
Locally Produced Survival Cytokines IL‐15 and IL‐7 may be Associated to the Predominance of CD8+...
10
Locally Produced Survival Cytokines IL-15 and IL-7 may be Associated to the Predominance of CD8 + T cells at Heart Lesions of Human Chronic Chagas Disease Cardiomyopathy S. G. Fonseca*, , M. M. Reisà, V. Coelho*, , L. G. Nogueira*, , S. M. Monteiro*, E. C. Mairena*, F. Bacal§, E. Bocchi§, L. Guilherme*, , X. X. Zheng–, F. Y. Liew**, M. L. Higuchià, J. Kalil*, , & E. Cunha-Neto*, , Introduction Chronic Chagas disease Cardiomyopathy (CCC) is the most important clinical consequence of the infection by the protozoan intracellular parasite Trypanosoma cruzi, affecting 16–18 million individuals in Latin America. T. cruzi replicates in the cytoplasm of host cells and establishes a chronic, low-grade, lifelong infection. CCC is an inflamma- tory-dilated cardiomyopathy that may lead to a fatal course, affecting around 10–30% of infected individuals 5–30 years after the primary infection. The remaining 60–70% chronically T. cruzi-infected individuals remain asymptomatic for many years. Histopathologically, CCC is characterized by a diffuse myocarditis with focal aspects; a mononuclear infiltrate, intense heart fibre damage, promin- ent fibrosis and scarcity of T. cruzi parasites [1–3]. Increased local expression of the cytokines IFN-c and TNF-a [4, 5], interleukin (IL)-6 and IL-4 [5], as well as HLA class I and II molecules and adhesion molecules has been reported [6]. Real-time PCR analysis showed that the *Laboratory of Immunology, Heart Institute of the Sao Paulo University Medical School, Sao Paulo, Brazil; Institute for Investigation in Immunology, Millennium Institutes, Sao Paulo, Brazil; àLaboratory of Pathology, Heart Institute of the Sao Paulo University Medical School, Sao Paulo, Brazil; §Heart Failure and Heart Transplantation Unit, Heart Institute of the Sao Paulo University Medical School, Sao Paulo, Brazil; –Division of Immunology, Beth Israel Deaconess Medical Center, Harvard Medical School, Boston, MA, USA; **Division of Immunology, Infection and Inflammation, Glasgow Biomedical Research Centre, University of Glasgow, Glasgow, UK; and Division of Clinical Immunology and Allergy, University of Sao Paulo, School of Medicine Sao Paulo, Brazil Received 13 April 2007; Accepted in revised form 24 May 2007 Correspondence to: Dr E. Cunha-Neto, Laboratorio de Imunologia, Instituto do Coracao, Universidade de Sao Paulo. Av Dr Eneas de Carvalho Aguiar, 44 BL 2, 9º andar, Sao Paulo, SP 05403-000, Brazil. E-mail: [email protected] Abstract Human chronic Chagas disease cardiomyopathy (CCC) is an inflammatory- dilated cardiomyopathy occurring years after infection by the protozoan Trypanosoma cruzi. The heart inflammatory infiltrate in CCC shows a 2:1 predominance of CD8 + in relation to CD4 + T cells, with a typical Th1-type cytokine profile. However, in vitro expansion of infiltrating T cells from heart biopsy-derived fragments with interleukin-2 (IL-2) and phyto- haemagglutinin leads to the outgrowth of CD4 + over CD8 + T cells. We hypothesized that survival cytokines, such as IL-2, IL-7 and IL-15 might be differentially involved in the growth and maintenance of heart-infiltra- ting and peripheral CD8 + T cells from CCC patients. We found that IL-7 and IL-15 were superior to IL-2 in the expansion and viability of CD8 + T cells from both PBMC and heart-infiltrating T-cell lines from CCC patients, and the combination of the three cytokines showed synergic effects. Heart-infiltrating CD8 + T cells showed higher expression of both IL-15Ra and c c chain than CD4 + T cells, which may explain the improve- ment of CD8 + T-cell growth in the presence of IL-2 + IL-7 + IL-15. Immunohistochemical identification of IL-15 and the higher mRNA expres- sion of IL-15Ra, IL-7 and c c chain in CCC heart tissues compared with control individuals indicate in situ production of survival cytokines and their receptors in CCC hearts. Together, our results suggest that local pro- duction of IL-7 and IL-15 may be associated with the maintenance and predominance of CD8 + T cells, the cells effecting tissue damage in CCC hearts. doi: 10.1111/j.1365-3083.2007.01987.x .................................................................................................................................................................. ȑ 2007 The Authors 362 Journal compilation ȑ 2007 Blackwell Publishing Ltd. Scandinavian Journal of Immunology 66, 362–371
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Locally Produced Survival Cytokines IL‐15 and IL‐7 may be Associated to the Predominance of CD8+...

Locally Produced Survival Cytokines IL-15 and IL-7may be Associated to the Predominance of CD8+ Tcells at Heart Lesions of Human Chronic ChagasDisease Cardiomyopathy
S. G. Fonseca*,�, M. M. Reis�, V. Coelho*,�, L. G. Nogueira*,�, S. M. Monteiro*, E. C. Mairena*,F. Bacal§, E. Bocchi§, L. Guilherme*,�, X. X. Zheng–, F. Y. Liew**, M. L. Higuchi�, J. Kalil*,�,��& E. Cunha-Neto*,�,��
Introduction
Chronic Chagas disease Cardiomyopathy (CCC) is the mostimportant clinical consequence of the infection by theprotozoan intracellular parasite Trypanosoma cruzi, affecting16–18 million individuals in Latin America. T. cruzireplicates in the cytoplasm of host cells and establishes achronic, low-grade, lifelong infection. CCC is an inflamma-tory-dilated cardiomyopathy that may lead to a fatalcourse, affecting around 10–30% of infected individuals
5–30 years after the primary infection. The remaining60–70% chronically T. cruzi-infected individuals remainasymptomatic for many years. Histopathologically, CCC ischaracterized by a diffuse myocarditis with focal aspects; amononuclear infiltrate, intense heart fibre damage, promin-ent fibrosis and scarcity of T. cruzi parasites [1–3].Increased local expression of the cytokines IFN-c andTNF-a [4, 5], interleukin (IL)-6 and IL-4 [5], as well asHLA class I and II molecules and adhesion molecules hasbeen reported [6]. Real-time PCR analysis showed that the
*Laboratory of Immunology, Heart Institute of
the Sao Paulo University Medical School, Sao
Paulo, Brazil; �Institute for Investigation in
Immunology, Millennium Institutes, Sao Paulo,
Brazil; �Laboratory of Pathology, Heart
Institute of the Sao Paulo University Medical
School, Sao Paulo, Brazil; §Heart Failure and
Heart Transplantation Unit, Heart Institute of
the Sao Paulo University Medical School, Sao
Paulo, Brazil; –Division of Immunology, Beth
Israel Deaconess Medical Center, Harvard
Medical School, Boston, MA, USA; **Division
of Immunology, Infection and Inflammation,
Glasgow Biomedical Research Centre,
University of Glasgow, Glasgow, UK; and
��Division of Clinical Immunology and Allergy,
University of Sao Paulo, School of Medicine
Sao Paulo, Brazil
Received 13 April 2007; Accepted in revisedform 24 May 2007
Correspondence to: Dr E. Cunha-Neto,
Laboratorio de Imunologia, Instituto do
Coracao, Universidade de Sao Paulo. Av Dr
Eneas de Carvalho Aguiar, 44 BL 2, 9º andar,
Sao Paulo, SP 05403-000, Brazil. E-mail:
Abstract
Human chronic Chagas disease cardiomyopathy (CCC) is an inflammatory-dilated cardiomyopathy occurring years after infection by the protozoanTrypanosoma cruzi. The heart inflammatory infiltrate in CCC shows a 2:1predominance of CD8+ in relation to CD4+ T cells, with a typicalTh1-type cytokine profile. However, in vitro expansion of infiltrating T cellsfrom heart biopsy-derived fragments with interleukin-2 (IL-2) and phyto-haemagglutinin leads to the outgrowth of CD4+ over CD8+ T cells. Wehypothesized that survival cytokines, such as IL-2, IL-7 and IL-15 mightbe differentially involved in the growth and maintenance of heart-infiltra-ting and peripheral CD8+ T cells from CCC patients. We found that IL-7and IL-15 were superior to IL-2 in the expansion and viability of CD8+
T cells from both PBMC and heart-infiltrating T-cell lines from CCCpatients, and the combination of the three cytokines showed synergiceffects. Heart-infiltrating CD8+ T cells showed higher expression of bothIL-15Ra and cc chain than CD4+ T cells, which may explain the improve-ment of CD8+ T-cell growth in the presence of IL-2 + IL-7 + IL-15.Immunohistochemical identification of IL-15 and the higher mRNA expres-sion of IL-15Ra, IL-7 and cc chain in CCC heart tissues compared withcontrol individuals indicate in situ production of survival cytokines andtheir receptors in CCC hearts. Together, our results suggest that local pro-duction of IL-7 and IL-15 may be associated with the maintenance andpredominance of CD8+ T cells, the cells effecting tissue damage in CCChearts.
doi: 10.1111/j.1365-3083.2007.01987.x..................................................................................................................................................................
� 2007 The Authors
362 Journal compilation � 2007 Blackwell Publishing Ltd. Scandinavian Journal of Immunology 66, 362–371

gene expression levels of IFN-c-inducible chemokinesMCP-1, IP-10 and MIG, as well as the chemokine recep-tors CCR2 and CXCR3, were selectively upregulated inCCC heart tissue [7]. The heart infiltrate in CCC shows a2:1 ratio of CD8+ over CD4+ T cells [2, 3, 5] and granzymeA is detected in the heart lesions of CCC patients, suggest-ing a role for cytotoxicity in heart tissue damage [8]. It hasbeen observed that CD8+ T cells accumulate in the scarceareas of heart tissue where T. cruzi is found [9]. Recently,we observed that CD8+ T cells derived from a heart-infil-trating T-cell line from a CCC patient were specific toT. cruzi epitopes [10]. The factors leading to the accumula-tion of CD8+ T cells in CCC heart tissue are still unknown.It can be hypothesized that survival cytokines, such asIL-2, IL-7 and IL-15, play a role in this phenomenon.
Interleukin-2, IL-15 and IL-7, are known to have effectson CD4+ and CD8+ T-cell development, survival and func-tion. IL-15, IL-7 and IL-2 bind to multimeric receptorsthat share the common c chain (cc) subunit (CD132). Eachcytokine, however, interacts with a unique, ligand-specifica-chain receptor. IL-2 and IL-15 receptors also share ab chain (CD122) and distinct associations of cytokinereceptor chains generate receptors with high or intermedi-ate affinity for each cytokine [11]. Even though IL-15 andIL-2 share several properties, it is known that IL-2 and IL-15 can have very distinct roles in T-cell biology [12, 13].IL-2 promotes T-cell activation and proliferation, and sig-nalling through the IL-2 receptor (IL-2R) complex mayalso trigger the death or suppression of activated lympho-cytes [14–16]. On the other hand, signalling through theIL-15R complex is necessary for the maintenance of mem-ory CD8+ T cells and for NK-cell development [17, 18].IL-15 also exists as a membrane-associated cytokine andthe membrane IL-15 ⁄ IL-15Ra can be internalized andrecycled back to cell surface promoting longer IL-15 activ-ity. This process may be involved in the long-term CD8+
memory T-cell survival [19]. It has been shown that themaintenance of CD8+ naıve T cells require T-cell receptor(TCR) signalling through MHC and IL-7, whereas CD8+
memory T cells require IL-15 or IL-7 [20–23].Considering the role of IL-2, IL-7 and IL-15 in the
growth and maintenance of CD8+ T cells, in the presentstudy we investigated the involvement of such cytokinesand their receptors in the accumulation of CD8+ T cellsin CCC heart lesions.
Materials and methods
PBMC and PBMC-derived T-cell lines. Peripheral bloodsamples of Chagas disease patients with CCC were collec-ted with anticoagulant (heparin or EDTA) for this study.Sample collection procedures have been approved by theInternal Review Board. Mononuclear cells from hepari-nized peripheral blood samples were obtained by Ficollgradient centrifugation. Thawed PBMC were resuspended
in DMEM medium with 10% of the human serum.Viability was assessed and generally exceeded 90%. TheT-cell line from a CCC patient (P#12) was obtained fromPBMC stimulated with phytohaemagglutinin (PHA)(5 lg ⁄ ml) and 106 feeder cells (50-Gy irradiated PBMC)in 1 ml of Dulbecco’s modified Eagle’s medium supple-mented with 2 mM of L-glutamine, 1 mM sodium pyru-vate, MEM’s non-essential amino acids and MEM’svitamins (all from GIBCO, Grand Island, NY, USA),50 lg ⁄ ml of gentamicin, 10 mM HEPES buffer, 10% ofnormal human serum (complete medium) plus 40 UI ⁄ mlhuman recombinant interleukin-2 (Proleukin IL-2;Chiron, Emeryville, CA, USA) in a humidified, 5% ofCO2 atmosphere. After two rounds of expansion (10–20 days), lymphoblasts were re-stimulated and culturedin DMEM 10% human serum supplemented with IL-2(100 UI ⁄ ml), IL-7 (5 ng ⁄ ml; PeproTech, Rocky Hill,NJ, USA) and IL-15 (5 ng ⁄ ml; PeproTech) and stimula-ted by PHA (Sigma, St Louis, MO, USA). CD8+ cellswere purified using magnetic beads attached to mono-clonal antibody against human CD8 (M450; Dynal, Oslo,Norway). The beads were removed using Detach-a-Beadantibodies (Dynal). The resulting CD8+ T-cell line had94% of CD8+ cells and 0.3% of CD4+ cells, the CD8),pass-through T-cell line had 90.4% of CD4+ and 1.2%of CD8+ cells and was referred as a CD4+ T-cell line.
Heart-infiltrating T-cell lines. Right ventricle endomyo-cardial biopsy fragments from 11 CCC patients wereminced and cultured in 96-well flat-bottom cultureplates in DMEM (Gibco, Grand Island, NY, USA) with10% of inactivated human serum, supplemented with40 U ⁄ ml IL-2 (Hoffman-La Roche, Nutley, NJ, USA), inthe presence of autologous irradiated (50 Gy) PBMC(105 ⁄ well) as described [4]. Lymphoblasts were laterexpanded by two 15-day rounds of re-stimulation withPHA (2.5 lg ⁄ ml), 50-Gy irradiated PBMC (106 ⁄ ml) incomplete medium plus 40 U ⁄ ml of IL-2. After two tothree rounds of expansion, lymphoblasts were cultured inDMEM, 10% human serum supplemented by IL-2(100 U ⁄ ml), IL-7 (5 ng ⁄ ml) and IL-15 (5 ng ⁄ ml) and sti-mulated by PHA. No Trypanosoma cruzi parasite growthwas observed during endomyocardial biopsy explant cul-tures, as detected by direct visualization of the highlymobile trypomastigote forms.
Immunohistochemistry for the detection of IL-15 in the heartlesion. Fragments of myocardium of 15 CCC patients andof four control individuals, without inflammatory cardiacdisease, were frozen in isopentane and maintained in liquidnitrogen. Sections (5 lm) of frozen myocardial fragmentswere fixed in acetone and incubated with mouse anti-CD68primary antibody (1:200) from Dako-Patts (Glostrup,Denmark) to define macrophages or mouse anti-IL-15(1:100) from R&D. The secondary antibody was biotinyl-ated rabbit anti-mouse IgG (Dako, Ely, UK), diluted to1:400 and incubated during 30 min at 37 �C. The reaction
S. G. Fonseca et al. IL-15 and IL-7 in Chagas’ Cardiomyopathy 363..................................................................................................................................................................
� 2007 The Authors
Journal compilation � 2007 Blackwell Publishing Ltd. Scandinavian Journal of Immunology 66, 362–371

product was detected with avidin–biotin–peroxidase com-plex according to ABC kit protocol (Vector Laboratories,Burlingame, CA, USA), and the reaction was developedwith diamino-benzidine (Sigma). The negative control wasassessed by elimination of primary antibody. The slideswere counterstained with haematoxylin and eosin. CD68+
for macrophages or IL-15+ mononuclear cells were countedin 25 fields, 400· optical magnification. In the standard-ization of the anti-human IL-15 antibody, we utilizedmononuclear cells of peripheral blood adhered in slidesand stimulated with lipopolysaccharide (LPS), 10 lg ⁄ ml,during 48 h.
Flow cytometry assays. The surface phenotype of lympho-blasts was identified after immunostaining with monoclo-nal antibodies directed to the CD3, CD4, CD8, TCRab,TCRcd, CD56, CD16 surface markers (BD Biosciences,San Jose, CA, USA) labelled with fluorochromes and flowcytometric analysis (FACScan; Becton-Dickinson, SanJose, CA, USA). The phycoerythrin-conjugated (PE) anti-bodies against CD25 (IL-2Ra), CD122 (IL-2Rb), CD132(IL-2Rc) were all purchased from BD PharMingen (SanDiego, CA, USA). For IL-15Ra-chain staining, cells wereincubated with an IL-15 mutant ⁄ Fcc2a fusion that bindsto the a-chain [24], followed by staining with biotinylat-ed goat anti-mouse IgG antibody. Cells were then washedand stained with PE-streptoavidin. Twenty thousandevents were collected and analysed on a FACScan flow cy-tometer and CellQuest software (Becton-Dickinson; Bio-sciences). Based on the characteristics of the forward andside-scatter patterns, a gate was set on the lymphocytesand the expression of IL-2R and IL-15R chains was fol-lowed separately on CD3+CD4+ and CD3+CD8+ cells.
Assessment of in vitro growth. T cells from long-termPBMC or heart-infiltrating T-cell lines from CCCpatients (2 · 105 cells ⁄ well) were cultured with 1 · 106
irradiated PBMC plus PHA (2.5 lg ⁄ ml) in the presenceof different cytokines: IL-2 (100 IU ⁄ ml), IL-7 and IL-15(10 ng ⁄ ml) or their combination. Fresh medium supple-mented with cytokines was added every 3 days. To deter-mine the survival and expansion of T cells, every2–3 days from day 6, the numbers of viable cells wereestimated using trypan blue exclusion, and the cells werere-suspended in fresh medium containing the correspond-ing cytokines.
Long-term effects of cytokines on heart-infiltrating CD8+
T-cell growth. T-cell lines were established from endomyo-cardial biopsy of CCC patients and cultured in the presenceof human recombinant IL-2 (100 lg ⁄ ml). After two tothree stimuli with PHA (2.5 lg ⁄ ml), the lines were cul-tured in the presence of IL-2 + IL-7 + IL-15. The pheno-type of T-cell line was monitored after two or more stimuliwith PHA in the presence of IL-2 + IL-7 + IL-15.
Real-time reverse transcriptase-PCR analysis. RNA sam-ples for the real-time reverse transcriptase (RT)-PCRexperiments came from explanted left ventricular free wall
heart samples obtained at transplantation from 11 CCC(serological diagnosis and positive epidemiology) end-stageheart failure patients and three non-failing donor heartsfrom normal adult heart tissue not used for cardiac trans-plantation. The total RNA was extracted using the Trizol�
method (Life Technologies Inc., Grand Island, NY, USA).The integrity of the RNA was confirmed by gel electro-phoresis. For cDNA synthesis, 5 lg of total RNA, 1 lloligodT (500 lg ⁄ ml), 1 ll dNTP (10 mM of dATP,dCTP, dGTP and dTTP), DEPC water q.s.p. 12 ll, wereincubated for 5 min at 40 �C in a 200-ll tube. Later, 4 llTranscription Buffer 5x (Tris-HCl 250 mM pH 8.3, KCl375 mM, MgCl2 15 mM), 2 ll DTT 0.1 M, 1 ll inhibitorRNase (40 U ⁄ ll) (RNAse OUT�; Invitrogen, Carlsbad,CA, USA) and 1 ll Super-script II� Reverse Transcriptase(200 U ⁄ ll) (Invitrogen, Carlsbad, CA, USA) were added;the reaction was heated for 50 min at 42�C and 15 min at70�C. We designed forward and reverse primers for quanti-tative real-time RT-PCR using Primer Express (PEApplied Biosystems, Foster City, CA, USA), for amplifica-tion of the desired gene with the following primersequences: IL-2 (Acc. no. V00564) sense 5¢-GACCCAGG-GACTTAATCAGCAAT-3¢, anti-sense 5¢-CTGTCTCATC-AGCATATTCACACATG-3¢; IL-2Ra (Acc. no. X01057)sense 5¢-GCGTCTGCAAAATGACCCAC-3¢, anti-sense5¢-CTGAGGCTTCTCTTCACCTGGA-3¢; IL-2Rb (Acc.no. M26062) sense 5¢-ATCAACTGCAGGAACACCGG-3¢, anti-sense 5¢-CTGGGAAAAGAACTTCGAGGG-3¢;IL-2Rc (Acc. no. D11086) sense 5¢-TCTGGCTGGAACG-GACGAT-3¢, anti-sense 5¢-ACACCACTCCAGGCCG-AAA-3¢; IL-7 (Acc. no. J04156) sense 5¢-TGAACTGCACTGGCCAGGTTAA-3¢, anti-sense 5¢-CTTCCAAACTCT-TTGTTGGTTGGG-3¢; IL-7Ra (Acc. no. M29696) sense5¢-GATGAAAACAAATGGACGCATGT-3¢, anti-sense5¢-GTGATCAGGGATGGATCGAACTT-3¢; IL-15 (Acc.no. U14407) sense 5¢-GGTTCACCCCAGTTGCAAAG-TAAC-3¢, anti-sense 5¢-TATCATGAATACTTGCATCT-CCGGA-3¢; IL-15Ra (Acc. no. U31628) sense 5¢-TGAGCGCTGTGTCTCTCCTG-3¢, anti-sense 5¢-CCATGGCTTCCATTTCAACG -3¢; glyceraldehyde-3-phosphate dehy-drogenase (GAPDH; Acc. no. M33197) sense 5¢-TGGT-CTCCTCTGACTTCAACA-3¢, anti-sense 5¢-AGCCAAATTCGTTGTCATACC-3¢. Real-time PCR reactions werecarried out in an ABI Prism 7500 Sequence Detection sys-tem (PE Applied Biosystems, Foster City, CA, USA). Real-time PCR was performed using the SYBR Green PCRMaster Mix (PE Applied Biosystems), according to themanufacturer’s instructions, using 5 ll of diluted cDNAas a template. All the samples were tested in triplicate withthe reference gene GAPDH, a housekeeping gene for nor-malization of data. For all genes we constructed standardcurves and determined the slope to calculate the PCR effi-ciency according to Pfaffl [25]. Normalization and foldchange were calculated with the DDCt method withGAPDH as the reference mRNA [26].
364 IL-15 and IL-7 in Chagas’ Cardiomyopathy S. G. Fonseca et al...................................................................................................................................................................
� 2007 The Authors
Journal compilation � 2007 Blackwell Publishing Ltd. Scandinavian Journal of Immunology 66, 362–371

Statistical analysis. The non-parametric Mann–Whitneyrank sum test was used to compare the numbers ofIL-15+ and CD68+ cells in heart tissues from CCCpatients and non-inflammatory heart disease samples.
Results
Phenotypic characterization of derived heart-infiltrating
T-cell lines from CCC patients grown on IL-2
To investigate the expansion of heart-infiltrating T cellsby T-cell trophic cytokines, heart-infiltrating T-cell lineswere established culturing small fragments of heart tissuebiopsies from 10 CCC patients in the presence of IL-2and autologous PBMC-irradiated feeder cells. The average±standard deviation of the percentage of CD3+ T cellsamong these T-cell lines was 94.0 ± 5.1% and an over-whelming majority was TCRab+ (99.0 ± 0.4%). Lessthan 1% of cells were TCRcd+ or CD56+CD16+ (NKcells). A predominance of 6:1 of CD4+ (74.6 ± 11.0%)over CD8+ T cells (11.3 ± 10.4%) was observed. This isin sharp contrast to the 0.5:1 CD4:CD8 T-cell ratiosfound on direct immunohistochemistry analysis of CCCendomyocardial biopsies [2, 3].
Effect of IL-7 and IL-15 on the expansion of CD8+ and
CD4+ T cells from PBMC-derived T-cell line from a CCC
patient
We thus compared the effect of IL-15 and IL-7 withIL-2 on the expansion of CD4+ and CD8+ T cells fromT-cell lines obtained from a CCC patient. For that mat-ter, we used isolated CD8+ and CD4+ T cells, obtained
from a PBMC-derived T-cell line from a CCC patient.The CD8+ and CD4+ T-cell lines were stimulated withPHA and irradiated feeder cells plus IL-2, IL-7, IL-15alone or in combination, and the cell growth was fol-lowed for 31 days. Fresh culture medium with cytokineswas added twice a week. Figure 1 shows that in thepresence of IL-2 alone, the number of viable CD4+ Tcells was stable until 31 days after stimulation withPBMC-irradiated cells plus PHA. In contrast, thegrowth of the CD8+ T-cell line was clearly reduced15 days after stimulus, in comparison to CD4+ T cellsunder IL-2 alone (Fig. 1). Both CD4+ and CD8+ Tcells show increased expansion under IL-7 and IL-15,when compared with IL-2 alone; while the combinationof IL-2, IL-7 and IL-15 increased both expansion andmaintenance of viability over time.
Expression of chains of IL-2R and cc chain on PBMC-derived
CD8+ and CD4+ T lymphocytes
We investigated whether the failure of IL-2 to inducethe growth of long-term PBMC-derived CD8+ T cellsfrom a CCC patient was dependent on the differentialexpression of IL-2R chains. For that matter, we evaluatedthe kinetics of the expression of IL-2Ra (CD25),IL-2 ⁄ IL-15Rb (CD122) and cc chain (CD132) on thePBMC-derived CD8+ and CD4+ T-cell lines culturedwith IL-2, 15 days after the last stimulation with PHAand irradiated feeder cells. We observed that the medianof fluorescence intensities for IL-2Ra and cc chain weresubstantially higher on CD4+ T cells than on CD8+ Tcells (Fig. 2A). This pattern did not change at day 10after PHA stimulation (data not shown). Similarly, we
IL-2
0 5 10 15 20 25 30 35 400
5
10
15
20
25
30CD8+ T cellsCD4+ T cells
Days after stimulus
Cel
ls x
10
6C
ells
x 1
06
IL-7
0 5 10 15 20 25 30 35 400
5
10
15
20
25
30
Days after stimulus
IL-15
0 5 10 15 20 25 30 35 400
5
10
15
20
25
30
Days after stimulus
IL-2+IL-7+IL-15
0 5 10 15 20 25 30 35 400
5
10
15
20
25
30
Days after stimulus
Cel
ls x
10
6C
ells
x 1
06
Figure 1 Growth of CD8+ and CD4+ T cells
from PBMC-derived T cell lines in presence
of cytokines. CD8+ and CD4+ T-cell lines
(2 · 105) isolated from a PBMC-derived
T-cell line from a Chagas disease patient were
cultured with 106 irradiated allogeneic
PBMC as feeder cells and PHA (2.5 lg ⁄ ml)
in presence of IL-2 (100 IU ⁄ ml), IL-7
(5 ng ⁄ ml) and IL-15 (5 ng ⁄ ml) or with a
combination of the three cytokines. Viable
cells were counted in a Neubauer chamber
from day 6 to day 31. Each time-point repre-
sents averages of cell numbers from three dif-
ferent experiments.
S. G. Fonseca et al. IL-15 and IL-7 in Chagas’ Cardiomyopathy 365..................................................................................................................................................................
� 2007 The Authors
Journal compilation � 2007 Blackwell Publishing Ltd. Scandinavian Journal of Immunology 66, 362–371

also observed that the percentages of IL-2Ra- and cc
chain-positive cells were much lower on CD8+ T cellsthan on CD4+ T cells from the PBMC-derived T-cell linefrom the CCC patient (Fig. 2B).
Effect of IL-2, IL-7 and IL-15 on the expansion of
heart-infiltrating T-cell lines from Chagas disease patients
After the observation that IL-15, IL-2 and IL-7 wereimportant for the long-term growth of peripheral CD8+
T-cell lines, we investigated the effect of the combinationof IL-2 + IL-7 + IL-15 on the expansion of heart-infiltra-ting T-cell lines from CCC patients. Figure 3 shows thegrowth curves of distinct heart-infiltrating T-cell linesfrom five CCC patients, after PHA and irradiated feederstimuli. Although there was some interindividual vari-ation of expansion levels, the combination of IL-2, IL-7and IL-15 promoted an increase in the number of viableT cells in comparison to IL-2 alone, visible on day 12after stimulus in all T-cell lines.
Differential expansion of CD8+ T cells from heart-infiltrating
T-cell lines from CCC patients in the presence of IL-2, IL-15
and IL-7
To address whether IL-15 and IL-7 were related with themaintenance of CD8+ T cells in the heart infiltrate of Cha-gas disease patients, we analysed the long-term growth ofheart-infiltrating T-cell lines from Chagas disease patients
in the presence of IL-2, IL-7 and IL-15. These heart-infil-trating T-cell lines were established initially with IL-2and autologous-irradiated feeder cells, and the average pro-portion of CD8+ T cells was 11.34%, ranging from 0.4%to 38%. The effect of long-term culture with IL-2, IL-7and IL-15 on the expansion of CD8+ T cells from heart-infiltrating T-cell lines from CCC patients is shown inFigure 4. Half of the heart-derived T-cell lines showed anincrease in the proportion of CD8+ T cells (Fig. 4; CD8+
T cells after expansion ⁄ CD8+ T cells before expansion ran-ging from 2 to 72).
In situ expression of IL-15 on heart tissue from CCC patients
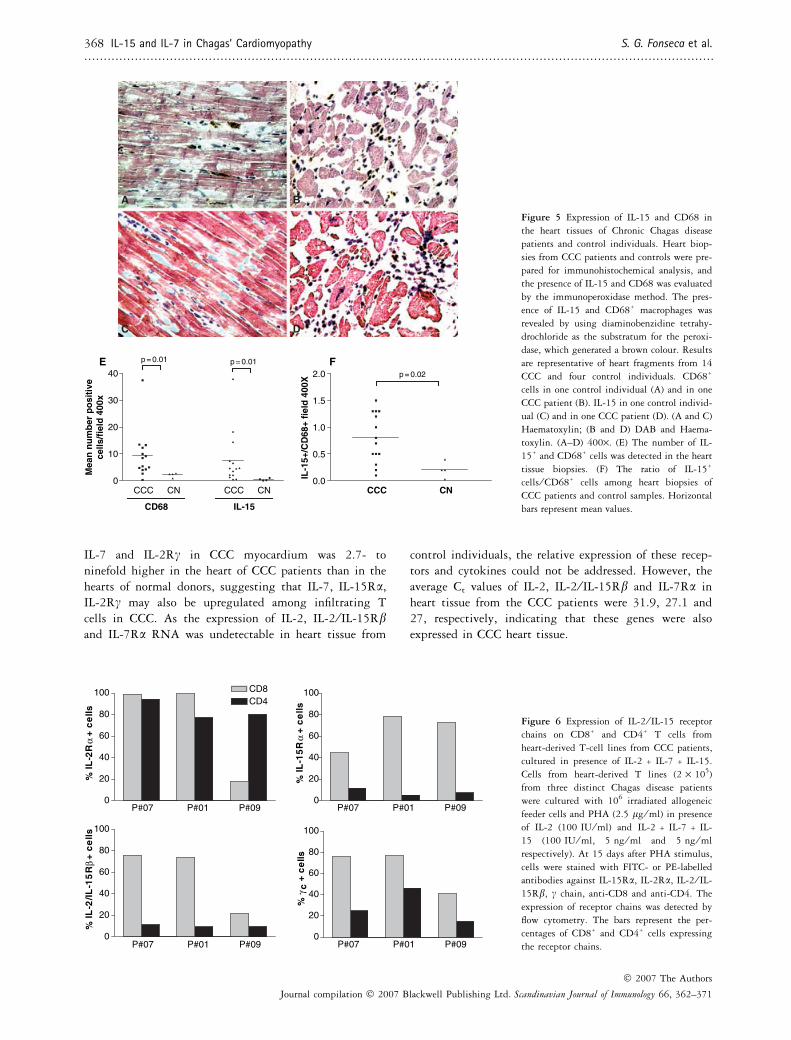
Considering the importance of IL-15 in the growth andmaintenance of CD8+ T cells from heart-infiltrating T-celllines and the predominance of CD8+ T over CD4+ T cellin the heart infiltrate of Chagas disease patients, we nextinvestigated the expression of IL-15 in the endomyocardialbiopsies of 14 CCC patients. As IL-15 can be secreted bymacrophages, the number of CD68+ macrophages was alsoquantified in the same heart tissues. Immunohistochemis-try analysis showed that the cell infiltrates from Chagasdisease patients have a significantly higher numbers of bothCD68+ cells and IL-15+ cells when compared with thehearts from control donors (Fig. 5). The average number ofIL-15+ cells ⁄ 400· field among CCC patients was 7.7 ±10.1, compared with 0.48 ± 0.4 cells among control sam-ples, a 16-fold difference (P = 0.01); the average number
100
500
Cou
nts
FL2-H FL2-H FL2-H
500
Cou
nts
500
Cou
nts
101 102 103 104 100 101 102 103 104 100 101 102 103 104
0
25
50
75
100B
A
0 10 days 0 10 days 0 10 days
% IL
-2R
α+ c
ells
0
25
50
75
% IL
-2/IL
-15R
β+ c
ells
0
25
50
75
100 CD8CD4
%γ c
+ ce
lls
IL-2Rα IL-2Rβ γ c
Figure 2 Expression of IL-2 receptor chains on CD8+ and CD4+ T cells from PBMC-derived T-cell lines from a Chagas disease patient, cultured in
presence of IL-2. Cells from CD8+ and CD4+ T-cell lines (2 · 105) isolated from PBMC-derived T-cell line from a Chagas disease patient were cul-
tured with 106 irradiated allogeneic feeder cells and PHA (2.5 lg ⁄ ml) in presence of IL-2 (100 IU ⁄ ml). (A) The relative expression of IL-2Ra,
IL-2 ⁄ IL-15Rb and cc on CD8 (shaded histogram) and on CD4 (solid line) T-cell lines were measured on day 0 by double stained with FITC or
PE-labelled antibodies to IL-2R chains and CD8 and CD4 detected by flow cytometry. (B) The percentage of positive cells to IL-2Ra, IL-2 ⁄ IL-15Rband cc in CD8+ and CD4+ T-cell lines were measured on day 0 and 10 after PHA stimulus and are represented by columns in the graph.
366 IL-15 and IL-7 in Chagas’ Cardiomyopathy S. G. Fonseca et al...................................................................................................................................................................
� 2007 The Authors
Journal compilation � 2007 Blackwell Publishing Ltd. Scandinavian Journal of Immunology 66, 362–371

of CD68+ cells ⁄ 400· field among CCC patients was9.3 ± 9.1, compared with 2.2 ± 0.8 among control sam-ples, a fourfold difference (P = 0.01) (Fig. 5E). The ratio ofIL-15+ ⁄ CD68+ cells among heart samples of Chagas dis-ease patients was fourfold higher compared with controlsamples (Fig. 5F; P = 0.02), suggesting an increasedactivation state.
Expression of IL-2, IL-7, IL-15 and their receptors on
intralesional T-cell lines and in the heart tissue from
CCC patients
Considering that heart-infiltrating T-cell lines expandedafter long-term culture with a combination of IL-2 + IL-7 + IL-15, we investigated the expression of IL-2 andIL-15 receptor chains in heart-derived T-cell lines. Weevaluated the surface expression of IL-2Ra, IL-15Ra andIL-2 ⁄ IL-15Rb and cc chain in CD8+ and CD4+ T cellsfrom heart-infiltrating T-cell lines from three CCCpatients, cultured with a combination of IL-2, IL-7 and IL-15. At day 15 post-stimulus, the proportion of IL-15Ra+
and IL-2 ⁄ IL-15Rb+ cells was higher on the CD8+ T cellsin all three heart-infiltrating T-cell lines than on CD4+ Tcells (Fig. 6). However, no difference was detected in theexpression of IL-2Ra and cc chain on the CD8+ and CD4+
T cells in the same heart-infiltrating T-cell lines culturedin the same conditions (Fig. 6). In addition, the median offluorescence intensity of IL-15Ra was higher on CD8+ Tcells (9.5, 62.3 and 155.3 respectively) than on CD4+ Tcells (3.4, 3.2 and 5.42 respectively).
To assess whether IL-2, IL-7 and IL-15 and theirreceptors are also expressed in CCC heart tissue, we usedreal-time PCR in heart tissue samples from 11 CCCpatients in comparison to heart tissues from three non-inflammatory heart diseases. Figure 7 shows that theexpression average of mRNA levels of genes IL-15Ra,
P#07
0 3 6 9 12 15 18 21 240
10
20
30
4075
125 IL-2IL2+7+15
Days after stimulus
Cel
l nu
mb
er x
105
Cel
l nu
mb
er x
105
Cel
l nu
mb
er x
105
Cel
l nu
mb
er x
105
Cel
l nu
mb
er x
105
P#04
0 3 6 9 12 15 18 21 240
10
20
30
4075
125
Days after stimulus
P#09
0 3 6 9 12 15 18 21 240
10
20
30
4075
125
Days after stimulus
P#05
0 3 6 9 12 15 18 21 240
10
20
30
4075
125
Days after stimulus
P#01
0 3 6 9 12 15 18 21 240
10
20
30
40
75125
Days after stimulus
Figure 3 Effect of IL-2 + IL-7 + IL-15 in
the growth of heart-infiltrating T-cell lines.
After three to five stimuli with 106 irradiated
autologous PBMC plus PHA (2.5 lg ⁄ ml)
and IL-2 (100 IU ⁄ ml), the heart-infiltrating
T-cell lines from five distinct CCC patients
were washed and split into two different cul-
ture conditions, one with IL-2 (100 IU ⁄ ml)
and the other with IL-2 + IL-7 + IL-15
(100 IU ⁄ ml, 5 ng ⁄ ml and 5 ng ⁄ ml respect-
ively) in presence of PHA and feeder cells,
counted every 3 days.
0
10
20
30
40
P#01
P#02
P#03
P#04
P#05
P#06
80
90
100
IL-2 IL-2+7+15
2-3 stimuli 2-5 stimuli
P#07
P#08
P#09
P#10
% C
D8+
Figure 4 Chronic exposure to IL-2 + IL-7 + IL-15 rescues the growth
of CD8+ T cells from heart-infiltrating T-cell lines. Heart-derived T-cell
lines were initially obtained from heart biopsies from 10 CCC patients
were initially stimulated with 106 irradiated autologous PBMC plus
PHA (2.5 lg ⁄ ml) and IL-2 (100 IU ⁄ ml). After three to five stimuli,
the T-cell lines were re-stimulated with IL-2 + IL-7 + IL-15
(100 IU ⁄ ml, 5 ng ⁄ ml and 5 ng ⁄ ml respectively) in presence of PHA
and feeder cells and the percentile of CD8+ T cells was measured by
flow cytometry.
S. G. Fonseca et al. IL-15 and IL-7 in Chagas’ Cardiomyopathy 367..................................................................................................................................................................
� 2007 The Authors
Journal compilation � 2007 Blackwell Publishing Ltd. Scandinavian Journal of Immunology 66, 362–371
IL-7 and IL-2Rc in CCC myocardium was 2.7- toninefold higher in the heart of CCC patients than in thehearts of normal donors, suggesting that IL-7, IL-15Ra,IL-2Rc may also be upregulated among infiltrating Tcells in CCC. As the expression of IL-2, IL-2 ⁄ IL-15Rband IL-7Ra RNA was undetectable in heart tissue from
control individuals, the relative expression of these recep-tors and cytokines could not be addressed. However, theaverage Ct values of IL-2, IL-2 ⁄ IL-15Rb and IL-7Ra inheart tissue from the CCC patients were 31.9, 27.1 and27, respectively, indicating that these genes were alsoexpressed in CCC heart tissue.
0
10
20
30
40
CD68
p = 0.01 p = 0.01
Mea
n n
um
ber
po
siti
vece
lls/f
ield
400
x
0.0
0.5
1.0
1.5
2.0
CCC CN
p = 0.02
IL-1
5+/C
D68
+ fi
eld
400
X
E
C D
A B
F
CNCCCCNCCC
IL-15
Figure 5 Expression of IL-15 and CD68 in
the heart tissues of Chronic Chagas disease
patients and control individuals. Heart biop-
sies from CCC patients and controls were pre-
pared for immunohistochemical analysis, and
the presence of IL-15 and CD68 was evaluated
by the immunoperoxidase method. The pres-
ence of IL-15 and CD68+ macrophages was
revealed by using diaminobenzidine tetrahy-
drochloride as the substratum for the peroxi-
dase, which generated a brown colour. Results
are representative of heart fragments from 14
CCC and four control individuals. CD68+
cells in one control individual (A) and in one
CCC patient (B). IL-15 in one control individ-
ual (C) and in one CCC patient (D). (A and C)
Haematoxylin; (B and D) DAB and Haema-
toxylin. (A–D) 400·. (E) The number of IL-
15+ and CD68+ cells was detected in the heart
tissue biopsies. (F) The ratio of IL-15+
cells ⁄ CD68+ cells among heart biopsies of
CCC patients and control samples. Horizontal
bars represent mean values.
0
20
40
60
80
100 CD8CD4
%R
2-LI
αsll
ec
+
P#07 P#01 P#09
P#07 P#01 P#09
P#07 P#01 P#09
P#07 P#01 P#09
0
20
40
60
80
100
%R
51-
LIα
sl le
c +
0
20
40
60
80
100
%R
51-
LI/2-
LIβ
slle
c +
0
20
40
60
80
100
%γ c
slle
c +
Figure 6 Expression of IL-2 ⁄ IL-15 receptor
chains on CD8+ and CD4+ T cells from
heart-derived T-cell lines from CCC patients,
cultured in presence of IL-2 + IL-7 + IL-15.
Cells from heart-derived T lines (2 · 105)
from three distinct Chagas disease patients
were cultured with 106 irradiated allogeneic
feeder cells and PHA (2.5 lg ⁄ ml) in presence
of IL-2 (100 IU ⁄ ml) and IL-2 + IL-7 + IL-
15 (100 IU ⁄ ml, 5 ng ⁄ ml and 5 ng ⁄ ml
respectively). At 15 days after PHA stimulus,
cells were stained with FITC- or PE-labelled
antibodies against IL-15Ra, IL-2Ra, IL-2 ⁄ IL-
15Rb, c chain, anti-CD8 and anti-CD4. The
expression of receptor chains was detected by
flow cytometry. The bars represent the per-
centages of CD8+ and CD4+ cells expressing
the receptor chains.
368 IL-15 and IL-7 in Chagas’ Cardiomyopathy S. G. Fonseca et al...................................................................................................................................................................
� 2007 The Authors
Journal compilation � 2007 Blackwell Publishing Ltd. Scandinavian Journal of Immunology 66, 362–371

Discussion
In the present work, we showed that the combination ofIL-15, IL-7 and IL-2 promoted the in vitro expansion ofCD8+ T cells from both PBMC-derived and heart-infil-trating T-cell lines from CCC patients. Such expansionmight be dependent on the selective expression ofIL-15Ra, and cc chain on CD8+ T cells from heart-infil-trating T-cell lines. Accordingly, higher heart tissueexpression of IL-15Ra, IL-15, and IL-7 was observed inCCC patients. Our results are consistent with the ideathat the accumulation of CD8+ T cells in the heart infil-trate of CCC patients may be dependent on the local pro-duction of IL-15 and IL-7.
The finding that IL-7 and ⁄ or IL-15 can rescue theproliferation and maintenance of the viability of IL-2-cul-tured CD8+ T cells from a PBMC-derived T-cell linefrom a CCC patient suggests that they play a major rolein the in vitro growth of CD8+ T cells (Fig. 1). In accord-ance with our results, it has been shown by others thatIL-7 [27, 28] and IL-15 [29] preferentially expand theCD8+ T-cell population. The mechanisms by whichIL-15 may contribute to the maintenance of memoryT cells are not fully understood. It has been shown thatthe cell surface IL-15Ra ⁄ IL-15 complex can undergoendosomal internalization, survive lysosomal degradationand recycle to the cell surface [19], contributing to thelong-term survival of CD8+ memory T cells. In our sys-tem, while a proliferative effect of IL-7, IL-15 or theircombination with IL-2 was obvious in the first 2 weeks,until the peak of growth, we cannot ascertain whetherincreased proliferation, inhibition of apoptosis or bothwere operative during the last 2 weeks of the culture,where the cell numbers were stable or decreased. The find-ing that CD8+ T cells from a PBMC-derived T-cell line
from a CCC patient express less IL-2Ra and cc thanCD4+ T cells (Fig. 2) could partially explain the relativerefractoriness of CD8+ T cells to IL-2 (Fig. 1), in linewith the published findings [30–32]. Moreover, the find-ing that CD4+ T cells from the PBMC-derived T-cell linealso increased in numbers after incubation with IL-7,IL-15 or IL-2 + IL-7 + IL-15 in comparison to IL-2 alone(Fig. 2) is also in line with the published literature [33].
The combination of IL-2 + IL-7 + IL-15 also preferen-tially expanded CD8+ T lymphocytes from intralesionalT-cell lines (Fig. 4). In addition, we have also shown thatCD8+ T lymphocytes from the three distinct intralesionalT-cell lines present higher expression of the IL-15Ra andIL-2 ⁄ IL-15Rb, when compared with CD4+ T lympho-cytes (Fig. 5) as observed by others [19, 30]. This is con-sistent with the selective in vitro growth of the heart-infiltrating CD8+ T lymphocytes in the presence of IL-2 +IL-7 + IL-15 by a receptor-driven mechanism. The mech-anism of increased responsiveness to IL-15 observedamong heart-infiltrating CD8+ T cells (increased IL-15Raand cc) seems thus to be distinct from that ofPBMC-derived CD8+ T cells, which showed reducedIL-2Ra and cc expression instead (Fig. 2). We cannotexclude that the refractoriness to IL-2 observed amongheart-infiltrating T cells could be due to distinct post-transductional signalling of IL-2.
The finding of increased mRNA expression ofIL-15Ra and cc in the CD8+ T-cell rich CCC myocardialtissues (Fig. 7) is in line with the surface expression dataon heart-infiltrating T-cell lines (Fig. 6), suggesting thatin situ T cells effectively express such receptors in vivo.The observation of increased levels of IL-7 mRNA(Fig. 7) and an increased number of IL-15+ cells, in thehearts from CCC patients (Fig. 5), indicates that bothIL-7 and IL-15, ligands for the above-mentioned recep-tors, are indeed expressed in the CCC heart tissue. Thefinding that IL-15 mRNA was expressed at similar levelsin control and the CCC heart tissue was in conflict withthe increased expression of immunologically detectableIL-15 protein in the CCC heart tissue. Nevertheless, ithas been reported that IL-15 mRNA is constitutivelyexpressed in many tissues, and its regulation is mainlypost-transcriptional [19, 34]. Together, our results areconsistent with locally produced IL-7 and IL-15 beingmajor growth factors for CD8+ T lymphocytes in CCCheart tissue.
Interleukin-15 has also been identified as an import-ant mediator in several chronic inflammatory diseases[34–40]. IL-15 can synergize with the IL-12 for IFN-cproduction [41], and IL-15 synergizes with IL-7 toenhance the ability of antigen-specific CD8+ and CD4+
T cells to produce IFN-c [42]. Moreover, IFN-c itselfinduces the expression of IL-15 [34]. The finding thatthe ratio of IL-15+ ⁄ CD68+ cells is increased in CCCheart tissue suggests that more CD68+ macrophages are
0.1
1
10
100F
old
ch
ang
e (v
s. n
orm
al h
eart
)
IL-15 IL-15Rα IL-7 IL-2Rα IL-2Rγ
Figure 7 Real-time RT-PCR analysis of expression of mRNA levels of
genes IL-15, IL-15Ra, IL-7, IL-2Ra and IL-2Rc in CCC myocardium.
Normalization and fold change were calculated with the DDCt method,
the data are presented as fold change in gene expression normalized to
the housekeeping gene GAPDH and relative to the three normal hearts.
Data represent mean of triplicate samples obtained from 11 CCC myo-
cardium samples. The horizontal bar stands for the median. Genes
whose median fold-change was above 2 or below 0.5 of control tissues
were considered differentially expressed.
S. G. Fonseca et al. IL-15 and IL-7 in Chagas’ Cardiomyopathy 369..................................................................................................................................................................
� 2007 The Authors
Journal compilation � 2007 Blackwell Publishing Ltd. Scandinavian Journal of Immunology 66, 362–371

induced to produce IL-15, possibly due to the inflamma-tory milieu. It is thus likely that the locally expressedIL-15, possibly produced by CD68+ macrophages, maycontribute to the increased production of proinflammato-ry cytokines, such as IFN-c and TNF-a, observed inCCC heart tissue [4, 5].
Taken together, our results suggest that the accumula-tion of CD8+ T cells in the heart tissue of CCC patientsmay be due, at least in part, to the local expression ofsurvival cytokines, possibly induced by IFN-c. IL-7 andIL-15 which may act on CD8+ cells bearing specific cyto-kine receptors, allowing their selective growth, survivaland maintenance. Their ongoing production of IFN-ccould in turn upregulate local production of IL-15, in apositive feedback loop. Furthermore, this mechanismwould lead to the maintenance of Th1-type inflammatorycells in heart tissue. Whether such granzyme+CD8+ Tcells have a role in heart tissue damage in CCC, or aremerely a collection of exhausted T cells remains to beestablished.
Acknowledgments
We thank Ms Priscila Teixeira for helping in heart tissuecollection. This work was supported by grants from FAP-ESP nº 97 ⁄ 13431-5 (to S.G.F.), CNPq nº 42.0006 ⁄ 99-0and nº 520533 ⁄ 97-6 (to E.CN).
References
1 Higuchi ML, De MC, Pereira BA et al. The role of active myocarditis
in the development of heart failure in chronic Chagas’ disease: a study
based on endomyocardial biopsies. Clin Cardiol 1987;10:665–70.
2 Higuchi ML, De Brito T, Reis MM et al. Correlation between Try-
panosoma cruzi parasitism and myocardial inflammatory infiltrate in
human chronic chagasic myocarditis: light microscopy and immu-
nohistochemical finding. Cardiovasc Pathol 1993;2:101–6.
3 Higuchi ML, Gutierrez PS, Aiello VD, Palomino S. Immunohisto-
chemical characterisation of infiltrating cells in human chronic myo-
carditis: comparison with myocardial rejection process. Virchows
Arch Pathol Anat 1993;4233:157–60.
4 Abel LC, Rizzo LV, Ianni B et al. Chronic Chagas’ disease cardi-
omyopathy patients display an increased IFN-c response to Trypano-soma cruzi infection. J Autoimmun 2001;17:99–107.
5 Reis MM, Higuchi ML, Benvenuti LA et al. An in situ quantitative
immunohistochemical study of cytokines and IL-2R+ in chronic
human chagasic myocarditis: correlation with the presence of myo-
cardial Trypanosoma cruzi antigens. Clin Immunol Immunopathol
1997;83:165–72.
6 Reis DD, Jones EM, Tostes S et al. Expression of major histocom-
patibility complex antigens and adhesion molecules in hearts of
patients with chronic Chagas’ disease. Am J Trop Med Hyg
1993;49:192–200.
7 Cunha-Neto E, Dzau VJ, Allen PD et al. Cardiac gene expression
profiling provides evidence for cytokinopathy as a molecular mech-
anism in Chagas’ disease cardiomyopathy. Am J Pathol
2005;167:305–13.
8 Reis DD, Jones EM, Tostes S Jr et al. Characterization of inflamma-
tory infiltrates in chronic chagasic myocardial lesions: presence of
tumor necrosis factor-alpha+ cells and dominance of granzyme A+,
CD8+ lymphocytes. Am J Trop Med Hyg 1993;48:637–44.
9 Higuchi ML, Reis MM, Aiello VD et al. Association of an increase
in CD8+ T cells with the presence of Trypanosoma cruzi antigens in
chronic, human, chagasic myocarditis. Am J Trop Med Hyg
1997;56:485–9.
10 Fonseca SG, Moins-Teisserenc H, Clave E et al. Identification of
multiple HLA-A*0201-restricted cruzipain and FL-160 CD8+ epi-
topes recognized by T cells from chronically Trypanosoma cruzi-infec-
ted patients. Microbes Infect 2005;7:688–97.
11 Grabstein KH, Eisenman J, Shanebeck K et al. Cloning of a T cell
growth factor that interacts with the beta chain of the interleukin-2
receptor. Science 1994;264:965–8.
12 Waldmann TA, Dubois S, Tagaya Y. Contrasting roles of IL-2 and
IL-15 in the life and death of lymphocytes: implications for immu-
notherapy. Immunity 2001;14:105–10.
13 Waldmann TA. The biology of interleukin-2 and interleukin-15:
implications for cancer therapy and vaccine design. Nat Rev Immunol2006;6:595–601.
14 Refaeli Y, Van Parijs L, London CA, Tschopp J, Abbas AK. Bio-
chemical mechanisms of IL-2-regulated Fas-mediated T cell apopto-
sis. Immunity 1998;8:615–23.
15 Ku CC, Murakami M, Sakamoto A, Kappler J, Marrack P. Control
of homeostasis of CD8+ memory T cells by opposing cytokines.
Science 2000;288:675–8.
16 Shevach EM. Regulatory T cells in autoimmmunity. Annu Rev
Immunol 2000;18:423–49.
17 Tan JT, Ernst B, Kieper WC, LeRoy E, Sprent J, Surh CD. Inter-
leukin (IL)-15 and IL-7 jointly regulate homeostatic proliferation of
memory phenotype CD8+ cells but are not required for memory
phenotype CD4+ cells. J Exp Med 2002;195:1523–32.
18 Waldmann TA, Tagaya Y. The multifaceted regulation of interleu-
kin-15 expression and the role of this cytokine in NK cell differen-
tiation and host response to intracellular pathogens. Annu Rev
Immunol 1999;17:19–49.
19 Sato N, Patel HJ, Waldmann TA, Tagaya Y. The IL-15 ⁄ IL-15Ral-
pha on cell surfaces enables sustained IL-15 activity and contributes
to the long survival of CD8 memory T cells. Proc Natl Acad Sci
USA 2007;104:588–93.
20 Tanchot C, Lemonnier FA, Perarnau B, Freitas AA, Rocha B. Dif-
ferential requirements for survival and proliferation of CD8 naive or
memory T cells. Science 1997;276:2057–62.
21 Tan JT, Tutl E, Leroy E et al. IL-7 is critical for homeostatic prolif-
eration and survival of naıve T cells. Proc Natl Acad Sci U S A
2001;98:8732–7.
22 Kirberg J, Berns A, von Boehmer H. Peripheral T cell survival
requires continual ligation of the T cell receptor to major histocom-
patibility complex-encoded molecules. J Exp Med 1997;186:1269–
75.
23 Schluns KS, Kieper WC, Jameson SC, Lefrancois L. Interleukin-7
mediates the homeostasis of naive and memory CD8 T cells in vivo.
Nat Immunol 2000;1:426–32.
24 Kim YS, Maslinski W, Zheng XX et al. Targeting the IL-15 recep-
tor with an antagonist IL-15 mutant ⁄ Fc gamma2a protein blocks
delayed-type hypersensitivity. J Immunol 1998;160:5742–8.
25 Pfaffl MW. A new mathematical model for relative quantification in
real-time RT-PCR. Nucleic Acids Res 2001;29:e45.
26 Livak KJ, Schmittgen TD. Analysis of relative gene expression data
using real-time quantitative PCR and the 2(-Delta Delta C(T))
Method. Methods 2001;25:402–8.
27 Komschlies KL, Gregorio TA, Gruys ME, Back TC, Faltynek CR,
Wiltrout RH. Administration of recombinant human IL-7 to mice
alters the composition of B-lineage cells and T cell subsets, enhan-
ces T cell function, and induces regression of established metastases.
J Immunol 1994;152:5776–84.
370 IL-15 and IL-7 in Chagas’ Cardiomyopathy S. G. Fonseca et al...................................................................................................................................................................
� 2007 The Authors
Journal compilation � 2007 Blackwell Publishing Ltd. Scandinavian Journal of Immunology 66, 362–371

28 Geiselhart LA, Humphries CA, Gregorio TA, Mou S, Subleski J,
Komschlies KL. IL-7 administration alters the CD4:CD8 ratio,
increases T cell absence of activation. J Immunol 2001;166:3019–27.
29 Weng NP, Liu K, Catalfamo M, Li Y, Henkart PA. IL-15 is a
growth factor and an activator of CD8 memory T cells. Ann N Y
Acad Sci 2002;975:46–56.
30 Zhang X, Sun S, Hwang I, Tough DF, Sprent J. Potent and select-
ive stimulation of memory-phenotype CD8+ T cells in vivo by IL-
15. Immunity 1998;8:591–9.
31 Lodolce JP, Boone Dl, Chai S et al. IL-15 receptor maintains lym-
phoid homeostasis by supporting lymphocyte homing and prolifer-
ation. Immunity 1998;9:669–76.
32 Kanegane H, Tosato G. Activation of naive and memory T cells by
interleukin-15. Blood 1996;88:230–5.
33 Geginat J, Sallusto F, Lanzavecchia A. Cytokine-driven proliferation
and differentiation of human naive, central memory, and effector
memory CD4(+) T cells. J Exp Med 2001;194:1711–9.
34 Liu Z, Geboes K, Colpaert S, DHaens GR, Rutgeerts P, Ceuppens
JL. IL-15 is highly expressed in inflammatory bowel disease and
regulates local T cell-dependent cytokine production. J Immunol
2000;164:3608–15.
35 Khan IA, Kasper LH. IL-15 augments CD8+ T cell-mediated
immunity against Toxoplasma gondii infection in mice. J Immunol
1996;157:2103–8.
36 McInnes IB, Al-Mughales J, Field M et al. The role of interleukin-
15 in T-cell migration and activation in rheumatoid arthritis. Nat
Med 1996;2:175–82.
37 Fehninger TA, Caligiuri MA. Interelukin 15: biology and relevance
to human disease. Blood 2001;97:14–32.
38 Jullien D, Sieling PA, Uyemura K, Mar ND, Rea TH, Modlin RL.
IL-15, an immunomodulator of T cell responses in intracellular
infection. J Immunol 1997;158:800–6.
39 Agostini C, Trentin L, Facco M et al. Role of IL-15, IL-2, and their
receptors in the development of T cell alveolitis in pulmonary sarco-
idosis. J Immunol 1996;157:910–18.
40 Wuttge DM, Eriksson P, Sirsjo A, Hanson GK, Stemme S. Expres-
sion of interleukin-15 in mouse and human atherosclerotic lesions.
Am J Pathol 2001;159:417–23.
41 Avice MN, Demeure CE, Delespesse G, Rubio M, Armant M,
Sarfati M. IL-15 promotes IL-12 production by human monocytes
via T cell-dependent contact and may contribute to IL-12-mediated
IFN-gamma secretion by CD4+ T cells in the absence of TCR liga-
tion. J Immunol 1998;161:3408–15.
42 Gu XX, Yue FY, Kovacs CM, Ostrowski MA. The role of cytokines
which signal through the common gamma chain cytokine receptor
in the reversal of HIV specific CD4 and CD8 T cell anergy. PLoS
ONE 2007;2:e300.
S. G. Fonseca et al. IL-15 and IL-7 in Chagas’ Cardiomyopathy 371..................................................................................................................................................................
� 2007 The Authors
Journal compilation � 2007 Blackwell Publishing Ltd. Scandinavian Journal of Immunology 66, 362–371




















