
The seroepidemiology of Bordetella pertussis in Israel—Estimate of incidence of infection
Upload
independentCategory
view
1download
0
Ii
EDa
b
c
a
ARRAA
KBIV
1
pbpTcor5fad
oipbbSF
0d
Vaccine 27 (2009) 542–548
Contents lists available at ScienceDirect
Vaccine
journa l homepage: www.e lsev ier .com/ locate /vacc ine
mmunoproteomic analysis of Bordetella pertussis and identification of newmmunogenic proteins
mrah Altındis a, Burcu E. Tefona, Volkan Yıldırıma, Erkan Özcengizb,örte Becherc, Michael Heckerc, Gülay Özcengiza,∗
Department of Biological Sciences, Middle East Technical University, 06531 Ankara, TurkeyVaccine Biologicals Research Company, Ankara, TurkeyInstitut für Mikrobiologie, Ernst-Moritz Arndt-Universität Greifswald, 17487 Greifswald, Germany
r t i c l e i n f o
rticle history:eceived 22 August 2008
a b s t r a c t
Bordetella pertussis is the causative agent of highly communicable respiratory infection whooping cough(pertussis) which remains one of the world’s leading causes of vaccine-preventable deaths. In the present
eceived in revised form 30 October 2008ccepted 3 November 2008vailable online 24 November 2008
eywords:ordetella pertussismmunoproteomics
study, total soluble proteins extracted from two B. pertussis strains, Tohama I and the local isolate Saadetwere separated by two-dimensional gel electrophoresis and analyzed by Western blotting for their reac-tivity with the antisera obtained from the mice immunized with inactivated whole cells as well asthose collected from the mice challenged intraperitoneally with live cells of each strain. Of a total of25 immunogenic proteins identified, 21 were shown to be the novel antigens for B. pertussis.
© 2008 Elsevier Ltd. All rights reserved.
tm
Bposrappc
2
2
accine candidates
. Introduction
Whooping cough (pertussis) is a highly contagious, acute res-iratory illness of humans that is caused by the gram-negativeacterium Bordetella pertussis. This bacterium is a strict humanathogen with no known animal or environmental reservoirs [1].he introduction of mass vaccination of children in the mid-20thentury decreased the incidence of pertussis; however, it is still onef the world’s leading causes of vaccine-preventable deaths [2]. Aseported by the World Health Organization (WHO), an estimated0 million cases and 300,000 deaths occur every year; case-atality rates in developing countries are estimated to be as highs 4% in infants (http://www.who.int/immunization monitoring/iseases/pertussis surveillance/en/, as of August 2008).
A new revolution in vaccine design has emerged from the usef post-genomic technologies [3,4]. Much of information aboutmmunogenic components can be derived from proteomics cou-led to Western blotting, namely immunoproteomics which has
een successfully applied for the discovery of antigens from variousacterial pathogens; examples including Helicobacter pylori [5,6],taphylococcus aureus [7], Bacillus anthracis [8], Shigella flexneri [9],rancisella tularensis [10], Corynebacterium diphtheriae [11], Strep-∗ Corresponding author. Tel.: +90 3122105170; fax: +90 3122107976.E-mail address: [email protected] (G. Özcengiz).
SS4cfTgv
264-410X/$ – see front matter © 2008 Elsevier Ltd. All rights reserved.oi:10.1016/j.vaccine.2008.11.020
ococcus pyogenes [12], Chlamydia pneumoniae [13] and Neisseriaeningitidis [14].
Sequencing of the genomes of B. pertussis, B. parapertussis and. bronchiseptica [15] provided an essential database for applyingroteomics approach to these organisms. First proteomics studyn B. pertussis involved an analysis of differential protein expres-ion under iron-limitation and iron-excess conditions [16]. Veryecently, a comparative surfaceome analysis of the vaccine strainnd the clinical strains of this pathogen was reported [17]. In theresent study, we report the first immunoproteomic approach to B.ertussis Tohama I and Saadet strains which involved the identifi-ation of immunogenic proteins of their total soluble proteomes.
. Material and methods
.1. Bacterial strains and preparation of total soluble proteomes
The strains of B. pertussis used in this study were Tohama I andaadet, the latter being a local strain. B. pertussis Tohama I andaadet strains were grown on Cohen-Wheeler agar medium [18] for8 h at 37 ◦C. For the preparation of whole cell lysates, the cells were
ollected and suspended in cold TE buffer and centrifuged at 4500 gor 10 min. The pellet was resuspended in 1 ml 0.04 M lysosyme inE buffer and incubated at 37 ◦C for 35 min, followed by a centrifu-ation at 16,000 g for 10 min. The pellet was solubilized in 8 M urea,ortexed at 4 ◦C for 30 min and centrifuged at 16,000 g for 10 min.
ccine 2
Tw
2
daf(Ra18Ttw5iaRsb(sD
2
WsiItepisp7sa
Sftrtc
2
fTsmtwaTt1p
s1Biwb
2
ewttasctT4eiti
2
f(e“mnwrstnfgeso(p
3
3
sgfsc
3
E. Altındis et al. / Va
he supernatant was stored at −20 ◦C. The protein concentrationas determined by Bradford technique [19].
.2. 2-D gel electrophoresis
IPG strips were passively rehydrated by applying 400 �l of rehy-ration buffer (8 M urea, 2 M thiourea, 2% w/v CHAPS, 28 mM DTTnd 0.5% v/v ampholyte 3-10) containing 350 �g protein sampleor 14 h. IEF was performed with commercially available IPG strips17 cm, pH 3–10, Bio-Rad) and the Protean IEF Cell (Bio-Rad USA).ehydrated strips were taken to the IEF process. The following volt-ge profile was used for IEF: 1 h 100 V; 1 h 300 V; 1 h 600 V; 1 h000 V; 2 h 3000 V; 2 h 5000 V followed by a linear increase to000 V. The final phase of 8000 V was terminated after 50,000 Vh.he IPG strips were equilibrated for 15 min each in 5 mL of solu-ion 1 (6 M urea, 50 mM Tris–HCl (pH 8.8), 30% v/v glycerin, 2%/v SDS, 50 mg DTT) and then in 5 mL of solution 2 (6 M urea,
0 mM Tris–HCl (pH 8.8), 30% v/v glycerin, 2% w/v SDS, 225 mgodacetamide) [20]. The isolated proteins were separated in 12%crylamide/bis-acrylamide gels with a Bio-Rad Cell system (Bio-ad, USA), applying approximately 25 mA per gel. To visualize theeparated proteins, each gel was stained with colloidal Coomassielue [21]. Coomassie stained gels were digitized by using a scannerHP Scanjet 4070 Photosmart scanner, USA). Spot pattern analy-es were accomplished by using the 2-D image analysis softwareelta2D version 3.3 (Decodon, Germany).
.3. Preparation of antisera against B. pertussis
B. pertussis Tohama and Saadet strains were grown on Cohen-heeler agar media for 48 h. The cells were suspended in 0.85%
aline solution to contain ca. 4 × 1010 bacteria/ml. For subcutaneousmmunization, the suspension was inactivated at 56 ◦C for 30 min.nactivated bacterial cells were used as the antigen and anti-B. per-ussis polyclonal antibodies were raised by immunizing mice. Forach strain, 10 mice received two subcutaneous injections of 0.5 mler animal at two week intervals between the first and second
njections. Their sera were collected and pooled 14 days after theecond injection. For intraperitoneal challenge, live cells were sus-ended in 5% sodium glutamate containing 1% casamino acid (pH:.4) solution containing approximately 4 × 1010 cells/ml. For eachtrain, 10 mice received one intraperitoneal injection of 50 �l pernimal and after 14 days their sera were collected and pooled.
In Western Blot analysis, the antisera Th (sc), Sa (sc), Th (ip) anda (ip) were used. The antisera Th (sc) and Sa (sc) were obtainedrom the mice which were subcutaneously immunized with inac-ivated whole cells of B. pertussis Tohama I and Saadet strains,espectively. The antisera Th (ip) and Sa (ip) were obtained fromhe mice which were intraperitoneally challenged with live wholeells of B. pertussis Tohama I and Saadet strains, respectively.
.4. Western blotting of 2-DE gels
Proteins from the identical, but non-stained gels were trans-erred to a NC membrane for 1 h at 400 mA in transfer buffer (25 mMris, 192 mM glycine, 2% w/v SDS and 20% v/v methanol) by usingemi dry blotter (Cleaver Scientific Ltd.). After the transfer, theembrane was blocked for 2 h with 10% skim milk in TBS solu-
ion (20 mM Tris and 5 M NaCl) at 37 ◦C. After rinsing for 10 minith TBS-Tween 20, the membrane was incubated with primary
ntibody, mouse anti-B. pertussis, at a dilution of 1:300 in 0.05%ween-20 in TBS (TTBS) containing 5% skim milk for 1 h at roomemperature on a gentle shaker. The membrane was rinsed for0 min and incubated with rabbit anti-mouse IgG-alkaline phos-hatase (Sigma), at a dilution of 1:15,000 in TTBS containing 5%
Tbo
7 (2009) 542–548 543
kim milk for 1 h. The membrane was then washed with TBS for0 min and developed with substrate (AP Conjugate Substrate Kit,io-Rad) until optimum color was developed. The immunoreactiv-
ty of each spot that gave positive signal in Western blot analysisas verified through their excision from 2-D gels followed by dot-lotting.
.5. Protein identification
The identifications were accomplished by mass spectrom-try according to established protocols. Briefly, protein spotsere excised from stained 2-D gels, destained and digested with
rypsin (Promega, Madison, WI, USA). For the extraction of pep-ides, the gel pieces were covered with 60 �l 0.1% trifluoroaceticcid in 50% CH3CN and incubated for 30 min at 40◦C. Peptideolutions were mixed with an equal volume of saturated �-yano-3-hydroxycinnamic acid solution in 50% acetonitrile-0.1%rifluoroacetic acid (v/v) and applied to a sample plate for MALDI-OF-MS. Mass analyses were carried out on the Proteome-Analyzer800 (Applied Biosystems). The three most abundant peptides inach MS spectrum were chosen for MS/MS experiment. The result-ng sequence data were included for the database search to increasehe reliability of protein identification. Mass accuracy was usuallyn the range between 10 and 30 ppm.
.6. Database searches
Amino acid sequences for B. pertussis proteins were obtainedrom Sanger Institute organism’s genome project web sitehttp://www.sanger.ac.uk/Projects/B pertussis/). The peak lists ofach protein spot were analyzed with the aid of “PMF” andMS/MS Ion Search” engines of MASCOT software (http://www.atrixscience.com/). The searches considered oxidation of methio-
ine and modification of cysteine by carbamidomethylation asell as partial cleavage leaving one internal cleavage site. Of the
esults given by the MASCOT software, those having a probabilitycore value higher than 53 were considered for successful pro-ein identification. To find out putative functions, protein accessionumbers of the identified spots were searched in the website
or B. pertussis. To predict epitopes of the identified immuno-enic proteins of B. pertussis, the artficial network based B-cellpitope prediction server ABCpred [22] was used. The PSORTb ver-ion 2.0.4 [23] was used for prediction of subcellular localizationf the identified immunogenic proteins. The SignalP webserverhttp://www.cbs.dtu.dk/services/signalp/) was employed for signaleptide prediction.
. Results
.1. Total soluble proteome
Total soluble proteomes of B. pertussis Tahoma I and Saadettrains were resolved by 2-DE. Three technical replicates of 2-DEels for each of the three independent biological samples were runor each strain. 350 �g of extracted proteins were applied to the IPGtrips. After being stained with colloidal CBB G250, over 600 spotsould be detected on the gels within the pI range 3–10 (Fig. 1).
.2. Identification of immunoreactive proteins
Western blotting of the 2-D gels using antisera designated ash (sc), Sa (sc), Th (ip) and Sa (ip), respectively as primary anti-ody and anti-mouse IgG as secondary antibody revealed a totalf 45 immunoreactive protein spots. These spots matched well to

544 E. Altındis et al. / Vaccine 2
Fig. 1. Dual channel 2-D imaging of B. pertussis strains Tohama I (green) and Saadet(red) (For interpretation of the references to color in this figure legend, the readeris referred to the web version of the ariticle.).
tiMc
3p
siaiic2s
4
bgs((Lh
Fig. 2. 2-D Western blot analysis of the total soluble proteome of B. pertussis strain (A) Tohagainst antiserum Sa (sc) and Saadet against antiserum Sa (ip).
7 (2009) 542–548
he spot patterns on the Coomassie-stained gels. The correspond-ng stained spots were excised, digested and analyzed by using
ALDI-TOF/MS. Forty out of 45 spots could be identified whichorresponded to 25 different gene products (Fig. 2A–D; Table 1).
.3. Epitope prediction, subcellular localization and signaleptides
The server ABCpred was used for B-cell epitope prediction. In ourtudy, the accuracy scores of the predicted epitopes of all identifiedmmunogenic proteins were within a meaningful range. PSORTbnalysis predicted that 13 of the 25 immunogenic proteins are local-zed in cytoplasm, 4 proteins on outer membrane and 1 proteinn the periplasm of the organism. The localization of 7 proteinsould not be determined. According to SignalP analysis, 5 of the5 immunogenic proteins were predicted to have a signal peptideequence (Table 1).
. Discussion
To date, a number of virulence determinants of B. pertussis haveeen identified to be immunogenic, including filamentous hemag-lutinin (FHA) and fimbriae (FIM) as two adhesins, pertactin (PRN),
erum resistance protein BrkA and tracheal colonization factorTcfA) as the autotransporters, three toxins, namely pertussis toxinPT), adenylate cyclase (CyaA) and dermonecrotic toxin (DNT), andPS, as extensively reviewed by Mattoo and Cherry [24]. Also, theeat shock protein 60 (Hsp60), which represents approximately 2%ama I against antiserum Th (sc), (B) Tohama I against antiserum Th (ip), (C) Saadet
E.Altındis
etal./Vaccine
27(2009)
542–548
545
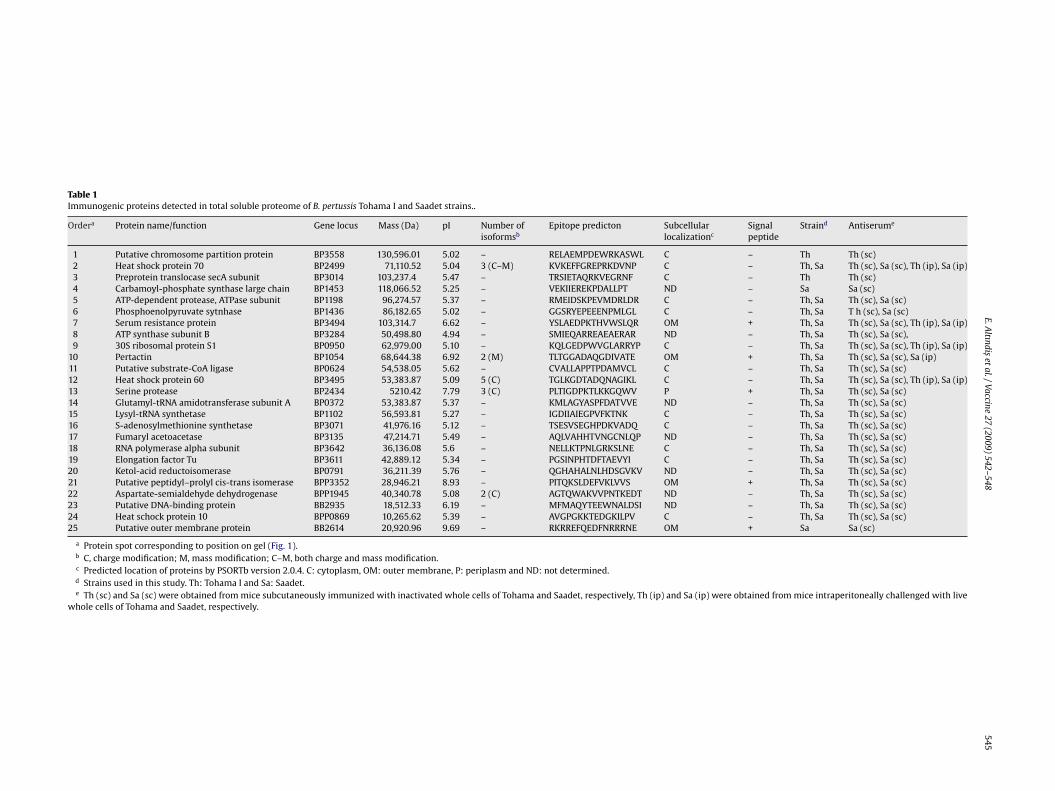
Table 1Immunogenic proteins detected in total soluble proteome of B. pertussis Tohama I and Saadet strains..
Ordera Protein name/function Gene locus Mass (Da) pI Number ofisoformsb
Epitope predicton Subcellularlocalizationc
Signalpeptide
Straind Antiserume
1 Putative chromosome partition protein BP3558 130,596.01 5.02 – RELAEMPDEWRKASWL C – Th Th (sc)2 Heat shock protein 70 BP2499 71,110.52 5.04 3 (C–M) KVKEFFGREPRKDVNP C – Th, Sa Th (sc), Sa (sc), Th (ip), Sa (ip)3 Preprotein translocase secA subunit BP3014 103,237.4 5.47 – TRSIETAQRKVEGRNF C – Th Th (sc)4 Carbamoyl-phosphate synthase large chain BP1453 118,066.52 5.25 – VEKIIEREKPDALLPT ND – Sa Sa (sc)5 ATP-dependent protease, ATPase subunit BP1198 96,274.57 5.37 – RMEIDSKPEVMDRLDR C – Th, Sa Th (sc), Sa (sc)6 Phosphoenolpyruvate sytnhase BP1436 86,182.65 5.02 – GGSRYEPEEENPMLGL C – Th, Sa T h (sc), Sa (sc)7 Serum resistance protein BP3494 103,314.7 6.62 – YSLAEDPKTHVWSLQR OM + Th, Sa Th (sc), Sa (sc), Th (ip), Sa (ip)8 ATP synthase subunit B BP3284 50,498.80 4.94 – SMIEQARREAEAERAR ND – Th, Sa Th (sc), Sa (sc),9 30S ribosomal protein S1 BP0950 62,979.00 5.10 – KQLGEDPWVGLARRYP C – Th, Sa Th (sc), Sa (sc), Th (ip), Sa (ip)
10 Pertactin BP1054 68,644.38 6.92 2 (M) TLTGGADAQGDIVATE OM + Th, Sa Th (sc), Sa (sc), Sa (ip)11 Putative substrate-CoA ligase BP0624 54,538.05 5.62 – CVALLAPPTPDAMVCL C – Th, Sa Th (sc), Sa (sc)12 Heat shock protein 60 BP3495 53,383.87 5.09 5 (C) TGLKGDTADQNAGIKL C – Th, Sa Th (sc), Sa (sc), Th (ip), Sa (ip)13 Serine protease BP2434 5210.42 7.79 3 (C) PLTIGDPKTLKKGQWV P + Th, Sa Th (sc), Sa (sc)14 Glutamyl-tRNA amidotransferase subunit A BP0372 53,383.87 5.37 – KMLAGYASPFDATVVE ND – Th, Sa Th (sc), Sa (sc)15 Lysyl-tRNA synthetase BP1102 56,593.81 5.27 – IGDIIAIEGPVFKTNK C – Th, Sa Th (sc), Sa (sc)16 S-adenosylmethionine synthetase BP3071 41,976.16 5.12 – TSESVSEGHPDKVADQ C – Th, Sa Th (sc), Sa (sc)17 Fumaryl acetoacetase BP3135 47,214.71 5.49 – AQLVAHHTVNGCNLQP ND – Th, Sa Th (sc), Sa (sc)18 RNA polymerase alpha subunit BP3642 36,136.08 5.6 – NELLKTPNLGRKSLNE C – Th, Sa Th (sc), Sa (sc)19 Elongation factor Tu BP3611 42,889.12 5.34 – PGSINPHTDFTAEVYI C – Th, Sa Th (sc), Sa (sc)20 Ketol-acid reductoisomerase BP0791 36,211.39 5.76 – QGHAHALNLHDSGVKV ND – Th, Sa Th (sc), Sa (sc)21 Putative peptidyl–prolyl cis-trans isomerase BPP3352 28,946.21 8.93 – PITQKSLDEFVKLVVS OM + Th, Sa Th (sc), Sa (sc)22 Aspartate-semialdehyde dehydrogenase BPP1945 40,340.78 5.08 2 (C) AGTQWAKVVPNTKEDT ND – Th, Sa Th (sc), Sa (sc)23 Putative DNA-binding protein BB2935 18,512.33 6.19 – MFMAQYTEEWNALDSI ND – Th, Sa Th (sc), Sa (sc)24 Heat schock protein 10 BPP0869 10,265.62 5.39 – AVGPGKKTEDGKILPV C – Th, Sa Th (sc), Sa (sc)25 Putative outer membrane protein BB2614 20,920.96 9.69 – RKRREFQEDFNRRRNE OM + Sa Sa (sc)
a Protein spot corresponding to position on gel (Fig. 1).b C, charge modification; M, mass modification; C–M, both charge and mass modification.c Predicted location of proteins by PSORTb version 2.0.4. C: cytoplasm, OM: outer membrane, P: periplasm and ND: not determined.d Strains used in this study. Th: Tohama I and Sa: Saadet.e Th (sc) and Sa (sc) were obtained from mice subcutaneously immunized with inactivated whole cells of Tohama and Saadet, respectively, Th (ip) and Sa (ip) were obtained from mice intraperitoneally challenged with live
whole cells of Tohama and Saadet, respectively.

5 ccine 2
ohpp
adkatcmtanwt
dStHhissftlBCakciatobasRiotaT[nrw
ihlitsatpS[pQ
vahpAwsaqpppwiPctBWhl
�astHuspob
pytacsoiAwmtftaSaTser
hpRws
46 E. Altındis et al. / Va
f the total cytoplasmic protein of B. pertussis is a target for theost immune response [25]. Some of these proteins are potentialrotective antigens that were used as the components of acellularertussis vaccines.
Our immunoproteomics approach led to the identification oftotal of 25 immunogenic proteins of B. pertussis which can be
ivided into three main groups. The first group comprised 3 well-nown antigens of B. pertussis; pertactin, serum resistance proteinnd Hsp60. Two very well-known antigenic proteins, FHA and PT ofhis group could not be detected in the present work. Mostly extra-ellular location and highly hydrophobic nature of these proteinsight have caused their elimination in the first steps of extrac-
ion procedure. The second group comprised 10 proteins that havelready been shown antigenic in certain pathogenic bacteria, butot in B. pertussis before and the third group comprised 12 proteinshich have not been shown to be immunogenic in any pathogen
ill the present study.The immunogenic proteins of the second group included ATP-
ependent protease, 30S ribosomal protein S1, RNAP � subunit,-adenosylmethionine synthetase, EF-Tu, glutamyl t-RNA amido-ransferase subunit A, ketol acid reductoisomerase, serine protease,sp10 and Hsp70. It is well known that heat shock proteinselp pathogens to penetrate host defenses and cause disease. The
mmunogenic activity of Hsp10 [26–31] and Hsp70 [10,32–34] washown in various pathogens. ATP-dependent proteases of the heathock group such as Lon and Clp are involved in turnover of mis-olded proteins, degradation of regulatory proteins and contributeo stress tolerance and virulence, as recently shown for Campy-obacter jejuni [35], Staphylococcus epidermidis [36], S. aureus [37],rachyspira pilosicoli [38] and two pathogenic Leptospira spp. [39].old inducible factors are also heat shock proteins and usually haverelation with virulence. A recent proteomics study showed that
etol acid reductoisomerase identified in the present study is aold inducible factor of B. bronchiseptica [40]. The immunogenic-ty of this protein was shown earlier in Candida albicans [41]. Inddition to their key roles in regulation of virulence by controllinghe levels of signal transduction pathways in cells, serine proteasesf pathogens facilitate the bacterial colonization of mucous mem-ranes, confer resistance to lysosomal proteolysis by phagocytesnd help dissemination in the course of infections [42]. A putativeerine protease identified in our study might play such functions.ibosomal protein S1 and RNA polymerase � subunit also detected
n this work were found to be immunogenic in total proteomesf Chlamydia trachomatis elementary bodies [43]. The latter pro-ein as well as S-adenosylmethionine synthatase were identifieds immunoreactive in H. pylori [44,45]. The immunogenicity of EF-u was shown for S. epidermidis [46], H. pylori [45], Leptospira spp.39] and Chlamydia pneumoniae [13]. Immunogenicity of cell wallon-lectin protein glutamyl-tRNA amidotransferase subunit A waseported for Streptococcus pneumoniae and its potential as a vaccineas discussed [47].
The proteins of the third group were those identified asmmunogenic for the first time in this study. Some of these proteinsave known to be related with pathogenesis in various bacteria, like
ysyl tRNA synthetase shown to be important for Salmonella enter-ca infection [48], preprotein translocase secA subunit engaged inhe export of main virulence factors of B. pertussis [49], aspartate-�-emialdehyde dehydrogenase required for survival in macrophagesnd protozoa in Legionella pneumophila and suggested as a targeto design attenuated L. pneumophila vaccines [50] and carbamoyl-
hosphate synthetase shown to be critical for the virulence ofalmonella typhimurium [51] as well as avian pathogenic E. coli52]. Interestingly, we detected the latter protein in the immuno-roteome of B. pertussis Saadet, but not in that of Tahoma I strain.uinolone signaling in cell-to-cell communication system regulatePooeu
7 (2009) 542–548
irulence in two gram-negative human pathogens, Pseudomonaseruginosa [53] and Burkholderia spp. [54]. The pqsABCDE operonas been identified necessary for Pseudomonas quorum signal (PQS)roduction, the pqsA gene encoding a putative substrate-CoA ligase.s demonstrated very recently, the PQS is a 3 hydroxy-4-quinoloneith a 2-alkyl substitution which is synthesized by the conden-
ation of anthranilic acid with a 3-keto-fatty acid by the enzymenthranilate-CoA ligase [55]. Although there are no reports onuinolone signaling system in other gram-negative pathogens, autative substrate-CoA ligase detected in immunoproteome of B.ertussis might be indicative of the presence of the same system in B.ertussis. The periplasm of gram-negative bacteria is well equippedith ATP-independent chaperones and folding catalysts, includ-
ng peptidyl–prolyl isomerases (PPIases). A newly identified PPIasear27 (the corresponding gene BP3561) displayed both PPIase andhaperone activities in vitro in B. pertussis and was shown to pro-ect FHA from proteolytic degradation [56]. PPIase B, encoded byP1906, was found to be an iron-responsive OMP in B. pertussis [16].e found another PPIase protein which was immunogenic and gave
ighest homology to BPP3352, with a predicted outer membraneocation.
Of above-mentioned proteins, only Hsp 60, RNA polymerase-subunit and putative peptidyl–prolyl cis-trans isomerase weremong a total of 19 differentially expressed proteins in a recenttudy of B. pertussis Tohama I total proteome during iron starva-ion [16]. In the present study Hsp 60, pertactin, serine protease,sp 70 and aspartate-semialdehyde dehydrogenase were found tondergo charge and/or mass modification. Hsp 60, pertactin anderine protease were also found in multiple spots in the surfaceroteome of some strains of B. pertussis [17] and three isoformsf Hsp 70 were also detected in the tick-borne spirochete Borreliaurgdorferi [57].
B. pertussis Saadet, isolated in 1948 in Turkey, had been therimary strain to manufacture cellular pertussis vaccine for manyears in the country. In this study, the identified immunogenic pro-eins were common to both Tohama I and Saadet strains, except forputative chromosome partition protein and preprotein translo-
ase secA subunit found only in Tohama I and carbamoyl-phosphateynthase large chain and a putative outer membrane protein foundnly in Saadet. The latter protein was of particular interest withts location, very strong reactivity and specificity to Saadet strain.s expected, the antisera obtained from mice immunized twiceith inactivated whole cells of Tahoma I or Saadet strains detecteduch higher number of immunogenic proteins as compared to
he antisera obtained by challenging which could identify only aew proteins, namely serum resistance protein, 30S ribosomal pro-ein S1, pertactin, Hsp60 and Hsp70. It is to be noted that only 4nd 5 out of 10 mice could survive following the challenge withaadet and Tohama I, respectively, and the survivors demonstratedpparent weight loss and signs of illness after bacterial injection.herefore, the proteins identified by the antisera collected fromuch mice might be considered among the most important antigensxpressed during in vivo infection and triggered primary humoralesponse
In spite of the widespread availability of effective vaccines andigh levels of vaccination coverage, a significant resurgence inertussis has been observed during the past 2 decades [58,59].emaining non-immunized children and older individuals withaning immunity may serve as reservoirs for the infection, occa-
ionally transmitting B. pertussis to unimmunized young infants.
ertussis which has long been considered to be only a diseasef childhood is now being reported in adolescents and adults. Itften goes underrecognized in such individuals because their dis-ase manifestations are frequently atypical. Contraindication to these of whole cell pertussis vaccines in individuals over 7 years of
ccine 2
akpaaeotav
2fsicpgfc
A
nh
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
E. Altındis et al. / Va
ge as well as the short-lived duration of protection afforded is wellnown. The licensed adolescent and adult formulations of acellularertussis vaccine combined with an adult formulation of diphtheriand tetanus toxoids (Tdap) are recorded to be safe, immunogenic,nd well tolerated [60,61]. On the other hand, in view of the mod-rate changes that have been observed in the genomic sequencesf the bacterial PRN and PT over time, there are concerns abouthe gradual loss of the efficacy of the current pertussis vacciness a result of antigenic drift and continuous selection of the leastaccine-sensitive clones [62].
In this study, there were many highly abundant proteins in the-DE gels which did not found as immunoreactive. It is there-ore rational to suggest that the immunogenic ones may be theignificant vaccine candidates. The protective capacities of themmunogenic proteins identified either by alone or in differentombinations remain to be determined in further studies. The dataresented in this work are expected to aid in development of newereneration vaccines against B. pertussis. Our current studies haveocused on both gel-based and gel-free sub-proteomic analysesoupled to Western blotting.
cknowledgements
This research was granted in part by TUBITAK-TBAG (projecto.107T444). We would like to acknowledge Dr. Dirk Albrecht foris helps in MALDI-TOF/MS analysis.
eferences
[1] Locht C, Antoine R, Jacob-Dubuisson F. Bordetella pertussis, molecular patho-genesis under multiple aspects. Curr Opin Microbiol 2001;4:82–9.
[2] Crowcroft NS, Britto J. Whooping cough-a continuing problem. BMJ 2002;324:1537–8.
[3] Khan AS, Mujer CV, Alefantis TG, Connolly JP, Mayr UB, Walcher P, et al.Proteomics and bioinformatics strategies to design countermeasures againstinfectious treat agents. J Chem Inf Model 2006;46(1):111–5.
[4] Serruto D, Rappuoli R. Post-genomic vaccine development. FEBS Lett2006;580:2985–92.
[5] Nilsson CL, Larsson T, Gustafsson E, Karlsson K, Davidson P. Identification ofprotein vaccine candidates from Helicobacter pylori using a preparative two-dimensional electrophoretic procedure and mass spectrometry. Anal Chem2000;72:2148–53.
[6] Backert S, Kwok T, Schmid M, Selbach M, Moese S, Peek R, et al. Subpro-teomes of soluble and structure-bound Helicobacter pylori proteins analyzedby two dimensional gel electrophoresis and mass spectrometry. Proteomics2005;5:1331–45.
[7] Vytvytska O, Nagy E, Bluggel M, Meyer HE, Kurzbauer R, Huber LA, et al. Iden-tification of vaccine candidate antigens of Staphylococcus aureus by serologicalproteome analysis. Proteomics 2002;2:580–90.
[8] Ariel N, Zvi A, Makarava KS, Chitlaru T, Elhanany E, Velan B, et al. Genome-basedbioinformatic selection of chromosomal Bacillus anthracis putative vaccine can-didates coupled with proteomic identification of surface-associated antigens.Infect Immun 2003;71:4563–79.
[9] Peng X, Ye X, Wang S. Identification of novel immunogenic proteins of Shigellaflexneri 2a by proteomic methodologies. Vaccine 2004;22(21–22):2750–6.
10] Havlasová J, Hernychová L, Halada P, Pellantová V, Krejsek J, Stulík J, et al. Map-ping of immunoreactive antigens of Francisella tularensis live vaccine strain.Proteomics 2002;2(7):857–67.
11] Hansmeier N, Chao T, Kalinowski J, Pühler A, Tauch A. Mapping and compre-hensive analysis of the extracellular and cell surface proteome of the humanpathogen Corynebacterium diphtheriae. Proteomics 2006;6:2465–76.
12] Rodríguez-Ortega MJ, Norais N, Bensi G, Liberatori S, Capo S, Mora M, et al.Characterization and identification of vaccine candidate proteins through anal-ysis of the group A Streptococcus surface proteome. Nat Biotechnol 2006;24(2):191–7.
13] Bunk S, Susnea I, Rupp J, Summersgill JT, Maass M, Stegmann W, et al.Immunoproteomic identification and serological responses to novel Chlamy-dia pneumoniae antigens that are associated with persistent C. pneumoniaeinfections. J Immunol 2008;180(8):5490–8.
14] LinksHsu CA, Lin WR, Li JC, Liu YL, Tseng YT, Chang CM, et al. Immuno-
proteomic identification of the hypothetical protein NMB1468 as a novellipoprotein ubiquitous in Neisseria meningitidis with vaccine potential. Pro-teomics 2008;8(10):2115–25.15] Parkhill J, Sebaihia M, Preston A, Murphy LD, Thomson N, Harris DE, et al. Com-parative analysis of the genome sequences of Bordetella pertussis, Bordetellaparapertussis and Bordetella bronchiseptica. Nat Genet 2003;35:32–40.
[
[
7 (2009) 542–548 547
16] Vidakovics ML, Paba J, Lamberti Y, Ricart CA, Sousa MV, Rodriguez ME. Pro-filing the Bordetella pertussis Proteome during Iron Starvation. J Proteome Res2007;6(7):2518–28.
17] Bottero D, Gaillard ME, Fingermann M, Weltman G, Fernández J, Sisti F, et al.Pulsed-field gel electrophoresis, pertactin, pertussis toxin S1 subunit polymor-phisms, and surfaceome analysis of vaccine and clinical Bordetella pertussisstrains. Clin Vaccine Immunol 2007;14(11):1490–8.
18] Sato Y, Arai H. Leucocytosis promoting factor of Bordetella pertussis. InfectImmun 1972;6:899–904.
19] Ramagli LS, Rodrigez LV. Quantitation of microgram amounts of protein in two-dimensional polyacrylamide gel electrophoresis sample buffer. Electrophoresis1985;6:559–63.
20] Gorg A, Weiss W, Dunn MJ. Current two-dimensional electrophoresis technol-ogy for proteomics. Proteomics 2004;4:3665–85.
21] Neuhoff V, Arold N, Taube D, Ehrhardt W. Improved staining of proteins inpolyacrylamide gels including isoelectric focusing gels with clear backgroundat nanogram sensitivity using Coomassie Brilliant Blue G-250 and R-250. Elec-trophoresis 1988;9:255–62.
22] Saha S, Raghava GP. Prediction of continuous B-cell epitopes in an antigen usingrecurrent neural network. Proteins 2006;65(1):40–8.
23] Gardy JL, Laird MR, Chen F, Rey S, Walsh CJ, Ester M, et al. PSORTb v.2. 0:expanded prediction of bacterial protein subcellular localization and insightsgained from comparative proteome analysis. Bioinformatics 2005;21(5):617–23.
24] Mattoo S, Cherry JD. Molecular pathogenesis, epidemiology, and clinical man-ifestations of respiratory infections due to Bordetella pertussis and otherBordetella subspecies. Clin Microbiol Rev 2005;18(2):326–82.
25] Del Giudice G, Gervaix A, Costantino P, Wyler CA, Tougne C, de Graeff-MeederER, et al. Priming to heat shock proteins in infants vaccinated against pertussis.J Immunol 1993;150:2025–32.
26] Barnes PN, Mehra V, Hirschfield CR, et al. Characterisation of T cell antigensassociated with the cell wall protein-peptidoglycan complex of Mycobucteriumtuberculosis. J Immunol 1989;143:2656–62.
27] Ragno S, Winrow VR, Mascagni P, Lucietto P, Di Pierro F, Morris CJ, et al. Asynthetic 10-kD heat shock protein (hsp10) from Mycobacterium tuberculosismodulates adjuvant arthritis. Clin Exp Immunol 1996;103(3):384–90.
28] Mehra V, Bloom BR, Bajardi AC, Grisso CL, Sieling PA, Alland D, et al. A majorT cell antigen of Mycobacterium leprae is a 10-kD heat-shock cognate protein. JExp Med 1992;175(1):275–84.
29] Chua-Intra B, Peerapakorn S, Davey N, Jurcevic S, Busson M, Vordermeier HM, etal. T-cell recognition of mycobacterial GroES peptides in Thai leprosy patientsand contacts. Infect Immun 1998;66(10):4903–9.
30] Pérez-Pérez GI, Thiberge JM, Labigne A, Blaser MJ. Relationship of immuneresponse to heat-shock protein A and characteristics of Helicobacter pylori-infected patients. J Infect Dis 1996;174(5):1046–50.
31] Wu YL, Lee LH, Rollins DM, Ching WM. Heat shock- and alkaline pH-inducedproteins of Campylobacter jejuni: characterization and immunological proper-ties. Infect Immun 1994;62(10):4256–60.
32] Adams E, Britton WJ, Morgan A, Goodsall AL, Basten A. Identification of humanT cell epitopes in the Mycobacterium leprae heat shock protein 70-kD antigen.Clin Exp Immunol 1993;94(3):500–6.
33] Kakeya H, Udono H, Maesaki S, Sasaki E, Kawamura S, Hossain MA, et al.Heat shock protein 70 (hsp70) as a major target of the antibody responsein patients with pulmonary cryptococcosis. Clin Exp Immunol 1999;115(3):485–90.
34] Zysk G, Bongaerts RJ, ten Thoren E, Bethe G, Hakenbeck R, Heinz HP. Detectionof 23 immunogenic pneumococcal proteins using convalescent-phase serum.Infect Immun 2000;68(June (6)):3740–3.
35] Cohn MT, Ingmer H, Mulholland F, Jørgensen K, Wells JM, Brøndsted L. Contri-bution of conserved ATP-dependent proteases of Campylobacter jejuni to stresstolerance and virulence. Appl Environ Microbiol 2007;73(24):7803–13.
36] Wang C, Li M, Dong D, Wang J, Ren J, Otto M, et al. Role of ClpP inbiofilm formation and virulence of Staphylococcus epidermidis. Microbes Infect2007;9(11):1376–83.
37] Michel A, Agerer F, Hauck CR, Herrmann M, Ullrich J, Hacker J, et al. Global reg-ulatory impact of ClpP protease of Staphylococcus aureus on regulons involvedin virulence, oxidative stress response, autolysis, and DNA repair. J Bacteriol2006;188(16):5783–96.
38] Movahedi A, Hampson DJ. Distribution of the clpX gene in Brachyspira speciesand reactivity of recombinant Brachyspira pilosicoli ClpX with sera from miceand humans. J Med Microbiol 2007;56(Pt 7):930–6.
39] Sakolvaree Y, Maneewatch S, Jiemsup S, Klaysing B, Tongtawe P, Srimanote P,et al. Proteome and immunome of pathogenic Leptospira spp. revealed by 2DEand 2DE-immunoblotting with immune serum. Asian Pac J Allergy Immunol2007;25(1):53–73.
40] Stübs D, Fuchs TM, Schneider B, Bosserhoff A, Gross R. Identification andregulation of cold-inducible factors of Bordetella bronchiseptica. Microbiology2005;151:1895–909.
41] Fernández-Arenas E, Gloria M, César N, Rosalía DO, Concha G. Low virulent
strains of Candida albicans: unravelling the antigens for a future vaccine. Pro-teomics 2004;4:3007–20.42] Ehrmann M, Clausen T. Proteolysis as a regulatory mechanism. Annu Rev Genet2004;38:709–24.
43] Sanchez-Campillo M, Bini L, Comanducci M, Raggiaschi R, Marzocchi B, PalliniV, et al. Identification of immunoreactive proteins of Chlamydia trachomatis by

5 ccine 2
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
48 E. Altındis et al. / Va
Western blot analysis of a two-dimensional electrophoresis map with patientsera. Electrophoresis 1999;20(11):2269–79.
44] Bumann D, Holland P, Siejak F, Koesling J, Sabarth N, Lamer S, et al. A compar-ison of murine and human immunoproteomes of Helicobacter pylori validatesthe preclinical murine infection model for antigen screening. Infect Immun2002;70(11):6494–8.
45] Lin YF, Wu M, Lin JT, Sun Y, Chen D, Chow L. Comparative immunoproteomics ofidentification and characterization of virulence factors from Helicobacter pylorirelated to gastric cancer. Mol Cell Proteomics 2006;5:1484–96.
46] Sellman BR, Howell AP, Kelly-Boyd C, Baker SM. Identification of immuno-genic and serum binding proteins of Staphylococcus epidermidis. Infect Immun2005;73:6591–600.
47] Portnoi M, Ling E, Feldman G, Dagan R, Mizrachi-Nebenzahl Y. The vaccinepotential of Streptococcus pneumoniae surface lectin- and non-lectin proteins.Vaccine 2006;24(11):1868–73.
48] Kaniga K, Compton MS, Curtiss 3rd R, Sundaram P. Molecular and func-tional characterization of Salmonella enterica serovar Typhimurium poxA gene:effect on attenuation of virulence and protection. Infect Immun 1998;66(12):5599–606.
49] Rambow-Larsen AA, Weiss AA. Temporal expression of pertussis toxin and Ptlsecretion proteins by Bordetella pertussis. J Bacteriol 2004;186(1):43–50.
50] Harb OS, Abu Kwaik Y. Identification of the aspartate-beta-semialdehyde dehy-drogenase gene of Legionella pneumophila and characterization of a null mutant.
Infect Immun 1998;66(5):1898–903.51] Mahan MJ, Slauch JM, Mekalanos JJ. Selection of bacterial virulence genes thatare specifically induced in host tissues. Science 1993;259:686–768.
52] Kwaga JK, Allan BJ, van der Hurk JV, Seida H, Potter AA. A carAB mutant of avianpathogenic Escherichia coli serogroup O2 is attenuated and effective as a live oralvaccine against colibacillosis in turkeys. Infect Immun 1994;62(9):3766–72.
[
[
7 (2009) 542–548
53] Pesci EC, Milbank JB, Pearson JP, McKnight S, Kende AS, Greenberg EP, et al.Quinolone signaling in the cell-to-cell communication system of Pseudomonasaeruginosa. Proc Natl Acad Sci U S A 1999;96(20):11229–34.
54] Diggle SP, Lumjiaktase P, Dipilato F, Winzer K, Kunakorn M, Barrett DA, et al.Functional genetic analysis reveals a 2-alkyl-4-quinolone signaling system inthe human pathogen Burkholderia pseudomallei and related bacteria. Chem Biol2006;13(7):701–10.
55] Coleman JP, Hudson LL, McKnight SL, Farrow 3rd JM, Calfee MW, Lindsey CA,et al. Pseudomonas aeruginosa PqsA is an anthranilate-coenzyme A ligase. JBacteriol 2008;190(4):1247–55.
56] Hodak H, Wohlkönig A, Smet-Nocca C, Drobecq H, Wieruszeski JM, SénéchalM, et al. The peptidyl-prolyl isomerase and chaperone Par27 of Bordetella per-tussis as the prototype for a new group of parvulins. J Mol Biol 2008;376(2):414–26.
57] Cluss RG, Goel AS, Rehm HL, Schoenecker JG, Boothby JT. Coordinate synthesisand turnover of heat shock proteins in Borrelia burgdorferi: degradation of DnaKduring recovery from heat shock. Infect Immun 1996;64(5):1736–43.
58] de Carvalho AP, Pereira EM. Acellular pertussis vaccine for adolescents. J Pediatr(Rio J) 2006;82(3):15–24.
59] Leung AK, Robson WL, Davies HD. Pertussis in adolescents. Adv Ther2007;24(2):353–61.
60] Halperin SA, Smith B, Russell M, Hasselback P, Guasparini R, Skowronski D, et al.An adult formulation of a five-component acellular pertussis vaccine combined
with diphtheria and tetanus toxoids is safe and immunogenic in adolescentsand adults. Vaccine 2000;18(14):1312–9.61] Pichichero ME, Blatter MM, Kennedy WA, Hedrick J, Descamps D, FriedlandLR. Acellular pertussis vaccine booster combined with diphtheria and tetanustoxoids for adolescents. Pediatrics 2006;117(4):1084–93.
62] Pertussis vaccines-WHO position paper. Weekly Epid Record 2005;80(4):31–9.
Copyright © 2022 FDOKUMEN




















