
Autologous Pancreatic Islet Transplantation in Human Bone Marrow
Upload
independentCategory
view
1download
0
UNCO
RRECTED P
RO
OF
1 Low dose naltrexone (LDN) enhances maturation of bone marrow dendritic2 cells (BMDCs)
3 JingjuanQ1 Meng a, Yiming Meng b, Nicholas P. Plotnikoff c, Gene Youkilis c, Noreen Grif!n c, Fengping Shan b,!4 a Central Laboratory, China Medical University, Shenyang 110001, China5 b Department of Immunology, School of Basic Medical Science, China Medical University, Shenyang 110001, China6 c TNI Bio Tech. Inc., 6701 Democracy Blvd., Suite 300, Bethesda, MD 20817, USA
7
8
a b s t r a c ta r t i c l e i n f o
9 Article history:10 Received 30 September 201311 Received in revised form 15 October 201312 Accepted 15 October 201313 Available online xxxx14151617 Keywords:18 Low dose naltrexone19 Bone marrow derived dendritic cells20 Modulation21 Maturation22 Immune system
23It has been demonstrated previously that immune cell activation and proliferationwere sensitive to the effects of24naltrexone, a non-peptidic "-opioid receptor selective antagonist and opioid receptors on BMDCs have been25detected [1]. However, there is little prior data published on naltrexone and DCs. Therefore, we hypothesized26that LDN could exert modulating effect on BMDCs. In present study, we studied in"uence of LDN on both27phenotypic and functional maturation of BMDCs. Changes of BMDC Q8post-treatment with LDN were evaluated28using conventional light microscope and transmission electron microscopy (TEM); "ow cytometry (FCM);29cytochemistry; acid phosphatase activity (ACP) test; FITC-dextran bio-assay; mixed lymphocytes and enzyme-30linked immunosorbent assay (ELISA). We have found that LDN enhances maturation of BMDCs as evidenced31by 1) up-regulating the expression of MHC II, CD40, CD83, CD80 and CD86 molecules on BMDCs; 2) down-32regulating the rates of pinocytosis and phagocytosis accompanied by the results of decreased ACP, and FITC-33dextran bio-assay; 3) mounting potential of BMDCs to drive T cell; and 4) inducing secretion of higher levels34of IL-12 and TNF-!. It is therefore concluded that LDN can ef!ciently promote the maturation of BMDCs via35precise modulation inside and outside BMDCs. Our study has provided meaningful mode of action on the role36of LDN in immunoregulation, and rationale on future application of LDN for enhancing host immunity in cancer37therapy and potent use in the design of DC-based vaccines for a number of diseases.38© 2013 Published by Elsevier B.V.
3940
41
42
43 1. Introduction
44 Naltrexone is an opioid receptor antagonist used primarily in45 the management of alcohol dependence and opioid addiction [2].46 Accumulating evidence suggests that LDN can promote health sup-47 porting immune modulation which may reduce various oncogenic48 and in"ammatory autoimmune processes [3]. There have been previous49 reports that kappa-opioid receptors are found onDCs [1]. Since LDN can50 upregulate endogenous opioid activity, it may also have a role in51 promoting stress resilience, exercise, social bonding, and emotional52 well-being, as well as amelioration of psychiatric problems such asQ10
53 autism and depression. It is proposed that LDN can be used effectively54 as a buffer for a large variety of bodily and mental ailments through55 its ability to bene!cially modulate both the immune system and the56 brain neurochemistry that regulates positive affect [4].
57Research following the discovery of dendritic cells (DCs) is58profoundly changing the science of immunology and its many59interfaces with medicine. These previously unknown cells are now60recognized as controllers that both create and curtail immunity [5].61DCs are a critical, and previously missing, link in the immune system.62As sentinels, dendritic cells patrol the body seeking out foreign invaders,63whether these are bacteria, viruses, or dangerous toxins. After capturing64the invaders, often termed antigens, dendritic cells convert them into65smaller pieces and display the antigenic fragments on their cell surfaces66to initiate T cell responses [6]. However, so far there is no report on67LDN's in"uence on DCs' maturation and related mechanisms remain68unclear. Due to the importance of LDN in modulation of immune loop69we hereby conducted following work to try to provide evidence to70elucidate these mechanisms.
712. Materials and methods
722.1. Chemicals
73LDN (naltrexonehydrochloride, C20H23NO4·HCl,MW: 377.86)was74made in France. The concentrations of LDN from 15.63#g/ml to 1mg/ml75on the proliferation of BMDCs in vitro were tested [7], and the optimal76concentration was found to be 125 #g/ml. Based on these data, the
International Immunopharmacology xxx (2013) xxx–xxx
Abbreviations: LDN, low dose naltrexone; GM-CSF, granulocyte macrophage colonystimulating factor; IL-4, interleukin-4; ACP, acidic phosphatase; BMDCs, bone marrowderived dendritic cells; APCs, antigen presenting cells; LPS, lipopolysaccharide;TEM, transmission electron microscopy; DAB, 3,3"-diaminobenzidineQ9 ; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide; FCM, "ow cytometry; MACS,magnetic activated cell sorting; CTL, cytotoxic lymphocyte.! Corresponding author. Tel.: +86 24 23256666; fax: +86 24 23989860.
E-mail address: [email protected] (F. Shan).
INTIMP-03054; No of Pages 6
1567-5769/$ – see front matter © 2013 Published by Elsevier B.V.http://dx.doi.org/10.1016/j.intimp.2013.10.012
Contents lists available at ScienceDirect
International Immunopharmacology
j ourna l homepage: www.e lsev ie r .com/ locate / in t imp
Please cite this article as: Meng J, et al, Low dose naltrexone (LDN) enhances maturation of bone marrow dendritic cells (BMDCs), IntImmunopharmacol (2013), http://dx.doi.org/10.1016/j.intimp.2013.10.012

UNCO
RRECTED P
RO
OF
77 optimal concentration was used in present study. The mAbs for this78 study include FITC-conjugated anti-CD40, PE-anti-MHC-II, PE-anti-79 CD80, PE-anti-CD83, and PE-anti-CD86, all of which were products of80 eBioscience and BD Pharmingen. The ELISA kits for IL-12 and TNF-!81 were purchased from eBioscience. LPS prepared from Escherichia coli82 (serotype 055:B5) was purchased from Sigma-Aldrich and was tested83 with tachypleus amebocyte lysate to prove that LPS is endotoxin free.84 Recombinantmurine cytokines of IL-4 andGM-CSFwere products of85 PeproTech Inc. (USA). Other chemicals which are frequently used at our86 laboratory were all purchased from Sigma-Aldrich or BD Pharmingen.
87 2.2. The preparation of BMDCs
88 BMDCs were prepared with the method described previously [8].89 All animals were treated kindly per the Guide for the Care and Use90 of Laboratory Animals of ChinaMedical University and all the surgical91 procedures were approved by The Committee of Experimental Animals92 of China Medical University. All mice were anesthetized by intra-93 peritoneal injection of Inactin (thiobutabarbital, 100mg/kg; RBI, Natick,94 MA) and ventilated with oxygen (35%) and air mixture.95 Bone marrow cells from the femurs and tibias of female C57BL/696 mice (4–6 weeks old) were depleted of red cells with lyses buffer97 solution. About 107/ml cells were obtained and placed in 24-well plates98 in RPMI 1640 media supplemented with 10% fetal bovine serum99 (10 ng/ml recombinant murine GM-CSF, 10 ng/ml IL-4, 100 units/ml100 penicillin, 100 #g/ml streptomycin and 2 mM L-glutamine). After101 incubation for 4 h, the medium containing non-adherent cells was102 replaced with fresh medium. Then, after 7 d of culture, the cells were103 kept with LPS (1 #g/ml) to generate more cells before puri!cation.104 CD11c+ BMDCs were then puri!ed by magnetic sorting, using anti-105 CD11c-coated magnetic beads and the auto-MACS system per the106 manufacturer's instruction (MiltenyiQ11 Biotec, CA, USA). The purity of107 the sorted cells was con!rmed with FCM (N90% for CD11c+ cells).
108 2.3. Transmitted electron microscopy (TEM) for intracellular changes
109 The cultured BMDCs, post-treatment with125 #g/ml LDN for 48 h,110 were collected for morphology study by TEM. Next the BMDCs were111 spun down, resuspended in 0.5 ml 0.05 M pH 7.2 PBS, and !xed112 overnight in a 2.5% solution of aqueous glutaraldehyde. This process113 was followed by a treatment with 1% osmium tetroxide dehydrated in114 ethanol and embedded in epon. The sections were cut on a Reichert-115 JungQ12 Ultracut E, stained with uranyl acetate and lead citrate. Finally,116 the sample was checked under TEM (JEOL JEM-1200EX).
117 2.4. Analysis of phenotypic marker by FCM
118 The cultured BMDC cells treated with125 #g/ml LDN for 48 h were119 collected for analysis of phenotypic marker. The BMDCs were rinsed120 with PBS and incubated with anti-CD40, anti-CD80, anti-CD86, anti-121 CD83 and anti-MHC II antibodies for 20 min at 4 °C. After extensive122 rinsing, the stained cells were analyzed using FACS Calibur (Becton123 Dickinson, San Diego, CA).
124 2.5. Cytokine assay of IL-12p40, IL-12p70 and TNF-!
125 The BMDC cells incubated with 125 #g/ml LDN for 96 h were126 collected for the assay of production of IL-12p70 and TNF-! respectively127 according to instruction included in the ELISA kit. The absorbance at128 450 nm (A450) was determined using a bichromatic microplate reader129 (BIO-TEK, USA).
130 2.6. Mixed lymphocyte reaction (MLR)
131 For themixed lymphocyte reaction (MLR) the BMDCpost-treatment132 with125 #g/ml LDN was harvested on 7 d. The CD8+ T cells from the
133splenocytes were separated and puri!ed with magnetic beads under134sterile condition and subsequently the puri!ed CD8+ T cells were135grown with 10 #g/ml conA for 48 h to an enriched concentration of136107/ml. The puri!ed CD8+ T cells (5! 105/well) were incubated with137graded numbers of BMDCs (at a ratio of 1:1, 1:5, 1:10, 1:50, 1:100) in13896-well culture plates (Corning-Costar) for 5 d. The proliferation of T139cells was monitored by measuring the absorbance at 492 nm (A492)140using a bichromatic microplate reader. The results were expressed as141mean±sd from triplicate wells.
1422.7. Immunohistochemical staining of phagocytic process by BMDCs
143BMDCs (1 ! 105/ml), treated with125 #g/ml LDN for 48 h were144incubatedwith 0.08mg/ml horseradish peroxidase,!xedwithmethanol145for 4h, stained with DAB, and examined under the microscope to locate146the phagocytosis process.
1472.8. Phagocytosis study by FCM
148The BMDCs incubated with 125 #g/ml LDN for 48 h were collected149and 100 #l FITC-Dextran (40,000D) 28–30 was added to the culture for1502 h at 4 °C and subsequently for 1 h at 37 °C. Finally the sample151underwent analysis using the FACS Calibur (Becton Dickinson, San152Diego, CA).
1532.9. ACP activity detection
154The number of BMDCswas adjusted to 1!106/ml. ACP activity in the155BMDCswith 1.25#g/ml IFN-$ for 48hwas tested at OD520nm (A520) by156the phenol-4-AAP (amino anti-pyrine) method combined with ACP157testing kit (Jiancheng Bio-engineering Institute of the South) [18].
1582.10. Statistical analysis
159All results were processed using statistical program SPSS (Statistical160Package for Social Sciences, Version 16.0) for Windows. The variables161were presented as mean ± sd. The differences evaluated by ANOVA162when p value was b0.05, indicate signi!cance.
1633. Results
1643.1. BMDC responses to a range of LDN dosage and time
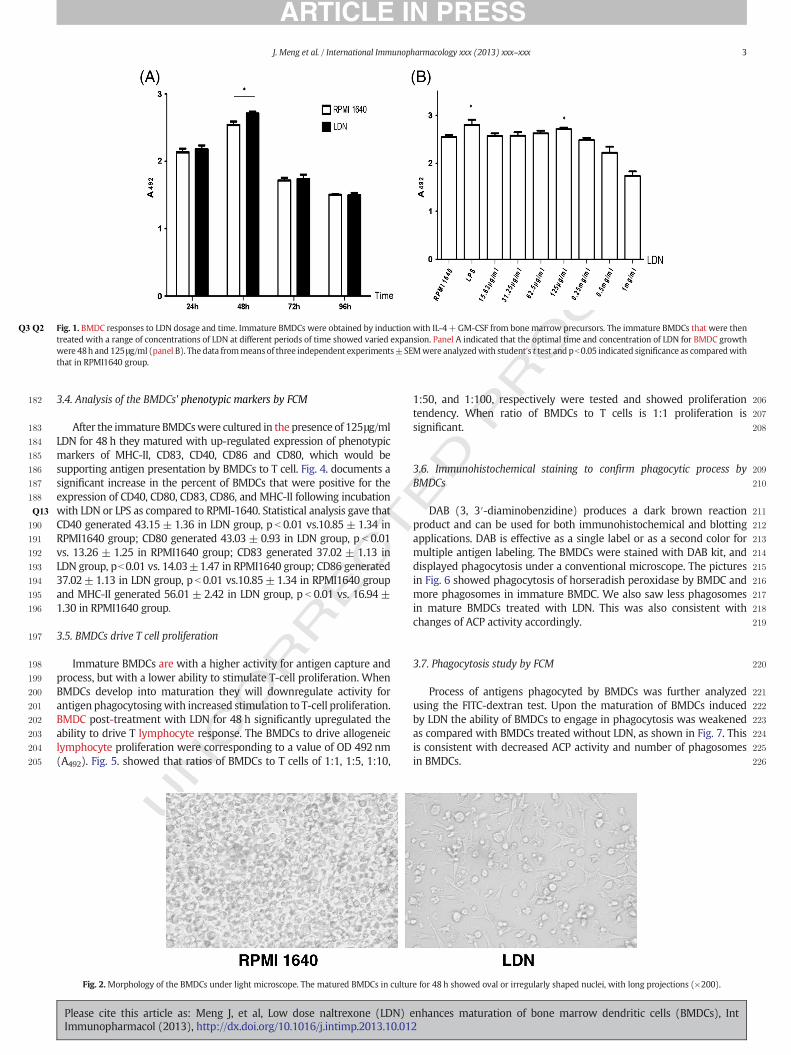
165A kinetic study of BMDC expansion was assessed and immature166BMDCs in the presence of a range of concentrations of LDN expand167into differential number at different time. The optimal boosting time168shown in Fig. 1A was 48 h and the best concentration of LDN to exert169stimulation on BMDC growth was 125 #g/ml (Fig. 1B).
1703.2. Changes of BMDCs' morphology under microscope
171After culture with 125 #g/ml LDN for 48 h, the BMDCs displayed a172typical matured morphology. Under light microscope, the BMDCs173exhibited many dendrites with rough surfaces and long protrusions174and extensions (Fig. 2).
1753.3. Intracellular changes inside the BMDCs under TEM
176Immature BMDCs possess a strong ability to phagocyte antigen and177there are a larger number of phagosomes inside the immature BMDC.178However, when BMDCsmature, the number of phagosomeswill reduce,179accompanying increased antigen presentation supported by higher180expression of phenotypic markers. Fig. 3 showed that there were181reduced numbers of phagosomes inside BMDCs.
2 J. Meng et al. / International Immunopharmacology xxx (2013) xxx–xxx
Please cite this article as: Meng J, et al, Low dose naltrexone (LDN) enhances maturation of bone marrow dendritic cells (BMDCs), IntImmunopharmacol (2013), http://dx.doi.org/10.1016/j.intimp.2013.10.012
UNCO
RRECTED P
RO
OF
182 3.4. Analysis of the BMDCs' phenotypic markers by FCM
183 After the immature BMDCswere cultured in the presence of 125#g/ml184 LDN for 48 h they matured with up-regulated expression of phenotypic185 markers of MHC-II, CD83, CD40, CD86 and CD80, which would be186 supporting antigen presentation by BMDCs to T cell. Fig. 4. documents a187 signi!cant increase in the percent of BMDCs that were positive for the188 expression of CD40, CD80, CD83, CD86, and MHC-II following incubation189 with LDN or LPS as compared to RPMI-1640. Statistical analysis gaveQ13 that190 CD40 generated 43.15± 1.36 in LDN group, p b 0.01 vs.10.85 ± 1.34 in191 RPMI1640 group; CD80 generated 43.03 ± 0.93 in LDN group, p b 0.01192 vs. 13.26 ± 1.25 in RPMI1640 group; CD83 generated 37.02 ± 1.13 in193 LDN group, pb0.01 vs. 14.03±1.47 in RPMI1640 group; CD86 generated194 37.02±1.13 in LDN group, p b 0.01 vs.10.85±1.34 in RPMI1640 group195 and MHC-II generated 56.01 ± 2.42 in LDN group, p b 0.01 vs. 16.94 ±196 1.30 in RPMI1640 group.
197 3.5. BMDCs drive T cell proliferation
198 Immature BMDCs are with a higher activity for antigen capture and199 process, but with a lower ability to stimulate T-cell proliferation. When200 BMDCs develop into maturation they will downregulate activity for201 antigen phagocytosingwith increased stimulation to T-cell proliferation.202 BMDC post-treatment with LDN for 48 h signi!cantly upregulated the203 ability to drive T lymphocyte response. The BMDCs to drive allogeneic204 lymphocyte proliferation were corresponding to a value of OD 492 nm205 (A492). Fig. 5. showed that ratios of BMDCs to T cells of 1:1, 1:5, 1:10,
2061:50, and 1:100, respectively were tested and showed proliferation207tendency. When ratio of BMDCs to T cells is 1:1 proliferation is208signi!cant.
2093.6. Immunohistochemical staining to con!rm phagocytic process by210BMDCs
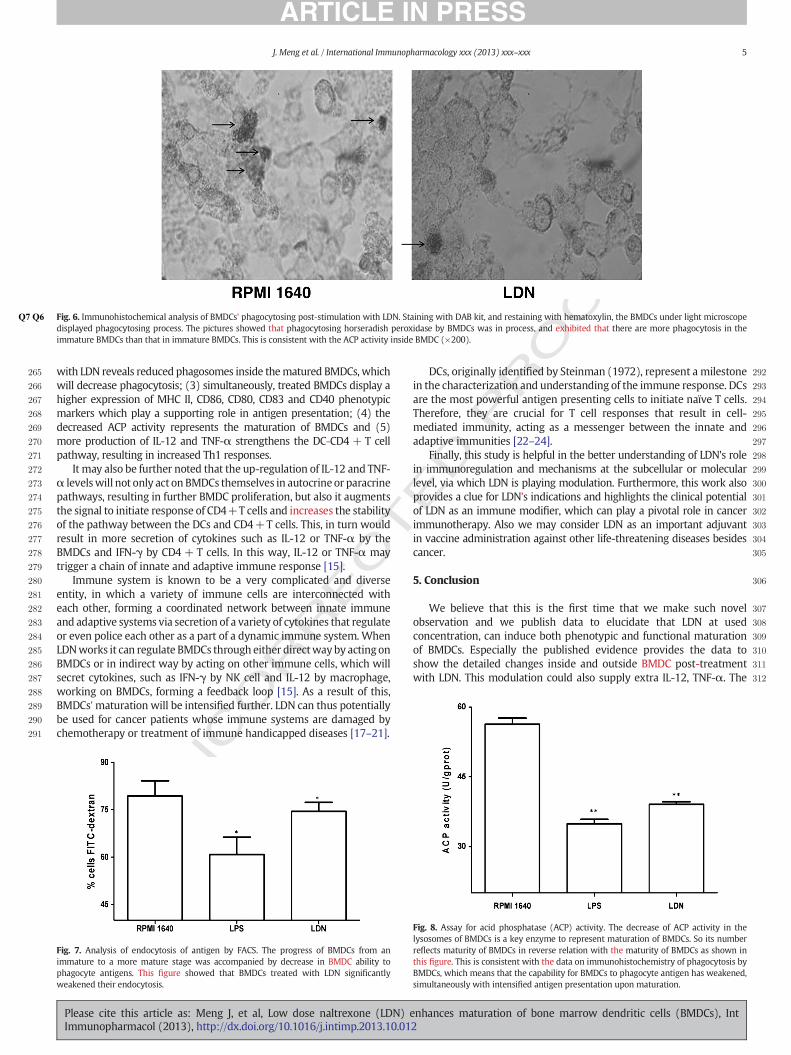
211DAB (3, 3"-diaminobenzidine) produces a dark brown reaction212product and can be used for both immunohistochemical and blotting213applications. DAB is effective as a single label or as a second color for214multiple antigen labeling. The BMDCs were stained with DAB kit, and215displayed phagocytosis under a conventional microscope. The pictures216in Fig. 6 showed phagocytosis of horseradish peroxidase by BMDC and217more phagosomes in immature BMDC. We also saw less phagosomes218in mature BMDCs treated with LDN. This was also consistent with219changes of ACP activity accordingly.
2203.7. Phagocytosis study by FCM
221Process of antigens phagocyted by BMDCs was further analyzed222using the FITC-dextran test. Upon the maturation of BMDCs induced223by LDN the ability of BMDCs to engage in phagocytosis was weakened224as compared with BMDCs treated without LDN, as shown in Fig. 7. This225is consistent with decreased ACP activity and number of phagosomes226in BMDCs.
Fig. 1.Q2Q3 BMDC responses to LDN dosage and time. Immature BMDCs were obtained by induction with IL-4+GM-CSF from bonemarrow precursors. The immature BMDCs that were thentreated with a range of concentrations of LDN at different periods of time showed varied expansion. Panel A indicated that the optimal time and concentration of LDN for BMDC growthwere 48h and 125#g/ml (panel B). The data frommeans of three independent experiments±SEMwere analyzedwith student's t test and pb0.05 indicated signi!cance as comparedwiththat in RPMI1640 group.
Fig. 2.Morphology of the BMDCs under light microscope. The matured BMDCs in culture for 48 h showed oval or irregularly shaped nuclei, with long projections (!200).
3J. Meng et al. / International Immunopharmacology xxx (2013) xxx–xxx
Please cite this article as: Meng J, et al, Low dose naltrexone (LDN) enhances maturation of bone marrow dendritic cells (BMDCs), IntImmunopharmacol (2013), http://dx.doi.org/10.1016/j.intimp.2013.10.012
UNCO
RRECTED P
RO
OF
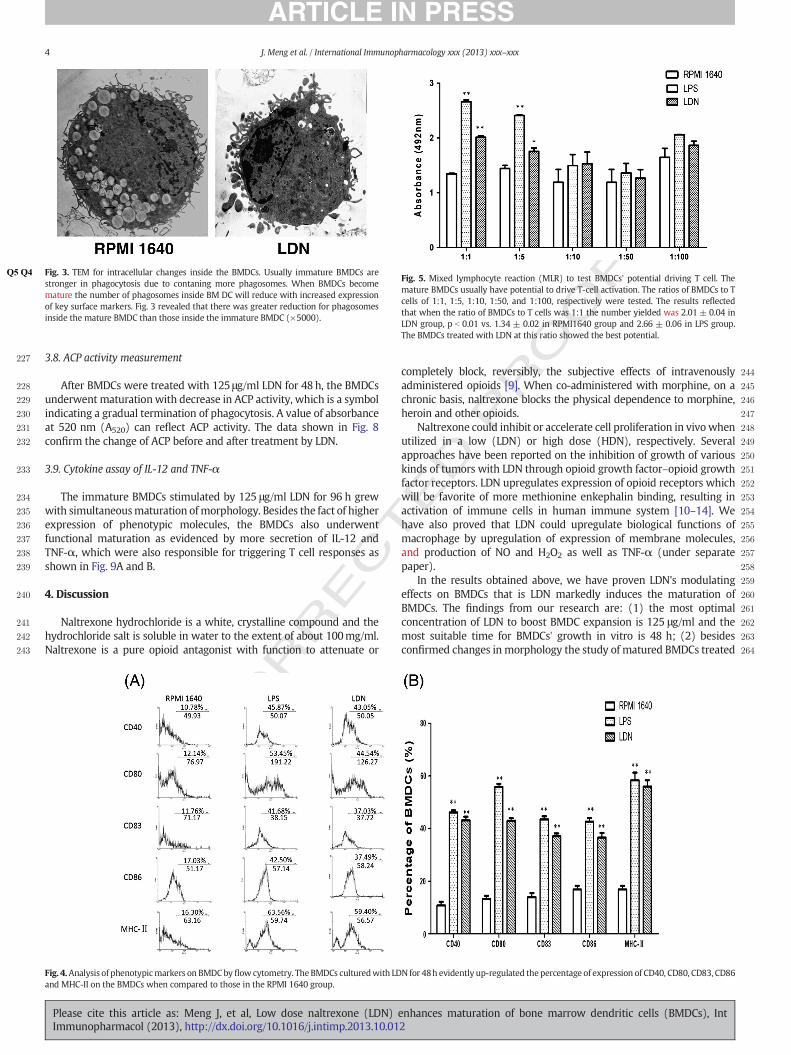
227 3.8. ACP activity measurement
228 After BMDCs were treated with 125 #g/ml LDN for 48 h, the BMDCs229 underwent maturationwith decrease in ACP activity, which is a symbol230 indicating a gradual termination of phagocytosis. A value of absorbance231 at 520 nm (A520) can re"ect ACP activity. The data shown in Fig. 8232 con!rm the change of ACP before and after treatment by LDN.
233 3.9. Cytokine assay of IL-12 and TNF-!
234 The immature BMDCs stimulated by 125 #g/ml LDN for 96 h grew235 with simultaneousmaturation ofmorphology. Besides the fact of higher236 expression of phenotypic molecules, the BMDCs also underwent237 functional maturation as evidenced by more secretion of IL-12 and238 TNF-!, which were also responsible for triggering T cell responses as239 shown in Fig. 9A and B.
240 4. Discussion
241 Naltrexone hydrochloride is a white, crystalline compound and the242 hydrochloride salt is soluble in water to the extent of about 100mg/ml.243 Naltrexone is a pure opioid antagonist with function to attenuate or
244completely block, reversibly, the subjective effects of intravenously245administered opioids [9]. When co-administered with morphine, on a246chronic basis, naltrexone blocks the physical dependence to morphine,247heroin and other opioids.248Naltrexone could inhibit or accelerate cell proliferation in vivo when249utilized in a low (LDN) or high dose (HDN), respectively. Several250approaches have been reported on the inhibition of growth of various251kinds of tumors with LDN through opioid growth factor–opioid growth252factor receptors. LDN upregulates expression of opioid receptors which253will be favorite of more methionine enkephalin binding, resulting in254activation of immune cells in human immune system [10–14]. We255have also proved that LDN could upregulate biological functions of256macrophage by upregulation of expression of membrane molecules,257and production of NO and H2O2 as well as TNF-! (under separate258paper).259In the results obtained above, we have proven LDN's modulating260effects on BMDCs that is LDN markedly induces the maturation of261BMDCs. The !ndings from our research are: (1) the most optimal262concentration of LDN to boost BMDC expansion is 125 #g/ml and the263most suitable time for BMDCs' growth in vitro is 48 h; (2) besides264con!rmed changes in morphology the study of matured BMDCs treated
Fig. 3.Q4Q5 TEM for intracellular changes inside the BMDCs. Usually immature BMDCs arestronger in phagocytosis due to contaning more phagosomes. When BMDCs becomemature the number of phagosomes inside BM DC will reduce with increased expressionof key surface markers. Fig. 3 revealed that there was greater reduction for phagosomesinside the mature BMDC than those inside the immature BMDC (!5000).
Fig. 4.Analysis of phenotypicmarkers on BMDCby "ow cytometry. The BMDCs culturedwith LDN for 48h evidently up-regulated the percentage of expression of CD40, CD80, CD83, CD86and MHC-II on the BMDCs when compared to those in the RPMI 1640 group.
Fig. 5. Mixed lymphocyte reaction (MLR) to test BMDCs' potential driving T cell. Themature BMDCs usually have potential to drive T-cell activation. The ratios of BMDCs to Tcells of 1:1, 1:5, 1:10, 1:50, and 1:100, respectively were tested. The results re"ectedthat when the ratio of BMDCs to T cells was 1:1 the number yielded was 2.01± 0.04 inLDN group, p b 0.01 vs. 1.34 ± 0.02 in RPMI1640 group and 2.66 ± 0.06 in LPS group.The BMDCs treated with LDN at this ratio showed the best potential.
4 J. Meng et al. / International Immunopharmacology xxx (2013) xxx–xxx
Please cite this article as: Meng J, et al, Low dose naltrexone (LDN) enhances maturation of bone marrow dendritic cells (BMDCs), IntImmunopharmacol (2013), http://dx.doi.org/10.1016/j.intimp.2013.10.012
UNCO
RRECTED P
RO
OF
265 with LDN reveals reduced phagosomes inside thematured BMDCs,which266 will decrease phagocytosis; (3) simultaneously, treated BMDCs display a267 higher expression of MHC II, CD86, CD80, CD83 and CD40 phenotypic268 markers which play a supporting role in antigen presentation; (4) the269 decreased ACP activity represents the maturation of BMDCs and (5)270 more production of IL-12 and TNF-! strengthens the DC-CD4 + T cell271 pathway, resulting in increased Th1 responses.272 It may also be further noted that the up-regulation of IL-12 and TNF-273 ! levelswill not only act on BMDCs themselves in autocrine or paracrine274 pathways, resulting in further BMDC proliferation, but also it augments275 the signal to initiate response of CD4+T cells and increases the stability276 of the pathway between the DCs and CD4+T cells. This, in turn would277 result in more secretion of cytokines such as IL-12 or TNF-! by the278 BMDCs and IFN-$ by CD4 + T cells. In this way, IL-12 or TNF-! may279 trigger a chain of innate and adaptive immune response [15].280 Immune system is known to be a very complicated and diverse281 entity, in which a variety of immune cells are interconnected with282 each other, forming a coordinated network between innate immune283 and adaptive systems via secretion of a variety of cytokines that regulate284 or even police each other as a part of a dynamic immune system. When285 LDNworks it can regulate BMDCs through either directwayby acting on286 BMDCs or in indirect way by acting on other immune cells, which will287 secret cytokines, such as IFN-$ by NK cell and IL-12 by macrophage,288 working on BMDCs, forming a feedback loop [15]. As a result of this,289 BMDCs' maturation will be intensi!ed further. LDN can thus potentially290 be used for cancer patients whose immune systems are damaged by291 chemotherapy or treatment of immune handicapped diseases [17–21].
292DCs, originally identi!ed by Steinman (1972), represent a milestone293in the characterization and understanding of the immune response. DCs294are the most powerful antigen presenting cells to initiate naïve T cells.295Therefore, they are crucial for T cell responses that result in cell-296mediated immunity, acting as a messenger between the innate and297adaptive immunities [22–24].298Finally, this study is helpful in the better understanding of LDN's role299in immunoregulation and mechanisms at the subcellular or molecular300level, via which LDN is playing modulation. Furthermore, this work also301provides a clue for LDN's indications and highlights the clinical potential302of LDN as an immune modi!er, which can play a pivotal role in cancer303immunotherapy. Also we may consider LDN as an important adjuvant304in vaccine administration against other life-threatening diseases besides305cancer.
3065. Conclusion
307We believe that this is the !rst time that we make such novel308observation and we publish data to elucidate that LDN at used309concentration, can induce both phenotypic and functional maturation310of BMDCs. Especially the published evidence provides the data to311show the detailed changes inside and outside BMDC post-treatment312with LDN. This modulation could also supply extra IL-12, TNF-!. The
Fig. 6.Q6Q7 Immunohistochemical analysis of BMDCs' phagocytosing post-stimulation with LDN. Staining with DAB kit, and restaining with hematoxylin, the BMDCs under light microscopedisplayed phagocytosing process. The pictures showed that phagocytosing horseradish peroxidase by BMDCs was in process, and exhibited that there are more phagocytosis in theimmature BMDCs than that in immature BMDCs. This is consistent with the ACP activity inside BMDC (!200).
Fig. 7. Analysis of endocytosis of antigen by FACS. The progress of BMDCs from animmature to a more mature stage was accompanied by decrease in BMDC ability tophagocyte antigens. This !gure showed that BMDCs treated with LDN signi!cantlyweakened their endocytosis.
Fig. 8. Assay for acid phosphatase (ACP) activity. The decrease of ACP activity in thelysosomes of BMDCs is a key enzyme to represent maturation of BMDCs. So its numberre"ects maturity of BMDCs in reverse relation with the maturity of BMDCs as shown inthis !gure. This is consistent with the data on immunohistochemistry of phagocytosis byBMDCs, which means that the capability for BMDCs to phagocyte antigen has weakened,simultaneously with intensi!ed antigen presentation upon maturation.
5J. Meng et al. / International Immunopharmacology xxx (2013) xxx–xxx
Please cite this article as: Meng J, et al, Low dose naltrexone (LDN) enhances maturation of bone marrow dendritic cells (BMDCs), IntImmunopharmacol (2013), http://dx.doi.org/10.1016/j.intimp.2013.10.012
UNCO
RRECTED P
RO
OF
313 importance of this work should be further emphasized so that more314 work would be done in depth toward the application of LDN for a315 number of immune related diseases.
316 6.Q14 Uncited reference
317 [16]
318 Acknowledgment
319 This study was supported by the Natural Science Foundation of320 Liaoning Province (No. 201102250). We apologize to the colleagues321 whose work could not be discussed due to space limitation.
322 References
323 [1] Kirst A, Wack C, Lutz WK, Eggert A, Kämpgen E, Fischer WH. Expression of324 functional kappa-opioid receptors on murine dendritic cells. Immunol Lett325 2002 Oct 21;84(1):41–8.326 [2] Mazlan M, Schottenfeld RS, Chawarski MC. New challenges and opportunities in327 managing substance abuse in Malaysia. Drug 2006 Sep;25(5):473–8.328 [3] Brown N, Panksepp J. Low-dose naltrexone for disease prevention and quality of life.329 Med Hypotheses 2009 Mar;72(3):333–7.330 [4] McLaughlin PJ, Rogosnitzky M, Zagon IS. Inhibition of DNA synthesis in mouse331 epidermis by topical imiquimod is dependent on opioid receptors. Exp Biol Med332 (Maywood) 2010 Nov;235(11):1292–9.333 [5] Steinman R, Cohn Z. Identi!cation of a novel cell type in peripheral lymphoid organs334 of mice. J Exp Med 1973;137:1142–62 (Integr Cancer Ther. 2009 Dec;8(4):416-22).335 [6] Wojas-Turek J, Pajtasz-Piasecka E, Rossowska J, Piasecki E, Du! D. Antitumor effect of336 murine dendritic and tumor cells transduced with IL-2 gene. Folia Histochem337 Cytobiol 2012 Oct 8;50(3):414–9.338 [7] Gopalan Soman, Xiaoyi Yang, Hengguang Jiang, Steve Giardina, Vinay Vyas,339 George Mitra, et al. MTS dye based colorimetric CTLL-2 cell proliferation340 assay for product release and stability monitoring of interleukin-15: assay341 quali!cation, standardization and statistical analysis. J Immunol Methods Aug.342 2009;348(1–2):83–94.343 [8] Segovia M, Cuturi MC, Hill M. Preparation of mouse bone marrow-derived dendritic344 cells with immunoregulatory properties. Methods Mol Biol 2011;677:161–8.345 [9] Sakurada S, Sawai T, Mizoguchi H, Watanabe H, Watanabe C, Yonezawa A, et al.346 Possible involvement of dynorphin A release via mu1-opioid receptor on347 supraspinal antinociception of endomorphin-2. Peptides 2008 Sep;29(9):1554–60.
348[10] Berkson BM, Rubin DM, Berkson AJ. Revisiting the ALA/N (alpha-lipoic acid/low-349dose naltrexone) protocol for people with metastatic and nonmetastatic pancreatic350cancer: a report of 3 new cases. Methods Mol Biol 2011;677:161–8.351[11] Amirshahrokhi K, Dehpour AR, Hadjati J, SotoudehM, Ghazi-KhansariM.Methadone352ameliorates multiple-low-dose streptozotocin-induced type 1 diabetes in mice.353Toxicol Appl Pharmacol 2008;232(1):119–24. http://dx.doi.org/10.1016/j.taap.3542008.06.020 (Epub 2008 Jul 11.).355[12] Zagon IS, Rahn KA, Turel AP, McLaughlin PJ. Endogenous opioids regulate expression356of experimental autoimmune encephalomyelitis: a new paradigm for the treatment357of multiple sclerosis. Exp Biol Med (Maywood) 2009 Nov;234(11):1383–92.358[13] Davis M, Goforth HW, Gamier P. Oxycodone combined with opioid receptor359antagonists: ef!cacy and safety. Expert Opin Drug Saf 2013 May;12(3):389–402.360[14] Donahue RN, McLaughlin PJ, Zagon IS. Low-dose naltrexone targets the opioid361growth factor-opioid growth factor receptor pathway to inhibit cell proliferation:362mechanistic evidence from a tissue culture model. Exp Biol Med (Maywood) 2011363Sep 1;236(9):1036–50.364[15] Shan F, Xia Y, Wang N, Meng J, Lu C, Meng Y, et al. Functional modulation of the365pathway between dendritic cells (DCs) and CD4+T cells by the neuropeptide:366methionine enkephalin (MENK). Peptides 2011 May;32(5):929–37.367[16] Jiang CY, Wang W, Tang JX, Yuan ZR. The adipocytokine resistin stimulates368production of proin"ammatory cytokines TNF-! and IL-6 in pancreatic acinar cells369via NF-%B activation. J Endocrinol Invest 2013 Jun 10. Q15370[17] Donahue RN, McLaughlin PJ, Zagon IS. Low-dose naltrexone suppresses ovarian371cancer and exhibits enhanced inhibition in combination with cisplatin. Exp Biol372Med (Maywood) 2011 Jul 1;236(7):883–95.373[18] Donahue RN, McLaughlin PJ, Zagon IS. The opioid growth factor (OGF) and low dose374naltrexone (LDN) suppress human ovarian cancer progression in mice. Gynecol375Oncol 2011 Aug;122(2):382–8.376[19] Berkson BM, Rubin DM, Berkson AJ. The long-term survival of a patient with377pancreatic cancer with metastases to the liver after treatment with the378intravenous alpha-lipoic acid/low-dose naltrexone protocol. Integr Cancer379Ther 2006 Mar;5(1):83–9.380[20] McLaughlin PJ, Immonen JA, Zagon IS. Topical naltrexone accelerates full-thickness381wound closure in type 1 diabetic rats by stimulating angiogenesis. Exp Biol Med382(Maywood) 2013 Jun 20 [Epub ahead of print]. Q16383[21] Colquhoun RM. Open label trial of naltrexone implants: measuring blood serum384levels of naltrexone. Subst Abus 2013 May 15;7:75–84.385[22] Liu J, Chen W, Meng J, Lu C, Wang E, Shan F. Induction on differentiation and386modulation of bone marrow progenitor of dendritic cell by methionine enkephalin387(MENK). Cancer Immunol Immunother 2012 Oct;61(10):1699–711.388[23] Xue M, Zhu L, Meng Y, Wang L, Sun H, Wang F, et al. Detailed modulation of389phenotypes and functions of bone marrow dendritic cells (BMDCs) by interferon-390gamma (IFN-$). Int Immunopharmacol 2013 Jul 16;17(2):366–72.391[24] Meng Y, Wang Q, Zhang Z, Wang E, Plotnikoff NP, Shan F. Synergistic effect of392methionine encephalin (MENK) combined with pidotimod (PTD) on thematuration393of murine dendritic cells (DCs). Hum Vaccin Immunother 2013 Mar 7;9(4).
394
Fig. 9. Secretion of IL-12 and TNF-! determined by ELISA. Uponmaturation of BMDCs, besides upregulated expression of keymembranemolecules they secrete higher levels of cytokinesIL-12 and TNF-!. This !gure showed that under the in"uence of LDN the BMDCs increased secretion of IL-12 and TNF-!, which will be involved in intensifying T cell response.
6 J. Meng et al. / International Immunopharmacology xxx (2013) xxx–xxx
Please cite this article as: Meng J, et al, Low dose naltrexone (LDN) enhances maturation of bone marrow dendritic cells (BMDCs), IntImmunopharmacol (2013), http://dx.doi.org/10.1016/j.intimp.2013.10.012
Copyright © 2022 FDOKUMEN




















