
Lipopolysaccharide-independent radioimmunoprecipitation and identification of structural and in vivo...
8
Lipopolysaccharide-independent radioimmunoprecipitation and identification of structural and in vivo induced immunogenic surface proteins of Salmonella typhi in typhoid fever Lieselotte Aron*, Gustavo Faundez*, Carlos Gonzalez*, Emilio Roessler* and Felipe Cabello *++ The humoral response to Salmonella typhi is important for protective immunity against typhoidfever, as indicated by the protection obtained with killed cell vaccines and component vaccines (outer membrane proteins, Vi antigen) in animals and human beings. Nonetheless, analysis and interpretation of host humoral immune response to S. typhi surface antigens have been difficult because of the complex structure of the S. typhi envelope and the lack of purified reagents for detection of immune response to individual surface components. Normal and convalescent human sera from typhoid fever patients were absorbed with S. typhi lipopolysaccharide. These sera were used in radioimmunoprecipitation assays of whole S. typhi cells and S. typhi membranes labelled with either 12si or 35S-methionine. This strategy has permitted the unequivocal identification of a humoral immune response to structural and in vivo induced outer membrane proteins of S. typhi. In this manner, we have identified the porins, lipoprotein, the iron-starvation-induced proteins, and three proteins of 30, 18.5 and 15 kDa as surface-exposed immunogens of S. typhi in patients with typhoid fever. These studies suggest that further experimental work is needed to characterize the relevance of both anti-S, typhi outer membrane protein and anti- lipopolysaccharide antibodies in recovery from S. typhi infections andprotective immunity. Keywords: Salmonella typhi; outer membrane proteins; typhoid fever; radioimmunoprecipitation; lipopolysaccharide INTRODUCTION Outer membrane proteins (OMPs), encoded by both chromosomal and plasmid genes, have increasingly been the focus of attention because of their potential role in bacterial pathogenicity of Gram-negative bacteria, especi- ally in Salmonella infections ~ v. These Salmonella OMPs may also be differentially induced by the environment of extracellular and intracellular niches in the host 5'8. The immunogenicity of S. typhi porins and iron-deprivation- induced proteins of S. typhi in typhoid fever 3'8'9 suggests the possibility of their use as immunizing or diagnostic agents. Active and passive immunization by OMPs and antibodies directed against them has been achieved with Salmonella typhimurium, Salmonella typhi, Pseudomonas *Department of Microbiology and Immunology, New York Medical College, Valhalla, NY 10595, USA. tDepartment of Medicine, School of Medicine, University of Chile, Santiago, Chile. tTo whom correspondence should be addressed. (Received 10 February 1992; revised 7 April 1992; accepted 7 April 1992) 0264-410X/93/010010-08 © 1993 Butterworth-Heinemann Ltd 10 Vaccine, Vol. 11, Issue 1, 1993 aeruginosa and Haemophilus influenzae infections in animals10-14. Unfortunately, the interpretation of these results has been obscured by the finding that protection induced by porin preparations is due to antibodies against contaminating LPS 1°, by the use of denatured OMP preparations contaminated with LPS as antigens in immunoblots 3'8 and sera containing anti-LPS anti- bodies 3's'1°, and by the presence in normal sera of antibodies crossreacting with antigens of Gram-negative pathogens 8. Moreover, in vitro experiments have sug- gested that monoclonal antibodies against OMPs do not reach their targets in Gram-negative bacteria because of steric hindrance produced by LPS and capsular poly- saccharides; this rules out a role for them in protective immunity against infection through the stimulation of phagocytosis and complement activation 15.16. Immuno- protection against typhoid fever has been achieved with killed parenteral vaccines and live oral vaccines with different degrees of success 17, suggesting that both humoral and cellular host immune responses are involved in the ability of humans to deal with S. typhi ~7. The relevance of the humoral response in immunoprevention of typhoid is highlighted by the high effectiveness of
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Lipopolysaccharide-independent radioimmunoprecipitation and identification of structural and in vivo...

Lipopolysaccharide-independent radioimmunoprecipitation and identification of structural and in vivo induced immunogenic surface proteins of Salmonella typhi in typhoid fever
Lieselotte Aron*, Gustavo Faundez*, Carlos Gonzalez*, Emilio Roessler* and Felipe Cabello *++
The humoral response to Salmonella typhi is important for protective immunity against typhoid fever, as indicated by the protection obtained with killed cell vaccines and component vaccines (outer membrane proteins, Vi antigen) in animals and human beings. Nonetheless, analysis and interpretation of host humoral immune response to S. typhi surface antigens have been difficult because of the complex structure of the S. typhi envelope and the lack of purified reagents for detection of immune response to individual surface components. Normal and convalescent human sera from typhoid fever patients were absorbed with S. typhi lipopolysaccharide. These sera were used in radioimmunoprecipitation assays of whole S. typhi cells and S. typhi membranes labelled with either 12si or 35S-methionine. This strategy has permitted the unequivocal identification of a humoral immune response to structural and in vivo induced outer membrane proteins of S. typhi. In this manner, we have identified the porins, lipoprotein, the iron-starvation-induced proteins, and three proteins of 30, 18.5 and 15 kDa as surface-exposed immunogens of S. typhi in patients with typhoid fever. These studies suggest that further experimental work is needed to characterize the relevance of both anti-S, typhi outer membrane protein and anti- lipopolysaccharide antibodies in recovery from S. typhi infections andprotective immunity.
Keywords: Salmonella typhi; outer membrane proteins; typhoid fever; radioimmunoprecipitation; lipopolysaccharide
I N T R O D U C T I O N
Outer membrane proteins (OMPs), encoded by both chromosomal and plasmid genes, have increasingly been the focus of attention because of their potential role in bacterial pathogenicity of Gram-negative bacteria, especi- ally in Salmonella infections ~ v. These Salmonella OMPs may also be differentially induced by the environment of extracellular and intracellular niches in the host 5'8. The immunogenicity of S. typhi porins and iron-deprivation- induced proteins of S. typhi in typhoid fever 3'8'9 suggests the possibility of their use as immunizing or diagnostic agents. Active and passive immunization by OMPs and antibodies directed against them has been achieved with Salmonella typhimurium, Salmonella typhi, Pseudomonas
*Department of Microbiology and Immunology, New York Medical College, Valhalla, NY 10595, USA. tDepartment of Medicine, School of Medicine, University of Chile, Santiago, Chile. tTo whom correspondence should be addressed. (Received 10 February 1992; revised 7 April 1992; accepted 7 April 1992)
0264-410X/93/010010-08 © 1993 Butterworth-Heinemann Ltd
10 Vaccine, Vol. 11, Issue 1, 1993
aeruginosa and Haemophilus influenzae infections in animals10-14. Unfortunately, the interpretation of these results has been obscured by the finding that protection induced by porin preparations is due to antibodies against contaminating LPS 1°, by the use of denatured OMP preparations contaminated with LPS as antigens in immunoblots 3'8 and sera containing anti-LPS anti- bodies 3's'1°, and by the presence in normal sera of antibodies crossreacting with antigens of Gram-negative pathogens 8. Moreover, in vitro experiments have sug- gested that monoclonal antibodies against OMPs do not reach their targets in Gram-negative bacteria because of steric hindrance produced by LPS and capsular poly- saccharides; this rules out a role for them in protective immunity against infection through the stimulation of phagocytosis and complement activation 15.16. Immuno- protection against typhoid fever has been achieved with killed parenteral vaccines and live oral vaccines with different degrees of success 17, suggesting that both humoral and cellular host immune responses are involved in the ability of humans to deal with S. typhi ~7. The relevance of the humoral response in immunoprevention of typhoid is highlighted by the high effectiveness of

Radioimmunoprecipitation
acetone-killed whole-cell vaccine 17.18, and the effectivity of the Vi antigen vaccine17,1 s,19. Nonetheless, the relative importance of humoral and cellular immunity in the protection and recovery from S. typhi infections remains uncharacterized 17,1 s, and there is no information on the antigens responsible for these responses17,1a. Improved characterization of the immune response to different S. typhi OMP antigens may be important in order to identify new S. typhi immunogens and to develop new vaccines 14'17'18'2°. This information is essential if attenu- ated live S. typhi vaccines are going to be used both to prevent typhoid fever and as carriers for presentation of cloned antigens to the immune system 2°-22. The unique pathogenesis, epidemiology and clinical presentation of S. typhi infections suggest that the host immune response to S. typhi may also have unique features whose characterization cannot be replaced by the study of other Salmonellae 3,7,18.
Serological assays based on immune responses to O, H and Vi antigens to diagnose typhoid fever patients and S. typhi carriers in endemic areas lack specificity and sensitivity, stressing the need for improved immuno- logical assays to diagnose the diseased and carrier state 17'18'23'24. The immunogenicity of S. typhi OMPS suggests the potential for developing diagnostic assays to detect antibodies against these proteins in typhoid fever.
Thus, the potential role of Salmonella OMPs in virulence and their immunogenicity suggests that a characterization of the host immune response to Salmonella OMPs will improve the understanding of the biology of this pathogen.
MATERIALS AND M E T H O D S
Bacterial strain and growth conditions S. typhi Ty2, a variant from our laboratory that lacks
the Vi antigen, was grown in 100 ml of modified Hershey medium z5 at 37°C to an absorbance at 600 nm (A60o) of 0.5 in the presence or absence of dipyridyl (200 #M, Sigma) and will be referred to as induced and uninduced cells, respectively 8.
Buffers The following buffers were used: phosphate-buffered
saline (PBS), pH7.4; phosphate-buffered iodide (PBI, similar to PBS, except that 50 mM of KI was substituted for NaCI), and PBI~0.02% sodium azide (PBIN). The OMP solubilization buffer was prepared as described previously z6 except that 0.1% (w/vol) sodium deoxy- cholate (Sigma) was always present.
Sera A pool was made with the sera obtained from eight
blood samples of typhoid fever patients, whose diagnosis was confirmed by blood culture. Serum from an individual typhoid fever patient was obtained immediately and 1, 2 and 3 weeks after the diagnosis of the disease. The normal human serum was a pool made from the blood of six healthy volunteers from a non-endemic area, without a history of typhoid fever. Another convalescent serum and one normal serum were stored unpooled. All sera were incubated 30min at 56°C to inactivate complement. S. typhi-absorbed sera were prepared as described previously eT. Immunoglobulin G (IgG) of normal and convalescent sera were prepared by three
of Salmonella typhi outer membrane proteins: L. Aron et al.
rounds of ammonium sulphate precipitation followed by ion-exchange chromatography on DEAE-cellulose (DE- 52, Whatman).
Removal of anti-S, typhi LPS antibodies Anti-S. typhi LPS antibodies were removed by re-
peated passages over purified S. typhi LPS coupled to Sepharose, and will be described elsewhere (Aron, Di Fabio and Cabello, unpublished results). To ensure complete removal of anti-S, typhi LPS antibodies, a second method was used after column absorption. This method can only be used with DEAE-cellulose-purified IgG. Purified smooth S. typhi LPS (1 mg) was added to an aliquot of convalescent IgG, incubated for 48 h at 4°C and centrifuged for 1 h at 20000 rev min-1 at 4°C in a SS-34 Sorvall centrifuge rotor. The supernatant was dialysed against 75 mM phosphate-Tris, pH 8.0. In this buffer, at least 0.8 mg of purified LPS could be bound to 1 ml of DEAE-cellulose, while the IgG remained unbound. To remove excess LPS, the IgG + LPS sample was passed over a DEAE-cellulose column equilibrated in this buffer. Complete removal of anti-lipid A and anti-Salmonella LPS core antibodies was achieved by absorption to nitrocellulose strips loaded with Salmonella typhimurium TVll9 Ra LPS (Sigma), and Salmonella minnesota 595 Re LPS (Sigma) and will be described elsewhere (unpublished results). The lack of anti-LPS antibodies after absorption was assessed by ELISA and immunodot assay (unpublished results). LPS-ELISA was performed in the following way: purified smooth S. typhi LPS (100/A/well, 5/zg/m1-1) was used to coat the wells of disposable polystyrene microtitre plates (Nunc Immunoplates II, Denmark) in 50 mM sodium carbonate buffer, pH 9.6. After an overnight incubation (37°C), the wells were washed three times with 0.05% Tween 20 (Bio-Rad) in phosphate-buffered saline (PBS-Tween). The plates were incubated for 2h at 37°C with 100#1 of serial dilutions of the sera in PBS-Tween+4% bovine serum albumin (BSA, Sigma), and then washed three times with PBS-Tween (200 #1 each time). The second antibody used was the alkaline phosphatase conjugate of goat anti-human IgG (Kirkegaard and Perry, 100/A for each well, 1:3000 dilution in PBS-Tween + 1% BSA, 2 h at 37°C). The second antibody was removed, the wells washed with 200/~1 of PBS-Tween and the wells were finally incubated with 100 #1 of p-nitrophenylphosphate (lmgm1-1, Sigma) in diethanolamine (Sigma) buffer (9.7% diethanolamine, pH9.8, 9.5mM MgCI:). The reaction was stopped with 25/A of 3 M NaOH. Assays were carried out in duplicate for each dilution and read at 405 nm against a reagent blank in a microtitre well reader (titertek Multiscan MCC/340). Purified immuno- globulin concentrations were measured by their extinc- tion at 280 nm (Spectronic 21 spectrophotometer) or by reaction with BCA Protein Assay Reagent (Pierce). The final antibody concentrations were adjusted to that present in their sera of origin.
Radiolabelling of S.typhi cells Induced cells (100ml) were intrinsically and extrinsi-
cally radiolabelled with 35S-methionine and Na125I, respectively. For intrinsic radiolabeUing, cells were grown as indicated in the presence of 4mCi 3SS-methionine (1470 Ci mmol- 1, Amersham); for extrinsic radiolabelling, the procedure described by Hansen et al. was used 2s.
Vaccine, Vol. 11, Issue 1, 1993 11

Radioimmunoprecipitation of Salmonella typhi outer membrane proteins: L. Aron et al.
Briefly, after growth, the cells were collected by centri- fugation at 7000 rev min-1 for 10 min at 4°C in a Sorvall SS-34 rotor, washed twice with PBS and resuspended in 2ml ice-cold PBS. Next, 2nmol KI and 4mCi Na125I (17.4mCiktg -1, Amersham) were added to the cell suspension, and the radioiodination started by the addition of 50/~g of lactoperoxidase (Sigma) and 50 #1 of 0.03% hydrogen peroxide (Sigma). The same quantities of both reagents were added to the reaction mixture after 4, 8, 12 and 16min. At 20min, radioiodination was stopped by the addition of 20ml cold PBIN. Both intrinsically and extrinsically radiolabelled cells were collected by centrifugation and washed five times with 10ml PBS or PBI, respectively.
Whole-cell radioimmunoprecipitation (whole-cell RIP) The procedure described by Hansen et al. was
followed 28. The final pellets of radiolabelled cells were each resuspended in 6 ml cold PBS supplemented with Aprotinin (10/~g ml- 1, Sigma), human plasma ~- 1 anti- trypsin (10#gm1-1, Sigma), and a-toluene sulphonyl fluoride (0.1 mM, Eastman). Next, 0.5 ml aliquots of cells were mixed with 400#1 of S. typhi-absorbed patient sera, patient and normal sera IgG, the last two containing or devoid of anti-S, typhi LPS, and incubated for 16 h at 4°C. A sixth aliquot was mixed with PBS and used as a control. The remaining 3.0ml were kept for membrane preparation. Subsequently, the cells were washed five times with PBS and lysed by a 6.89 × 10 7 p.s.i. cycle on a French Press (American Instrument Co.), and the outer membrane isolated as described 29, except that dithiothreitol was omitted and the sucrose gradients were centrifuged at 42000revmin -1 for 8h at 5°C in a SW50.1Ti rotor (Beckman). The solutions used were always supplemented with the above-mentioned mixture of protease inhibitors. After collection (Haake Buchler Auto Densi-Flow), the fractions containing the outer membrane vesicles were pooled, resuspended in I ml O M P solubilization buffer, and incubated for 2 h at 37°C. The unsolubilized membranes were removed by centrifugation for l h at 13000revmin -1 in a Sorvall HB-4 rotor with adaptors for Eppendorf tubes. The supernatants were added to 50/d of either Protein A-Sepharose (Sigma) or Staphylococcus aureus Cowan I strain (10%, Sigma) suspensions. After incubation for 1 h at 4°C, the Protein A-Sepharose incubates were transferred to 10ml syringes provided with sintered glass filters, washed ten times with 2 ml solubilization buffer and three times with 62.5 mM Tris-HC1, pH 6.8, and 10% glycerol, and transferred to Eppendorf tubes. The supernatants were carefully discarded after centri- fugation and 35/~1 of 62.5mM Tris-HC1, pH6.8, 10% glycerol, 5% mercaptoethanol,0.04% bromophenol blue and 5% SDS were added to each tube, mixed with the Sepharose and boiled for 10min. The tubes were centrifuged in an Eppendorf microfuge, the supernatants withdrawn, and the extraction repeated once. Both ex- tracts were pooled, and aliquots ( ~ 20 000 counts min- 1 for 35S and 10 000 counts min -1 for 1251) were loaded onto 10-20% acrylamide gradient gels and electro- phoresed at 75V for 20h, using the discontinuous dissociating buffer system of Laemmli 3°. The S. aureus incubates were processed essentially as described for protein A-Sepharose, except that the washings were performed in Eppendorf tubes, followed each time by a centrifugation step. Cold molecular weight standards
(Sigma), plus added Aprotinin as a 6.5 kDa standard 31 and cold membrane standards were always included on every gel to facilitate identification and ensure repro- ducibility. Staining and destaining of gels was done according to Weber and Osborn 32. Gels containing 35S samples were soaked for 1 h in 3% glycerol-5% methanol, followed by 1 h in 1M sodium salicylate. Both 1251 and 35S gels were dried (Bio-Rad Slab Dryer), followed by radioautography on Kodak X-OMAT Ar film. Exposure lasted from 3 to 7 days at -70°C.
Outer-membrane radioimmunoprecipitation (outer-membrane RIP)
The remaining 3.0 ml of extrinsically and intrinsically radiolabelled induced cells were lysed and outer mem- brane vesicles prepared as described above. Aliquots (5x 106-107 counts min -1) were incubated for 16h at 4°C with 50-400#1 of the same sera and IgG used for whole-cell RIP. Unreacted serum was removed after centrifugation for 1 h in a Sorvall HB-4 rotor, followed by three washings with PBS and centrifugation. Solubil- ization of the pellet and the following steps were essentially as described above, except that 50/A of S. aureus Cowan I strain were used instead of Protein A-Sepharose.
R E S U L T S
Characterization of labelled and unlabelled outer membrane proteins of S. typhi
As a preliminary step in these studies We completed the characterization of S. typhi OMPs. Previous work indicated that the analysis of the OMPs of S. typhi by SDS-PAGE reveals a pattern similar to that of other Gram-negative Enterobacteriaceae 3'33, where the major proteins are represented by the porins and lipoprotein (Figure la). As described previously, the growth of S. typhi in iron-starvation conditions induces the appear- ance of three new proteins of 83, 78 and 69kDa 8. Subsequent SDS-PAGE analysis of the OMPs ofS. typhi strains of different origin has suggested that the expres- sion of these proteins is conserved in all S. typhistudied 33. The characterization of the 1251- and 35S-labelled outer and inner membranes of S. typhi shown in Figure lb, indicates that the labelled OMPs are the same as in the unlabelled preparations and that they are different from the inner membrane proteins. However, the 80 kDa range proteins are poorly labelled with 35S-methionine when compared both with the rest of the proteins and with the relative intensity of the proteins labelled with 125I. Moreover, these results indicate that there are only small differences between the protein patterns of outer mem- branes solubilized directly with loading buffer and after extraction of the outer membranes with OMP-solubilizing buffer.
Absorption of anti-S, typhi lipopolysaccharide antibodies The anti-S, typhi LPS antibodies present in con-
valescent and normal sera were absorbed to ensure the specificity of the radioimmunoprecipitations of the OMPs of S. typhi. Figure 2a shows an immunodot assay for smooth S. typhi LPS. It can be seen that the convalescent IgG pool had a positive titre of at least 1:3200. LPS-ELISA gave the same results (data not shown). The titre of the individual typhoid fever convalescent serum remained constant at up to 1:800
12 Vaccine, Vol. 11, Issue 1, 1993

a a b
Radioimmunoprecipitation of S a l m o n e l l a typhi outer membrane proteins: L. Aron et al.
c b a b c d e f g h i
9 4 . 7 -
6 6 . 7 -
4 2 . 7 -
31.0 -
21.5-
14 .4 -
9 4 . 7 ,
6 6 . 2 - 4
4 2 . 7 ,
31
2 1 . 5 - - -
1 4 . 4
6 . 5
Figure 1 Salmonella typhi outer membrane proteins. (a) Unlabelled outer membrane proteins of S. typhi. Acrylamide gradient (10-20%) SDS gel of uninduced (lane b) and induced (lane c) S. typhi OMP. Lane a contains protein molecular weight markers, indicated on the left in kDa. (b) Autoradiography of a 10-20% acrylamide gradient SDS gel of induced S. typhi membrane proteins. Lane a: unlabelled protein molecular weight markers, indicated on the left in kDa; lane c and g: cold outer membranes; lanes b, d and e: labelled with1251; lanes f, h and i: labelled with 3SS-methionine; lanes b and f: inner membrane; lanes d and h: outer membranes solubilized directly in loading buffer; lanes e and i: aliquots of outer membrane proteins extracted in solubilizing buffer prior to loading
over the evolution of the disease. After absorption of the anti-S, typhi LPS antibodies from the convalescent serum pool, no reaction could be detected at a 1:2 dilution. This result was confirmed by LPS-ELISA (data not shown).
Figure 2b shows an immunodot assay with S. typhi- murium TVll9 R, LPS. The serum of the individual patient showed a constant titre of l:1600 over the evolution of the disease. The convalescent IgG pool had a low titre of 1:200, which was completely abolished by passage over LPS-Sepharose followed by strip absorption.
Figure 2C shows a Western blot of S. typhi smooth LPS and S. typhimurium TV119 R, LPS, reacted with typhoid fever convalescent IgG pool dilutions with another convalescent serum (anti-S. typhi smooth LPS and R, LPS titres were 1:3200 and 1:400, respectively, before absorption) after three and six absorptions with S. typhi cells and with the convalescent IgG pool after LPS-Sepharose and R a LPS strip absorption. These Western blots confirmed the high anti-S, typhi smooth LPS and the low anti-R a LPS titres of the convalescent IgG pool. They also show that extensive absorption with whole bacteria removed anti-O antigen antibodies, whereas it was incapable of doing so for anti-R, LPS antibodies. Only absorption with purified LPS seemed capable of eliminating both antibodies. Section 2 of Figure 2c confirmed the low titre of the individual patient serum pool obtained during the evolution of the disease
against smooth S. typhi LPS and its high titre against R a LPS.
Whole-cell and membrane radioimmunoprecipitations Finally the identification of the immunogenic OMPs of
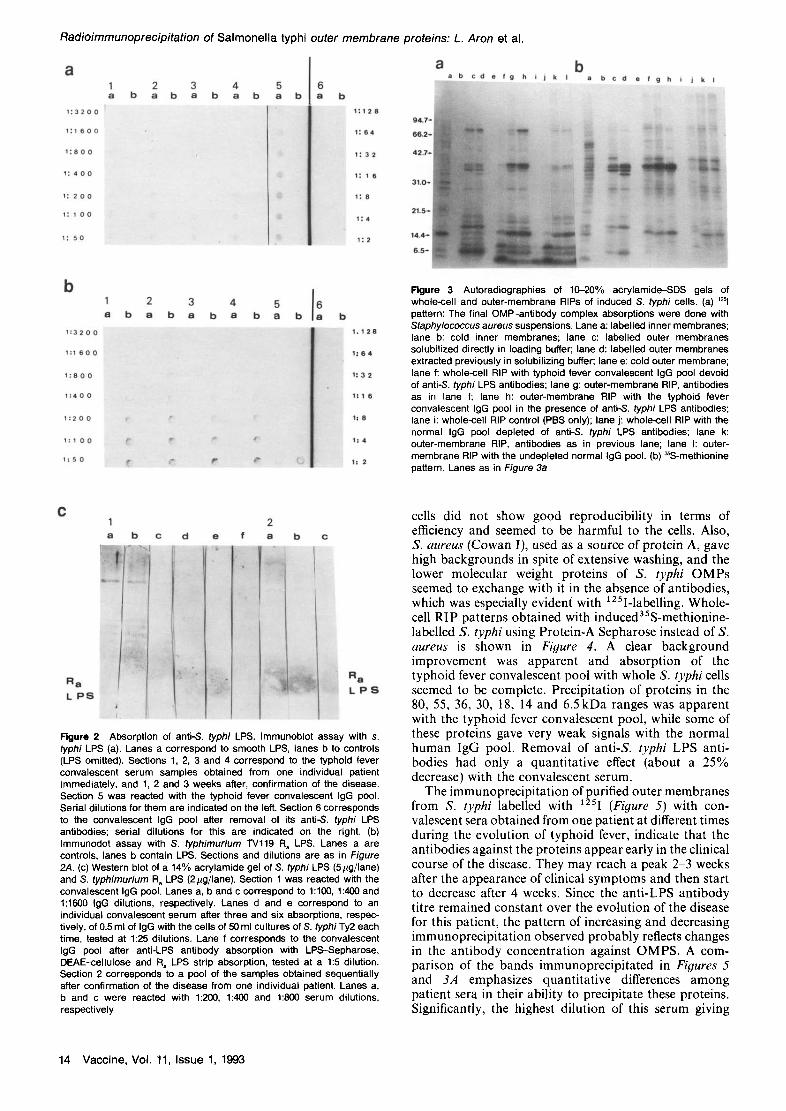
S. typhi was performed using radioimmunoprecipitation assays of labelled whole cells and membranes. Patterns obtained with 125I (Figure 3a) and 35S-methionine labelling (Figure 3b) were qualitatively similar except for the 80 kDa and 6.5 kDa range. Both the iron-starvation- induced proteins and lipoprotein were poorly labelled with 35S-methionine. In both cases, outer and inner membrane protein patterns were qualitatively different, although some cross-contamination was apparent. Removal of anti-& typhi LPS antibodies from the convalescent sera seemed to have mainly a quantitative effect, except for the 69kDa iron-starvation-induced protein, which was not apparent when normal human serum was used, and its precipitation seemed to be independent from the presence of anti-LPS antibodies. Otherwise, normal and convalescent sera exhibited similar qualitative patterns, although large quantitative differences are apparent for the 36 kDa range proteins. Patterns obtained with whole-cell and outer-membrane RIP were qualitatively identical. The 125I-pattern also showed quantitative increases in the precipitation of 30, 18 and 15kDa proteins when performed with con- valescent sera. Unfortunately, radioiodination of S. typhi
Vaccine, Vol. 11, Issue 1, 1993 13
Radioimmunoprecipitation of S a l m o n e l l a typhi outer membrane proteins: L. Aron et al.
a 1 =1
1 : 3 2 o o
1 : 1 6 0 0
1 ; 8 0 0
1 : 4 0 0
1 : 2 0 0
1 : 1 0 0
1 : 5 0
2 b a b
3 4 5 a b a b a b
i
i i
ii
6 a b
1 : 1 2 8 9 a L 7 -
1 : 6 4 6 6 . 2 -
1:3 = 42.7-
1 : 1 6 3 1 . 0 "
21 .5~ 1:4
14 .4~ 1 : 2
6 .S ~
a b a b c rl • f g h i j k I a b c d a f ¢m h i i u m
b
1 : 3 2 0 0
1 : 1 6 0 0
1 : 8 0 0
1 : 4 0 0
1 : 2 0 0
1:1 O 0
1 ; 5 0
b 2 3 4 5 a b a b a b a
6 a b
1 . 1 2 6
1 ; 6 4
1 : 3 2
1 : 1 6
1 : 8
1=4
1 : 2
Figure 3 Autoradiographies of 10-20% acrylamide-SDS gels of whole--cell and outer-membrane RIPs of induced S. typhi cells. (a) ~251 pattern: The final OMP-antibody complex absorptions were done with Staphylococcus aureus suspensions. Lane a: labelled inner membranes; lane b: cold inner membranes; lane c: labelled outer membranes solubilized directly in loading buffer; lane d: labelled outer membranes extracted previously in solubilizing buffer; lane e: cold outer membrane; lane f: whole-cell RIP with typhoid fever convalescent IgG pool devoid of anti-S, typhi LPS antibodies; lane g: outer-membrane RIP, antibodies as in lane f; lane h: outer-membrane RIP with the typhoid fever convalescent IgG pool in the presence of anti-S, typhi LPS antibodies; lane i: whole-cell RIP control (PBS only); lane j: whole-cell RIP with the normal IgG pool depleted of anti-S, typhi LPS antibodies; lane k: outer-membrane RiP, antibodies as in previous lane; lane I: outer- membrane RIP with the undepleted normal IgG pool. (b) ~S-methionine pattern. Lanes as in Figure 3a
C 1
a b
~i!i ~i!' !i I
L P 8
C d e f
. . . .
........ /
} ;
2 a b c
R 8
L P S
Figure 2 Absorption of anti-S, typhi LPS. Immunoblot assay with s. typhi LPS (a). Lanes a correspond to smooth LPS, lanes b to controls (LPS omitted). Sections 1, 2, 3 and 4 correspond to the typhoid fever convalescent serum samples obtained from one individual patient immediately, and 1, 2 and 3 weeks after, confirmation of the disease. Section 5 was reacted with the typhoid fever convalescent IgG pool. Serial dilutions for them are indicated on the left. Section 6 corresponds to the convalescent IgG pool after removal of its anti-S, typhi LPS antibodies; serial dilutions for this are indicated on the right. (b) Immunodot assay with S. typhimurium TVl19 R a LPS. Lanes a are controls, lanes b contain LPS. Sections and dilutions are as in Figure 2,4. (c) Western blot of a 14% acrylamide gel of S. typhi LPS (5#g/lane) and S. typhimurium Ra LPS (2 #g/lane). Section 1 was reacted with the convalescent IgG pool. Lanes a, b and c correspond to 1:100, 1:400 and 1:1600 IgG dilutions, respectively. Lanes d and e correspond to an individual convalescent serum after three and six absorptions, respec- tively, of 0.5 ml of IgG with the cells of 50 ml cultures of S. typhi Ty2 each time, tested at 1:25 dilutions. Lane f corresponds to the convalescent IgG pool after anti-LPS antibody absorption with LPS-Sepharose, DEAE-cellulose and R, LPS strip absorption, tested at a 1:5 dilution. Section 2 corresponds to a pool of the samples obtained sequentially after confirmation of the disease from one individual patient• Lanes a, b and c were reacted with 1:200, 1:400 and 1:800 serum dilutions, respectively
cells did not show good reproducibility in terms of efficiency and seemed to be harmful to the cells. Also, S. aureus (Cowan I), used as a source of protein A, gave high backgrounds in spite of extensive washing, and the lower molecular weight proteins of S. typhi OMPs seemed to exchange with it in the absence of antibodies, which was especially evident with 125I-labelling. Whole- cell RIP patterns obtained with induced35S-methionine- labelled S. typhi using Protein-A Sepharose instead of S. aureus is shown in Figure 4. A clear background improvement was apparent and absorption of the typhoid fever convalescent pool with whole S. typhi cells seemed to be complete. Precipitation of proteins in the 80, 55, 36, 30, 18, 14 and 6.5kDa ranges was apparent with the typhoid fever convalescent pool, while some of these proteins gave very weak signals with the normal human IgG pool. Removal of anti-S, typhi LPS anti- bodies had only a quantitative effect (about a 25% decrease) with the convalescent serum.
The immunoprecipitation of purified outer membranes from S. typhi labelled with 1251 (Figure 5) with con- valescent sera obtained from one patient at different times during the evolution of typhoid fever, indicate that the antibodies against the proteins appear early in the clinical course of the disease. They may reach a peak 2-3 weeks after the appearance of clinical symptoms and then start to decrease after 4 weeks. Since the anti-LPS antibody titre remained constant over the evolution of the disease for this patient, the pattern of increasing and decreasing immunoprecipitation observed probably reflects changes in the antibody concentration against OMPS. A com- parison of the bands immunoprecipitated in Figures 5 and 3A emphasizes quantitative differences among patient sera in their ability to precipitate these proteins. Significantly, the highest dilution of this serum giving
14 Vaccine, Vol. 11, i ssue 1, 1993

Radio immunoprec ip i ta t ion of Salmone l la typhi outer membrane proteins: L. Aron et al.
a b c d e f g h i a b c d e f g h i j k I m n o
94
6 6
4 2
31.~
21.
1 4 . ,
6.1
Figure 4 Autoradiography of ~S-methionine whole-cell RIP of induced S. typhi cells. The final OMP-antibody complex absorption was done with protein A~Sepharose. Lane a: molecular weight markers; lane b: labelled outer membranes; lane c: cold outer membranes; lane d: reaction with the typhoid fever convalescent IgG pool; lane e: reaction with the same antibody pool after six absorptions with S. typhi Ty2 cells; lane f: reaction with normal human IgG pool; lane g: reaction with the typhoid fever convalescent IgG pool devoid of anti-S, typhi LPS antibodies; lane h: reaction with the normal human IgG pool depleted of anti S. typhi LPS activity; lane i: control, antiserum substituted with PBS
a detectable signal for anti-S, typhi LPS antibodies by immunodot assay was 1:400, whereas for the con- valescent IgG pool it was 1:3200. Nevertheless, this serum had high titres against S. typhimurium R a LPS (1:1600) as opposed to the convalescent pool (1:200) when assayed by immunodot blotting, suggesting that the increased immunoprecipitation of iron-starvation- induced proteins observed with unabsorbed sera was due to anti-S, typhi O-chain antibodies.
The results of serum absorption with S. typhi whole cells for another typhoid fever convalescent serum are shown in Figure 5. The titre of this serum against S. typhi LPS was also 1:3200 when tested by immunodot blotting. After six absorptions of this serum, the x25I-outer membrane-RIP pattern is indistinguishable from the control, while the pattern obtained for the same un- absorbed serum (lane m), is very similar to the one corresponding to the unabsorbed convalescent IgG pool (Figure 3a, lane h). The bands immunoprecipitated after three absorptions are reminiscent of the pattern obtained with the convalescent IgG pool after removal of the anti-S, typhi LPS antibodies (Figure 3a, lane g), and the pattern corresponding to the individual patient with a low anti-S, typhi O-antigen titre (Figure 4).
9 4 . 7
6 6 . 2
4 2 . 7
31
21.5
14 .4
6 .5
Figure 5 Evolution of antibody response to S. typhi OMPs in typhoid fever: autoradiography of a 10-20% gel of induced 12Sl outer-membrane RIP. The final OMP-antibody complex absorptions were done with Staphylococcus aureus suspensions. Lane a: cold molecular weight markers; lanes b and c: I~I-S. typhi outer membranes without and with prior extraction with solubilizing buffer; lane d: cold outer membranes; lanes e, f, g and h: reacted with 50#1 of serum samples obtained from one individual patient immediately after the typhoid fever diagnosis was confirmed and after 1, 2 and 3 weeks, respectively; lanes i and o: l~l-outer membrane controls (no serum added); lane j; cold outer membranes; lane k: ~l-outer membranes reacted with 50pl of an individual normal human serum; lanes I, m and n: l~l-outer membranes reacted with 50pl of another convalescent individual serum after six, none, and three absorptions with 50pl of S. typhi, respectively
DISCUSSION
This series of experiments indicates that the human host is able to mount a humoral immune response against OMP antigens on the surface of S. typhi cells. Our RIP experiments with LPS absorbed and unabsorbed sera indicate that OMPs of S. typhi can be carried over in the assay, both by antibodies against LPS, as happens mostly with the iron-starvation-induced FepA- like proteins, and more specifically by antibodies against protein epitopes, as occurs with the porins, and the 30, 18.5, 15 and 6.5 kDa proteins. The immunoprecipitation of similar proteins in experiments where the cells have been labelled with 1251 and 35S.methionine indicates that the labelling of these proteins is not an artefact produced by the iodination. Thus, the immunoprecipitation of S. typhi OMPs after absorption of anti-LPS antibodies and their interaction with the antibodies in whole cells suggest that these proteic antigens are exposed during the infective process as occurs in the bloodstream of the human host x8'29. Alternatively, it could be postulated that the cell preparations contain death cells debris that, by reacting with the antibodies, are immunoprecipitated in the assay. Immunofluorescence experiments with S. typhi whole-cell preparations and monoclonal antibodies against the 36kDa porin indicate that, under the experimental conditions used, there appears to be minimal disruption of the cell structure and leakage of cellular debris (data not shown), ruling out the possibility that the proteins precipitated in the assay are only the result of alterations in the cell architecture 34. The experi- ments indicating total absorption of anti-LPS O-antigen antibodies present in convalescent sera by whole S. typhi
V a c c i n e , Vol. 11, Issue 1, 1993 15

Radioimmunoprecipitation of Sa lmone l la typhi outer membrane proteins: L. Aron et al.
Ty2 cells can be explained by the lack of Vi antigen in our Ty2 variant. The accessibility to antibodies of OMPs of whole smooth S. typhi cells suggests that these OMPs of S. typhi and their antibodies show a different behaviour than the OMPs and antibodies of Escherichia coli and other Enterobacteriaceae, where the interaction between the antibodies and their target proteins in the outer membrane is hindered by the presence of LPS and capsular polysaccharide ~La6. It would seem that the surface of S. typhi resembles that of capsulated H. influenzae and P. aeruginosa, where antibodies are able to react with the OMP in whole cells without hindrance by the capsular polysaccharides and LPS, suggesting that general rules cannot be applied to the study of the interactions between bacterial surface antigens and their antibodies 35'36. Alternatively, these different results could be produced by variations in the density of the LPS and capsular polysaccharide of bacteria growing in dissimilar conditions 37, and by differences in the affinity and behaviour of human and animal antibodies in their interaction with the baterial cell surface 37. The former hypothesis is supported by experiments demonstrating that in E. coli iron deprivation, a condition used in our experiments, generates a differential expression of core and O-polysaccharide antigens 38.
The immunoprecipitation observed with IgG prepared from sera from normal people suggests that it is due to cross-reactivity between the OMPs of S. typhi, E. coli and other Gram-negative bacteria that are part of the normal flora 39. Some of the immune response observed in typhoid fever patients could be due to anamnestic clonal expansion of B cells able to recognize cross- reacting Gram-negative OMP epitopes. The existence of this cross-reactivity has been demonstrated by the ability of anti-E-coli FepA protein antibodies to recognize the FepA-like proteins of S. typhi 8. Nonetheless, in our experiments the specificity of the immunoprecipitation reactions is confirmed by the lack of immunoprecipi- tation when the sera are absorbed with whole S. typhi cells. Moreover, these experiments also suggest that the cross-reactivity observed by various investigators among Gram-negative OMPs could be partially produced by antibodies against cross-reacting LPS contaminating these preparations 39'4°. In summary, these results allow an improved interpretation of findings of previous work using immunoblots with isolated proteins and extracted OMPs, because they permit the detection of the humoral immune response to LPS and undenatured and exposed surface OMPs independently of each other 3'~ 1.4L42.
The immunoprecipitation of proteins induced by iron starvation and probably involved in iron transport, confirms previous observations by us that the iron uptake system seems to be fully induced in typhoid fever, suggesting a role for iron uptake in the pathogenesis of the disease and a role for these antibodies in host defence 8. These results also confirm the immunogenicity of proteins in S. typhi that are induced in vivo in the human host and they suggest that this methodology could be applied to study the immunogenicity of other in vivo induced proteins 5'8'43. More experimental work will be required to ascertain whether the immunoprecipitation of these surface proteins of S. typhi in reactions with whole cells could indicate that these antibodies may have relevance in the recovery from infection in humans by their ability to stimulate phagocytosis, complement activation and antibody-dependent cell killing ~8. This
may be an important point, because it has been shown that with antibody preparations able to recognize OMPs and protect against Gram-negative infections, the protection is in reality provided by antibodies directed against the small amounts of LPS contaminating the OMP preparations 1°. In the case of the antibodies against the FepA-like protein, it could be postulated that they play a role in blocking iron uptake and helping the human host to overcome the infection by inhibition of bacterial growth. Similarly, antibodies against porins may block the virulence potential of these proteins in S. typhi infections, as work in S. typhimurium indicates a role for porins in oral virulence for mice 6. Moreover, anti-FepA and anti-porin antibodies could also play a role in complement-mediated lysis, phagocytosis and antibody-dependent cell-mediated cytotoxicity. The use of monoclonal antibodies against these proteins in in vitro assays to detect their binding to whole S. typhi cells and studies of the interaction of purified OMPs with the immune system will complete this characterization. These experiments and the identical nature of the OMPs of S. typhi isolates of different origin indicate that the protein composition of the OMPs is quite conserved, as is the immunoresponsiveness to them 3'14'3a. Potential diagnostic or preventive methods based on these structural proteins will not have to overcome problems posed by variation of these structures among different S. typhi strains.
Radioimmunoprecipitation experiments with un- absorbed and differentially absorbed sera, such as those described here, may also be useful to characterize protective antibodies induced by the effective acetone- killed whole-cell vaccine 17'18. Moreover, they could be used to investigate the humoral immune response to intracellular S. typhi antigens and to foreign antigens expressed in Salmonella vaccines T M . The analysis of the host immune response to surface proteins of S. typhi should be complemented by the study of the response to S. typhi proteins that, as in S. typhimurium, may be induced during its interactions with epithelial cells and inside macrophages TM.
A C K N O W L E D G E M E N T S
This work was supported by grant RO1 AI33613 from the National Institute of Allergy and Infectious Diseases and grants from the American Heart Association and from the Eastern Paralyzed Veterans Association to Felipe Cabello. Carlos Gonzalez was the recipient of a faculty fellowship from the Educational Commission for Foreign Medical Graduates. The authors are grateful to Mrs Harriett V. Harrison for her excellent secretarial assistance.
REFERENCES
1 Fernandez-Beros, M.E., Kissel, V., Lior, H. and Cabello, F.C. Virulence-related genes in ColV plasmids of Escherichia coil isolated from human blood and intestines. J. C/in. Microbiol. 1990, 28, 742-746
2 Aguero, ME., Aron, L., De Luca, A.C., Timmis, K.N. and Cabello, F.C. A plasmid-encoded outer membrane protein, TraT, enhances resistance of Escherichia coil to phagocytosis. Infect. Immun. 1984, 46, 740-746
3 Aguero, J., Mora, G., Mroczenski-Wildey, M.J., Fernandez-Beros, M.E., Aron, L. and Cabello, F.C. Cloning, expression and character- ization of the 36 kDal Salmonella typhi porin gene in Escherichia coil Microb. Pathog. 1987, 3, 388-407
16 Vaccine, Vol. 11, Issue 1, 1993

Radio immunoprec ip i ta t ion of S a l m o n e l l a typhi outer membrane proteins: L. Aron et al.
4 Hacket, J., Wyck, P., Reeves, P. and Mathan, V. Mediation of serum resistance in Salmonella typhimurium by an 11 kilodalton poly- peptide encoded by the cryptic plaemid. J. Infect. Dis. 1987, 55, 540-549
5 Miller, S.I., Kutral, A. and Mekalanos, J.J. A two component regu- latory system (phoP and phoQ) controls Salmonella typhimurium virulence. Proc. Natl Acad. Sci. USA 1989, 88, 5054-5058
6 Chatfield, S.N., Dorman, D.J., Hayward, C. and Dougan, D. Role of ompR- dependent genes in Salmonella typhimurium virulence: Mutants deficient in both OmpC and OmpF are attenuated in vivo. Infect. Immun. 1991, 59, 449-452
7 Pulkkinen, W.S. and Miller, S.I. A Salmonella typhimurium virulence protein is similar to Yersinia enterocolitica invasion protein and bacteriophage lambda outer membrane protein. J. Bacteriol. 1991, 173, 86-93
8 Fernandez-Beros, ME., Gonzalez, C., Mclntosh, M. and Cabello, F.C. Immune response to the iron-deprivation induced proteins of Salmonella typhi in typhoid fever. Infect. Immun. 1989, 57, 127%1275
9 Calderon, I., Lobos, S.R., Rojas, H.A., Palominos, C., Rodriguez, L.H. and Mora, G. Antibodies to porin antigens of Salmonella typhi induced during typhoid fever in humans. Infect. Immun. 1986, 52, 209-212
10 Saxen, H., Nurminen, M., Kuusi, N., Soerison, S.B. and Makela, P.H. Evidence for the importance of O antigenic antibodies in mouse- protective Salmonella outer membrane (porin) antisera. Microb. Pathog. 1986, 1,433-441
11 Sharma, P., Ganguly, N.E., Sharma, B.K., Sharma, S. and Seghal, R. Specific immunoglobulin response in mice immunized with porins and challenged with Salmonella typhi. Microb. Immunol. 1989, 33, 519-525
12 Gilleland, H.E., Parker, M.G, Matthews, JM. and Berg, R.D. Use of outer membrane protein F (porin) preparation of Pseudomonas aeruginosa as a protective vaccine in mice. Infect. Immun. 1984, 44, 49-54
13 Granoff, D.M. and Munson, R.S. Prospects for prevention of Haemophilus influenzae type b disease by immunization. J. Infect. Dis. 1986, 153, 44~461
14 Silva, B.A., Gonzalez, C., Mora, G.C. and Cabello, F. Genetic characteristics of the Salmonella typhi strain Ty21a vaccine. J. Infect. Dis. 1987, 155, 1077-1078
15 Bentley, A.T. and Klebba, P.E. Effect of lipopolysaccharide structure on reactivity of antiporin monoclonal antibodies with the bacterial cell surface. J. Bacteriol. 1988, 170, 1(363-1068
16 van der Ley, P., Kuipers, O., Tommassen, J. and Lugtenberg, B. O-antigenic chains of lipopolysaccharide prevent binding of antibody molecules to an outer membrane por protein in Enterobacteriaceae. Microb. Pathog. 1986, 1, 43-49
17 Levine, M.M., Ferreccio, C., Black, R.E., Tacket, C.O., Germanier, R. and The Chilean Typhoid Committee. Progress in vaccines against typhoid fever. Rev. Infect. Dis. 1989, 11(Suppl. 3), S552~$567
18 Edelman, R. and Levine, M.M. Summary of an international workshop on typhoid fever. Rev. Infect. Dis. 1986, 8, 329-349
19 Robbins, J.B., Schneerson, R., Acharya, I.L., Lowe, C.U., Szu, S.C., Daniels, E. et al. Protective roles of mucosal and serum immunity against typhoid fever. Monogr. Allergy 1988, 24, 315-320
20 Schultz, C.L., Kaufman, B., Hamilton, D., Hartman, A., Ruiz, M., Powell, C. and Berman, B. Cell wall structures which may be important for successful immunization with Salmonella-Shigella hybrid vaccines. Vaccine 1990, 8, 115-120
21 Formal, S.B., Baron, L.S., Kopecko, D.J., Washington, O., Powell, C. and Life, C.A. Construction of a potentially bivalent vaccine strain: Introduction of Shigella sonnei form I antigen genes into the Salmonella typhi Ty21a typhoid vaccine strain. Infect. Immun. 1981, 34, 746-750
22 Sadoff, J.C., Ballou, W.R., Baron, L.S., Majarian, W.R., Brey, R.N., Hockmayer, W.T. et al. Oral Salmonella typhimurium vaccine expressing circumsporozoite protein protects against malaria. Science 1988, 240, 336-338
23 Nolan, C.M., White, P.C., Felley, J.C., Brown, S.L. and Harnbie, E.A. Vi serology in the detection of typhoid carriers. Lancet 1981, I, 583-585
24 Widal, MF. Serodiagnostic de la fi~vre typhoide. Bull. Mem. Soc. Med. Hop. Paris 1986, 13, 881-568
25 Sancar, A., Hack, A.M. and Rupp, W.D. A simple method for identification of plasmid coded proteins. J. Bacteriol. 1979, 137, 692-693
26 Gulig, P.A., McCracken, G.H., Jr, Frisch, C.F., Johnston, K.H. and Hansen, E.C. Antibody response of infants to cell surface-exposed outer membrane proteins of Haemophilus influenzae type b after systemic Haemophilus disease. Infect. Immun. 1982, 37, 82~38
27 Aguero, M.E. and Cabello, F.C. Relative contribution of ColV plasmid and K1 antigen to the pathogenicity of Escherichia coll. Infect. Immun. 1983, 40, 359-388
28 Hansen, E.J., Frisch, C.F., McDade, R.L. Jr and Johnston, K.H. Identification of immunogenic outer membrane proteins of Haemo- philus influenzae type b in the infant rat model system. Infect. Immun. 1981, 32, 1084-1092
29 Osborn, M.J., Gander, J.E., Parisi, E. and Carson, J. Mechanism of assembly of the outer membrane of Salmonella typhimurium. Isolation and characterization of cytoplasmic and outer membrane. J. Biol. Chem. 1972, 247, 3962--3972
30 Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature (London) 1970, 227,
31 Anderer, F.A. and Hornie, S. The disulfide linkages in kallikrein inactivator of bovine lung. J. Biol, Chem. 1966, 241, 1568-1577
32 Weber, K. and Osborn, M.J. The reliability of molecular weight determination by dodecyl sulfate polyacrylamide gel electro- phoresis. J. Biol. Chem. 1989, 244, ~ 1 2
33 Faundez, G., Aron, L. and Cabello, F.C. Chromosomal DNA, iron-transport system, outer membrane proteins and enterotoxin (heat labile) production in Salmonella typhi strains. J. Clin. Microbiol. 1990, 28, 894-897
34 Hoekstra, D., Van Der Laan, J.W., De Leij, L. and Witholt, B. Release of outer membrane fragments from normal growing Escherichia coll. Biochim. Biophys. Acta 1978, 455, 88!)-899
35 Kimura, A., Gulig, P.A., McCraken, G.H. Jr, Loftus, A. and Hansen, E.J. A minor high-molecular-weight outer membrane protein of Haemophilus influenzae type B is a protective antigen. Infect. Immun. 1985, 34, 320-332
36 Mutharia, L and Hancock, R.E. Surface localization of Pseudomonas aeruginosa outer membrane porin protein F by using monoclonal antibodies. Infect. Immun. 1983, 42, 1027-1033
37 Grossman, N.A., Schnertz, M.A., Foulds, J., Klima, E.N., Jimenez, V., Leibe, L.L. and Joiner, K.A. Lipopolysaccharide size and distribution determine serum resistance in Salmonella montevideo. J. Bacteriol. 1987, 169, 856-863
38 Nelson, D., Bathgate, A.J. and Poxton, I.R. Monoclonal antibodies as probes for detecting lipopolysaccharide expression on Escherichia coil from different growth conditions. J. Gen. Microbiol. 1991, 137, 2741-2751
39 Hofstra, J., van Tol, M.J.D. and Dankert, J. Crossreactivity of major outer membrane proteins of Enterobacteriaceae, studied by crossed immunoelectrophoresis. J. Bacteriol. 1980, 143, 328-337
40 Isibasi, A., Ortiz, V., Vargas, M., Paniagua, J., Gonzalez, C., Moreno, J. and Kumate, J. Protection against Salmonella typhi infection in mice after immunization with outer membrane proteins isolated from Salmonella typhig, 12, d, Vi. Infect. Immun. 1988, 56, 2953-2959
41 Udhayakumar, V. and Muthukkarupan, V.R. Protective immunity induced by outer membrane proteins of Salmonella typhimurium in mice. Infect. Immun. 1987, 55, 816-821
42 Brown, A. and Hormaeche, C.E. The antibody response to Salmonellae in mice and humans: studies by immunoblots and ELISA. Microb. Pathog. 1989, 6,
43 Lee, C.A. and Falkow, A. The ability of Salmonella to enter mammalian cells is affected by bacterial growth state. Proc. Natl Acad. Sci. USA 1990, 87, 4304-4308
44 Wu, J.Y., Newton, S., Judd, A., Stocker, B. and Robinson, W.S. Expression of immunogenic epitopes of hepatitis B surface antigen with hybrid flagellin proteins by a vaccine strain of Salmonella. Proc. Natl Acad. Sci. USA 1989, 86, 4726-4730
45 Brown, A., Hormaeche, C.E., Demarco de Hormaeche, R., Winther, M., Dougan, G., Maskell, D.J. and Stocker, B.A.D. An attenuated aroA Salmonella typhimurium vaccine elicits humoral and cellular immunity to cloned/~-galactosidase. J. Infect. Dis. 1987,155, 86-92
46 Finlay, B.B. and Falkow, S. Salmonella as an intracellular parasite. Mol. Microbiol. 1989, 3, 1833-1841
Vaccine, Vol. 11, Issue 1, 1993 17




















