
Despite Corona, Haryana govt maintained good financial ... - Yugmarg
Upload
independentCategory
view
1download
0
Loss of Oocytes in Dazl Knockout Mice Results inMaintained Ovarian Steroidogenic Function but AlteredGonadotropin Secretion in Adult Animals
J. R. MCNEILLY, P. T. K. SAUNDERS, M. TAGGART, M. CRANFIELD,H. J. COOKE, AND A. S. MCNEILLY
Medical Research Council Human Reproductive Sciences Unit, University of Edinburgh Center forReproductive Biology (J.R.M., P.T.K.S., A.S.M.), Edinburgh, Scotland EH3 9ET; Medical ResearchCouncil Human Genetics Unit, Western General Hospital (M.T., H.J.C.), Edinburgh, Scotland EH42XU; and Center for Proteins and Peptides, School of Biological and Molecular Sciences, OxfordBrookes University (M.C.), Headington, Oxford, United Kingdom OX3 0BP
ABSTRACTWithin 2 days of birth, the mouse ovary is mainly composed of
oocytes surrounded by a few pregranulosa cells forming primordialfollicles that remain quiescent until they are recruited by intraovar-ian or other unknown factors to initiate growth of the oocyte andproliferation of the attendant granulosa cells. However, the role of theoocyte in this early development and organization of the follicle ispoorly understood. The Dazl knockout (2/2) mouse in which there istotal ablation of oocytes in fetal life has allowed us to address thisissue. Ovaries from 2/2 females lack any follicular structure andhave no cells positive for either Mullerian inhibiting factor or sulfatedglycoprotein-1, indicating a lack of small follicles or corpora lutea.However, by immunocytochemistry, there are cells positive for 3b-hydroxysteroid dehydrogenase, 17a-hydroxylase, and aromatase, in-dicating the presence of steroidogenically active cells capable of pro-ducing estrogen. This was confirmed by the presence of hypertrophied
uterine endometrium expressing both estrogen receptor a (ERa) andERb together with normal levels of plasma estradiol. In addition,these steroidogenically active cells contain ERb, inhibin a, and bB-subunits, and 2/2 mice have low measurable plasma inhibin A andB levels. The ovarian steroids and inhibins had no significant effecton either plasma or pituitary gonadotropin levels, with significantly(P , 0.01) lower LH and FSH in intact 1/1 and 1/2 females. How-ever, significantly (P , 0.05) increased plasma inhibin B togetherwith significantly (P , 0.05) lower FSH were observed in the 1/2females. In conclusion, our data showed that despite oocyte loss infetal life, the adult ovaries contained steroidogenically active cellscapable of producing estradiol and inhibin. Furthermore, in the 1/2mice, the enhanced plasma inhibin B implies a role for Dazl proteinwithin the oocyte either from more small follicles or increased inhibinB production from each follicle. (Endocrinology 141: 4284–4294,2000)
IN THE MAMMALIAN female, fertility requires the pro-duction of mature oocytes, and it is by the process of
folliculogenesis that mature fertilizable oocytes are pro-duced. Within the ovary, each follicle consists of an oocytesurrounded by somatic cells. A complex cascade of eventsinvolving both intra- and extraovarian factors leads to ovu-lation. During this period, there is coordination between thegrowth and development of the oocyte, differentiation of thesomatic cells to become granulosa cells or thecal cells, fol-lowed by proliferation and development to produce the ste-roidogenically active follicle (1, 2).
In the mouse during fetal life, the ovary consists of somaticcells arranged in cords, with each cord surrounded by mes-enchymal cells and filled with masses of rete ovary-derivedcells and oocytes (3). Within the first 3 days of birth, pri-mordial follicles are formed that consist of oocytes with at-tendant somatic squamous pregranulosa cells (4, 5). Almostimmediately, folliculogenesis is initiated, and a subset ofthese primordial follicles is recruited and begins to grow. The
mechanisms involved in this initial recruitment are as yetunknown; however, studies in the mouse, rat, cow, and hu-man have demonstrated that although follicle growth can beinitiated in the absence of gonadotropins, few follicles de-velop beyond two layers of granulosa cells (6–9). However,the availability of transgenic mouse models of ovarian failuremay help to identify the intraovarian factors involved (10).The rate of recruitment of primordial follicles into the growthphase is greatest between 7–21 days and thereafter decreasesand remains stable for the rest of the reproductive life of theanimal (11). Until the formation of an antrum, regulation ofdevelopment appears to be intrafollicular with various cru-cial factors, e.g. growth differentiation factor-9 (GDF-9) andKit ligand, involved in signaling from the oocyte to the gran-ulosa cells and vice versa (12–14). In fact, an oocyte is neces-sary for preantral growth up to the induction of a thecal layer,which occurs once the follicle has achieved two layers ofgranulosa cells. This thecal layer provides a source of aro-matizable androgen to the adjacent granulosa cells essentialfor estrogen production (15–17).
Available evidence suggests that the presence of oocytesis critical for the induction of effective thecal-granulosa cellinteraction and hence for the initiation of steroidogenesis(18). The requirement of oocytes for the maintenance of ste-roidogenesis is not clear. The Dazl knockout (2/2) mouse
Received April 17, 2000.Address all correspondence and requests for reprints to: Dr. J. R.
McNeilly, Medical Research Council, Human Reproductive SciencesUnit, University of Edinburgh Center for Reproductive Biology, 37Chalmers Street, Edinburgh, Scotland EH3 9ET. E-mail: [email protected].
0013-7227/00/$03.00/0 Vol. 141, No. 11Endocrinology Printed in U.S.A.Copyright © 2000 by The Endocrine Society
4284

offers us a unique opportunity to address this question. Al-though on embryonic day 15 the ovaries appear normal in2/2 females, there is a marked reduction in oocyte numberson E19 and a complete absence of follicles and ova in theadult ovary (19). The aim of this study was to determine theeffect of early oocyte loss on the capacity of the ovary toproduce steroids and inhibins and the consequences onplasma and pituitary gonadotropin secretion using intactand ovariectomized adult 2/2, heterozygous (1/2), andwild-type (1/1) Dazl females.
Materials and MethodsAnimals and experimental design
In all studies the female mice used were from lines generated by aconventional knock out approach in which the Dazl gene was disruptedby a neomycin-resistant marker after homologous recombination inembryonic stem (ES) cells, causing the loss of exons 6 and 7 and all butfive amino acids of exon 5. Chimeric males were generated by selectionof ES cells containing the targeted mutation using a PCR-based assay.Mating these males to MF1 females generated heterozygotes that whenintercrossed gave homozygous (2/2) animals (7). Animals wereweaned at 3 weeks of age and genotyped at 6 weeks by extraction of tailDNA and subsequent PCR. They were maintained on a 14-h light, 10-hdark photoperiod under normal animal facility conditions. All studieswere approved by the Home Office (United Kingdom) and undertakenunder license. Adult females (8–12 weeks of age) expressing all threegenotypes (1/1, 1/2, and 2/2 for the Dazl gene) were caged insibling groups and age matched before this study. Vaginal smears werecollected daily for at least 7 days, equivalent to at least one estrous cyclefrom intact females of all three genotypes (n 5 6–10/genotype), todetermine whether they were exhibiting normal estrous cycles. Duringthe next cycle, 1/2 and 1/1 animals only were killed by CO2 as-phyxiation when an estrogenic type smear, similar to that observed inall 2/2 animals, was observed. The final vaginal smear was air-dried,fixed for 1–2 min in 70% ethanol, then stained with hematoxylin, de-hydrated, and mounted in Pertex (Cellpath, Hemel Hempstead, UK) asa record of vaginal cytology at the time of tissue collection. Bloodsamples were collected by cardiac puncture after killing the animals withCO2, and plasma was separated by centrifugation at 3000 rpm for 15 minand frozen at 220 C until assayed for LH, FSH, inhibin A and B,estradiol, and progesterone. Pituitaries were collected and stored at 220C until homogenized in 0.5 ml PBS (Sigma, St. Louis, MO) before assayfor LH and FSH. Ovaries and uteri were removed, weighed, and fixedin Bouin’s fixative for 2–5 h, then transferred to 70% ethanol. Concur-rently, females (n 5 6–10) from the three genotypes had the stage of theestrous cycle determined by vaginal smear before being ovariectomizedunder IsoFlo anesthesia (Mallinckrodt, Inc., Harefield, UK). After re-moval, the ovaries were weighed and fixed in Bouin’s as previously
described. Seven days after surgery the animals were killed by CO2, andplasma, pituitaries, and uteri were collected as previously described.Ovaries were also removed from day 16 females after animals werekilled by CO2 and were fixed in Bouin’s fixative; the animals weregenotyped retrospectively from tail DNA and PCR as describedpreviously.
Immunocytochemistry
After processing, ovaries and uteri were embedded in paraffin waxand sectioned at 5 mm thickness before detection of the antigens listedin Table 1 (20–24). Antigen retrieval (25) was required for the successfuldetection of some antigens using 0.01 m citrate pH 6, or 0.05 m glycine-EDTA, pH 3.5. Before incubation of the sections with primary antibody,all slides were blocked with either 1:5 normal rabbit serum/Tris-buff-ered saline if the primary antibody was monoclonal or 1:5 normal swineserum/Tris-buffered saline if it was polyclonal. Similarly, primary an-tibodies, diluted in the appropriate blocking buffer, were added to thesections, which were coverslipped with Gelbond (FMC Bioproducts,Rockland, ME) before incubation overnight at 4 C. Detection using abiotinylated second antibody (swine antirabbit or rabbit antisheep forpolyclonal primary antibodies; rabbit antimouse for monoclonal pri-mary antibodies; DAKO Corp., Copenhagen, Denmark) was followed bythe avidin-biotin-horseradish peroxidase system (DAKO Corp.) andvisualized by 39,39-diaminobenzidine. For sections incubated with anti-Mullerian hormone/Mullerian inhibiting substance (AMH/MIS) anti-serum, the avidin-biotin-alkaline phosphatase system (DAKO Corp.)was used with nitro blue tetrazolium (Sigma) and x-phosphatase (5-bromo-4-chloro-3-indolyl-phosphate; Sigma) visualization. Control sec-tions were incubated with normal rabbit serum or normal mouse serumin place of the primary antibody. The specificity of the antibodies wasconfirmed in previous studies using antigen preadsorption. All sectionswere counterstained with hematoxylin, then dehydrated and mountedin Pertex (Cell Path, Hemel Hempstead, UK). Sections were photo-graphed using an Olympus Corp. Provis microscope (New Hyde Park,NY), and Kodak 420 digital camera (Eastman Kodak, Inc., Rochester,NY), and montages were assembled using Photoshop 5 (Adobe Systems,Inc., San Jose, CA).
Immunoassays
The concentrations of plasma and pituitary LH and FSH were mea-sured by RIA using reagents supplied by the NIDDK with all samplesfor each hormone assayed in duplicate in one assay. The referencepreparations used were rat LH RP-1 and rat FSH RP-3, and the minimumdetectable concentrations were 0.2 and 1.2 ng/ml for LH and FSH,respectively. The intraassay coefficients of variation were less than 6%(26). As the volume of plasma available for assay was limited, not allsamples were assayed for inhibin A and B, estradiol, and progesterone;however, there were sufficient samples to allow meaningful values (n 54–6) to be obtained in each treatment group. In some cases the samples
TABLE 1. Antisera used for immunocytochemical studies on ovarian and uterine tissues
Antibody against Source (Ref. no.) Polyclonal ormonoclonal
Antigenretrieval Dilution Tissue
ERa Novocastro M Yes, c 1:20 UterusERb Saunders (20) P Yes, ge 1:1000 Uterus/ovaryPCNA Dako M No 1:100 Ovary3bHSD Mason (21) P No 1.1000 Ovary17a-Hydroxylase Mason (21) P No 1:500 OvaryAMH/MIS Rey (22)a P Yes, c 1:500 OvarySGP-1 Sylvester (23)b P Yes, c 1:100 OvaryAromatase Groomec M No 1:10 OvaryInhibin a Groome (24) M Yes, c 1:450 OvaryInhibin bB Groome (24) M Yes, c 1:3000 Ovary
ERa, Estrogen receptor a; ERb, estrogen receptor b; PCNA, proliferating cell nuclear antigen; 3bHSD, 3b-hydroxysteroid dehydrogenase;AMH, anti-Mullerian hormone; SGP-1, sulfated glycoprotein-1; c, 0.01m citrate buffer, pH 6.0; ge, 0.05 M Glycine and EDTA, pH 3.5.
a Gift to Dr. P. T. K. Saunders.b Gift to Dr. R. M. Sharpe.c Saunders, P. T. K., and A. S. McNeilly, unpublished observations.
OVARIAN STEROIDOGENIC FUNCTION IN DAZL KNOCKOUT MICE 4285

were assayed as single samples. Inhibins A and B were measured usingtwo-site enzyme-linked immunosorbent assays (27, 28) and were vali-dated for mouse plasma (29). All samples were assayed in a single assay,and the minimum detectable concentrations were 1 pg/ml (inhibin A)and 8 pg/ml (inhibin B), respectively, with coefficients of variation ofless than 10%.
Estradiol and progesterone were determined after solvent extractionusing sensitive RIA methods modified for use with mouse and sheepplasma, as previously described (30, 31). All samples were measured inthe same assay with a coefficients of variation of less than 8%, and theminimum detectable concentrations for estradiol and progesterone were4.6 pg/ml and 0.5 ng/ml, respectively.
Statistical analysis
All data were analyzed by ANOVA and Student’s paired t tests usingthe GB Stat program (Dynamic Microsystem, Inc., Silver Spring, MD).P # 0.05 compared with the appropriate control was considered sta-tistically significant.
ResultsEffect of genotype on uterine morphology
The degree of hypertrophy of the uterine endometriumgives an indication of the circulating estrogenic steroid level.
FIG. 1. A, Changes in uterine wet weights in wild-type (1/1), heterozygous (2/1), and homozygous (2/2) Dazl-1 knockout mice before(f) and after (M) ovariectomy. a– c, Significant differences between uteri from ovary-intact mice; d–f, significant differences afterovariectomy. B, Immunostaining for ERa (B, a and b) and ERb (B, c and d) in the uteri removed from these intact wild-type (WT) 1/1and knockout (KO) 2/2 adult mice. There was no difference among 1/1, 1/2 (results not shown), and 2/2 in the distribution of ERaor ERb immunostaining.
4286 OVARIAN STEROIDOGENIC FUNCTION IN DAZL KNOCKOUT MICE Endo • 2000Vol. 141 • No. 11

FIG. 2. Morphological appearance of ovaries in adult 1/1, 1/2, and 2/2 Dazl mice. Sections were immunostained with antibodies directedagainst PCNA (a and b), AMH/MIS (c, e, and g), and SGP-1 (d, f, and h). Granulosa cells surrounding preantral as well as antral (*) follicleswere present in ovaries from both 1/1 (a, c, and d) and 1/2 (e and f) females, whereas no follicular structures were present in ovaries lackingDazl (2/2; b, g, and h). Granulosa cells in developing follicles were immunopositive for PCNA (a, arrows) consistent with a high rate of celldivision in the somatic cells of the follicle, whereas very few PCNA-positive cells (b, arrows) were present in ovaries from 2/2 mice. Thehistological appearance of ovaries from 1/1 and 1/2 mice were indistinguishable; both contained small follicles (arrowheads) in which granulosacells were immunopositive for AMH/MIS and contained antral follicles (*), the granulosa cells of which did not express the protein. Stromalcells (S) and corpora lutea were immunopositive for SGP-1. In contrast, ovaries from 2/2 females did not contain organized structures expressingeither AMH/MIS (g) or SGP-1 (h). Magnifications: a and b, 320; c, d, e, f, g, h, 310.
OVARIAN STEROIDOGENIC FUNCTION IN DAZL KNOCKOUT MICE 4287

FIG. 3. Expression of steroidogenic enzymes and ERb within the cytoplasm of cells in ovaries from both 1/1 and 2/2 mice. Immunoexpressionof 3bHSD in the 1/1 ovary (a) was intense both within the stroma and in thecal cells surrounding the developing follicles, and expression inGC was initiated in antral follicles (*). Expression of 3bHSD was maintained in the 2/2 ovaries (b) and was localized to groups of cells, thesize and abundance of which varied within the ovarian structures between animals. Similarly, 17a-hydroxylase expression was in only thecaland interstitial stroma (s) in 1/1 ovaries (c), with no immunopositive cells observed in the granulosa cells (*); however, in the 2/2 animals
4288 OVARIAN STEROIDOGENIC FUNCTION IN DAZL KNOCKOUT MICE Endo • 2000Vol. 141 • No. 11

Despite a lack of oocytes, uteri from intact 2/2 females weregrossly similar to those from intact estrous 1/2 females,with no differences in endometrial thickness between thegenotypes noted. However, uteri from intact estrous 1/1females were significantly heavier (P , 0.01 and P , 0.05,respectively) compared with those from 2/2 and 1/2 an-imals (Fig. 1A, wet weight). There was a similar pattern ofexpression of estrogen receptor a (ERa; Fig. 1B, a and b) andERb (Fig. 1B, c and d) in all three genotypes, with intensestaining of both glandular and stromal endometrium.
After ovariectomy, there was a highly significant (P , 0.01)decrease in uterine wet weights in all three genotypes (Fig.1A). Interestingly, uteri from 2/2 females weighed signif-icantly (P , 0.05) less than those from both 1/1 and 1/2after ovariectomy. There was no significant difference inuterine weight between ovariectomized 1/2 and 1/1 fe-males, although the 1/2 females tended to be lighter. Therewas no difference in body weight among the three genotypes.
Functional competence of ovarian cells and expression ofsteroidogenic enzymes
Evidence that the 2/2 ovary, despite a lack of oocytes,secretes sufficient estrogenic steroid to maintain a prolifer-ative uterine endometrium led us to investigate the func-tional activity of the cells in the 2/2 ovaries compared withthose in the other two genotypes. Cellular proliferation, asdetermined by immunocytochemistry using proliferatingcell nuclear antigen (PCNA) antiserum (Fig. 2, a and b)showed few dividing cells in the 2/2 ovary, whereas in the1/1 and 1/2 ovaries (1/2 data not shown), extensivecellular proliferation was observed in all follicular tissues,consistent with the presence of actively proliferating gran-ulosa cells with a much lower rate of cell division in theovarian stroma. This absence of functionally competent fol-licular tissue was confirmed by the presence of few MIS-positive cells in the 2/2 ovaries (Fig. 2g), with MIS beingexpressed only in granulosa cells of small follicles in normal1/1 and 1/2 ovaries (Fig. 2, c and e). Similarly, stromalcells and corpora lutea were immunopositive for sulfatedglycoprotein-1 (SGP-1) in 1/1 and 1/2 ovaries (Fig. 2, dand f), whereas ovaries from 2/2 females did not containany organized structures that expressed SGP-1 (Fig. 2h).
Ovarian sections from 1/1 and 1/2 females, stained for3b-hydroxysteroid dehydrogenase (3bHSD), showed in-tense cytoplasmic staining in both stromal and thecal cells.The first appearance of 3bHSD-immunopositive granulosacells was in antral follicles (Fig. 3a; data for 1/2 ovaries notshown). In ovarian tissue from 2/2 females (Fig. 3b) groupsof intensely stained cells were dispersed throughout the tis-sue, consistent with a lack of structural organization. Theintensity of staining and the variation in size of these ste-roidogenically active cells varied among 2/2 animals (data
not shown). The pattern of expression of 17a-hydroxylasewas identical to that of 3bHSD in 1/1 (Fig. 3c), 1/2, and2/2 (Fig. 3d) mouse ovaries.
In ovaries from 1/1 and 1/2 females, aromatase cyto-chrome P450-positive cells were present in the peripheralgranulosa cells of large preovulatory follicles, with the mostintense staining observed at the granulosa/thecal junction(Fig. 3e, arrowhead). In corpora lutea, there was variable ex-pression of aromatase ranging from immunonegative tostrongly immunopositive cells (Fig. 3g, arrows). The potentialof cells within the 2/2 ovaries to synthesize estrogen wasendorsed by the detection of aromatase-positive cells in 2/2ovarian tissue (Fig. 3, f and h). The intensity of staining andits distribution in the ovarian mass varied between ovaries,with some positive cells forming clusters as shown in Fig. 3g.ERb was expressed in the nuclei of granulosa cells through-out follicular development in both 1/1 (Fig. 3i) and 1/2ovaries and in occasional clusters of cells within 2/2 ovaries(Fig. 3j).
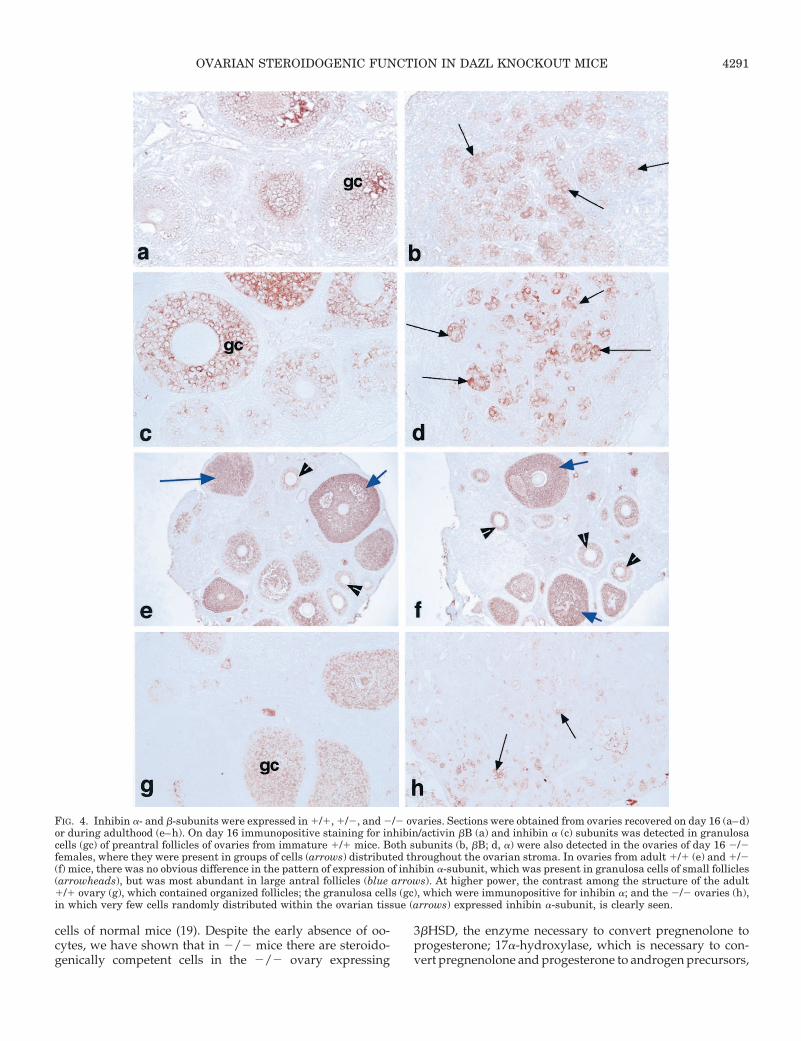
In ovaries collected from 1/1 and 1/2 females on day16 postpartum, there were many preantral follicles with in-tensely positive stained granulosa cells for both activin/inhibin bB and inhibin a (Fig. 4, a and c; data for 1/2 notshown). Both subunits were also detected in 2/2 ovaries,with groups of immunopositive staining cells distributedthroughout the ovarian stroma (Fig. 4, b and d). In adult 2/2ovaries, although a few inhibin a-positive cells were present(Fig. 4h), they were weakly stained and tended to lack thecluster formation observed in the day 16 postpartum tissue.In both 1/1 and 1/2 ovaries, inhibin a was strongly ex-pressed in granulosa cells (Fig. 4, e–g).
Plasma gonadotropin concentrations
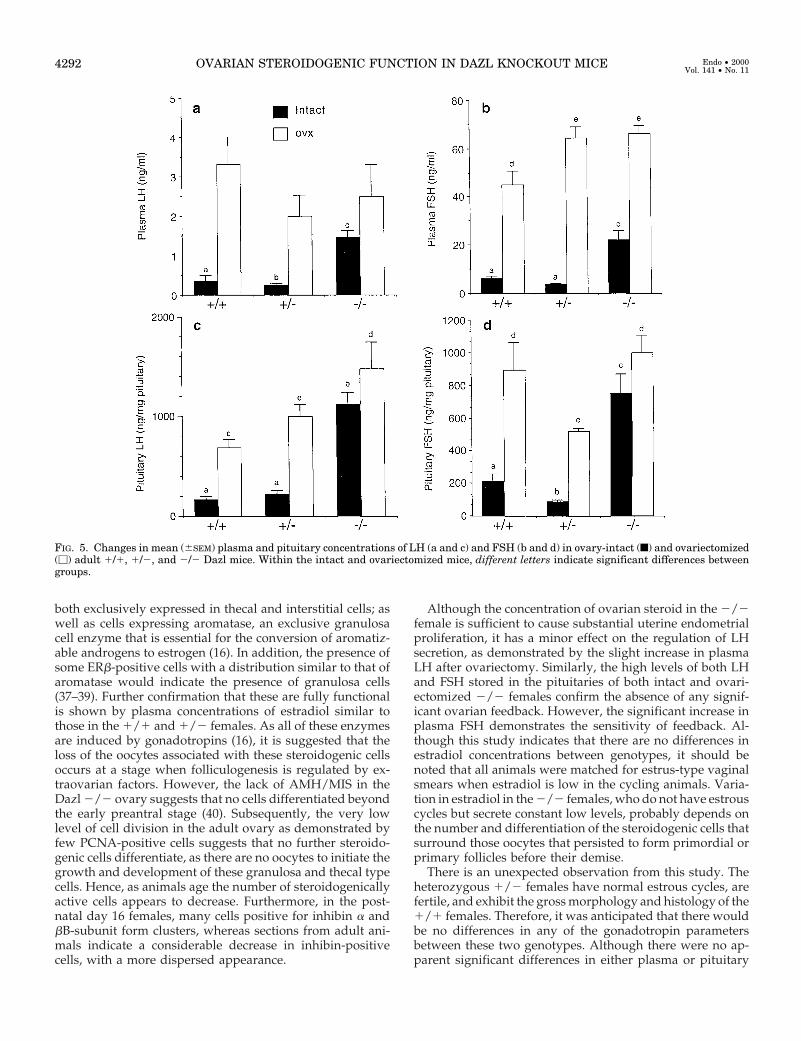
LH. Plasma and pituitary LH concentrations in intact andovariectomized females are shown in Fig. 5, a and c. In intact2/2 females, plasma LH concentrations were significantly(P , 0.01) elevated compared with those in both 1/1 intactand 1/2 intact females (Fig. 4a). After ovariectomy, plasmaLH concentrations in 1/1 and 1/2 females were signifi-cantly (intact 1/1 vs. ovariectomized 1/1, P , 0.01; intact1/2 vs. ovariectomized 1/2, P , 0.01) higher than theirrespective intact levels. The increase in plasma LH in ovari-ectomized 1/2 females tended to be lower than those in theother two groups. Furthermore, ovariectomy in the 2/2females did not lead to an increase in plasma LHconcentrations.
Pituitary LH in intact females from all three genotypesmirrored the secretion pattern of plasma LH with pituitariesfrom intact 2/2 females containing highly significantly (P ,0.01) greater amounts of LH than those from 1/1 and 1/2females. After ovariectomy, pituitary LH levels in 1/1 and1/2 females rose significantly (P , 0.01) compared with
(d), groups of positive cells were observed throughout the ovarian tissue. In ovaries from 1/1 females, expression of aromatase was observedwithin GC of large antral follicles, where it was more abundant in those cells close to the thecal layer (e, arrowheads). In corpora lutea (g), variableimmunopositive staining was observed, ranging from immunonegative (§) to strongly immunopositive (arrows). Expression of aromatase wasobserved in groups of cells in all ovaries from 2/2 females, but the intensity of immunopositive staining varied between ovaries from weak (f,arrowheads) to strong (h, arrows). Granulosa cells (*) from 1/1 ovaries 1) expressed abundant ERb from small preantral to large preovulatoryfollicles. In 2/2 ovaries (j), expression of ERb was localized to clusters of cells (arrowheads), which varied throughout the tissue. ERb-immunopositive cells were also observed in the oviduct (od). Magnification, all 340.
OVARIAN STEROIDOGENIC FUNCTION IN DAZL KNOCKOUT MICE 4289

levels in their respective intact groups. As observed in thepattern of plasma LH secretion, only in ovariectomized 2/2females was there no significant increase in pituitary LH. Nodifferences in pituitary LH were observed between 1/1 and1/2 females, either intact or ovariectomized; however, therewas significantly (P , 0.05) less LH in 1/1 pituitaries afterovariectomy than in pituitaries from 2/2 females.
FSH. Plasma and pituitary FSH concentrations are shown inFig. 4, b and d, respectively. A pattern of secretion similar tothat of LH was noted, with no difference in plasma FSH in1/1 and 1/2 intact females, but intact 2/2 females hadsignificantly (P , 0.01) elevated circulating FSH. Despite thisincreased FSH secretion, there was a significant (P , 0.01)rise in plasma FSH in 2/2 females after ovariectomy, whichwas similar to that observed in 1/2 females. Although therewas a significant (P , 0.01) increase in plasma FSH in ovari-ectomized 1/1 females, it was significantly (P , 0.05) lowerthan that observed in both 1/2 and 2/2 females. It isinteresting to note that the greatest postovariectomy rise inplasma FSH concentrations was observed in the 1/2females.
Pituitary FSH was significantly (P , 0.01) higher in intact2/2 females than in intact 1/1 and 1/2 females. Al-though both plasma and pituitary FSH in 1/2 females ap-peared lower than the 1/1 value, they failed to achievesignificance. Ovariectomy resulted in a rise in pituitary FSHin all genotypes; however, only in the 1/1 and 1/2 femaleswas this rise significant (P , 0.01) compared with those intheir respective intact groups. Although there appeared to bean increase in pituitary FSH after ovariectomy in the 2/2females compared with their intact value, this was not sig-nificant. However, the increase in pituitary FSH in 1/2females after ovariectomy was significantly (P , 0.01) lowerthan that in the 1/1 females. There was no difference be-tween FSH content in 1/1 and 2/2 pituitaries afterovariectomy.
Inhibin and ovarian steroids. Plasma levels of inhibin A and Bin intact females from all genotypes are shown in Fig. 6, a andb. In 2/2 females, both inhibin A and B concentrations wereextremely low, with some samples being below the mini-mum detectable limit of the assays (inhibin A, 3 of 10; inhibinB, 3 of 16). The remaining values were significantly (P , 0.01)lower than those detected in the other two groups. Althoughthere was no difference in inhibin A between 1/1 and 1/2females, significantly (P , 0.05) higher levels of inhibin Bwere present in 1/2 females compared with those in both1/1 and 2/2 animals.
Despite the lack of oocytes in 2/2 females, there was nosignificant difference in the plasma concentrations of eitherestradiol or progesterone between the groups, with bothsteroids being detectable in all samples in intact females fromall three genotypes (Fig. 6, c and d).
Discussion
In the present study we have shown that in the completeabsence of oocytes, there are cells present within the adultovary that can produce estradiol and progesterone togetherwith low measurable inhibin A and B and cause the prolif-
eration of the uterine endometrium. This shows for the firsttime that some development of the granulosa and thecal-likecells can be maintained in the absence of factors from theoocyte, although no functional organization of the follicleoccurs. In fact, it is possible that once these cells are differ-entiated they maintain their steroidogenic function until theydie, as there appear to be fewer inhibin-positive cells in theadult ovary compared with those in day 16 females. In theDazl 2/2 mouse, oocyte loss starts and chronologicallywould appear to be completed before the initiation of fol-licular growth. In the absence of Dazl protein expression, thedeveloping oocyte on embryonic day 15 enters meiosis, butis unable to progress further, and most are lost by embryonicday 19 with no follicular development seen on postnatal day4 (our unpublished observation). It should be noted that theDazl gene is a member of the DAZ/SPGY family of genes,which, like the RBM genes, are located on the Y-chromosomein Old World primates and apes, but are represented by theautosomal gene Dazl in others mammals. The function ofDazl protein is unknown; however, as it is cytoplasmic andhas RNA binding motifs it may be involved in translationalcontrol (19).
At birth in the mouse, only “naked” oocytes are present,which by 2 days postpartum associate with three to fivepregranulosa cells and become primordial follicles (4, 5).Follicular growth proceeds with a change in the morphologyof these somatic cells followed by an increase in the numberof granulosa cells, leading to the formation of multiple layers.When there are two layers of follicle cells present, fibroblast-like cells are recruited from the interstitium to encircle thefollicle and form the thecal layer. All of these events appearto be regulated primarily by intraovarian factors and requirethe presence of an oocyte (2), and this oocyte-granulosa cellinteraction is exemplified by the expression of c-Kit ligand ingranulosa cells and c-Kit receptor in the oocyte (12, 13, 32).The importance of the oocyte in the proliferation and dif-ferentiation of granulosa cells is illustrated in the GDF-9knockout mouse, where follicles arrest after two mitotic di-visions of granulosa cells but remain at this stage, with theoocyte overgrowing until cell death (33). Interestingly, al-though granulosa cells express FSH receptor, and thecal cellsexpress LH receptor, there is no evidence for any gonado-tropin requirement for follicle growth and development atthis preantral stage (2, 34). However, in large preantral andsmall antral follicles, it is probably the presence of thesefunctional receptors that marks the change from predomi-nately intra- to both intra- and extraovarian gonadotropinregulation. The absence of normal large antral follicles andthe dramatic reduction in the number of early antral folliclesin both FSH-knockout and hpg mice support the concept thatfollicles do not require FSH for development to the earlyantral stage (35, 36). In addition, only those follicles able torespond to rising FSH levels continue to develop, while themajority of early antral follicles become atretic (2). In normalmice, all oocytes have entered prophase of meiosis 1 bypostnatal day 2, and the minority that do not become atreticcomplete meiosis at ovulation. However, in the Dazl 2/2mouse, oocytes enter meiosis and then arrest and are de-stroyed, possibly by atresia, due to the lack of Dazl protein,which is found in the cytoplasm of fetal ovarian pachytene
4290 OVARIAN STEROIDOGENIC FUNCTION IN DAZL KNOCKOUT MICE Endo • 2000Vol. 141 • No. 11
cells of normal mice (19). Despite the early absence of oo-cytes, we have shown that in 2/2 mice there are steroido-genically competent cells in the 2/2 ovary expressing
3bHSD, the enzyme necessary to convert pregnenolone toprogesterone; 17a-hydroxylase, which is necessary to con-vert pregnenolone and progesterone to androgen precursors,
FIG. 4. Inhibin a- and b-subunits were expressed in 1/1, 1/2, and 2/2 ovaries. Sections were obtained from ovaries recovered on day 16 (a–d)or during adulthood (e–h). On day 16 immunopositive staining for inhibin/activin bB (a) and inhibin a (c) subunits was detected in granulosacells (gc) of preantral follicles of ovaries from immature 1/1 mice. Both subunits (b, bB; d, a) were also detected in the ovaries of day 16 2/2females, where they were present in groups of cells (arrows) distributed throughout the ovarian stroma. In ovaries from adult 1/1 (e) and 1/2(f) mice, there was no obvious difference in the pattern of expression of inhibin a-subunit, which was present in granulosa cells of small follicles(arrowheads), but was most abundant in large antral follicles (blue arrows). At higher power, the contrast among the structure of the adult1/1 ovary (g), which contained organized follicles; the granulosa cells (gc), which were immunopositive for inhibin a; and the 2/2 ovaries (h),in which very few cells randomly distributed within the ovarian tissue (arrows) expressed inhibin a-subunit, is clearly seen.
OVARIAN STEROIDOGENIC FUNCTION IN DAZL KNOCKOUT MICE 4291
both exclusively expressed in thecal and interstitial cells; aswell as cells expressing aromatase, an exclusive granulosacell enzyme that is essential for the conversion of aromatiz-able androgens to estrogen (16). In addition, the presence ofsome ERb-positive cells with a distribution similar to that ofaromatase would indicate the presence of granulosa cells(37–39). Further confirmation that these are fully functionalis shown by plasma concentrations of estradiol similar tothose in the 1/1 and 1/2 females. As all of these enzymesare induced by gonadotropins (16), it is suggested that theloss of the oocytes associated with these steroidogenic cellsoccurs at a stage when folliculogenesis is regulated by ex-traovarian factors. However, the lack of AMH/MIS in theDazl 2/2 ovary suggests that no cells differentiated beyondthe early preantral stage (40). Subsequently, the very lowlevel of cell division in the adult ovary as demonstrated byfew PCNA-positive cells suggests that no further steroido-genic cells differentiate, as there are no oocytes to initiate thegrowth and development of these granulosa and thecal typecells. Hence, as animals age the number of steroidogenicallyactive cells appears to decrease. Furthermore, in the post-natal day 16 females, many cells positive for inhibin a andbB-subunit form clusters, whereas sections from adult ani-mals indicate a considerable decrease in inhibin-positivecells, with a more dispersed appearance.
Although the concentration of ovarian steroid in the 2/2female is sufficient to cause substantial uterine endometrialproliferation, it has a minor effect on the regulation of LHsecretion, as demonstrated by the slight increase in plasmaLH after ovariectomy. Similarly, the high levels of both LHand FSH stored in the pituitaries of both intact and ovari-ectomized 2/2 females confirm the absence of any signif-icant ovarian feedback. However, the significant increase inplasma FSH demonstrates the sensitivity of feedback. Al-though this study indicates that there are no differences inestradiol concentrations between genotypes, it should benoted that all animals were matched for estrus-type vaginalsmears when estradiol is low in the cycling animals. Varia-tion in estradiol in the 2/2 females, who do not have estrouscycles but secrete constant low levels, probably depends onthe number and differentiation of the steroidogenic cells thatsurround those oocytes that persisted to form primordial orprimary follicles before their demise.
There is an unexpected observation from this study. Theheterozygous 1/2 females have normal estrous cycles, arefertile, and exhibit the gross morphology and histology of the1/1 females. Therefore, it was anticipated that there wouldbe no differences in any of the gonadotropin parametersbetween these two genotypes. Although there were no ap-parent significant differences in either plasma or pituitary
FIG. 5. Changes in mean (6SEM) plasma and pituitary concentrations of LH (a and c) and FSH (b and d) in ovary-intact (f) and ovariectomized(M) adult 1/1, 1/2, and 2/2 Dazl mice. Within the intact and ovariectomized mice, different letters indicate significant differences betweengroups.
4292 OVARIAN STEROIDOGENIC FUNCTION IN DAZL KNOCKOUT MICE Endo • 2000Vol. 141 • No. 11

LH, surprisingly, both pituitary and plasma concentrationsof FSH were significantly lower in 1/2 than 1/1 animals.This was related to a large increase in plasma concentrationsof inhibin B. As inhibin B is produced mainly by small fol-licles (41), the present results suggest that there may be anincrease in the pool of small follicles in 1/2 ovaries. How-ever, the lack of change in inhibin A, secreted preferentiallyby the larger follicles (41), suggests that the increased num-bers of small follicles do not all progress to become preovu-latory follicles. However, a significant (P , 0.05) increase inthe ovulation rate in second litters from 1/2 compared withthat in the 1/1 females (McNeilly, J. R., unpublished data)indicates that more follicles may become ovulatory in 1/2than 1/1 animals. The reasons for the increase in inhibin Band the relation to the numbers of small follicles requirefurther investigation.
As a model to investigate follicular steroidogenesis in theabsence of oocytes, the Dazl 2/2 mouse is unique, in thatthere is maintenance in the adult of some thecal and gran-ulosa steroidogenically and inhibin-active cells in the com-plete absence of oocytes from the early neonatal period. Thiscontrasts with other mouse knockout and mutant models,which are associated with ovarian failure [Kit ligand-Steel1(42), Kit ligand-Steel panda (12), connexin 43 knockout (43),ERa knockout (44), FSH knockout (35), FSH receptor knock-out (45), and GDF-9 knockout (46)] in which oocytes remainpresent throughout adult life, and the cellular organizationof small follicles remains intact. In contrast, in the absence ofoocytes in the Dazl 2/2 mouse, the ovary becomes disor-
ganized, and the steroidogenically and inhibin-active cellsappear randomly distributed throughout the ovary.
In conclusion, the present study indicates, firstly, that de-spite the absence of factors from the oocyte, once steroido-genically active follicle cells have become differentiated toproduce estrogen, they can maintain their steroid outputthroughout the lifetime of the cell. Furthermore, even thoughoocytes die from embryonic day 15 on, surrounding somaticcells differentiate sufficiently to become steroidogenicallycompetent. Secondly, we have shown that low levels of Dazlprotein in the 1/2 females are associated with the apparentsurvival of more small follicles. Additional studies to addressthis issue by counting the number of follicles are planned.
Acknowledgments
The authors acknowledge the expert technical assistance of Ian Swan-ston, Ted Pinner, and Tom McFetters for graphics, and the AnimalHouse staff, especially Denis Doogan and Maureen Ross. Some of thereagents for the RIAs were provided by the NIDDK through the NationalHormone and Pituitary Program and Dr. A. F. Parlow.
References
1. Richards JS 1980 Maturation of ovarian follicles: actions and interaction ofpituitary and ovarian hormones on follicular cell differentiation. Physiol Rev60:51–69
2. Richards JS 1994 Hormonal control of gene expression Endocr Rev 15:725–7513. Byskov AG 1978 The anatomy and ultrastructure of the rete system in the fetal
mouse ovary. Biol Reprod 19:720–7354. Hirschfield AN, DeSanti AM 1995 Patterns of ovarian cell proliferation in rats
during the embryonic period and the first three weeks postpartum. Biol Re-prod 53:1208–1221
FIG. 6. Changes in mean (6SEM) plasma concentrations of inhibin A (a), inhibin B (b), estradiol (E2; c), and progesterone (d) in intact adultfemale 1/1, 1/2, and 2/2 Dazl mice. Different letters indicate significant differences between groups.
OVARIAN STEROIDOGENIC FUNCTION IN DAZL KNOCKOUT MICE 4293

5. Lintern-Moore S, Moore GPM 1979 Initiation of follicle and oocyte growth inthe mouse ovary. Biol Reprod 20:773–778
6. Wang XN, Greenwald GS 1993 Hypophysectomy of the cyclic mouse. I. Effectson folliculogenesis, oocyte growth and follicle stimulating hormone and hu-man chorionic gonadotropin receptors. Biol Reprod 45:585–594
7. Braw-Tal R, Yossefi S 1997 Studies in vivo and in vitro on the initiation offollicle growth in the bovine ovary. J Reprod Fertil 109:165–171
8. Oktay K, Newton H, Mullan J, Gosden RG 1998 Development of humanprimordial follicles to antral stages in SCID/hpg mice stimulated with follicle-stimulating hormone. Hum Reprod 13:1133–1138
9. McGee EA, Hsueh AJW 2000 Initial and cyclic recruitment of ovarian follicles.Endocr Rev 21:200–214
10. Elvin JS, Matzuk MM 1998 Mouse models of ovarian failure Rev Reprod3:183–195
11. Greenwald GS, Roy SK 1994 Follicular selection and its control. In: Knobil E,Neill J (eds) The Physiology of Reproduction. Raven Press, New York, pp629–724
12. Huang E, Manova K, Packer A, Sanchez S, Bachvarova R, Besmer P 1993 Themurine steel panda mutation affects kit ligand expression and growth of earlyovarian follicles. Dev Biol 157:100–109
13. Joyce IM, Pendola FL, Wigglesworth K, Eppig JJ 1999 Oocyte regulation ofkit ligand expression in mouse ovarian follicles. Dev Biol 214:342–353
14. Elvin JA, Changning Y, Wang P, Nishimori K, Matzuk MM 1999 Molecularcharacterization of the follicle defects in the growth differentiation factor-9deficient ovary Mol Endocrinol 13:1018–1034
15. Fortune JE, Kito S, Bird DD 1999 Activation of primordial follicles in vitro.J Reprod Fertil [Suppl] 54:439–448
16. Gore-Langton RE, Armstrong DT 1994 Follicular steroidogenesis and its con-trol. In: Knobil E, Neill J (eds) The Physiology of Reproduction. Raven Press,New York, pp 571–627
17. Magoffin D, Weitsman SR 1993 Differentiation of ovarian theca-interstitialcells in vitro: regulation of 17a hydroxylase messenger ribonucleic acid ex-pression by LH and insulin-like growth factor-1. Endocrinology 132:1945–1951
18. Hirshfield A 1991 Development of follicles in the mammalian ovary. Int RevCytol 124:43–101
19. Ruggiu M, Speed R, Taggart M, McKay SJ, Kilanowski F, Saunders P, DorinJ, Cooke HJ 1997 The mouse Dazla gene encodes a cytoplasmic protein es-sential for gametogenesis Nature 389:77–79
20. Saunders PTK, Maguire SM, Gaughan J, Millar MR 1997 Expression ofoestrogen receptor beta (ERb) in multiple rat tissues visualised by immuno-histochemistry J Endocrinol 154:R13–R16
21. Bird IM, Pasquarette MM, Rainey WE, Mason JI 1996 Differential control of17a-hydroxylase and 3b-hydroxysteroid dehydrogenase expression in humanadrenocortical H295R cells. J Clin Endocrinol Metab 81:2171–2178
22. Frojdman K, Pelliniemi LJ, Rey R, Virtanen I 1999 Presence of anti-Mullerianhormone correlates with absence of laminin alpha 5 chain in differentiating rattestis and ovary. Histochem Cell Biol 111:367–373
23. Sylvester SR, Morales C, Oko R, Griswold MD 1989 Sulfated glycoprotein-1(saposin precursor) in the reproductive tract of the male rat. Biol Reprod41:941–948
24. Majdic G, McNeilly AS, Sharpe RM, Evans LR, Groome NP, Saunders PTK1997 Testicular expression of inhibin and activin subunits and follistatin in therat and human fetus and neonate and during postnatal development in the rat.Endocrinology 138:2136–2147
25. Norton AJ, Jordon S, Yeomans P 1994 Brief high-temperature heat denatur-ation (pressure cooking): a simple and effective method of antigen retrieval forroutinely processed tissues. J Pathol 173:371–377
26. McNeilly JR, Brown P, Mullins JJ, Clark AJ, McNeilly AS 1996 Characteri-sation of the ovine LHb-subunit gene: the promoter is regulated by GnRH andgonadal steroids in transgenic mice. J Endocrinol 151:35–43
27. Groome NP, Illingworth PJ, O’Brien M, Cooke I, Ganesan TS, Baird DT,McNeilly AS 1994 Detection of dimeric inhibin throughout the human men-
strual cycle by two-site enzyme immunoassay. Clin Endocrinol (Oxf)40:717–723
28. Groome NP, Illingworth PJ, O’Brien M, Pai R, Rodger FE, Mather J, Mc-Neilly AS 1996 Measurement of dimeric inhibin-B throughout the humanmenstrual cycle. J Clin Endocrinol Metab 81:1401–1405
29. Kananen K, Markkula M, el-Hefnawy T, Zhang FP, Paukku T, Su JG, HsuehAJ, Huhtaniemi I 1996 The mouse inhibin alpha-subunit promoter directsSV40 T-antigen to Leydig cells in transgenic mice. Mol Cell Endocrinol119:135–146
30. Mann GE, Campbell BK, McNeilly AS, Baird DT 1993 Follicular develop-ment and ovarian hormone secretion following passive immunization of ewesagainst inhibin or estradiol. J Endocrinol 136:225–253
31. McNeilly AS, Fraser HM 1987 Effect of gonadotrophin-releasing hormoneagonist-induced suppression of LH and FSH on follicle growth and corpusluteum function in the ewe. J Endocrinol 115:273–282
32. Parrot JA, Skinner MK 1999 Kit-ligand/stem cell factor induces primordialfollicle development and initiates folliculogenesis Endocrinology 140:4262–4271
33. Elvin JA, Clark AT, Wang P, Wolfman NM, Matzuk MM 1999 Paracrineactions of growth differentiation factor-9 in the mammalian ovary Mol En-docrinol 13:1035–1048
34. McNatty KP, Heath DA, Lundy T, Fidler AE, Quirke L, O’Connell A, SmithP, Groome NP, Tisdall DJ 1999 Control of early ovarian follicular develop-ment. J Reprod Fertil Suppl 54:3–16
35. Kumar TR, Wang Y, Lu N, Matzuk MM 1997 Follicle stimulating hormone isrequired for ovarian follicle maturation but not male fertility. Nat Genet15:201–204
36. Halpin DMG, Charlton HM, Faddy MJ 1986 Effects of gonadotrophin defi-ciency on follicular development in hypogonadal (hpg) mice J Reprod Fertil78:119–125
37. Drummond AE, Baillie AJ, Findlay JK 1999 Ovarian estrogen receptor a andb mRNA expression: impact of development and estrogen. Mol Cell Endo-crinol 149:153–161
38. Saunders PTK, Millar M, Williams K, MacPherson S, Harkiss D, AndersonRA, Orr B, Groome NP, Scobie G, Fraser HM, Differential expression ofestrogen receptor-[alph]a and –b and androgen receptor in the ovaries ofmarmoset and human. Biol Reprod, in press
39. Sar M, Welsch F 1999 Differential expression of estrogen receptor-b andestrogen receptor-a in the rat ovary. Endocrinology 140:963–971
40. Uneo S, Takahashi M, Manganaro TF 1989 Cellular localization of Mullerianinhibiting-substance in the developing rat ovary. Endocrinology 124:1000–1006
41. Woodruff TK, Bescke LM, Groome N, Draper LB, Schwartz NB, Weiss J 1996Inhibin A and inhibin B are inversely correlated to follicle-stimulating hor-mone, yet are discordant during the follicular phase of the rat estrous cycle,and inhibin A is expressed in a sexually dimorphic manner. Endocrinology137:5463–5467
42. Kuroda H, Terada N, Nakayama H, Matsuomoto K, Kitamura Y 1988 Infer-tility due to growth arrest of ovarian follicles in the SI/SI1 mice. Dev Biol126:71–79
43. Juneja SC, Barr KJ, Enders GC, Kidder GM 1999 Defects in the germ line andgonads of mice lacking connexin 43. Biol Reprod 60:1263–1270
44. Lubahn DB, Moyer JS, Golding TS, Couse JF, Korach KS, Smithies O 1993Alteration of reproductive function but not prenatal sexual development afterinsertional disruption of the mouse estrogen receptor gene. Proc Natl Acad SciUSA 90:11162–11166
45. Abel MH, Wooton AN, Wilkins V, Huhtaniemi I, Knight P, Charlton HM2000 The effect of a null mutation in the FSH receptor gene on mouse repro-duction Endocrinology. 141:1795–1803
46. Dong J, Albertini DF, Nishimori K, Kumar TR, Lu N, Matzuk MM 1996Growth differentiation factor-9 is required during early ovarian folliculogen-esis. Nature 383:531–535
4294 OVARIAN STEROIDOGENIC FUNCTION IN DAZL KNOCKOUT MICE Endo • 2000Vol. 141 • No. 11
Copyright © 2022 FDOKUMEN




















