
Regulation of the Innate Immune Response during the Human ...
Upload
independentCategory
view
1download
0
of November 28, 2013.This information is current as
Innate Immune SystemIFN, On Cytokine Production by Cells of the
, A Novel Type IκRegulatory Effect of IFN-
A. MooreUllrich, Giampiero Girolomoni, Cristina Albanesi and PaulYun Hee Cho, David W. LaFleur, Devanshi Shah, Stephen Bernardetta Nardelli, Liubov Zaritskaya, Mark Semenuk,
http://www.jimmunol.org/content/169/9/48222002; 169:4822-4830; ;J Immunol
Referenceshttp://www.jimmunol.org/content/169/9/4822.full#ref-list-1
, 25 of which you can access for free at: cites 46 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2002 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on Novem
ber 28, 2013http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on N
ovember 28, 2013
http://ww
w.jim
munol.org/
Dow
nloaded from

Regulatory Effect of IFN-�, A Novel Type I IFN, On CytokineProduction by Cells of the Innate Immune System
Bernardetta Nardelli,1* Liubov Zaritskaya,* Mark Semenuk,* Yun Hee Cho,*David W. LaFleur,* Devanshi Shah,* Stephen Ullrich,* Giampiero Girolomoni,†
Cristina Albanesi,† and Paul A. Moore*
IFN-� is a recently identified type I IFN that exhibits both structural and functional homology with the other type I IFN subclasses.In this study, we have investigated the effect of IFN-� on cells of the innate immune system by comparing cytokine release followingtreatment of human cells with either IFN-� or two recombinant IFN subtypes, IFN-� and IFN-�2a. Although IFN-�2a failed tostimulate monocyte cytokine secretion, IFN-�, like IFN- �, induced the release of several cytokines from both monocytes anddendritic cells, without the requirement of a costimulatory signal. IFN-� was particularly effective in inhibiting inducible IL-12release from monocytes. Unlike IFN-�, IFN-� did not induce release of IFN-� by PBL. Expression of the IFN-� mRNA wasobserved in resting dendritic cells and monocytes, and it was up-regulated by IFN-� stimulation in monocytes, while IFN-� mRNAwas minimally detectable under the same conditions. Monocyte and dendritic cell expression of IFN-� was also confirmed in vivoin chronic lesions of psoriasis vulgaris and atopic dermatitis. Finally, biosensor-based binding kinetic analysis revealed that IFN-�,like IFN- �, binds strongly to heparin (Kd: 2.1 nM), suggesting that the cytokine can be retained close to the local site of production.The pattern of cytokines induced by IFN-� in monocytes, coupled with the unique induction of IFN-� mRNA by IFN- �, indicatesa potential role for IFN- � in the regulation of immune cell functions. The Journal of Immunology, 2002, 169: 4822–4830.
T ype I IFNs are a large family of homologous cytokinesconserved in many species, from fish to reptiles to mam-mals, and were first discovered for their antiviral activity.
Subsequently, it was established that they possess a wide range ofbiological effects including the activation of immune cell popula-tions (1–5). In humans, 12 different IFN-� proteins, one IFN-�,and one IFN-� have been identified. Consequently, there is muchdiscussion on the significance of the existence of so many similarcytokines. The main question at issue is whether their activity isredundant or they are characterized by unique functions (6). Stud-ies of the biological effects of natural IFN subtypes have shownthat their activities can vary greatly (7–10). IFN-�8, for example,is the most potent antiviral protein (11), while IFN-� has a higherpotency in inhibiting the growth of certain tumors and is used inthe treatment of multiple sclerosis (12–14).
We recently reported the identification and preliminary biolog-ical characterization of IFN-�, a 180-aa human cytokine belongingto the type I IFN family (15). The coding sequence of IFN-� doesnot have preferential homology with any of the other family mem-bers found in humans, displaying �30% homology to IFN-�,IFN-�, or IFN-�. It is characterized by the presence of an addi-tional cysteine at the C terminus following the four conservedcysteines, and by an insertion of 12 amino acids between the C andD region. Alignment of the type I proteins shows that IFN-� has anoverall structure typical of the family. Molecular modeling of the
protein displays an organization consisting of five � helices andindicates that two disulfide bonds link the four conserved cysteinesas in the other family members. Although the IFN-� gene is lo-cated on the short arm of chromosome 9, where the other type IIFN genes are located, it resides �6 Mb away from the cluster,suggesting it evolved separately.
A distinguishing feature of IFN-� is the detectable constitutiveexpression of its transcript in uninduced cells, particularly kera-tinocytes (15). The mRNA expression level of IFN-� can be up-regulated by viral infection, by other type I IFNs and, character-istically, by IFN-�. IFN-� binds to the type I IFN receptorcomplex, and such interaction induces signaling events typical ofthe other type I IFNs, such as the activation of the IFN-stimulatedresponse elements. The previous study has also demonstrated thatIFN-� can protect human cells of fibroblastic origin from infectionwith two types of viruses, vesicular stomatitis virus and encepha-lomyocarditis virus.
The current work was undertaken to evaluate the biological ef-fects of IFN-� on cells of the immune system. In particular, be-cause the protein is up-regulated after viral challenge, we havefocused our attention on cells of the innate immunity, which rep-resent the first line of defense in pathological conditions.
Materials and MethodsReagents
Recombinant human IFN-� was expressed and purified from Escherichiacoli, as described (15). The endotoxin level in the preparation was �2EU/mg of protein as determined by the limulus amebocyte lysate method.Recombinant human IFN-�2a and IFN-� purified from E. coli were pur-chased from PBL Biomedical Laboratories (New Brunswick, NJ); IFN-�,IL-2, IL-4, and GM-SCF were obtained from PeproTech (Rocky Hill, NJ);LPS, heparin, chondroitin sulfates A, B, and C, and fucoidan were obtainedfrom Sigma-Aldrich (St. Louis, MO). The neutralizing mAbs 23738.111(anti-IL-10), 1825.121 (anti-TNF-�), and 24822.111 (anti-monocyte
*Human Genome Sciences, Rockville, MD 20850; and †Istituto Dermopaticodell’Immacolata, Rome, Italy
Received for publication December 21, 2001. Accepted for publication September 4,2002.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 Address correspondence and reprint requests to Dr. Bernardetta Nardelli, HumanGenome Sciences, 9410 Key West Avenue, Rockville, MD 20850. E-mail address:[email protected]
The Journal of Immunology
Copyright © 2002 by The American Association of Immunologists, Inc. 0022-1767/02/$02.00
by guest on Novem
ber 28, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

chemoattractant protein (MCP)2-1) were purchased from R&D Systems(Minneapolis, MN). The mAbs specific for IFN-�, 20E07 and 17B09, weregenerated by the fusion of mouse myeloma cells P3 � 63Ag8.653 withsplenocytes from BALB/c mice immunized with rIFN-�.
Cell cultures
Monocytes were obtained from human PBMC by centrifugation of leuka-pheresis preparations (BRT, Baltimore, MD) through Histopaque (Sigma-Aldrich) gradients followed by counterflow centrifugal elutriation. Cellpurity was �90%. Cells were cultured in complete medium consisting ofRPMI 1640 medium (Life Technologies, Rockville, MD) supplementedwith 10% heat-inactivated FBS, 2 mM L-glutamine, and 50 �g/ml genta-micin (Biofluids, Rockville, MD). In some experiments, monocytes wereprepared by magnetic beads separation: PBMC were first depleted of plas-macytoid dendritic cells by direct magnetic labeling with anti-bromodichloroacetate-4 mAb-conjugated microbeads and a high-gradientmagnetic cell sorting device (MiniMACS; Miltenyi Biotec, Auburn, CA).CD14� cells were then isolated by two-step immunomagnetic cell sorting(MACS). Monocytes obtained by this procedure were �95% pure.
Monocyte-derived dendritic cells were obtained by culturing monocytesfor 7–10 days in complete medium supplemented with 1% nonessentialamino acids, 1% sodium pyruvate (Biofluids), 5 � 105 M 2-ME (Sigma-Aldrich), 50 ng/ml GM-CSF, and 20 ng/ml IL-4. Over 90% of the cellsdemonstrated by flow cytometry to express high levels of CD1a and lowlevels of CD14 and showed characteristic dendrite formations on exami-nation with phase-contrast light microscopy.
Cytokine ELISAs
For measurement of the cytokines TNF-�, IL-10, MCP-1, macrophage-inflammatory protein (MIP)-1�, or IFN-�, monocytes (1 � 106 cells/ml),dendritic cells (0.5 � 106 cells/ml), or PBL (2 � 106 cells/ml) were cul-tured for 24 h with complete medium alone or with varying concentrationsof IFNs. For measurement of IL-12 production, monocytes were incubatedfor 18 h with IFN-� (5 ng/ml) in the presence or absence of varying con-centrations of IFNs. LPS (100 ng/ml) was then added and the cells wereincubated for an additional 24 h. Cell-free supernatants from the cultureswere then harvested and stored at �20°C until use. ELISAs were per-formed using kits obtained from BioSource International (Camarillo, CA)according to the manufacturer’s instructions. The ELISAs were comprisedof a specific capture mAb immobilized on plates, a specific biotinylated Abas detector, and streptavidin-conjugated peroxidase. Conversion of the sub-strate by the enzyme was measured at 405 nm. Each value was calculatedas the mean � SE of triplicate samples.
Immunohistochemistry
Punch skin biopsies were obtained after informed consent from healthyadult volunteers (n � 2), and from adult patients with chronic plaque pso-riasis (n � 3) or chronic atopic dermatitis (n � 3). Specimens were em-bedded in OCT compound, frozen in liquid nitrogen, and stored at �80°Cuntil use. Four-micrometer cryostatic sections were fixed in 5% parafor-maldehyde for 5 min, treated with 0.3% hydrogen peroxide to quenchendogenous peroxidase activity, incubated with normal horse serum (Vec-tastain ABC kit; Vector Laboratories, Burlingame, CA) for 20 min, andfinally permeabilized with 0.05% Triton X-100. Single staining was per-formed with mAbs against IFN-�, 20E07, or 17B09, (5 �g/ml), developedusing the avidin-biotin-peroxidase system (Vector Laboratories) and the3-amino-9-ethylcarbazole substrate and counterstained with hematoxylin.Double immunostaining was performed with the same anti-IFN-� mAbsand mAbs anti-CD1a (1:20) (BD PharMingen, San Diego, CA), anti-CD14(1:10), anti-CD3 (BD Biosciences, Mountain View, CA), or anti-FVIII/vWF (1:25) (DAKO, Glostrup, Denmark), using the avidin-biotin-peroxi-dase or avidin-biotin-alkaline phosphatase system together with the chro-mogens 3-amino-9-ethylcarbazole and Blue Vector (Vector Laboratories)to reveal the peroxidase and alkaline phosphatase activity, respectively. Nocounterstain was applied. As negative control, primary mAbs were omittedor replaced with an irrelevant isotype-matched mAb.
Heparin-binding ELISA
ELISA microplates (Maxisorp; Nalge Nunc International, Naperville, IL)were coated with heparin-BSA conjugate (Sigma-Aldrich) at 5 �g/ml inbicarbonate buffer (0.05 M sodium bicarbonate, pH 9.5) overnight at 4°Cand then blocked for 1 h with 1% BSA (Sigma-Aldrich) in the same buffer.Serial dilutions of IFN-� (250 ng/ml) were added to the wells in diluent
buffer (PBS containing 0.1% Tween 20 and 1% BSA) for 2 h at roomtemperature. Three sequential steps followed with intervening wash stepswith PBS 0.1% Tween. First, detection of bound IFN-� was achieved byadding an affinity purified biotinylated rabbit polyclonal anti IFN-�-spe-cific Ab at 0.125 �g/ml for 1 h at room temperature. Peroxidase strepta-vidin (Vector Laboratories) was then added at a dilution of 1/2000 in di-luent buffer for 1 h. Finally, tetramethylbenzidine substrate (Kirkegaard &Perry, Gaithersburg, MD) was added and the incubation was continued for20 min. Plates were read at 450 nm after stopping the reaction with sulfuricacid. In some experiments, a competitive ELISA was used in which IFN-�(83.3 ng/ml) was preincubated for 2 h in the absence or presence of varyingconcentrations of soluble competitors before the addition of the mixture tothe heparin-BSA coated plates.
BIAcore analysis
The technology uses surface plasmon resonance to measure changes inrefractive index when a ligand binds to an immobilized moiety. Heparin-binding kinetic analysis was performed on a BIAcore 3000 (BIAcore, Pis-cataway, NJ). Two flow cells (high and low density surfaces) of a CM5chip (BIAcore AB) were activated with a mixture of 0.4 M N-ethyl-N-(3dimethyl-aminopropyl) carbo-diimide and 0.1 M N-hydroxysuccinimide atpH 8.0, after which streptavidin (Sigma-Aldrich) in 10 mM acetate buffer,pH 5.0, was injected. Unreacted groups were blocked with injection of 1 Methanolamine at pH 9.0, followed by a single injection of 10 mM HCl.Biotinylated heparin-BSA (Sigma-Aldrich), resuspended in HBS-EP buffer(0.01 M HEPES, 0.15 M NaCl, 3 mM EDTA, and 0.005% surfactant P20,pH 7.4) with 0.2 M NaCl, was flowed over the surface to allow bindingonto the streptavidin surface. Both flow cells were then conditioned withtwo wash steps with 10 mM HCl. The high and low density flow cells hadnet densities of 1000 and 300 resonance units, respectively. Kd determi-nation was over a low density flow cell (16). For kinetic analysis, IFN testsamples were dialyzed in HBS-EP and flowed over a low density surfaceat 25°C and a flow rate of 15 �l/min. The complexes were washed at 15�l/min with HBS-EP to study the dissociation phase. The surface wasregenerated using 10 mM glycine-HCl, pH 1.5, in the presence of 1.5 MNaCl. Background binding was subtracted by flowing the sample over ablank flow cell. For each IFN subtype, a complete set of sensorgrams wasrecorded with a series of different concentrations (range � 5–0.31 �g/ml).Sets of sensorgrams were analyzed with BIAevaluation Software, version3.1 (BIAcore), provided with the instrument.
Quantitative PCR analysis
Total RNA (50 ng) was used in a one-step, 50 �l, quantitative RT-PCR. Asa control for genomic contamination, parallel reactions were set up withoutreverse transcriptase. The abundance of specific mRNAs was measuredrelative to 18S rRNA using the Applied Biosystems Prism 7700 SequenceDetection System. Reactions were conducted at 48°C for 30 min, 95°C for10 min, followed by 40 cycles of 95°C for 15 s, 60°C for 1 min. Reactionswere performed in triplicate. Using Primer Express Software (AppliedBiosystems, Foster City, CA), primer and probe sets were designed to targetthe following human sequences, where the gene named is followed by thesequences of the two primers and then the probes: IFN-� (GCCCCAAGAGTTTCTGCAATAC, GGCCTGTAGGGACATTTCATAGA, CAACCTATGAAGAGGGACATCAAGAAGGCC); IFN-� (GACATCCCTGAGGAGATTAAGCA,CTGGAGCATCTCATAGATGGTCAA, CAGTTCCAGAAGGAGGACGCCGC); 18S (CGGCTACCACATCCAAGGAA, GCTGGAATTACCGCGGCT, TGCTGGCACCAGACTTGCCCTC).
Probes were labeled at the 5-end with the reporter dye 6-FAM and onthe 3-end with the quencher dye TAMRA (BioSource International). ThemRNA level of IFN-� or IFN-� in each sample, determined in triplicate,are expressed as the mean level and SD relative to the 18S ribosomal RNAlevel (endogenous reference).
Statistical analysis
The statistical significance of differences among groups was analyzed usingthe unpaired, two-tailed Mann-Whitney test.
ResultsInduction of cytokine release in monocytes
The production of cytokines by phagocytic cells at the onset ofinfections initiates a series of events leading to the activation of theother effector populations of the immune system. In our study, weexamined the effect of IFN-�, IFN-�2a and IFN-� on cytokinerelease by human monocytes. The other two IFNs were chosenbecause they are subtypes currently in clinical use (17, 18) and are
2 Abbreviations used in this paper: MCP, monocyte chemoattractant protein; MIP,macrophage-inflammatory protein; IFNAR, IFN-� receptor.
4823The Journal of Immunology
by guest on Novem
ber 28, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

highly effective in inhibiting viral infection and cell proliferation.Monocytes were purified from peripheral blood and cultured at aconcentration of 1 � 106 cells/ml for 1 day in the presence ofrecombinant IFN-�, IFN-�2a, or IFN-� (Fig. 1). Both IFN-� andIFN-� were able to induce release of TNF-� even in the absenceof a costimulus, such as IFN-� that is often required in this ex-perimental system. In contrast, IFN-�2a was ineffective in pro-moting TNF-� release. To avoid that plasmacytoid dendritic cellspresent in the elutriated monocytes (in average �1%) would ac-count for the observed cytokine release, in several experimentsmonocytes were purified using magnetic separation. Plasmacytoiddendritic cells were first completely removed from the PBMC us-ing blood dendritic cell Ag-4 mAb-conjugated microbeads (19)
followed by immunomagnetic enrichment of CD14� cells. Theresulting cell preparations (�95% monocytes) released TNF-� fol-lowing stimulation with IFN-� and IFN-�, as observed with thedonor reported in Fig. 1B. Cytokine release following IFN-� in-cubation was consistently observed in all donors tested (n � 10);however, the concentration of cytokine detected in the conditionedmedia was donor-dependent. For example, following treatmentwith 100 ng/ml, cells from the donors reported in Figs. 1A and 2Areleased 166 and 423 ng/ml of TNF-�, respectively. IFN-�strongly synergized with IFN-� to stimulate TNF-� release whenthe two factors were simultaneously added to the cultures (Fig.2B). In contrast, IFN-� did not increase LPS-induced cytokine re-lease (Fig. 2C). Chemokines, such as MCP-1 and MIP-1�, werefound to be up-regulated in IFN-�-stimulated monocytes (Table I).Interestingly, IL-10 production was strongly induced by the novelIFN, with an increase up to 40-fold.
Inhibition of inducible IL-12 release in monocytes
Among cytokines produced by monocytes, IL-12 plays an impor-tant role in immunoregulation, being an inducer of IFN-� and agenerator of Th1 T cell responses. IL-12 production is thereforenormally tightly regulated. The ability of IFN-� to modulate itsrelease was examined. To obtain release of IL-12 from humanmonocytes, the stimulation protocol described by Hayes et al. (20)was used. Freshly isolated monocytes were cultured overnight withIFN-� in the presence of either IFN-� or IFN-�, with LPS thenadded to the cultures for an additional 24 h. In the donor reportedin Fig. 3A, cell stimulation induced the release of 2231 pg/ml ofIL-12, which was inhibited by IFN-� in a concentration-dependentmanner. A consistent pattern of inhibition of inducible IL-12 re-lease was observed in all the donors (n � 7), and the level ofreduction observed with IFN-� was greater than that observed withIFN-�.
The ability of IFN-� treatment to suppress IL-12 release is de-pendent on IL-10 release because it could be blocked by an IL-10-specific mAb (Fig. 3B and Table II). To determine whetherother cytokines produced by monocytes, such as TNF-� or
FIGURE 1. Induction by IFN-� of TNF-� release from monocytes.Monocytes were treated with varying concentrations of IFN-�, IFN-�2a, orIFN-� (ng/ml) for 1 day; conditioned media were collected and assayed forTNF-� content by ELISA. A, Monocytes from healthy donors were puri-fied by centrifugal elutriation; representative experiment (one donor of sev-en). B, Monocytes were purified by magnetic separation; representativeexperiment (one donor of three).
FIGURE 2. Synergistic effect of IFN-� on IFN-�-induced TNF-� release from monocytes. Monocytes were treated with IFN-� (ng/ml) alone or incombination with IFN-� (5 ng/ml) or LPS (100 ng/ml). TNF-� content in the media, conditioned for 24 h, was analyzed by ELISA. One representativeexperiment is shown (one of three).
Table I. Cytokine release from IFN-�-treated monocytesa
IL-10 MCP-1 MIP-1�
Untreated 5 � 0.0 203 � 0.6 1130 � 0.1IFN-� 100 204 � 0.2 1389 � 205 3020 � 0.1IFN-� 30 95 � 1.1 687 � 0.2 1480 � 0.1IFN-� 10 39 � 1.0 372 � 10 1240 � 0.0
a Monocytes (1 � 106/ml) were treated with IFN-� nanograms per milliliter for 1 day.Conditioned media from the cell cultures were then collected and analyzed by ELISA forcytokine contents picograms per milliliter; one representative experiment of three.
4824 IFN-� AND CELLS OF THE INNATE IMMUNE SYSTEM
by guest on Novem
ber 28, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

MCP-1, participate in the observed inhibitory activity (21, 22),neutralizing mAbs against TNF-� or MCP-1 were added to theexperimental system (Table II). Anti-IL-10 mAb completely abol-ished the inhibitory effect of 10 ng/ml IFN-� and partially inhibitedthe effect of 100 ng/ml. Anti-TNF-� or MCP-1 mAbs partiallyblocked the inhibitory effect of 10 ng/ml IFN-�, but did not haveany effect in neutralizing the activity of 100 ng/ml IFN-�. Whenthe neutralizing mAbs against TNF-� and MCP-1 were used to-gether with the anti-IL-10 mAb to block the activity of 100 ng/mlIFN-�, no additive neutralizing effect was observed.
Effect on dendritic cells
IFN-� activity was then analyzed on dendritic cells. As with mono-cytes, a direct stimulation of cytokine release was mediated byIFN-� (Fig. 4). Significant levels of TNF-� and IL-10 were de-tected in the supernatants following culture for 1 day. In addition,IFN-�, like IFN-�, enhanced the expression of Ags correlated withactivation and increased Ag-presenting capacity, such as MHCclass II and CD54 (data not shown). Contrary to the experimentswith monocytes, we did not observe inhibition of IL-12 productionfrom LPS-stimulated dendritic cells (data not shown). Interest-
ingly, McRae et al. (23) have reported that, on dendritic cells, typeI IFNs inhibited IL-12 production induced by CD40 ligation, butnot by LPS stimulation.
Effect on IFN-� release
Having established a modulatory activity of IFN-� on cytokinerelease in monocytes, we explored the effect of type I IFNs on
FIGURE 3. Inhibition by IFN-� of IL-12 release from monocytes. A,Comparison of the effect of IFN-� and IFN-�. Human monocytes weretreated with IFN-� (5 ng/ml) and with IFN-� or IFN-� (ng/ml). After 16 hof incubation, cells were stimulated by the addition of LPS (100 ng/ml).Following additional 24 h in culture, conditioned media were collected andassayed for IL-12 content by ELISA. A representative experiment (one ofseven) is shown. B, IFN-�-induced inhibition is IL-10-dependent. The neu-tralizing anti-IL-10 mAb 23738.111 (1 �g/ml) was added to the cultures to-gether with the IFNs. One representative experiments is shown (one of three).
FIGURE 4. Induction by IFN-� of cytokine release from dendritic cells.Dendritic cells (5 � 105/ml) were treated for 1 day with IFN-�. Condi-tioned media were collected and assayed for TNF-� or IL-10 content byELISA. Representative experiment of four.
Table II. Effect of cytokine-specific Abs on IFN-�-induced inhibition ofIL-12 releasea
IFN-� (ng/ml) Neutralizing mAb Specificity IL-12 (pg/ml)
268 � 2100 2 � 010 136 � 1100 IL-10 58 � 110 IL-10 333 � 9100 TNF-� 2 � 010 TNF-� 95 � 1100 MCP-1 2 � 010 MCP-1 112 � 1100 IL-10 � TNF-� 51 � 010 IL-10 � TNF-� 265 � 1100 IL-10 � TNF-� � MCP-1 66 � 110 IL-10 � TNF-� � MCP-1 332 � 0
a Monocytes (1 � 106/ml) were primed with IFN-� (5 ng/ml) for 18 h and thenstimulated with LPS (100 ng/ml). IFN-� and the mAbs (1 �g/ml) were added to thecultures together with IFN-�. IL-12 release was evaluated in the culture supernatants1 day post LPS stimulation; one representative experiment of two.
4825The Journal of Immunology
by guest on Novem
ber 28, 2013http://w
ww
.jimm
unol.org/D
ownloaded from
IFN-� release from PBL (Fig. 5A). In four donors, IFN-� induceda strong dose-dependent release of IFN-�, while IFN-� produceda minimal effect only at 300 ng/ml ( p � 0.001 IFN-� vs IFN-�treatment). To define the population responsible for the release ofIFN-� following stimulation with the two type I IFNs, PBL fromfour additional donors were depleted of T cells using CD3-conju-gated magnetic beads (following separation, T cell content �2%,NK content �30%). The resulting cell populations enriched forNK cells were stimulated with IFN-� or IFN-�. As observed in theprevious experiments, IFN-� strongly induced IFN-� release andIFN-� did not have effect (Fig. 5B). However, IFN-� was able toincrease NK cytotoxic activity of PBL, although at a lower levelthan IFN-� or IFN-�2a (not shown).
IFN-� expression in myeloid cells
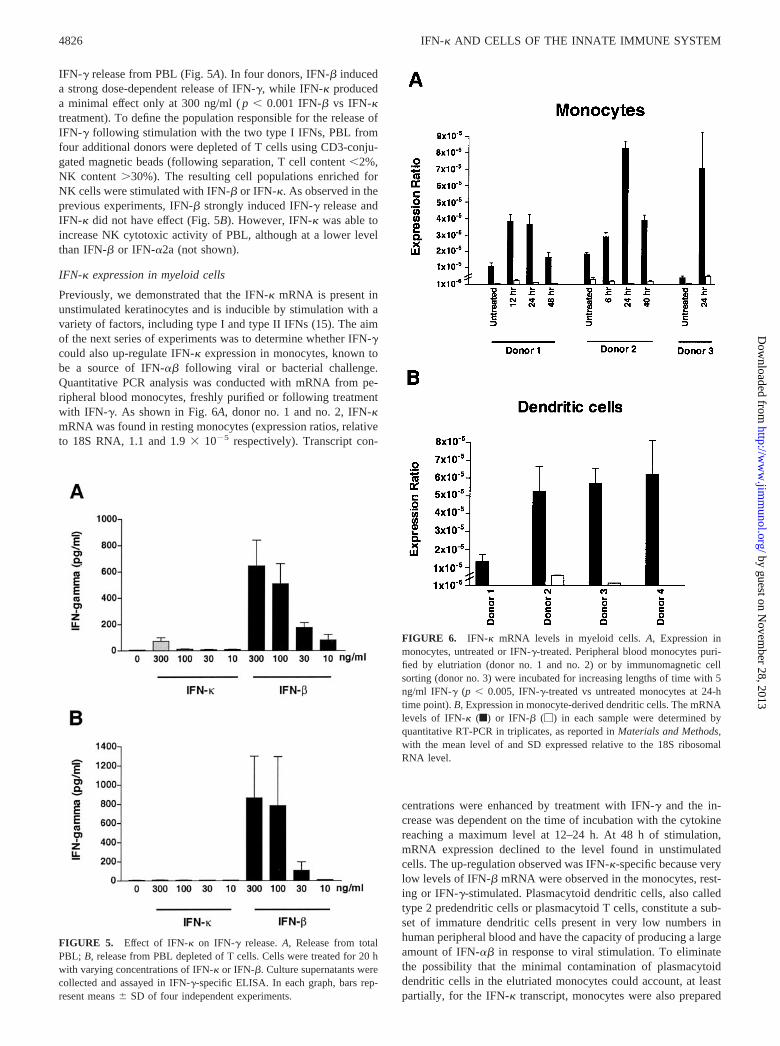
Previously, we demonstrated that the IFN-� mRNA is present inunstimulated keratinocytes and is inducible by stimulation with avariety of factors, including type I and type II IFNs (15). The aimof the next series of experiments was to determine whether IFN-�could also up-regulate IFN-� expression in monocytes, known tobe a source of IFN-�� following viral or bacterial challenge.Quantitative PCR analysis was conducted with mRNA from pe-ripheral blood monocytes, freshly purified or following treatmentwith IFN-�. As shown in Fig. 6A, donor no. 1 and no. 2, IFN-�mRNA was found in resting monocytes (expression ratios, relativeto 18S RNA, 1.1 and 1.9 � 10�5 respectively). Transcript con-
centrations were enhanced by treatment with IFN-� and the in-crease was dependent on the time of incubation with the cytokinereaching a maximum level at 12–24 h. At 48 h of stimulation,mRNA expression declined to the level found in unstimulatedcells. The up-regulation observed was IFN-�-specific because verylow levels of IFN-� mRNA were observed in the monocytes, rest-ing or IFN-�-stimulated. Plasmacytoid dendritic cells, also calledtype 2 predendritic cells or plasmacytoid T cells, constitute a sub-set of immature dendritic cells present in very low numbers inhuman peripheral blood and have the capacity of producing a largeamount of IFN-�� in response to viral stimulation. To eliminatethe possibility that the minimal contamination of plasmacytoiddendritic cells in the elutriated monocytes could account, at leastpartially, for the IFN-� transcript, monocytes were also prepared
FIGURE 5. Effect of IFN-� on IFN-� release. A, Release from totalPBL; B, release from PBL depleted of T cells. Cells were treated for 20 hwith varying concentrations of IFN-� or IFN-�. Culture supernatants werecollected and assayed in IFN-�-specific ELISA. In each graph, bars rep-resent means � SD of four independent experiments.
FIGURE 6. IFN-� mRNA levels in myeloid cells. A, Expression inmonocytes, untreated or IFN-�-treated. Peripheral blood monocytes puri-fied by elutriation (donor no. 1 and no. 2) or by immunomagnetic cellsorting (donor no. 3) were incubated for increasing lengths of time with 5ng/ml IFN-� (p � 0.005, IFN-�-treated vs untreated monocytes at 24-htime point). B, Expression in monocyte-derived dendritic cells. The mRNAlevels of IFN-� (f) or IFN-� (�) in each sample were determined byquantitative RT-PCR in triplicates, as reported in Materials and Methods,with the mean level of and SD expressed relative to the 18S ribosomalRNA level.
4826 IFN-� AND CELLS OF THE INNATE IMMUNE SYSTEM
by guest on Novem
ber 28, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

by immunomagnetic cell sorting. PBMC were completely depletedof BDCA-2� cells (24) and then were positively selected for CD14binding. Monocytes obtained from three donors showed constitu-tive levels of IFN-� (mean expression ratio � 2.7 � 10�5 � 1.8)and it was inducible by IFN-� treatment. A representative donor(donor no. 3) is shown in Fig. 6A.
IFN-� mRNA was also found in monocyte-derived dendriticcells (mean expression ratio � 4.6 � 10�5 � 2.2 in four donors),while IFN-� mRNA was minimally detectable (mean expressionratio � 0.3 � 10�5 � 0.3) in two of four donors (Fig. 6B). How-ever, in this cell type treatment with IFN-� did not significantlyenhance the level of the transcript (not shown).
Expression of IFN-� in inflamed human tissues
To test whether IFN-� is expressed in vivo at inflammatory sitesrich in monocytes and dendritic cells, skin lesions from patientswith psoriasis or atopic dermatitis were assayed for IFN-� immu-noreactivity. Normal skin from healthy subjects showed only scat-tered IFN-�-positive cells in the dermis (Fig. 7A). In contrast, nu-merous IFN-�-positive cells were observed in sections frompsoriasis and atopic dermatitis, in both the epidermis and dermis(Fig. 7, B and C). In particular, double-staining showed that thecells expressing IFN-� were mostly CD14� monocytes and part ofCD1a� dendritic cells in both diseases (Fig. 7, D and E, and G andH). IFN-� was also expressed by basal and suprabasal keratino-cytes, with the positivity localized along the cell membrane ofthese cells, in their cytoplasm and in the intercellular spaces. Fi-nally, anti-IFN-� mAb reacted with the majority of FVIII� venuleslocalized in both the superficial and reticular dermis but not withCD3� T lymphocytes (Fig. 7, F and I).
IFN-� is a heparin-binding protein
The intriguing observation of IFN-� constitutive expression in ker-atinocytes (15) suggests that this IFN may have a local function inthe epidermis. To determine whether IFN-� can be retained at itssite of production through interaction with the glycosaminoglycanspresent in many tissues, we developed an ELISA-based assay tomeasure IFN-� binding to heparin. As shown in Fig. 8A, dose-
FIGURE 7. In vivo detection of IFN-� in in-flamed human skin. Single-staining for IFN-� (red)was performed on normal skin (A) and psoriatic (B)or atopic dermatitis (C) lesions. Double-stainingshows that cells positive for IFN-� (red) are CD1a�
dendritic cells (blue) (D and E, psoriasis and atopicdermatitis, respectively) and CD14� monocytes(blue) (G and H, psoriasis and atopic dermatitis, re-spectively). Anti-IFN-� Ab also stains most of theFVIII� dermal microvessels (I) but not CD3� Tlymphocytes (F) (blue) in psoriatic skin. A mem-brane and cytoplasmic immunoreactivity for IFN-�is also present in basal and suprabasal keratinocytes.Arrows in D and E and G and H indicate the IFN-�/CD1a and IFN-�/CD14 double-positive cells, re-spectively; representative experiment shown. Thestaining and number of donors are as reported inMaterials and Methods.
FIGURE 8. IFN-� is a heparin-binding protein. A, Binding of rIFN-� toimmobilized heparin-BSA complex. Increasing concentrations of IFN-� wereadded to ELISA plate wells coated with 5 �g/ml heparin-BSA; binding wasdetected by biotinylated rabbit polyclonal anti-IFN-� Ab. B, Inhibition of bind-ing by glycosaminoglycans. Before addition to the heparin-BSA-coated plate,IFN-� was incubated in absence or presence of 1 �g/ml heparin, chondroitinsulfate A (CSA), chondroitin sulfate B (CSB), chondroitin sulfate C (CSC), orfucoidan. Absorbance was measured after a 20-min incubation with substrate.The experiment was performed twice with similar results.
4827The Journal of Immunology
by guest on Novem
ber 28, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

dependent binding of IFN-� to the heparin-BSA complex was ob-served. The binding is detected with high sensitivity because it wasreadily observed with as little as 1 ng of IFN-�. In competitionexperiments, heparin and fucoidan, a nonmammalian polysaccha-ride, strongly inhibited interaction, while chondroitin sulfate A andB were very weak inhibitors, and chondroitin sulfate C was inef-fective (Fig. 8B). Surface plasmon resonance (BIAcore analysis)allowed the determination of the affinity binding. Biotinylated hep-arin-BSA was immobilized on a streptavidin-activated sensor chipand then IFN-� was injected over the heparin-BSA surface. Atypical sensorgram was obtained (Fig. 9A) with an associationphase and, when IFN-� was replaced by running buffer alone, adissociation phase. Similar sensorgrams were obtained with IFN-�(Fig. 9B) and IFN-� (not shown). The heparin-binding affinity val-ues of the two type I IFNs were similar: 2.1 nM and 1.4 nM forIFN-� and IFN-�, respectively. On the contrary, IFN-�2a showeda minimal binding to heparin (not shown). The affinity value forIFN-�, 1.1 nM, was within the range of previously published re-sults (25).
In summary, we have established for the first time, to our knowl-edge, that two type I IFNs, IFN-� and IFN-�, are heparin-bindingproteins. IFN-� binding is specific because no interaction withother glycosaminoglycans, such as chondroitin sulfate C, was ob-served. The strong competing effect of fucoidan was observed also
in other studies of heparin-binding proteins and is believed to be dueto the highly sulfated and branched structure of the polymer (26).
DiscussionIn this study, we have extended the biological characterization ofIFN-�, a newly identified subclass of human type I IFN. The func-tional profile of the cytokine resembles that of IFN-� more thanthat of IFN-�2a, one of the IFN-� subclasses. Like IFN-� but incontrast to IFN-�2a, IFN-� is able to directly modulate cytokinerelease from monocytes and dendritic cells, and to strongly bindheparin. However, IFN-� did not stimulate IFN-� release from NKcells.
Two distinct receptor subunits for type I IFNs have been iden-tified, IFN-� receptor (IFNAR)-1 and IFNAR-2 (reviewed in Refs.27 and 28). IFNAR-1 has an important role in signal transduction,whereas IFNAR-2 seems to be the major ligand-binding compo-nent. Although all the type I proteins, including IFN-� (15), bindto the same receptor complex, there is much experimental evidencedemonstrating that the interactions with the receptor or the induc-tion of signaling pathways can vary, thus influencing the biologicalresponses (9, 29–32). The large differences observed in the im-munological activities of IFN subspecies are suggestive of specificfunctions characterizing the different cytokines (7). Our results in-dicate that IFN-�2a does not have the same modulatory activity onmonocytes that IFN-� and IFN-� do. In addition, the effect ofIFN-� on monocytes, qualitatively similar to that of IFN-�, raisesthe possibility that this novel IFN shares with IFN-� the bindingsites and/or the signaling pathways regulating the monocyticresponse.
Of particular interest is the ability of IFN-� to mediate a strongincrease in IL-10 release and a concomitant inhibition of inducibleIL-12 release. The decrease in IL-12 level was found to be IL-10-dependent, as demonstrated by the blocking activity of an IL-10-neutralizing Ab. There are conflicting reports of the role of type IIFNs on IL-12 production. IFN-� and IFN-� were found to in-crease IL-12 mRNA in human macrophages (4) and up-regulatethe expression of the IL-12R �2 chain in CD4� T cells (33). How-ever in other studies, type I IFNs suppressed IL-12 production ina mixed leukocyte population and in dendritic cells (23, 34). Fur-ther support for an inhibitory role of class I IFNs comes from invivo studies. In virally infected mice, endogenously expressedIFN-�� inhibited the ability of LPS to induce IL-12, while neu-tralization of IFN-�� enhanced IL-12 and IFN-� production (34).Lastly, Wang et al. (35) demonstrated that IFN-� inhibits IL-12production in Staphylococcus aureus Cowan- plus IFN-�-stimu-lated human PBMC in an IL-10-dependent mechanism. In thisstudy, the authors did not observe enhancement by IFN-� of basalIL-10 production in monocytes and suggested that the IFN-� effectwas mediated by inhibition of IFN-� priming. In our experiments,IFN-� directly induced IL-10 release from monocytes in the ab-sence of other stimulatory factors, such as LPS/S. aureus Cowanor IFN-�.
Our findings suggest a potential therapeutic use of IFN-� forautoimmune disorders in which the pathogenesis is linked to anoverproduction of IL-12, including multiple sclerosis, rheumatoidarthritis, insulin-dependent diabetes, and experimental colitis. Ithas been shown that decreased levels of IL-10 and increased IL-12mRNA in cells of multiple sclerosis patients are associated withdisease progression (36). IFN-� is, indeed, an approved therapy forthis disease and was found able to slow disease progression andreduce relapses (13, 14). IFN-� might also be beneficial in thetreatment of inflammatory syndromes, such as sepsis, where IL-10plays a down-regulatory role and IL-12 has the opposing effect
FIGURE 9. BIAcore analysis of type I IFN binding to immobilized hep-arin. The sensorgrams show the on- and off-rate regions of varying con-centrations of IFN-� (A) or IFN-� (B), as described in Material and Meth-ods. The binding response in resonance units was recorded as a function ofthe time. Kd values were calculated using the BIAevaluation kinetic soft-ware program. The experiment was performed twice with similar results.
4828 IFN-� AND CELLS OF THE INNATE IMMUNE SYSTEM
by guest on Novem
ber 28, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

(37, 38). These potential therapeutic indications are further sup-ported by the very low stimulatory effect of IFN-� on IFN-� re-lease from PBMC. IFN-� is a principal player in the regulation ofboth autoimmune and inflammatory disorders, thus a low level ofIFN-� secretion might benefit such disease states.
Another interesting characteristic of IFN-�, shared with IFN-�,is the high-affinity interaction with heparin. Numerous growth fac-tors, chemokines, and cytokines bind to heparin or heparan sul-fates, including IFN-�, TGF�-1, IL-8, IL-2, the p40 subunit ofIL-12, and IL-10 (25, 39–43). Due to the highly acidic property ofheparin, it is believed that clusters of basic amino acids in thecytokine sequences promote the interaction with glycosaminogly-cans (44). The sequence of IFN-� does not display clusters ofarginines and lysines in the primary sequence. However, it is pos-sible that the folding of the protein in the secondary structurebrings into proximity discontinuous basic residues. For example, asingle amino acid mutation in the sequence of MIP-1� abolishedthe glycosaminoglycan binding activity of the chemokine (45).The binding of the two cytokines, IFN-� and IFN-�, to polysac-charides is likely to keep them close to the production site and tostimulate antiviral and immune responses focused at the local siteof infection.
A distinct property of IFN-� is that its transcript is readily in-duced by type II IFN. Monocytic cells are among the main pro-ducers of type I IFNs following viral challenge or stimulation withbacterial endotoxin. Although secretion of IFN-� was observed inmurine peritoneal macrophages treated with IFN-� (46), it wasshown that freshly isolated human monocytes did not produce typeI IFNs in response to single IFN-� or LPS stimulation, but neededto be primed by either IFN-� or GM-CSF (47). In agreement withthese results, we have demonstrated by quantitative RT-PCR anal-ysis that human monocytes and dendritic cells have a very low orundetectable level of IFN-� mRNA that is not inducible by IFN-�treatment alone. On the contrary, a significantly higher constitutivelevel of IFN-� mRNA was observed in the cells. Moreover, theexpression was inducible in monocytes by IFN-� treatment. Thepresence of IFN-� in monocytes and dendritic cells was confirmedin vivo in the skin affected by psoriasis or atopic dermatitis.
In conclusion, our findings suggest that IFN-� may play a rolein the regulation of systemic or local immune functions through itseffect on cells of the innate immune system. The enhancement ofIFN-� expression by IFN-� could be a negative feedback mecha-nism to suppress further activation of T lymphocytes and NK cellsvia enhancement of IL-10 and inhibition of IL-12 release in ac-cessory cells. To completely define the function of IFN-� in theimmune system, the effect of this cytokine on the cells regulatingadaptive immunity needs to be characterized.
AcknowledgmentsWe thank Drs. Tim A. Coleman and Guoxian Chen for providing the re-combinant IFN-� and Dr. Viktor Roschke for providing the mAbs anti-IFN-�. We thank Tatiana Tsareva, Don Mather, and Victor Kao fortechnical help.
References1. Roberts, R. M., L. Liu, Q. Guo, D. Leaman, and J. Bixby. 1998. The evolution
of type I interferons. J. Interferon Cytokine Res. 18:805.2. Grander, D., O. Sangfelt, and S. Erickson. 1997. How does interferon exert its
cell growth inhibitory effect? Eur. J. Haematol. 59:129.3. Leaman, D. W. 1998. Mechanism of interferon action. Prog. Mol. Subcell. Biol.
20:101.4. Belardelli, F., and I. Gresser. 1996. The neglected role of type I interferon in the
T cell response: implications for its clinical use. Immunol. Today 17:369.5. Marrack, P., J. Kappler, and T. Mitchell. 1999. Type I interferons keep activated
T cells alive. J. Exp. Med. 189:521.6. Foster, G. R., and N. B. Finter. 1998. Are all type I human interferons equivalent?
J. Viral. Hepat. 5:143.
7. Fish, E., K. Banerjee, and N. Stebbing. 1983. Human leukocyte interferon sub-types have different antiproliferative and antiviral activities on human cells. Bio-chem. Biophys. Res. Commun. 112:537.
8. Runkel, L., L. Pfeffer, M. Lewerenz, D. Monneron, C. H. Yang, A. Murti,S. Pellegrini, S. Goelz, G. Uze, and K. Mogensen. 1998. Differences in activitybetween � and � type I interferons explored by mutational analysis. J. Biol.Chem. 273:8003.
9. Hibbert, L., and G. R. Foster. 1999. Human type I interferons differ greatly intheir effects on the proliferation of primary B cells. J. Interferon Cytokine Res.19:309.
10. Pestka, S. 1997. The human interferon-� species and hybrid proteins. Semin.Oncol. 24(Suppl. 9):4.
11. Foster, G., O. Rodrigues, F. Ghouze, E. Schulte-Frohlinde, D. Testa, M. J. Liao,G. R. Stark, L. Leadbeater, and H. C. Thomas. 1996. Different relative activitiesof human cell derived interferon-� subtypes: IFN-�8 has very high antiviralpotency. J. Interferon Cytokine Res. 16:1027.
12. Rosenblum, M. G., W. K. Yung, P. J. Kelleher, F. Ruzicka, P. A. Steck, andE. C. Borden. 1990. Growth inhibitory effects of interferon-� but not interferon-�on human glioma cells: correlation of receptor binding, 2,5-oligoadenylate syn-thetase and protein kinase activity. J. Interferon Res. 10:141.
13. The IFN�-1b Multiple Sclerosis Study Group. 1993. Interferon �-1b is effectivein relapsing-remitting multiple sclerosis. Neurology 43:655.
14. Jacobs, L. D., D. L. Cookfair, R. A. Rudick, R. M. Herdon, J. R. Richert,A. M. Salazar, J. S. Fischer, D. E. Goodkin, C. V. Granger, J. H. Simon, et al.1996. Intramuscular IFN �-1a for disease progression in relapsing multiple scle-rosis. Ann. Neurol. 39:285.
15. La Fleur, D. W., B. Nardelli, T. Tsareva, D. Mather, P. Feng, M. Semenuk,K. Taylor, M. Buergin, D. Chincilla, V. Roschke, et al. 2001. Interferon-�, anovel type I IFN expressed in human keratinocytes. J. Biol. Chem. 276:39765.
16. Myszka, D. G. 1999. Improving biosensor analysis. J. Mol. Recognit. 12:279.17. Pfeffer, L. M., C. A. Dinarello, R. B. Herberman, B. R. Williams, E. C. Borden,
R. Bordens, M. R. Walker, T. L. Nagabhushan, P. P. Trotta, and S. Pestka. 1998.Biological properties of recombinant �-interferons: 40th anniversary of the dis-covery of interferons. Cancer Res. 58:2489.
18. Rudick, R. A., R. M. Ransohoff, J. C. Lee, R. Peppler, M. Yu, P. M. Mathisen,and V. K. Tuohy. 1998. In vivo effects of interferon �-1a on immunosuppressivecytokines in multiple sclerosis. Neurology 50:1294.
19. Dzionek, A., A. Fuchs, P. Schmidt, S. Cremer, M. Zysk, S. Miltenyi, D. W. Buck,and J. Schmitz. 2000. BDCA-2, BDCA-3, and BDCA-4: three markers for dis-tinct subsets of dendritic cells in human peripheral blood. J. Immunol. 165:6037.
20. Hayes, M. P., J. Wang, and M. A. Norcross. 1995. Regulation of IL-12 expressionin human monocytes: selective priming by IFN-� of LPS-inducible p35 and p40genes. Blood 86:646.
21. Ma, X., J. Sun, E. Papasavvas, H. Riemann, S. Roberton, J. Marshall,R. T. Bailer, A. Moore, R. P. Donnelly, G. Trinchieri, and L. J. Montaner. 2000.Inhibition of IL-12 production in human monocyte-derived macrophages byTNF. J. Immunol. 165:1722.
22. Chensue, S. W., K. S. Warmington, J. H. Ruth, P. S. Sanghi, P. Lincoln, andS. L. Kunkel. 1996. Role of monocyte chemoattractant protein-1 (MCP-1) in Th1(mycobacterial) and Th2 (schistosomal) antigen-induced granuloma formation.J. Immunol. 157:4602.
23. McRae, B. J., B. A. Beilfuss, and G. A. van Seventer. 2000. IFN-� differentiallyregulates CD40-induced cytokine secretion by human dendritic cells. J. Immunol.164:23.
24. Dzionek, A., Y. Sohma, J. Nagafune, M. Cella, M. Colonna, F. Facchetti,G. Gunther, I. Johnson, A. Lanzavecchia, T. Nagasaka, et al. 2001. BDCA-2, anovel plasmacytoid dendritic cell-specific type II C-type lectin, mediates antigencapture and is a potent inhibitor of interferon �/� induction. J. Exp. Med. 194:1823.
25. Lortat-Jacob, H., and J. A. Grimaud. 1992. Binding of IFN-� to heparan sulfateis restricted to the heparin-like domains and involves carboxylic- but not N-sulfated groups. Biochim. Biophys. Acta 1117:126.
26. Patankar, M. S., S. Oehninger, T. Barnett, R. L. Williams, and G. F. Clark. 1993.A revised structure for fucoidan may explain some of its biological activities.J. Biol. Chem. 268:21770.
27. Pestka, S. 1997. The interferon receptors. Semin. Oncol. 24(Suppl. 9):18.28. Mogensen, K. E., M. Lewerenz, J. Reboul, G. Lutfalla, and G. Uze. 1999. The
type I interferon receptor: structure, function and evolution of a family business.J. Interferon Cytokine Res. 19:1069.
29. Hu, R., J. Bekisz, M. Hayes, S. Audet, J. Beeler, E. Petricoin, and K. Zoon. 1999.Divergence of binding, signaling and biological responses to recombinant humanhybrid IFN. J. Immunol. 163:854.
30. Petricoin, E. F., S. Ito, B. L. Williams, S. Audet, L. F. Stancato, A. Gamero,K. Clouse, P. Grimley, A. Weiss, J. Beeler, et al. 1997. Antiproliferative actionof interferon-� requires components of T-cell receptor signaling. Nature 390:629.
31. Domanski, P., O. W. Nadeau, L. C. Platanias, E. Fish, M. Kellum, P. Pitha, andO. R. Colamonici. 1998. Differential use of the �L subunit of the type I interferon(IFN) receptor determines signaling specificity for IFN-�2 and IFN-�. J. Biol.Chem. 273:3144.
32. Lewerenz, M., K. E. Mogensen, and G. Uze. 1998. Shared receptor componentsbut distinct complexes for � and � interferons. J. Mol. Biol. 282:585.
33. Rogge, L., L. Barberis-Maino, N. Passini, D. H. Presky, U. Gubler, andF. Sinigaglia. 1997. Selective expression of an interleukin-12 receptor componentby human T helper 1 cells. J. Exp. Med. 185:825.
34. Cousens, L. P., J. S. Orange, H. C. Su, and C. A. Biron. 1997. Interferon-�/�inhibition of interleukin-12 and interferon-� production in vitro and endog-enously during viral infection. Proc. Natl. Acad. Sci. USA 94:634.
4829The Journal of Immunology
by guest on Novem
ber 28, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

35. Wang, X., M. Chen, K. P. Wandinger, G. Williams, and S. Dhib-Jalbut. 2000.IFN-�-1b inhibits IL-12 production in peripheral blood mononuclear cells in anIL-10-dependent mechanism: relevance to IFN-�-1b therapeutic effects in mul-tiple sclerosis. J. Immunol. 165:548.
36. Van Boxel-Dezaire, A. H., S. C. Hoff, B. W. van Oosten, C. L. Verweij,A. M. Drager, H. J. Ader, J. C. van Houwelinger, F. Barkhof, C. H. Polman, andL. Nagelkerken. 1999. Decreased IL-10 and increased IL-12 p40 mRNA areassociated with disease activity and characterize different disease stages in mul-tiple sclerosis. Ann. Neurol. 45:695.
37. Howard, M., T. Muchamuel, S. Andrade, and S. Menon. 1993. IL-10 protectsmice from lethal endotoxemia. J. Exp. Med. 177:1205.
38. Wysocka, M., M. Kubin, L. Q. Vieria, L. Ozmen, G. Garotta, P. Scott, andG. Trinchieri. 1995. IL-12 is required for IFN-� production and lethality in LPS-induced shock in mice. Eur. J. Immunol. 25:672.
39. McCaffrey, T. A., D. J. Falcone, and B. Du. 1992. Transforming growth factor-�1is a heparin binding protein: identification of putative heparin-binding regionsand isolation of heparins with varying affinity for TGF-�1. J. Cell. Physiol. 152:430.
40. Spillmann, D., D. Witt, and U. Lindahl. 1998. Defining the IL-8-binding domainof heparan sulfate. J. Biol. Chem. 273:15487.
41. Hasan, M., S. Najjam, M. Y. Gordon, R. V. Gibbs, and C. C. Rider. 1999. IL-12is a heparin-binding cytokine. J. Immunol. 162:1064.
42. Najjam, S., B. Mulloy, J. Theze, M. Gordon, R. Gibbs, and C. C. Rider. 1998.Further characterization of the binding of human recombinant IL-2 to heparin andidentification of putative binding sites. Glycobiology 8:509.
43. Salek-Ardakani, S., J. R. Arrand, D. Shaw, and M. Mackett. 2000. Heparin andheparan sulfate bind IL-10 and modulate its activity. Blood 96:1879.
44. Hileman, R. E., J. R. Fromm, J. M. Weiler, and R. J. Linhardt. 1998. Glyco-saminoglycan-protein interactions: definition of consensus sites in glycosamino-glycan binding proteins. BioEssays 20:156.
45. Koopman, W., C. Ediriwickrema, and M. S. Krangel. 1999. Structure and func-tion of the glycosaminoglycan binding site of chemokine macrophage-inflamma-tory protein-1�. J. Immunol. 163:2120.
46. Gessani, S., F. Belardelli, A. Pecorelli, P. Puddu, and C. Baglioni. 1989. Bacteriallipopolysaccharide and � interferon induce transcription of � interferon mRNAand interferon secretion in murine macrophages. J. Virol. 63:2785.
47. Hayes, M. P., J. C. Enterline, T. L. Gerrard, and K. C. Zoon. 1991. Regulation ofinterferon production by human monocytes: requirements for priming for lipopo-lysaccharide-induced production. J. Leukocyte Biol. 50:176.
4830 IFN-� AND CELLS OF THE INNATE IMMUNE SYSTEM
by guest on Novem
ber 28, 2013http://w
ww
.jimm
unol.org/D
ownloaded from
Copyright © 2022 FDOKUMEN




















