
• Mediator-s in -typhoid fever · - Radboud Repository
171
PDF hosted at the Radboud Repository of the Radboud University Nijmegen The following full text is a publisher's version. For additional information about this publication click this link. http://hdl.handle.net/2066/146563 Please be advised that this information was generated on 2022-07-10 and may be subject to change.
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of • Mediator-s in -typhoid fever · - Radboud Repository

PDF hosted at the Radboud Repository of the Radboud University
Nijmegen
The following full text is a publisher's version.
For additional information about this publication click this link.
http://hdl.handle.net/2066/146563
Please be advised that this information was generated on 2022-07-10 and may be subject to
change.

• Mediator-s in -typhoid fever · •сΙίη¡Όari and experimental'studies
nique Ceuter ·


Mediators in typhoid fever
clinical and experimental studies

ISBN 90-9011526-9
©1998 Monique Keuter. No part of this publication may be reproduced, stored in a
retrieval system, or transmitted in any form or by any means without the prior written
permission of the author, or, where appropriate, of the publishers of the publications.

Mediators in typhoid fever clinical and experimental studies
Een wetenschappelijke proeve
op het gebied van de Medische Wetenschappen
Proefschrift
ter verkrijging van de graad van doctor aan de
Katholieke Universiteit Nijmegen,
volgens besluit van het College van Decanen in het
openbaar te verdedigen op dinsdag 26 mei 1998
des namiddags om 1.30 uur precies
door
Monique Keuter
geboren op 13 augustus 1956 te Utrecht

Promotores Prof. Dr. J.W.M, van der Meer Prof. Dr. R. Djokomoeljanto (Diponegoro University)
Co-promotores Dr. B.J. Kullberg
Dr. W.M.V. Dolmans
Manuscriptcommissie Prof. Dr. D. Ruiter
Prof. Dr. J.A.A. Hoogkamp-Korstanje
Dr. W.P.M. Eling
The studies presented in this thesis were performed at the Division of General Internal
Medicine, Department of Medicine, University Hospital Nijmegen, The Netherlands,
and at the Dr. Kariadi Hospital, Diponegoro University, Semarang, Indonesia

To my mother
In memory of my father

6

Mediators in typhoid fever Clinical and experimental studies
Chapter 1 Introduction
Chapter 2 Experimental studies on the pathogenesis of Salmonella infections
Chapter 3 Patterns of pro-inflammatory cytokines and inhibitors during
typhoid fever
Chapter 4 Phospholipase A2 is a circulating mediator in typhoid fever
Chapter 5 A semi-quantitative reverse transcriptase polymerase chain reaction
method for measurement of mRNA for TNFa and IL-Iß in whole
blood cultures: Its application in typhoid fever and eccentric
exercise
Chapter 6 Pro-inflammatory cytokine profile in Salmonella typhimurium
infection in mice and the effect of recombinant IL-la
Chapter 7 G-CSF enhances survival of CBA/J mice with Salmonella
typhimurium infection
Chapter 8 Hyperlipoproteinemia protects low-density-lipoprotein receptor
deficient mice against lethal Salmonella typhimurium infection
Chapter 9 General discussion
Chapter 10 Summary
Chapter 11 Ringkasan
Chapter 12 Samenvatting
Dankwoord en Curriculum Vitae

8

Chapter 1
Introduction

10 Chapter 1

Introduction 11
Typhoid fever Typhoid fever is a severe systemic infection usually caused by Salmonella typhi
and occasionally by Salmonella paratyphi. The disease is confined to humans and
spreading is by the fecal-oral route. Great improvements in Western countries have
been achieved with controlled sewage systems and better living conditions. Still,
typhoid fever is a cause of morbidity in many countries of the world, where water is not
properly purified [1]. The World Health Organization (WHO) have estimated that annu
ally, 16.6 million cases of typhoid fever occur, with nearly 600 000 deaths. In South
East Asia, the incidence is highest in Indonesia, with more than 1000 cases per 100 000
inhabitants [2]. Most afflicted are children and young adults, with resulting economic
loss.
Pathogenesis of typhoid fever and other Salmonella infections involves several
processes. In a natural infection, Salmonellae enter via the intestinal tract. First, the
bacteria have to survive the acidic pH of the stomach. The outcome of the infection
depends upon a number of factors including size of the inoculum, the virulence of the
strain, and the ability to mobilize defense mechanisms [1]. The virulence of the Salmo
nella will influence bacterial invasion, survival and replication mechanisms [3, 4]. On
the side of the host, cellular immunity has always been considered the most important
defense mechanism in Salmonella infection [5]. These host factors will be discussed in
Chapter 2.
Clinical manifestations of infection with S. typhi start after an incubation time from
5-30 days. Symptoms range from asymptomatic infection to death, but most often
patients suffer a severe febrile disease with abdominal pain and constipation. Cough
and diarrhea may be prominent in the second week. Complications may occur in the
third week, with gastrointestinal perforation or bleeding and cerebral abnormalities or
even coma. When untreated, typhoid fever begins to wane in the fourth week, with
relapses occurring in 10-30% of the patients.
The definite diagnosis is made by isolating 5. typhi or S. paratyphi from blood or
bone marrow, the latter being frequently positive even after the use of antibiotics [6].
The diagnosis of typhoid fever with the Widal test alone is considered prone to error
[7]. Other serological tests are currently being developed [8].
The incidence of complications has decreased after the introduction of antimicro
bial therapy [9]. In Indonesia antibiotics reduced the mortality from 18% to 4%. Resis
tance to antibiotics is becoming a serious problem globally, and has spread from the
Indian subcontinent to other countries in Asia, northern Africa and Latin America [2].
Still, chloramphenicol can be used as first line drag in many places; third generation

12 Chapter 1
cephalosporines and quinolones are important, although expensive alternatives. In Indo
nesia, resistance to chloramphenicol is not wide-spread [10].
Vaccination with either parenteral Vi polysaccharide or live oral Ty21a Salmonella
vaccines is protective in 65-70%. However, a study conducted in Indonesia with the
latter Salmonella vaccine reported only 53% protection [11]. Newer strains of attenua
ted Salmonella typhi seem promising as oral vaccines, but these are still under develop
ment [12].
Mediators of fever With few exceptions, patients with full-blown typhoid fever have a continuous
fever. The final pathway in the pathogenesis of fever is the prostaglandin E2 (PGE2)
production in the organum vasculosum of the lamina terminalis (OVLT) in the central
nervous system (CNS) [13, 14]. The manner in which this is achieved is not completely
clear. It is generally believed that the release of PGE2 is induced by circulating cyto
kines (also referred to as endogenous pyrogens) which are produced at the site of infec
tion or inflammation and reach the CNS by fenestrated capillaries. Cytokines involved
in the pathogenesis of fever are interleukin-lß (IL-Iß), IL-6, tumor necrosis factor-α
(TNFct) and interferons, which act independently as endogenous pyrogens. Experimen
tal studies have indeed shown, that potent febrile responses could be induced by injec
tion of cytokines directly in the hypothalamus or in the circulation [13]. However, it
may be questioned whether this classical concept of the origin of fever is valid in all
diseases [15].
Regarding the symptoms in typhoid fever, pyrogenic cytokines or other pyrogenic
mediators would be expected to be present in the circulation during weeks, as these
patients have a continuous fever. Since this is not the case, it is suggested that other
mechanisms are involved. This may include other, yet unidentified circulating endoge
nous pyrogens or different mechanisms, including the local release of cytokines.
We asked the question whether secretory phospholipase A2 (sPLA2) may play a
role as a circulating pyrogen. Indeed, intraventricular injection of inhibitors of sPLA2
were shown to suppress fever in rabbits [16]. Phospholipases are lipolytic enzymes
which catalyse the degradation of phospholipids. To date, three varieties of PLA2 have
been characterized: group I (pancreatic) and group II (non-pancreatic) 14kD sPLA2 and
a cytosolic (85kD) PLA2. The group Π sPLA2 is present in and is secreted by a variety
of cells, and this form has been implicated in the generalized inflammatory responses
found in several experimental models and clinical syndromes such as sepsis and adult
respiratory distress syndrome (ARDS) [17, 18]. Its release is induced by IL-1 and

Introduction 13
TNFa and the enzyme is able to mediate the production of arachidonic acid (AA) [19,
20]. If the metabolic effects of these cytokines during infection are caused by sPLA2,
this would imply a central role for sPLA2 in inflammation as an intermediate between
IL-1 and TNFa and AA [18]. Support for a role of sPLA2 as a mediator of the systemic
inflammatory response came from studies in rabbits in which sPLA2 induced a fall in
blood pressure similar to that found with endotoxin infusion, and administration of
sPLA2 inhibitor p-bromophenacylbromide protected against the hypotensive effect of
purified rabbit sPLA2 [16,21].
Collaboration with Indonesia Joint research between the Catholic University Nijmegen, the Netherlands and the
Diponegoro University, Semarang, Java Indonesia was initiated in 1989. The purpose
of this collaboration was to investigate clinical and immunological features of patients
with typhoid fever, as well as to study the pathogenesis of Salmonella infection in an
experimental animal model.
A prospective study was conducted in adult patients with culture proven typhoid
fever. The aim of this project was to determine the role of mediators i.e., cytokines
present in the circulation and of importance otherwise in both complicated and uncom
plicated typhoid fever. Clinical data were collected and blood samples for cytokines
were processed locally according to a slightly adapted whole blood method, which can
be applied in countries where sophisticated laboratory facilities are not readily available
[22]. Since the immune response to many infections is also determined by HLA type
[23], the genetic characteristics of patients were also subject of study.
Furthermore, a scoring system was developed in order to predict the occurrence of
complications in patients with typhoid fever on admission. This typhoid fever admis
sion severity score (TYFASS) was derived from the clinical data collected in a pilot
study of 112 patients and will be validated by a running study. To determine risk factors
for contracting typhoid fever, social factors and micronutrients of patients and age and
sex-matched controls were determined. Salmonella typhi strains from the patients with
typhoid fever were investigated on resistance [10] and phage typed.
Finally, a therapeutic trial was performed to optimize antimicrobial therapy in
patients with typhoid fever. From the pilot study on clinical aspects, it had become clear
that bone marrow cultures had a reasonably good yield. Especially after the use of anti
biotics, bone marrow cultures remained positive as many as seven days after the start of
chloramphenicol [6]. As this permanent presence of bacteria in the bone marrow may
be the reason for relapse and carrierstate after treatment with chloramphenicol, a rando-

14 Chapter 1
mized clinical trial was done to study the sterilization-of bone marrow after treatment
with chloramphenicol compared to quinolones.
To gain insight in the pathogenesis and host defense of Salmonella infections, we
investigated whether experimental Salmonella infections in mice could be modulated
by cytokines.
Outline of this thesis In Chapter 2, the pathogenesis of Salmonella infections is described and the role of
cytokines involved in these infections are reviewed.
Chapter 3 addresses the question which concentrations of proinflammatory cyto
kines are detectable in plasma of patients with typhoid fever. We measured circulating
cytokines (IL-lß, TNFa, TNFß (lymphotoxin-a) and IL-6), and the concentrations of
interleukin-8 and the cytokine inhibitors interleukin-1 receptor antagonist (IL-IRA) and
the soluble TNF receptors p55 and p75 (sTNF-R). Moreover, the capacity of circulating
blood cells to produce cytokines after stimulation by lipopolysaccharide (LPS) ex vivo
was determined. For this purpose, we have used the whole blood cytokine test as descri
bed by us and Nerad [24, 25].
In Chapter 4, we report investigations to answer the question whether sPLA2 is a
circulating mediator in typhoid fever and whether its presence could be related to cyto
kine responses and severity of disease. Therefore, we sequentially measured circulating
pyrogenic cytokines and their inhibitors and sPLA2. In addition, we examined the
capacity of blood cells to produce IL-lß, TNFa, IL-IRA and sPLA2, upon LPS stimu
lation in samples obtained from patients with typhoid fever at various stages of the dis
ease.
In Chapter 5, the role of messenger RNA of IL-lß and TNFa in the whole blood
procedure is described, to contribute further to the understanding of how the production
of cytokines during typhoid fever is regulated.
In order to better understand the host defense against Salmonella, we investigated
the cytokine responses in mice (Chapter 6) and compared these with those found in
patients with typhoid fever (Chapters 3, 4 and 5) In line with other studies from our
laboratory, we asked the question whether administration of recombinant IL-1 would
exert a protective effect in a murine model of 5. typhimurium infection (Chapter 6).
Neutrophils are able to ingest and kill salmonellae, and they do play a role in the
outcome of Salmonella infection in vivo (see Chapter 2). We asked the question whe
ther stimulating neutrophil production and function by administering rG-CSF before
and during a lethal S.t yphimurium infection would affect survival, outgrowth of micro-

Introduction 15
organisms, histology and cytokines (Chapter 7)
Although Salmonella is a Gram-negative rod, the role of endotoxin in the pathoge
nesis of infection caused by these organism is controversial. We approached this pro
blem by using low density lipoprotein receptor knock-out (LDLR-/-) mice, of which the
circulating lipoproteins are able to bind and neutralize bacterial LPS, thereby reducing
the induction of proinflammatory cytokines (Chapter 8).
References start on page 122

16 Chapter 1

Chapter 2
Experimental studies on the pathogenesis of Salmonella infections

18 Chapter 2

Experimental Salmonella infecüon 19
Whereas Salmonella typhi is not pathogenic to mice, experimental Salmonella
typhimurium infection in mice leads to replication of bacteria in the spleen and the liver
and induction of a disease similar to human typhoid fever. Therefore the mouse model
with S. typhimurium is used to study typhoid fever.
Genetic differences in host defense Inbred mouse strains differ in their susceptibility to S. typhimurium infection. The
most important gene involved is Ity (Immunity to typhimurium), which encodes for the
recently described natural resistance associated membrane protein 1 (Nrampl) on pro
fessional phagocytes [26-28]. The gene is located on chromosome 1. Macrophages of
Ity resistant (R) mice are better able to restrict the growth of Salmonella compared to
those of Ity susceptible (S) mice, that lack the Nramp function. Genetic control of the
innate resistance of mice to S. typhimurium (Ity) is noticeable 24 hours after the start of
infection [29]. Infections in ItyR and Ity S mice can be influenced by recombinant cyto
kines [30-33]. It is not clear whether Ity itself has a regulatory role on cytokine produc
tion [34, 35].
The role of endotoxin Salmonella is a facultative intracellular Gram-negative rod; it contains lipopoly-
saccharide (LPS) but Salmonella spp. do not induce an illness similar to Gram-negative
extracellular bacteremia such as Klebsiella or Escherichia coli do. In animal models of
salmonellosis, TNFoc was detectable in the circulation only several days after S. typhi
murium infection, whereas TNFa did rise 1 hour after LPS administration in these ani
mals [36, 37]. Free endotoxin has never been found in the circulation in S. typhi
infection and endotoxin tolerance in humans did not prevent the development of
typhoid fever [38, 39]. However, LPS in Salmonella is important, as shown by
СЗН/HeJ mice in which cytokines can not be induced. СЗН/HeJ mice are Lps-d and
lack the Lps-gene. The Lps-gene is located on chromosome 4. These mice are not able
to respond to LPS and therefore they have no cytokine production upon infection with
Salmonella. They die after a low dose of Salmonella, although they are ItyR i.e. they
possess the Nramp function. This Lps-d strain benefits from TNFa and IL-1 post
Salmonella infection [40]. LPS of the Salmonella has also been reported to be a viru
lence factor in colonization of the gut and in vaccine immunity [41].

20 Chapter 2
Phases of experimental Salmonella infection The host defense against S. typhimurium infection requires several effector cells,
such as neutrophils, monocytes/macrophages and Τ lymphocytes. These cells are acti
vated and communicate via cytokines. A systemic S. typhimurium infection in mice is
thought to proceed in phases [42,43].
The first phase of an intraperitoneal (i.p.) or intravenous (i.v.) infection takes only
a few hours, in which initial distribution of the microorganisms take place. Whether
macrophages are already activated before this infection seems important, as Salmonella
injected into mice with activated macrophages by a previous infection with S. typhimu
rium or bacillus Calmette-Guerin (BCG), led to a significant lower numbers of micro
organisms in the liver and spleen after 3 and 8 hours respectively, compared to those in
organs of normal previously uninfected mice [44,45]. The transition phase of the bacte
rium, lipids and circulating antibodies are probably also able to influence this stage.
The second phase, during the first days of infection is called the exponential
growth stage and takes place before the acquired immunity has developed. Genetic
factors of the host (such as Ity) play a role [46,47] as well as virulence and inoculum of
the microorganisms. Silica treatment to destroy macrophages leads to a deterioration of
this phase of the infection [48].
Neutrophils are crucial in this stage. Salmonellae invade hepatocytes, which are
destroyed by neutrophils [49]. Natural killer (NK) cells and Τ lymphocytes are probably
not involved in the lysis of hepatocytes. Earlier studies have shown that the liver and
spleen of mice infected with S. typhimurium contained many neutrophils and micro-
abcesses after 24 hours of infection. Hardly any monocytes were found at that point of
time [49-51]. Blocking the migration of neutrophils through administration of monoclo
nal antibodies to the leukocyte adhesion molecule ß-integrin, lethally exacerbated the
infection. 100 times more salmonellae were recovered from the livers of these mice
compared to control mice [49]. In addition, neutropenic mice developed a lethal syste
mic salmonellosis after infection with low numbers of microorganisms. In these mice,
especially the hepatic microvasculature was invaded with salmonellae [50, 52]. There
fore it appears, that neutrophils are crucial for effective host defense against Salmonella
infection. Neutrophils act early in the infection to restrict Salmonella growth to a level
that can be adequately dealt with by other host defenses later.
After 3-7 days, the outgrowth in liver and spleen during Salmonella infection

Experimental Salmonella infection 21
reaches a plateau under the influence of activated macrophages producing proinflam
matory cytokines In this third phase, Τ lymphocytes do not have an important func
tion, as it was shown that athymic mice could reach the plateau phase without difficul
ties [53,54].
There is vast evidence that macrophages play an important role in the defense
against facultative intracellular infections such as Salmonella infection [5, 55-58]
Treatment of mice with silica, leading to destruction of macrophages in experimental
Salmonella infections, led to deterioration of the infection [48] Before the discovery of
the Ity locus on chromosome 1, genetic differences (ItyS and ItyR) in susceptibility in
mice were already found to be based on the difference ш ability of macrophages and
granulocytes to kill S typhimunum [46,47].
Whether the activation of macrophages is crucial for the host defense (killing)
against salmonellae is matter of much debate Although during an infection with S.
typhimunum, macrophages became activated, as was shown by their enhanced capacity
to inhibit proliferation of Toxoplasma gondii and production of nitric oxide, these
activated macrophages were unable to kill Salmonella better in vitro [44, 59] Also
macrophages activated by BCG infection did not have enhanced intracellular killing
abilities for Salmonella [45], although BCG pretreatment in vivo increased survival
from 5. typhimunum infection [60] Macrophages might be activated not to better kill,
but to enhance killing by other cells by producing cytokines. These other cells could be
NK cells, as these cells were activated during vaccination with a Salmonella strain [61]
or granulocytes [49, 51, 52]. The latter would imply that granulocytes do not only play
a role in the early phases of Salmonella infection
Some investigators query that Salmonella is an intracellular pathogen in the tradi
tional definition of being able to multiply inside macrophages Studies with murine and
human macrophages showed that survival of S typhi or S. typhimunum within macro
phages was dependent on host specificity and virulence of the strain [62] There is also
evidence that Salmonella are readily killed by neutrophils and peritoneal macrophages
[63] Intracellular proliferation of Salmonella proved possible within hepatocytes and
other parenchymal cells [50, 51]. Other bacterial and protozoal pathogens currently
believed to be intramacrophage pathogens in fact may parasitize non-phagocytic cells in
vivo. Integrity of these cells might also be influenced by cytokines and this could be an
important defense mechanism with a role for macrophages It has been shown for
cultured HEp-2 cells, that TNFa inhibited invasion of Salmonella [64] In L mono
cytogenes infection a similar phenomenon has also been shown for interferon-gamma
(IFN-γ) [65, 66]

22 Chapter 2
Following the plateau phase, there is the clearance phase during the third week of
infection, which involves the activity of Τ lymphocytes. Athymic mice or T-cell deple
ted animals show an increase in the number of microorganisms in the liver and spleen
after the plateau phase. In this phase, CD4+ T-lymphocytes mediate bacterial clearance
and IFN-γ mRNA expression was seen in CD4+, but also in CD4- (NK) cells. CD4+
cells are required in this phase as infection with S. typhimurium dependent on aromatic
precursors (aroA strain) could not be cleared by mice that lacked these cells or the IFN-
γ-receptor [67].
Cytokines In animal models of extracellular Gram-negative septicemia, high circulating
concentrations of proinflammatory cytokines such as IL-1 and TNFa have been consi
dered the main cause of organ damage and death. In contrast, against infections with
intracellular pathogens like Leishmania spp., Listeria monocytogenes or Mycobacte
rium spp., TNFa is supposed to be needed locally (in small amounts) and adminis
tration of TNFa inhibited the outgrowth of the microorganisms, whereas administration
of antibodies to this cytokine proved to be detrimental [68-73]. Moreover, mice without
TNFa receptor I (TNFRI-knockout mice) were resistant against endotoxin-induced
shock, but succumbed to infection with L monocytogenes [74, 75].
It is generally accepted that the main role of T-cells in resistance to intracellular
pathogens, is to produce IFN-γ to increase bactericidal abilities of macrophages [58]. In
the past 10 years, a concept has been developed in dividing Τ helper cells according to
the cytokines they produce (Thl versus Th2 response) [76]. Thl cells produce IFN-γ
and TNF-ß (lymphotoxin-α) inducing cellular immunity, whereas Th2 cells produce IL-
4, IL-5, IL-10 and IL-13, which are responsible for strong antibody responses. The
differentiation between Thl/Th2 is also under control of cytokines as IL-12 induces
Thl, and IL-4 induces Th2 development. In general, for the host defense of intracellular
infections a type 1 response is required.
Indeed, Thl-cell-produced cytokines such as IL-1, TNFa and IFN-γ have been
shown to be beneficial to animals with Salmonella infections and blocking these sub
stances deteriorated the infection. In contrast, antibodies to most Th2 produced cyto
kines, such as IL-4 and IL-10 increased resistance to S. typhimurium infection. How
ever, this division in Thl and Th2 should not become dogma, as there are conflicting
reports on both sides [77]. The role of the cytokines, regarded the most important in
Salmonella infections, will be described.

Experimental Salmonella infection 23
Tumor necrosis factor-α
The family of tumor necrosis factors comprises three members: TNFa, TNF-ß
(also known as lymphotoxin-a, LT-a) and LT-ß. TNFa has a key role in inflammatory
diseases. Mononuclear phagocytes are the major source of TNFa. The 26 kDa propro-
tein is cleaved by TNFa converting enzyme (TACE) and yields the 17kDa protein [78].
The effects of TNFa are transmitted via crosslinking of the membrane bound receptor I
(TNFRI, p55) and TNFRII (p75). The extracellular parts of these receptors can be shed
and these soluble receptors can bind TNFa and limit some of its effects [79]. TNFa is
produced after a stimulus such as LPS. The sepsis syndrome that develops after
injection of LPS can be mimicked by intravenous administration of IL-1 or TNFa. The
two proinflammatory cytokines, TNFa and IL-1 can induce each other's release.
Although TNFa in large amounts can be deleterious in extracellular Gram-negative
infections, TNFa is required for host defense in intracellular infections. TNFa induces
IFN-γ production by NK-cells in the early phases of the infection [80]. In infections
with intracellular pathogens like Leishmania spp, Listeria monocytogenes and Toxo
plasma gondii or Mycobacterium spp., administration of TNFa inhibited the outgrowth
of the microorganisms, whereas injection of antibodies to this cytokine proved to be
detrimental [70-72, 81, 82]
In experimental S. typhimurium infection in mice, the role of TNFa is similar to
that in other intracellular infections [83-85]. Inhibition of TNFa during Salmonella
infection has worsened outcome [84, 86], whereas administration of TNFa resulted in
better survival [83, 87].
The exact function of TNFa in Salmonella infections however, is not clear. In-vivo
studies of S. typhimurium infection in mice showed that TNFa was involved after the
early exponential growth phase. TNFa was especially required in the plateau and clear
ance phases. On histopathology, the effect of anti-TNFa antibodies was also seen not
earlier than after 4 days. In these anti-TNFa treated mice, most microorganisms were
seen in Kupffer cells and it was shown that the recruitment of monocytes was impaired.
There was less granuloma formation with fewer mononuclear cells. Administration of
antibodies on day 7 resulted in relapse of the infection and regression of established
granulomas [86]. No apparent effect on neutrophils was shown in the latter study, while
in other experiments with anti-TNFa antibodies fewer neutrophils were seen in the liver
[88]. TNFa is also important for immunity. Anti-TNFa antibodies prior to oral chal
lenge completely abolished protection conferred by the vaccine [85]. From these studies
one can conclude that TNFa acts primarily on monocytes (granuloma formation) and

24 Chapter 2
these cells have their function in the plateau and clearance phase in Salmonella infec
tions. TNFa is essential for the activation of monocytes in an autocrine fashion [81].
Furthermore, cultured hepatocytes (HEp-2 cells) were resistant against invasion of Sal
monella after incubation with TNFa in vitro, which suggests a role for TNFa on hepa
tocytes too [64].
Interferon-γ
IFN-γ is produced by CD4+ Τ lymphocytes, CD8+ T-cells, natural killer (NK)
cells and γδ Τ lymphocytes. IFN-γ has profound effects on host resistance and affects
the function of polymorphonuclear leukocytes and macrophages. Macrophage activa
tion is necessary to kill intracellular pathogens [58]. IFN-γ was shown to be the key
lymphokine in macrophage activation in in vitro animal experiments, as well as in trials
with patients with leprosy or AIDS [89]. IFN-γ increases the respiratory burst and pro
duction of nitric oxide, thereby enhancing the capacity to kill intracellular microorga
nisms. It also induces expression of Fc receptors and surface markers such as the cyto
kine IL-2 and major histocompatibility complex (MHC) class-Π antigens [90].
IFN-γ has been shown to have a beneficial effect on infections with intracellular
pathogens such as Toxoplasma gondii, Leishmania spp. and Listeria [91]. In infections
with Listeria monocytogenes, the mRNA for IFN-γ correlated with antibacterial resis
tance and was lower in susceptible mice [35,92, 93]. Mice that lack the IFN^-receptor
displayed increased susceptibility to Listeria infections, but had normal lymphocyte
subset development and MHC antigen presentation [91].
IFN-γ is also an important mediator in Salmonella infections. Mouse strain differ
ences in cytokine production have been reported previously [94] and in mice with S.
typhimurium infection, lower IFN-γ production by isolated spleen cells stimulated with
live S. typhimurium from ItyS mice has been shown compared to ItyR mice [95 ]. Viru
lence of the bacteria could play a role next to genotype in this difference [96]. A diffe
rence in the ability of production of IFN-γ is denied by other studies, in which IFN-γ
mRNA appeared at the same time in susceptible and resistant mice during a S.typhimu-
rium infection, and kinetics of this cytokine were not different between the strains [35].
Cytokine production in vivo after S. dublin infection was not different either between
ItyS and ItyR mice [34]. In contrast with Listeria infections, mRNA of IFN-γ correlated
with numbers of Salmonella cfu in the spleen and not with enhanced killing abilities
[35,92].
Anti-IFN-γ antibodies decreased host defense to Salmonella infection in mice.
These antibodies exerted a maximal effect during the plateau-phase of the infection [54,

Experimental Salmonella infection 25
84]. During this phase, IFN-γ is presumably produced by NK cells and not by T-cells
[53, 54, 83]. In orally challenged mice, anti-IFN-γ antibodies abrogated the resistance
and caused more mortality and outgrowth of Salmonella in the tissues [97]. There are
conflicting results concerning the role of IFN-γ in the clearance phase. Anti IFN-γ
antibodies administered in the late stage of salmonellosis did not alter the elimination of
certain strains of Salmonella [98], whereas infection with S. typhimurium dependent on
aromatic precursors (aroA strain) could not be cleared by mice that lacked the IFN-γ-
receptor [67].
Pretreatment of mice with exogenous rIFN-γ before injection of LPS resulted in
higher circulating concentrations of TNFa [99]. Some of the functions of activated
macrophage by rlFN-yare also mediated by the production of endogenous TNFa [100].
This might have been the case in studies where pretreatment with exogenous rIFN-γ
had an inhibiting effect on multiplication on the first day of the in vivo infection, but
not on the growth rate of Salmonella during days 1 to 4 [54]. In vitro macrophage acti
vation by IFN-γ with decreased multiplication of Salmonella has been shown by some
[30], but denied by other investigators, albeit in shorter observation periods [101, 102].
Orally administered recombinant murine IFN-γ had a suppressive effect on the
development of a natural S. typhimurium infection [103].
Interleukin-1
IL-1 is the name for two polypeptides (IL-la and IL-Iß) that possess a wide
spectrum of inflammatory, metabolic, physiologic, hematopoietic and immunologic
properties. These forms are distinct gene products, but they bind to the same receptor
and share various biologic activities [104]. The most potent stimulus for IL-1 is LPS.
When LPS is injected in mice or human volunteers, IL-1 concentrations are found in
the circulation after 3 hours. A similar time course was found after injection of Salmo
nella LPS [36]. However, in experimental S. typhimurium infection, circulating IL-la
was not found [37], showing that Salmonella infection and LPS injection is pathophy
siological^ distinct.
Intervention studies with antibodies against IL-1 in animals with Salmonella infec
tions have not been done. Pretreatment with recombinant IL-1 ameliorates a variety of
infections. It prolongs survival in Gram-negative and Gram-positive infections in
normal and neutropenic mice and increases resistance against candidal and plasmodial
infections in mice [105-108].
Intraperitoneal recombinant IL-1, given 24 hours before a lethal S. typhimurium
infection, increased survival in ItyR mice and reduced outgrowth in the organs [32].

26 Chapter 2
Addition of rTNFa to rIL-1 had a synergistic effect in ityR mice.
The mechanisms of protection by rIL-1 are not completely known yet. In Gram-
negative Pseudomonas and Klebsiella infections, reduction of proinflammatory cyto
kine production by peritoneal macrophages of Swiss mice has been found after admi
nistration of rIL-1 and this ameliorated the lethal cytokinemia [109]. In addition, cyto
kine receptors were modulated.
Whether IL-1 is involved in the microbicidal activity of murine macrophages is a
matter of debate. Incubation with rIL-1 did not enhance the capacity of peritoneal
macrophages to kill Toxoplasma gondii and to produce nitric oxide [100]. However, in
another study LPS-stimulated peritoneal macrophages were found to produce more
nitric oxide after pretreatment of mice in vivo with rIL-1 [110]. Induction of nitric
oxide synthesis is considered to be important for the killing of S. typhimurium in mice
and granuloma formation in the livers of these mice was impaired when nitric oxide
production was inhibited [111].
InterIeukin-4
Interleukin-4 (a 18-20 kDa glycoprotein) is a multifunctional cytokine, which dis
plays both inhibitory and stimulatory properties. IL-4 is considered to be produced by
the Th2 subset of Τ cells. IL-4 is also able to promote this Th2 type of response and
synthesis of IgE and it inhibits the production of proinflammatory cytokine [112].
In systemic Salmonella infection, IL-4 is detrimental. In a recent study with S.
typhimurium infection in IL-4 knock-out mice, these mice had longer survival and less
abcesses in the liver than their IL-4 +/+ littermates [113]. Expression of the murine IL-4
gene in an attenuated strain of S. typhimurium caused more outgrowth of salmonellae
when used as a live vaccine. The killing of Salmonella by macrophages of these
vaccinated mice was impaired [114].
Interleukin-6
IL-6 is a pleiotropic cytokine, involved in the regulation of immune responses, the
acute phase response and haematopoiesis. Infusion of rIL-6 results in fever, but not in
shock. IL-6 is produced by mononuclear phagocytes and other cell types. IL-1 induces
production of IL-6, and some properties of IL-1 such as synthesis of acute phase pro
teins are mediated by IL-6. In various diseases with cytokine activation, IL-6 can be
detected in the circulation, as it has a long halflife. IL-6 induced IL-IRA and sTNFRs
and inhibited production of IL-1 and TNFct and [115] is therefore considered to be an
antiinflammatory cytokine [116].

Experimental Salmonella infection 27
Interleukin-6-knock-out mice proved highly susceptible to Listeria monocytogenes
infection. In this study, the absence of IL-6 impaired neutrophilia in the peripheral
blood [117]. Treatment with recombinant IL-6 could not elevate the resistance against
L monocytogenes infection in normal mice in contrast to rIL-1 and rTNFa [118].
In S. typhimurium infection in mice, a gradual increase of circulating concentra
tions of IL-6 has been found during the first week of infection [37]. The effect of anti
bodies to IL-6 or recombinant IL-6 has not been investigated in Salmonella infection.
Interleukin-10
IL-10 is a product of T-cells, В cells and macrophages. Like IL-4 it is produced by
the Th2 subset of CD4+ Τ cells and it suppresses Thl cells [119]. IL-10 is a potent anti
inflammatory cytokine. It was reported to be able to downregulate IFN-γ production by
NK cells and CD4+ Τ lymphocytes, presumably by its inhibition of TNFa and IL-1
production by macrophages [120]. IL-4 together with IL-10 can inhibit the delayed type
hypersensitivity responses, which are Thl properties [121].
IL-10 gene expression in spleen cells in ItyS mice was reported to be higher than in
ItyR mice during the early phase of S. typhimurium infection. This difference correlated
with outgrowth and was a consequence rather than a cause of Ity susceptibility [35]. In
S. cholerasuis infection, the administration of anti-IL-10 antibodies was benificial for
the mice and resulted in increased proinflammatory cytokine production of infected
peritoneal cells. Bacterial growth in the liver and in the peritoneal cavity was reduced in
these mice [122]. However, anti-IL-10 antibodies did not influence the outcome of 5.
typhimurium infection in ItyS mice in another study [35].
Interleukin-12
Interleukin-12 is a heterodimeric cytokine, composed of 40- and 35-kDa subunits.
IL-12 is produced by mononuclear cells, macrophages, neutrophils, B-cells and dendri
tic cells, in response to a variety of stimuli [123]. IL-12 activates NK cells and Τ lym
phocytes to produce IFN-γ. This combination of IL-12 and IFN-γ shifts the immune
response towards Thl type immunity. Down-regulation of IL-12 can be achieved by IL-
4, IL-10, IL-13, TGFß and PGE2 and possibly IL-12 p40 homodimers [124, 125]
Recombinant IL-12 is being investigated as a potential stimulant of the immune
responses to malaria, tuberculosis, leishmaniasis and possibly AIDS (and as a vaccine
additive). For instance in experimental leishmaniasis, IL-12 exerted therapeutic activity
and induced IFN^and IL-10 and suppressed IL-4 independently of IFN-γ [126, 127].
However, high doses of IL-12 induced a toxic shock like syndrome.

28 Chapter 2
Endogenous IL-12 is required for the host defense against Salmonella. After immu
nization with attenuated Salmonella spp. or killed organisms, protection correlated with
IL-12 p40 mRNA in Peyer's patches and in peritoneal macrophages [128]. In mice chal
lenged orally with S. dublin, endogenous IL-12 was found in the mesenteric lymph
nodes. The administration of antibodies to IL-12 increased the outgrowth of microorga
nisms and reduced survival times [129]. In S. typhimurium infection in ItyR mice, anti-
IL-12 antibodies were shown to impair the establishment of the plateau phase leading to
higher number of Salmonella cfu in spleen and liver at day 7 of infection [130].
The neuropeptide substance Ρ has been shown to increase secretion of IL-12 p40
and expression of IL-12 p40 and p35 mRNA in cultured murine macrophages [131].
Spantide Π, a substance Ρ antagonist reduced survival of mice infected orally with
Salmonella spp., reducing IFN-yand IL-12 p40 expression at mucosal sites [132].
After incubation with S. dublin LPS in vitro, murine macrophages released IL-
12p40, but not IL-12p70 [133]. The IL-12p40 homodimers competed with the IL-12
(p35 and p40) heterodimers for binding to the high affinity IL-12 receptor and therefore
inhibited bioactivity. These Salmonella LPS-induced homodimers of IL-12p40 might
attenuate the cytokine response in acute endotoxemia [134].
Granulocyte colony-stimulating factor
Granulocyte colony-stimulating factor (G-CSF) is a hematopoietic growth factor,
which stimulates activation and proliferation of neutrophils [135]. The effects on matu
re neutrophils include enhanced phagocytosis and respiratory burst as well as reduced
apoptosis. Administration of rG-CSF has been studied in many neutropenic, as well as
non-neutropenic animal models [136]. In a peritonitis model in non-neutropenic mice,
rG-CSF was highly effective in improving survival [137, 138].
In models of intracellular infection, experience with rG-CSF is scarce. In mycobac
terial infections there are divergent results [139]. rG-CSF was beneficial in Listeria
monocytogenes infection in mice, five days of pretreatment with high doses of human
rG-CSF reducing lethality and outgrowth of microorganisms [140,141].
Natural resistance in murine salmonellosis is necessary in the period before acqui
red immunity. Neutrophils are important in the early SalmonelL· infection [49] and
therefore it may be envisaged that administration of rG-CSF is beneficial.
Apart from its capacity to increase neutrophil function, G-CSF has antiinflamma
tory properties. rG-CSF has been shown to be able to impair the LPS-induced IFN-γ
excretion, and increase the production of the anti-inflammatory cytokine IL-10. Fur
thermore, rG-CSF has been shown to be able to influence TNFa production [142,143].

Experimental Salmonella infection 29
With intracellular pathogens, proinflammatory cytokines are required to overcome the
infection Therefore with a resulting decreased TNFa production by treatment with rG-
CSF, administration of rG-CSF could harm animals with Salmonella infection Since
we do not know, how neutrophil-activating and antiinflammatory functions of rG-CSF
are balanced, it is interesting to study the administration of rG-CSF in a mouse model
of S typhtmunum infection
References start on page 122

30 Chapter 2

Chapter 3
Patterns of pro-inflammatory cytokines and inhibitors during typhoid fever
Monique Keuter, Edi Dharmana, M. Hussein Gasem, Johanna van der Ven-Jongekrijg,
Robert Djokomoeljanto, Wil M.V. Dolmans, Piene N.M. Demacker, Robert Sauerwein,
Harald Gallati and Jos W.M. van der Meer.
J Infect Dis 1994;169:1306-1311

32 Chapter 3
Abstract Cytokines and inhibitors in plasma were measured in 44 patients with typhoid
fever. Ex-vivo production of the cytokines was analyzed in a whole blood culture sys
tem with or without lipopolysaccharide (LPS). Acute phase circulating concentrations
of cytokines (± SD) were: interleukin-lß (IL-lß) < 140 pg/mL, tumor necrosis factor α
(TNFa) 130 ± 50 pg/mL, interleukin-6 (IL-6) 96 ± 131 pg/mL and interleukin-8 (IL-8)
278 ± 293 pg/mL. Circulating inhibitors were elevated in the acute phase: Interleukin-1
receptor antagonist (IL-IRA) 2304 ± 1427 pg/mL and soluble TNF receptors 55 and 75
4973 ±2644 and 22865 ±15143 pg/mL, respectively. LPS-stimulated production of
cytokines was lower during the acute phase than during convalescence (mean values:
IL-lß: 2547 vs. 6576 pg/mL; TNFa: 2609 vs. 6338 pg/ml; IL-6: 2416 vs. 7713 pg/mL).
LPS-stimulated production of IL-IRA was higher in the acute than during the convales
cent phase (5608 vs 3977 pg/mL). Inhibited production of cytokines in the acute phase
may be due to a switch from a proinflammatory to an antiinflammatory mode.

Cytokines in typhoid fever 33
Introduction
Typhoid fever is caused by the facultative intracellular Gram-negative bacillus
Salmonella typhi and occasionally by Salmonella paratyphi. Although salmonellae con
tain lipopolysaccharide (bacterial endotoxin, LPS), the clinical picture of typhoid fever
differs from Gram-negative sepsis, and the role of endotoxin in the pathophysiology of
typhoid fever is controversial [1].
The proinflammatory cytokines interleukin-lß (IL-lß), tumor necrosis factor alpha
(cachectin, TNFa), interleukin-6 (IL-6), and interleukin-8 (IL-8) have been implicated
in the pathogenesis of sepsis caused by Gram-negative microorganisms [2-4]. When
LPS is injected intravenously into animals or human volunteers, elevated concentrations
of these cytokines can be detected and the symptoms and signs of sepsis are mimicked
[5-7]. Elevated circulating levels of TNFa have been shown to correlate with poor
prognosis in sepsis, meningococcemia and also in cerebral malaria [7-10]. In contrast,
in infections with intracellular pathogens like Leishmania spp, Listeria monocytogenes
or mycobacteria administration of TNFa inhibited the outgrowth of the microorga
nisms, whereas administration of antibodies to this cytokine proved to be detrimental
[11-16]. In experimental Salmonella typhimurium infection in mice, the role of TNFa
seems to be similar to that in other intracellular infections [17-19]. In calves with
Salmonella typhimurium sepsis, the cytokine pattern appears to differ from that seen
after intravenously administered LPS: where TNFa did rise 1 hour after LPS admini
stration, Salmonella sepsis caused a hardly detectable increase of TNFa [20].
In contrast to these animal studies, circulating cytokines (TNFa, IL-6 and IL-lß)
have been found elevated in children with typhoid fever in Chile [21]. Butler et al.
studied outcome of typhoid fever in adult patient in Nepal and found higher values of
IL-6 and soluble TNF receptor p55 related to poorer outcome [22].
From 1989 joint research on several aspects of typhoid fever was started between
Nijmegen University, the Netherlands and Semarang University, Indonesia. To obtain
more insight in the pathophysiology of typhoid fever, not only did we measure
circulating pyrogenic cytokines (IL-lß, TNFa, TNFß (lymphotoxin) and IL-6), but also
the concentrations of interleukin-8 and the cytokine inhibitors interleukin-1 receptor
antagonist (IL-1RA) and the soluble TNF receptors p55 and p75 (sTNF-R). In addition,
we have investigated the capacity of blood cells to produce IL-lß, TNFa, IL-6, and
IL-IRA ex vivo in the acute and convalescent phase of hospitalized patients with
typhoid fever.
We used the whole blood cytokine test as described by van Deuren [23] and Nerad

34 Chapter 3
et al.[24]. This assay is simple, reproducible and especially suitable for use in laborato
ries that are not particularly well-equipped for work with cytokines. In addition, the
method may be less artificial than isolating mononuclear cells over a gradient, and pro
bably is a more natural mirror of what is happening in vivo, because plasma factors and
other cells are left in situ.
Patients and methods
The study was done in Dr. Kariadi Hospital, Diponegoro University, Semarang,
Indonesia beginning in December 1990. Blood and bone-marrow cultures were done for
all adult patients (> 14 years old) hospitalized with suspected typhoid fever (defined as
patients with fever > 38.5° С and at least one of the following signs: relative brady
cardia, abdominal complaints, mental changes, signs of complicated typhoid fever, an
enlarged liver or spleen and no apparent other disease). If blood or bone marrow cul
tures were positive for Salmonella typhi or paratyphi A or patients were found to have
perforated ilea at surgical exploration, typhoid fever was considered proven. A total of
44 patients were studied. Patient characteristics are shown in Table 1. Complications of
typhoid fever were defined as: gastrointestinal bleeding; intestinal perforation; shock;
delirium, stupor or coma; pneumonia; diffuse intravascular coagulation.
Table 1. Characteristics of 44 patients with culture-proven typhoid fever
Characteristic complicated disease uncomplicated disease
Number of patients
Mean age (range), years
Males/females
Median (range) of days with
fever before admission
Leukocyte count at admission
(range)
Complications
pneumonia
delirium
perforation
bleeding
16
20(14-34)
7/9
10 (4-20)
7.3xl0 9/L(2.6-37)
16
10
2
5
1
28
24 (14-60)
12/16
8.5 (4-30)
4.4xl09/L(1.6-7.4)
0

Cytokines in typhoid fever 35
Treatment consisted of chloramphenicol (40 mg/kg/day orally) if leukocyte counts
were > 2 χ 109/L. If fever did not subside within 6 days, treatment was changed to sul
famethoxazole (800 mg) and trimethoprim (160 mg) twice daily or ampicillin (4 χ lg).
Surgical patients received ampicillin, metronidazole and gentamicin during and after
surgery. No cyclooxygenase inhibitors were given. Only 2 patients received a single
dose of 120 mg dexamethasone, but not before blood was obtained for cytokine mea
surement. Most patients were discharged 7-10 days after defervescence, which was the
definition of convalescence. No patients died.
Cytokine measurements
On admission and at convalescence blood was drawn for cytokine measurements.
Venous blood samples were aseptically collected into sterile 4 mL tubes containing
EDTA (Vacutainer, Becton Dickinson, Rutherford, NJ). Unless stated otherwise, 3
tubes were taken from each patient [23]. To each tube 250 \ÍL aprotinin (Trasylol,
Bayer, Leverkusen, Germany; final concentration 625 kallikreine inactivating units/mL)
was added through the stopper by a tuberculin needle and syringe. One tube was
centriraged directly (1250 g for 10 minutes) the platelets from the supernatant plasma
were removed by second centrifugation (15000 g, 1 min) and the plasma was collected
and stored at -20°C until assayed for cytokines. To one of the two remaining tubes 50
μ ι LPS (E. coli serotype 055:B5; Sigma, St Louis, USA; final concentration 10 μ^πτί)
were added to stimulate cytokine production. Unstimulated samples contained only
aprotinin, but no LPS. Both tubes were incubated at 37°C for 24 hours.
For 17 (random) patients 1 tube was added containing indomethacin (0.5 μg/mL
final concentration) in the acute phase. Furthermore, for 26 (random) patients we added
2 tubes and removed the plasma and replaced this with a same amount of phosphate
buffered saline.
TNFa was determined by a radioimmunoassay (RIA) as described earlier (detec
tion level 100 pg/mL) [25]. Normal values for our laboratory: circulating concentrations
and ex vivo production without LPS below detection limit, ex vivo production after 24
hours stimulation with LPS 3780 ± 950 pg/mL. IL-lß was measured by RIA according
to Lisi et al., but without chloroform extraction (detection level 140 pg/mL) [26]. Nor
mal values for our laboratory: circulating concentrations and ex vivo production without
LPS below detection limit, ex vivo production after 24 hours stimulation with LPS 6930
±3160 pg/mL. IL-6 was measured by an ELISA as described (detection level 20
pg/mL) [27]. Normal values for our laboratory: circulating concentrations and ex vivo
production without LPS below detection limit. IL-8 was measured by ELISA (Quanti-

36 Chapter 3
kine, R&D Systems, Abingdon, UK), detection limit 45 pg/mL, normal values below
detection limit. TNFß (lymphotoxin-α) was measured by ELISA (Quantikine). In our
laboratory, we did not ever measure any detectable TNFß. IL-IRA was determined by a
RIA according to Poutsiaka et al. [28] (detection level 300 pg/mL). Normal values for
our laboratory: circulating concentrations and ex vivo production without LPS below
detection limit, ex vivo production after 24 hours stimulation with LPS 5757 ± 1060
pg/mL. sTNF-R were measured by an enzyme linked inumino binding assay ELIBA
(Hoffmann-La Roche, Basel, Switzerland) (detection level 80 pg/mL for p55 and 300
pg/ml for p75). Normal values: circulating concentrations 1.50 ng/mL (p55) and 2.51
ng/mL (p75). All samples from the same patient were analyzed in the same run in dupli
cate to minimize analytical errors.
Statistics
When frequency distribution was paramedical, paired and unpaired Student's t-test
were used. If not, Wilcoxon signed-rank test or Mann-Whitney U test were used. Ρ <
.05 was considered significant.
Results
Circulating cytokines and inhibitors during the acute phase and convalescence
Concentrations of pyrogenic cytokines in the acute phase (IL-Iß, IL-6, TNFa, lym-
photoxin) are shown in figure 1. IL-Iß concentrations were below the detection limit in
both acute and convalescent phases. IL-6 concentrations ranged from undetectable (<
20 pg/mL) to 600 pg/mL (median 73). TNFa concentrations ranged from below detec
tion limit to 310 pg/mL (median 110) in the acute phase and to 300 pg/mL (median was
below detection limit) during convalescence. All lymphotoxin concentrations were
below detection limit as well during the acute as during the convalescent phase. IL-8
concentrations were detectable in the acute phase (median 145, range 47 - 998 pg/mL),
but lower during convalescence (median 46, range 46 - 180).
However, nanograms per mL of inhibitors like IL-IRA and sTNF-R (p55 and p75)
were measured in the acute phase, concentrations being significantly higher than during
convalescence: IL-IRA 2304 ± 1427 pg/mL vs 469 ± 324 and sTNF-R55 4973 ± 2644
vs 1671 ± 532 and sTNF-R75 22865 ± 15143 vs 5971 ± 2750 pg/mL (figure 2).
No differences were found in circulating cytokines or inhibitors between the 16
patients with complicated and the 28 with uncomplicated disease course.
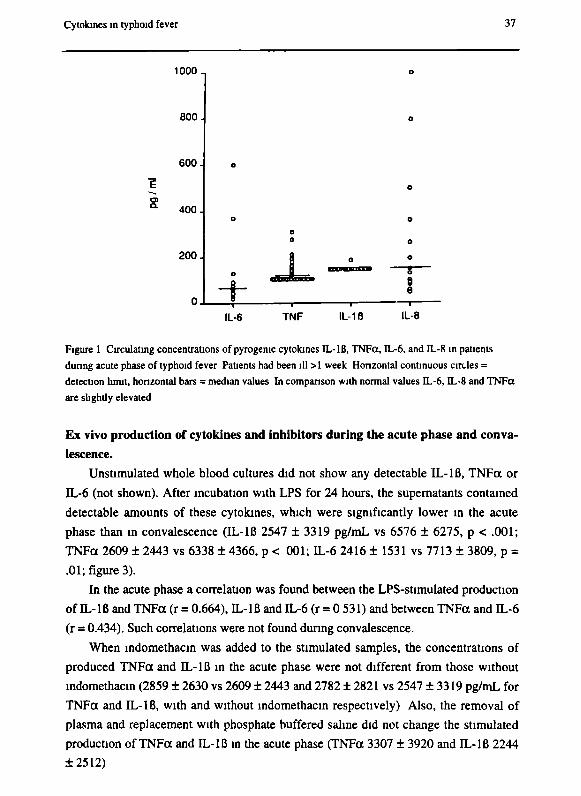
Cytokines in typhoid fever 37
1000-,
800.
600.
400.
200.
0 .
о
о
О о
» ^ J I +
о
о
о
о
*» — 8 — θ 8
IL-6 TNF IL-1 ß IL-8
Figure 1 Circulating concentrations of pyrogenic cytokines IL-IB, TNFct, IL-6, and IL-8 in patients
during acute phase of typhoid fever Patients had been ill >1 week Horizontal continuous circles =
detection limit, horizontal bars = median values In comparison with normal values IL-6, IL-8 and TNFct
are slightly elevated
Ex vivo production of cytokines and inhibitors during the acute phase and conva
lescence.
Unstimulated whole blood cultures did not show any detectable IL-Iß, TNFa or
IL-6 (not shown). After incubation with LPS for 24 hours, the supematants contained
detectable amounts of these cytokines, which were significantly lower in the acute
phase than in convalescence (IL-lß 2547 ± 3319 pg/mL vs 6576 ± 6275, ρ < .001;
TNFa 2609 ± 2443 vs 6338 ± 4366, ρ < 001; IL-6 2416 ± 1531 vs 7713 ± 3809, ρ =
.01; figure 3).
In the acute phase a correlation was found between the LPS-stimulated production
of IL-lß and TNFa (r = 0.664), IL-lß and IL-6 (r = 0 531) and between TNFa and IL-6
(r = 0.434). Such correlations were not found during convalescence.
When indomethacin was added to the stimulated samples, the concentrations of
produced TNFa and IL-lß in the acute phase were not different from those without
indomethacin (2859 ± 2630 vs 2609 ± 2443 and 2782 ± 2821 vs 2547 ±3319 pg/mL for
TNFa and IL-lß, with and without indomethacin respectively) Also, the removal of
plasma and replacement with phosphate buffered saline did not change the stimulated
production of TNFa and IL-lß in the acute phase (TNFa 3307 ± 3920 and IL-lß 2244
±2512)
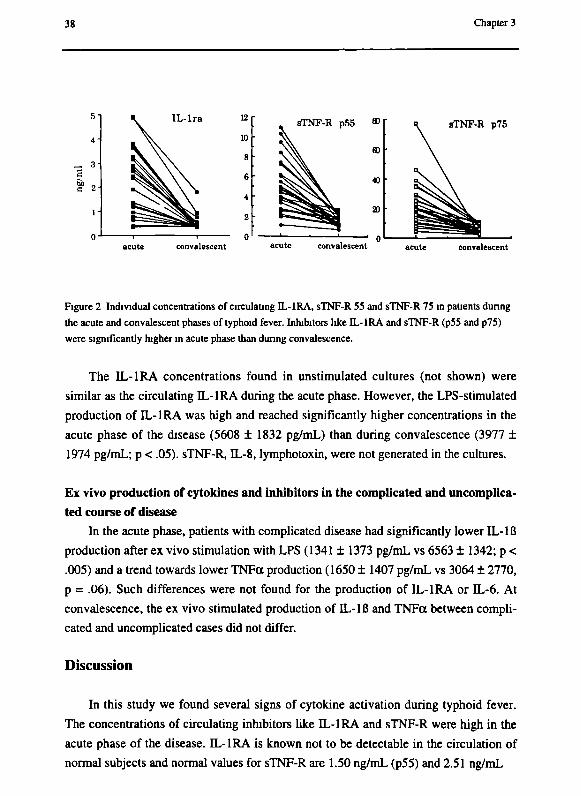
38 Chapter 3
acute convalescent acute convalescent acute convalescent
Figure 2 Individual concentrations of circulating IL-1RA, sTNF-R 55 and sTNF-R 75 in patients during
the acute and convalescent phases of typhoid fever. Inhibitors like IL-IRA and sTNF-R (p55 and p75)
were significantly higher in acute phase than during convalescence.
The IL-IRA concentrations found in unstimulated cultures (not shown) were
similar as the circulating TL-IRA during the acute phase. However, the LPS-stimulated
production of IL-IRA was high and reached significantly higher concentrations in the
acute phase of the disease (5608 ± 1832 pg/mL) than during convalescence (3977 ±
1974 pg/mL; ρ < .05). sTNF-R, IL-8, lymphotoxin, were not generated in the cultures.
Ex vivo production of cytokines and inhibitors in the complicated and uncomplica
ted course of disease
In the acute phase, patients with complicated disease had significantly lower IL-Iß
production after ex vivo stimulation with LPS (1341 ± 1373 pg/mL vs 6563 ± 1342; ρ <
.005) and a trend towards lower TNFa production (1650 ± 1407 pg/mL vs 3064 ± 2770,
ρ = .06). Such differences were not found for the production of IL-IRA or IL-6. At
convalescence, the ex vivo stimulated production of IL-Iß and TNFa between compli
cated and uncomplicated cases did not differ.
Discussion
In this study we found several signs of cytokine activation during typhoid fever.
The concentrations of circulating inhibitors like IL-IRA and sTNF-R were high in the
acute phase of the disease. IL-IRA is known not to be detectable in the circulation of
normal subjects and normal values for sTNF-R are 1.50 ng/mL (p55) and 2.51 ng/mL
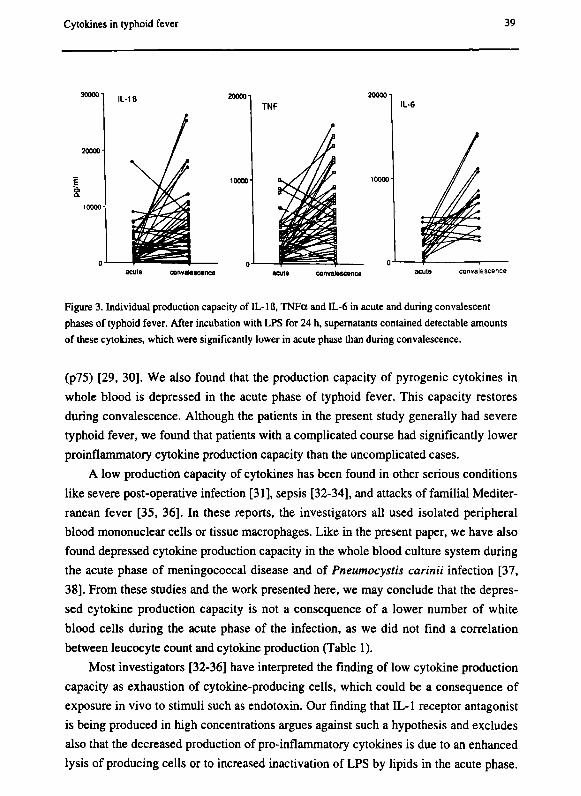
Cytokines in typhoid fever 39
convalescence
Figure 3. Individual production capacity of IL-IB, TNFa and IL-6 in acute and during convalescent
phases of typhoid fever. After incubation with LPS for 24 h, supernatants contained detectable amounts
of these cytokines, which were significantly lower in acute phase than during convalescence.
(p75) [29, 30]. We also found that the production capacity of pyrogenic cytokines in
whole blood is depressed in the acute phase of typhoid fever. This capacity restores
during convalescence. Although the patients in the present study generally had severe
typhoid fever, we found that patients with a complicated course had significantly lower
proinflammatory cytokine production capacity than the uncomplicated cases.
A low production capacity of cytokines has been found in other serious conditions
like severe post-operative infection [31], sepsis [32-34], and attacks of familial Mediter
ranean fever [35, 36]. In these reports, the investigators all used isolated peripheral
blood mononuclear cells or tissue macrophages. Like in the present paper, we have also
found depressed cytokine production capacity in the whole blood culture system during
the acute phase of meningococcal disease and of Pneumocystis carinii infection [37,
38]. From these studies and the work presented here, we may conclude that the depres
sed cytokine production capacity is not a consequence of a lower number of white
blood cells during the acute phase of the infection, as we did not find a correlation
between leucocyte count and cytokine production (Table 1).
Most investigators [32-36] have interpreted the finding of low cytokine production
capacity as exhaustion of cytokine-producing cells, which could be a consequence of
exposure in vivo to stimuli such as endotoxin. Our finding that IL-1 receptor antagonist
is being produced in high concentrations argues against such a hypothesis and excludes
also that the decreased production of pro-inflammatory cytokines is due to an enhanced
lysis of producing cells or to increased inactivation of LPS by lipids in the acute phase.

40 Chapter 3
Although we have not been able to demonstrate that the proinflammatory cytokines and
IL-IRA are being produced by the same kind of cells, it is our hypothesis that after the
initial phase of infection, cytokine-producing cells switch from a balanced proinflam
matory to an anti-inflammatory repertoire. Our findings that patients in the acute phase
of typhoid fever have high concentration of soluble TNF receptors in their blood is in
agreement with this notion.
Since the cultures were performed with whole blood, we investigated whether
some circulating common factor could be responsible for the correlated low production
capacity of the cytokines IL-Iß, TNFa and IL-6 in the acute phase. Cyclooxygenäse
products, such as PGE2, which are known to inhibit production of IL-1 and TNFa [39]
were not responsible, since addition of indomethacin to the whole blood cultures did not
lead to significant changes in cytokine production. Likewise, removal of plasma and
addition of saline before incubation did not overcome the suppression in the acute phase
of the disease.
It is possible that exposure in vivo to other inhibitory factors explains the low cyto
kine production capacity. Schindler et al [40] demonstrated that exposure of isolated
mononuclear cells to IL-6 inhibits the production of IL-1 and TNFa. In the present
study we could not find any correlation between IL-6 concentrations in plasma and the
magnitude of the production of IL-lß and TNFa (r = 0.041, г = 0.035 respectively).
Exposure to other cytokines such as IL-4, IL-10 and TGFß could, however, play a role
here. Recently, Vannier et al provided evidence that exposure of cells to IL-4 suppres
ses the IL-1 production but upregulates the synthesis of IL-Ira [41].
With few exceptions, patients with typhoid fever have a continuous fever. Hence,
pyrogenic cytokines would be expected to be present in the circulation in the acute
phase of the disease. In our series of febrile patients with typhoid fever, we were not
able to detect appreciable concentrations of the pyrogenic cytokines IL-lß, TNFa and
lymphotoxin. The concentrations of IL-6, generally considered a relatively weak pyro
gen [42], were low as compared to other febrile conditions [3, 9]. We could however
detect elevated concentrations of IL-8, but this cytokine is considered non-pyrogenic
[43]. Thus, the question which pyrogens are responsible for the continuous fever in
typhoid fever remains unanswered at the present time.
Acknowledgments
We would like to thank Dr. James Vannice (Synergen) for his supply of reagents
for IL-1RA measurements.

Cytokines in typhoid fever 41
References
1. Greisman SE. The role of endotoxin during typhoid fever and tularemia in man. J
Clin Invest 1969; 48: 613-629.
2. Cannon JG, Thompkins RG, Gelfand JA, Michie HR, Stanford GG, Van der Meer
JWM, Endres S, Lonnemann G, Corsetti J, Chemow B, Wilmore DW, Wolff SM,
Burke JF, Dinarello CA. Circulating interleukin-1 and tumor necrosis factor in
septic shock and experimental endotoxin fever. J Infect Dis 1990; 161: 79-84.
3. Hack CE, De Groot ER, Felt-Bersma RJF. Increased plasma levels of interleukin-6
in sepsis. Blood 1989; 74: 1704-1710.
4. Mitchie HR, Spriggs DR. Manogue KR. Tumor necrosis factor and endotoxin indu
ce similar metabolis responses in human beings. Surgery 1988; 104: 280-285.
5. Fischer E, Marano MA, Barber AE. Comparison between the effects of interleukin-
1 alpha administration and sublethal endotoxemia in primates. Am J Physiol 1991;
261: R442-452.
6. Okusawa S, Gelfand JA, Ikejima T, Connolly RJ, Dinarello CA. Interleukin-1 indu
ces a shock-like state in rabbits. J Clin Invest 1988; 81:1162-1172.
7. Calandra Τ, Baumgarten JD, Grau GE. Prognostic values of tumor necrosis factor/
cachectin, interleukin-1 interferon-alpha and interferon-gamma in the serum of pa
tients with septic shock. J Infect Dis 1990; 161: 982-987.
8. Damas Ρ, Reuter A, Gysen Ρ, Demonty J, Lamy M, Franchimont P. Tumor necrosis
factor and interleukin-1 serum levels during severe sepsis in humans. Crit Care
Med 1989; 17: 975-978.
9. Waage A, Brandtzaeg P, Halstensen A, Kierulf P, Espevik T. The complex pattern
of cytokines in serum from patients with meningococcal septic shock. Association
between interleukin-6, interleukin-1 and fatal outcome. J Exp Med 1989; 169: 333-
338.
10. Grau GE, Fajardo LF, Piguet PF, Allet В, Lambert PH, Vassalli P. Tumor necrosis
factor (cachectin) as an essential mediator in murine cerebral malaria. Science
1987;237: 1210-1212.
11. Barnes PF, Chatterjee D, Brennan PJ, Rea TH, Modlin RL. Tumor necrosis factor
production in patients with leprosy. Infect Immun 1992; 60: 1441-1446.
12. Havell EA. Production of tumor necrosis factor during murine listeriosis. J Immu
nol 1987; 139: 4225-4231.
13. Liew FJ, Parkinson C, Millot S, Severn A, Carrier M. Tumor necrosis factor (TNF)
in leishmaniasis 1 .TNF mediates host protection against cutaneous leishmaniasis.

42 Chapter 3
Immunology 1990; 69: 570-573.
14. Nakane A, Minagawa T, Kato K. Endogenous tumor necrosis factor (cachectin) is
essential to host resistance against Listeria monocytogenes infection. Infect Immun
1988; 56: 2563-2569.
15. Silva CL, Foss NT. Tumor necrosis factor in leprosy patients. J Infect Dis 1989;
159: 787-790.
16. Titus RG, Sherry B, Cerami A. Tumor necrosis factor plays a protective role in ex
perimental murine cutaneous leishmaniasis. J Exp Med 1989; 170: 2097-2104.
17. Nakano Y, Onozuka К, Terada Y, Shinomiya H, Nakano M. Protective effect of re
combinant tumor necrosis factor-α in murine salmonellosis. J Immunol 1990; 144:
1935-1941.
18. Nauciel C, Espinasse-Maes F. Role of gamma-interferon and tumor necrosis factor
alpha in resistance to Salmonella typhimurium infection. Infect Immun 1992; 60:
450-454.
19. Tite JP, Dougan G, Chatfield SN. The involvement of tumor necrosis factor in im
munity to Salmonella infection. J Immunol 1991; 147: 3161-3164.
20. Peel JE. Induction of circulating tumor necrosis factor cannot be demonstrated
during septicemic salmonellosis in calves. Infect Immun 1990; 58: 439-42.
21. Roine I, Herrera Ρ, Ledermann W, Peltola Η. Tumor necrosis factor-α, interleukin-
lß and interleukin-6 levels in typhoid fever. 30th Interscience Conference on Anti
microbial Agents and Chemotherapy. Atlanta, GA, 1990: Abstract 299.
22. Butler T, Ho M, Acharya G, Tiwari M, Gallati H. Interleukin-6, gamma interferon,
and tumor necrosis factor receptors in typhoid fever related to outcome of anti
microbial therapy. Antimicrob Agents Chemother 1993; 37: 2418-2421.
23. Van Deuren M, Van der Ven-Jongekrijg J, Keuter M, Demacker PNM, Van der
Meer JWM. Cytokine production in whole blood cultures. J Int Fed Clin Chem
1993;5:216-221.
24. Nerad JL, Griffiths JK, Van der Meer JWM, Endres S, Poutsiaka DD, Keusch GT,
Bennish M, Salam MA, Dinarello CA, Cannon JG. Interleukin-lß (IL-lß), IL-1 re
ceptor antagonist, and TNF-α production in whole blood. J Leukocyte Biol 1992;
52: 687-692.
25. Van der Meer JWM, Endres S, Lonnemann G, Cannon JG, Dcejima T, Okusawa S,
Gelfand JA, Dinarello CA. Concentrations of immunoreactive human tumor necro
sis factor alpha produced by human mononuclear cells in vitro. J Leucocyte Biol
1988; 43: 216-223.
26. Lisi PJ, Chu CW, Koch GA, Endres S, Lonnemann G, Dinarello CA. Development

Cytokines in typhoid fever 43
and use of radioimmunoassay for human interleukin-lß. Lymphokine Res 1987; 6:
229-244.
27. Barrera Ρ, Boerbooms AMT, Janssen EM. Circulating soluble TNF receptors and
interleukin-2 receptors, tumor necrosis factor-alpha and interleukin-6 in rheumatoid
arthritis. Longitudinal evaluation during methotrexate and azathioprine therapy.
Arthritis Rheuma 1993; 36: 1072-1079.
28. Poutsiaka DD, Clark BD, Vannier E, Dinarello CA. Production of interleukin-1 re
ceptor antagonist and interleukin-lß by peripheral blood mononuclear cells is diffe
rentially regulated. Blood 1991; 78: 1275-1281.
29. Dinarello CA. Interleukin-1 and interleukin-1 antagonism. Blood 1991; 77: 1627-
1652.
30. Shapiro L, Clark BD, Orencole SF, Poutsiaka DD, Granowitz EV, Dinarello CA.
Detection of tumor necrosis factor soluble receptor p55 in blood samples from
healthy and endotoxemic humans. J Infect Dis 1993; 167: 1344-1350.
31. Luger A, Graf H, Schwarz HP, Stummvoll HK, Luger TA. Decreased serum inter
leukin-1 activity and monocyte interleukin-1 production in patients with fatal
sepsis. Crit Care Med 1986; 14:458-461.
32. Simpson SQ, Modi H, Balk RA, Bone RC, Casey LC. Reduced alveolar macropha
ge production of tumor necrosis factor during sepsis in mice and man. Crit Care
Med 1991; 19: 1060-1066.
33. Srugo I, Berger A, Lapidot Z, Katz R, Pollak S. Interleukin-1 secretion by blood
monocytes of septic premature infants. Infection 1991; 3: 150-154.
34. Helminen M. Interleukin-1 production from peripheral blood monocytes in septic
infections in children. Scand J Infect Dis 1991; 23: 607-611.
35. Rozenbaum M, Katz R, Rozner I, Pollack S. Decreased interleukin-1 activity relea
sed from circulating monocytes of patients with familial Mediterranean fever
during in vitro stimulation by lipopolysaccharide. J Rheumatol 1992; 19: 416-418.
36. Schattner A, Lachmi M, Livneh A, Pras M, Hahn T. Tumor necrosis factor in fami
lial Mediterranean fever. Am J Med 1991; 90: 434-438.
37. Van Deuren M, Van der Ven-Jongekrijg J, Demacker PNM, Bartelink AKM, Van
Dalen R, Sauerwein RW, Gallati H, Vannice JL, Van der Meer JWM. Differential
expression of proinflammatory cytokines and their inhibitors during the course of
meningococcal infections. J Infect Dis 1994; 169: 157-161.
38. Perenboom RM, Van Schijndel ACHW, Beckers P, Sauerwein R, Van Hamersvelt
HW, Festen J, Gallati H, Van der Meer JWM. Cytokine profiles in broncho-
alveolar lavage fluid and blood in HIV-seronegative patients with Pneumocystis

44 Chapter 3
carinii pneumonia. Eur J Clin Invest 1994; 26: 159-166.
39. Endres S, Cannon JG, Dempsey RA, Sissson S, Ghorbani R, Lonnemann G,
Dinarello CA. In vitro production of IL-Iß, IL-la, TNF and IL-2 in healthy
subjects: distribution, effect of oral cyclooxygenase inhibitors and evidence of
independent gene regulation. Eur J Immunol 1989; 19: 2327-2333.
40. Schindler R, Mancilla J, Endres S. Correlations and interactions in the production
of interleukin-6 (IL-6), interleukin-1 (IL-1) and tumor necrosis factor (TNF) in
human blood mononuclear cells: IL-6 suppresses IL-1 and TNF. Blood 1990; 75:
40-47.
41. Vannier E, Miller LC, Dinarello CA. Coordinated antiinflammatory effects of inter-
leukin 4: interleukin 4 suppresses interleukin 1 production but up-regulates gene
expression and synthesis of interleukin 1 receptor antagonist. Proc Natl Acad Sci
USA 1992; 89: 4076-4080.
42. Dinarello CA, Cannon JG, Mancilla J, Bishai I, Lees J, Coceani F. Interleukin-6 as
an endogenous pyrogen: induction of prostaglandin E2 in brain but not in periphe
ral blood mononuclear cells. Brain Res 1991; 562: 199-206.
43. Van Damme J. Interleukin-8 and related molecules. In: Thomson AW, ed. The
cytokine handbook. London: Academic Press, 1991.

Chapter 4
Phospholipase A2 is a circulating mediator in typhoid fever
Monique Keuter, Edi Dhaimana, Bart-Jan Kullberg, Casper Schalkwijk, M. Hussein
Gasem, Liesbeth Seuren, Robert Djokomoeljanto, Wil M. V. Dolmans, Henk Van den
Bosch and Jos W.M. van der Meer
J Infect Dis 1995;172:305-308

46 Chapter 4
Abstract
Circulating proinflammatory mediators have not been found in studies on typhoid
fever, although the patients suffer from a systemic disease with characteristic protracted
fever. The 14-kDa group Π extracellular phospholipase A2 (PLA2) is induced by inter-
leukin-1 (IL-1) and tumor necrosis factor (TNF), and may mediate some of the effects
of these cytokines. Circulating PLA2 concentrations were measured in 12 typhoid fever
patients on various days after admission and after recovery. On admission, mean
concentrations of PLA2 were elevated (1444 ± 1560 ng/ml) and decreased gradually
and significantly to day 14 (55 ± 48 ng/ml). Patients with complicated disease had
significantly higher PLA2 levels on admission. PLA2 was not produced in a lipopoly-
saccharide-stimulated whole blood culture, indicating that PLA2 originates from other
types of cells. These data indicate that PLA2 may be a mediator of disease in protracted
inflammatory diseases such as typhoid fever.

PLA2 in typhoid fever 47
Introduction
Despite the presence of protracted fever and other generalized signs of illness in
typhoid fever, several studies have failed to demonstrate circulating proinflammatory
mediators in these patients. In a previous report, we presented evidence for cytokine
activation in terms of raised interleukin-1 receptor antagonist (IL-IRA) and soluble
tumor necrosis factor receptors (sTNF-r) in patients with typhoid fever, but no apparent
rise in circulating concentrations of the proinflammatory cytokines interleukin-1 ß (IL-
lß), tumor necrosis factor alpha (TNFa) and interleukin-6 (IL-6) [1]. This is in agree
ment with other observations in patients with typhoid fever in Nepal and Indonesia [2,
3], although one study revealed elevated cytokine levels in a minority of patients [4].
Phospholipases are lipolytic enzymes which catalyze the degradation of phospho
lipids. To date, three varieties of phospholipase A2 (PLA2) have been characterized:
group I (pancreatic) and group Π (non-pancreatic) 14kD PLA2 and a cytosolic (85kD)
PLA2. The group Π PLA2 occurs in and is secreted by a variety of cells, and has been
implicated in the generalized inflammatory responses in several experimental models
and clinical syndromes such as sepsis and adult respiratory distress syndrome (ARDS)
[5]. Its release is induced by IL-1 and TNFa and the enzyme mediates the production of
arachidonic acid [6, 7]. Some of the metabolic effects of these cytokines during infec
tion may therefore be mediated by PLA2, potentially giving this enzyme a central role
in inflammation [8].
The absence of sizable proinflammatory cytokinemia in typhoid fever made us
consider extracellular PLA2 a candidate circulating mediator of the systemic response.
Besides this we wanted to relate its presence to cytokine responses and severity of
disease. Therefore we measured circulating PLA2 and pyrogenic cytokines and their
inhibitors sequentially. In addition, we examined the capacity of peripheral blood cells
to produce PLA2, IL-Iß, TNFa, and IL-Ira ex vivo of patients with typhoid fever ad
mitted to the hospital.
Patients and Methods
The study was conducted in Dr. Kariadi Hospital, Diponegoro University, Sema-
rang, Indonesia, from December 1990 onward. Patient selection and treatment has been
described elsewhere [1]. In brief, the diagnosis of typhoid fever was confirmed by posi
tive blood or bone marrow culture in all patients, and treatment was with chlorampheni
col. Most patients were discharged 7 to 10 days after defervescence, which was the defi-

48 Chapter 4
nition of convalescence. None of the patients died.
Measurements of cytokines and other mediators
On admission and on day 2, 4 and 6 and after recovery (day 14), blood was drawn
for cytokine and PLA2 measurements. Circulating concentrations and ex vivo produc
tion of cytokines were determined as described earlier[l, 9]. Endotoxin was measured in
platelet-rich plasma by a chromogenic limulus amoebocyte lysate assay (Kabi Vitrum,
Stockholm, Sweden). TNFa was measured by RIA as described [10] (detection level,
30 pg/ml). This RIA detects both free TNFa and TNFa bound to its soluble receptors.
IL-Iß was measured by RIA according to Lisi et al but without chloroform extraction
(detection level 30 pg/ml)[ll]. IL-1RA was determined by a RIA as described by
Poutsiaka et al. [12] (detection level 300 pg/ml). sTNF-R were measured by an enzyme
linked immuno binding assay ELIBA (Hoffmann-La Roche, Basel, Switzerland)
(detection level 80 pg/ ml for p55 and 300 pg/ml for p75).
In a group of healthy controls normal values for our laboratory were as follows.
TNFa: circulating concentrations and ex vivo production without LPS 106 ± 25 pg/ml,
ex vivo production after 24 hours stimulation with LPS 3780 ± 950 pg/ml. IL-lß:
circulating concentrations and ex vivo production without LPS below detection limit, ex
vivo production after 24 hours stimulation with LPS 6930 ±3160 pg/ml. IL-IRA:
circulating concentrations and ex vivo production without LPS below detection limit, ex
vivo production after 24 hours stimulation with LPS 5757 ± 1060 pg/ml. For sTNF-R:
Normal values: circulating concentrations 1.50 ng/ml (p55) and 2.51 ng/ml (p75).
PLA2 measurements
PLA2 activity was assayed with 0.2 mM l-acyl-2[l-C14]linoleoylphosphatidyl-
ethanolamine (specific activity 1000 dpm/nmol in 0.1M Tris-HCl (pH 8.5) containing
10 mM CaC12 and 0.05% Triton X-100. After incubation for 30 min at 37°C the [C14]
linoleate released was extracted by a modified Dole extraction procedure[13]. The
radioactive substrate was prepared biosynthetically as described previously [13].
Immunoreactive group II PLA2 was determined with an ELISA modified from
Smith et al. [14]. Two different monoclonal antibodies against human group Π PLA2
(kindly provided by Dr FB Taylor jr., Oklahoma Medical Research Foundation, Okla
homa City, OK) were used as coating and catching antibodies. Results were compared
with those obtained with culture medium from HepG2 cells stimulated with human IL-
6. The amount of group II PLA2 in this culture medium was assessed by comparison
with purified recombinant human group II PLA2. The lower limit of detection was 1

PLA2 in typhoid fever 49
ng/ml and normal plasma values in 19 healthy volunteers amounted to 20 ± 7 ng/ml
(range 9-30 ng/ml).
Statistics
All samples from the same patient were analyzed in the same run in duplicate to
minimize analytical errors. Values are expressed as mean ± SD unless otherwise indi
cated. When frequency distribution was parametrical, paired and unpaired Student's t-
test were used. If not, Wilcoxon signed-rank test or Mann-Whitney U test were used.
Analysis of variance was done when considered suitable. Ρ < .05 was considered signi
ficant.
Results
Patients
Of 12 patients studied, 4 had a complicated course of disease (3 pneumonia and 1
delirium). The 9 female and 3 male (3:1) patients had a mean age of 16 years (range 14-
53). There was no difference in white blood cell count, female/male ratio or hemoglobin
between patients with a complicated or uncomplicated disease course. No circulating
endotoxin was found in any of the samples.
Circulating PLA2
On admission, mean concentrations of immunoreactive PLA2 were elevated (1444
± 1560 ng/ml) and decreased gradually and significantly to day 14 (55 ± 48 ng/ml);
figure 1). Patients with complicated disease had significantly higher mean PLA2 values
on admission (2520 ± 1284 ng/ml) than patients with uncomplicated disease (235 ± 206
ng/ml; p=.001). Immunoreactive PLA2 concentrations correlated well with PLA2 found
in the bioassay (r = 0.894; figure 1).
Ex vivo production capacity of PLA2
Unstimulated whole blood cultures (data not shown) showed PLA2 concentrations
virtually equal to circulating concentrations. After incubation with LPS for 24 hours,
the concentrations of enzyme in the supernatants did not change, indicating that no
additional PLA2 had been produced by peripheral blood cells. Cells incubated with LPS
after removal of plasma did not produce PLA2 either (data not shown).
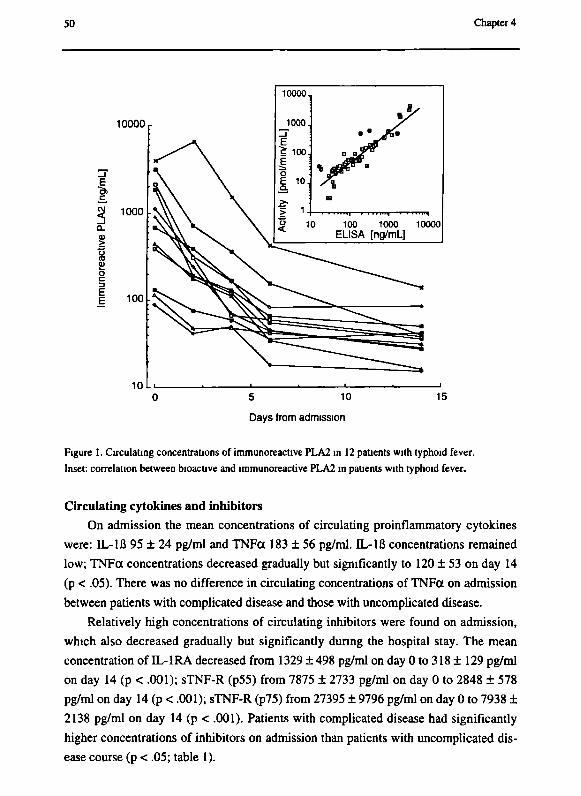
50 Chapter 4
Days from admission
Figure 1. Circulating concentrations of immunoreactive PLA2 in 12 patients with typhoid fever.
Inset: correlation between bioactive and immunoreactive PLA2 in patients with typhoid fever.
Circulating cytokines and inhibitors On admission the mean concentrations of circulating proinflammatory cytokines
were: IL-Iß 95 ± 24 pg/ml and TNFa 183 ± 56 pg/ml. IL-Iß concentrations remained
low; TNFa concentrations decreased gradually but significantly to 120 ± 53 on day 14
(p < .05). There was no difference in circulating concentrations of TNFa on admission
between patients with complicated disease and those with uncomplicated disease.
Relatively high concentrations of circulating inhibitors were found on admission,
which also decreased gradually but significantly during the hospital stay. The mean
concentration of IL-IRA decreased from 1329 ± 498 pg/ml on day 0 to 318 ± 129 pg/ml
on day 14 (p < .001); sTNF-R (p55) from 7875 ± 2733 pg/ml on day 0 to 2848 ± 578
pg/ml on day 14 (p < .001); sTNF-R (p75) from 27395 ± 9796 pg/ml on day 0 to 7938 ±
2138 pg/ml on day 14 (p < .001). Patients with complicated disease had significantly
higher concentrations of inhibitors on admission than patients with uncomplicated dis
ease course (p < .05; table 1).
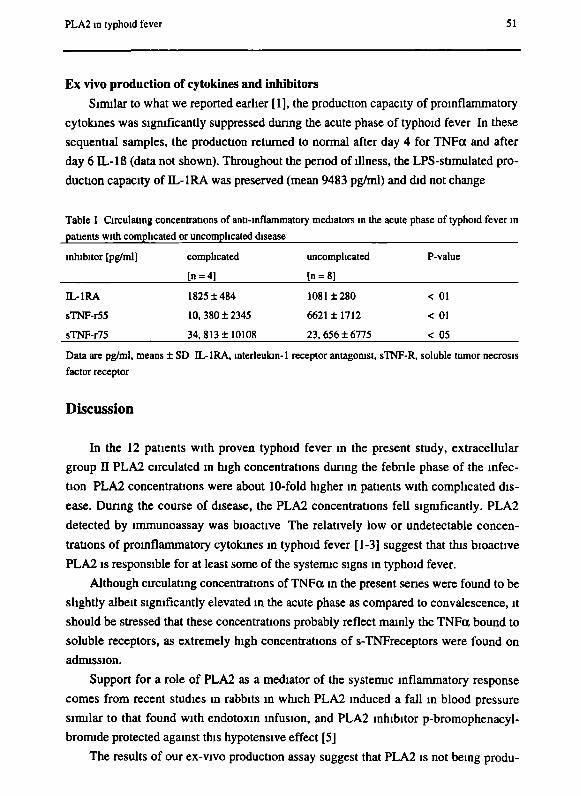
PLA2 in typhoid fever 51
Ex vivo production of cytokines and inhibitors Similar to what we reported earlier [1], the production capacity of proinflammatory
cytokines was significantly suppressed during the acute phase of typhoid fever In these
sequential samples, the production returned to normal after day 4 for TNFa and after
day 6 IL-lß (data not shown). Throughout the penod of illness, the LPS-stimulated pro
duction capacity of IL-1RA was preserved (mean 9483 pg/ml) and did not change
Table 1 Circulating concentrations of anti-inflammatory mediators in the acute phase of typhoid fever in
patients with complicated or uncomplicated disease
inhibitor [pg/ml]
IL-1RA
sTNF-r55
sTNF-r75
complicated
[n = 4]
1825 ±484
10, 380 ±2345
34, 813 ±10108
uncomplicated
[n = 8]
1081 ±280
6621 ±1712
23,656 ±6775
P-value
< 01
< 01
< 05
Data are pg/ml, means ± SD IL-IRA, interleukin-1 receptor antagonist, sTNF-R, soluble tumor necrosis
factor receptor
Discussion
In the 12 patients with proven typhoid fever in the present study, extracellular
group II PLA2 circulated m high concentrations during the febrile phase of the infec
tion PLA2 concentrations were about 10-fold higher in patients with complicated dis
ease. Dunng the course of disease, the PLA2 concentrations fell significantly. PLA2
detected by immunoassay was bioactive The relatively low or undetectable concen
trations of proinflammatory cytokines in typhoid fever [1-3] suggest that this bioactive
PLA2 is responsible for at least some of the systemic signs in typhoid fever.
Although circulating concentrations of TNFa in the present senes were found to be
slightly albeit significantly elevated m the acute phase as compared to convalescence, it
should be stressed that these concentrations probably reflect mainly the TNFa bound to
soluble receptors, as extremely high concentrations of s-TNFreceptors were found on
admission.
Support for a role of PLA2 as a mediator of the systemic inflammatory response
comes from recent studies m rabbits in which PLA2 induced a fall in blood pressure
similar to that found with endotoxin infusion, and PLA2 inhibitor p-bromophenacyl-
bromide protected against this hypotensive effect [5]
The results of our ex-vivo production assay suggest that PLA2 is not being produ-

52 Chapter 4
ced by circulating cells, but rather outside the bloodstream (e.g. the liver or in endo
thelial cells). As IL-lß and TNFa are proximal signals for PLA2 [6,7], it is conceivable
that in typhoid fever, proinflammatory cytokines are first produced.
Production of these cytokines may be confined to the site of the infection in the tis
sues, rather than in the circulation. PLA2 locally induced by these cytokines may reach
the circulation and mediate the systemic signs of the disease [6, 15]. This concept of
PLA2 as a circulating mediator could explain why circulating cytokines are not found in
clinical syndromes that are thought to be cytokine-mediated and that can easily be
mimicked by injection of cytokines.
Important issues remain to be investigated. It is for instance not clear how intravas
cular release of PLA2 is counter regulated, nor has the role of PLA2 in the production
of arachidonate derivatives been satisfactorily defined, especially in view of the lack of
specificity of group II PLA2 for arachnoidate. Moreover, the role of other secreted
(group Γ) PLA2 as a mediator of inflammatory processes has to be elucidated. Our study
suggests that PLA2 is a crucial mediator in a protracted inflammatory disease such as
typhoid fever. Inhibition of PLA2 may provide a new therapeutic intervention for
febrile diseases mediated by this enzyme.
References 1. Keuter M, Dharmana E, Gasem MH, Van der Ven-Jongekrijg J, Djokomoeljanto R,
Dolmans WMV, Demacker PMN, Sauerwein R, Gallati H, Van der Meer JWM.
Patterns of proinflammatory cytokines and inhibitors during typhoid fever. J Infect
Dis 1994; 169: 1306-1311.
2. Butler T, Ho M, Acharya G, Tiwari M, Gallati H. Interleukin-6, gamma interferon,
and tumor necrosis factor receptors in typhoid fever related to outcome of anti
microbial therapy. Antimicrob Agents Chemother 1993; 37: 2418-2421.
3. McGladderly S, Larasati R, Silitonga N, Punjabi N, Lesmana M, Pulungsih S,
O'Hanley P. Acute inflammatory cytokine responses in typhoid fever. Clin Infect
Dis 1993; 17: 578.
4. Roine I, Herrera Ρ, Ledermann W, Peltola Η. Tumor necrosis factor-α, interleukin-
lß and interleukin-6 levels in typhoid fever. 30th Interscience Conference on Anti
microbial Agents and Chemotherapy. Atlanta, GA, 1990: Abstract 299.
5. Vadas P, Hay JB. Involvement of circulating phospholipase A2 in the pathogenesis
of the hemodynamic changes in endotoxin shock. Can J Physiol Pharmacol 1983;
61: 561-566.
6. Crawl RM, Stoller TJ, Conroy RR, Stoner CR. Induction of phospholipase A2 gene

PLA2 in typhoid fever 53
expression in human hepatoma cells by mediators of the acute phase response. J
Biol Chem 1991; 266: 2647-2651.
7. Schalkwijk С, Pfeilschifter J, Marki F, Van den Bosch H. Interleukin-lß, tumor ne
crosis factor and forskolin stimulate the synthesis and secretion of group II phos-
pholipase A2 in rat mesangial cells. Biochem Biophys Res Commun 1991; 174:
268-275.
8. Pruzanski W, Vadas P. Phospholipase A2. A mediator between proximal and distal
effectors of inflammation. Immunol Today 1991; 12: 143-146.
9. Van Deuren M, Van der Ven-Jongekrijg J, Keuter M, Demacker PNM, Van der
Meer JWM. Cytokine production in whole blood cultures. J Int Fed Clin Chem
1993;5:216-221.
10. Van der Meer JWM, Endres S, Lonnemann G, Cannon JG, Dcejima T, Okusawa S,
Gelfand JA, Dinarello CA. Concentrations of immunoreactive human tumor necro
sis factor alpha produced by human mononuclear cells in vitro. J Leucocyte Biol
1988; 43: 216-223.
11. Lisi PJ, Chu CW, Koch GA, Endres S, Lonnemann G, Dinarello CA. Development
and use of radioimmunoassay for human interleukin-lß. Lymphokine Res 1987; 6:
229-244.
12. Poutsiaka DD, Clark BD, Vannier E, Dinarello CA. Production of interleukin-1 re
ceptor antagonist and interleukin-lß by peripheral blood mononuclear cells is diffe
rentially regulated. Blood 1991; 78: 1275-1281.
13. Van den Bosch H, Aarsman AJ. A review on methods of phospholipase A determi
nation. Agents Actions 1979; 9: 382-389.
14. Smith GM, Ward RL, McGuigan L, Rajkovic IA, Scott KF. Measurement of phos
pholipase A2 in arthritis plasma using a newly developed sandwich ELISA. Br J
Rheum 1992; 31: 175-178.
15. Inada M, Tojo H, Kawata S, Tarui S, Okamoto M. Induction of group Il-like phos
pholipase A2 by lipopolysaccharide in the liver of BCG-primed rats. Biochem
Biophys Res Commun 1991; 174: 1077-1083.

54 Chapter 4

Chapter 5
A semi-quantitative reverse transcriptase polymerase chain reaction method for measurement of mRNA for TNFa and IL-Iß in whole blood cultures: Its application in typhoid fever and eccentric exercise
Mihai G. Netea, Joost P. H. Drenth, Natasja de Bont, Anneke Hijmans, Monique
Keuter, Edi Dharmana, Pierre N.M. Demacker and Jos W.M. van der Meer
Cytokine 1996; 8:739-744

56 Chapter 5
Abstract
Whole blood cultures are used to study cytokine stimulation and release ex-vivo. In
the present study we compared this method with a more direct approach and a quan
titative reverse transcriptase polymerase chain reaction (RT-PCR) was used to assess
mRNA expression for interleukin (IL)-Iß and tumor necrosis factor (TNF)ct and mRNA
in whole blood.
Stimulation of whole blood from normal donors with lipopolysaccharide (LPS) at
various time intervals showed a parallel rise of immunogenic IL-Iß and TNFct as well
as a rise of mRNA expression for IL-Iß and TNFct with peak levels for IL-Iß after 4-6h
stimulation and for mRNA TNFct expression after 2h stimulation. These methods were
used to explore cytokine production during the course of typhoid fever and after a 5 km
run.
In both conditions circulating cytokine concentrations were not influenced, but the
TNFa and IL-Iß mRNA gene expression in circulating whole blood cells was increased
in patients with typhoid fever. The LPS stimulated production of TNFa and IL-Iß was
decreased in both but there was no change for the mRNA content in whole blood for
these cytokines. These findings demonstrate that RT-PCR is an attractive method to
study the gene expression of cytokines in whole blood, an increased TNFa and IL-Iß
gene expression is present in typhoid fever, and that the LPS stimulated down regula
tion of cytokines in exercise and typhoid fever may be mediated by post transcriptional
processes.

RT-PCR for cytokine mRNA 57
Introduction
Whole blood culture (WBC) is a practical and reproducible method for assessment
of cytokine production from human cells [1, 2]. Although a variety of stimulants have
been used, we prefer the addition of bacterial lipopolysaccharide (LPS) which is added
as an immunostimulus to venous blood. After a stationary incubation for 24h at 37°C,
the release of cytokines is measured and compared to the concentration in a control
tube. Minimal amounts of blood are required and insight is provided in the capacity to
produce cytokines [3]. In healthy volunteers addition of LPS to whole blood will lead to
measurable cytokine concentrations within few hours after incubation, certainly as a
result of an increased production and or cellular release into plasma [3]. The time
course of ex-vivo production of cytokines in WBC is comparable to that in healthy vo
lunteers after a single LPS injection [4]. We have used this method to assess the cyto
kine response in various clinical situations. Application of WBC revealed that the LPS
stimulated ex-vivo production of the inflammatory cytokines tumor necrosis factor-α
(TNFoc) and interleukin-lß (EL-Iß) becomes down regulated not only in the acute phase
of infections such as typhoid fever [5], meningococcal sepsis [6], and Pneumocystis
carinii infections [7], during attacks of familial Mediterranean fever [8] and after major
surgery [9] but also after strenuous physical exercise [10]. In all these clinical situa
tions, the production capacity of these cytokines restores during convalescence.
The mechanism of the depressed cytokine production is incompletely understood.
In LPS-stimulated whole blood from patients with sepsis, Northern blot analysis
showed a decreased expression of mRNA for TNFa and IL-6 and led to the conclusion
that transcription is a decisive mechanism for the modulation of cytokine production
[И].
Apart from this study in sepsis, the molecular mechanism of down-modulation of
cytokines in the above mentioned conditions is not known. In addition, the kinetics of
cytokine mRNA expression in WBC is not fully elucidated. The present study describes
a simple and reproducible method developed to assess the IL-Iß and TNFa mRNA in
whole blood with a semi-quantitative reverse transcriptase polymerase chain reaction
(RT-PCR). This method appears to be suitable to determine the kinetics of cytokine
mRNA expression in circulating cells after stimulation with LPS. Furthermore, with this
method it is possible to evaluate the transcription of mRNA in the down-modulation of
cytokine production capacity in various clinical conditions such as physical exercise
and typhoid fever.

58 Chapter 5
Materials and methods
Controls
Four healthy volunteers participated in the study in order to obtain data for study of
the kinetics of TNFa and IL-Iß protein production and gene expression in LPS stimu
lated whole blood.
Typhoid fever
Four patients with culture proven typhoid fever were recruited in Dr. Kariadi Hos
pital, Diponegoro University, Semarang, Indonesia as part of a project on cytokines in
the pathophysiology of typhoid fever. Samples were obtained on admission (acute) and
after 7-10 days after defervescence (recovery).
Exercise
Seven well-trained males were recruited in Wijchen, The Netherlands to perform a
5 km exercise run. The distance was covered between 18 min 40 sec and 22 min 16 sec.
Samples were drawn before (recovery) and immediately after the exercise (acute).
Blood samples
Blood for cytokine measurements was collected into 2-mL or 4-mL (ex-vivo pro
duction) endotoxin-free tubes containing EDTA (Vacutainer Systems, Becton and Dick
inson, Rutherford, NJ). Cytokine production was measured using a whole blood culture
system as described elsewhere [3]. Briefly, two 2-mL tubes containing 24μL EDTA-K3
(Bayer, Leverkusen, Germany) (4 ml; 48μί EDTA-K3) were drawn. One tube was
incubated immediately, the other tube was incubated after addition of 25μί LPS
(Escherichia coli serotype 055 :B5; Sigma, St Louis, MO; final concentration 10 μg per
mL blood).
After 24 hours of incubation at 37°C the tube was centrifuged at 2250 χ g for 10
minutes and secondly at 15 000 χ g for 5 minutes to obtain platelet-poor plasma. After
4h (exercise volunteers) or 2h (typhoid fever patients) after LPS-stimulation, aliquots of
500 [¡L blood were taken, and after addition of an equivalent amount of guanidinium-
isothiocyanate (GITC) stored at -70°C until RNA isolation. For studies on kinetics of
mRNA in healthy volunteers the 4-mL EDTA tube was incubated for 24 hours and after
0,0.5, 1, 2,4, 6, 8, and 24h 500μί whole blood was taken and subsequently centrifuged
at 2250 χ g for 10 minutes; plasma was used for cytokine protein determination and the
cell pellet was dissolved in 500 μί, (4M) GITC enriched with 7 μ ι ß-mercaptoethanol

RT-PCR for cytokine mRNA 59
for isolation of total RNA. Aliquots were stored at -70°C until assay.
Materials
M-MLV Reverse Transcriptase, DTT, RT buffer, Agarose and Taq DNA polyme
rase were all purchased from Life Technologies (Breda, The Netherlands) RNAsin was
purchased from Promega (Leiden, The Netherlands). dNTPs and pd(N)6 were purcha
sed from Pharmacia (Woerden, The Netherlands). DNAse I was purchased from Boeh-
ringer Mannheim. (Almere, The Netherlands) Ethidium-bromide was purchased from
Sigma (St. Louis, MO). PCR primers for human TNFot, TL-Iß and ß2M were obtained
from Dr. E. Mensink, (Division of Hematology, University Hospital Nijmegen, the
Netherlands) Quantitative densitometry of the gels was performed on a Molecular Ana-
lyst™/PC densitometer (Model GS-670, Biorad, Veenendaal, The Netherlands).
RNA isolation
Total RNA was isolated by the method of Chomczynski and Sacchi with minor
modifications [12]. Briefly, 500 μ ι of whole blood was resuspended in 1 mL GITC,
and sonicated for 10 min, followed by the addition of 2M sodium acetate, phenol and
chloroform / isoamylalcohol (49:1). After centrifugation, the RNA was precipitated
twice from aqueous phase with acidified ethanol. The RNA was dissolved in RNAase-
free sterile water. The amount and quality of RNA were determined by spectrophoto
metry and analysis by agarose gel electrophoresis.
RT-PCR Analysis of cytokine mRNA and ß2-microglobuIin (ß2M) mRNA
For each sample 0.5 μg of total RNA was reverse transcribed in a volume of 20 μL·
reverse transcriptase buffer (50mM Tris-HCL pH 8.3, 75 mM KCl, 3 mM MgCl2) con
taining lOmM dithiothreitol, 5μΜ random hexamers, 250μΜ dNTPs, 20U RNAsin,
200U M-MLV reverse transcriptase. Reaction mixtures were overlaid with mineral oil.
RT reaction was performed for 10 minutes at 20°C, followed by 45 minutes at 42°C,
and finishing for 10 minutes at 95°C using a Mastercycler 5330 (Eppendorf, Hamburg,
Germany), and the samples were stored at -20°C until PCR analysis was performed.
Sequences of the PCR primers for human TNFa, IL-Iß and ВгМ are listed in Table
1. Each primer pair was tested to determine the annealing temperature and the linear
range of the reaction (Table 1). PCR reactions consisted of 3 μΐ cDNA in 50 μΐ^ PCR
buffer (20mM Tris-HCL, pH 8.4, 50 mM KCl, 1.5 mM MgCl2 , 0.001% gelatin) con
taining ΙΟΟμΜ dNTP's, 0.3μΜ of each primer and 1.25U Taq polymerase. Reaction
mixtures were overlaid with mineral oil. PCR cycles were performed (30 sec denatu-
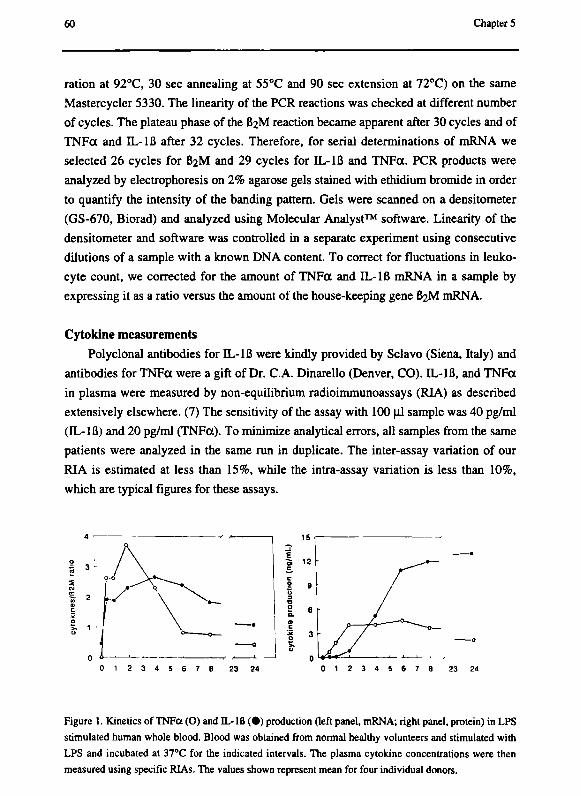
60 Chapter 5
ration at 92°C, 30 sec annealing at 55°C and 90 sec extension at 72°C) on the same
Mastercycler 5330. The linearity of the PCR reactions was checked at different number
of cycles. The plateau phase of the ВгМ reaction became apparent after 30 cycles and of
TNFa and IL-Iß after 32 cycles. Therefore, for serial determinations of mRNA we
selected 26 cycles for ВгМ and 29 cycles for IL-Iß and TNFa. PCR products were
analyzed by electrophoresis on 2% agarose gels stained with ethidium bromide in order
to quantify the intensity of the banding pattern. Gels were scanned on a densitometer
(GS-670, Biorad) and analyzed using Molecular Analyst™ software. Linearity of the
densitometer and software was controlled in a separate experiment using consecutive
dilutions of a sample with a known DNA content. To correct for fluctuations in leuko
cyte count, we corrected for the amount of TNFa and IL-Iß mRNA in a sample by
expressing it as a ratio versus the amount of the house-keeping gene ВгМ mRNA.
Cytokine measurements
Polyclonal antibodies for IL-Iß were kindly provided by Sciavo (Siena, Italy) and
antibodies for TNFa were a gift of Dr. C.A. Dinarello (Denver, CO). EL-Iß, and TNFa
in plasma were measured by non-equilibrium radioimmunoassays (RIA) as described
extensively elsewhere. (7) The sensitivity of the assay with 100 μΐ sample was 40 pg/ml
(IL-Iß) and 20 pg/ml (TNFa). To minimize analytical errors, all samples from the same
patients were analyzed in the same run in duplicate. The inter-assay variation of our
RIA is estimated at less than 15%, while the intra-assay variation is less than 10%,
which are typical figures for these assays.
Figure 1. Kinetics of TNFa (O) and IL-lfl ( · ) production (left panel, mRNA; right panel, protein) in LPS
stimulated human whole blood. Blood was obtained from normal healthy volunteers and stimulated with
LPS and incubated at 37°C for the indicated intervals. The plasma cytokine concentrations were then
measured using specific RIAs. The values shown represent mean for four individual donors.
0 1 2 3 4 5 6 7 8 23 24

RT-PCR for cytokine mRNA 61
Table 1 List of pnmer pairs used for mRNA amplification, with data regarding sequence, annealing tem
perature (Tm°C), size of the PCR product (bp) and number of PCR cycles within the exponential phase of
the reaction
pnmer sequence bp Tm(°C) cycles
ß2M sense S'-CCAGCAGAGAATGGAAAGTC-З· 268 55 26
antisense 5'-GATGCTGCTTACATGTCTCG-3'
TNFa sense 5'-ACAAGCCTGTAGCCCATGTT-3' 427 55 29
antisense 5'-AAAGTAGACCTGCCCAGACT-3'
IL-IB sense 5'-GGATATGGAGCAACAAGTGG-3' 263 55 29
antisense 5'-ATGTACCAGTTGGGGAACTG-3'
Statistical analysis
The non-paired non-parametric Mann-Whitney test was used for statistical compa
rison of results. Probability (p) values were calculated on the basis of two-tailed tests. A
ρ value of < 0.05 considered to be the lowest level of significance.
Results
Time course of IL-lß and TNFa gene expression
The induction of TNFa mRNA in healthy volunteers was very rapid, and high
levels were attained after 30 min. Maximum expression was found after 2h of incu
bation with LPS. After this time point, the TNFa mRNA levels declined and after 24h
incubation values were still higher compared to those at baseline. IL-lß mRNA expres
sion became increased 2h after LPS stimulation, but maximum values were reached
now until only after 4-6h. Compared to TNFa, IL-lß mRNA decreased slower (Fig la).
The kinetics of TNFa and IL-lß protein secretion production and release follow that of
the mRNA transcription (Fig lb).
Table 2 Circulating concentrations (circ) and ex-vivo production of IL-IB and TNFa in supematants of
whole blood stimulated for 24 h at 37°C with LPS in patients suffering from typhoid fever and healthy
volunteers subjected to a 5 km run
Clinical situation
Typhoid fever
Physical exercise
circ
ex-vivo
circ
ex-vivo
TNFa
acute
0 1 6 ± 0 0 2
4 4 ± 1 1
0 0 9 ± 0 0 2
3 0 ± 1
(ng/ml)
recovery
0 I 9 ± 0 1 2
7 1 ± 3 4
0 0 9 ± 0 0 3
6 9 ± 1 8
IL-lß (ng/ml)
acute
0 17±003
3 5 ± 2 8
0 0 5 ± 0 0 1
1 7 2 ± 9 9
recovery
0 14 ± 0 02
5 2 ± 4 8
0 0 6 ± 0 1 8
22±8 5

62 Chapter 5
Ladder Acute circulating B2M Acute circulating TNFo Acute circulating IL-lfl
: Recovery circulating B2M Recovery circulating TNFa Recovery circulating DL-lß
Figure 2. RT-PCR analysis of whole blood from a patient during and after the acute stage of typhoid
fever. Whole blood was stimulated with LPS and at the 2h time point mRNA was extracted and subjected
to RT-PCR. The lanes show mean values for the ratio of TNFa and IL-Iß to ВгМ expression for four
volunteers.
Circulating and ex-vivo production of cytokines
Typhoidfever
In patients with typhoid fever, circulating concentrations of IL-Iß and TNFa
remained unchanged regardless of the phase of the disease (Table 2). However, mRNA
analysis from the circulating uncultured cells of typhoid fever patients in the acute
phase of the disease revealed an increase of both IL-lß/ß2M and TNFa/ß2M ratios
when compared to the recovery phase (Fig 2).
LPS stimulated WBC of patients in the acute phase of typhoid fever showed a decreased
production of IL-1 ß and TNFa, but the production of these proteins restored during
convalescence. In contrast to the decreased ex-vivo LPS-stimulated production of the
DL-lß and TNFa proteins, the ПЛВ/ВгМ and TNFct/BîM mRNA ratios did not differ
significantly between the acute and recovery phase of typhoid fever (Table 3).
Exercise
The 5 km run did not influence the circulating IL-Iß or TNFa concentrations, nor
was there any effect on mRNA content for these cytokines in uncultured whole blood
(Table 2, 3). Similar to typhoid fever, we observed an inhibition of LPS-stimulated ex-
vivo production of both IL-Iß and TNFa in blood samples from athletes after a 5 km
run compared to pre-exercise values but the difference was larger for TNFa. Remarka
bly, the IL-Iß production capacity obtained before exercise was substantially higher in
the athletes compared to the recovered typhoid fever patients and to the values obtained
from healthy sedentary controls for our laboratory. The mRNA values for IL-Iß and
TNFa were increased by the 5 km run, although these differences were not statistically
significant (Table 3).
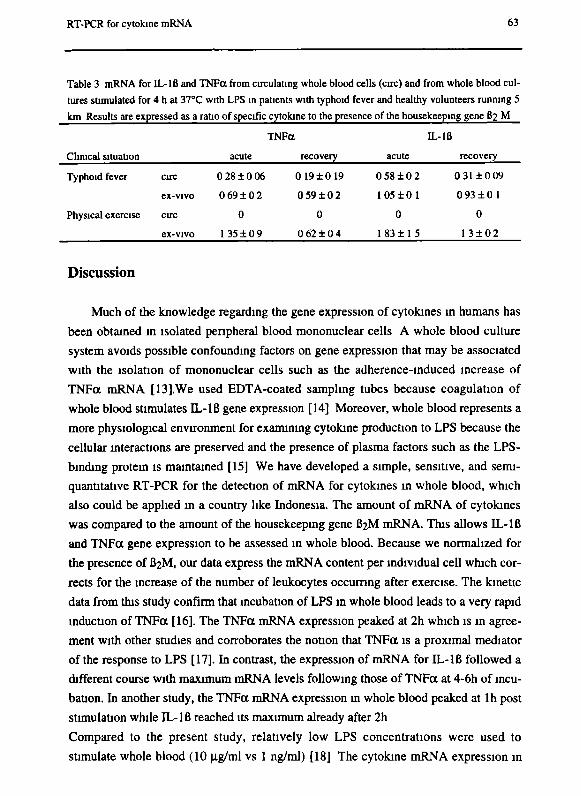
RT-PCR for cytokine mRNA 63
Table 3 mRNA for IL-Iß and TNFa from circulating whole blood cells (arc) and from whole blood cul
tures stimulated for 4 h at 37°C with LPS in patients with typhoid fever and healthy volunteers running 5
km Results are expressed as a ratio of specific cytokine to the presence of the housekeeping gene B2 M
TNFa IL-lß
Clinical situation acute recovery acute recovery
Typhoid fever arc 0 28 ± 0 06 0 19±0 19 0 58 ± 0 2 0 31 ± 0 09
ex-vivo 0 6 9 ± 0 2 0 5 9 ± 0 2 1 0 5 ± 0 1 0 9 3 ± 0 1
Physical exercise arc 0 0 0 0
ex-vivo 1 3 5 ± 0 9 0 6 2 ± 0 4 1 83 ± 1 5 13 + 0 2
Discussion
Much of the knowledge regarding the gene expression of cytokines in humans has
been obtained in isolated peripheral blood mononuclear cells A whole blood culture
system avoids possible confounding factors on gene expression that may be associated
with the isolation of mononuclear cells such as the adherence-induced increase of
TNFa mRNA [13].We used EDTA-coated sampling tubes because coagulation of
whole blood stimulates IL-lß gene expression [14] Moreover, whole blood represents a
more physiological environment for examining cytokine production to LPS because the
cellular interactions are preserved and the presence of plasma factors such as the LPS-
binding protein is maintained [15] We have developed a simple, sensitive, and semi
quantitative RT-PCR for the detection of mRNA for cytokines in whole blood, which
also could be applied in a country like Indonesia. The amount of mRNA of cytokines
was compared to the amount of the housekeeping gene ВгМ mRNA. This allows IL-lß
and TNFa gene expression to be assessed in whole blood. Because we normalized for
the presence of ВгМ, our data express the mRNA content per individual cell which cor
rects for the increase of the number of leukocytes occurring after exercise. The kinetic
data from this study confirm that incubation of LPS in whole blood leads to a very rapid
induction of TNFa [16]. The TNFa mRNA expression peaked at 2h which is in agree
ment with other studies and corroborates the notion that TNFa is a proximal mediator
of the response to LPS [17]. In contrast, the expression of mRNA for IL-lß followed a
different course with maximum mRNA levels following those of TNFa at 4-6h of incu
bation. In another study, the TNFa mRNA expression m whole blood peaked at lh post
stimulation while IL-lß reached its maximum already after 2h
Compared to the present study, relatively low LPS concentrations were used to
stimulate whole blood (10 μg/ml vs 1 ng/ml) [18] The cytokine mRNA expression in

64 Chapter S
circulating whole blood cells in typhoid fever patients could be assessed. Despite the
absence of differences of cytokine concentrations, there is an increase of mRNA expres
sion for IL-Iß and TNFa in patients with acute typhoid fever. This presence of cytokine
mRNA in circulating cells of these patients suggests that the cytokine network is activa
ted. The pathophysiological significance of these findings for typhoid fever are present
ly unclear. The absence of increased cytokine proteins in acute typhoid fever could be
due to an increased turnover of protein for these cytokines. However, it remains possi
ble that the increased expressed mRNA is not translated and thus does not lead to the
appearance of these proteins in the circulation. The absence of any change in the circu
lating TNFa and IL-Iß concentrations in athletes is associated with absence of mRNA
for these cytokines. Apparently, the stress related to a 5 km run is not intense enough to
elicit detectable cytokines in the circulation of these athletes.
Study of the mRNA expression of cytokines in LPS-stimulated whole blood in
patients with typhoid fever and athletes allowed us to make an important observation.
Despite a decreased cytokine production in the acute phase of typhoid fever and post-
exercise, both IL-Iß and TNFa mRNA expression was similar in the acute and recovery
phase. These findings suggest that under the conditions studied, the inhibition of the
LPS-stimulated ex-vivo cytokine production is a post-transcriptional event. In this
respect our results differ from those in a recent study in sepsis patients, where a decrea
sed TNFa and IL-6 mRNA expression in LPS stimulated whole blood was found [11].
This suggests that the molecular mechanism of down-modulation of cytokine produc
tion may be influenced by type of infection.
The rate of mRNA degradation is the most important mechanism of post transcrip
tional deactivation [19]. Several additional post-transcriptional processes are potentially
able to influence the presence of cytoplasmic mRNA, including capping, splicing, poly-
adenylation, nuclear export and compartmentalization [20-24]. Furthermore, kinetic
influences may play a role and the rate of degradation of IL-Iß and TNFa proteins may
precede the degradation of mRNA. Nevertheless, our results are in line with ex-vivo
experiments with LPS-tolerant mice, showing that peritoneal macrophages did not pro
duce TNFa when restimulated, while its mRNA was still induced. This strongly suggest
a posttranscriptional regulation of TNF down-regulation [25].
Acknowledgments J.P.H. Drenth is a recipient of a Dutch Organization for Scientific Research fellow
ship for Clinical Investigators. (KWO 900-716-065) The authors wish to thank the ath
letes from "Athletiekvereniging Wijchen" (Wijchen, The Netherlands) for their coope-

RT-PCR for cytokine mRNA 65
ration with this study. R. Krebbers and Dr. S.H.M. van Uum are thanked for their help
in performing the study.
References
1. Nerad JL, Griffiths JK, Van der Meer JWM, Endres S, Poutsiaka DD, Keusch GT,
Bennish M, Salam MA, Dinarello CA, Cannon JG. Interleukin-lß (IL-lß), IL-1
receptor antagonist, and TNF-a production in whole blood. J Leukocyte Biol 1992;
52: 687-692.
2. Finch-Arietta MB, Cochran FR. Cytokine production in whole blood ex vivo.
Agents Actions 1991; 34:49-52.
3. Van Deuren M, Van der Ven-Jongekrijg J, Keuter M, Demacker PNM, Van der
Meer JWM. Cytokine production in whole blood cultures. J Int Fed Clin Chem
1993;5:216-221.
4. Van Zee KJ, Kohno T, Fischer E, Rock CS, Moldawer LL, Lowry SF. Tumor
necrosis factor soluble receptors circulate during experimental and clinical inflam
mation and can protect against excessive tumor necrosis factor-alpha in vitro and in
vivo. Proc Natl Acad Sci 1992; 89: 4845-4849.
5. Keuter M, Dharmana E, Gasem MH, Van der Ven-Jongekrijg J, Djokomoeljanto R,
Dolmans WMV, Demacker PMN, Sauerwein R, Gallati H, Van der Meer JWM.
Patterns of proinflammatory cytokines and inhibitors during typhoid fever. J Infect
Dis 1994; 169: 1306-1311.
6. Van Deuren M, Van der Ven-Jongekrijg J, Demacker PNM, Bartelink AKM, Van
Dalen R, Sauerwein RW, Gallati H, Vannice JL, Van der Meer JWM. Differential
expression of proinflammatory cytokines and their inhibitors during the course of
meningococcal infections. J Infect Dis 1994; 169: 157-161.
7. Perenboom RM, Van Schijndel ACHW, Beckers P, Sauerwein R, Van Hamersvelt
HW, Festen J, Gallati H, Van der Meer JWM. Cytokine profiles in broncho-alveo
lar lavage fluid and blood in Ш -seronegative patients with Pneumocystis carinii
pneumonia. Eur J Clin Invest 1994; 26: 159-166.
8. Rozenbaum M, Katz R, Rozner I, Pollack S. Decreased interleukin-1 activity relea
sed from circulating monocytes of patients with familial Mediterranean fever
during in vitro stimulation by lipopolysaccharide. J Rheumatol 1992; 19: 416-418.
9. Kruimel JW, Naber AHJ, Van der Vliet AJ, Buskens FGM, Pesman G, Sweep F,
Liem T, Kho HG, Van der Meer JWM, Jansen JBMJ. Effects of abdominal surgery
on cytokines in a clinical trial designed to study the effects of parenteral admini-

66 Chapter S
stration of lipid emulsions. Neth J Med 1994; 45: A17-A18.
10. Drenth JPH, Van Uum SHM, Van Deuren M, Pesman GJ, Van der Ven-Jongekrijg
J, Van der Meer JWM. Endurance run increases circulating IL-6 and IL-Ira but
downregulates ex vivo TNF-a and IL-lß production. J Appi Physiol 1995; 79:
1497-1503.
11. Ertel W, Kremer JP, Kenney J, Steckholzer U, Jarrar D, Trentz O, Schildberg FW.
Downregulation of proinflammatory cytokine release in whole blood from septic
patients. Blood 1995; 85: 1341-1347.
12. Chomczynski P, Sacchi N. Single step method of RNA isolation by guanidinum
thiocyanate phenol chloroform extraction. Anal Biochem 1987; 162: 156-159.
13. Haskill S, Johnson C, Eierman D, Becker S, Warren K. Adherence induces selec
tive mRNA expression of monocyte mediators and proto-oncogenes. J Immunol
1988; 140: 1690-1695.
14. Mileno MD, Margolis NH, Clark BD, Dinarello CA, Burke JF, Gelfand JA. Coagu
lation of whole blood stimulates interleukin-lß gene expression. J Infect Dis 1995;
172:308-311.
15. Wright SD, Ramos RA, Tobias PS, Ulevitch RJ, Mathison JC. CD14, a receptor for
complexes of lipopolysaccharide (LPS) and LPS binding protein. Science 1990;
249: 1431-1433.
16. Oliver JC, Bland LA, Oettinger CW, Arduino MJ, McAllister SK, Agüero SM,
Favero MS. Cytokine kinetics in an in vitro whole blood model following an endo
toxin challenge. Lymphokine and Cytokine Research 1993; 12: 115-120.
17. DeForge LE, Kenney JS, Jones ML, Warren JS, Remick DG. Biphasic production
of IL-8 in lipopolysaccharide (LPS)-stimulated human whole blood. J Immunol
1992; 148:2133-2141.
18. Dedrick RL, Conlon PJ. Prolonged expression of lipopolysaccharide (LPS)-induced
inflammatory genes in whole blood cultures requires continual exposure to LPS.
Infect Immun 1995; 63: 1362-1368.
19. Sachs AB. Messenger RNA degradation in eukaryocytes. Cell 1993; 74:
20. Konarska MM, Padgett RA, Sharp PA. Recognition of cap structure in splicing in
vitro mRNA precursors. Cell 1984; 38: 731-736.
21. Amrein H, Hedley ML, Maniatis R. The role of specific protein-RNA and protein-
protein interactions in positive and negative control of pre-mRNA splicing by
transformer 2. Cell 1994; 76: 735-746.
22. Sachs A, Wähle E. Poly(A) tail metabolism and function in eukaryocytes. J Biol
Chem 1993; 268: 22955-22958.

RT-PCR for cytokine mRNA 67
23. Jarmolowski A, Boelens WCM, Izauralde E, Mattaj IW. Nuclear export of different
classes of RNA is mediated by specific factors. J Cell Biol 1994; 124: 627-635.
24. Hill MA, Schedlich L, Gunning P. Serum-induced signal transduction determines
the peripheral location of ß-actin mRNA within the cell. J Cell Biol 1994; 126:
1221-1230.
25. Zuckerman SH, Evans GF, Snyder YM, Roeder WD. Endotoxin-macrophage inter
action: posttranslational regulation of tumor necrosis factor expression. J Immunol
1989; 143: 1223-1237.

68 Chapter 5

Chapter 6
Pro-inflammatory cytokine profile in Salmonella
typhimurium infection in mice and the effect of recombinant IL-la
Monique Keuter, Edi Dharmana, В art-Jan Kullberg, Ineke Verschueren and
Jos W.M. van der Meer
Submitted

70 Chapter 6
Abstract
The endogenous cytokine response and the effect of recombinant human interleu-
kin-loc (rhIL-la) pretreatment in S. typhimurium infection was investigated in CBA/J
mice. Administration of 80 ng rhIL-la i.p. 24 h before infection significantly decreased
bacterial outgrowth in organs on days 3 and 7.
In plasma, circulating concentrations of tumor necrosis factor-α (TNFcc), IL-la
and IL-Iß before and after infection were low. On day 7, TNFa was 145 ± 7 pg/mL
(mean ± SD). Ex vivo LPS-induced cytokine production of blood without infection was
negligible; after Salmonella infection, production increased to day 7. Bone marrow
cells produced low cytokine concentrations without infection; in cells obtained from
infected mice, production increased. Peritoneal cells produced cytokines without infec
tion: IL-la 4081 ± 1554 pg/mL, IL-lß 68 ± 25 pg/mL and TNFa 414 ± 185 pg/mL. In
infected cells, production increased, except for IL-la.
Pretreatment with rhIL-la did not affect circulating cytokines but increased LPS-
induced IL-lß production of blood ex vivo at day 7 (1234 ± 765 vs 2137 ± 581 pg/mL
(p < .005)). RhIL-la pretreatment without infection significantly increased production
of IL-la by peritoneal (to 6190 ± 2190 pg/mL: ρ < .01) and bone marrow macrophages
(from 57 ± 14 pg/mL to 161 ± 69 pg/mL: ρ < .0001), respectively.
In conclusion, production of IL-lß and TNFa increased during S. typhimurium in
fection. Pretreatment with rhIL-la decreased outgrowth of microorganisms and increa
sed endogenous IL-la production of uninfected bone marrow and peritoneal cells.

rIL-1 in S. typhimunum infection 71
Introduction
Antibiotic therapy has reduced complications and mortality in typhoid fever, the
most important Salmonella infection in man, but the mortality of the disease is still 5%
[1]. Therefore, methods are sought to increase resistance of the host against S. typhi. To
this end, more insight in the pathophysiology of Salmonella infections is required. In
studies in patients with typhoid fever, we have measured production of cytokines after a
stimulus such as lipopolysaccharide (LPS) in a whole blood culture system [2]. Howe
ver, sequential cytokine responses in the blood and information on cytokines at the
tissue level cannot be obtained in humans.
Therefore, the mouse model of typhoid fever, induced with S. typhimurium, can
provide useful information on host defense in typhoid fever. Low concentrations of
circulating proinflammatory cytokines have been reported in this experimental infection
model in C57B1/6J mice [3]. However, no data exist on the cytokine production of mu
rine macrophages infected with Salmonella in the experimental model of typhoid fever.
Pretreatment with recombinant interleukin-1 (rIL-1) has been shown to ameliorate
the outcome of Gram-negative and Gram-positive infections in normal and neutropenic
mice. It also increases resistance against candidal and plasmodial infections in mice [4-
7]. The mechanism of this protection is multifactorial, the main mechanism in Gram-
negative infections being a down-regulation of TNFcc production and a modulation of
cytokine receptors [8]. In addition IL-1 pretreatment augments antifungal activity by a
mechanism that has not been fully elucidated [4].
Recent studies on experimental S. typhimurium infection in mice have shown that
rIL-1 pretreatment increased survival of CBA/J mice [9]. This finding is remarkable,
since rIL-1-induced inhibition of TNFa production might be deleterious for the defense
against facultative intracellular Gram-negative bacteria such as the salmonellae. TNFa
has been shown to contribute to the intracellular killing of Salmonella by macrophages
[10,11].
In the present study, the cytokine profiles and bacterial outgrowth during S. typhi
murium infection were investigated in CBA/J mice, with and without pretreatment with
recombinant human (rh)IL-la.
Material and methods
Animals
Specific pathogen-free female CBA/J mice, weighing 20-25 grams, were housed

72 Chapter 6
under standard laboratory conditions and fed sterilized laboratory chow (Hope Farms,
Woerden, The Netherlands) and water ad libitum. The experiments were approved by
the ethics committee for animal experiments at the Catholic University Nijmegen.
Materials
rhIL-la, a gift from Hoffmann-La Roche, was diluted in pyrogen-free saline to a
final concentration of 0.8 \ig/n\L. The endotoxin content was < 20 pg/mg.
Salmonella typhimurium infection
A serum-resistant strain of S. typhimurium (phage type 510) was grown to statio
nary phase culture by overnight incubation at 37CC in nutrient broth (BHI Oxoid). The
LD50 in the ItyR CBA/J mice was 5 χ IO3 bacteria. СВАЯ mice were injected i.p. with
5 χ 103 cfu of S. typhimurium.
rhIL-la was given as an i.p. injection of 80 ng in 100 цЬ of pyrogen free saline at
24 hours before the infection (day -1). Control mice received pyrogen free saline.
Just before the infection with S. typhimurium and on days 1, 3 or 7 after the injec
tion, mice were killed by cervical dislocation and outgrowth of the microorganisms
from the liver and spleen was quantified. For this purpose, the liver and spleen were
removed aseptically, weighed, and homogenized in sterile saline in a tissue grinder. The
number of viable salmonellae was determined by plating several dilutions on Brilliant
Green agar (BGA) plates and cfu were counted after overnight incubation at 37°C. The
results were expressed as the log cfu per gram of tissue.
Cytokine studies
Circulating cytokines
Before infection and on days 1, 3 and 7 of infection, separate groups of mice were
anesthetized with ether to collect blood from the intraorbital plexus for the measu
rement of circulating cytokine concentrations. TNFa, IL-la and IL-Iß concentrations
were determined using specific radioimmunoassays (RIA) developed in our laboratory,
as previously described [12].
Ex-vivo cytokine production
Whole blood cells, resident peritoneal macrophages and bone marrow macro
phages were stimulated ex vivo with LPS before infection and on day 1, 3 and 7 of in
fection with S. typhimurium. Production of cytokines before and during infection of
control mice was compared with that of rhIL-la -pretreated mice.

rIL-1 in S. typhimunum infection 73
For that purpose, EDTA blood was diluted thrice in RPMI 1640 (Dutch modi
fication; Flow Laboratories, Irvine, UK) and stimulated with lipopolysaccharide (LPS)
(E. coli serotype 055:B5; Sigma, St Louis, USA; final concentration l(^g/mL). After
incubation for 24 hours at 37°C, the tubes were centrifuged and the supernatant was
stored at -70°C until assayed.
Before infection, and on days 1, 3 and 7 after the injection of S. typhimurium, peri
toneal macrophages were harvested by rinsing the peritoneal cavity aseptically with 2
ml cold PBS containing 0.38% (w/v) sodium citrate. Bone marrow cells were flushed
from the femora of the mice with 1 ml PBS. After centrifugation, cells were resuspen-
ded in RPMI 1640, containing 1 mM pyruvate, 2 mM L-glutamine and 100 μg gentami-
cin per mL, and incubated for 1 h at 37°C (lO^well) in 96-wells microtiter plates (Co-
star Corporation, Cambridge, MA). The non-adherent cells were discarded and the re
maining cell population consisted of more than 90% macrophages, as assessed by light
microscopy.
Cells (105/ well) were incubated with LPS (1 ng in 100μί of RPMI) at 37°C in 5%
CO2. After 24h, the supernatants were collected and stored at -70°C until assayed. To
the remaining macrophages, 200 \iL of RPMI was added and the cells were disrupted
by three freeze-thaw cycles to determine the cell-associated cytokines. The samples
were stored at -70°C until assayed for cytokines as described above.
Statistical analysis
Differences in concentrations of cytokines and in numbers of the microorganisms
were analyzed by the Mann-Whitney U test. Differences were considered significant at
ρ < .05. All the experiments were at least performed in duplicate.
Results
Outgrowth of Salmonella in the organs
After infection with 5 χ 103 cfu S. typhimurium, the numbers of microorganisms in
the livers and spleens rose rapidly in control mice (Figure 1). Administration of 80 ng
rhfL-la i.p. 24 hours before injection of S. typhimurium significantly decreased
bacterial outgrowth in liver and spleen on day 3 and 7 of infection (Figure 1).
Effect of rhIL-la pretreatment on peripheral blood and peritoneal cell counts
There was no difference in numbers of peripheral blood neutrophils between rhlL-
loc pretreated animals and control mice before infection and on day 3 of infection (data
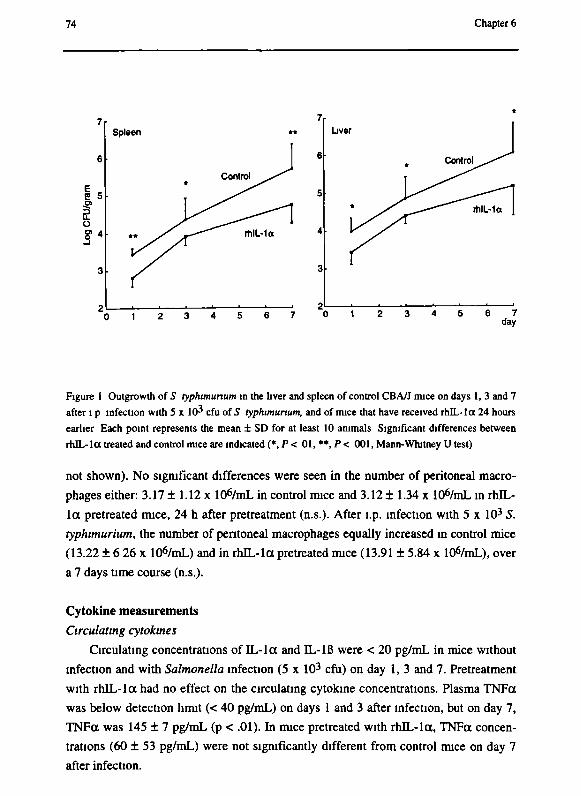
74 Chapter 6
Is f о Ç4
Spleen Liver
0 1 2 3 4 5 6 7 0 1 2 3 4 5 6 7 day
Figure 1 Outgrowth of S typhimunum in the liver and spleen of control СВАЛ mice on days 1, 3 and 7
after ι ρ infection with 5 χ ICH cfu of S typhimunum, and of mice that have received rhIL-la 24 hours
earlier Each point represents the mean ± SD for at least 10 animals Significant differences between
rhIL-la treated and control mice are indicated (*,Ρ < 0l,**,P< 001, Mann-Whitney U test)
not shown). No significant differences were seen in the number of peritoneal macro
phages either: 3.17 ± 1.12 χ lOfymL in control mice and 3.12 ± 1.34 χ lOfymL in rhlL-
lot pretreated mice, 24 h after pretreatment (η.s.). After l.p. infection with 5 χ 103 S.
typhimunum, the number of peritoneal macrophages equally increased in control mice
(13.22 ± 6 26 χ lOfymL) and in rhIL-la pretreated mice (13.91 ± 5.84 χ lOfymL), over
a 7 days time course (n.s.).
Cytokine measurements
Circulating cytokines
Circulating concentrations of IL-la and IL-Iß were < 20 pg/mL in mice without
infection and with Salmonella infection (5 χ 103 cfu) on day 1, 3 and 7. Pretreatment
with rhIL-la had no effect on the circulating cytokine concentrations. Plasma TNFot
was below detection limit (< 40 pg/mL) on days 1 and 3 after infection, but on day 7,
TNFcc was 145 ± 7 pg/mL (p < .01). In mice pretreated with rhIL-la, TNFa concen
trations (60 ± 53 pg/mL) were not significantly different from control mice on day 7
after infection.

rIL-1 in S. typhimuríum infection 75
Ex vivo cytokine production in whole blood
Before infection, LPS-induced cytokine production of whole blood cells ex vivo
was negligible. During Salmonella infection, production of IL-la and IL-Iß significant
ly increased from day 3 to day 7 (p < .001; Figure 2). Production of TNFa significantly
rose from day 0 to day 3 (p < .01) and from day 3 to day 7 (p < .001).
The same pattern was seen after rbJL-Ια pretreatment. Pretreatment with rhIL-Ια
led to a significant increase of IL-lß production at day 7 (p < .01), but did not affect
production of IL-la or TNFa (Figure 2).
Ex vivo cytokine production by bone marrow macrophages
LPS induced IL-lß production by bone marrow cells in mice without infection was
below the detection limit. IL-la production was 57 ± 14 pg/mL and TNFa production
was 206 ± 99 pg/mL. During 5. typhimuríum infection, IL-la production significantly
increased from day 0 (without infection) to day 3 (p < .0001), whereas IL-lß and TNFa
rose non-significantly during infection. Pretreatment with rhIL-Ια significantly increa
sed the production of cell-associated IL-la before infection (p = .0001; Figure 2).
Ex vivo cytokine production by peritoneal macrophages
Production of IL-lß and TNFa was significantly increased 3 days after Salmonella
infection (p < .001). In contrast, IL-la production was significantly decreased at day 3
(p < .0001). Pretreatment with rhIL-Ια only increased the IL-la production before in
fection (p = 0.01 ; Figure 2).
Discussion
In the present study, we demonstrate that treatment with rhIL-Ια strongly reduces
outgrowth of S. typhimuríum in the organs of mice, and this is probably an explanation
for the improved survival of rhIL-Ια-treated animals. The microbiological effect of a
single dose of rhIL-Ια was established early in the infection, and lasted throughout the
study.
A possible mechanism of the reduction in outgrowth of S. typhimuríum in the
organs includes recruitment and activation of neutrophils with enhanced killing of
microorganisms. Since granulocyte numbers were not found to be increased in our ex
periments (data not shown) and in those of others [13], the effect is probably not due to
induction of G-CSF by IL-1. This is underscored by our recent finding that recombinant
murine G-CSF increases survival of mice with 5. typhimuríum infection, but does not
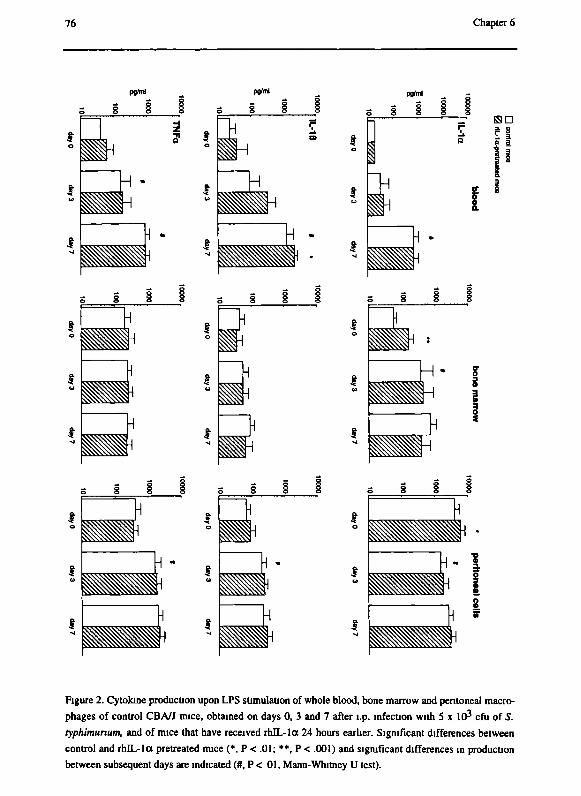
76 Chapter 6
pgM
J i_
H
ч ¡1
I i
Ш
H
li
ss
pgM
i I * t
si 4
4
Ш
I i
sa
го**
S §
ш
^
ч · ?
ш Η
Figure 2. Cytokine production upon LPS stimulation of whole blood, bone marrow and peritoneal macro
phages of control СВАЛ mice, obtained on days 0, 3 and 7 after l.p. infection with 5 χ 1(P cfu of S.
typhimunum, and of mice that have received rhIL-la 24 hours earlier. Significant differences between
control and rhEL-la pretreated mice (*, Ρ < .01; **, Ρ < .001) and significant differences in production
between subsequent days are indicated (#, Ρ < 01, Mann-Whitney U test).

rIL-1 in S. typhimurium infection 77
reduce outgrowth of Salmonella in the organs [14].
Thus, we addressed the question whether the increased microbicidal effect was due
to an influence on cytokines, especially as it is known that 1L-1 is able to stimulate its
own production and that of TNFa [15]. TNFa is capable to inhibit the outgrowth of
intracellular salmonellae, probably by augmenting macrophage function [10, 11, 16].
The increased nitrite production by peritoneal macrophages after pretreatment with rlL-
1 in vivo may also play a role [17].
The effect of rhIL-la pretreatment on cytokine production was found to occur at
24 hours after administration, leading to significantly increased cell-associated IL-la
production by peritoneal and bone marrow macrophages at that time point. This sug
gests that the rhIL-Ια-induced modulation of cytokine production may have caused the
reduction of microorganisms, since the microbiological effect was also established early
in the infection.
Peritoneal macrophages of rhIL-la treated mice might have reduced the numbers
of 5. typhimurium by killing the microorganisms, before the distribution of Salmonella
to the circulation and the subsequent invasion of the organs. This would mean confi
nement of the infection to the peritoneal space and a direct effect of rhIL-la on distri
bution of Salmonella in the early phase of the infection. The outgrowth pattern does
suggest such a scenario, as the rate of outgrowth in the organs of control mice and rhlL-
l a pretreated mice is similar.
RhIL-la had no effect on outgrowth of microorganisms during infection with
Pseudomonas aeruginosa in neutropenic mice [8]. In those experiments, it has been
shown that administration of either rIL-1 or rTNFa increased survival of mice, which
was partially mediated by reduction of circulating concentrations of proinflammatory
cytokines and reduction of LPS-stimulated cytokine production by peritoneal macro
phages [8]. In contrast to extracellular Pseudomonas infections, the cause of death of
mice infected with the facultative intracellular pathogen Salmonella is most probably
organ dysfunction rather than cytokinemia. In the present animal study, concentrations
of TNFa remained low throughout the infection.
Although it has been reported previously, that TNFa is required in Salmonella
infection [10, 11, 16], we did not find an effect of rhIL-la on LPS-stimulated produc
tion of TNFa by peritoneal cells or bone marrow macrophages either before or during
S. typhimurium infection. Moreover, the beneficial effect of rhIL-la on numbers of
microorganisms slightly increased during 7 days of infection, whereas an inhibitory
effect on macrophage activation through reducing TNFa production would have led to
increased outgrowth.

78 Chapter 6
Whereas LPS-stimulated cytokine production during S. typhimurium infection was
generally increased, the IL-la production by infected peritoneal cells was decreased.
This is an important observation, since peritoneal macrophages are easy to obtain and
frequently used as representatives for the resident or activated macrophage population
of the mouse. The present study shows that the cytokine response of peritoneal and
other macrophages are divergent. The attention to peritoneal cells alone, might not yield
information on cytokine production at other sites of infection.
There are important differences in cytokine patterns between these mouse
experiments and the studies in patients with typhoid fever. During S. typhimurium in
fection in mice, the circulating TNFa concentration was increased on day 7, as well as
the stimulated production of IL-la, IL-Iß and TNFa in whole blood. Production of IL-
lß and TNFa by peritoneal and bone marrow cells was also increased during infection.
This general upregulation of proinflammatory cytokine production in this animal model
is in contrast with what we have found in humans with typhoid fever [2]. In these
patients, downregulation of LPS-stimulated production of cytokines in whole blood was
found. These differences seem not to be caused by the time point of measuring cytokine
production, as the patients with typhoid fever had also been ill for a week. More early
down-regulation of cytokine production was shown in patients with Gram-negative
infection such as meningococcemia [18]. It is obviously impossible to draw conclusions
from these animal experiments and extrapolate the results to the human situation. In
conclusion, rhIL-la pretreatment decreased outgrowth of S. typhimurium, partly by a
modest influence on peritoneal cell cytokine production.
References
1. Van den Bergh ET AM, Hussein Gasem M, Keuter M, Dolmans WMV. Outcome in
three groups of patients with typhoid fever treated in Indonesia between 1948 and
1990. 1998; submitted.
2. Keuter M, Dharmana E, Gasem MH, Van der Ven-Jongekrijg J, Djokomoeljanto R,
Dolmans WMV, Demacker PMN, Sauerwein R, Gallati H, Van der Meer JWM.
Patterns of proinflammatory cytokines and inhibitors during typhoid fever. J Infect
Dis 1994; 169: 1306-1311.
3. Jotwani R, Tanaka Y, Watanabe K, Tanaka K, Kato N, Ueno К. Cytokine stimula
tion during Salmonella typhimurium sepsis in ItyS mice. J Med Microbiol 1995;
42: 348-52.
4. Kullberg BJ, Van 't Wout JW, Van Fürth R. Role of granulocytes in enhanced host

rIL-1 in S. typhimurium infection 79
resistance to Candida albicans induced by recombinant interleukin-1. Infect Im
mun 1990;58:3319-3324.
5. Czuprynski CJ, Brown JF. Recombinant murine interleukin-la enhancement of
nonspecific antibacterial resistance. Infect Immun 1987; 55: 2061-2065.
6. Van der Meer JWM, Barza M, Wolff SM, Dinarello CA. A low dose of recom
binant interleukin 1 protects granulocytopenic mice from lethal Gram-negative in
fection. Proc Natl Acad Sci USA 1988; 85: 1620-1623.
7. Curfs JHAJ, van der Meer JWM, Sauerwein R, Eling WMC. Low dosages of inter
leukin-1 protect mice against lethal cerebral malaria. In: Dinarello CA, Kluger M,
Oppenheim J, Powanda M, ed. Interleukin-1 and related cytokines. New York:
AlanLiss, 1990.
8. Vogels MTE, Mensink EJBM, Ye K, Boerman OC, Verschuren CMM, Dinarello
CA, Van der Meer JWM. Differential gene expression for IL-1 receptor antagonist,
IL-1, and TNF receptors and IL-1 and TNF synthesis may explain IL-1-induced
resistance to infection. J Immunol 1994; 153: 5772-5780.
9. Morrissey PJ, Charrier К. Treatment of mice with IL-1 before infection increases
resistance to a lethal challenge with Salmonella typhimurium - the effect correlates
with the resistance allele at the ity locus. J Immunol 1994; 153: 212-219.
10. Mastroeni P, Skepper JN, Hormaeche CE. Effect of anti-tumor necrosis factor
alpha antibodies on histopathology of primary Salmonella infections. Infect Immun
1995; 63: 3674-3682.
11. Nauciel C, Espinasse-Maes F. Role of gamma-interferon and tumor necrosis factor
alpha in resistance to Salmonella typhimurium infection. Infect Immun 1992; 60:
450-454.
12. Drenth JPH, Van Uum SHM, Van Deuren M, Pesman GJ, Van der Ven-Jongekrijg
J, Van der Meer JWM. Endurance run increases circulating IL-6 and IL-Ira but
downregulates ex vivo TNF-oc and IL-lß production. J Appi Physiol 1995; 79:
1497-1503.
13. Kullberg BJ, Van 't Wout JW, Van Fürth R. No effect of recombinant human inter
leukin-1 on the numbers of peripheral blood and peritoneal leukocytes during an
acute inflammation. Inflammation 1991; 15:457-470.
14. Keuter M, Dharmana E, Kullberg BJ, Curfs J, Djokomoeljanto R, Van der Meer
JWM. G-CSF enhances survival of Salmonella typhimurium infection in CBA/J
mice, submitted 1998;
15. Dinarello CA, Ikejima T, Warner SJC, Orencole SF, Lonnemann G, Cannon JG,
Libby P. Interleukin 1 induces interleukin 1.1. Induction of interleukin 1 in rabbits

80 Chapter 6
in vivo and in human mononuclear cells in vitro. J Immunol 1987; 139:1902-1910.
16. Nakano Y, Onozuka К, Terada Y, Shinomiya H, Nakano M. Protective effect of re
combinant tumor necrosis factor-α in murine salmonellosis. J Immunol 1990; 144:
1935-1941.
17. Vogels MTE, Van Rooyen N, Bemelmans MHA, Jansen M, Van der Meer JWM.
Interleukin-1 (IL-l)-induced resistance to bacterial infection: role of the macropha
ge and soluble TNF receptors. In: Vogels MTE, ed. Mechanisms of enhanced natu
ral resistance to infection induced by interleukin-1. Thesis: Catholic University
Nijmegen, 1994.
18. Van Deuren M, Van der Ven-Jongekrijg J, Bartelink AKM, Van Dalen R,
Sauerwein RW, Van der Meer JWM. Correlation between proinflammatory cyto
kines and antiinflammatory mediators and the severity of disease in meningococcal
infections. J Infect Dis 1995; 172: 433-439.

Chapter 7
G-CSF enhances survival of CBA/J mice with Salmonella typhimurium infection
Monique Keuter, Edi Dhaimana, Bart-Jan Kullberg, Jo H.A.J. Curfs
and Jos W.M. van der Meer
Submitted

82 Chapter 7
Abstract
This study shows that recombinant murine granulocyte colony-stimulating factor
(rmG-CSF) improves survival of CBA/J mice with a potentially lethal Salmonella
typhimurium infection. Of the control mice, 8% survived, compared with 33% survival
in mice treated with a single dose of rmG-CSF one day before infection (p < .0001).
The outgrowth of S. typhimurium in liver, spleen, bone marrow, lung, brain, kidney and
heart on day 1, 3 and 5 was not different between the groups. Circulating cytokine con
centrations did not differ between the groups either. One day after of rmG-CSF treat
ment, the lipopolysaccharide (LPS) stimulated tumor necrosis factor-α (TNFa) produc
tion of resident peritoneal macrophages was significantly higher in treated mice: 1.125
± 0.476 ng/mL vs 0.621 ± 0.173 ng/mL in control mice (p < .01). Histological
examination of the liver on day 1 and 5 showed more localized inflammatory foci in the
rmG-CSF treated animals. These results show that activation of neutrophils by rmG-
CSF is beneficial in the facultative intracellular S. typhimurium infection in mice.

rG-CSF in S. typhimurium infection 83
Introduction
Salmonella infections, especially typhoid fever, remain significant causes of mor
bidity and mortality in tropical countries. More insight in the host response to Sal
monella is necessary to develop more effective therapeutic strategies, e.g. immunother
apy, which might improve the outcome of infection. Patients with typhoid fever usually
present with a relative leukopenia. In these patients, we found a correlation between
leukopenia and the development of complications (unpublished results). These observa
tions may suggest that granulocytes play a role in the host defense against Salmonella
infection in man.
In experimental S. typhimurium infection in mice, the bacteria induce a disease
similar to human typhoid fever. The survival of mice during the first days of infection
with the facultative intracellular S. typhimurium , before the development of specific
acquired immunity, depends on natural immunity. Neutrophils are thought to play an
important part in this phase of S. typhimurium infection in mice. Earlier studies have
shown that liver and spleen of mice infected with Salmonella typhimurium contained
many polymorphonuclear phagocytes (PMN's) and microabcesses after 24 hours of in
fection. Hardly any macrophages were found at that time point [1-3]. Blocking the mi
gration of PMN's through administration of mAb to leukocyte adhesion molecules,
lethally exacerbated the infection [1]. In addition, neutropenic mice developed a lethal
systemic salmonellosis after infection with low numbers of microorganisms [2].
Granulocyte colony-stimulating factor (G-CSF) is a hematopoietic growth factor,
which stimulates proliferation and activation of neutrophils and their precursors [4].
The effects on mature neutrophils include enhanced phagocytosis and respiratory burst
as well as reduced apoptosis. rG-CSF treatment has been shown to be beneficial in
many neutropenic animal models, but also in infections with non-neutropenic animals
[5]. In a peritonitis model in non-neutropenic mice, rG-CSF was highly effective in
improving survival [6, 7].
In models of intracellular infection, experience with rG-CSF is scarce, and most
studies have been performed with human rG-CSF, of which the activity is considered
sub optimal for mice. The present study reports survival, outgrowth of microorganisms,
and cytokines in CBA/J mice with a potentially lethal S. typhimurium infection after
treatment with murine rG-CSF (rmG-CSF), which has recently become available.

84 Chapter 7
Materials and methods
Animals
Specific pathogen-free female Ity resistant (ItyR) CBA/J mice, weighing 20-25 g
were used. The course of Salmonella infections in mice is regulated by the Ity locus on
chromosome 1, which most likely encodes for the so-called natural resistance associa
ted membrane protein 1 (Nrampl) on professional phagocytes [8-10]. The CBA/J mice
belong to the ItyR, or Salmonella resistant mice.
Animals were housed under standard laboratory conditions and fed sterilized labo
ratory chow (Hope Farms, Woerden, The Netherlands) and water ad libitum. The expe
riments were approved by the ethical committee for animals experiments at the Catholic
University Nijmegen.
Materials
Recombinant murine G-CSF, a gift from Amgen (Thousand Oaks, CA)was diluted
in pyrogen-free saline to a final concentration of 2.5 μ^πύ.
Salmonella typhimurium infection
A serum-resistant strain of S. typhimurium (phage type 510) was grown to statio
nary phase culture by overnight incubation at 37°C in nutrient broth (BHI Oxoid). The
LD50 in the ItyR CBA/J mice was 0.5 χ IO4 bacteria. Mice were injected i.p. with 1 χ
IO4 cfu of S. typhimurium. rmG-CSF was given as a s.c. injection of 500 ng (25 |ig/kg)
in 100 mL of pyrogen free saline. One group of mice received 500 ng s.c. 24 hours
before the infection (Pretreatment group). The multiple treatment group received 500
ng s.c. once daily from 24 hours before infection until day 7 during infection. Control
mice received pyrogen free saline s.c. daily. Survival was assessed twice daily for 21
days in groups of at least 20 animals.
On day 1, 3 or 5 after the injection of S. typhimurium, mice were killed by cervical
dislocation and outgrowth of the microorganisms from the liver, spleen and bone
marrow was quantified in groups of at least 8 mice. For this purpose, the liver and
spleen were removed aseptically, weighed, and homogenized in sterile saline in a tissue
grinder. Bone marrow was flushed from the femur aseptically with 1 mL of sterile
saline. In 4 mice per group, outgrowth in lung, heart, kidney and brain was also quanti
fied. The number of viable salmonellae was determined by plating serial dilutions on
Brilliant Green agar (BGA) plates and cfu were counted after overnight incubation at
37°C. The results were expressed as the log cfu per gram of tissue.

iG-CSF in S. typhimuríum infection 85
Cytokine studies
Cytokine measurements
On days 0,1,3 or 5, separate groups of mice (5 mice per group) were anesthetized
with ether to collect blood from the intraorbital plexus for the measurement of circu
lating cytokine concentrations. TNFa, IL-la and IL-lß concentrations were determined
using specific radioimmunoassays (RIA) developed in our laboratory, as previously
described [11].
Ex-vivo cytokine production
On day 0 (24h after rmG-CSF injection) and on day 3 after infection, resident peri
toneal macrophages of mice were harvested by rinsing the peritoneal cavity aseptically
with 2 ml cold PBS containing 0.38% (w/v) sodium citrate. After centrifugation, cells
were resuspended in RPMI1640 (Dutch modification; Flow Laboratories, Irvine, UK),
containing 1 mM pyruvate, 2 mM L-glutamine and 100 μg of gentamicin per mL, and
105 cells /well were incubated for 1 h at 37°C in 96-wells microtiter plates (Costar
Corporation, Cambridge, MA). The non-adherent cells were discarded and the remai
ning cell population consisted of more than 90% macrophages, as assessed by light
microscopy. LPS (E. coli serotype 055:B5; Sigma, St Louis, USA; final concentration
10μg/mL) was added to the peritoneal macrophages to a final volume of 100 μ ι and
incubated at 37°C in 5% C02. After 24h, the supematants were collected and stored at
-70°C until assayed. To the remaining macrophages, 200 μ ι of RPMI were added and
the cells were disrupted by three freeze-thaw cycles to determine the cell-associated
cytokine contents. The samples were stored at -70°C until assayed for TNFa, IL-la and
IL-lß concentrations as described above.
Blood samples
Blood samples (20 \iL) were collected from the retroorbital plexus and the leuko
cytes were counted in a Coulter Counter (Coulter Electronics, Luton, England). The
total numbers of granulocytes, lymphocytes and monocytes per mm3 were calculated
from the total numbers of leukocytes per mm3 and the differential counts of 200 leuko
cytes in 2 Giemsa-stained blood smears.
Histology
From other groups of animals, the liver was removed, fixed in buffered formalin
(4%), and serial sections were examined microscopically after staining with hematoxy
lin and eosin.
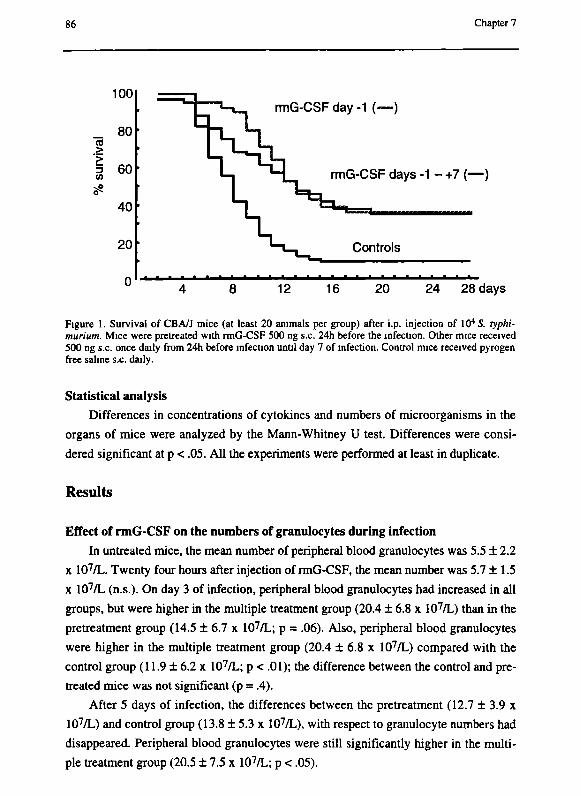
86 Chapter 7
100
_ 80 л .> i 60 (Я
40 •
20 '
rmG-CSF day -1 (—)
rmG-CSF days -1 - +7 (—)
Controls
12 16 20 24 28 days
Figure 1. Survival of СВАЛ mice (at least 20 animals per group) after i.p. injection of 104 S. typhi-murium. Mice were pretreated with rmG-CSF 500 ng s.c. 24h before the infection. Other mice received 500 ng s.c. once daily from 24h before infection until day 7 of infection. Control mice received pyrogen free saline s.c. daily.
Statistical analysis
Differences in concentrations of cytokines and numbers of microorganisms in the
organs of mice were analyzed by the Mann-Whitney U test. Differences were consi
dered significant at ρ < .05. All the experiments were performed at least in duplicate.
Results
Effect of rmG-CSF on the numbers of granulocytes during infection
In untreated mice, the mean number of peripheral blood granulocytes was 5.5 ± 2.2
χ 107/L. Twenty four hours after injection of rmG-CSF, the mean number was 5.7 ±1.5
χ 107/L (n.s.). On day 3 of infection, peripheral blood granulocytes had increased in all
groups, but were higher in the multiple treatment group (20.4 ± 6.8 χ 107/L) than in the
pretreatment group (14.5 ± 6.7 χ 107/L; ρ = .06). Also, peripheral blood granulocytes
were higher in the multiple treatment group (20.4 ± 6.8 χ 107/L) compared with the
control group (11.9 ± 6.2 χ 107/L; ρ < .01); the difference between the control and pre
treated mice was not significant (p = .4).
After 5 days of infection, the differences between the pretreatment (12.7 ± 3.9 χ
107/L) and control group (13.8 ± 5.3 χ 107/L), with respect to granulocyte numbers had
disappeared. Peripheral blood granulocytes were still significantly higher in the multi
ple treatment group (20.5 ± 7.5 χ 107/L; ρ < .05).
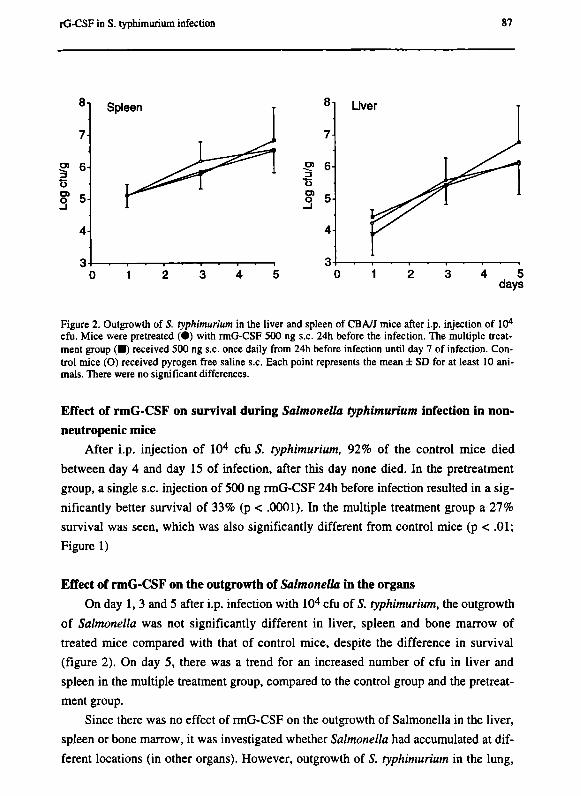
rG-CSF in S. typhimuríum infection 87
!> 6 Ь Q 5
Spleen 8-1
7-
3
t5 σι _
3 5
4
3
Liver τ
^À , ^
^ l Г ^
Γ" • 1 » Π 1 — — I — · — I
4 5 days
Figure 2. Outgrowth of S. typhimurium in the liver and spleen of CBA/J mice after i.p. injection of 104
cfu. Mice were pretreated ( · ) with rmG-CSF 500 ng s.c. 24h before the infection. The multiple treatment group (•) received 500 ng s.c. once daily from 24h before infection until day 7 of infection. Control mice (O) received pyrogen free saline s.c. Each point represents the mean ± SD for at least 10 animals. There were no significant differences.
Effect of rmG-CSF on survival during Salmonella typhimuríum infection in non-
neutropenic mice
After i.p. injection of IO4 cfu S. typhimurium, 92% of the control mice died
between day 4 and day 15 of infection, after this day none died. In the pretreatment
group, a single s.c. injection of 500 ng rmG-CSF 24h before infection resulted in a sig
nificantly better survival of 33% (p < .0001). In the multiple treatment group a 27%
survival was seen, which was also significantly different from control mice (p < .01;
Figure 1)
Effect of rmG-CSF on the outgrowth of Salmonella in the organs
On day 1, 3 and 5 after i.p. infection with 104 cfu of S. typhimurium, the outgrowth
of Salmonella was not significantly different in liver, spleen and bone marrow of
treated mice compared with that of control mice, despite the difference in survival
(figure 2). On day 5, there was a trend for an increased number of cfu in liver and
spleen in the multiple treatment group, compared to the control group and the pretreat
ment group.
Since there was no effect of rmG-CSF on the outgrowth of Salmonella in the liver,
spleen or bone marrow, it was investigated whether Salmonella had accumulated at dif
ferent locations (in other organs). However, outgrowth of S. typhimurium in the lung,

88 Chapter 7
heart, kidney and brain was similar in control and rmG-CSF treated animals (data not
shown).
Circulating cytokines Circulating concentrations of TNFct, IL-la and IL-Iß were determined on day 0
before infection (24 hours after rmG-CSF) and day 1, 3 and 5 after infection of 104 cfu
S. typhimurium. Before infection and on day 1 of infection, cytokine concentrations in
all samples were under the detection limits. No detectable concentrations of IL-Iß (<
0.020 ng/mL) were found at any time point during the infection.
On day 3, TNFa was 0.109 + 0.061 ng/mL (mean ± SD) in rmG-CSF pretreated
mice, 0.115 ± 0.075 ng/mL in the multiple treatment group and 0.127 ± 0.105 ng/mL in
control mice (n.s.). The IL-la concentrations were not significantly different in the
three groups either: 0.067 ± 0.067 ng/mL in pretreated mice, 0.049 ± 0.025 ng/mL in
the multiple treatment group and 0.062 ± 0.039 ng/mL in control mice.
On day 5, circulating concentrations of TNFa and IL-la were higher, but not
significantly different in rmG-CSF multiple dose treated groups than in control mice:
for TNFa 0.323 ± 0.232 ng/mL vs 0.264 ± 0.160 ng/mL (n.s.) and for IL-la 0.093 ±
0.085 ng/mL in the multiple treatment group vs 0.070 ± 0.042 ng/mL in the control
mice (n.s.).
Ex-vivo cytokine production To investigate whether a difference in cytokine production of macrophages after
rmG-CSF could explain the difference in outcome, resident peritoneal macrophages of
the groups of mice were harvested before infection (24h after rmG-CSF administration)
and on day 3 of infection, and cells were stimulated with LPS in vitro.
Before infection, LPS stimulated TNFa concentrations in supematants of macro
phages were significantly higher in the rmG-CSF pretreated group than in controls (p <
.01). On the same day, cell-associated IL-la production tended to be lower in the rmG-
CSF treated group than in controls (n.s.). IL-Iß concentrations in supematants of
macrophages were not different in the 2 groups (Figure 3).
On day 3 during infection, TNFa concentrations in supematants of macrophages
were not significantly higher in the rmG-CSF pretreated group (2.856 ± 2.252 ng/mL)
and multiple treatment group (3.529 ± 2.964 ng/mL) than in controls (2.112 ± 0.897
ng/mL).
Cell-associated IL-la production tended to be higher (but did not reach signifi
cance) in the rmG-CSF pretreated (2.593 ± 1.241 ng/mL) and multiple treatment group
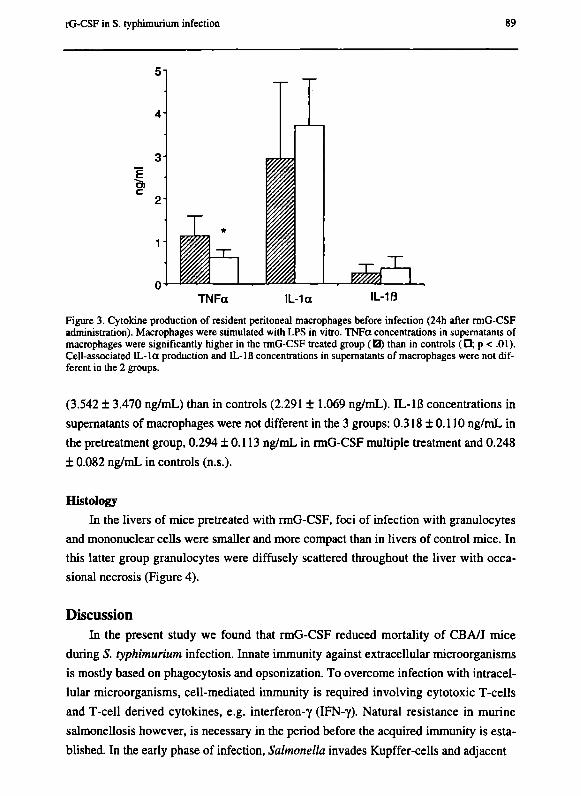
rG-CSF in S. typhimurium infection 89
51
Ш ém A l TNFa IL-1a IL-1B
Figure 3. Cytokine production of resident peritoneal macrophages before infection (24h after rmG-CSF administration). Macrophages were stimulated with LPS in vitro. TNFa concentrations in supematants of macrophages were significantly higher in the rmG-CSF treated group (И) than in controls (•; ρ < .01). Cell-associated IL-la production and IL-IB concentrations in supematants of macrophages were not different in the 2 groups.
(3.542 ± 3.470 ng/mL) than in controls (2.291 ± 1.069 ng/mL). IL-Iß concentrations in
supematants of macrophages were not different in the 3 groups: 0.318 ±0.110 ng/mL in
the pretreatment group, 0.294 ± 0.113 ng/mL in rmG-CSF multiple treatment and 0.248
± 0.082 ng/mL in controls (n.s.).
Histology
In the livers of mice pretreated with rmG-CSF, foci of infection with granulocytes
and mononuclear cells were smaller and more compact than in livers of control mice. In
this latter group granulocytes were diffusely scattered throughout the liver with occa
sional necrosis (Figure 4).
Discussion In the present study we found that rmG-CSF reduced mortality of CBA/J mice
during S. typhimurium infection. Innate immunity against extracellular microorganisms
is mostly based on phagocytosis and opsonization. To overcome infection with intracel
lular microorganisms, cell-mediated immunity is required involving cytotoxic T-cells
and T-cell derived cytokines, e.g. interferon-γ (IFN-γ). Natural resistance in murine
salmonellosis however, is necessary in the period before the acquired immunity is esta
blished. In the early phase of infection, Salmonella invades Kupffer-cells and adjacent

90 Chapter 7
Suf' І * I ^* .* "I"*-'¡ г
Figure 4. Histology of the liver of mice pretreated with rmG-CSF (A magnification χ 10; В magnifica
tion χ 40) and control mice (C magnification χ 10; D magnification χ 40). In rmG-CSF treated mice,
more granulocytes can be seen in compact foci in the liver.

rG-CSF in S. typhimurìum infection 91
hepatocytcs [1]. Neutrophils are important for the killing of Salmonella in that stage.
Blocking the migration of these cells through administration of mAb to leukocyte adhe
sion molecules such as CD lib/CD 18 in a sublethal S. typhimurìum infection in mice,
resulted in extracellular outgrowth of salmonellae in the hepatic sinusoids and in a fail
ure to destroy Salmonella-lozded hepatocytes, leading to a lethal infection [12]. Natural
killer (NK) cells and Τ lymphocytes are not involved in the lysis of hepatocytes. There
fore it appears, that neutrophils play an important role in the early host defense against
Salmonella infection. Enhancing the recruitment or activation of neutrophils by rG-CSF
may be helpful at this stage of the infection.
In the present study, administration of rmG-CSF had a beneficial effect on survi
val, but not on the outgrowth of Salmonella in the organs. In general the numbers of
Salmonella cfu in the liver and the spleen correlate with mortality in experimental S.
typhimurìum infection in mice [13-15], however the number of bacteria may not reflect
the extent of organ destruction. In the rmG-CSF treated animals we have seen that mi
croorganisms were contained in a granuloma, whereas there was more diffuse granulo
cytosis and necrosis in the control group.
Although rmG-CSF had increased the granulocytes, this was not the only reason
for the difference in survival on day 5, as the number of PMN's were highest in the mul
tiple treatment group and similar in the control and pretreated mice at that time point.
Apart from its effect on neutrophil function, G-CSF has antiinflammatory properties
and has been shown to be able to influence TNFa production [16, 17]. In this respect, it
is important to note that in intracellular infection, the proinflammatory cytokines play a
beneficial role, partly through activation of macrophages, resulting in better survival
[18]. Inhibition of TNFa during Salmonella infection in mice has been shown to wor
sen outcome [19, 20]. We found that the LPS-stimulated TNFa production by peri
toneal macrophages was increased 24 hours after s.c. administration of rmG-CSF.
Granulocyte-macrophage colony-stimulating factor (GM-CSF) has been known to indu
ce fever as a side-effect, which possibly is associated with its ability to induce produc
tion of TNFa by macrophages in vitro [21]. In contrast, G-CSF, which is supposed not
to induce TNFa is not associated with fever. Moreover, G-CSF protected against a
lethal injection of LPS in rodents [22]. In human volunteers pretreated with rG-CSF,
endotoxin infusion increased the peak of circulating TNFa [23]. However, it was found
that ex vivo production of TNFa in human blood was lower after G-CSF treatment [16,
17]. Apparently, differences in species (man-mouse-rat), strains, time points and way of
delivery of G-CSF can play a role in these discrepancies.
In previous studies we have shown that the clinical picture and pattern of circula-

92 Chapter 7
ting cytokines of typhoid fever in humans differs from that of Gram-negative sepsis or
endotoxinemia [24]. Also in animal models, TNFoc was detectable in the circulation
only several days after S. typhimurium infection, whereas TNFa did rise 1 hour after
LPS administration in these animals [25, 26] .The circulating TNFa concentrations
found in our previous studies on murine salmonellosis almost always seemed to be a re
sult of the outgrowth of microorganisms in the tissues [27].
It could have been envisaged that the increased local TNFa production, as we have
found in our experiments, may have led to increased activation of macrophages and
subsequently to a reduction of the outgrowth in the organs. However, we only found
significantly increased TNFa production 24 hours after administration of rmG-CSF and
not at later time points during the infection. In contrast, TNFa plays a role especially
after 4-5 days of Salmonella infection [19]. Besides, cytokine production of peritoneal
macrophages might not be representative for production by macrophages in the liver.
It may be questioned whether rG-CSF has a beneficial effect on intracellular infec
tions through enhanced (early) neutrophil recruitment, whereas the (late) antiinflamma
tory properties of rG-CSF may be deleterious, through inhibition of TNFa mediated
macrophage activation. The trend for an increased number of cfu in liver and spleen in
the multiple treatment group could be an illustration of this suggestion and may be due
to such an adverse late effect. Pretreatment alone seems beneficial as has been shown in
Listeria monocytogenes infection in mice, in which pretreatment with high dose human
rG-CSF during 5 days, reduced mortality as well as outgrowth of microorganisms [28,
29].
In conclusion, rmG-CSF protects mice against a lethal 5. typhimurium infection in
mice, although no effect was found on the outgrowth of microorganisms in the organs.
Further studies are needed to elucidate the mechanism, afore rG-CSF can be used as im
munotherapy in Salmonella infection.
References 1. Conlan JW, North RJ. Early pathogenesis of infection in the liver with the faculta
tive intracellular bacteria Listeria monocytogenes, Francisella tularensis, and
Salmonella typhimurium involves lysis of infected hepatocytes by leukocytes.
Infect Immun 1992; 60: 5164-5171.
2. Conlan JW, North RJ. Neutrophil-mediated lysis of infected hepatocytes. ASM
News 1993; 59: 563-567.
3. Hsu HS. Pathogenesis and immunity in murine salmonellosis. Microbiol Rev 1989;
53: 390-409.

rG-CSF in S. typhimurium infection 93
4. Dale DC, Liles WC, Summer WR, Nelson S. Review: Granulocyte colony-stimula
ting factor. Role and relationships in infectious diseases. J Infect Dis 1995; 172:
1061-1075.
5. Kullberg BJ, Netea MG, Curfs JHAJ, Keuter M, Meis JFGM, Van der Meer JWM.
Recombinant murine granulocyte colony-stimulating factor protects against acute
disseminated Candida albicans infection in non-neutropenic mice. J Infect Dis
1998; 177: 175-181.
6. Nelson S. Role of granulocyte colony-stimulating factor in the immune response to
acute bacterial infection in the nonneutropenic host: an overview. Clin Infect Dis
1994;18(suppl2):S197-204.
7. Barsig J, Bundschuh DS, Härtung Τ, Bauhofer Α, Sauer A, Wendel A. Control of
fecal peritoneal infection in mice by colony-stimulating factors. J Infect Dis 1996;
174: 790-799.
8. Vidal S, Tremblay ML, Govoni G, Gauthier S, Sebastiani G, Malo D, Skamene E,
Olivier M, Jothy S, Gros P. The Ity/Lsh/Bcg locus: Natural resistance to infection
with intracellular parasites is abrogated by disruption of the Nrampl gene. J Exp
Med 1995; 182: 655-666.
9. Vidal S, Lepage Ρ, Gauthier S, Gros P. Natural resistance to intracellular infec
tions. J of Imm 1996; 157: 3559-3568.
10. Govoni G, Vidal S, Gauthier S, Skamene E, Malo D, Gros P. The BcgЯty/Lsh
locus: Genetic transfer of resistance to infections in C57B1/6J mice transgenic for
the N r a m p l G l y 1 6 9 allele. Infect Immun 1996; 64: 2923-2929.
11. Drenth JPH, Van Uum SHM, Van Deuren M, Pesman GJ, Van der Ven-Jongekrijg
J, Van der Meer JWM. Endurance run increases circulating IL-6 and IL-lra but
downregulates ex vivo TNF-a and IL-Iß production. J Appi Physiol 1995; 79:
1497-1503.
12. Conlan JW. Neutrophils prevent extracellular colonization of the liver microvascu-
lature by Salmonella typhimurium. Infect Immun 1996; 64: 1043-1047.
13. Morrissey PJ, Charrier К. Interleukin-1 administration to СЗН/HeJ mice after but
not prior to infection increases resistance to Salmonella typhimurium. Infect Immun
1991;59:4729-4731.
14. Morrissey PJ, Charrier К. Treatment of mice with IL-1 before infection increases
resistance to a lethal challenge with Salmonella typhimurium - the effect correlates
with the resistance allele at the ity locus. J Immunol 1994; 153: 212-219.
15. Van Dissel JT, Stikkelbroeck JJM, Sluiter W, Leijh PCJ, Van Fürth R. Differences
in initial rate of intracellular killing of Salmonella typhimurium by granulocytes of

94 Chapter 7
salmonella susceptible C57B1/10 mice and salmonella resistant CBA mice. J
Immunol 1986; 136: 1074-1080.
16. Kitabayashi A, Hirokawa M, Hatano Y, Lee M, Kuroki J, Niitsu H, Miura AB.
Granulocyte colony-stimulating factor dowmegulates allogeneic immune responses
by posttranscriptional inhibition of tumor necrosis factor-alpha production. Blood
1995; 86: 2220-2227.
17. Härtung Τ, Docke WD, Gantner F, Krieger G, Sauer A, Stevens Ρ, Volk HD,
Wendel Α. Effect of granulocyte colony-stimulating factor treatment on ex vivo
blood cytokine response in human volunteers. Blood 1995; 85: 2482-2489.
18. Nakano Y, Onozuka К, Terada Y, Shinomiya H, Nakano M. Protective effect of
recombinant tumor necrosis factor-α in murine salmonellosis. J Immunol 1990;
144: 1935-1941.
19. Mastroeni P, Skepper JN, Hormaeche CE. Effect of anti-tumor necrosis factor
alpha antibodies on histopathology of primary Salmonella infections. Infect Immun
1995; 63: 3674-3682.
20. Nauciel C, Espinasse-Maes F. Role of gamma-interferon and tumor necrosis factor
alpha in resistance to Salmonella typhimurium infection. Infect Immun 1992; 60:
450-454.
21. Sisson SD, Dinarello CA. Production of interleukin-1 alpha, interleukin-1 beta and
tumor necrosis factor by human mononuclear cells stimulated with granulocyte-
macrophage colony-stimulating factor. Blood 1988; 72: 1368-1374.
22. Gorgen I, Härtung Τ, Leist M, Niehorster M, Tiegs G, Uhlig S, Weitzel F, Wendel
A. Granulocyte colony-stimulating factor treatment protects rodents against lipo-
polysaccharide-induced toxicity via suppression of systemic tumor necrosis factor.
J Immunol 1992; 149: 918-924.
23. Pajkrt D, Manten A, Van der Poll T, Van Buul TMM, Jansen J, Ten Cate WJ, Van
Deventer SJ. Modulation of cytokine release and neutrophil function by granulo
cyte colony-stimulating factor during endotoxemia in humans. Blood 1997; 90:
1415-1424.
24. Keuter M, Dharmana E, Gasem MH, Van der Ven-Jongekrijg J, Djokomoeljanto R,
Dolmans WMV, Demacker PMN, Sauerwein R, Gallati Η, Van der Meer JWM.
Patterns of proinflammatory cytokines and inhibitors during typhoid fever. J Infect
Dis 1994; 169: 1306-1311.
25. Peel JE. Induction of circulating tumor necrosis factor cannot be demonstrated
during septicemic salmonellosis in calves. Infect Immun 1990; 58:439-42.
26. Jotwani R, Tanaka Y, Watanabe K, Tanaka K, Kato N, Ueno К. Cytokine stimula-

rG-CSF in S. typhimurium infection 95
tion during Salmonella typhimurium sepsis in ItyS mice. J Med Microbiol 1995;
42: 348-52.
27. Keuter M, Netea MG, Kullberg BJ, Van Dissel JT, Demacker PNM, Stalenhoef
AFH, Van der Meer JWM. Hyperlipoproteinemia protects low-density-lipoprotein
receptor deficient mice against lethal Salmonella typhimurium infection. 1998;
submitted.
28. Serushago BA, Yoshikai Y, Handa T, Mitsuyama M, Muramori K, Nomoto K.
Effect of recombinant human granulocyte colony-stimulating factor (rh G-CSF) on
murine resistance to Listeria monocytogenes. Immunology 1992; 75: 475-480.
29. Shinomiya N, Tsuru S, Katsura Y, Kayashima S, Nomoto K. Enhanced resistance
against Listeria monocytogenes achieved by pretreatment with granulocyte colony-
stimulating factor. Infect Immun 1991; 59: 4740-4743.

96 Chapter 7

Chapter 8
Hyperlipoproteinemia protects low-density-lipoprotein receptor deficient mice against lethal Salmonella typhimurium infection
Monique Keuter, Minai G. Netea, Bart-Jan Kullberg, Jaap T. van Dissel, Pierre N.M.
Demacker, Anton F.H. Stalenhoef and Jos W.M. van der Meer
Submitted

98 Chapter 8
Abstract
Lipoproteins improve the outcome of severe Gram-negative infections through
neutralizing lipopolysaccharides (LPS) and thus reducing the induction of proinflam
matory cytokines. During a lethal Salmonella typhimurium infection in C57B1/6J mice,
comparing low density lipoprotein receptor deficient (LDLR-/-) and LDLR+/+ animals,
survival in LDLR-/- mice was greater when compared with LDLR+/+ (100% vs 5%, ρ
< .001). The outgrowth of S. typhimurium in the liver, spleen and bone marrow of
LDLR-/- mice was 100-1000 times lower than in controls on day 3 and 7 after infec
tion, with lower circulating cytokine concentrations. Upon intravenous high dose Sal
monella infection of LDLR+/+ mice, the bacteria disappeared from the blood within 30
minutes with rapid invasion of liver and spleen. In contrast, in LDLR-/- mice, Salmo
nella remained in the circulation for up to 2h, with a significantly decreased (70-80%)
organ count. Distribution differences appear to be a reason for improved survival to
salmonellosis of mice with hyperlipoproteinemia.

S. typhimurium in LDLR-/- mice 99
Introduction
Salmonella infections, especially typhoid fever, pose important public health pro
blems in tropical countries. Despite preventive measures such as hygiene and vaccina
tion, and the availability of antibiotics, typhoid fever remains a significant cause of
morbidity and mortality. To develop more effective therapeutic strategies, e.g. immuno
therapy, more insight in the host response to Salmonella is necessary.
The role of endotoxin in typhoid fever has been debated [1]. Although salmonellae
contain LPS, the clinical picture and pattern of circulating cytokines of typhoid fever in
humans differs from that in other Gram-negative sepsis [2]. Also in animal models of
typhoid fever, TNFcc was undetectable in the circulation until several days after 5.
typhimurium infection, whereas TNFa did rise 1 hour after LPS administration or extra
cellular Gram-negative infection in these animals [3,4]. salmonellae behave as faculta
tive intracellular parasites. It has been shown that in models of intracellular infection,
the proinflammatory cytokines play a beneficial role, resulting in better survival [5] and
inhibition of TNFa during Salmonella infection has worsened outcome [6, 7].
Circulating lipoproteins are able to bind and neutralize bacterial lipopolysaccha-
rides (LPS), thereby reducing its toxic properties by decreasing the induction of proin
flammatory cytokines such as IL-1 and TNFa [8,9]. In low density lipoprotein receptor
deficient (LDLR-/-) mice, the loss of the receptor results in а 7 to 9 times higher LDL-
cholesterol level than in their wild-type LDLR+/+ littermates. Our group has shown
that LDLR-/- mice survive longer and have lower proinflammatory cytokine concen
trations compared to control mice after bacterial endotoxin injection and after infection
with Klebsiella pneumoniae [10].
We realized that, although hyperlipoproteinemic LDLR-/- mice are able to with
stand extracellular Gram-negative infections like with Klebsiella pneumoniae, the sus
ceptibility to Salmonella infection would be more complex to predict in view of the
protective role of TNFa and the limited role of Salmonella endotoxin in these infec
tions. In the present study, we have investigated the susceptibility of LDLR knock out
mice to S. typhimurium.
Methods
Animals
Homozygous C57B1/6J mice lacking low density lipoprotein receptors (LDLR-/-)
and their wild-type littermates (LDLR+/+) were obtained from Jackson Laboratory (Bar

100 Chapter 8
Harbour, ME) as mating pairs and bred in our local facility. For the experiments, 6-8
weeks old mice, weighing 20-25 grams were used. The animals were fed standard labo
ratory chow (Hope Farms, Woerden, The Netherlands) and housed under specific
pathogen free conditions. The experiments were approved by the ethical committee for
animal experiments at the Catholic University Nijmegen.
Salmonella typhimurium infection
A serum-resistant strain of 5. typhimurium (phage type 510) was grown to statio
nary phase culture by overnight incubation at 37°C in nutrient broth (BHI Oxoid). The
LD50 after either intraperitoneal (i.p.) or intravenous (i.v.) injection into C57B1/6J
(LDLR+/+) mice was < 102 bacteria. Mice were injected i.v. with 1 χ IO2 cfu of 5.
typhimurium. Survival was assessed daily for 21 days in groups of at least 20 animals.
On days 1, 3 and 7, separate groups of mice were anesthetized with ether to collect
blood from the intraorbital plexus for the measurement of circulating cytokine concen
trations. On day 1, 3 and 7 after the injection of S. typhimurium, mice were killed by
cervical dislocation and outgrowth of the microorganisms from the liver, spleen and
bone marrow was quantified. For this purpose, the liver and spleen were removed asep-
tically, weighed, and homogenized in sterile saline in a tissue grinder. Bone marrow
was flushed from the femur aseptically with 1 ml of sterile saline. The number of viable
salmonellae was determined by plating several dilutions on Brilliant Green agar (BGA)
plates and cfu were counted after overnight incubation at 37°C. The results were ex
pressed as the log cfu per gram of tissue. Previous experiments had shown that i.p. and
i.v. injection of the microorganisms led to an identical outcome.
Distribution of Salmonella cfu
In a separate experiment, Salmonella cfu (105 /mouse) were injected i.v. Distri
bution was determined in blood, liver and spleen after 30, 60, 120 and 360 minutes by
plating serial dilutions of blood and homogenized tissue samples on BGA. After over
night incubation at 37°C the cfu were counted. Groups of 5 mice were used for each
time point.
In vitro cytokine production
Resident peritoneal macrophages of LDLR-/- and LDLR+/+ mice were harvested
by rinsing the peritoneal cavity aseptically with 2 ml cold PBS containing 0.38% (w/v)
sodium citrate. After centrifugation, cells were resuspended in RPMI 1640 (Dutch

S. typhimurium in LDLR-/- mice 101
modification; Flow Laboratories, Irvine, UK), containing 1 гпМ pyruvate, 2 mM L-
glutamine and 100 μg gentamicin per ml, and incubated for 1 h at 37°C (1 Orwell) in
96-wells microtiter plates (Costar Corporation, Cambridge, MA). The non-adherent
cells were discarded and the remaining cell population consisted of more than 90%
macrophages, as assessed by light microscopy.
Heat-killed (30 min, 100°C) S. typhimurium were washed three times in RPMI.
Fifty uL of suspension containing 106 microorganisms in RPMI were added to perito
neal macrophages (105/ well) to a final volume of 100 \iL and incubated at 37°C in 5%
C02. After 24h, the supematants were collected and stored at -70°C until assayed. To
the remaining macrophages, 200 uJ_ of RPMI was added and the cells were disrupted
by three freeze-thaw cycles to determine the cell-associated cytokine contents. The
samples were stored at -70°C until assayed.
In order to assess the effect of the various lipoprotein subfractions on the produc
tion of cytokines, macrophages of LDLR+/+ mice were incubated with LPS-free lipo
protein subfractions of the plasma from LDLR-/- mice: LDL, VLDL and HDL respec
tively. Isolation was performed as described before [11]. Final concentration of isolated
cholesterol was 0.15 mmol/L. Heat-killed Salmonella were used to stimulate these
macrophages in the same procedure as described above.
Cytokine measurements
TNFa, IL-la and IL-Iß concentrations were determined using specific radioim
munoassays (RIA) developed in our laboratory, as previously described [12].
Cytokine mRNA expression
Total RNA from spleen cells 24 hours after infection was isolated by the method of
Chomczynski and Sacchi [13]. For each sample 0.5 μg of total RNA was reverse trans
cribed and PCR was performed as described [14]. The following primers were used for
the PCR reactions (from a 0.5 μg sample RNA): GAPDH, sense, 5'-AACTCCCTCAA
GATTGTCAGCA-3', and antisense, 5'-TCCACCACCCTGTTGCTGTA-3'; TNFa,
sense, 5'-TCTCATCAGTTCTATGGCCC-3', and antisense, 5-GGGAGTAGACAAG
GTACAAC- 3'; IL-la, sense, 5'-CAGTTCTGCCATTGACCATC-3', and antisense, 5'-
TCTCACTGAAACTCAGCCGT-3·, IL-lß, sense, 5-TTGACGGACCCCAAAAGAT
G-3', and antisense, 5'-AGAAGGTGCTCATGTCCTCA-3' (Eurogentec, Seraing, Bel
gium). After checking the reactions to be in the log phase, thirty PCR cycles were per
formed with sets at 92°C for 30 sec, 55°C for 30 sec, and 72°C for 90 sec, using a
Mastercycler 5330 (Eppendorf, Hamburg, Germany). PCR products were run on 2%

102 Chapter 8
agars gels stained with ethidium bromide. The gels were scanned on a densitometer
(GS-670, Biorad, Veenendaal, The Netherlands) and analyzed using Molecular Ana
lyst software (Biorad). The relative amount of TNFa, IL-la and IL—1 β mRNA in a
sample was expressed as a ratio versus the amount of mRNA for the housekeeping gene
GAPDH.
Growth of Salmonella in vitro
To investigate the effect of lipoproteins on the in vitro growth, 0.5 χ IO3 cfu S.
typhimurium in 0.5 mL BHI were incubated with 0.5 mL of plasma obtained from
LDLR+/+ mice (cholesterol concentration 0.2 mM) or plasma obtained from LDLR-/-
mice (cholesterol concentration 1.5 mM). After 2, 7, 12 and 24 hours, aliquots of 0.1
mL were removed, serial dilutions were plated on BGA agar, and cfu were counted
after overnight incubation at 37°C.
Intracellular killing of S. typhimurium by peritoneal macrophages
Intracellular killing of S. typhimurium by macrophages in vitro [described in 15]
was determined as a decrease in the number of viable intracellular bacteria during incu
bation of macrophages containing ingested bacteria after phagocytosis in vivo.
For the in vivo phagocytosis, a suspension (1 mL) containing 2xl08 Salmonella cfu
in broth suppleted with 10% newborn calf serum was injected i.p. into anesthetized
mice (LDLR-/- and LDLR+/+). After 15 minutes, the animals were killed by cervical
dislocation; the peritoneal macrophages were collected after injection of 2 ml cold PBS
containing 50 U/ml heparin. After differential centrifugation and three washes to re
move extracellular bacteria, macrophages containing bacteria were reincubated with
RPMI containing 10% newborn calf serum and 10% autologous (LDLR-/- or LDLR
+/+) serum at 37°C and 5% CO2 and rotated (4 rpm). After 30, 60, and 90 minutes, 50
|lL aliquots of the suspension were administered to 450 uL distilled water with 0.01%
bovine serum in order to lyse the macrophages. The number of viable bacteria was de
termined by plating dilutions of this solution on BGA plates and counting cfu after
overnight incubation at 37°C.
Statistical analysis
Survival curves in control and LDLR-/- mice were compared by the Kaplan-Meyer
log-rank test. Differences in concentrations of cytokines and in organ counts of the
microorganisms were analyzed by the Mann-Whitney U test. Differences were consi
dered significant at ρ < .05. All the experiments were at least performed in duplicate.

S. typhimuríum in LDLR7- mice 103
100
(0 > ¡> (Λ
80
60
40
20
-η η η
LDLR-/-
LDLR+/+
12 16 20 days
Figure 1. Survival of LDLR-/- and LDLR+/+ C57B1/6J mice (at least 20 animals per group) after i.v. in
jection of 102 S. typhimuríum.
Results
Survival of mice and outgrowth oí Salmonella in the organs
After i.v. infection with 102 cfu of S. typhimuríum, only 5% of the LDLR-/- mice
died (p < .001), whereas all LDLR+/+ died within 12 days of infection (fig. 1). Out
growth of Salmonella 3 days after infection with 102 microorganisms was at least 10-
fold lower in liver, spleen and bone marrow of LDLR-/- mice compared with control
LDLR+/+ mice (fig. 2). These differences were significant at ρ <.001 (liver) and ρ < .01
(spleen and bone marrow). On day 7 the differences were even greater (p < .001; fig. 2).
The outgrowth after i.p. infection did not significantly differ from that after i.v. infec
tion.
Distribution of Salmonella
Since the differences in outgrowth could be due to a difference in distribution, we
determined disappearance of microorganisms from the circulation after i.v. injection of
105 S. typhimuríum cfu. Blood from LDLR+/+ mice contained significantly less Salmo-
nella cfu than that of LDLR-/- mice 30 minutes after injection (fig. 3, ρ < .01). The
numbers of cfu in the liver and spleen were 70-80% lower in LDLR-/- mice than that in
LDLR+/+ mice at that point in time, and remained lower throughout the experiment,
being significant at 30, 60, 120 and 360 minutes for the liver (p < .05) and at 30 and 60
minutes for the spleen (p <.05 ; fig. 3).
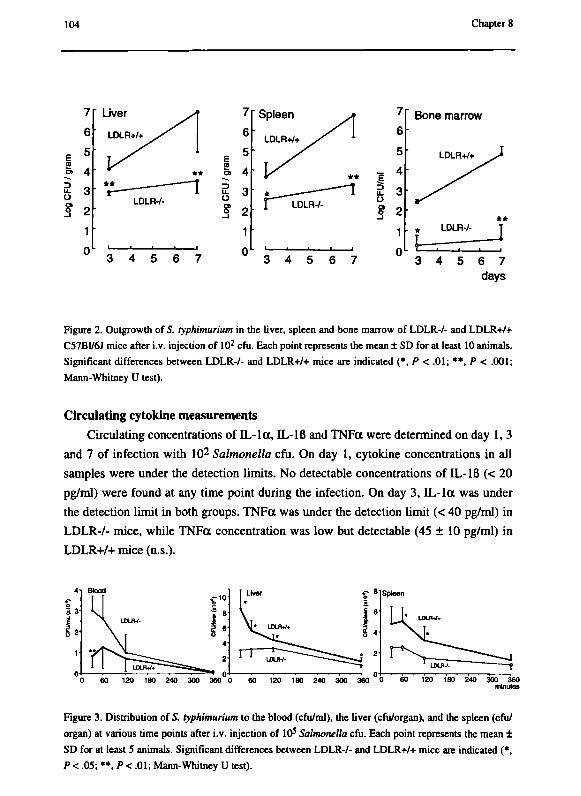
104 Chapter 8
υ α> 3
Bone marrow
.E э υ
Figure 2. Outgrowth of S. typhimurium in the liver, spleen and bone marrow of LDLR-/- and LDLR+/+
C57B1/6J mice after i.v. injection of 102 cfu. Each point represents the mean ± SD for at least 10 animals.
Significant differences between LDLR-/- and LDLR+/+ mice are indicated (*, Ρ < .01; **, Ρ < .001;
Mann-Whitney U test).
Circulating cytokine measurements
Circulating concentrations of IL-la, IL-Iß and TNFa were determined on day 1, 3
and 7 of infection with 102 Salmonella cfu. On day 1, cytokine concentrations in all
samples were under the detection limits. No detectable concentrations of IL-Iß (< 20
pg/ml) were found at any time point during the infection. On day 3, IL-la was under
the detection limit in both groups. TNFa was under the detection limit (< 40 pg/ml) in
LDLR-/- mice, while TNFa concentration was low but detectable (45 ± 10 pg/ml) in
LDLR+/+ mice (n.s.).
Figure 3. Distribution of S. typhimurium to the blood (cfu/ml), the liver (cfu/organ), and the spleen (cfu/
organ) at various time points after i.v. injection of 10s Salmonella cfu. Each point represents the mean ±
SD for at least 5 animals. Significant differences between LDLR-/- and LDLR+/+ mice are indicated (*,
Ρ < .05; **, Ρ < .01; Mann-Whitney U test).

S. typhimunum in LDLR-/- mice 105
On day 7, circulating concentrations of IL-Ια and TNFa were significantly higher
in LDLR+/+ than in LDLR-/- mice: 95 ± 63 pg/ml vs 30 ± 10 pg/ml for IL-la (p < .02)
and 1140 ± 290 pg/ml vs 43 ± 6 pg/ml for TNFa (p < .02) (Figure 4).
Cytokines in the early stages of the infection with 105 Salmonella cfu were asses
sed as well. After 60 minutes, IL-la concentrations in the plasma were higher in the
LDLR-/- mice compared to those in the LDLR+/+ mice: 910 ± 370 pg/ml vs 310 ± 140
pg/ml (p < .05). This difference had disappeared after 2 hours. There were no differen
ces in the TNFa concentrations between the 2 strains (Figure 4).
In vitro cytokine production
To investigate whether a difference in cytokine production capacity in response to
Salmonella could explain the difference in outcome, resident (uninfected) peritoneal
macrophages of LDLR-/- and LDLR+/+ mice were stimulated with heat-killed Salmo
nella in vitro. TNFa concentrations in supernatants of macrophages were similar in
LDLR-/ mice and LDLR+/+ mice: In unstimulated cultures 194 ± 91 pg/ml and 186 ±
55 pg/ml and in macrophage cultures incubated with heat-killed Salmonella 550 ± 166
pg/ml and 520 ±135 pg/ml for LDLR-/- and LDLR+/+ mice, respectively.
Figure 4. Circulating concentrations of TNFa (Δ,Α) and IL-la (•,•) on day 1, 3, and 7 of infection
with of 1 χ 102 CFU S. typhimurium in LDLR-/- (open symbols) and LDLR+/+ (closed symbols) mice.
Significant differences are indicated (*, Ρ < 05; Mann-Whitney U test)
Inset: Circulating concentrations of TNFa (Д,А) and IL-la (C,B) at the early stage (60 and 120
minutes) of infection with 1 χ 105 CFU of S typhimunum in LDLR-/- (open symbols) and LDLR+/+
(closed symbols) mice. Significant differences are indicated (*, Ρ < .05; Mann-Whitney U test).
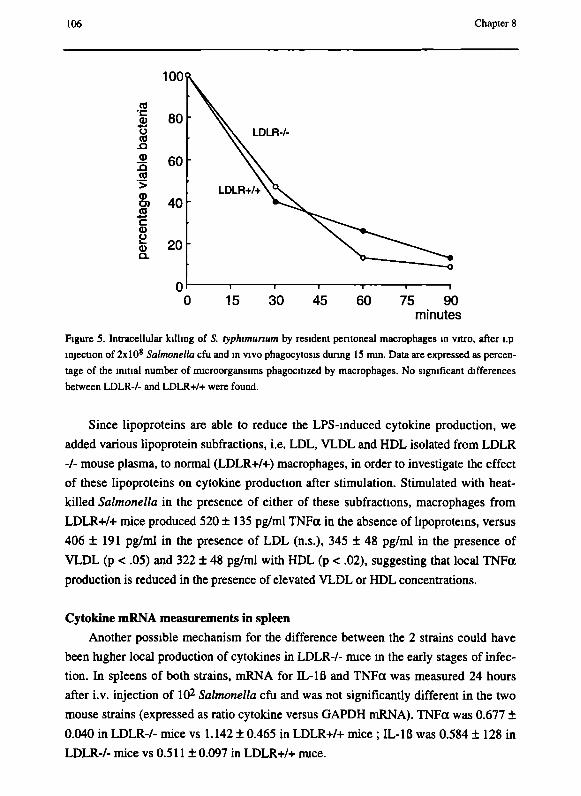
106 Chapter 8
100
fl ш 80
• ^ υ to
Л 60 со '> (D
S) 40 СО
• ^ с φ и Ё 20 о.
0 0 15 30 45 60 75 90
minutes
Figure S. Intracellular killing of S. typhimunum by resident peritoneal macrophages in vitro, after l.p
injection of 2xl0 8 Salmonella cfu and in vivo phagocytosis during IS min. Data are expressed as percen
tage of the initial number of microorgansims phagocitized by macrophages. No significant differences
between LDLR-/- and LDLR+/+ were found.
Since lipoproteins are able to reduce the LPS-induced cytokine production, we
added various lipoprotein subfractions, i.e. LDL, VLDL and HDL isolated from LDLR
-/- mouse plasma, to normal (LDLR+/+) macrophages, in order to investigate the effect
of these lipoproteins on cytokine production after stimulation. Stimulated with heat-
killed Salmonella in the presence of either of these subfractions, macrophages from
LDLR+/+ mice produced 520 ±135 pg/ml TNFa in the absence of lipoproteins, versus
406 ±191 pg/ml in the presence of LDL (n.s.), 345 ± 48 pg/ml in the presence of
VLDL (p < .05) and 322 ± 48 pg/ml with HDL (p < .02), suggesting that local TNFa
production is reduced in the presence of elevated VLDL or HDL concentrations.
Cytokine mRNA measurements in spleen
Another possible mechanism for the difference between the 2 strains could have
been higher local production of cytokines in LDLR-/- mice in the early stages of infec
tion. In spleens of both strains, mRNA for IL-lß and TNFa was measured 24 hours
after i.v. injection of 102 Salmonella cfu and was not significantly different in the two
mouse strains (expressed as ratio cytokine versus GAPDH mRNA). TNFa was 0.677 ±
0.040 in LDLR-/- mice vs 1.142 ± 0.465 in LDLR+/+ mice ; IL-lß was 0.584 ± 128 in
LDLR-/- mice vs 0.511 ± 0.097 in LDLR+/+ mice.

S. typhimurium in LDLR-/- mice 107
Growth of Salmonella in vitro We investigated whether there was an effect of lipoproteins on in vitro growth of S.
typhimurium. The growth curves of Salmonella was similar after 2, 7, 12 and 24 hours
in broth with plasma obtained from LDLR-/- mice and broth with plasma obtained from
LDLR+/+ mice (data not shown).
Intracellular killing of Salmonella in vitro A difference in killing capacity of the respective macrophages could account for a
difference in outgrowth of Salmonella cfu in the organs. After in vivo phagocytosis, the
numbers of cfu phagocytized by. macrophages of LDLR-/- and LDLR+/+ mice were
similar (data not shown), which shows that uptake of salmonellae was not enhanced by
peritoneal macrophages from one of the strains examined. The results of the intracel
lular killing experiments showed that resident peritoneal cells of LDLR-/- mice and
LDLR+/+ mice do not differ in their ability to kill S. typhimurium. The killing rate did
not differ when lipoprotein-rich serum of LDLR-/- mice was added to LDLR+/+ control
macrophages, and likewise, serum from LDLR+/+ mice did not affect the killing of
Salmonella by LDLR-/- macrophages (Figure 5).
Discussion
In the present study we have shown that hyperlipoproteinemic LDLR-/- mice are
protected against S. typhimurium infection. In LDLR-/- mice, survival was significantly
greater than that of control mice, outgrowth of microorganisms in the organs was redu
ced, and circulating proinflammatory cytokine concentrations at late stages of infection
were lower.
In previous studies, it has been shown that lipoproteins are able to bind and neutra
lize LPS, and the subsequently reduced induction of circulating proinflammatory cyto
kines is the main mechanism for the beneficial effect of lipoproteins in Gram-negative
infection [10].
The situation in the intracellular S. typhimurium infection appears to be more com
plicated. First of all, in contrast to an extracellular Gram-negative infection such as that
caused by K. pneumoniae, endotoxin release as judged by the cytokine response is not
prominent. More important is the finding that the outgrowth of Salmonella is reduced in
LDLR-/- mice and the lower bacterial load may be the reason for the lesser degree of
cytokinemia. In lethal K. pneumoniae infection, there was no difference in outgrowth of
microorganisms in the organs between the two strains of mice [10]. In view of the bene-

108 Chapter 8
ficial effect of TNFa in host defense against intracellular infections [5-7, 16], we consi
dered that the reduced outgrowth of bacteria in the tissues could be due to increased
cytokine production at the tissue level not reflected in the circulation. Therefore we
investigated whether the decreased outgrowth of microorganisms in the organs could be
a result of higher local TNFa production. In peritoneal macrophages of LDLR-/- mice,
a slight increase in TNFa and especially IL-Iß production after stimulation with LPS
has been observed previously [10]. In the present study, when peritoneal macrophages
of LDLR-/- mice were stimulated in vitro with heat-killed Salmonella, no increase of
cytokine production by macrophages was detected. These in vitro experiments are
further sustained by our observations that no increased local production of messenger
RNA in the spleens of LDLR-/- mice could be found during infection with S. typhi-
murium. This implies that the decreased outgrowth of Salmonella in the liver and spleen
is unlikely to be explained by a local increase of proinflammatory cytokines. Another
explanation could be a difference in microbicidal capacity of phagocytic cells.
High concentrations of lipoproteins may induce changes at the cell-membrane level
[17, 18], thereby changing the susceptibility to Salmonella infection. Although diver
gent rates of S. typhimurium outgrowth in the organs of infected mice may suggest that
differences either in killing or in virulence of Salmonella might play a role, we found
that both phagocytosis and intracellular killing of S. typhimurium by macrophages from
LDLR-/- mice were similar to that of control mice.
A direct effect of lipoproteins on the bacteria may also have influenced the decrea
sed outgrowth of Salmonella in LDLR-/- mice. In this respect, it is of interest that HDL
has been found to be cidal against Trypanosoma cruzi [14]. However, the growth of
Salmonella was similar in LDL-rich medium and in control medium, excluding direct
involvement of lipoproteins as an inhibiting factor for Salmonella growth in LDLR-/-
mice.
An important finding in our studies is the difference in distribution of Salmonella
during the first hours after infection. It is conceivable that these short term distribution
differences determine the short and long term outgrowth and subsequent better survival
of LDLR-/- mice to a considerable extent. To explain this phenomenon there are two
possibilities: either the salmonellae are kept in the circulation or the organs have im
paired capabilities for uptake of the microorganisms. It could be that lipoproteins
directly interact with Salmonella uptake, for example by coating the bacterial surface
structures with lipids. Alternatively, the absence of the LDL receptor could be the
reason for an altered uptake of bacteria. In this respect it is interesting that malaria
parasites compete for the same receptor with certain lipoproteins [19], and that Strepto-

S. typhimurìum in LDLR-Λ mice 109
coccus pneumoniae has been shown to bind to a lipid receptor [20]. However, the ex
periments in which the in vivo phagocytosis of salmonellae by peritoneal macrophages
was investigated argue against such a mechanism, since equal uptake was found by
LDLR-A and LDLR+/+ macrophages. Still, further studies are necessary before this
hypothesis can be rejected.
Acknowledgments The authors thank Ineke Verschueren for performing the cytokine determinations
and Margo van de Brink, Monique Bakker, Yvette Brom, Theo van der Ing, and Maria
van den Barselaar for assistance with the animal experiments.
References
1. Greisman SE. The role of endotoxin during typhoid fever and tularemia in man. J
Clin Invest 1969; 48: 613-629.
2. Keuter M, Dharmana E, Gasem MH, Van der Ven-Jongekrijg J, Djokomoeljanto R,
Dolmans WMV, Demacker PMN, Sauerwein R, Gallati H, Van der Meer JWM.
Patterns of proinflammatory cytokines and inhibitors during typhoid fever. J Infect
Dis 1994; 169: 1306-1311.
3. Peel JE. Induction of circulating tumor necrosis factor cannot be demonstrated
during septicemic salmonellosis in calves. Infect Immun 1990; 58: 439-42.
4. Jotwani R, Tanaka Y, Watanabe K, Tanaka K, Kato N, Ueno К. Cytokine stimula
tion during Salmonella typhimurium sepsis in ItyS mice. J Med Microbiol 1995;
42: 348-52.
5. Nakano Y, Onozuka К, Terada Y, Shinomiya H, Nakano M. Protective effect of
recombinant tumor necrosis factor-α in murine salmonellosis. J Immunol 1990;
144: 1935-1941.
6. Mastroeni P, Skepper JN, Hormaeche CE. Effect of anti-tumor necrosis factor
alpha antibodies on histopathology of primary Salmonella infections. Infect Immun
1995; 63: 3674-3682.
7. Nauciel C, Espinasse-Maes F. Role of gamma-interferon and tumor necrosis factor
alpha in resistance to Salmonella typhimurium infection. Infect Immun 1992; 60:
450-454.
8. Harris HW, Grunfeld C, Feingold KR, et al. Chylomicrons alter the fate of endo
toxin, decreasing tumor necrosis factor release and preventing death. J Clin Invest
1993; 91: 1028-1034.

по Chapter S
9. Flegel WA, Wolpl A, Mannel DN, Northoff H. Inhibition of endotoxin-induced
activation of human monocytes by human lipoproteins. Infect Immun 1989; 57:
2237-2245.
10. Netea MG, Demacker PNM, Kullberg BJ, Boerman OC, Verschueren I, Stalenhoef
AFH, Van der Meer JWM. Low-density-lipoprotein receptor deficient mice are
protected against lethal endotoxinemia and severe Gram-negative infections. J Clin
Invest 1996; 97: 1366-1372.
11. Demacker PNM, Vos-Jansen HE, Jansen AP, Van 't Laar A. Evaluation of the dual
precipitation method by comparison with the ultracentrifugation methods for the
measurement of lipoproteins in serum. Clin Chem 1977; 23: 1238-1244.
12. Drenth JPH, Van Uum SHM, Van Deuren M, Pesman GJ, Van der Ven-Jongekrijg
J, Van der Meer JWM. Endurance run increases circulating IL-6 and IL-Ira but
downregulates ex vivo TNF-ct and IL-Iß production. J Appi Physiol 1995; 79:
1497-1503.
13. Chomczynski P, Sacchi N. Single step method of RNA isolation by guanidinum
thiocyanate phenol chloroform extraction. Anal Biochem 1987; 162: 156-159.
14. Smith AB, Esko JD, Hajduk SL. Killing of trypanosomes by the human haptoglo-
bin-related protein. Science 1995; 268: 284-286.
15. Van Dissel JT, Stikkelbroeck JJM, Sluiter W, Leijh PCJ, Van Fürth R. Differences
in initial rate of intracellular killing of Salmonella typhimurium by granulocytes of
salmonella susceptible C57B1/10 mice and salmonella resistant CBA mice. J
Immunol 1986; 136: 1074-1080.
16. Tite JP, Dougan G, Chatfield SN. The involvement of tumor necrosis factor in im
munity to Salmonella infection. J Immunol 1991; 147: 3161-3164.
17. Esfahani M, Bigler RD, Alfieri JL, Lund-Katz S, Baum JD, Scerbo L. Cholesterol
regulates the cell surface expression of glycophospholipid-anchored CD 14 antigen
on human monocytes. Biochim Biophys Acta 1993; 1149: 217-223.
18. Rothberg KG, Ying YS, Kamen BA, Anderson RGW. Cholesterol controls the
clustering of the glycophospholipid-anchored membrane receptor for 5-methyl-
tetrahydrofolate. J Cell Biol 1990; 111: 2931-2938.
19. Sinnis P, Willnow TE, Briones MRS, Herz J, Nussenzweig V. Remnant lipopro
teins inhibit malaria sporozoite invasion of hepatocytes. J Exp Med 1996; 184:
945-954.
20. Cundell DR, Gerard NP, Gerard C, Idanpaan-Heikkila I, Tuomanen EI. Strepto
coccus pneumoniae anchor to activated human cells by the receptor for platelet-
activating factor. Nature 1995; 377: 435^38.

Chapter 9
General Discussion

112 Chapter 9

General Discussion ИЗ
Mediators of fever in patients with typhoid fever In the classical ideas about the induction of fever, pyrogenic cytokines circulate
and stimulate the synthesis of PGE2 in the lamina terminalis of the central nervous
system (CNS), which results in behavioral, endocrine and autocrine responses [13]. In
the study described in Chapter 3, we were not able to detect appreciable concentrations
of the pyrogenic cytokines IL-Iß, TNFa and LTa in patients with typhoid fever. The
concentrations of IL-6, generally considered a relatively weak pyrogen [144], were low
as compared to other febrile conditions [145, 146]. The suggestion that synthesis of
prostaglandins such as PGE2 in the CNS might be induced by other substances or in
other ways than by circulating endogenous pyrogenic cytokines, should be considered
[15].
Many years ago, Cranston's group showed that intraventricular injection of inhi
bitors of sPLA2 suppressed fever in rabbits [16]. Since sPLA2 release is induced by IL-
1 and TNFa and the enzyme could mediate the production of arachidonic acid (AA)
[20], sPLA2 might be this intermediate.
In Chapter 4, we found elevated concentrations of extracellular group II 14-kDa
sPLA2 during the acute phase of typhoid fever in 12 patients. In patients with compli
cated disease the bioactive sPLA2 concentrations were 10-fold higher than in those
with uncomplicated disease. sPLA2 concentrations fell significantly towards recovery.
We considered that this bioactive sPLA2 might be responsible for the symptoms of
typhoid fever, as sPLA2 is locally induced by proinflammatory cytokines at the site of
the infection and reaches the circulation. In this manner sPLA2 could mediate the sys
temic signs of the disease [19, 147].
However, this concept of sPLA2 as a circulating mediator of fever has become
more doubtful lately. The role of sPLA2 in the generation of AA and other eicosanoids
which takes place intracellularly, is not clear. It has been questioned why cells should
secrete an enzyme involved in intracellular functions, when there is already an intracel
lular enzyme for AA formation. This enzyme is the cytosolic 85 kDa PLA2. Further
more, we were not able to induce fever in rabbits with purified rat sPLA2 (unpublished
results). Recombinant human sPLA2 did not express inflammatory activity in normal
rat tissues either [148].
Recent publications suggest a role for sPLA2 together with C-reactive protein
(CRP) in the removal of injured cells. For instance in myocardial infarction, it is hypo
thesized that CRP binds to muscle cells via lyso-phosphatidylcholine, generated in the
outer leaflet of the cell membrane by sPLA2. Inner leaflet phosphatidylserine and etha-
nolamine move to the outer leaflet in "flip-flopped" membranes, making phosphatidyl-

114 Chapter 9
choline more susceptible to phospholipolysis by sPLA2. This damage or flip-flop
occurs, when the cells are ischemic. Complement is bound and neutrophils activated
when CRP binds to these lysophospholipids [149]. In this hypothesis sPLA2 has a func
tion in waste disposal probably to allow quick repair. sPLA2 can only be damaging in
situations, where membranes of cells are already injured. This might explain the ab
sence of biological effects when recombinant sPLA2 is injected into healthy animals
[148].
sPLA2 is relevant to the pathogenesis of various clinical conditions, including
rheumatoid arthritis (RA) [150]. As females tended to have higher group IIPLA2 acti
vity in plasma and cells, the authors of this study suggested that sPLA2 activity might
be a risk factor for RA in view of the increased prevalence of the disease in women
[151]. So, although the function of sPLA2 in myocardial infarction [18], RA and
septicemia could be the removal of injured cells, which might be increased by an excess
of sPLA2, the pathobiology of excess sPLA2 production is not completely understood.
In the concept of fever, it is important to note that although concentrations of circu
lating pyrogenic cytokines were low in the acute febrile phase of typhoid fever, mRNA
for TNFa and IL-Iß was found to be present in circulating whole blood cells (Chapter
J). Thus these circulating cells could themselves produce or induce pyrogenic cytokines
in the endothelial cells of the OVLT [15]. Moreover, the recent finding of membrane-
bound TNFa, might explain the absence of free circulating pyrogens in the circulation.
One could hypothesize that this membrane bound TNFa could be released at the site
(OVLT) [152].
Cytokine profile in patients with typhoid fever In Chapter 3, we report low concentrations of circulating proinflammatory cytoki
nes, but high concentrations of the antiinflammatory IL-IRA and sTNFRI and sTNFRII
during the acute phase of typhoid fever. At that time point, the capacity of LPS-induced
production of proinflammatory cytokines in whole blood of patients with typhoid fever
was down-regulated. This depressed cytokine production was not a consequence of a
lower number of white blood cells during the acute phase of the infection.
Down-regulation of cytokine production In Chapter 3, we describe down-regulation of the production of the proinflam
matory cytokines IL-1, IL-6 and TNFa, but not of the anti-inflammatory products
sTNF-receptors and IL-IRA. This is in accordance with later studies on endotoxemia in
volunteers, in which production of proinflammatory cytokines (but also interestingly of

General Discussion 115
the antiinflammatory cytokine IL-10) was found to be suppressed, whereas production
of receptors of TNFa and IL-IRA was increased [153].
It has been described that exposure to endotoxin leads to tolerance [154]. Tolerance
is a general biological phenomenon observed for many ligand-receptor interactions. A
low production of cytokines has not only been found in serious conditions like severe
post-operative infection [155] and sepsis [156-158], but also in patients with familial
Mediterranean fever, during attacks which are not endotoxin-mediated [159,160]. Also
after major surgery or blood loss and strenuous exercise [161, 162], down-regulation of
cytokines was observed and it has been suggested that any major stress can give an im
paired inflammatory response.
This down-regulation of cytokine production occurs only a short time after the in
jury. At 2 hours after endotoxin infusion in volunteers, cytokine production upon LPS
stimulation of whole blood was down-regulated [153]. Also in isolated peripheral blood
mononuclear cells (PBMCs), down-regulation of proinflammatory cytokine production
was apparent few hours after the first injection of endotoxin in volunteers [163].
Decreased cytokine production capacity as a consequence of exposure in vivo to
substances such as for instance endotoxin, has often been interpreted as exhaustion of
cytokine-producing cells [155-160]. Our finding that IL-1 receptor antagonist and
sTNFRs are being produced in high concentrations in whole blood argues against
exhaustion of cells, although these antiinflammatory cytokines have different kinetics
and might be produced by different cell types. PBMCs of patients with typhoid fever
produced fewer proinflammatory cytokines, but were still able to produce EL-IRA in
the acute phase of typhoid fever (unpublished results), indicating that the IL-IRA found
in the whole blood method was at least partly produced by monocytes and not only by
granulocytes.
To investigate the mechanism of down-regulated cytokine production in typhoid
fever further, messenger RNA for IL-Iß and TNFa in whole blood was measured and
this study was described in Chapter 5. The finding that the amounts of mRNA for IL-Iß
and TNFa in the acute phase of typhoid fever were similar to those in the recovery
phase, also denies an exhausted production and is in favor of post-transcriptional inhibi
tion of cytokine production.
LPS tolerance may in part be reproduced by post-endotoxin infusion plasma, sug
gesting that soluble factors produced in response to the LPS challenge are involved
[153]. These soluble factors might include glucocorticosteroids, norepinephrin, lipo
proteins and antiinflammatory cytokines [112, 164, 165]. For instance antiinflammatory
IL-4, IL-10 and TGF-ß are able to down-regulate the production of IL-1, TNFa, IL-6,

116 Chapter 9
IL-8, and to induce the production of IL-IRA [120]. The mechanisms of endotoxin
tolerance are not clear so far. In acute Gram-negative sepsis, down-regulation of cyto
kine production was shown to be caused partly by norepinephrin, which is able to
attenuate cytokine production [165]. In addition, proinflammatory cytokines themselves
may be responsible for the down-regulation, as was shown in mice treated with a single
low dose of rIL-1 [109]. Apparently, after the initial phase of infection there is a post-
transcriptional switch-off of the production of proinflammatory cytokines and a switch-
on to an anti-inflammatory repertoire in mononuclear cells. Theoretically one could
hypothesize that within the mononuclear cells there are two subtypes: those that are sus
ceptible to Thl stimulation and those that are more Th2-sensitive.
Down-regulation and the concept of a switch have been assented by several inves
tigators and have also been shown in vitro in isolated peripheral blood mononuclear
cells. In such experiments, preincubation of PBMCs with a low dose of LPS rendered
these cells tolerant for subsequent stimulation with LPS, resulting in lower TNFa as
well as lower IL-10 responses, but not lower IL-RA production [166]. IL-10 mRNA up-
regulation has been reported in these LPS-tolerant monocytes, whereas TNFa mRNA
was found to be reduced [167]. The CD14 receptor was still expressed in these cells as
well as the transcription factor NF-κΒ, which suggests that LPS could reach its receptor
and cytokine production was not inhibited by a transcription factor [168]. In vitro, anti
inflammatory cytokines such as IL-10 and TGF-ß were able to induce the process of
down-regulation, and neutralizing these cytokines could prevent it [166]. These expe
riments show that IL-10 and TGF-ß are important in the switch to antiinflammatory
cytokine production. However, in vitro down-regulation of cultured cell populations,
which are repeatedly stimulated with LPS is not similar to ex vivo stimulation of whole
blood from patients with for instance endotoxemia.
It was also found that endotoxin tolerance could be antagonized by IFN-γ in vitro
[167]. Recent clinical studies have shown that this LPS tolerance or "immunoparalysis",
in which monocytes of septic patients were neither able to express HLA-DR nor to
produce TNFa, could be reversed in these patients by treatment with IFN-γ [169]. Re
covery of this immune function was reported to lead to survival in 8 of 9 patients.
In conclusion, in-vivo down-regulation of cytokine production with preserved
mRNA of proinflammatory cytokines, which we found during the acute phase of
typhoid fever, is a common phenomenon in many circumstances. Apparently, it is
found in endotoxin-mediated diseases [153, 164, 170, 171] as well as in the intracellular
Salmonella infection. Down-regulation is most likely a protection against overwhel-

General Discussion 117
ming cytokinemia, but can impair immune functions.
The benefit of down-regulation in patients with typhoid fever is difficult to under
stand in view of experimental animal studies with Salmonella infection, in which the
proinflammatory cytokines are required to survive the infection [82-84, 86, 87] (see
Chapter 2). Possibly the deeper downregulated proinflammatory cytokines in compli
cated cases could reflect a less effective host defense and immunotherapy might be
needed to improve host defense.
Cytokines during experimental S. typhimurium infection In Chapter 6, the cytokine profile during experimental S. typhimurium infection in
mice is described. Without infection, circulating cytokine concentrations but also LPS-
stimulated ex-vivo production in whole blood were undetectable. The latter inability to
produce cytokines is the reverse of the human situation, since human peripheral blood
cells can be stimulated to produce large amounts of proinflammatory cytokines without
infection. In contrast, uninfected bone marrow and peritoneal macrophages were able to
produce proinflammatory cytokines after LPS stimulation.
During S. typhimurium infection, the circulating TNFa concentration was increa
sed on day 7, as was the LPS-stimulated production of IL-la, IL-lß and TNFa in
whole blood. Production of IL-lß and TNFa by peritoneal and bone marrow cells was
also increased during infection. Whereas LPS-stimulated cytokine production during S.
typhimurium infection was generally increased, the IL-la production by infected peri
toneal cells was decreased. This is an important observation, since peritoneal macro
phages are easy to obtain and frequently used as representatives for the resident or acti
vated macrophage population of the mouse. This shows that the cytokine response of
peritoneal and other macrophages are divergent. The attention to peritoneal cells alone
might not yield information on cytokine production at other sites of infection.
The general upregulation of proinflammatory cytokine production in this animal
model is also in contrast with what we have found in humans with typhoid fever, des
cribed in Chapter 3. In these patients, down-regulation of LPS-stimulated production of
cytokines in whole blood was found. This phenomenon was even more profound in
complicated cases. These differences seem not to be caused by the time point of
measuring cytokine production: The patients with typhoid fever had been ill for a week
before admission, and down-regulated production was found at that time point. In the
experimental study, mice were followed for 7 days and upregulation of cytokine
production was predominantly found at day 7. It is obvious that one should be careful to
draw conclusions from animal experiments and extrapolate the results to the human

118 Chapter 9
situation.
The effect of IL-1 pretreatment In Chapter 6, the effect of pretreatment of mice with rhIL-la on S. typhimurium
infection has been described. Administration of rhIL-la led to a significantly decreased
outgrowth of microorganisms in the liver and spleen of mice. This effect of rhIL-la
pretreatment was seen as early as 24 hours after the onset of the infection. The micro
biological effect of a single dose of rhIL-la lasted throughout the study.
Pretreatment with rhIL-la had no effect on circulating cytokine concentrations.
The ex-vivo cell-associated IL-la production by peritoneal and bone marrow macro
phages however, was significantly increased 24 hours after pretreatment.
RhIL-la had no effect on outgrowth of microorganisms during infection with
Pseudomonas aeruginosa in neutropenic mice [172]. In those experiments, it has been
shown that administration of either rIL-1 or rTNFa increased survival of mice. This
effect was partially due to reduction of circulating concentrations of proinflammatory
cytokines and reduction of LPS-stimulated cytokine production by peritoneal macro
phages [109]. It is very unlikely that the mechanism of reducing TNFa production
applies to both Salmonella and Pseudomonas infections. It is known that lacking the
TNFa receptor-1 is deleterious for mice with intracellular Listeria infection whereas it
is life-saving for the same mice with endotoxemia [74, 75]. Besides, we did not find
downregulation of TNFa production after rhIL-la treatment in our experiments with
Salmonella-infected mice.
Possible mechanisms of the reduction in outgrowth of S. typhimurium in the organs
include neutrophil recruitment and activation. Intraperitoneally administered IL-1 in
creased the numbers of neutrophils [106]. IL-1 is able to induce granulocyte colony-
stimulating factor (G-CSF) in vitro, although concentrations of G-CSF have never been
measured in vivo after rIL-1 pretreatment. However, granulocyte numbers were not
found to be increased in our experiment and in those of others at the time point that the
effect on microbial outgrowth was seen [173]. We have shown in Chapter 7 that recom
binant murine G-CSF increased survival, but did not reduce outgrowth of S. typhim
urium in the organs of mice with a sublethal infection. This suggests that induction of
G-CSF is not likely to be the mechanism behind the protection by rIL-1 pretreatment.
There is some debate whether administration of rIL-1 leads to activation of macro
phages [100, 110], and if so, whether these activated macrophages are important in the
earliest stage of the infection, including the distribution phase, or as more efficient
killers later during the infection. Studies with macrophages activated through a prece-

General Discussion 119
ding S. typhìmurìum or BCG infection showed that this led to lower numbers of salmo-
nellae in the organs during the first hours of a subsequent infection [44,45]. However,
these previously activated macrophages, though able to inhibit T. gondii did not display
a significant effect on growth rate of microorganisms in the organs during this Salmo
nella infection [44, 45]. The outgrowth pattern in our experiment also suggests a distri
bution effect of rhIL-la pretreatment, as the rate of outgrowth in the organs of control
mice and rhIL-la pretreated mice is similar between day 1 and 7. This early effect may
be established by several mechanisms. First, antibodies (specific or non-specific) could
be induced by previous infections and these could influence the distribution of bacteria.
Second, the macrophages themselves may be involved, although the unaffected killing
rate argues against this hypothesis. Third, the activated macrophages may produce
cytokines that exert their effect on other cells, such as granulocytes or NK cells, to
enhance killing. Alternatively, these cytokines may influence invasion or intracellular
killing of microorganisms in hepatocytes or other cells [64,174].
The fact that IL-1 pretreatment protects against different types of infection is in
favor of a general phenomenon such as macrophage activation. The hypothesis that this
effect of rIL-1 is exerted through an effect on distribution is supported by the obser
vation that a similar rate of outgrowth in the organs of control mice and rIL-1 pretreated
mice is also found in Candida infection [173]. Also the beneficial effect of BCG-activa-
ted macrophages on Candida infection was seen in the first hours of infection [175].
The effect of G-CSF treatment In Chapter 7, we described that recombinant murine granulocyte colony-stimula
ting factor (rmG-CSF) improved survival of mice with a potentially lethal Salmonella
typhimurium infection. However, the outgrowth of S. typhimurium in the liver, spleen,
bone marrow, lungs, brain, kidneys and heart of rmG-CSF mice was not different from
that of control mice. Histological examination of liver and spleen on day 1 and 5
showed more localized inflammatory foci in the rmG-CSF treated animals. These
results show that pretreatment with rmG-CSF is beneficial in the facultative intracel
lular 5. typhimurium infection in mice.
The effect of rG-CSF on proinflammatory cytokines has been studied by several
investigators. In contrast to granulocyte-macrophage colony-stimulating factor (GM-
CSF), rG-CSF did not induce fever, suggesting that the induced TNFa production in
GM-CSF treated macrophages [176] was absent with rG-CSF. In human volunteers,
rG-CSF treatment increased plasma concentrations of IL-IRA, TNFa and sTNFRs
without any side effects such as fever [177]. It was found that ex vivo production of

120 Chapter 9
TNFcc in human blood was lower after rG-CSF treatment [142, 143], although Pajkrt et
al. found a biphasic response [178]. In human volunteers pretreated with rG-CSF 2
hours or 12 hours before endotoxin infusion, the peak of circulating TNFa was increa
sed [177, 178]. In rodents, pretreatment with rG-CSF 1 hour before endotoxin infusion
protected against LPS-induced death, but rG-CSF did not prevent TNFa-induced death,
suggesting an effect on TNFa production [179].
In our experiments in mice, basal circulating cytokine concentrations did not differ
between the rmG-CSF treated and control group. One day after treatment, LPS-stimu-
lated TNFa production of resident peritoneal macrophages was significantly higher in
rmG-CSF treated mice. Apparently, differences in species, time points and way of ad
ministration of G-CSF (in vivo or in vitro) play a role in these discrepancies.
It may be questioned, whether rG-CSF has a beneficial effect on intracellular infec
tions through enhanced (early) neutrophil recruitment, and whether the (late) antiin
flammatory properties of rG-CSF are deleterious, e.g., through inhibition of TNFa
mediated macrophage activation. The trend for an increased number of cfu in liver and
spleen in the group, that received continuous rmG-CSF treatment, could be an illus
tration of the latter suggestion and may be due to such an adverse late effect. Late treat
ment with rmG-CSF was also deleterious in experimental Candida infection in mice
(Kullberg, unpublished results).
Pretreatment alone is beneficial in several infections such as with Candida [136].
In L. monocytogenes infection in mice, pretreatment with high dose human rG-CSF
during 5 days, reduced mortality as well as outgrowth of microorganisms [140,141].
In conclusion, rmG-CSF pretreatment protects mice against a lethal S. typhimurium
infection in mice, although no effect was found on the outgrowth of microorganisms in
the organs. Further studies are needed to elucidate the mechanism, before rG-CSF can
be used as immunotherapy in Salmonella infection.
In recent studies, rG-CSF increased the in vitro antimicrobial activity of neutro
phils with ciprofloxacin [180]. Neutrophils treated with rG-CSF for 1 hour were incuba
ted with ciprofloxacin. rG-CSF increased the intracellular concentration of cipro
floxacin in these neutrophils. Inhibition of microorganisms was greatest when neutro
phils, rG-CSF and quinolones were combined. As ciprofloxacin is an important antimi
crobial against Salmonella infection, the combination of these substances is worth
investigating in Salmonella infection.
Lipids, endotoxin and Salmonella infection In patients with Gram-negative bacteremia, lipid infusions are considered for

General Discussion 121
treatment, since lipoproteins can bind and neutralize LPS and thereby reduce proinflam
matory cytokine production. Therefore we thought it interesting to investigate the effect
of lipoproteins on infection with the Gram-negative intracellular Salmonella typhimu-
rium.
In Chapter 8, the presence of high concentrations of lipoproteins were shown to
improve the outcome of S. typhimurium infection. We show, that neutralization of
endotoxin, which in Klebsiella infection is the beneficial effect of lipids, is not the
mechanism behind the better survival of S. typhimurium infection.
During a lethal S. typhimurium infection in C57B1/6J mice, survival in LDLR-/-
mice was greater when compared with LDLR+/+. The outgrowth of S. typhimurium in
the liver, spleen and bone marrow of LDLR-/- mice was 100-1000 times lower than in
controls on day 3 and 7 after infection. Remarkably, circulating cytokine concentrations
were lower in LDLR-/- mice.
A most important finding in this study is the difference in distribution of micro
organisms during the first hours of infection. After intravenous injection of Salmonella
into LDLR+/+ mice, the bacteria disappeared from the blood within 30 minutes with
rapid invasion of liver and spleen. In contrast, in LDLR-/- mice, Salmonella remained
in the circulation for up to 2h, with a significantly decreased (70-80%) organ count.
How these short term distribution differences relate to the long term outgrowth and
subsequent better survival of LDLR-/- mice, has still to be elucidated. Apparently, two
phenomena play a role: on the one hand the difference in early distribution and on the
other hand the difference in growth rate of salmonellae in the organs of mice.
Distribution effects may be explained by lipids in the circulation, as these are able
to influence the uptake of for instance endotoxin by various cells. Infusion of chylo
microns into rats has been shown to reduce the endotoxin uptake in Kupffer cells and to
increase uptake by hepatocytes [181]. Although LPS is not the same as Salmonella bac
teria, it could well be that lipoproteins directly interfere with Salmonella uptake, for
example by coating Salmonella surface structures with lipids. Also, it may be hypothe
sized that the LDL-receptor influences the uptake of bacteria. In this respect it is inter
esting that malaria parasites compete for the same receptor with certain lipoproteins
[182], and that Streptococcus pneumoniae has been shown to bind to a lipid receptor
[183].
Still, the difference in growth rate is not explained by any of these phenomena. We
found that phagocytosis of Salmonella by peritoneal macrophages, as well as the subse
quent intracellular killing did not differ between LDLR+/+ and LDLR-/- mice. This
suggests that the observed difference in growth rate is to be explained by an effect

122 Chapter 9
through other phagocytic cells, e.g. neutrophils, by induction of humoral antibacterial
factors or by divergent effects of LDLR-/- on macrophage types other than the perito
neal cells, which we studied.
References
1. Jones B, Falkow S. Salmonellosis: Host immune responses and bacterial virulen
ce. Annu Rev Immunol 1996; 14: 533-561.
2. Pang T, Bhutta ZA, Finlay BB, Altwegg M. Typhoid fever and other salmonello
sis: a continuing challenge. Trends Microbiol 1995; 3: 253-255.
3. Rathman M, Barker LP, Falkow S. The unique trafficking pattern of Salmonella
typhimurium-conlaining phagosomes in murine macrophages is independent of
the mechanism of bacterial entry. Infect Immun 1997; 65: 1475-1485.
4. Tang P, Foubister V, Pucciarelli MG, Finlay BB. Methods to study bacterial inva
sion. J Microbiol Meth 1993; 18: 227-240.
5. Mackaness GB, Blanden RV, Collins FM. Host-parasite relations in mouse
typhoid. J Exp Med 1966; 124: 573-583.
6. Hussein Gasem M, Dolmans WMV, Bambang Isbandrio B, Hendro Wahyono,
Keuter M, Djokomoeljanto R. Culture of Salmonella typhi and Salmonella para
typhi from blood and bone marrow in suspected typhoid fever. Trop Geograph
Med 1995; 47: 164-167.
7. Onuigbo MAC. Diagnosis of typhoid fever in Nigeria: misuse of the Widal test.
Trans R Soc Trop Med Hyg 1990; 84: 129-131.
8. Butler T, Rolfe RD, James GM, Hentges DJ. Soluble plasma antigen in experi
mental Salmonella typhimurium infection in mice. FEMS Immunol Med
. Microbiol 1994; 9: 325-332.
9. Van den Bergh ET AM, Hussein Gasem M, Keuter M, Dolmans WMV. Outcome
in three groups of patients with typhoid fever treated in Indonesia between 1948
and 1990. 1998; submitted.
10. Bambang Isbandrio B, Hussein Gasem M, Dolmans WMV, Hoogkamp-Korstanje
JAA. Comparative activities of three quinolones and seven comparison standard
drugs against Salmonella typhi from Indonesia. J Antimicrob Chemother 1994;
33: 1055-1056.
11. Simanjuntak CH, Paleólogo FP, Punjabi NH, Darmowigoto R, Harjining Toto-
sudirjo S, Haryanto P, Suprijanto E, Witham ND, Hoffman SL. Oral immuniza
tion against typhoid fever in Indonesia with Ty21a vaccine. Lancet 1991; 338:

General Discussion 123
1055-1059.
12. Sztein MB, Wasserman SS, Tacket CO, Edelman R, Hone D, Lindberg AA,
Levine MM. Cytokine production patterns and lymphoproliferative responses in
volunteers orally immunized with attenuated vaccine strains of Salmonella typhi.
J Infect Dis 1994; 170: 1508-1517.
13. Dinarello CA, Cannon JG, Wolff SM. New concepts on the pathogenesis of fever.
Rev Infect Dis 1988; 10: 168-189.
14. Dinarello CA, Wolff SM. Pathogenesis of fever in man. N Engl J Med 1979; 298:
607-612.
15. Netea MG, Kullberg BJ, Van der Meer JWM. Fever: Do endogenous pyrogens
circulate in the bloodstream? Clin Infect Dis 1998; in press:
16. Cranston WI, Hellon RF, Mitchell D, Townsend Y. Intraventricular injections of
drugs which inhibit phospholipase A2 suppress fever in rabbits. J Physiol 1983;
339: 97-105.
17. Vadas P, Pruzanski W, Stefanski E, Sternby В, Mustard R, Bohnen J, Fraser I,
Farewell V, Bombardier C. Pathogenesis of hypotension in septic shock: Correla
tion of circulating phospholipase A2 levels with circulatory collaps. Crit Care
Med 1988; 16: 1-7.
18. Pruzanski W, Vadas P. Phospholipase A2. A mediator between proximal and
distal effectors of inflammation. Immunol Today 1991; 12: 143-146.
19. Crowl RM, Stoller TJ, Conroy RR, Stoner CR. Induction of phospholipase A2
gene expression in human hepatoma cells by mediators of the acute phase respon
se. J Biol Chem 1991; 266: 2647-2651.
20. Schalkwijk С, Pfeilschifter J, Marki F, Van den Bosch H. Interleukin-lß, tumor
necrosis factor and forskolin stimulate the synthesis and secretion of group II
phospholipase A2 in rat mesangial cells. Biochem Biophys Res Commun 1991;
174: 268-275.
21. Vadas P, Hay JB. Involvement of circulating phospholipase A2 in the pathogene
sis of the hemodynamic changes in endotoxin shock. Can J Physiol Pharmacol
1983;61:561-566.
22. Nerad JL, Griffiths JK, Van der Meer JWM, Endres S, Keusch GT, Bennish M,
Dinarello CA, Cannon JG. A simple rapid method for studying cytokine produc
tion by endotoxin-stimulated leukocytes. Cytokine 1989; 1: 139.
23. Bendtzen K, Morling N, Fomsgaard A, Svenson M, Jakobson В, Odum Ν,
Svejgaard A. Association between HLA-DR2 and production of tumor necrosis
factor alpha and interleukin-1 by mononuclear cells activated by lipopoly-

124 Chapter 9
saccharide. Scand J Immunol 1988; 28: 599-606.
24. Nerad JL, Griffiths JK, Van der Meer JWM, Endres S, Poutsiaka DD, Keusch
GT, Bennish M, Salam MA, Dinarello CA, Cannon JG. Interleukin-lß (IL-lß),
IL-1 receptor antagonist, and TNF-a production in whole blood. J Leukocyte Biol
1992; 52: 687-692.
25. Van Deuren M, Van der Ven-Jongekrijg J, Keuter M, Demacker PNM, Van der
Meer JWM. Cytokine production in whole blood cultures. J Int Fed Clin Chem
1993;5:216-221.
26. Vidal S, Tremblay ML, Govoni G, Gauthier S, Sebastiani G, Malo D, Skamene E,
Olivier M, Jothy S, Gros P. The Ity/Lsh/Bcg locus: Natural resistance to infection
with intracellular parasites is abrogated by disruption of the Nrampl gene. J Exp
Med 1995; 182: 655-666.
27. Vidal S, Lepage Ρ, Gauthier S, Gros P. Natural resistance to intracellular infec
tions. J of Imm 1996; 157: 3559-3568.
28. Govoni G, Vidal S, Gauthier S, Skamene E, Malo D, Gros P. The Bcg/Ity/Lsh
locus: Genetic transfer of resistance to infections in C57B1/6J mice transgenic for
the N r a m p l G l y 1 6 9 allele. Infect Immun 1996; 64: 2923-2929.
29. Swanson RN, O'Brien AD. Genetic control of the innate resistance of mice to
Salmonella typhimurium: Ity gene is expressed in vivo by 24 hours after infection.
J of Immunol 1983; 131: 3014-3020.
30. Kagaya K, Watanabe K, Fukazawa Y. Capacity of recombinant gamma interferon
to activate macrophages for Salmonella-killing activity. Infect Immun 1989; 57:
609-615.
31. Mastroeni P, Arena A, Costa GB, Liberto MC, Bonina L, Hormaeche CE. Serum
TNFalpha in mouse typhoid and enhancement of a Salmonella infection by anti-
TNFalpha antibodies. Microb Pathogen 1991; 11: 33-38.
32. Morrissey PJ, Charrier К. Treatment of mice with IL-1 before infection increases
resistance to a lethal challenge with Salmonella typhimurium - the effect correla
tes with the resistance allele at the ity locus. J Immunol 1994; 153: 212-219.
33. Morrissey PJ, Charrier К. GM-CSF administration augments the survival of ΓΓΥ-
resistant A/J mice, but not ΓΤΥ-susceptible C57B1/6 mice to a lethal challenge
with Salmonella typhimurium. J Immunol 1990; 14: 557-561.
34. Eckmann L, Fierer J, Kagnoff MF. Genetically resistant (ItyR) and susceptible
(ItyS) congenie mouse strains show similar cytokine responses following infec
tion with Salmonella Dublin. J Immunol 1996; 156: 2894-2900.
35. Pie S, Matsiota-Bernard P, Truffa-Bachi Ρ, Nauciel С. Gamma-interferon and

General Discussion 125
interleukin-10 gene expression in innately susceptible and resistant mice during
the early phase of Salmonella typhimurium infection. Infect Immun 1996; 64:
849-854.
36. Peel JE. Induction of circulating tumor necrosis factor cannot be demonstrated
during septicemic salmonellosis in calves. Infect Immun 1990; 58: 439^42.
37. Jotwani R, Tanaka Y, Watanabe K, Tanaka K, Kato N, Ueno К. Cytokine stimu
lation during Salmonella typhimurium sepsis in ItyS mice. J Med Microbiol 1995;
42: 348-52.
38. Greisman SE, Hornick RB, Wagner HN, Woodward TE. The integrity of the
endotoxin tolerance mechanisms during typhoid fever in man. Trans Assoc Am
Physicians 1967; 80: 250-258.
39. Greisman SE. The role of endotoxin during typhoid fever and tularemia in man. J
Clin Invest 1969; 48: 613-629.
40. Morrissey PJ, Charrier К, Vogel SN. Exogenous tumor necrosis factor alpha and
interleukin-1 alpha increase resistance to Salmonella typhimurium: efficacy is
influenced by the Ity and LPS loci. Infect Immun 1995; 63: 3158-3165.
41. Licht TR, Krogfelt KA, Cohen PS, Poulsen LK, Urbance J, Molin S. Role of lipo-
polysaccharide in colonization of the mouse intestine by Salmonella typhimurium
studied by in situ hybridization. Infect Immun 1996; 64: 3811-3817.
42. Hormaeche CE, Maskell DJ, Harrington К, Joysey H, Brock J. Mechanisms of
natural resistance to mouse typhoid. Bull Europ Physiopath Resp 1983; 19: 137-
142.
43. Hormaeche CE, Villareal В, Mastroeni Ρ, Dougan G, Chatfield SN. Immunity
mechanisms in experimental salmonellosis. In: Cabello F, Hormaeche С,
Mastroeni Ρ, Bonina L, ed. Biology of Salmonella. New York: Plenum Press,
1993:223-235.
44. Langermans JAM, Van der Hulst MB, Nibbering PH, Van Fürth R. Activation of
mouse peritoneal macrophages during infection with Salmonella typhimurium
does not result in enhanced intracellular killing. J Immunol 1990; 144: 4340-
4346.
45. Van Dissel JT, Stikkelbroeck JJM, Van den Barselaar MT, Sluiter W, Leijh PCJ,
Van Fürth R. Divergent changes in antimicrobial activity after immunologic acti
vation of mouse peritoneal macrophages. J Immunol 1987; 139: 1665-1672.
46. Van Dissel JT, Leijh PCJ, Van Fürth R. Differences in initial rate of intracellular
killing of Salmonella typhimurium by resident peritoneal macrophages from
various mouse strains. J Immunol 1985; 134: 3404-3410.

126 Chapter 9
47. Van Dissel JT, Stikkelbroeck JJM, Sluiter W, Leijh PCJ, Van Fürth R. Differen
ces in initial rate of intracellular killing of Salmonella typhimurium by granulo
cytes of salmonella susceptible C57B1/10 mice and salmonella resistant CBA
mice. J Immunol 1986; 136: 1074-1080.
48. O'Brien AD, Scher I, Formal SB. Effect of silica on the innate resistance of inbred
mice to Salmonella typhimurium infection. Infect Immun 1979; 25: 513-520.
49. Conlan JW, North RJ. Early pathogenesis of infection in the liver with the facul
tative intracellular bacteria Listeria monocytogenes, Francisella tularensis, and
Salmonella typhimurium involves lysis of infected hepatocytes by leukocytes.
Infect Immun 1992; 60: 5164-5171.
50. Conlan JW, North RJ. Neutrophil-mediated lysis of infected hepatocytes. ASM
News 1993; 59: 563-567.
51. Hsu HS. Pathogenesis and immunity in murine salmonellosis. Microbiol Rev
1989; 53: 390-409.
52. Conlan JW. Neutrophils prevent extracellular colonization of the liver micro-
vasculature by Salmonella typhimurium. Infect Immun 1996; 64: 1043-1047.
53. Hormaeche CE, Mastroeni P, Arena A, Uddin J, Joysey HS. T-cells do not media
te the initial suppression of a salmonella infection in the RES. Immunology 1990;
70: 247-250.
54. Muotiala A, Makela PH. The role of IFN-γ in murine Salmonella typhimurium
infection. Microb Pathogen 1990; 8: 135-141.
55. Mackaness GB. Cellular resistance to infection. J Exp Med 1962; 116: 381-417.
56. Mackaness GB. The immunological basis of acquired cellular resistance. J Exp
Med 1964; 120: 105-120.
57. Mackaness GB, Blanden RV. Cellular immunity. Progr Allergy 1967; 11: 89-140.
58. Mackaness GB. Resistance to intracellular infection. J Infect Dis 1971; 123: 439-
445.
59. Langermans JAM, Nibbering PH, Van Fürth R. Activation of murine peritoneal
macrophages by Salmonella typhimurium. Agents Actions 1989; 26: 208-210.
60. Senterfitt VC, Shands JW. Salmonellosis in mice infected with Mycobacterium
tuberculosis BCG. I. Role of endotoxin in infection. J Bacterid 1968; 96: 287-
292.
61. Schafer R, Eisenstein TK. Induction of natural killer cell activity by a Salmonella
vaccine. FASEB J 1988; 2: A677.
62. Vladoianu IR, Chang HR, Pechere JC. Expression of host resistance to
Salmonella typhi and Salmonella typhimurium: bacterial survival within macro-

General Discussion 127
phages of murine and human origin. Microb Pathogen 1990; 8: 83-90.
63. Lin FR, Hsu HS, Mumaw VR, Moncure CW. Confirmation of destruction of
salmonellae within murine peritoneal exudate cells by immunocytochemical
technique. Immunology 1989; 67: 394^00.
64. Degre M, Bukholm G, Czamlecki CW. In vitro treatment of HEp-2 cells with
human tumor necrosis factor-alpha and human interferons reduces invasiveness of
Salmonella typhimurium. J Biol Regul Homeost Agents 1989; 3: 1-7.
65. Portnoy DA, Schreiber RD, Connelly P, Tilney LG. γ-Interferon limits access of
Listeria monocytogenes to the macrophage cytoplasm. J Exp Med 1989; 170:
2141-2146.
66. Gregory SH, Wing EJ. IFN-gamma inhibits the replication of Listeria
monocytogenes in hepatocytes. J Immunol 1993; 151: 1401-1409.
67. Hess J, Ladel C, Miko D, Kaufmann SH. Salmonella typhimurium aroA- infection
in gene targeted immunodeficient mice: major role of CD4+ TCR-alpha beta cells
and IFN-gamma in bacterial clearance independent of intracellular location. J
Immunol 1996; 156: 3321-3326.
68. Barnes PF, Chatterjee D, Brennan PJ, Rea TH, Modlin RL. Tumor necrosis factor
production in patients with leprosy. Infect Immun 1992; 60: 1441-1446.
69. Havell EA. Production of tumor necrosis factor during murine listeriosis. J
Immunol 1987; 139: 4225-4231.
70. Liew FJ, Parkinson C, Millot S, Severn A, Carrier M. Tumor necrosis factor
(TNF) in leishmaniasis l.TNF mediates host protection against cutaneous leish
maniasis. Immunology 1990; 69: 570-573.
71. Nakane A, Minagawa T, Kato K. Endogenous tumor necrosis factor (cachectin) is
essential to host resistance against Listeria monocytogenes infection. Infect
Immun 1988; 56: 2563-2569.
72. Silva CL, Foss NT. Tumor necrosis factor in leprosy patients. J Infect Dis 1989;
159: 787-790.
73. Titus RG, Sherry B, Cerami A. Tumor necrosis factor plays a protective role in
experimental murine cutaneous leishmaniasis. J Exp Med 1989; 170: 2097-2104.
74. Pfeffer К, Matsuyama Τ, Kundig TM, Wakeham A, Kishihara К, Shahinian A,
Wiegmann К, Ohashi PS, Kronke M, Мак TW. Mice deficient for the 55 kDa
tumor necrosis factor receptor are resistant to endotoxic shock, yet succumb to L.
monocytogenes infection. Cell 1993; 73: 457-467.
75. Rothe J, Lesslauer W, Lotscher Η, Lang Y, Koebel Ρ, Kontgen F, Althage A,
Zinkernagel R, Steinmetz M, Bluethmann H. Mice lacking the tumour necrosis

128 Chapter 9
factor receptor-1 are resistant to TNF-mediated toxicity but highly susceptible to
infection by Listeria monocytogenes. Nature 1993; 364: 798-802.
76. Mosmann TR, Cherwinski H, Bond MW, Giedlin MA, Coffman RL. Two types
of murine helper T-cell clone. I. Definition according to profiles of lymphokine
activities and secreted proteins. J Immunol 1986; 136: 2348-2357.
77. Allen JE, Maizels RM. Thl-Th2: reliable paradigm or dangerous dogma. Immu
nol Today 1997; 18: 387-392.
78. Black RA, Rauch CT, Kozlosky CJ, Peschon JJ, Slack JL, Wolfson MF, Castner
BJ, Stocking KL, Reddy P, Srinivasan S, Nelson N, Boiani N, Schooley KA,
Gerhart M, Davis R, Fitzner JN, Johnson RS, Paxton RJ, March CJ, Cerretti DP.
A metalloproteinase disintegrin that releases tumour necrosis factor-alpha from
cells. Nature 1997; 385: 729-733.
79. Eigler A, Sinha B, Hartmann G, Endres S. Taming TNF: strategies to restrain this
proinflammatory cytokine. Immunology Today 1997; 18: 487-492.
80. Bancroft GJ, Sheehan KCF, Schreiber RD, Unanue ER. Tumor necrosis factor is
involved in the Τ cell-independent pathway of macrophage activation in SCID
mice. J Immunol 1989; 143: 127-130.
81. Langermans JAM, Van der Hulst MEB, Nibbering PH, Van Fürth R. Endogenous
tumor necrosis factor alpha is required for enhanced antimicrobial activity against
Toxoplasma gondii and Listeria monocytogenes in recombinant gamma
interferon-treated mice. Infect Immu 1992; 60: 5107-5112.
82. Mastroeni P, Villareal-Ramos B, Hormaeche CE. Effect of late administration of
anti-TNFalpha antibodies on a Salmonella infection in the mouse model. Microb
Pathogen 1993; 14: 473^80.
83. Nakano Y, Onozuka К, Terada Y, Shinomiya H, Nakano M. Protective effect of
recombinant tumor necrosis factor-α in murine salmonellosis. J Immunol 1990;
144: 1935-1941.
84. Nauciel C, Espinasse-Maes F. Role of gamma-interferon and tumor necrosis
factor alpha in resistance to Salmonella typhimurium infection. Infect Immun
1992; 60:450-454.
85. Tite JP, Dougan G, Chatfield SN. The involvement of tumor necrosis factor in
immunity to Salmonella infection. J Immunol 1991; 147: 3161-3164.
86. Mastroeni P, Skepper JN, Hormaeche CE. Effect of anti-tumor necrosis factor
alpha antibodies on histopathology of primary Salmonella infections. Infect
Immun 1995; 63: 3674-3682.
87. Degré M, Bukholm G. Effect of tumor necrosis factor-alpha on infection with

General Discussion 129
Salmonella typhimurium in a mouse model. J Biol Regul Homeost Agents 1990;
4: 157-161.
88. Langermans JAM. The cytokine network, macrophage activation, and enhanced
antimicrobial activity. Leiden: thesis, 1992:
89. Murray HW. Interferon-gamma, the activated macrophage, and host defense
against microbial challenge. Ann Int Med 1988; 108: 595-608.
90. Cavaillon JM. Cytokines and macrophages. Biomed & Pharmacother 1994; 48:
445-453.
91. Huang S, Hendriks W, Althage A, Hemm S, Bluetmann H, Kamijo R, Vilcek J,
Zinkernagell RM, Aguet M. Immune response in mice that lack the interferon-γ
receptor. Science 1993; 259: 1742-1745.
92. Pie S, Truffa-Bachi Ρ, Pia M, Nauciel С. Thl response in Salmonella
typhimurium-inkctcd mice with a high or low rate of bacterial clearance. Infect
Immun 1997; 65:4509-4514.
93. Iizawa Y, Wagner RD, Czuprynski CJ. Analysis of cytokine mRNA expression in
Listeria-resistant C57BL/6 and Listeria-susceptible A/J mice during Listeria
monocytogenes infection. Infect Immun 1993; 61: 3739-44.
94. Neta R, Salvin SB. Resistance and susceptibility to infection in inbred murine
strains. Cell Immunol 1983; 75: 173-180.
95. Ramarathinam L, Niesei DW, Klimpel GR. Ity influences the production of IFN-y
by murine plenocytes stimulated in vitro with Salmonella typhimurium. J of
Immunol 1993; 150: 3965-3972.
96. Benbernou N, Nauciel С Influence of mouse genotype and bacterial virulence in
the generation of interferon-gamma-producing cells during the early phase of
Salmonella typhimurium infection. Immunol 1994; 83: 245-249.
97. Ramarathinam L, Shaban RA, Niesei DW, Klimpel GR. Interferon gamma (IFN-
y) production in the gut-associated lymphoid tissue and spleen following oral
Salmonella typhimurium challenge. Microb Pathogen 1991; 11: 347-356.
98. Muotiala A, Makela PH. Role of gamma interferon in late stages of murine sal
monellosis. Infect Immun 1993; 61: 4248^253.
99. Heinzel FP. The role of interferon-gamma in the pathology of experimental endo-
toxemia. J Immunol 1990; 145: 2920-2924.
100. Langermans JAM, Van der Hulst MEB, Nibbering PH, Hiemstra PS, Fransen L,
Van Fürth R. IFN-y-induced L-arginine-dependent toxoplasmastatic activity in
murine peritoneal macrophages is mediated by endogenous tumor necrosis factor-
a. J Immunol 1992; 148: 568-574.

130 Chapter 9
101. Langermans JAM, Nibbering PH, Van der Hulst MEB, Van Fürth R. Microbici
dal activities of Salmonella typhimurium- and interferon gamma activated mouse
peritoneal macrophages. Pathobiology 1991; 59: 189-193.
102. Osanto S, Van den Barselaar MT, Van Dissel JT. Increased intracellular killing of
bacteria in vitro by monocytes of patients with metastatic melanoma before and
during treatment with interferon-gamma and interferon-alpha. Eur j Cancer 1991;
27: 478-482.
103. Degre M, Bukholm G. Orally administered interferon-gamma but not tumor
necrosis factor-alpha suppress infection with Salmonella typhimurium in a mouse
model. J Biol Reg Homeost Agents 1995; 9: 15-20.
104. Dinarello CA. Interleukin-1 and interleukin-1 antagonism. Blood 1991; 77: 1627-
1652.
105. Kullberg BJ, Van 't Wout JW, Van Fürth R. Role of granulocytes in enhanced
host resistance to Candida albicans induced by recombinant interleukin-1. Infect
Immun 1990; 58: 3319-3324.
106. Czuprynski CJ, Brown JF. Recombinant murine interleukin-la enhancement of
nonspecific antibacterial resistance. Infect Immun 1987; 55: 2061-2065.
107. Van der Meer JWM, Barza M, Wolff SM, Dinarello CA. A low dose of recombi
nant interleukin 1 protects granulocytopenic mice from lethal Gram-negative
infection. Proc Natl Acad Sci USA 1988; 85: 1620-1623.
108. Curfs JHAJ, van der Meer JWM, Sauerwein R, Eling WMC. Low dosages of
interleukin-1 protect mice against lethal cerebral malaria. In: Dinarello CA, Klu
ger M, Oppenheim J, Powanda M, ed. Interleukin-1 and related cytokines. New
York: Alan Liss, 1990.
109. Vogels MTE, Mensink EJBM, Ye K, Boerman OC, Verschuren CMM, Dinarello
CA, Van der Meer JWM. Differential gene expression for IL-1 receptor anta
gonist, IL-1, and TNF receptors and IL-1 and TNF synthesis may explain IL-1-
induced resistance to infection. J Immunol 1994; 153: 5772-5780.
110. Vogels MTE, Van Rooyen N, Bemelmans MHA, Jansen M, Van der Meer JWM.
Interleukin-1 (IL-l)-induced resistance to bacterial infection: role of the macro
phage and soluble TNF receptors. In: Vogels MTE, ed. Mechanisms of enhanced
natural resistance to infection induced by interleukin-1. Thesis: Catholic Univer
sity Nijmegen, 1994.
111. Umezawa K, Akaike T, Fujii S, Suga M, Setoguchi К, Ozawa A, Maeda H.
Induction of nitric oxide synthesis and xanthine oxidase and their roles in the anti
microbial mechanism against Salmonella typhimurium infection in mice. Infect

General Discussion 131
Immun 1997; 65: 2932-2940.
112. Vannier E, Miller LC, Dinarello CA. Coordinated antiinflammatory effects of
interleukin 4: interleukin 4 suppresses interleukin 1 production but up-regulates
gene expression and synthesis of interleukin 1 receptor antagonist. Proc Natl
Acad Sci USA 1992; 89: 4076-W80.
113. Everest P, Allen J, Papakonstantinopoulou A, Mastroeni P, Roberts M, Dougan
G. Salmonella typhimurium infections in mice deficient in Interleukin-4 produc
tion: role of IL-4 in infection associated pathology. J Immunol 1997; 159: 1820-
1827.
114. Denich K, Borlin P, O'Hanley PD, Howard M, Heath AW. Expression of the
murine interleukin-4 gene in an attenuated aroA strain of Salmonella typhimu
rium: persistence and immune response in BALB/c mice and susceptibility to
macrophage killing. Infect Immun 1993; 61:4818-4827.
115. Schindler R, Mancilla J, Endres S. Correlations and interactions in the production
of interleukin-6 (IL-6), interleukin-1 (IL-1) and tumor necrosis factor (TNF) in
human blood mononuclear cells: IL-6 suppresses IL-1 and TNF. Blood 1990; 75:
40-47.
116. Tilg H, Dinarello CA, Mier JW. IL-6 and APPs: anti-inflammatory and immuno-
suppresive mediators. Immunol Today 1997; 18: 429-432.
117. Dalrymple SA, Lucían LA, Slattery R, McNeil Τ, Aud DM, Fuchino S, Lee F,
Murray R. Interleukin-6-deficient mice are highly susceptible to Listeria mono
cytogenes infection: correlation with inefficient neutrophilia. Infect Immun 1995;
63: 2262-2268.
118. Czuprynski CJ, Haak-Frendscho M, Maroushek N, Brown JF. Effects of recom
binant human interleukin-6 alone and in combination with recombinant inter
leukin-la and tumor necrosis factor-alpha on antibacterial resistance in mice.
Antimicrob Agents Chemother 1992; 36: 68-70.
119. Howard M, O'Garra A, Ishida H, de Waal Malefyt R, de Vries J. Biological
properties of interleukin-10. J Clin Immunol 1992; 12: 239-247.
120. Goldman M, Stordeur P. Interleukin-10 as an anti-stress cytokine. Eur Cytokine
Netw 1997; 8: 301-302.
121. Powrie F, Menon S, Coffman RL. Interleukin-4 and interleukin-10 synergize to
inhibit cell-mediated immunity in vivo. Eur J Immunol 1993; 23: 3043-3049.
122. Arai Τ, Hiromatsu К, Nishimura Η, Kimura Y, Kobayashi Ν, Ishida Η, Nimura
Y, Yoshikai Y. Effects of in vivo administration of anti-EL-10 monoclonal anti
body on the host defence mechanism against murine Salmonella infection. Immu-

132 Chapter 9
nology 1995; 85: 381-388.
123. Hall SS. IL-12 at the crossroads. Science 1995; 268: 1432-1434.
124. Van der Pouw Kraan ТСГМ, Boeije LCM, Smeenk RJT, Wijdenes J, Aarden LA.
Prostaglandin-E2 is a potent inhibitor of human interleukin-12 production. J Exp
Med 1995; 181:775-779.
125. Skeen MJ, Miller MA, Shinnick TM, Ziegler HK. Regulation of murine macro
phage IL-12 production. Activation of macrophages in vivo, restimulation in vitro
and modulation by other cytokines. J Immunol 1996; 156: 1196-1206.
126. Murray HW, Hariprashad J. Interleukin-12 is effective treatment for an esta
blished systemic intracellular infection: experimental visceral leishmaniasis. J
Exp Med 1995; 181: 387-391.
127. Wang Z, Zheng S, Cony DB, Dalton DK, Seder RA, Reiner SL, Locksley RM.
Interferon-gamma independent effects of interleukin-12 administered during acute
or established infection due to Leishmania major. Proc Natl Acad Sci USA 1994;
91: 12932-12936.
128. Chong C, Bost KL, Clements JD. Differential production of interleukin-12
mRNA by murine macrophages in response to viable or killed Salmonella spp.
Infect Immun 1996; 64: 1154-1160.
129. Kincy-Cain T, Clements JD, Bost KL. Endogenous and exogenous interleukin-12
augment the protective immune response in mice orally challenged with Salmo
nella dublin. Infect Immun 1996; 64: 1437-1440.
130. Mastroeni P, Harrison JA, Chabalgoity JA, Hormaeche CE. Effect of interleukin-
12 neutralization on host resistance and gamma-interferon production in mouse
typhoid. Infect Immun 1996; 64: 189-196.
131. Kincy-Cain T, Bost KL. Substance P-induced IL-12 production by murine macro
phages. J Immunol 1997; 158: 2334-2339.
132. Kincy-Cain T, Bost KL. Increased susceptibility of mice to Salmonella infection
following in vivo treatment with the substance Ρ antagonist, spantide Π. J Immu
nol 1996; 157: 255-264.
133. Bost KL, Clements JD. Intracellular Salmonella dublin induces sustantial secre
tion of the 40-kilodalton subunit of interleukin-12 (IL-12) but minimal secretion
of IL-12 as a 70-kilodalton protein in murine macrophages. Infect Immun 1997;
65:3186-3192.
134. Heinzel FP, Hujer AM, Ahmed FN, Rerko RM. In vivo production and function
of IL-12 p40 homodimers. J Immunol 1997; 158: 4381-4388.
135. Dale DC, Liles WC, Summer WR, Nelson S. Review: Granulocyte colony-stimu-

General Discussion 133
lating factor. Role and relationships in infectious diseases. J Infect Dis 1995; 172:
1061-1075.
136. Kullberg В J, Netea MG, Curfs JHAJ, Keuter M, Meis JFGM, Van der Meer
JWM. Recombinant murine granulocyte colony-stimulating factor protects
against acute disseminated Candida albicans infection in non-neutropenic mice. J
Infect Dis 1998; 177: 175-181.
137. Nelson S. Role of granulocyte colony-stimulating factor in the immune response
to acute bacterial infection in the nonneutropenic host: an overview. Clin Infect
Dis 1994; 18(suppl2): S197-204.
138. Barsig J, Bundschuh DS, Härtung Τ, Bauhofer Α, Sauer A, Wendel A. Control of
fecal peritoneal infection in mice by colony-stimulating factors. J Infect Dis 1996;
174: 790-799.
139. Appelberg R. G-CSF in a murine model of Mycobacterium avium infection. In:
Andresen J, ed. Abstracts G-CSF Research Meeting. Santa Monica, CA: Amgen,
1996;36-38.
140. Serushago BA, Yoshikai Y, Handa T, Mitsuyama M, Muramori K, Nomoto K.
Effect of recombinant human granulocyte colony-stimulating factor (rh G-CSF)
on murine resistance to Listeria monocytogenes. Immunology 1992; 75: 475-480.
141. Shinomiya N, Tsuru S, Katsura Y, Kayashima S, Nomoto K. Enhanced resistance
against Listeria monocytogenes achieved by pretreatment with granulocyte colo
ny-stimulating factor. Infect Immun 1991; 59:4740-4743.
142. Kitabayashi A, Hirokawa M, Hatano Y, Lee M, Kuroki J, Niitsu H, Miura AB.
Granulocyte colony-stimulating factor downregulates allogeneic immune respon
ses by posttranscriptional inhibition of tumor necrosis factor-alpha production.
Blood 1995; 86: 2220-2227.
143. Härtung Τ, Docke WD, Gantner F, Krieger G, Sauer A, Stevens Ρ, Volk HD,
Wendel Α. Effect of granulocyte colony-stimulating factor treatment on ex vivo
blood cytokine response in human volunteers. Blood 1995; 85: 2482-2489.
144. Dinarello CA, Cannon JG, Mancilla J, Bishai I, Lees J, Coceani F. Interleukin-6
as an endogenous pyrogen: induction of prostaglandin E2 in brain but not in peri
pheral blood mononuclear cells. Brain Res 1991; 562: 199-206.
145. Hack CE, De Groot ER, Felt-Bersma RJF. Increased plasma levels of interleukin-
6 in sepsis. Blood 1989; 74: 1704-1710.
146. Waage A, Brandtzaeg P, Halstensen A, Kierulf P, Espevik T. The complex
pattern of cytokines in serum from patients with meningococcal septic shock.
Association between interleukin-6, interleukin-1 and fatal outcome. J Exp Med

134 Chapter 9
1989; 169: 333-338.
147. biada M, Tojo H, Kawata S, Tarui S, Okamoto M. Induction of group Il-like
phospholipase A2 by lipopolysaccharide in the liver of BCG-primed rats.
Biochem Biophys Res Commun 1991; 174: 1077-1083.
148. Morgan DW, Anderson CM, Meyers KP. Recombinant human type Π phospholi
pase A2 lacks edema producing activity in rat. Eur J Pharmacol 1993; 235: 45-50.
149. Hack CE, Wolbink GJ, Schalkwijk С, Speijer H, Hermens WT, Van den Bosch
H. A role for secretory phospholipase A2 and C-reactive protein in the removal of
injured cells. Immunology Today 1997; 18: 111-115.
150. Pruzanski W, Vadas P. Secretory synovial fluid phospholipase A2 and its role in
the pathogenesis of inflammation in arthritis. J Rheumatol 1988; 15: 1601-1603.
151. Kuslys T, Vishwanath BS, Frey FJ, Frey BM. Differences in phospholipase A2
activity between males and females and Asian Indians and Caucasians. Eur J Clin
Invest 1996;26:310-315.
152. Lucas R, Juillard P, Decoster E, Redard M, Burger D, Donaty Y, Giroud C,
Monso-Hinard C, De Kesel T, Buurman WA, Moore MW, Dayer JM, Fiers W,
Bluethmann H, Grau GE. Crucial role of tumor necrosis factor (TNF) receptor 2
and membrane-bound TNF in experimental cerebral malaria. Eur J Immunol
1997; 27: 1719-1725.
153. Van der Poll T, Coyle SM, Moldawer LL, Lowry SF. Changes in endotoxin-indu-
ced cytokine production by whole blood after in vivo exposure of normal humans
to endotoxin. J Infect Dis 1996; 174: 1356-1360.
154. Mengozzi M, Ghezzi P. Cytokine down-regulation in endotoxin tolerance. Eur
Cytokine Netw 1993; 4: 89-98.
155. Luger A, Graf H, Schwarz HP, Stummvoll HK, Luger TA. Decreased serum
interleukin-1 activity and monocyte interleukin-1 production in patients with fatal
sepsis. Crit Care Med 1986; 14:458-461.
156. Simpson SQ, Modi H, Balk RA, Bone RC, Casey LC. Reduced alveolar macro
phage production of tumor necrosis factor during sepsis in mice and man. Crit
Care Med 1991; 19: 1060-1066.
157. Srugo I, Berger A, Lapidot Z, Katz R, Pollak S. Interleukin-1 secretion by blood
monocytes of septic premature infants. Infection 1991; 3: 150-154.
158. Helminen M. Interleukin-1 production from peripheral blood monocytes in septic
infections in children. Scand J Infect Dis 1991; 23: 607-611.
159. Rozenbaum M, Katz R, Rozner I, Pollack S. Decreased interleukin-1 activity
released from circulating monocytes of patients with familial Mediterranean fever

General Discussion 135
during in vitro stimulation by lipopolysaccharide. J Rheumatol 1992; 19: 416-
418.
160. Schattner A, Lachmi M, Livneh A, Pras M, Hahn T. Tumor necrosis factor in
familial Mediterranean fever. Am J Med 1991; 90:434-438.
161. Drenth JPH, Van Uum SHM, Van Deuren M, Pesman GJ, Van der Ven-
Jongekrijg J, Van der Meer JWM. Endurance run increases circulating IL-6 and
IL-lra but downregulates ex vivo TNF-a and IL-lß production. J Appi Physiol
1995; 79: 1497-1503.
162. Kruimel JW, Naber AHJ, Van der Vliet AJ, Buskens FGM, Pesman G, Sweep F,
Liem T, Kho HG, Van der Meer JWM, Jansen JBMJ. Effects of abdominal surge
ry on cytokines in a clinical trial designed to study the effects of parenteral admi
nistration of lipid emulsions. Neth J Med 1994; 45: A17-A18.
163. Granowitz EV, Porat R, Mier JW, Orencole SF, Kaplanski G. Lynch EA, Ye K,
Vannier E, Wolff SM, Dinarello CA. Intravenous endotoxin suppresses the cyto
kine response of peripheral blood mononuclear cells of healthy humans. J Immu
nol 1993; 151: 1637-1645.
164. Van der Poll T, de Waal Malefijt R, Coyle SM, Lowry SF. Antiinflammatory
cytokine responses during clinical sepsis and experimental endotoxemia: sequen
tial measurements of plasma soluble interleukin (IL)-l receptor type Π, IL-10 and
IL-13. J Infect Dis 1997; 175: 118-122.
165. Van der Poll T, Jansen J, Endert E, Sauerwein HP, Van Deventer SJH.
Noradrenaline inhibits lipopolysaccharide-induced tumor necrosis factor and
interleukin-6 production in human whole blood. Infect Immun 1994; 62: 2046-
2050.
166. Randow F, Syrbe U, Meisel C, Krausch D, Zuckermann H, Platzer С, Volk HD.
Mechanism of endotoxin desensitization: involvement of interleukin-10 and trans
forming growth factor-ß. J Exp Med 1995; 181: 1887-1892.
167. Frankenberger M, Pechumer H, Loms Ziegler-Heitbrock HW. Interleukin-10 is
upregulated in LPS tolerance. J Inflamm 1995; 45: 56-63.
168. Loms Ziegler-Heitbrock HW, Frankenberger M, Wedel A. Tolerance to lipopoly
saccharide in human blood monocytes. Immunobiol 1995; 193: 217-223.
169. Docke WD, Randow F, Krausch D, Khusru A, Reinke P, Volk HD, Кох W.
Monocyte deactivation in septic patients: restoration by IFN-gamma treatment.
Nature Med 1997; 3: 678-681.
170. Van Deuren M. Kinetics of tumour necrosis factor alpha, soluble tumour necrosis
factor receptors, interleukin-1-beta and its receptor antagonist during serious

136 Chapter 9
infections. Eur J Clin Microbiol Infect Dis 1994; 13 Suppl 1: 12-16.
171. Van Deuren M, Van der Ven-Jongekrijg J, Demacker PNM, Bartelink AKM, Van
Dalen R, Sauerwein RW, Gallati H, Vannice JL, Van der Meer JWM. Differential
expression of proinflammatory cytokines and their inhibitors during the course of
meningococcal infections. J Infect Dis 1994; 169: 157-161.
172. Vogels MTE, Van der Meer JWM. Use of immune modulators in nonspecific
therapy of bacterial infections. Antimicrob Agents Chemother 1992; 36: 1-5.
173. Kullberg BJ, Van 't Wout JW, Van Fürth R. No effect of recombinant human
interleukin-1 on the numbers of peripheral blood and peritoneal leukocytes during
an acute inflammation. Inflammation 1991; 15: 457-470.
174. Lajarin F, Rubio G, Galvez J, Garcia-Penarrubia P. Adhesion, invasion and intra
cellular replication of Salmonella typhimurium in a murine hepatocyte cell line.
Effect of cytokines and LPS on antibacterial activity of hepatocytes. Microb
Pathog 1996; 21: 319-329.
175. Van 't Wout JW, Poell R, Van Fürth R. The role of BCG-PPD-activated macro
phages in resistance against systemic candidiasis in mice. Scand J Immunol 1992;
36:713-719.
176. Sisson SD, Dinarello CA. Production of interleukin-1 alpha, interleukin-1 beta
and tumor necrosis factor by human mononuclear cells stimulated with granulo
cyte-macrophage colony-stimulating factor. Blood 1988; 72: 1368-1374.
177. Pollmacher T, Korth C, Mullington J, Schreiber W, Sauer J, Vedder H, Galanos
С, Holsboer F. Effects of granulocyte colony-stimulating factor on plasma cytoki
ne and cytokine receptor levels and on the in vivo host response to endotoxin in
healthy men. Blood 1996; 87: 900-905.
178. Pajkrt D, Manten A, Van der Poll T, Van Buul TMM, Jansen J, Ten Cate WJ,
Van Deventer SJ. Modulation of cytokine release and neutrophil function by gra
nulocyte colony-stimulating factor during endotoxemia in humans. Blood 1997;
90: 1415-1424.
179. Gorgen I, Härtung Τ, Leist M, Niehorster M, Tiegs G, Uhlig S, Weitzel F,
Wendel A. Granulocyte colony-stimulating factor treatment protects rodents
against lipopolysaccharide-induced toxicity via suppression of systemic tumor
necrosis factor. J Immunol 1992; 149: 918-924.
180. Andresen J. Enhanced antimicrobial activity of polymorphonuclear leukocytes
and antibiotics in the presence of filgrastim. In: Andresen J, ed. Abstracts G-CSF
Research Meeting. Santa Monica, CA: Amgen, 1996;36-38.
181. Harris HW, Grunfeld C, Feingold KR, Read TE, Kane JP, Jones AL, Eichbaum

General Discussion 137
EB, Bland GF, Rapp JH. Chylomicrons alter the fate of endotoxin, decreasing
tumor necrosis factor release and preventing death. J Clin Invest 1993; 91: 1028-
1034.
182. Sinnis P, Willnow TE, Briones MRS, Herz J, Nussenzweig V. Remnant lipopro
teins inhibit malaria sporozoite invasion of hepatocytes. J Exp Med 1996; 184:
945-954.
183. Cundell DR, Gerard NP, Gerard C, Idanpaan-Heikkila I, Tuomanen EI. Strepto
coccus pneumoniae anchor to activated human cells by the receptor for platelet-
activating factor. Nature 1995; 377: 435-438.

138 Chapter 9

Chapter 10
Summary

140 Chapter 10

Summary 141
Typhoid fever is a severe systemic infection, confined to humans and is usually
caused by Salmonella typhi. Mostly children and young adults may contract the disease
in places where water is not properly purified, as the spreading is by the fecal-oral
route. The incidence of typhoid fever is high in Indonesia, with more than 1000 cases
per 100 000 inhabitants. In Indonesia antibiotics reduced the mortality from 18% to 4%.
Joint research between the Catholic University Nijmegen, the Netherlands, and the
Diponegoro University, Semarang, Java, Indonesia was initiated in 1989. The purpose
of this collaboration was to investigate clinical and immunological features of patients
with typhoid fever, as well as to study the pathogenesis of Salmonella infection in an
experimental animal model.
In Chapter 2, the pathogenesis of Salmonella infections is described and the role of
cytokines involved in these infections is reviewed. Whereas Salmonella typhi is not
pathogenic to mice, experimental Salmonella typhimurium infection in mice leading to
a disease similar to human typhoid fever, is used to investigate pathogenesis. Therefore
studies with this mouse model are described in Chapter 2. In contrast to extra cellular
Gram-negative septicemia, where high circulating concentrations of proinflammatory
cytokines such as interleukin-1 (IL-1) and tumor necrosis factor (TNFa) have been con
sidered the main cause of organ damage and death and inhibition of these proinflamma
tory cytokines is beneficial, in intracellular infections this very treatment is deleterious.
TNFa is supposed to be needed locally (in small amounts) and administration of TNFa
inhibited the outgrowth of the microorganisms in Salmonella infections.
Pathogenesis of typhoid fever and other Salmonella infections involves several
processes. Regarding the symptoms especially fever in typhoid fever, pyrogenic cyto
kines or other pyrogenic mediators would be expected to be present in the circulation. It
is generally believed that the release of prostaglandin (PGE2) in the brain is the final
pathway in the pathogenesis of fever and this is thought to be induced by circulating
proinflammatory cytokines. These so-called endogenous pyrogens are produced at the
site of infection. In Chapter 3 we report low concentrations of circulating proinflam
matory cytokines, but high concentrations of the antiinflammatory and non-pyrogenic
interleukin-1 receptor antagonist (IL-IRA) and soluble tumor necrosis factor receptors
(sTNFRI and sTNFRII) during the acute phase of typhoid fever. Since pyrogenic cyto
kines were not found in febrile patients with typhoid fever, it was suggested that other
mechanisms were involved. These included other mediators such as secretory phospho-
lipase A2 (sPLA2). Phospholipases are lipolytic enzymes which catalyse the degrada-

142 Chapter 10
tion of phospholipids. The release of sPLA2 is induced by interleukin-1 (IL-1) and
TNFct and the enzyme is able to mediate the production of arachidonic acid and PGE2.
As intraventricular injection of inhibitors of sPLA2 had been shown to suppress fever
in rabbits, sPLA2 could play a role as a circulating pyrogen. In Chapter 4K we indeed
found elevated concentrations of extracellular group II 14-kDa sPLA2 during the acute
phase of typhoid fever. Concentrations of sPLA2 fell to recovery. We considered that
this bioactive sPLA2 might be responsible for the symptoms of typhoid fever.
In the concept of fever, it is important to note that although concentrations of circu
lating pyrogenic cytokines were low in the acute febrile phase of typhoid fever, mRNA
for TNFa and EL-18 was found to be present in circulating whole blood cells (Chapter
5). Another hypothesis is that these circulating cells themselves produce or induce
pyrogenic cytokines in the central nervous system.
Also in Chapter 3, we report down-regulation of the capacity of circulating blood
cells to produce proinflammatory cytokines after stimulation by lipopolysaccharide
(LPS) in the acute phase of typhoid fever. For this purpose, we have used a whole blood
cytokine test. This depressed cytokine production was not a consequence of a lower
number of white blood cells or exhaustion of these cells during the acute phase of the
infection. After the initial phase of infection there is a posttrancriptional switch-off of
the production of proinflammatory cytokines and a switch-on to an anti-inflammatory
repertoire in mononuclear cells, as antiinflammatory factors such as IL-IRA were not
suppressed.
To investigate the mechanism of down-regulated cytokine production in typhoid
fever further, messenger RNA for DL-lß and TNFa in whole blood was measured. In
Chapter 5, the finding of preserved mRNA for IL-Iß and TNFa in the acute phase of
typhoid fever, also denies an exhausted production and is in favor of post-transcriptio-
nal inhibition of cytokine production.
In-vivo down-regulation of cytokine production is a common phenomenon in many
circumstances. Most likely, this is meant to protect against overwhelming cytokinemia,
but down-regulation can impair immune functions. The benefit of down-regulation in
patients with typhoid fever is difficult to understand in view of experimental animal
studies with Salmonella infection, in which the proinflammatory cytokines are required
to survive the infection (Chapter 2). Possibly the deeper downregulated proinflam
matory cytokines in complicated cases could reflect a less effective host defense and
immunotherapy might be needed to improve host defense.

Summary 143
In order to better understand the host defense against Salmonella, we investigated
the cytokine responses in mice (Chapter 6) and compared these with those found in
patients with typhoid fever (Chapters 3, 4 and 5). Without infection, circulating cyto
kine concentrations but also LPS-stimulated ex-vivo production in whole blood were
undetectable. The latter inability to produce cytokines is the reverse of the human situa
tion, since human peripheral blood cells can be stimulated to produce large amounts of
proinflammatory cytokines without infection. We found a general upregulation of pro
inflammatory cytokine production in this animal model during infection, which is also
in contrast with what we have found in humans with typhoid fever, described in
Chapter 3.
In line with other studies from our laboratory, we asked the question whether admi
nistration of recombinant IL-1 would exert a protective effect in a murine model of
Salmonella typhimurium infection (Chapter 6). Administration of rhIL-la led to а
significantly decreased outgrowth of microorganisms in the liver and spleen of mice.
This effect of rhIL-la pretreatment was seen as early as 24 hours after the onset of the
infection and the microbiological effect of a single dose of rhIL-lot lasted throughout
the study. Peritoneal cells produced significantly more cell-associated IL-la 24 hours
after pretreatment with rhIL-la. The early beneficial effect of this pretreatment on
Salmonella infection might have been established by several mechanisms. The fact that
IL-1 pretreatment protects against different types of infection is in favor of a general
phenomenon. We considered this to be macrophage activation.
Neutrophils are able to ingest and kill Salmonella, and they are important in the
outcome of this infection in vivo (Chapter 2). In Chapter 7, we describe that recom
binant murine granulocyte colony-stimulating factor (rmG-CSF) improved survival of
mice with a potentially lethal S. typhimurium infection. However, the outgrowth of S.
typhimurium in the organs of rmG-CSF pretreated mice was not different from that of
control mice. In our experiments, LPS-stimulated TNFoc production of resident perito
neal macrophages was significantly higher in rmG-CSF treated mice, 24 hours after
pretreatment. Histological examination of the liver after infection showed more loca
lized inflammatory foci in the rmG-CSF treated animals. These results show that pre
treatment with rmG-CSF is beneficial in the facultative intracellular S. typhimurium
infection in mice.
As we found a trend for more outgrowth in continuously treated animals, it was
questioned, whether rG-CSF has a beneficial effect on intracellular infections through
enhanced (early) neutrophil recruitment, and whether the (late) antiinflammatory

144 Chapter 10
properties of rG-CSF are deleterious, e.g., through inhibition of TNFa mediated
macrophage activation.
Although Salmonella is a Gram-negative rod, the role of endotoxin in the patho
genesis of infection caused by these organism is controversial. We approached this
problem (Chapter 8) by using low density lipoprotein receptor knock-out (LDLR-/-)
mice, of which the circulating lipoproteins are able to bind and neutralize bacterial LPS,
thereby reducing the induction of proinflammatory cytokines. The presence of high
concentrations of lipoproteins were shown to improve the outcome of S. typhimurium
infection. We show, that neutralization of endotoxin, which in extracellular Klebsiella
infection is the beneficial effect of lipids, is not the mechanism behind the better survi
val of S. typhimurium infection.
We found effects on distribution and later outgrowth. The former may be explained
by lipoproteins directly interfering with Salmonella uptake, for example by coating
Salmonella surface structures with lipids. Also, it may be hypothesized that the LDL-
receptor influences the uptake of bacteria.
For the effect on outgrowth, we found that phagocytosis of Salmonella by perito
neal macrophages, as well as the subsequent intracellular killing did not differ between
mice with or without the LDL-receptor. This suggests that the observed difference in
growth rate is to be explained by an effect through other phagocytic cells than the
peritoneal cells, which we studied.
Conclusions In patients with typhoid fever, we found down-regulation of proinflammatory cyto
kine production, which has been shown to be a common phenomenon in infections and
other situations. This observation supports the notion that immunotherapy may be bene
ficial for Salmonella typhi infection and that such interventions should be aimed at sup-
pletion or upregulation of immunomodulators that enhance host defense to the infec
tion.
We studied immunomodulation in experimental Salmonella typhimurium infection
in mice. It was found that pretreatment with recombinant IL-1 had beneficial effects,
with less outgrowth of microorganisms in the organs of mice. It is hypothesized that
this is due to a distribution effect probably mediated by activated macrophages.
Pretreatment with rmG-CSF had also a favorable effect in mice in terms of survival
to S. typhimurium infection. Outgrowth of microorganisms in the organs however was
not affected. The early effect of rmG-CSF on neutrophil recruitment and activation was

Summary 145
considered the most important mechanism for the survival. In addition, rmG-CSF led to
an increase of the LPS-induced TNFa production by peritoneal macrophages at the
time of infection, which may have contributed to the outcome.
The beneficial effect of lipoproteins on S. typhimurium infection was observed on
both early distribution and subsequent outgrowth of the microorganisms. This indicates
that lipoproteins and their receptors are crucial players in the host defense against
Salmonella infections, in the early distribution as well as in the intracellular killing by
phagocytes.
These observations in experimental S. typhimurium infection in mice give direction
to development of immunotherapy for Salmonella typhi infection in patients.

146 Chapter 10

Chapter 11
Ringkasan

148 Chapter 11

Ringkasan 149
Demam tifoid merupakan penyakit infeksi sistemile yang menyerang manusia dan
pada umumnya disebabkan oleh Salmonella typhi. Penyakit ini terutama menyerang
anak-anak dan dewasa muda dengan cara penyebaran melalui jalur fekal-oral dan oleh
karenanya banyak ditemukan pada daerah yang purifikasi airaya tidak sempuma.
Insidensi demam tifoid di Indonesia tinggi mencapai 1000 kasus per 100.000 penduduk.
Dengan penggunaan antibiotika mortalitas dapat kurangi dari 18% menjadi 4%.
Kerjasama dalam bidang penelitian antara Universitas Katolik Nijmegen dengan
Universitas Diponegoro Semarang telah diawali sejak tahun 1989. Tujuan dari kerja
sama ini adalah untuk mempelajari secara lebih lengkap gambaran klinik dan imuno-
logik penderita demam tifoid serta penelitian patogénesis infeksi Salmonella pada bina-
tang coba.
Pada Bah 2 dibicarakan patogénesis dan peran sitokin pada infeksi Salmonella.
Berdasarkan pada kenyataan bahwa Salmonella typhi bersifat apatogen pada mencit
maka untuk mempelajari patogénesis demamtifoid pada binatang (mencit) dipergu-
nakan Salmonella typhimurium yang akan memberikan penyakit serupa dengan demam
tifoid pada manusia. Studi dengan mempergunakan mencit sebagai model dijelaskan
pada Bah 2. Berlawanan dengan septikemia kuman Gram-negatif ekstraseluler dimana
konsentrasi tinggi sitokin proinflamasi seperti interleukin-1 (IL-1) dan tumor nekrosis
faktor (TNFa) yang dianggap sebagai penyebab utama dari kerusakan organ dan kema-
tian serta penghambatan sitokin proinflamasi akan bermanfaat, pada infeksi kuman
intraseluler perlakuan semacam ini justru akan membahayakan. TNFa diperkirakan
diperlukan secara lokal (dalam jumlah kecil) dan pemberian TNFa dapat mencegah
pertumbuhan microorganisme pada infeksi Salmonella.
Patogénesis demam tifoid dan infeksi Salmonella lainnya melibatkan beberapa
proses. Dengan terdapatnya gejala febris pada demam tifoid diharapkan terdapatnya
sitokin pirogenik dan mediator pirogenik lainnya pada sirkulasi. Secara umum dianggap
bahwa pelepasan prostaglandin (PGE2) di otak yang dirangsang produksinya oleh
sitokin proinflamasi adalah merupakan jalur akhir dari patogénesis febris. Sitokin yang
disebut juga sebagai pirogen endogenik ini diproduksi secara lokal pada tempat terjadi-
nya infeksi.
Pada Bab 3 kami laporkan terdapatnya sitokin proinflamasi dengan kadar rendali di
dalam sirkulasi disertai dengan terdapatnya sitokin antiinflamasi (non-pirogenik) inter
leukin-1 reseptor antagonis (IL-1RA) serta "soluble tumor necrosis factor receptors"
(sTNFRI dan sTNFRII) dengan kadar tinggi pada fase akut demam tifoid. Dengan tidak

150 Chapter 11
diternukannya sitokin pirogenik pada fase febris pada demam tifoid maka diperkirakan
bahwa terdapat mekanisme lain berperan.
Termasuk diantaranya adalah "secretory phospholipase A2" (PLA2). Fosfolipase
adalah ensim lipolitik yang dapat memecah fosfolipid. Pelepasan sPLA2 dipacu oleh
IL-1 dan TNFcc. Ensim ini dapat berperan sebagai mediator untuk terbentuknya asam
arakbidonat dan PGE2. Pemberian injeksi inhibitor sPLA2 pada kelinci dapat mence-
gah timbulnya panas. Oleh karena itu sPLA2 diperkirakan merupakan zat pirogen pada
sirkulasi. Pada Bab 4 kami dapatkan peningkatan kadar Π 14-kDa sPLA2 pada fase
akut demam tifoid. Sedangkan kadar sPLA2 menurun pada fase menyembuhan. Berda-
sarkan hal tersebut di atas kami perkirakan bahwa sPLA2 merupakan penyebab terjadi-
nya panas pada demam tifoid.
Penting diketahui bahwa walaupun konsentrasi pirogen di dalam sirkulasi rendah
pada fase akut demam tifoid, namun mRNA dari TNFa dan IL-Iß dapat ditemukan
pada sei daran pada sirkulasi {Bab 5). Hipotesa lain yang dapat dikemukakan adalah
bahwa sel-sel ini menghasilkan atau menginduksi sitokin pirogenik di susunan saraf
pusat.
Pada Bab 3 kami juga melaporkan terdapatnya regulasi yang bersifat menekan
kapasitas produksi sitokin proinflamasi pada fase akut demam tifoid. Hal ini dapat
dilihat pada hasil pengukuran kadar sitokin pada sel darah yang diberi stimulasi lipo-
polisakharida (LPS). Depresi produksi sitokin tersebut tidak disebabkan karena jumlah
sel darah putih yang rendah ataupun sebagai akibat kelelahan sei pada fase akut infeksi.
Untuk penelitian ini kami pergunakan darah total.
Setelah fase permulaan infeksi, terjadi "switch-off pada fase pascatranskripsi dari
produksi sitokin proinflamasi dan "switch-on" sitokin antiinflamasi pada sel-sel mono-
nuklear yang dapat dibuktikan dengan tidak tersupresinya sintesa IL-IRA.
Untuk mengatui lebih lanjut mekanisme penekanan produksi sitokin pada demam
tifoid dilakukan pengukuran mRNA dari IL-lß dan TNFa dalam darah. Pada Bab 5
dilaporkan bahwa berdasarkan pemeriksaan mRNA dari IL-lß dan TNFa penekanan
produksi sitokin pada demam tifoid fase akut terjadi fase pascatranskripsi dan bukan
disebabkan karena akibat "kelelahan" sel yang sitokin tersebut.
Regulasi yang bersifat menekan produksi sitokin in vivo merupakan fenomena
yang umum terjadi pada beberapa keadaan. Proces penekanan ini kemungkinan meru
pakan suatu upaya tubuh untuk menghambat terjadinya sitokinemia yang berlebihan
waluapun hai ini dapat pula melumpuhkan fungsi imunitas. Keuntungan dari proses
penekanan pada demam tifoid sulit dimengerti sebab pada infeksi Salmonella binatang

Ringkasan 151
coba sitokin proinflamasi diperlukan hospes untuk bertahan hidup {Bab 2). Penekanan
lebih kuat terhadap sitokin proinflamasi pada penderita demam tifoid dengan komplika-
si dapat merupakan gambaran akan adanya sistem kekebalan tubuh yang kurang efektif
sehingga dengan demikian pemberian khemoterapi diperlukan untuk memperkuat perta-
hanan tubuh.
Dalam upaya untuk mengetahui lebih lanjut pertahanan tubuh terhadap Salmonella
kami melakukan studi respon sitokin pada mencit {Bab 6) untuk dibandingkan dengan
respon pada manusia yang menderita demam tifoid (Bab 3, 4 dan 5). Tanpa adanya in-
feksi, sitokin pada sirkulasi dan juga pada kultur darah (ex-vivo) tidak terdeteksi walau-
pun dengan pemberian stimulasi LPS. Ketidak mampuan untuk memproduksi sitokin
pada mencit tersebut diatas berlawanan dengan manusia dimana pemberian stimulasi
LPS pada sei darah tepi menghasilkan sitokin proinflamasi dalam jumlah yang besar
walaupun tanpa infeksi. Kami juga menemukan bahwa pada mensit yang terinfeksi ter-
jadi stimulasi sitokin proinflamasi. Hai ini juga berlawanan dengan yang ditemukan
pada demam tifoid pada manusia {Bab 3).
Searah dengan penelitian lain pada laboratorium kami, timbul suatu pertanyaan
tentang kemungkinan terdapatnya pengaruh yang bersifat protektif dengan tindakan
pemberian rekombinan IL-1 pada mencit yang terinfeksi Salmonella typhimurium {Bab
6). Pemberian rhIL-la secara signifikan dapat menurunkan pertumbuhan mikroorga-
nisme pada hepar dan lien. Efek pemberian rhIL-la ргарегіакиап terlihat pada 24 jam
pertama setelah infeksi dan efek mikrobiologik dari dosis tunggal rhIL-la bertahan
selama eksperimen berlangsung. Sel-sel peritoneum secara signifikan memproduksi
lebih banyak "cell associated" IL-Ια pada 24 jam setelah pemberian rhIL-la.
Efek menguntungkan pada stadium dini dari pemberian rhIL-la ргарегіакиап
mungkin melalui beberapa mekanisme. Kenyataan bahwa pemberian IL-1 praperlakuan
dapat memberikan perlindungan terhadap berbagai macam infeksi merupakan fenome-
na umum. Kami perkirakan hai ini disebabkan oleh aktivasi makrofag.
Netrofil mempunya kemampuan untuk memakan dan membunuh Salmonella dan
berpengaruh terhadap perjalanan penyakit in vivo {Bab 2). Pada Bab 7 dijelaskan
tentang efek rekombinan "murine granulocyte colony-stimulating factor (rmG-CSF)"
yang dapat meningkatkan daya hidup mencit yang mengalami infeksi letal kuman
Salmonella. Temyata, tidak terdapat perbedaan pertumbuan kuman S. typhimurium
mencit kontrol dibandingkan mencit yang diberi praperlakuan rmG-CSF. Pada
eksperimen kami, 24 jam setelah pemberian rmG-CSF praperlakuan, produksi TNFa

152 Chapter 11
oleh makrofag peritoneum (dengan stimulasi LPS) lebih tinggi secara signifikan.
Pada mencit yang diberi rmG-CSF dengan pemeriksaan histologik pada hepar di-
dapatkan fokus-fokus inflamasi yang lebih terlokalisir. Hasil ini menunjukkan bahwa
pemberian rmG-CSF praperlakuan memberikan hasil yang menguntungkan pada mencit
yang terinfeksi kuman fakultatif intaseluler S. typhimurium.. Yang menjadi pertanyaan
adalah, rG-CSF mempunyai efek yang menguntungkan pada infeksi intraseluler melalui
rekrutmen netrofll pada fase dim ataukah sebaliknya justru merugikan karena pada fase
lanjut rG-CSF mempunyai kemampuan antiinflamasi sehingga dapat mengambat
aktivasi makrofag.
Meskipun Salmonella merupakan kuman berbentuk batang Gram-negatif, akan
tetapi masih terdapat kontroversi mengenai peran endotoksin pada patogénesis infeksi.
Kami berusaha memecahkan masalah ini (Bab 8) dengan melakukan percobaan pada
mencit "low density lipoprotein receptor knock-out (LDLR-/-)". Pada mencit jenis ini
lipoprotein yang terdapat pada sirkulasi dapat mengikat dan menetralisasi LPS kuman
sehingga dapat mengurangi induksi sitokin proinflamasi. Konsentrasi tinggi lipoprotein
dapat memberikan pengaruh yang baik pada mencit yang terinfeksi S. typhimurium.
Efek menguntungkan dari lipid yang mempunyai kemampuan untuk menetralisasi
endotoksin seperti halnya yang terjadi pada infeksi kuman ekstraseluler Klebsiella ter-
nyata bukan merupakan mekanisme yang mendasari terjadinya peningkatan daya hidup
pada infeksi S. typhimurium.. Pada penelitian ini terlihat bahwa pengaruh lipoprotein
adalah terhadap distribusi dan pertumbuhan fase lanjut dari kuman. Efek pada distribusi
kuman dihubungkan dengan kemampuan lipoprotein yang secara langsung dapat meng-
ganggu "uptake" dari Salmonella misalkan dengan сага menyelubungi struktur permuka
kuman dengan lipid. Hypotesa lain adalah bahwa reseptor LDL mempengarahi
"uptake" bakteri.
Terhadap pertumbuhan kuman, tidak kami dapatkan perbedaan bermakna pada
makrofag peritoneum dal am kemampuan fagositosis dan membunuh kuman antara
mencit dengan atau tanpa reseptor LDL. Hal ini menunjukkan bahwa perbedaan dalam
kecepatan pertumbuhan kuman disebabkan karena peran dari fagosit lain sel perito
neum.
Kesimpulan
Pada penderita demam tifoid kami dapatkan adanya regulasi yang menekan pro-
duksi sitokin proinflamasi yang juga merupakan fenomena umum pada suatu infeksi

Ringkasan 153
dan keadaan lainnya. Observasi ini mendukung pendapat yang menyatakan bahwa
imunoterapi merupakan hal yang menguntungkan pada infeksi Salmonella dan inter-
vensi yang dilakukan harus ditujukan untuk peningkatan sistim kekebalan tubuh.
Kami mempelajari imunomodulasi pada suatu eksperimen yang dilakukan pada
mencit yang terinfeksi S. typhimurium.. Pada penelitian ini didapatkan adanya pengaruh
yang menguntungkan dari pemberian rekombinan IL-1 praperlakuan terbukti dengan
berkurangnya pertumbuhan kuman pada organ-organ tubuh mencit. Dapat dikemukakan
suatu hipotesa bahwa hai ini terjadi sebagai akibat terdapatnya efek distribusi yang
diperankan oleh makrofag yang teraktivasi.
Pemberian nnG-CSF praperlakuanjuga memberikan pengaruh yang menguntung
kan pada daya hidup mencit terhadap S. typhimurium walaupun pertumbuhan kuman
pada organ tidak terpengaruh. Efek rmG-CSF pada rekrutmen dan aktivasi netrofil yang
terjadi pada saat awal merupakan mekanisme yang paling utama untuk daya hidup.
Dapat juga dilihat bahwa rmG-CSF dapat memacu peningkatan produksi TNFa oleh
makrofag peritoneum yang dipacu oleh LPS. Keadaan ini ikut mempengaruhi perjala-
nan penyakit.
Efek menguntungkan dari lipoprotein pada infeksi S. typhimurium ditemukan baik
pada distribusi kuman pada saat awal maupun pertumbuhan selanjutnya dari mikroorga-
nisme. Hai ini menunjukkan bahwa lipoprotein beserta reseptomya berperan penting
dalam pertahanan tubuh terhadap infeksi Salmonella baik pada distribusi awal maupun
dalam kemampuan "intracellular killing" fagosit.
Observasi infeksi Salmonella typhimurium yang dilakukan secara eksperimental
dapat memberikan aran pada perkembangan imunoterapi pada infeksi S. typhi pada ma-
nusia.

154 Chapter 11

Chapter 12
Samenvatting

156 Chapter 12

Samenvatting 157
Buiktyfus is een ernstige gegeneraliseerde infektie veroorzaakt door Salmonella
typhi. De ziekte wordt van mens op mens orofecaal overgedragen. Meestal zijn het
kinderen en jonge volwassenen, die buiktyfus krijgen. In Indonesie is de incidentie
hoog, met meer dan 1000 gevallen per 100 000 inwoners. De mortaliteit van buiktyfus
is in Indonesie door antibiotica gedaald van 18% tot 4%.
In 1989 werd gezamelijk onderzoek opgezet tussen de Katholieke Universiteit Nij
megen en de Diponegoro Universiteit in Semarang, Java, Indonesië. Het doel van deze
samenwerking was om klinische en immunologische factoren te onderzoeken bij buik-
tyfuspatienten. Daarbij was het ook de bedoeling de Pathogenese van Salmonella infek-
ties te bestuderen in een experimenteel diermodel.
In Hoofdstuk 2 wordt de Pathogenese van Salmonella infekties beschreven alsmede
de rol van cytokinen die bij deze infekties betrokken zijn. Omdat S. typhi niet patho-
geen is voor muizen, wordt de experimentele S. typhimurium infektie gebruikt om de
Pathogenese te bestuderen, aangezien deze infektie eenzelfde ziektebeeld als buiktyfus
geeft in muizen. Studies die gedaan zijn met dit muizenmodel worden in Hoofdstuk 2
gerefereerd. Bij extracellulaire Gram-negatieve infekties, waar hoge circulerende
concentraties van proinflammatoire cytokinen zoals interleukiene-l (IL-1) en tumor
necrosis factor (TNFot) worden beschouwd als de oorzaak van orgaanfalen en dood, is
remming van deze substanties gunstig. Daarentegen is bij intracellulaire infekties
remming van proinflammatoire cytokinen meestal nadelig. TNFa lijkt bij deze infekties
lokaal nodig (in kleine hoeveelheden) en toediening van recombinant TNFa voorkomt
uitgroei van microorganismen bij Salmonella infekties.
Wanneer men de symptomen, met name de koorts overziet bij buiktyfus, is het te
verwachten dat endogene pyrogenen in het bloed circuleren. Aangenomen wordt dat het
vrijkomen van Prostaglandine (PGE2) in de hersenen de uiteindelijke stap is bij het
onstaan van koorts en dat die stap gebeurt onder invloed van circulerende proinflam
matoire cytokinen. Deze zogenaamde endogene pyrogenen worden gemaakt op de
plaats van de infektie. In Hoofdstuk 3 rapporteren we dat er in de acute fase van buik
tyfus lage concentraties circulerende proinflammatoire cytokinen gevonden worden en
daarbij hoge concentraties antiinflammatoire niet-pyrogene stoffen zoals interleukin-1
receptor antagonist (IL-IRA) and oplosbare receptoren van tumor necrosis factor
(sTNFRI and sTNFRII). Omdat bij buiktyfuspatienten met koorts pyrogene cytokinen
niet gevonden werden, leek het dat andere mechanismen een rol zouden kunnen spelen.
Deze andere mechanismen zouden andere mediatoren kunnen zijn zoals het secretoire

158 Chapter 12
phospholipase A2 (sPLA2). Phospholipases zijn lipolytische enzymen die de afbraak
van phospholipiden katalyseren. Het vrijkomen van sPLA2 wordt geïnduceerd door IL-
1 en TNFa en het enzym кап de produktie van arachidonzuur en PGE2 aanzetten. Aan
getoond werd dat intraventriculaire injektie van sPLA2-remmers in hersenen van konij
nen, koorts kon tegengaan. sPLA2 zou dus mogelijk een rol spelen als circulerend pyro-
geen. In Hoofdstuk 4 vonden we inderdaad verhoogde concentraties sPLA2 in de acute
fase van buiktyfus. Deze waarden daalden naarmate de patiënten verbeterden. We be
schouwen het als mogelijk dat dit bioactieve sPLA2 verantwoordelijk zou zijn voor een
gedeelte van de symptomen van buiktyfus.
Hoewel circulerende pyrogene cytokinen niet gevonden werden in de acute fase
van buiktyfus, is het belangrijk te vermelden in het "koortsconcept" dat het transport
RNA voor TNFa en IL-Iß aanwezig bleef in circulerende witte bloedcellen. Dit werd
beschreven in Hoofdstuk 5. Een andere hypothese (dan die aangaande PLA2) voor
koorts is dat zulke circulerende cellen zelf de pyrogene cytokines maken, wanneer ze in
het centraal zenuwstelsel komen.
In Hoofdstuk 3 rapporteren we in de acute fase van buiktyfus ook down-regulatie
van de produktie van proinflammatoire cytokines door circulerende cellen na stimulatie
door lipopolysaccharide (LPS). Om dit te bestuderen gebruikten we een vol-bloed cyto
kine test. De gevonden verminderde cytokine productie was niet het gevolg van leuko-
penie of uitputting van deze cellen gedurende de acute fase van de infektie. Er blijkt een
posttrancriptionele uitschakeling van de produktie van proinflammatoire cytokines te
bestaan in de beginfase van de infektie, terwijl tegelijkertijd de anti-inflammatoire
respons in de mononucléaire cellen behouden blijft, want de produktie van IL-IRA was
niet verminderd op dat moment.
Om dit mechanisme van verminderde cytokine produktie of down-regulatie verder
te bestuderen in buiktyfus werd transportRNA voor IL-Iß en TNFa bepaald in vol
bloed. De bevinding dat transport RNA voor IL-Iß en TNFa niet verlaagd was in de
acute fase van buiktyfus, beschreven in Hoofdstuk 5, verwerpt ook uitgeputte produktie
als verklaring en onderstreept posttranscriptionele remming van cytokine produktie.
Deze in-vivo down-regulatie van cytokine produktie is sind enige tijd een bekend
fenomeen ook in andere omstandigheden. Het is zeer waarschijnlijk bedoeld als be
scherming tegen overweldigende cytokinenstromen, maar de verminderde produktie
kan ook afweerfunkties verslechteren. Het voordeel van down-regulatie in patiënten
met buiktyfus is niet goed te rijmen met de experimentele dierproeven met Salmonella
infekties waar proinflammatoire cytokinen nodig zijn om te overleven {Hoofdstuk 2).

Samenvatting 159
Het zou kunnen dat de nog lagere proinflammatoire cytokines bij gecompliceerde ge
vallen een afspiegeling zijn van een minder effektieve afweer, en dat immunothérapie
de gastheerweerstand hier zou kunnen verbeteren.
Om de afweer tegen Salmonella, infektie beter te begrijpen, bestudeerden we de
cytokine produktie in muizen (Hoofdstuk 6) en vergeleken deze met de gegevens uit de
voorgaande hoofdstukken van patiënten met buiktyfus (Hoofdstukken 3, 4 en 5). Circu
lerende cytokinen concentraties maar ook LPS-gestimuleerde ex-vivo produktie in vol
bloed waren niet detecteerbaar zonder infektie. Het onvermogen om cytokinen te
maken is het omgekeerde van de humane situatie, aangezien gestimuleerde humane
perifere bloedcellen grote hoeveelheden proinflammatoire cytokinen produceren zonder
infektie. Tijdens infektie vonden we meer proinflammatoire cytokinen produktie in dit
diermodel, wat ook in tegenstelling is tot wat we vonden bij patiënten met buiktyfus,
beschreven in Hoofdstuk 3.
Overeenkomstig andere studies van ons laboratorium bekeken we of toediening
van recombinant IL-1 (rhIL-la) een beschermend effekt had in een muizenmodel van
Salmonella typhimurium infekties (Hoofdstuk 6). RhJL-la toediening leidde tot een sig
nificant verminderde uitgroei van microorganismen in de organen van de muizen. Dit
effekt van rhIL-la voorbehandeling werd al 24 uur na het begin van de infektie zicht
baar, en het microbiologische effekt van deze eenmalige dosis van гшЪ-Іос bleef gedu
rende de gehele studie bestaan. Peritoneaalcellen van muizen, voorbehandeld met rhlL-
1 α produceerden significant meer cel-geassocieerd IL-la na 24 uur. Verschillende
mechanismes zouden ten grondslag kunnen liggen aan dit gunstige effekt van voorbe
handeling op een Salmonella infektie. Het feit dat IL-1 voorbehandeling beschermt
tegen zeer uiteenlopende infekties doet vermoeden dat het om een algemeen mechanis
me gaat. Wij postuleren dat dit macrofaagactivatie zou kunnen zijn.
Neutrofiele granulocyten zijn in staat Salmonella te fagocyteren en te doden. Van
deze cellen is gebleken dat ze belangrijk zijn voor het verloop van de infektie in vivo
(Hoofdstuk 2). In Hoofdstuk 7 beschrijven we dat recombinant murine granulocyte
colony-stimulating factor (rmG-CSF) de overleving verbeterd van muizen met een
potentieel lethale S. typhimurium infektie. De uitgroei van 5. typhimurium in de orga
nen van rmG-CSF voorbehandelde muizen was echter niet verschillend van die van
controle muizen. De gestimuleerde TNFct produktie van peritoneaal macrofagen van
met rmG-CSF voorbehandelde muizen was hoger dan die van controle muizen na 24
uur. Histologische beoordeling van de lever na infektie liet meer gelokaliseerde (tegen-

160 Chapter 12
over diffuse) infektiehaarden zien in de met rmG-CSF behandelde dieren. Voorbehan
deling met rmG-CSF is dus van voordeel bij muizen met een infektie met de facultatief
intracellulaire S. typhimurium.
Omdat we bij doorbehandelde dieren een trend vonden naar meer uitgroei van
Salmonella vroegen we ons af, of het gunstige effekt van rG-CSF op intracellulaire
infekties tot stand komt via (vroege) verhoogde neutrofielen activiteit en aantallen en of
later de antiinflammatoire eigenschappen die rG-CSF heeft, mogelijk door remming
van TNFa gemedieerde macrofaag activatie, nadelig zouden zijn.
Hoewel Salmonella een Gram-negatieve staaf is, is de rol van endotoxine in de
Pathogenese van door dit microorganisme veroorzaakte infekties controversieel. We
benaderden dit probleem {Hoofdstuk 8) door studie van low density lipoprotéine recep
tor knock-out (LDLR-/-) muizen, waarvan de circulerende lipoproteinen in staat zijn
om bacterieel LPS te binden en te neutraliseren, waarbij de aanmaak van proinflam
matoire cytokinen geremd wordt. De hoge concentraties lipoproteinen bleken het ver
loop van een S. typhimurium infektie gunstig te beïnvloeden. We laten zien dat het
gunstige effekt op de overleving na een S. typhimurium infektie niet tot stand komt via
neutralisatie van endotoxine, wat bij infektie met de extracellulaire Klebsiella het me
chanisme is.
We vonden effekten op distributie en uitgroei. Het eerste zou verklaard kunnen
worden door lipoproteinen die direkt de Salmonella opname verhinderen doordat ze
zich bijvoorbeeld hechten op oppervlaktestrakturen van de bacterie. Ook zou het moge
lijk zijn dat de receptor voor LDL de opname van Salmonella direkt beïnvloed.
Voor het latere effekt op uitgroei vonden we niet meteen een verklaring. Zowel de
fagocytose van Salmonella als ook de intracellulaire bacteriocidie door peritoneaal-
macrofagen was niet verschillend tussen muizen met of zonder LDL-receptor. Deze
resultaten suggereren dat het gevonden verschil in groeisnelheid verklaard moet worden
door een effekt op andere fagocyterende cellen dan de peritoneaalcellen, die wij
onderzochten.
Conclusies Wij vonden down-regulatie van de produktie van proinflammatoire cytokinen bij
patiënten met buiktyfus. Deze bevinding onderstreept het idee dat immunothérapie
gunstig kan zijn voor infekties met Salmonella typhi en dat zulke interventies gericht
moeten zijn op het suppleren of het doen toenemen van die immuunmodulatoren, die de
afweer tegen deze infektie verhogen.

Samenvatting 161
We bestudeerden immuunmodulatie door middel van de experimentele Salmonella
typhimurium infektie bij muizen. Voorbehandeling met recombinant IL-1 bleek gunstig
te werken op deze infektie met minder uitgroei van microorganismen in de organen. Als
hypothese werd verdedigd, dat dit door een distributie effekt komt, gemedieerd door
geactiveerde macrofagen.
Voorbehandeling met rmG-CSF vergrootte ook de overleving van muizen geduren
de een S. typhimurium infektie. De uitgroei van microorganismen daarentegen veran
derde niet. Het vroege effekt op neutrofiele granulocyten aantallen en activatie werd het
meest belangrijke mechanisme geacht voor de overleving. Voorbehandeling met rmG-
CSF verhoogde de LPS-gestimuleerde TNFa produktie door peritoneaal macrofagen op
het tijdstip van infektie. Dit kan bijgedragen hebben tot de betere overleving.
Lipoproteinen verminderden de distributie en de latere uitgroei tijdens een S. typhimu
rium infektie. De resultaten geven aan dat lipoproteinen en hun receptoren cruciale
factoren zijn in de afweer tegen Salmonella infekties zowel in de vroege distributiefase
als bij de intracellulaire fagocytaire bacteriocidie. Deze bevindingen in experimentele S.
typhimurium infekties bij muizen geven richting aan de ontwikkeling van immunothé
rapie voor Salmonella typhi infekties in patiënten.

162
Dankwoord
Pertama-tama saya berterima kasih untuk pasien-pasien, yang beri daran untuk
penelitian ini di Semarang. Kemudian Edi Dharmana dan M. Hussein Gasem teman-
teman saya dan semua perawat, Jujuk dan dari bagian mikrobiologi Bambang Isban-
drio, Hendro Wahyono dan Pak Untung: terimah kasih. Di rumah (dan restorán) Ninik
Jamari dan begitu di pak Supardio keramahan besar.
Mijn promotor Prof. Dr. J. W. M van der Meer. Beste Jos, toen je jaren geleden mij
aannam op het buiktyfusprojekt had je niet kunnen vermoeden dat het zo lang en 2 kin
deren verder, zou duren tot de afsluiting met dit proefschrift. Met je kenmerkende opti
misme bleef je ons drieën (Hussein, Edi en mij) stimuleren. We zullen zorgen dat de
andere 2 proefschriften snel volgen.
Prof. Dr. R. Djokomoeljanto. Matumuun Prof. untuk undangan dari Semarang
kerjasama di penelitian tentang demam tifus. Ken a tidak selalu gampang, tetapi teman-
teman di Semarang dan makanan Java bisa mengumpan saya berkali-kali.
Mijn co-promotor Dr. B.J. Kullberg. Beste Bart-Jan, jij hebt zeer veel werk verricht
aan dit proefschrift vanaf 1993. Van je zorgvuldigheid heb ik veel geleerd. Heel veel
dank voor alles.
Mijn co-promotor Dr. W. M.V. Dolmans. Beste Wil, jij bent de grote facilitator
van deze samenwerking met Indonesie geweest en nog. Dank je zeer.
Mihai Netea, dank voor je hulp en stimulerende gesprekken over de muizenstudies.
Van het Laboratorium Algemene Interne Geneeskunde (Hoofd Dr. P. N. M. De-
macker) wil ik met name danken Ineke Verschueren, Johanna van der Ven-Jongekrijg
en Liesbeth Seuren en natuurlijk Pierre. Ook Gerard Pesman en Marielle Spruijtenburg
van het Laboratorium Endocrinologie en Voortplanting dank ik. Jullie weten beter dan
ik dat zonder jullie werk dit proefschrift er niet zou zijn.
Van het Centraal Dierenlaboratorium (Hoofd Dr. J. Koopman) dank ik Jos
Hendriks, Yvette Brom, Theo van der Ing, Gerrie Grutters, Margo van de Brink en
Monique Bakker voor hun hulp bij de dierproeven.
Joep van Oosterhout en Arthur de Meyer dank ik voor de hulp bij de voorberei
dingen van het dierproevenonderzoek.
Tenslotte zou alles niet zo zijn gelopen als de kinderen niet vertroeteld werden
door mijn schoonouders, mijn moeder en door Angela Scholten-Peeters en Miep Don
kers- Awater (in Kekerdom) en door Lucia Tenten, Ine van Hest en Hermine Dekkers
(in Heeswijk). Voor de prettige sfeer in een schoon huis ben ik jullie zeer dankbaar.

163
Curriculum Vitae De schrijfster van dit proefschrift werd geboren op 13 augustus 1956 te Utrecht. In
1974 behaalde zij het einddiploma gymnasium-ß aan het Christelijk Gymnasium te
Utrecht. Hetzelfde jaar werd gestart met de studie geneeskunde aan de Rijksuniversiteit
Utrecht. Het doctoraal werd behaald in 1980 en het artsexamen op 26 juni 1981. In
augustus 1981 werd begonnen met de specialisatie tot internist in het Diaconessenhuis
in Arnhem (opleider Dr. C. van Gastel). Van 1983 tot 1984 deed zij onderzoek naar de
relatie tussen neuropeptiden en circulatie aan het Rudolf Magnus Instituut voor farma
cologie te Utrecht (Prof. Dr. W. de Jong, Prof. Dr. D. de Wied). Vanaf 1984 werd de
opleiding tot internist vervolgd in het Onze Lieve Vrouwe Gasthuis te Amsterdam (op
leider Dr. K.J .Roosendaal, Dr. J. Silberbusch). Na de afsluiting met tropische genees
kunde in het Academisch Medisch Centrum (AMC), Amsterdam (Prof. Dr. A. de
Geus), werd zij op 1 januari 1987 geregistreerd als internist.
Van 1987 tot 1989 werkte zij als internist in Mumias, Kakamega Diocese, Western
Province Kenya en deed malaria-onderzoek in samenwerking met het AMC, het Kenya
Medical Research Institute (Dr. A.D. Brandling-Bennett en J. Were) en Wellcome Trust
(Dr. W. Watkins).
Na terugkeer in Nederland werkte zij 1 jaar op de afdeling Intensieve Zorg,
Radboud Ziekenhuis, Nijmegen (Hoofd: Dr. J. Gimbrere). Vanaf 1990 is zij gedeeltelijk
verbonden als universitair docent aan de facultaire werkgroep Gezondheidszorg
Ontwikkelingslanden (Thans: Nijmegen Institute of International Health; Hoofd: Dr.
W.M.V. Dolmans) en aan de afdeling Algemeen Interne Geneeskunde (Hoofd: Prof.
Dr. J.W.M, van der Meer) van de Katholieke Universiteit Nijmegen. In 1998 werd zij
geregistreerd als intemist-infectioloog (Opleider Dr. B.J. Kullberg).
Zij is gehuwd met Albert van Linge. Zij hebben twee dochters, Christine en Vita.


Stellingen
1. Down-regulatie van proinflammatoire cytokinenproduktie wordt gezien in de acute fase van
buiktyfus, maar dit fenomeen is een gebruikelijke reaktie bij infekties of acute stress zoals
operaties of excessieve inspanning.
2. Recombinant interleukine-1 verbetert de uitkomst van Salmonella infektie bij muizen
waarschijnlijk door in een vroege fase de distributie van bacteriën te beïnvloeden.
3. Het gunstige effekt van lipoproteinen op een infektie met Salmonella typhimurium bij
muizen berust op beïnvloeding van distributie en uitgroei van micro-organismen.
4. Recombinant granulocyte colony-stimulating factor (rG-CSF) heeft een gunstig effekt op de
overleving van muizen met een Salmonella infektie en bevordert de opname van chinolonen in
neutrofiele granulocytes Daarom zou de combinatiebehandeling van G-CSF en deze antibiotica
patiënten met buiktyfus van nut kunnen zijn.
Dit proefschrift en Andresen J, ed. Abstracts G-CSF Research Meeting. Santa Monica, CA: Amgen,
1996;36-38.
5. Het grote struikelblok in de ontwikkeling van IL-12 als immunothérapie is dat het mogelijk
werkt op ziekten die slechts in arme zuidelijke landen veelvuldig voorkomen.
Hail SS. IL-12 at the crossroads. Science 1995; 268:1432-1434.
6. Elk ziekenhuis zou willen ruilen met de accomodatie van ziektekostenverzekeraars.
7. Het verlies van stemmen van D66 illustreert het verschil in respect dat men heeft voor oudere
vrouwen tegenover dat voor oudere mannen.
8. Dat Indonesië het land met de hoogste incidentie van buiktyfus is, is pas afgelopen jaar voor
ieder aanschouwelijk gemaakt. (TU<j>oç=beneveling / тл)фоі=гоок maken)
9. Een сорго-motor is een milieuvriendelijke aandrijver van de (meestal juist papierverslin-
dende) promovendus.
10. De meeste vrouwen willen meer sexegenoten op hogere posities, mits zij het niet zelf
hoeven te zijn.
Nijmegen 26 mei 1998 Monique Keuter



с
G,




















