
Mapping a gene conferring resistance to Pseudocercosporella herpotrichoides on chromosome 4V of...
6
Mapping a gene conferring resistance to Pseudocercosporella herpotrichoides on chromosome 4V of Dasypyrum villosum in a wheat background Ahmet Yildirim, Stephen S. Jones, and Timothy D. Murray Abstract: The objectives of this study were to map and tag the previously undescribed eyespot resistance gene PchDv on chromosome 4V of Dasypyrum villosum in a wheat background. The 82 F 2 plants used for mapping were produced from a cross between a susceptible wheat ‘Yangmai–5’ (4V(4D)) substitution line and a resistant wheat ‘Chinese Spring’ disomic addition line of chromosome 4V of D. villosum. Segregation for resistance and susceptibility among F 2 plants was 3:1, indicating that resistance was controlled by a single dominant gene. PchDv mapped to the distal part of chromosome 4V and was bracketed by two RFLP markers, Xcdo949 and Xbcd588, in a 33-cM interval. This distance could not be reduced, owing to a lack of polymorphic loci in this region. Theoretically, double recombination in this region occurs in 3.3% of the individuals; therefore, 96.7% of the selected genotypes would have PchDv, with simultaneous selection for both flanking markers. Double recombination between the flanking markers was observed in 2 out of 82 (2.4%) F 2 individuals. Key words: eyespot, Haynaldia villosa, Triticum aestivum, RFLP, wild wheat. Résumé : L’objectif de cette étude était de déterminer l’emplacement chromosomique et de marquer un gène de résistance au piétin-verse qui n’avait pas encore été décrit, PchDv, sur le chromosome 4V du Dasypyrum villosum dans un fond génétique de blé. Les 82 plantes F 2 employées pour la cartographie ont été produites par croisement entre une lignée de substitution sensible du blé ‘Yangmai–5’ (4V(4D)) et une lignée d’addition résistante disomique du blé ‘Chinese Spring’ portant le chromosome 4V du D. villosum. Un ratio de ségrégation de 3 résistantes : 1 sensible parmi les plantes F 2 a révélé que la résistance était sous le contrôle d’un seul gène dominant. Le gène PchDv a été assigné à un intervalle de 33 cM sur le partie distale du chromosome 4V entre les marqueurs RFLP Xcdo949 et Xbcd588. La distance n’a pu être réduite davantage en raison de l’absence de loci polymorphes dans cette région. Théoriquement, des doubles recombinaisons dans cette région surviennent dans 3,3% des individus, ainsi 96,7% des génotypes sélectionnés à l’aide des deux marqueurs posséderont le gène PchDv. Parmi les 82 plantes F 2 examinées, deux double recombinants (2,4%) ont été observés. Mots clés : piétin-verse, Haynaldia villosa, Triticum aestivum, RFLP, blé sauvage. [Traduit par la Rédaction] Introduction Eyespot, caused by the facultative parasite Pseudocerco- sporella herpotrichoides (Fron.) Deighton, is an economically important disease of wheat in the U.S. Pacific Northwest and many other winter wheat growing areas of the world (Johnson 1992; Wiese 1991). Previously, there were only two resistance genes known for eyespot. The first, Pch1, was transferred from Aegilops ventricosa (Dosba and Dossinault 1973) and is used extensively in breeding programs because of tightly linked isozyme (McMillin et al. 1986) and RFLP markers (Chao et al. 1989). Pch1 alone does not provide complete resistance to the disease, but is more effective than the second gene, Pch2. Pch1 occurs on the long arm of chromosome 7D and, recently, Pch2 was mapped to the long arm of chromosome 7A (de la Peña et al. 1997). Occurrence of new pathotypes of the pathogen increases the possibility of circumventing existing resistance (Jones et al. 1995). Dasypyrum villosum (2n = 14, genome VV), a distant rela- tive of wheat, was identified as being highly resistant to eye- spot, and a resistance gene(s) was located in a chromosome 4V addition line (Murray et al. 1994). Subsequently, it was deter- mined that a disomic wheat – D. villosum 4V(4D) substitu- tion line was susceptible to eyespot (Yildirim et al. 1997; A. Yildirim, unpublished data). The existence of the suscepti- ble 4V substitution line makes it possible to map the resistance gene in a wheat background, which is advantageous, because the chromosome is homozygous in a disomic state. Dasypyrum villosum is a cross-pollinating diploid species (Frederiksen Genome, 41: 1–6 (1998) Corresponding Editor: J.P. Gustafson. Received May 28, 1997. Accepted September 16, 1997. A. Yildirim 1 and S.S. Jones. 2 Department of Crop and Soil Sciences, Washington State University, Pullman, WA 99164–6420, U.S.A. T.D. Murray. Department of Plant Pathology, Washington State University, Pullman, WA 99164–6430, U.S.A. 1 Present address: Gaziosmanpasa University Ziraat Fakültesé, 60110 Tokat, Turkey. 2 Author to whom all correspondence should be addressed (e-mail: [email protected]). 1 © 1998 NRC Canada
Transcript of Mapping a gene conferring resistance to Pseudocercosporella herpotrichoides on chromosome 4V of...

Mapping a gene conferring resistance to
Pseudocercosporella herpotrichoides on
chromosome 4V of Dasypyrum villosum in a
wheat background
Ahmet Yildirim, Stephen S. Jones, and Timothy D. Murray
Abstract: The objectives of this study were to map and tag the previously undescribed eyespot resistance gene PchDv on
chromosome 4V of Dasypyrum villosum in a wheat background. The 82 F2 plants used for mapping were produced from a
cross between a susceptible wheat ‘Yangmai–5’ (4V(4D)) substitution line and a resistant wheat ‘Chinese Spring’ disomic
addition line of chromosome 4V of D. villosum. Segregation for resistance and susceptibility among F2 plants was 3:1,
indicating that resistance was controlled by a single dominant gene. PchDv mapped to the distal part of chromosome 4V and
was bracketed by two RFLP markers, Xcdo949 and Xbcd588, in a 33-cM interval. This distance could not be reduced, owing
to a lack of polymorphic loci in this region. Theoretically, double recombination in this region occurs in 3.3% of the
individuals; therefore, 96.7% of the selected genotypes would have PchDv, with simultaneous selection for both flanking
markers. Double recombination between the flanking markers was observed in 2 out of 82 (2.4%) F2 individuals.
Key words: eyespot, Haynaldia villosa, Triticum aestivum, RFLP, wild wheat.
Résumé: L’objectif de cette étude était de déterminer l’emplacement chromosomique et de marquer un gène de résistance au
piétin-verse qui n’avait pas encore été décrit, PchDv, sur le chromosome 4V du Dasypyrum villosum dans un fond génétique
de blé. Les 82 plantes F2 employées pour la cartographie ont été produites par croisement entre une lignée de substitution
sensible du blé ‘Yangmai–5’ (4V(4D)) et une lignée d’addition résistante disomique du blé ‘Chinese Spring’ portant le
chromosome 4V du D. villosum. Un ratio de ségrégation de 3 résistantes : 1 sensible parmi les plantes F2 a révélé que la
résistance était sous le contrôle d’un seul gène dominant. Le gène PchDv a été assigné à un intervalle de 33 cM sur le partie
distale du chromosome 4V entre les marqueurs RFLP Xcdo949 et Xbcd588. La distance n’a pu être réduite davantage en
raison de l’absence de loci polymorphes dans cette région. Théoriquement, des doubles recombinaisons dans cette région
surviennent dans 3,3% des individus, ainsi 96,7% des génotypes sélectionnés à l’aide des deux marqueurs posséderont le gène
PchDv. Parmi les 82 plantes F2 examinées, deux double recombinants (2,4%) ont été observés.
Mots clés : piétin-verse, Haynaldia villosa, Triticum aestivum, RFLP, blé sauvage.
[Traduit par la Rédaction]
Introduction
Eyespot, caused by the facultative parasite Pseudocerco-sporella herpotrichoides (Fron.) Deighton, is an economicallyimportant disease of wheat in the U.S. Pacific Northwest andmany other winter wheat growing areas of the world (Johnson1992; Wiese 1991). Previously, there were only two resistancegenes known for eyespot. The first, Pch1, was transferred from
Aegilops ventricosa (Dosba and Dossinault 1973) and is usedextensively in breeding programs because of tightly linkedisozyme (McMillin et al. 1986) and RFLP markers (Chao et al.1989). Pch1 alone does not provide complete resistance to thedisease, but is more effective than the second gene, Pch2. Pch1occurs on the long arm of chromosome 7D and, recently, Pch2was mapped to the long arm of chromosome 7A (de la Peñaet al. 1997). Occurrence of new pathotypes of the pathogenincreases the possibility of circumventing existing resistance(Jones et al. 1995).
Dasypyrum villosum (2n = 14, genome VV), a distant rela-tive of wheat, was identified as being highly resistant to eye-spot, and a resistance gene(s) was located in a chromosome 4Vaddition line (Murray et al. 1994). Subsequently, it was deter-mined that a disomic wheat – D. villosum 4V(4D) substitu-tion line was susceptible to eyespot (Yildirim et al. 1997;A. Yildirim, unpublished data). The existence of the suscepti-ble 4V substitution line makes it possible to map the resistancegene in a wheat background, which is advantageous, becausethe chromosome is homozygous in a disomic state. Dasypyrumvillosum is a cross-pollinating diploid species (Frederiksen
Genome, 41: 1–6 (1998)
Corresponding Editor: J.P. Gustafson.
Received May 28, 1997. Accepted September 16, 1997.
A. Yildirim 1 and S.S. Jones.2 Department of Crop and SoilSciences, Washington State University, Pullman, WA99164–6420, U.S.A.T.D. Murray. Department of Plant Pathology, WashingtonState University, Pullman, WA 99164–6430, U.S.A.
1 Present address: Gaziosmanpasa University Ziraat Fakültesé,60110 Tokat, Turkey.
2 Author to whom all correspondence should be addressed(e-mail: [email protected]).
1
© 1998 NRC Canada
1991) and, theoretically, the genetic constitution is mostly het-erozygous, thus complicating standard F2 mapping protocols.In addition, inbreeding depression and self-incompatabilitymake the production of homozygous lines difficult. Moreover,the level of resistance in the disomic addition line of chromo-some 4V in a ‘Chinese Spring’ background was shown to beequal to D. villosum itself (Murray et al. 1994). The level ofresistance in a hexaploid background conferred by a gene ona D. villosum chromosome shows that this gene has potentialin wheat improvement programs.
The success and accuracy of a map strongly depend on theprecise identification of the individual genotypes in the map-ping population. Previously, the mapping of eyespot resistancegenes and the development of cultivars with this resistance wasslowed by an inability to recognize and select for resistantgenotypes with certainty. Field evaluations based on visualscores did not unambiguosly differentiate between highly re-sistant, resistant, and susceptible genotypes (Jones et al. 1995).Recently, an improved technique for evaluating resistance wasdescribed that uses a GUS (β-glucuronidase) transformedstrain of P. herpotrichoides (de la Peña and Murray 1994). Inthis technique, disease development in 4- to 8-week-old seed-lings is scored on the basis of the amount of GUS. The amountof fungal growth on young seedlings, and thus disease severity,is directly related to the amount of GUS activity. Therefore,the method clearly differentiates among highly resistant, resis-tant, and susceptible genotypes.
Identification and transfer of new genes for eyespot resis-tance is important, since neither Pch1 nor Pch2 completely
prevent disease. In fact, ‘Rendezvous’, which reportedly pos-sesses both genes (Hollins et al. 1988), and ‘Madsen’ and‘Hyak’, which are the only resistant cultivars in the U.S. (Allanet al. 1989, 1990), still sustain yield losses when disease issevere (Jones et al. 1995). Such losses demonstrate the needfor cultivars with improved eyespot resistance, produced eitherby identifying new sources of resistance or through pyramid-ing resistance genes in the same cultivar.
The objective of this study was to map PchDv on chromo-some 4V, with the ultimate goal of the rapid and successfultransfer of the gene into a stable wheat background.
Materials and methods
Genetic stocksA disomic chromosome 4V addition line of D. villosum in a ‘ChineseSpring’ background (CS + 4V) (2n = 44) (Hyde 1953; Sears 1953)was obtained from the Wheat Genetics Resource Center, Kansas StateUniversity, Manhattan, Kans. An experimental wheat line with chro-mosome 4V substituted for chromosome 4D in the cultivar ‘Yangmai–5’(4V(4D)) (2n = 42) was provided by Dr. P.D. Chen, CytogeneticsInstitute, Nanjing Agricultural University, Nanjing, China. The linesVPM–1 (resistant breeding line) and ‘Chinese Spring’ (susceptible)were used as controls for eyespot reaction.
Mapping populationA genetic map of chromosome 4V was constructed using a populationof 82 F2 plants produced from a cross between the resistant disomicCS + 4V addition line and the susceptible ‘Yangmai–5’ (4V(4D))substitution line. Initially, 98 F2 plants were produced, but only 82 ofthem were used in the construction of the map, owing to insufficient
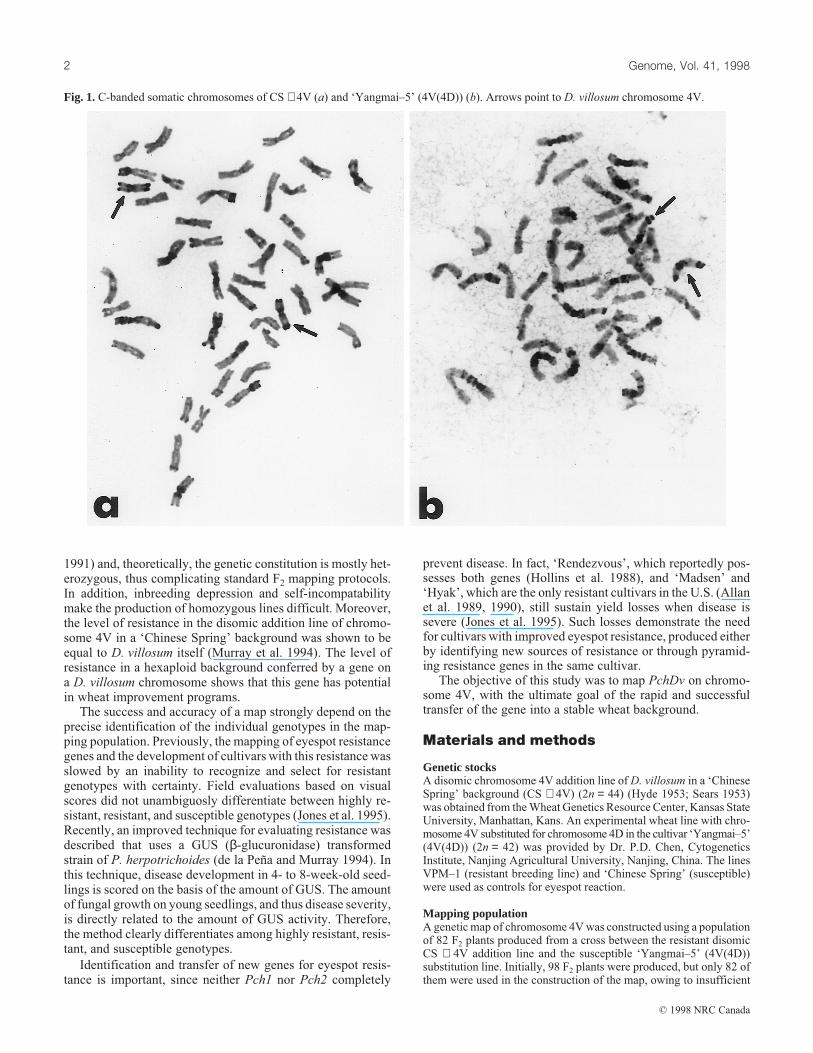
Fig. 1. C-banded somatic chromosomes of CS + 4V (a) and ‘Yangmai–5’ (4V(4D)) (b). Arrows point to D. villosum chromosome 4V.
Genome, Vol. 41, 19982
© 1998 NRC Canada
amounts of F2 DNA and (or) delayed germination. Chromosome 4Vfrom both parents was expected to pair in F1 hybrids, producing atrisomic individual with 2n = 43 chromosomes. Homologous recom-bination between the two 4V chromosomes was required for map-ping. Chromosome 4D from the addition line would be univalent inthe hybrids, and in most cases was expected to be lost in the nextgeneration. Aneuploid lines of ‘Chinese Spring’ (Sears 1954) wereused to verify chromosomes involved in polymorphic loci.
Eyespot reaction and statistical analysisA genetically modified strain of P. herpotrichoides, with growth rateand pathogenicity similar to its parental strain but expressing GUS,was used to identify resistant plants. Seedlings were inoculated at thetwo-leaf stage and the disease reaction was scored as described pre-viously by Yildirim et al. (1995). GUS values of F2 plants were dif-ferentiated by Fisher’s least significant difference (LSD) (P = 0.05),based on the standard error of mean differences of the parents (Littleand Hills 1978). F2 plants with GUS values that exceeded the mean ofthe resistant parent (CS + 4V) value plus the LSD value were consid-ered susceptible.
Isolation of genomic DNA and Southern hybridizationPrior to disease evaluation, leaves were harvested from each F2 plantfor DNA extraction. Genomic DNA isolation, from individual F2
plants and single-enzyme digests of genomic DNA with restrictionendonucleases BamHI, DraI, EcoRI, EcoRV, HindIII, PstI, XbaI, orXhoI, and Southern transfers were performed according to Cadle et al.(1994). The DNA blots were prehybridized for at least 6 h at 65°C ina mixture of 7% SDS, 0.25 M Na2HPO4 (pH 7.2), and 1 mM EDTA(pH 8.0) (Church and Gilbert 1984). After hybridization with[32P]dCTP labelled probe and washing, blots were exposed to x-rayfilm (Eastman Kodak Co., Rochester, N.Y.) for 5–10 days at –80ºC,depending upon the intensity of the signals.
Detection of polymorphism and linkage analysisSeventy-seven single-copy cDNA and genomic clones containing ho-mologous sequences on the long arm of group 4 chromosomes ofwheat were used to detect polymorphism between the parents. Probesthat revealed polymorphism were used for segregation analysis. The
MAPMAKER computer program was used to determine recombinationfrequencies and locus order (Lander et al. 1987).
C-bandingThe presence of chromosome 4V in the parents was confirmed byC-banding. Mitotic chromosomes were prepared for C-banding ac-cording to Cai et al. (1996). C-banding was performed as describedby Gill et al. (1991a).
In situ hybridizationThe pairing behaviour of 4V chromosomes in F1 plants was checkedby in situ hybridization (ISH). Anthers of F1 plants at meiotic meta-phase I (MI) were collected and fixed in ethanol – acetic acid (3:1) atroom temperature for 2–3 days, or at 4°C for longer periods of time(1–8 weeks). Anthers at MI were squashed in 45% acetic acid afterstaining in 2% acetocarmine for 10 min. Cover glasses were removedby freezing the slides at –80°C. A highly repetitive D. villosum spe-cific probe, p380 (obtained from C. De Pace, Department of Agricul-tural Biology and Chemistry, University of Tuscia, Viterbo, Italy),was labelled with biotin-16-dUTP by nick translation (Enzo Diagnos-tics Nick Translation Kit). Probe hybridization and signal detectionwere performed according to Friebe et al. (1993).
Results
Genetic constitution of the parents, meiotic pairing, andF2 plants
The cross between CS + 4V (2n = 44) and ‘Yangmai–5’
Fig. 3. In situ hybridization pattern of MI in pollen mother cells of
F1 hybrid between CS + 4V and ‘Yangmai–5’ (4V(4D)) hybridized
with D. villosum specific probe p380. The arrow indicates paired
4V chromosomes and the open arrowhead indicates the univalent
4D chromosome.
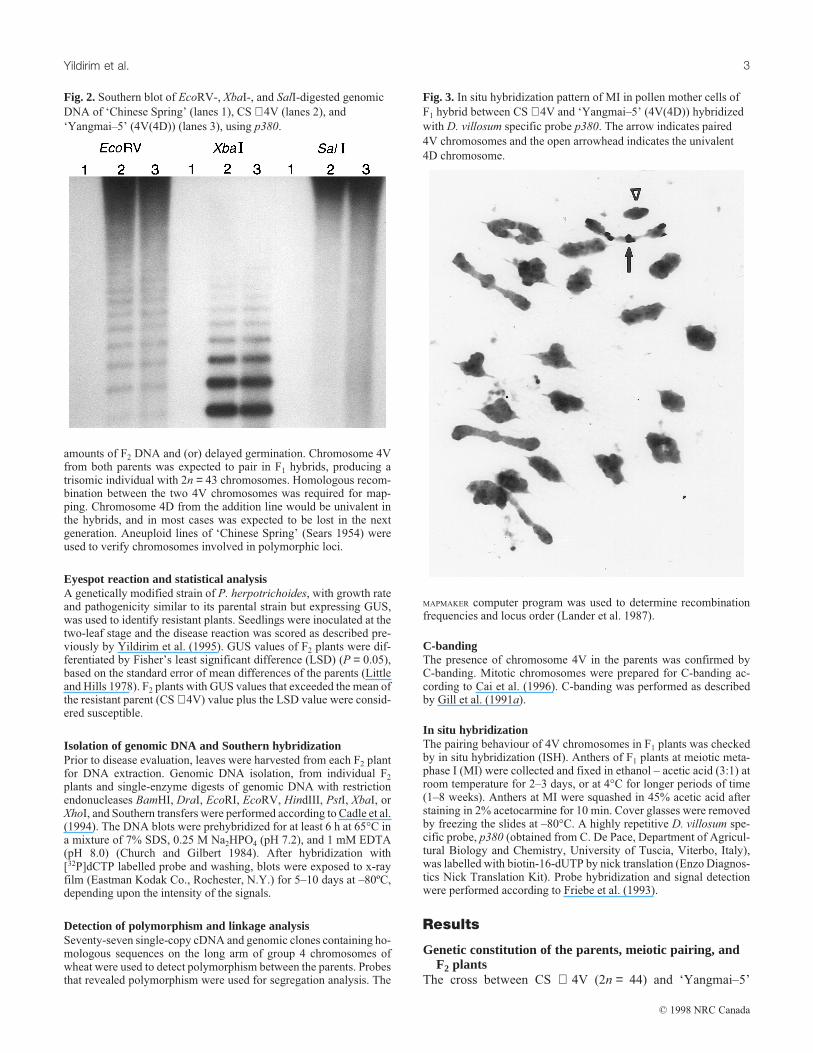
Fig. 2. Southern blot of EcoRV-, XbaI-, and SalI-digested genomic
DNA of ‘Chinese Spring’ (lanes 1), CS + 4V (lanes 2), and
‘Yangmai–5’ (4V(4D)) (lanes 3), using p380.
Yildirim et al. 3
© 1998 NRC Canada
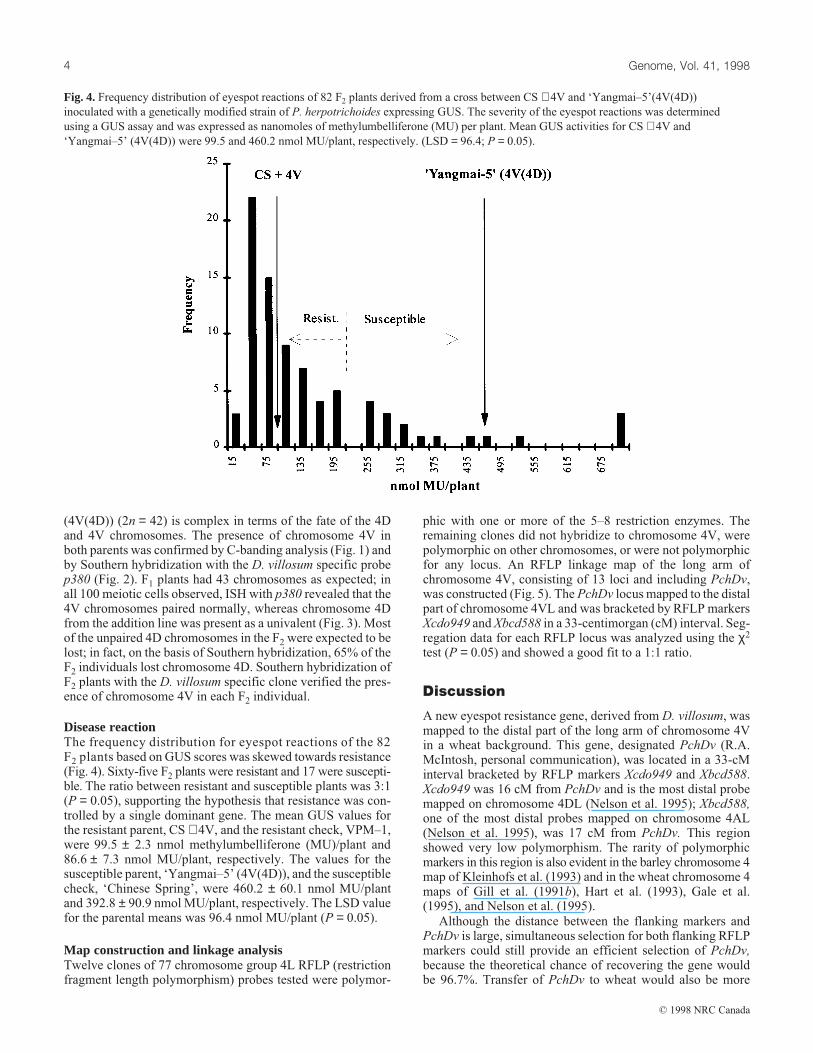
(4V(4D)) (2n = 42) is complex in terms of the fate of the 4Dand 4V chromosomes. The presence of chromosome 4V inboth parents was confirmed by C-banding analysis (Fig. 1) andby Southern hybridization with the D. villosum specific probep380 (Fig. 2). F1 plants had 43 chromosomes as expected; inall 100 meiotic cells observed, ISH with p380 revealed that the4V chromosomes paired normally, whereas chromosome 4Dfrom the addition line was present as a univalent (Fig. 3). Mostof the unpaired 4D chromosomes in the F2 were expected to belost; in fact, on the basis of Southern hybridization, 65% of theF2 individuals lost chromosome 4D. Southern hybridization ofF2 plants with the D. villosum specific clone verified the pres-ence of chromosome 4V in each F2 individual.
Disease reactionThe frequency distribution for eyespot reactions of the 82F2 plants based on GUS scores was skewed towards resistance(Fig. 4). Sixty-five F2 plants were resistant and 17 were suscepti-ble. The ratio between resistant and susceptible plants was 3:1(P = 0.05), supporting the hypothesis that resistance was con-trolled by a single dominant gene. The mean GUS values forthe resistant parent, CS + 4V, and the resistant check, VPM–1,were 99.5 ± 2.3 nmol methylumbelliferone (MU)/plant and86.6 ± 7.3 nmol MU/plant, respectively. The values for thesusceptible parent, ‘Yangmai–5’ (4V(4D)), and the susceptiblecheck, ‘Chinese Spring’, were 460.2 ± 60.1 nmol MU/plantand 392.8 ± 90.9 nmol MU/plant, respectively. The LSD valuefor the parental means was 96.4 nmol MU/plant (P = 0.05).
Map construction and linkage analysisTwelve clones of 77 chromosome group 4L RFLP (restrictionfragment length polymorphism) probes tested were polymor-
phic with one or more of the 5–8 restriction enzymes. Theremaining clones did not hybridize to chromosome 4V, werepolymorphic on other chromosomes, or were not polymorphicfor any locus. An RFLP linkage map of the long arm ofchromosome 4V, consisting of 13 loci and including PchDv,was constructed (Fig. 5). The PchDv locus mapped to the distalpart of chromosome 4VL and was bracketed by RFLP markersXcdo949 and Xbcd588 in a 33-centimorgan (cM) interval. Seg-regation data for each RFLP locus was analyzed using the χ2
test (P = 0.05) and showed a good fit to a 1:1 ratio.
Discussion
A new eyespot resistance gene, derived from D. villosum, wasmapped to the distal part of the long arm of chromosome 4Vin a wheat background. This gene, designated PchDv (R.A.McIntosh, personal communication), was located in a 33-cMinterval bracketed by RFLP markers Xcdo949 and Xbcd588.Xcdo949 was 16 cM from PchDv and is the most distal probemapped on chromosome 4DL (Nelson et al. 1995); Xbcd588,one of the most distal probes mapped on chromosome 4AL(Nelson et al. 1995), was 17 cM from PchDv. This regionshowed very low polymorphism. The rarity of polymorphicmarkers in this region is also evident in the barley chromosome 4map of Kleinhofs et al. (1993) and in the wheat chromosome 4maps of Gill et al. (1991b), Hart et al. (1993), Gale et al.(1995), and Nelson et al. (1995).
Although the distance between the flanking markers andPchDv is large, simultaneous selection for both flanking RFLPmarkers could still provide an efficient selection of PchDv,because the theoretical chance of recovering the gene wouldbe 96.7%. Transfer of PchDv to wheat would also be more
Fig. 4. Frequency distribution of eyespot reactions of 82 F2 plants derived from a cross between CS + 4V and ‘Yangmai–5’(4V(4D))
inoculated with a genetically modified strain of P. herpotrichoides expressing GUS. The severity of the eyespot reactions was determined
using a GUS assay and was expressed as nanomoles of methylumbelliferone (MU) per plant. Mean GUS activities for CS + 4V and
‘Yangmai–5’ (4V(4D)) were 99.5 and 460.2 nmol MU/plant, respectively. (LSD = 96.4; P = 0.05).
Genome, Vol. 41, 19984
© 1998 NRC Canada

efficient, because 4V chromatin, other than that containing thegene of interest, can be selected against. Double recombinationbetween the flanking markers was observed in only two F2
plants out of the 82 individuals. This closely matches the theo-retical value of successful selection based on flanking markers.The use of flanking markers for PchDv, and for the recentlymapped Pch2, will enhance opportunities for pyramiding eye-spot resistance genes, and could be used to improve the overalleffectiveness of resistant varieties.
In general, there was colinearity in the locus order and po-sition between the chromosome 4V map developed here andthe chromosome group 4 maps of wheat and barley (Gale et al.1995; Kleinhofs et al. 1993; Nelson et al. 1995). However,five markers in the chromosome 4V map, Xpsr104, Xpsr163,Xpsr139, Xpsr115, and Xpsr119, mapped on chromosome 4Aof wheat (Gale et al. 1995) in a reverse order but with similardistances. In the 4AL consensus map (Gale et al. 1995), thedistance between Xpsr139 and Xpsr115 was 71 cM and be-tween Xpsr115 and Xpsr119 was 35.2 cM, whereas in the 4V
map, these distances were 73 and 37.3 cM, respectively.Xpsr104 and Xpsr163 mapped to the short arm of chromosome4A, but mapped to the long arms of chromosomes 4B and 4D,with the same distance (2.7 cM), but in reverse order (Galeet al. 1995). We have also mapped these loci on the long armof chromosome 4V in an order that was consistent with thaton chromosomes 4BL and 4DL. These results are in agreementwith a previously reported pericentric inversion on chromo-some 4AL of wheat relative to chromosomes 4Am, 4B, and 4D(Naranjo 1990; Devos et al. 1995).
The mapping population, and the approach of using thistype of cross to map a non-wheat gene, made the cytologicalcharacterization of both 4V chromosomes extremely impor-tant. Mitotic C-banding patterns of the parents in this studyrevealed 4V chromosomes in both parents (Fig. 1). In addition,ISH with p380 verified that chromosome 4V was pairing nor-mally and that the extra univalent was a wheat chromosome(Fig. 3). These results eliminated any doubts about recombi-nation events on 4V chromosomes, an element needed for asuccessful map. This technique has proven useful for mappinghomozygous 4V chromosomes from different sources and haseliminated the need to deal with the heterozygosity of D. vil-losum at the diploid level. Mapping PchDv on chromosome4V in a wheat background will provide a more efficient ma-nipulation of the gene through molecular assisted selectiontechniques.
Acknowledgments
Special thanks are extended to Drs. B.S. Gill and P.D. Chen forproviding CS + 4V and ‘Yangmai–5’ (4V(4D)) seeds, and toA. Kleinhofs, C. De Pace, M.D. Gale, M.E. Sorrells, andP. Leroy for providing the clones used in this study.
References
Allan, R.E., Peterson, C.J., Jr., Rubenthaler, G.L., Line, R.F., andRoberts, D.E. 1989. Registration of ‘Madsen’ wheat. Crop Sci. 29:1575–1576.
Allan, R.E., Peterson, C.J., Jr., Rubenthaler, G.L., Line, R.F., andRoberts, D.E. 1990. Registration of ‘Hyak’ wheat. Crop Sci. 30:234.
Cadle, M.M., Rayfuse, L.M., Walker-Simmons, M.K.,and Jones, S.S.1994. Mapping abscisic acid responsive genes and vp1 to chromo-somes in wheat and Lophopyrum elongatum. Genome, 37:129–132.
Cai, X., Jones, S.S., and Murray, T.D. 1996. Characterization of anAgropyron elongatum chromosome conferring resistance tocephalosporium stripe in common wheat. Genome, 39: 56–62.
Chao, S., Sharp, P.J., Worland, A.J., Warham, A.J., Koebner, R.M.D.,and Gale, M.D. 1989. RFLP-based maps of wheat homoeologousgroup 7 chromosomes. Theor. Appl. Genet. 78: 495–504.
Church, G., and Gilbert, W. 1984. Genomic sequencing. Proc. Natl.Acad. Sci. U.S.A. 81:1991.
de la Peña, R.C., and Murray, T.D. 1994. Identifying wheat genotypesresistant to eyespot disease with a β-glucuronidase-transformedstrain of Pseudocercosporella herpotrichoides. Phytopathology,84: 972–977.
de la Peña, R.C., Murray, T.D., and Jones, S.S. 1997. Identification ofan RFLP interval containing Pch2 on chromosome 7AL in wheat.Genome, 40: 249–252.
Devos, K.M., Dubcovsky, J., Dvor ák, J., Chinoy, C.N., and Gale,M.D. 1995. Structural evolution of wheat chromosomes 4A, 5A,
Fig. 5. Genetic linkage map of the long arm of chromosome 4V of
D. villosum constructed from 82 F2 plants derived from CS + 4V בYangmai–5’ (4V(4D)). Map distances and locus order were
determined using MAPMAKER 3.0 (Lander et al. 1987).
Yildirim et al. 5
© 1998 NRC Canada

and 7B and its impact on recombination. Theor. Appl. Genet. 91:282–288.
Dosba, F., and Dossinault, G. 1973. Resistance to eyespot (Cerco-sporella herpotrichoides) introduced to bread wheat fromAegilops ventricosa. Proc. Int. Wheat Genet. Symp. 4: 409–413.
Frederiksen, S. 1991. Taxonomic studies in Dasypyrum (Poaceae).Nord. J. Bot. 11: 135–142.
Friebe, B., Jiang, J., Gill, B.S., and Dyck, P.L. 1993. Radiation in-duced nonhomoeologous wheat – Agropyron elongatum trans-locations conferring resistance to leaf rust. Theor. Appl. Genet.86: 141–149.
Gale, M.D., Atkinson, M.D., Chinoy, C.N., Harcourt, R.L., Jia, J., Li,Q.Y., and Devos, K.M. 1995. Genetic maps of hexaploid wheat.In Proceedings of the 8th International Wheat Genetics Sympo-sium, held at Beijing, China, 20–25 July 1993. Edited by Z.S. Liand Z.Y. Xin. China Agricultural Scientech Press, Beijing. pp.29–40.
Gill, B.S., Friebe, B., and Endo, T.R. 1991a. Standard karyotype andnomenclature system for description of chromosome bands andstructural aberrations in wheat (Triticum aestivum). Genome, 34:830–839.
Gill, K.S., Lubbers, E.L., Gill, B.S., Raupp, W.J., and Cox, T.S.1991b. A genetic linkage map of Triticum tauschii (DD) and itsrelationship to the D genome of bread wheat (AABBDD).Genome, 34: 362–374.
Hart, G.E., Gale, M.D., and McIntosh, R.A. 1993. Linkage maps ofTriticum aestivum (hexaploid wheat, 2n = 42, genomes A, B andD) and T. tauschii (2n = 14, genome D). In Genetic Maps, 6th ed.Edited by S.J. O’Brien. Cold Spring Harbor Laboratory, ColdSpring Harbor, N.Y. pp. 6204–6219.
Hollins, T.W., Lockley, K.D., Blackman, J.A., Scott, P.R., and Bing-ham, J. 1988. Field performance of Rendezvous, a wheat cultivarwith resistance to eyespot (Pseudocercosporella herpotrichoides)derived from Aegilops ventricosa. Plant Pathol. 37: 251–260.
Hyde, B.B. 1953. Addition of individual Haynaldia villosa chromo-somes to hexaploid wheat. Am. J. Bot. 40: 174–182.
Johnson, R. 1992. Reflections of a plant pathologist on breeding dis-ease resistance, with emphasis on yellow rust and eyespot ofwheat. Plant Pathol. 41: 239–254.
Jones, S.S., Murray, T.D., and Allan, R.E. 1995. Use of alien genesfor the development of disease resistance in wheat. Annu. Rev.Phytopathol. 33: 429–443.
Kleinhofs, A., Kilian, A., Saghai-Maroof, MA., Biyashev, K.M.,Hayes, P., Chen, F.Q., Lapitan, N., Fenwick, A., Blake, T.K.,Kanazin, V., Ananiev, E., Dahleen, L., Kudrna, D., Bollinger, J.,Knapp, S.J., Liu, B., Sorrells, M., Heun, M., Francowiak, J.D.,Hoffman, D., Skadsen, R., and Steffenson, B.J. 1993. A molecu-lar, isozyme, and morphological map of the barley (Hordeum vul-gare) genome. Theor. Appl. Genet. 85: 705–712.
Lander, E., Green, P., Abrahamson, J., Barlow, A., Daly, M., Lincoln,S., and Newburg, L. 1987. MAPMAKER: an interactive computerpackage for constructing primary genetic linkage maps and ex-perimental and natural populations. Genomics, 1: 174–181.
Little, T.M., and Hills, F.J. 1978. Agricultural experimentation: de-sign and analysis. John Wiley & Sons, N.Y.
McMillin, D.E., Allan, R.E., and Roberts, D.E. 1986. Association ofan isozyme locus and strawbreaker foot rot resistance derivedfrom Aegilops ventricosa in wheat. Theor. Appl. Genet. 72:743–747.
Murray, T.D., de la Peña, R.C., Yildirim, A., and Jones, S.S. 1994. Anew source of resistance to Pseudocercosporella herpotrichoides,cause of eyespot disease of wheat, located on chromosome 4V ofDasypyrum villosum. Plant Breed. 113: 281–286.
Naranjo, T. 1990. Chromosome structure of durum wheat. Theor.Appl. Genet. 79: 397–400.
Nelson, J.C., Sorrells, M.E., Van Deynze, A.E., Lu, Y.H., Atkinson,M.D., Bernard, M., Leroy, P., Faris, J., and Anderson, J. 1995.Molecular mapping of wheat. Major genes and rearrangements inhomoeologous groups 4, 5, and 7. Genetics, 141: 721–731.
Sears, E.R. 1953. Addition of the genome of Haynaldia villosa toTriticum aestivum. Am. J. Bot. 40: 168–174.
Sears, E.R. 1954. The aneuploids of common wheat. Mo. Agric. Exp.Stn. Res. Bull. No. 572. pp. 1–59.
Wiese, M.V. 1991. Compedium of wheat diseases. 2nd ed. APS Press,St. Paul, Minn. pp. 47–48.
Yildirim, A., Jones, S.S., Murray, T.D., Cox, T.S., and Line, R.F.1995. Resistance to stripe rust and eyespot diseases of wheat inTriticum tauschii. Plant Dis.79: 1230–1236.
Genome, Vol. 41, 19986
© 1998 NRC Canada




















