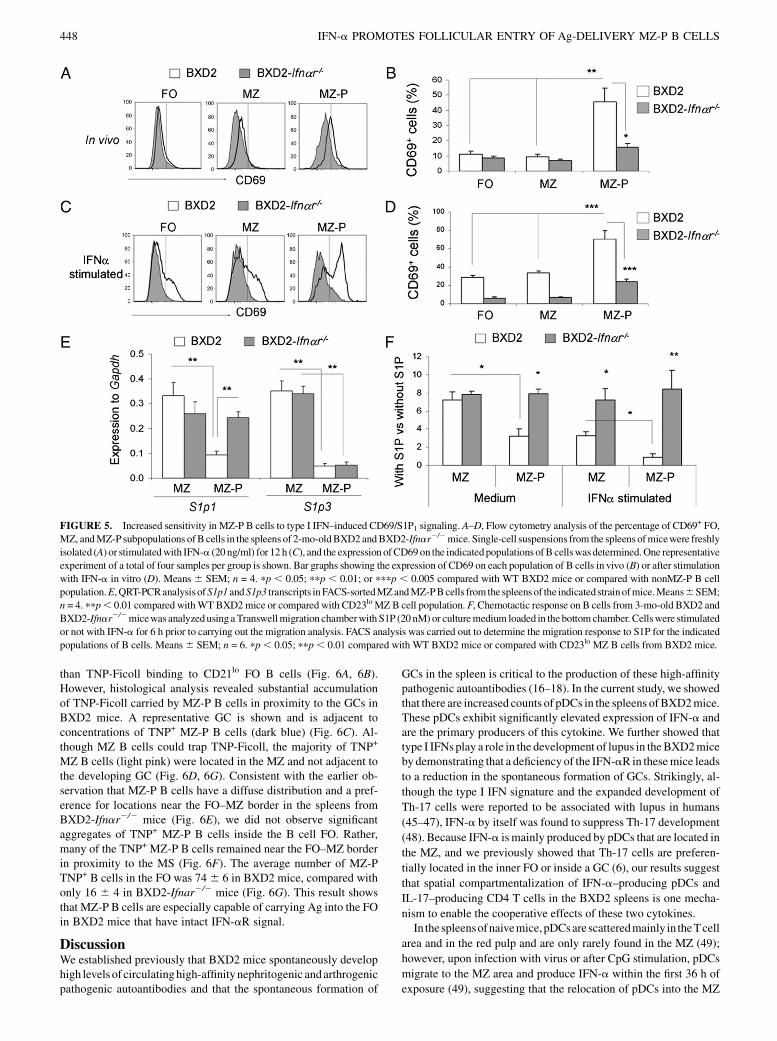
S1P1 surface expression is known to antagonize the surface ex-pression of CD69 (40), and its expression on B cells is known topromote their localization to theMZ (41). Because greater fractionsof MZ-P B cells are localized within the FO in BXD2 mice than inB6 or BXD2-Ifnar2/2 mice (Fig. 4H), we sought to determine theexpression of CD69 and S1p1. The highest surface expression ofCD69was observed on freshly isolatedMZ-PB cells comparedwithMZ and FO B cells (Fig. 5A). CD69 surface expression on MZ andMZ-P B cells was decreased in BXD2-Ifnar2/2 mice comparedwith their WT counterparts (Fig. 5A). The percentage of CD69+
IgMhiCD21hiCD23hi MZ-P B cells was significantly greater com-pared with the other two CD69+ B cell populations in BXD2 mice(Fig. 5B). In contrast, MZ-P B cells from BXD2-Ifnar2/2 miceexhibited significantly suppressed levels of CD69 (Fig. 5B).To further test whether type I IFNs can increase the surface
expression of CD69 on the individual B cell populations, cell-sorted FO, MZ, and MZ-P B cells were stimulated with IFN-a invitro. Culture with IFN-a led to significant induction of CD69surface expression on all three B cell populations from BXD2mice (Fig. 5C). More importantly, IFN-a significantly increasedCD69 surface expression on MZ-P B cells to a greater level thanFO or MZ B cells in BXD2 mice, indicating that MZ-P B cellshave a greater sensitivity to IFN-a (Fig. 5D). In contrast, CD69surface expression on MZ-P B cells from BXD2-Ifnar2/2 miceremained significantly suppressed (Fig. 5D).
Consistentwith thegreater percentageofMZ-PBcells fromBXD2mice that expressed CD69, the cell-sorted MZ-P B cells expressedlower levels ofS1p1 transcripts comparedwith theCD23loMZBcellsfrom these mice (Fig. 5E). In contrast, MZ B cells displayed com-parable levels of S1p1 transcripts (Fig. 5E), regardless of whethertype I IFN signaling was present. Furthermore, S1p3 transcript levelswere significantly suppressed in MZ-P B cells, independent of type IIFN signaling (Fig. 5E). Analysis of the chemotactic response towardS1P in vitro indicated that the migration of theMZB cell in responseto S1P was equivalent between BXD2 and BXD2-Ifnar2/2 mice(Fig. 5F).However, the S1P-induced chemoattractive effect onMZ-PB cells from the BXD2micewas lower than that of MZ-P cells fromBXD2-Ifnar2/2 mice (Fig. 5F). Consistent with the elevated ex-pression of CD69 on the BXD2 MZ-P cells, in vitro IFN-a stimu-lation of B cells from BXD2 mice led to further reduction of thechemotactic response toward S1P, especially in the CD23hi MZ-P Bcell population; this decrease was not observed in B cells isolatedfrom BXD2-Ifnar2/2 mice (Fig. 5F). Together, these results sug-gest a mechanistic model in which the production of type I IFNs,including IFN-a, by pDCs localized to the marginal periphery of Bcell FOs in BXD2 spleens can upregulate the surface expression ofCD69 on MZ-P B cells to induce intrafollicular aggregations thatmay collectively facilitate GC induction.
Type I IFNs promote Ag delivery to GCs by MZ-P B cellsin BXD2 mice
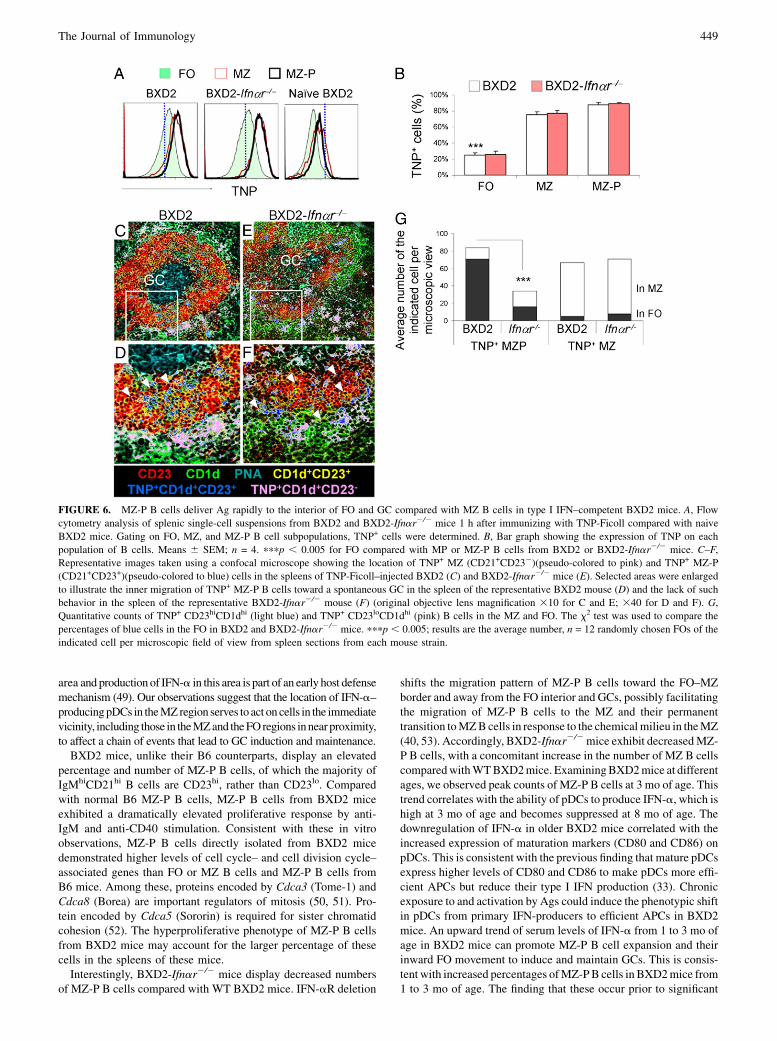
Onecritical function ofBcellmigration into theFOandGC region isto serve as Ag-delivery cells (42). To determine whether MZ-P Bcells could transport Ag and be influenced by type I IFNs, BXD2and BXD2-Ifnar2/2 mice were analyzed 1 h after TNP-Ficolladministration. TNP-Ficoll was shown to bind to MZ B cells viaa complement receptor (CD21–CD35)-dependent manner. Thisfacilitates the visualization of the shuffling of TNP capturingCD21+ B cells (43, 44). Because MZ and MZ-P B cells expresshigh levels of CD21, we determined whether MZ and MZ-P B cellsexhibit an equivalent ability to bind to and deliver TNP-Ficoll intothe FOs. Flow cytometry analysis indicated that there was nearequivalent binding of TNP-Ficoll to MZ and MZ-P B cells fromBXD2 and BXD2-Ifnar2/2 mice, which was significantly higher
FIGURE 4. Increased FO concen-
tration of MZ-P B cell aggregation in
proximity to GCs in type I IFN–
competent BXD2 mice. A–C, Con-
focal imaging analysis of spleen
sections from B6, BXD2, and BXD2-
Ifnar2/2 mice stained with fluores-
cent-conjugated anti-CD1d, anti-
CD23, PNA (GC), and anti-CD4 (T)
(original objective lens magnification
310). D–F, Confocal imaging anal-
ysis of the distribution of CD1dhi
CD23hi (yellow) MZ-P B cells in the
indicated rectangular area in panels
A–C (original objective lens magni-
fication 340). G, Quantitative counts
of CD23loCD1dhi (green) B cells in
MZ and FO. H, Quantitative counts
of CD23hiCD1dhi (yellow) B cells in
MZ and FO. Means 6 SEM; n = 4–7.
The x2 test was used to compare the
percentages of each B cell population
in the FO. ppp , 0.01; pppp , 0.005
compared with BXD2 mice.
The Journal of Immunology 447
than TNP-Ficoll binding to CD21lo FO B cells (Fig. 6A, 6B).However, histological analysis revealed substantial accumulationof TNP-Ficoll carried by MZ-P B cells in proximity to the GCs inBXD2 mice. A representative GC is shown and is adjacent toconcentrations of TNP+ MZ-P B cells (dark blue) (Fig. 6C). Al-though MZ B cells could trap TNP-Ficoll, the majority of TNP+
MZ B cells (light pink) were located in the MZ and not adjacent tothe developing GC (Fig. 6D, 6G). Consistent with the earlier ob-servation that MZ-P B cells have a diffuse distribution and a pref-erence for locations near the FO–MZ border in the spleens fromBXD2-Ifnar2/2 mice (Fig. 6E), we did not observe significantaggregates of TNP+ MZ-P B cells inside the B cell FO. Rather,many of the TNP+ MZ-P B cells remained near the FO–MZ borderin proximity to the MS (Fig. 6F). The average number of MZ-PTNP+ B cells in the FO was 746 6 in BXD2 mice, compared withonly 16 6 4 in BXD2-Ifnar2/2 mice (Fig. 6G). This result showsthat MZ-P B cells are especially capable of carrying Ag into the FOin BXD2 mice that have intact IFN-aR signal.
DiscussionWe established previously that BXD2 mice spontaneously develophigh levels of circulatinghigh-affinitynephritogenic andarthrogenicpathogenic autoantibodies and that the spontaneous formation of
GCs in the spleen is critical to the production of these high-affinitypathogenic autoantibodies (16–18). In the current study, we showedthat there are increased counts of pDCs in the spleens ofBXD2mice.These pDCs exhibit significantly elevated expression of IFN-a andare the primary producers of this cytokine. We further showed thattype I IFNs play a role in the development of lupus in theBXD2miceby demonstrating that a deficiency of the IFN-aR in thesemice leads
to a reduction in the spontaneous formation of GCs. Strikingly, al-
though the type I IFN signature and the expanded development of
Th-17 cells were reported to be associated with lupus in humans
(45–47), IFN-a by itself was found to suppress Th-17 development
(48). Because IFN-a is mainly produced by pDCs that are located in
the MZ, and we previously showed that Th-17 cells are preferen-
tially located in the inner FO or inside a GC (6), our results suggest
that spatial compartmentalization of IFN-a–producing pDCs and
IL-17–producing CD4 T cells in the BXD2 spleens is one mecha-
nism to enable the cooperative effects of these two cytokines.In thespleensofnaivemice,pDCsarescatteredmainly in theTcell
area and in the red pulp and are only rarely found in the MZ (49);
however, upon infection with virus or after CpG stimulation, pDCs
migrate to the MZ area and produce IFN-a within the first 36 h of
exposure (49), suggesting that the relocation of pDCs into the MZ
FIGURE 5. Increased sensitivity in MZ-P B cells to type I IFN–induced CD69/S1P1 signaling. A–D, Flow cytometry analysis of the percentage of CD69+ FO,
MZ, andMZ-P subpopulations ofB cells in the spleens of 2-mo-oldBXD2andBXD2-Ifnar2/2mice. Single-cell suspensions from the spleens ofmicewere freshly
isolated (A) or stimulatedwith IFN-a (20 ng/ml) for 12 h (C), and the expression ofCD69on the indicatedpopulations ofB cellswasdetermined.One representative
experiment of a total of four samples per group is shown. Bar graphs showing the expression of CD69 on each population of B cells in vivo (B) or after stimulation
with IFN-a in vitro (D). Means 6 SEM; n = 4. pp , 0.05; ppp , 0.01; or pppp , 0.005 compared with WT BXD2 mice or compared with nonMZ-P B cell
population.E,QRT-PCRanalysis ofS1p1 andS1p3 transcripts inFACS-sortedMZandMZ-PBcells from the spleens of the indicatedstrainofmice.Means6SEM;
n = 4. ppp, 0.01 compared withWTBXD2mice or compared with CD23loMZ B cell population. F, Chemotactic response on B cells from 3-mo-old BXD2 and
BXD2-Ifnar2/2micewas analyzedusing aTranswellmigration chamberwith S1P (20nM)or culturemedium loaded in the bottomchamber. Cellswere stimulated
or not with IFN-a for 6 h prior to carrying out the migration analysis. FACS analysis was carried out to determine the migration response to S1P for the indicated
populations of B cells. Means6 SEM; n = 6. pp, 0.05; ppp, 0.01 compared with WT BXD2 mice or compared with CD23lo MZ B cells from BXD2 mice.
448 IFN-a PROMOTES FOLLICULAR ENTRY OF Ag-DELIVERY MZ-P B CELLS
area and production of IFN-a in this area is part of an early host defensemechanism (49). Our observations suggest that the location of IFN-a–producingpDCs in theMZregion serves to act on cells in the immediatevicinity, including those in theMZand theFOregions innearproximity,to affect a chain of events that lead to GC induction and maintenance.BXD2 mice, unlike their B6 counterparts, display an elevated
percentage and number of MZ-P B cells, of which the majority ofIgMhiCD21hi B cells are CD23hi, rather than CD23lo. Comparedwith normal B6 MZ-P B cells, MZ-P B cells from BXD2 miceexhibited a dramatically elevated proliferative response by anti-IgM and anti-CD40 stimulation. Consistent with these in vitroobservations, MZ-P B cells directly isolated from BXD2 micedemonstrated higher levels of cell cycle– and cell division cycle–associated genes than FO or MZ B cells and MZ-P B cells fromB6 mice. Among these, proteins encoded by Cdca3 (Tome-1) andCdca8 (Borea) are important regulators of mitosis (50, 51). Pro-tein encoded by Cdca5 (Sororin) is required for sister chromatidcohesion (52). The hyperproliferative phenotype of MZ-P B cellsfrom BXD2 mice may account for the larger percentage of thesecells in the spleens of these mice.Interestingly, BXD2-Ifnar2/2 mice display decreased numbers
of MZ-P B cells compared with WT BXD2 mice. IFN-aR deletion
shifts the migration pattern of MZ-P B cells toward the FO–MZborder and away from the FO interior and GCs, possibly facilitatingthe migration of MZ-P B cells to the MZ and their permanenttransition toMZBcells in response to the chemicalmilieu in theMZ(40, 53). Accordingly, BXD2-Ifnar2/2mice exhibit decreasedMZ-P B cells, with a concomitant increase in the number of MZ B cellscomparedwithWTBXD2mice. ExaminingBXD2mice at differentages, we observed peak counts of MZ-P B cells at 3 mo of age. Thistrend correlates with the ability of pDCs to produce IFN-a, which ishigh at 3 mo of age and becomes suppressed at 8 mo of age. Thedownregulation of IFN-a in older BXD2 mice correlated with theincreased expression of maturation markers (CD80 and CD86) onpDCs. This is consistent with the previous finding that mature pDCsexpress higher levels of CD80 and CD86 to make pDCs more effi-cient APCs but reduce their type I IFN production (33). Chronicexposure to and activation by Ags could induce the phenotypic shiftin pDCs from primary IFN-producers to efficient APCs in BXD2mice. An upward trend of serum levels of IFN-a from 1 to 3 mo ofage in BXD2 mice can promote MZ-P B cell expansion and theirinward FO movement to induce and maintain GCs. This is consis-tent with increased percentages ofMZ-PB cells in BXD2mice from1 to 3 mo of age. The finding that these occur prior to significant
FIGURE 6. MZ-P B cells deliver Ag rapidly to the interior of FO and GC compared with MZ B cells in type I IFN–competent BXD2 mice. A, Flow
cytometry analysis of splenic single-cell suspensions from BXD2 and BXD2-Ifnar2/2 mice 1 h after immunizing with TNP-Ficoll compared with naive
BXD2 mice. Gating on FO, MZ, and MZ-P B cell subpopulations, TNP+ cells were determined. B, Bar graph showing the expression of TNP on each
population of B cells. Means 6 SEM; n = 4. pppp , 0.005 for FO compared with MP or MZ-P B cells from BXD2 or BXD2-Ifnar2/2 mice. C–F,
Representative images taken using a confocal microscope showing the location of TNP+ MZ (CD21+CD232)(pseudo-colored to pink) and TNP+ MZ-P
(CD21+CD23+)(pseudo-colored to blue) cells in the spleens of TNP-Ficoll–injected BXD2 (C) and BXD2-Ifnar2/2 mice (E). Selected areas were enlarged
to illustrate the inner migration of TNP+ MZ-P B cells toward a spontaneous GC in the spleen of the representative BXD2 mouse (D) and the lack of such
behavior in the spleen of the representative BXD2-Ifnar2/2 mouse (F) (original objective lens magnification 310 for C and E; 340 for D and F). G,
Quantitative counts of TNP+ CD23hiCD1dhi (light blue) and TNP+ CD23loCD1dhi (pink) B cells in the MZ and FO. The x2 test was used to compare the
percentages of blue cells in the FO in BXD2 and BXD2-Ifnar2/2 mice. pppp , 0.005; results are the average number, n = 12 randomly chosen FOs of the
indicated cell per microscopic field of view from spleen sections from each mouse strain.
The Journal of Immunology 449

development of autoantibody production, which occurs at 4 to 6 moof age (17), indicates that MZ-P B cell expansion and its regulationby type I IFN is an early event prior to disease onset.Importantly, our results suggest that type I IFNs exhibit an im-
portant effect to promote the FO-orientedmigration ofMZ-PB cells.BecauseCD69 surface expression is diminished inFO,MZ, andMZ-P B cells from BXD2-Ifnar2/2 mice, type I IFN can regulate S1p1expression within each of these B cell populations. Despite IFN-astimulation under in vitro conditions, CD69 surface expression in allthree B cell populations in BXD2-Ifnar2/2 mice remained sup-pressed. Interestingly, MZ-P B cells in IFN-aR intact BXD2 miceexpressed the highest levels of cell surface CD69. When stimulatedwith IFN-a, MZ-P B cells from BXD2 mice exhibited the highestsurface expression of CD69 compared with FO or MZ B cells.Consistent with the antagonistic relationship between CD69 andS1P1 expression, when IFN-aR is deleted, S1p1 transcript levelsbecome elevated. Furthermore, S1P3, known to be highly expressedin MZ B cells, had significantly depressed transcript levels in theMZ-PB cells, regardless ofwhether type I IFN signalingwas presentor absent. S1P3 stabilizes the positioning ofB cells in theMZ(41). Itsdepressed levels inMZ-PB cells further suggest that, comparedwithMZ cells, MZ-P B cells exhibit a higher tendency to migrate into theFO.We also determined the chemotactic responses ofMZ andMZ-Pfrom BXD2 and BXD2-Ifnar2/2 mice to CXCL12, CXCL13,CCL19, and CCL21; CXCL13 was the only chemokine to providedifferentially chemoattractive effects to MZ and MZ-P B cells fromthese two strains. MZ andMZ-P B cells from BXD2mice exhibiteda stronger migration response toward CXCL13 compared with thesame cells from BXD2-Ifnar2/2 mice (data not shown). Therefore,the results suggest that the dominant effects of IFN-a to regulate theFO entry of MZ-P B cells were via its suppression of the S1P che-motactic response. Potentially, pDCs residing in the MZ can gen-erate massive amounts of IFN-a to achieve this.McHeyzer-Williams et al. (54) reported that during the initial
phase of protein immunization, naive B cells specifically recog-nize the protein Ag and transport this Ag to initiate the T cell–dependent B cell response in checkpoint I. They further proposedthat these B cells contact cell-associated Ags for priming duringthis early initiation phase of adaptive immunity. In parallel, naiveTh cells expand, differentiate into effector Th cells, and migrate tothe T–B borders to contact peptide–MHC II–expressing Ag-primed B cells in checkpoint II. Despite this importance of B cellsas the initial Ag-transporting B cells in adaptive immunity, theidentity and unique property of these initial Ag-transportingB cells and how these B cells can affect the development of au-toimmune disease are not well known (55). Our present studysuggests that MZ-P B cells are the dominant B cells that transportAg directly into a GC in BXD2 mice. In the context of autoim-munity, this continuous supply of Ag to the GCs can potentiallydrive and maintain the generation of autoantibody-producingplasma cells. However, in BXD2-Ifnar2/2 mice, MZ-P B cellscarrying Ag are now relegated to the periphery near the FO–MZborder in proximity to the MS and, thereby, prevent the influx ofAg-delivery B cells into the inner FOs. The present results suggestthat production of IFN-a in the MS to drive the FO migration ofAg-transporting MZ-P B cells into the GC area exhibits the po-tential to break the closely monitored checkpoints I and II of theT-dependent humoral responses, leading to the development ofspontaneous GCs and pathogenic autoantibodies in BXD2 mice.
AcknowledgmentsWe thank Marion L. Spell of the UAB AIDS FACS Core Facility and Enid
Keyser of the Arthritis and Musculoskeletal Disease Center Analytic and
Preparative Cytometry Facility for operating the FACS instrument. We
thank Albert Tousson of the High Resolution Imaging Facility for assis-
tance with operating the confocal imaging equipment. We thank Dr. Joce-
lyn Demengeot, Instituto Gulbenkian de Ciencia, Oeiras, Portugal, for
providing B6-Ifnar2/2 mice. We thank Dr. Fiona Hunter for expert review
of the manuscript and Carol Humber for excellent secretarial assistance.
DisclosuresThe authors have no financial conflicts of interest.
References1. Duan, B., H. Niu, Z. Xu, A. H. Sharpe, B. P. Croker, E. S. Sobel, and L. Morel.
2008. Intrafollicular location of marginal zone/CD1d(hi) B cells is associatedwith autoimmune pathology in a mouse model of lupus. Lab. Invest. 88: 1008–1020.
2. Paul, E., A. Nelde, A. Verschoor, and M. C. Carroll. 2007. Follicular exclusion ofautoreactive B cells requires FcgammaRIIb. Int. Immunol. 19: 365–373.
3. Cyster, J. G., S. B. Hartley, and C. C. Goodnow. 1994. Competition for follicularniches excludes self-reactive cells from the recirculating B-cell repertoire. Na-ture 371: 389–395.
4. Schmidt, K. N., and J. G. Cyster. 1999. Follicular exclusion and rapid elimi-nation of hen egg lysozyme autoantigen-binding B cells are dependent oncompetitor B cells, but not on T cells. J. Immunol. 162: 284–291.
5. Corcione, A., L. Ottonello, G. Tortolina, P. Tasso, F. Ghiotto, I. Airoldi,G. Taborelli, F. Malavasi, F. Dallegri, and V. Pistoia. 1997. Recombinant tumornecrosis factor enhances the locomotion of memory and naive B lymphocytesfrom human tonsils through the selective engagement of the type II receptor.Blood 90: 4493–4501.
6. Hsu, H. C., P. Yang, J. Wang, Q. Wu, R. Myers, J. Chen, J. Yi, T. Guentert,A. Tousson, A. L. Stanus, et al. 2008. Interleukin 17-producing T helper cellsand interleukin 17 orchestrate autoreactive germinal center development in au-toimmune BXD2 mice. Nat. Immunol. 9: 166–175.
7. Moratz, C., V. H. Kang, K. M. Druey, C. S. Shi, A. Scheschonka, P. M. Murphy,T. Kozasa, and J. H. Kehrl. 2000. Regulator of G protein signaling 1 (RGS1)markedly impairs Gi a signaling responses of B lymphocytes. J. Immunol. 164:1829–1838.
8. Chan, O., and M. J. Shlomchik. 1998. A new role for B cells in systemic au-toimmunity: B cells promote spontaneous T cell activation in MRL-lpr/lpr mice.J. Immunol. 160: 51–59.
9. Datta, S. K. 2009. Anti-CD20 antibody is an efficient therapeutic tool for theselective removal of autoreactive T cells. Nat. Clin. Pract. Rheumatol. 5: 80–82.
10. Eisenberg, R. 2006. Targeting B cells in SLE: the experience with rituximabtreatment (anti-CD20). Endocr. Metab. Immune Disord. Drug Targets 6: 345–350.
11. Roth, R., R. J. Gee, and M. J. Mamula. 1997. B lymphocytes as autoantigen-presenting cells in the amplification of autoimmunity. Ann. N. Y. Acad. Sci. 815:88–104.
12. Weyand, C. M., and J. J. Goronzy. 2003. Ectopic germinal center formation inrheumatoid synovitis. Ann. N. Y. Acad. Sci. 987: 140–149.
13. Bystry, R. S., V. Aluvihare, K. A. Welch, M. Kallikourdis, and A. G. Betz. 2001.B cells and professional APCs recruit regulatory T cells via CCL4. Nat. Im-munol. 2: 1126–1132.
14. Goronzy, J. J., and C. M. Weyand. 2003. B cells as a therapeutic target in au-toimmune disease. Arthritis Res. Ther. 5: 131–135.
15. Kessel, A., I. Rosner, and E. Toubi. 2008. Rituximab: beyond simple B celldepletion. Clin. Rev. Allergy Immunol. 34: 74–79.
16. Mountz, J. D., P. Yang, Q. Wu, J. Zhou, A. Tousson, A. Fitzgerald, J. Allen,X. Wang, S. Cartner, W. E. Grizzle, et al. 2005. Genetic segregation of spon-taneous erosive arthritis and generalized autoimmune disease in the BXD2 re-combinant inbred strain of mice. Scand. J. Immunol. 61: 128–138.
17. Hsu, H. C., T. Zhou, H. Kim, S. Barnes, P. Yang, Q. Wu, J. Zhou, B. A. Freeman,M. Luo, and J. D. Mountz. 2006. Production of a novel class of polyreactivepathogenic autoantibodies in BXD2 mice causes glomerulonephritis and ar-thritis. Arthritis Rheum. 54: 343–355.
18. Hsu, H. C., Y. Wu, P. Yang, Q. Wu, G. Job, J. Chen, J. Wang, M. A. Accavitti-Loper, W. E. Grizzle, R. H. Carter, et al. 2007. Overexpression of activation-induced cytidine deaminase in B cells is associated with production of highlypathogenic autoantibodies. J. Immunol. 178: 5357–5365.
19. Braun, D., P. Geraldes, and J. Demengeot. 2003. Type I interferon controls theonset and severity of autoimmune manifestations in lpr mice. J. Autoimmun. 20:15–25.
20. Koutouzov, S., A. Mathian, and A. Dalloul. 2006. Type-I interferons and sys-temic lupus erythematosus. Autoimmun. Rev. 5: 554–562.
21. Santiago-Raber, M. L., R. Baccala, K. M. Haraldsson, D. Choubey, T. A. Stewart,D. H. Kono, and A. N. Theofilopoulos. 2003. Type-I interferon receptordeficiency reduces lupus-like disease in NZB mice. J. Exp. Med. 197: 777–788.
22. Jego, G., A. K. Palucka, J. P. Blanck, C. Chalouni, V. Pascual, and J. Banchereau.2003. Plasmacytoid dendritic cells induce plasma cell differentiation throughtype I interferon and interleukin 6. Immunity 19: 225–234.
23. Le Bon, A., G. Schiavoni, G. D’Agostino, I. Gresser, F. Belardelli, andD. F. Tough. 2001. Type i interferons potently enhance humoral immunity andcan promote isotype switching by stimulating dendritic cells in vivo. Immunity14: 461–470.
450 IFN-a PROMOTES FOLLICULAR ENTRY OF Ag-DELIVERY MZ-P B CELLS

24. Allman, D., and S. Pillai. 2008. Peripheral B cell subsets. Curr. Opin. Immunol.20: 149–157.
25. Srivastava, B., W. J. Quinn 3rd, K. Hazard, J. Erikson, and D. Allman. 2005.Characterization of marginal zone B cell precursors. J. Exp. Med. 202: 1225–1234.
26. Moratz, C., and J. H. Kehrl. 2004. In vitro and in vivo assays of B-lymphocytemigration. Methods Mol. Biol. 271: 161–171.
27. Cavanagh, L. L., A. Boyce, L. Smith, J. Padmanabha, L. Filgueira,P. Pietschmann, and R. Thomas. 2005. Rheumatoid arthritis synovium containsplasmacytoid dendritic cells. Arthritis Res. Ther. 7: R230–R240.
28. Yao, Y., L. Richman, C. Morehouse, M. de los Reyes, B. W. Higgs, A. Boutrin,B. White, A. Coyle, J. Krueger, P. A. Kiener, et al. 2008. Type I interferon:potential therapeutic target for psoriasis? PLoS One 3: e2737.
29. Lian, Z. X., K. Kikuchi, G. X. Yang, A. A. Ansari, S. Ikehara, andM. E. Gershwin. 2004. Expansion of bone marrow IFN-a-producing dendriticcells in New Zealand Black (NZB) mice: high level expression of TLR9 andsecretion of IFN-a in NZB bone marrow. J. Immunol. 173: 5283–5289.
30. Colonna, M., G. Trinchieri, and Y. J. Liu. 2004. Plasmacytoid dendritic cells inimmunity. Nat. Immunol. 5: 1219–1226.
31. Hanten, J. A., J. P. Vasilakos, C. L. Riter, L. Neys, K. E. Lipson, S. S. Alkan, andW. Birmachu. 2008. Comparison of human B cell activation by TLR7 and TLR9agonists. BMC Immunol. 9: 39.
32. Hughes, G. C., S. Thomas, C. Li, M. K. Kaja, and E. A. Clark. 2008. Cuttingedge: progesterone regulates IFN-a production by plasmacytoid dendritic cells.J. Immunol. 180: 2029–2033.
33. Liu, Y. J. 2005. IPC: professional type 1 interferon-producing cells and plas-macytoid dendritic cell precursors. Annu. Rev. Immunol. 23: 275–306.
34. Wu, P., J. Wu, S. Liu, X. Han, J. Lu, Y. Shi, J. Wang, L. Lu, and X. Cao. 2008.TLR9/TLR7-triggered downregulation of BDCA2 expression on human plas-macytoid dendritic cells from healthy individuals and lupus patients. Clin. Im-munol. 129: 40–48.
35. Saito, T., S. Chiba, M. Ichikawa, A. Kunisato, T. Asai, K. Shimizu,T. Yamaguchi, G. Yamamoto, S. Seo, K. Kumano, et al. 2003. Notch2 is pref-erentially expressed in mature B cells and indispensable for marginal zone Blineage development. Immunity 18: 675–685.
36. Srivastava, B., R. C. Lindsley, N. Nikbakht, and D. Allman. 2005. Modelsfor rperipheral B cell development and homeostasis. Semin. Immunol. 17: 175–182.
37. Ferguson, A. R., M. E. Youd, and R. B. Corley. 2004. Marginal zone B cellstransport and deposit IgM-containing immune complexes onto follicular den-dritic cells. Int. Immunol. 16: 1411–1422.
38. Allen, C. D., T. Okada, and J. G. Cyster. 2007. Germinal-center organization andcellular dynamics. Immunity 27: 190–202.
39. McHeyzer-Williams, L. J., D. J. Driver, and M. G. McHeyzer-Williams. 2001.Germinal center reaction. Curr. Opin. Hematol. 8: 52–59.
40. Shiow, L. R., D. B. Rosen, N. Brdickova, Y. Xu, J. An, L. L. Lanier, J. G. Cyster,and M. Matloubian. 2006. CD69 acts downstream of interferon-a/b to inhibitS1P1 and lymphocyte egress from lymphoid organs. Nature 440: 540–544.
41. Cinamon, G., M. Matloubian, M. J. Lesneski, Y. Xu, C. Low, T. Lu, R. L. Proia,and J. G. Cyster. 2004. Sphingosine 1-phosphate receptor 1 promotes B celllocalization in the splenic marginal zone. Nat. Immunol. 5: 713–720.
42. Fazilleau, N., L. Mark, L. J. McHeyzer-Williams, andM. G. McHeyzer-Williams.2009. Follicular helper T cells: lineage and location. Immunity 30: 324–335.
43. Cinamon, G., M. A. Zachariah, O. M. Lam, F. W. Foss Jr., and J. G. Cyster. 2008.Follicular shuttling of marginal zone B cells facilitates antigen transport. Nat.Immunol. 9: 54–62.
44. Guinamard, R., M. Okigaki, J. Schlessinger, and J. V. Ravetch. 2000. Absence ofmarginal zone B cells in Pyk-2-deficient mice defines their role in the humoralresponse. Nat. Immunol. 1: 31–36.
45. Pernis, A. B. 2009. Th17 cells in rheumatoid arthritis and systemic lupus er-ythematosus. J. Intern. Med. 265: 644–652.
46. Yang, J., Y. Chu, X. Yang, D. Gao, L. Zhu, X. Yang, L. Wan, and M. Li. 2009.Th17 and natural Treg cell population dynamics in systemic lupus eryth-ematosus. Arthritis Rheum. 60: 1472–1483.
47. Zhao, X. F., H. F. Pan, H. Yuan, W. H. Zhang, X. P. Li, G. H. Wang, G. C. Wu, H.Su, F. M. Pan, W. X. Li, et al. 2009. Increased serum interleukin 17 in patientswith systemic lupus erythematosus. Mol Biol Rep. In press.
48. Moschen, A. R., S. Geiger, I. Krehan, A. Kaser, and H. Tilg. 2008. Interferon-acontrols IL-17 expression in vitro and in vivo. Immunobiology 213: 779–787.
49. Asselin-Paturel, C., G. Brizard, K. Chemin, A. Boonstra, A. O’Garra, A. Vicari,and G. Trinchieri. 2005. Type I interferon dependence of plasmacytoid dendriticcell activation and migration. J. Exp. Med. 201: 1157–1167.
50. Ayad, N. G., S. Rankin, M. Murakami, J. Jebanathirajah, S. Gygi, andM. W. Kirschner. 2003. Tome-1, a trigger of mitotic entry, is degraded during G1via the APC. Cell 113: 101–113.
51. Ruchaud, S., M. Carmena, and W. C. Earnshaw. 2007. The chromosomal pas-senger complex: one for all and all for one. Cell 131: 230–231.
52. Schmitz, J., E. Watrin, P. Lenart, K. Mechtler, and J. M. Peters. 2007. Sororin isrequired for stable binding of cohesin to chromatin and for sister chromatidcohesion in interphase. Curr. Biol. 17: 630–636.
53. Rubtsov, A. V., C. L. Swanson, S. Troy, P. Strauch, R. Pelanda, and R. M. Torres.2008. TLR agonists promote marginal zone B cell activation and facilitateT-dependent IgM responses. J. Immunol. 180: 3882–3888.
54. McHeyzer-Williams, L. J., N. Pelletier, L. Mark, N. Fazilleau, andM. G. McHeyzer-Williams. 2009. Follicular helper T cells as cognate regulatorsof B cell immunity. Curr. Opin. Immunol. 21: 266–273.
55. Batista, F. D., and N. E. Harwood. 2009. The who, how and where of antigenpresentation to B cells. Nat. Rev. Immunol. 9: 15–27.
The Journal of Immunology 451
Copyright © 2022 FDOKUMEN




















