
Pectinolytic enzymes secreted by yeasts from tropical fruits

Seediscussions,stats,andauthorprofilesforthispublicationat:https://www.researchgate.net/publication/229927808
Lipidsofoleaginousyeasts.PartII:Technologyandpotentialapplications
ArticleinEuropeanJournalofLipidScienceandTechnology·August2011
DOI:10.1002/ejlt.201100015
CITATIONS
142
READS
277
2authors:
Someoftheauthorsofthispublicationarealsoworkingontheserelatedprojects:
MARINALGAE4AQUA(ERA-NETCOFASPprojectcoordinatedbyLuisaValente(CIIMAR/PT):Improving
bio-utilisationofmarinealgaeassustainablefeedingredientstoincreaseefficiencyandqualityof
aquacultureproduction.Viewproject
SeraphimPapanikolaou
AgriculturalUniversityofAthens
128PUBLICATIONS4,894CITATIONS
SEEPROFILE
GeorgeAggelis
UniversityofPatras
121PUBLICATIONS4,989CITATIONS
SEEPROFILE
AllcontentfollowingthispagewasuploadedbyGeorgeAggelison19May2014.
Theuserhasrequestedenhancementofthedownloadedfile.Allin-textreferencesunderlinedinblueareaddedtotheoriginaldocument
andarelinkedtopublicationsonResearchGate,lettingyouaccessandreadthemimmediately.

Review Article
Lipids of oleaginous yeasts. Part II: Technology andpotential applications
Seraphim Papanikolaou1 and George Aggelis2
1 Laboratory of Food Microbiology and Biotechnology, Department of Food Science and Technology,
Agricultural University of Athens, Iera Odos, Athens, Greece2 Unit of Microbiology, Division of Genetics, Department of Biology, Cell and Development Biology,
University of Patras, Patras, Greece
The process of lipid accumulation in the oleaginous yeasts cultivated in various fermentation configurations
when either sugars and related compounds or hydrophobic substances are used as substrates is presented
and kinetic models describing both de novo and ex novo lipid accumulation are analyzed. Technological
aspects related with single cell oil (SCO) produced by oleaginous yeasts are depicted. The influence of
culture parameters upon lipid production process is presented. Lipid production has been studied in batch,
fed-batch, and continuous cultivation systems using yeasts belonging to the species Lipomyces starkeyi,
Rhodosporidium toruloides, Apiotrichum curvatum, Candida curvata, Cryptococcus curvatus, Trichosporon
fermentans, and Yarrowia lipolytica. The potentiality of yeasts to produce SCO as starting material of
2nd generation biodiesel is indicated and discussed. Of significant importance is also the utilization of yeast
lipids as substitutes of high added value exotic fats (e.g., cocoa butter). Lipid produced by the various yeasts
presents, in general, similar composition with that of common vegetable oils being composed of
unsaturated fatty acids, whereas cocoa butter is principally composed of saturated fatty acids,
consequently the various strategies that are followed in order to increase the cellular saturated fatty
acid content of the yeast lipid are presented and comprehensively discussed.
Keywords: 2nd generation biodiesel / Cocoa butter substitute / Lipid biotechnology / Modeling / Oleaginous yeasts /
Single cell oil
Received: January 10, 2011 / Revised: March 19, 2011 / Accepted: April 12, 2011
DOI: 10.1002/ejlt.201100015
1 Introduction
Two major sectors of lipid biotechnology, the non-conven-
tional biocatalysis and the production of lipid deriving from
microbial sources (the so-called ‘‘single cell oil, SCO’’)
present a continuous expansion in the last years. In the former
sector, hydrolytic enzymes (e.g., lipases or other esterases,
glycosidases, etc.) in free and/or immobilized form are used in
media presenting a feeble water concentration (organic or
non-conventional media), in order to synthesize novel bio-
molecules of industrial, technological, and medical interest
(for reviews see: Buchholz and Bornscheuer [1]; Adlercreutz
[2]; Metzger and Bornscheuer [3]; Bottcher et al. [4]). A
significant part of this work refers to the synthesis of specific
types of structured TAGs that either present composition
similarities with various high-added value exotic fats like
cocoa butter, or have medical significance (i.e., medium-
chain TAGs) through reactions of trans- or inter-esterifica-
tion, in which in several cases low added value fatty materials
(e.g., lard, pomace olive oil, etc.) are used as substrates [3, 5–11].
Likewise, by using such types of reactions it is possible to
concentrate polyunsaturated fatty acids (PUFAs) of medical
importance, found in various fatty materials [12]. Moreover,
in recent developments, hydrolytic enzymes (principally
lipases) have been used in non-conventional media in order
to synthesize with an eco-friendly strategy biofuels (inmost cases
Correspondence: Dr. Seraphim Papanikolaou, Laboratory of Food
Microbiology and Biotechnology Department of Food Science and
Technology, Agricultural University of Athens, Athens, Greece
E-mail: [email protected]
Fax: þ30-210-5294700
Abbreviations: CBS, cocoa butter substitute; FAMEs, fatty acid methyl-
esters; g, of A substance formed per g of B substance consumed; Glc,
Glucose (in g/L); K, saturation constant (in g/L); L, total cellular lipid
(in g/L); q, specific rate of product formed (g/g/h); S, substrate (in g/L);
SCO, single cell oil; TSFAs, total saturated fatty acids; X, total biomass (in
g/L); Xf, lipid-free biomass (in g/L); YA/B, conversion yield; m, biomass
specific growth rate (h�1), subscript max indicates the maximum quantity
and O the initial quantity of the components
1052 Eur. J. Lipid Sci. Technol. 2011, 113, 1052–1073
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

biodiesel that principally refers to fatty acid methyl-esters –
FAMEs) (for reviews see: Adamczak et al. [13]; Szczesna-
Antczak et al. [14]).
The second sector of lipid biotechnology refers to the
production of SCOs. This sector is of particular interest
due to the capacity of various microorganisms (principally
yeasts, molds, and algae and to lesser extent bacteria) to
synthesize lipids with specific structure and/or composition
[15–22]. The continuously increasing demand of the 1st
generation biodiesel (FAMEs deriving from trans-esterifica-
tion of principally plant oils) has increased the cost of various
food-stuffs, and this situation has led to the necessity of
discovery of non-conventional sources of oils, that could
be subsequently converted into biodiesel. The oleaginous
microorganisms are considered as potential candidates for
the production of this lipid that would result in the generation
of the ‘‘2nd generation’’ biodiesel deriving from lipid pro-
duced by oleaginous microorganisms growing on wastes or
agro-industrial residues like sewage sludge, hemicelluloses
hydrolysates, waste glycerol, cheese whey, etc. [23–31], or
the 3rd generation biodiesel deriving from lipid produced by
oleaginous micro-algae, with carbon being offered by atmos-
pheric CO2 sequestration [22, 26, 32].
In general, the oleaginous yeasts produce lipid containing
unsaturated fatty acids similar to that found in common plant
oils [18, 20–22, 33]. Although the production cost of the
microbial lipids generally remains higher than that of the
conventional vegetable oils [21], the production of yeast
lipids with composition similarities with high added-value
specialty fatty materials (like the cocoa butter or other exotic
fats) has been considered as a process potentially economi-
cally viable [15, 17, 18, 20, 22, 34], specifically if various low-
or negative cost raw materials (e.g., whey, industrial-crude
fatty acids, waste glycerol, xylose, etc.) are utilized as sub-
strates [35–41]. Yeast lipid presenting composition sim-
ilarities with the cocoa butter has been produced in
industrial scale at the end of 1980s [42]. Furthermore, due
to the last crisis in the production and price of the various
comestible products, the cost of the various plant oils (e.g.,
rapeseed oil, soybean oil, etc.) has considerably increased the
last years [21]; this event has resulted in a non-negligible
increase of conventional biodiesel production cost. However,
the application of biofuels in a large commercial scale is
strongly recommended by various authorities; with the EU
directive 2003/30/EC, a quantity of 5.75% w/w of biofuel
was planned to be introduced in the conventional fuel by
2010 [26, 28]. Thus, discovery of novel sources for a massive
production of lipid presents significant importance, with the
oleaginous microorganisms being considered as potential
candidates for this purpose (for reviews see: Chisti [32];
Luque et al. [26]; Meng et al. [43]). Specifically, the yeasts
have the remarkable capability to be cultivated on a plethora
of renewable- or waste-type materials in various fermentation
configurations (e.g., xylose-based materials, hemi-cellulose
hydrolysates, (raw) glycerol, sewage sludge, various types of
whey permeate, etc.) [15, 24, 27, 33, 44–48] while they
present a significantly maximum higher specific growth rate
compared to molds and algae [17, 21, 34], therefore yeast
lipids can be considered as potential starting materials for the
synthesis of this 2nd generation biodiesel [28].
In the part I [151] of this work, we were interested in the
biochemical events related with the accumulation of storage
lipid during growth of oleaginous yeasts on hydrophilic or
hydrophobic substances utilized as substrates. Fundamental
differences on biochemical level exist between the two proc-
esses, since in the former case de novo lipid accumulation
occurs, while lipid is synthesized during the secondarymetab-
olism performed usually after nitrogen (or to lesser extent
after other essential nutrient like phosphorus or sulfate)
depletion from the medium, whilst in the later case (ex novo
lipid accumulation), lipid is synthesized together with the
production of lipid-free material, irrespective of the nitrogen
presence into the medium. Moreover, after carbon source
depletion from the medium, cellular lipid is subjected into
degradation, regardless of the mechanism that had been
previously used in order for its formation. In the present part
II of this review-article, modeling approaches of lipid
accumulation during growth of oleaginous microorganisms
on several substrates will be presented. Also, the principal
industrial applications of the utilization of oleaginous yeasts
for the production of lipid suitable for biodiesel production,
or the production of substitutes of the cocoa butter will be
discussed.
2 Biochemical engineering and potentialapplications of lipid produced by oleaginousyeasts
2.1 Technology and modeling of lipid accumulationin oleaginous yeasts
2.1.1 De novo lipid accumulation
A large variety of substrates have been used as carbon sources
for the oleaginous microorganisms performing de novo lipid
accumulation, in shake-flask, batch-bioreactor, fed-batch
bioreactor, and continuous culture modes. The substrates
used include analytical-grade or industrially derived sugars
[15, 23, 25, 29, 49–59], molasses [27, 58, 59], cheese whey
[31, 33, 36, 37, 44, 60], glucose-enriched tomato waste
hydrolysate [61–64], glucose-enriched sewage sludge [24],
polysaccharides [65–67], (raw) glycerol [45–47, 57, 68–73],
glycerol-enriched tomato waste hydrolysate [64], prickly pear
juice [74, 75], N-acetyl-glucosamine [30], starch hydroly-
sates [76], inulin or Jerusalem artichoke hydrolysates [77–
79], rice straw hydrolysates [48], sweet sorghum extracts
[80], organic acids [51, 81–84], and ethanol [85–87].
In general onset of lipid accumulation through the de
novo anabolic pathway is given after an essential nutrient
Eur. J. Lipid Sci. Technol. 2011, 113, 1052–1073 The biotechnology of oleaginous yeasts 1053
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

(in most cases nitrogen) depletion into the growth medium,
while in some cases (specifically through utilization of the
yeast Yarrowia lipolytica), instead of or simultaneously with
lipid accumulation, nitrogen depletion into the medium in
batch or fed-batch experiments leads also to the secretion of
citric acid into the culture medium [28, 68, 70]. Only in few
investigations so far, batch cultivations of Y. lipolytica yeasts
in nitrogen-limited sugar-based media have resulted in
noticeable accumulation of lipid inside the yeast cells (i.e.,
lipid >40% w/w, in dry yeast mass), without simultaneous
production of organic acids into the medium [78]. While the
number of investigations concerning lipid accumulation from
sugars or similarly metabolized substrates is indeed high,
kinetic modeling approaches related with this bioconversion
are relatively restricted. Moreover, in most of the case studies
performed, turnover of accumulated lipid was not taken into
consideration in the models proposed. In a first approach
appeared in early investigations, Glatz et al. [60] have pro-
posed a model describing cell growth and lipid synthesis of
the yeast Apiotrichum curvatum growing on whey-permeate in
batch-bioreactor trials. A similar approach has been estab-
lished by Ykema et al. [88] in which the same strain was
cultivated in continuous experiments with glucose utilized as
the sole substrate. It has been considered that lipid accumu-
Table 1. Representation of predicted parameter values estimated from numerical or analytical models describing de novo lipid accumulation
process (or the similar bioprocess of production of extra-cellular citric acid fromyeasts) or ex novo lipid accumulation process from oleaginous
microorganisms
Parameter Reference
Lmax (g/L) 2.1 (Aggelis et al. [96])a), 3.2 (Papanikolaou and Aggelis [107])b), 2.7 (Papanikolaou and Aggelis [107])c)
mXf ðSÞmax(h�1) 0.20 (Ykema et al. [88])d), 0.19 (Glatz et al. [60])e), 0.23 (Papanikolaou and Aggelis [107])b),c),
0.18–0.36 (Papanikolaou and Aggelis [93])f), 0.09 (Fakas et al. [69])g), 0.32 (Papanikolaou et al. [90])h),
0.24 (Anastassiadis et al. [92])i), 0.566 (Economou et al. [80])j)
qLmax (g/g/h) 0.06 (Aggelis and Sourdis [89])a), 0.025 (Glatz et al. [60])e), 0.16 (Papanikolaou and Aggelis [107])b),
0.10 (Papanikolaou and Aggelis [107])c), 0.028 (Fakas et al. [69])g), 2.73 � 10�3 (Papanikolaou et al. [90])h),
0.785 (Economou et al. [80])j)
mXf ðLÞmax(h�1) 1.4 � 10�2 (Aggelis and Sourdis [89])a), 5.1 � 10�3 (Papanikolaou and Aggelis [107])b),
4.0 � 10�3 (Papanikolaou and Aggelis [107])c)
YXf/S (g/g) 0.55 (Ykema et al. [88])d), 0.69 (Glatz et al. [60])e), 0.86 (Papanikolaou and Aggelis [107])b),
0.78 (Papanikolaou and Aggelis [107])c), 2.22 (Papanikolaou et al. [90])h), 0.08 (Fakas et al. [69])g),
0.345 (Economou et al. [80])j)
YL/S (g/g) 0.58–0.60 (Aggelis and Sourdis [89])a), 0.41 (Ykema et al. [88])d), 0.30 (Glatz et al. [60])e),
0.63 (Papanikolaou and Aggelis [107])b), 0.86 (Papanikolaou and Aggelis [107])c), 0.43 (Fakas et al. [69])g),
0.52 (Papanikolaou et al. [90])h), 0.242 (Economou et al. [80])j)
YXf/N (g/g) 38.6 (Glatz et al. [60])e), 60.4 (Papanikolaou et al. [90])h), 31.9–33.6 (Papanikolaou and Aggelis [93])f),
39.3 (Fakas et al. [60])g), 18.21 (Economou et al. [80])j)
YXf/L (g/g) 0.63–0.66 (Aggelis and Sourdis [89])a), 1.37 (Papanikolaou and Aggelis [107])b),
0.70 (Papanikolaou and Aggelis [107])c)
KN (g/L) 0.12 (Glatz et al. [60])e), 0.196 (Papanikolaou and Aggelis [93])f), 0.15 (Fakas et al. [69])g),
0.047 (Anastassiadis et al. [92])i), 0.085 (Economou et al. [80])j), 0.179 (Arzumanov et al. [91])k)
KS (g/L) 59 (Glatz et al. [60])e), 20 (Fakas et al. [69])g), 1.256 (Economou et al. [80])j)
KLS (g/L) 69.27 (Economou et al. [80])j)
Lmax, maximum concentration of SCO produced; mXf ðSÞmax, maximum specific growth rate attributed to the extra-cellular carbon source;
qLmax , maximum specific rate of storage lipids production; mXf ðLÞmax, maximum specific growth rate attributed to the consumption of storage
lipids; YXf/S, conversion yield of lipid-free biomass formed per carbon substrate consumed; YL/S, conversion yield of storage lipid formed per
carbon substrate consumed; YXf/N, conversion yield of lipid-free biomass formed per nitrogen consumed; YXf/L, conversion yield of lipid-free
biomass formed per storage lipid consumed; KN, KLS, and KS are saturation constants.a) Mucor circinelloides on sunflower oil, batch flask culture, SCO production.b) Y. lipolytica on stearin, batch flask culture, SCO production.c) Y. lipolytica on stearin/hydrolyzed rapeseed oil 50/50, batch flask culture, SCO production.d) A. curvatum on glucose, single-stage continuous culture, SCO production.e) A. curvatum on whey permeate, batch bioreactor culture, SCO production.f) Y. lipolytica on biodiesel-derived glycerol, batch flask culture, citric acid production.g) T. elegans on biodiesel-derived glycerol, batch flask culture, SCO production.h) Y. lipolytica on mixture of commercial-industrial glucose and stearin, batch flask culture, simultaneous SCO and citric acid production.i) Candida oleophila on analytical-grade glucose, batch flask culture, citric acid production.j) M. isabellina on sweet sorghum extract, batch flask culture, SCO production.k) Y. lipolytica on ethanol, fed-batch bioreactor culture, citric acid production.
1054 S. Papanikolaou and G. Aggelis Eur. J. Lipid Sci. Technol. 2011, 113, 1052–1073
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

lation process was a combination of two different
mechanisms:
The first mechanism corresponds principally at balanced
cell growth phase, in which lipid accumulation is proportional
to the production of non-lipid cell mass production. In the
second phase, lipid biosynthesis, being independent from the
production of non-lipid cell mass, was performed through the
formation of a rate-controlling intermediate of lipids I,
between sugar and storage lipid [60, 88], describing, thus,
the nitrogen-limited phase (unbalanced growth phase). Not
any accumulation of storage materials other than lipophilic
compounds was assumed in the above approach during the
nitrogen-limited period.
The process of lipid accumulation is performed
when extra-cellular nitrogen is the limiting factor of cell
growth. Therefore, the specific growth rate of lipid-free
material (mXf) was illustrated by the following formula:
mXf ¼ mXfmaxN=ðKN þ NÞð Þ, where KN was the saturation
constant for nitrogen (in g/L) [60]. In this assumption is
considered that lipid-free material is the ‘‘active’’ biomass
portion [60, 88–90].
It should be stressed that similar Monod-type models
have also been used in order to simulate biomass production
during citric acid fermentation using sugars or alcohols [91–
93], or the production of fatty and lipid-free material during
growth of the fungus Thamnidium elegans CCF-1465 on
glycerol-based nitrogen-limited cultures [69]. On the other
hand, in similar types of nitrogen-limited growth of the fun-
gus Mortierella isabellina ATHUM 2935 on sweet sorghum
extracts in shake-flask experiments, the specific growth rate of
lipid-free material synthesized was considered to be influ-
enced by both the sugar and the nitrogen, being described by
a type of double-substrate limitation with sugar inhibition,
according to Andrews’ equation:
mXf ¼ mXfmax
N
KN þ N
� �S
KS þ S þ S2=Ki1
� �;
where KN and KS are the saturation constants for sugar and
nitrogen, respectively (in g/L), and Ki1 is the inhibition con-
stant [80].
A power function f ¼ mXf =mXfmaxpresented the transition
of the microbial metabolism toward the accumulation of
cellular lipids. Specifically, a type function ð1�fÞm was uti-
lized for this purpose [60], therefore, the more the extra-
cellular nitrogen concentration decreased, the more the value
of the function f became lower. The specific formation rate of
the intermediate qI (in g/g/h), related to extra-cellular nitro-
gen exhaustion was presented by the following formula:
qI ¼ ð1�fÞmqImax [60].
The consumption of sugar (substrate – S) contributed to
generation of Xf and I, while the specific formation rate of the
intermediate qI was expressed by aMichaelis–Menten expres-
sion [qI ¼ qImax S=ðKS þ SÞð Þ], where KS was the saturation
constant for sugar substrate (in g/L) [60]. Under the same
optics, the accumulated fat was generated from the inter-
mediate product I through its specific formation rate (qL),
that, again, was expressed by aMichaelis–Menten expression
[qL ¼ qLmax I=ðKI þ IÞð Þ], where I was the intermediate con-
centration (g/L), and KI was the saturation constant for I (in
g/L) [60].
Although intra-cellular lipid storage through de novo lipid
accumulation process is a non-growth associated process, the
specific production rate of cellular lipids is not expressed as
constant. In a similar type of modeling approach recently
appeared in the literature (nitrogen-limited cultures of
T. elegans on glycerol), qL was expressed as combination of
Blackman- and Michaelis–Menten-type expressions:
qL ¼ qLmax
S
KS þ S
� �N0�N
N0
� �;
where KS was the saturation constant for glycerol (in g/L) and
N0 was the initial quantity of assimilable nitrogen [69].
Economou et al. [80] while performing shake-flask cultures
of the fungus M. isabellina on sweet sorghum extracts con-
taining variable initial quantities of assimilable nitrogen and
sugars, proposed the expression
qL ¼ qLmax
S
KLS þ S þ S2=Ki2
� �k2
k2 þ N
� �;
where KLS is the saturation constant (in g/L), Ki2 the inhi-
bition constant (in g/L), and k2 is a constant ensuring that at
high nitrogen concentration lipid production is low [80].
Nevertheless, in similar types of processes (e.g., production
of citric acid from glucose or glycerol), the specific production
rate of citric acid was considered to be a constant (qCit ¼ a)
[90, 92, 93].
2.1.2 Ex novo lipid accumulation
A remarkable plethora of hydrophobic carbon sources have
been used as substrates for ex novo lipid accumulation. These
substrates are various vegetable oils like olive oil, corn oil,
sunflower oil, etc. [60, 89, 94–99], fatty by-products or
wastes such as crude fish oils, soap-stocks, ‘‘stearin’’ (a
low-cost derivative of tallow composed of saturated free-fatty
acids), hydrolyzed rapeseed oil, etc. [39, 100–109], pure free-
fatty acids [110–112], fatty esters [113], and n-alkanes [114,
115]. In some investigations utilization of mixtures of hydro-
(i) Sugar (substrate) Lipid-free biomass (Xf) þ accumulated lipid (L)
(ii) Sugar (substrate) Intermediate (I) Accumulated lipid (L).
Eur. J. Lipid Sci. Technol. 2011, 113, 1052–1073 The biotechnology of oleaginous yeasts 1055
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

philic substrates (glucose and/or glycerol) with various fatty
materials has been employed [40, 90, 113, 116–120],
whereas with some exceptions in which batch bioreactor
experiments were performed [94, 105, 106, 108, 113,
116], in most trials shake-flask trials have been carried out.
As in the case of de novo lipid accumulation process, a
scarce number of modeling studies of growth and lipid
accumulation in oleaginous microorganisms growing on
hydrophobic materials have been realized so far. Storage lipid
accumulation occurs during primary metabolic growth when
fatty materials are used as substrate and therefore after the
exhaustion of the substrate fat the culture environment is still
favorable for growth [96, 97, 105]. For this reason, in all cases
in which kinetic studies have been performed on fatty sub-
strates lipid accumulated was always re-consumed in favor of
lipid-free material generation regardless of the fatty acid
composition of the fat used as substrate [39, 89, 104–
108]. Therefore, storage lipid degradation should obligatorily
be taken into consideration in order to correctly simulate the
bioprocess [89, 107]. It is considered that the microbial dry
matter (total biomass – X) corresponds to the sum of total
cellular accumulated lipids (L) and lipid-free biomass (Xf).
Likewise, Xf should be capable of consuming from the two
available lipid pools, the extra- as well as the intra-cellular one
[89, 107]. Under this optics, it is considered that the micro-
organism grows with two specific growth rates (diauxic
growth-type model), with mXf(S) being the specific rate of
lipid-free biomass generation from the extra-cellular lipid
substrate (in h�1) and mXf(L) being the specific rate of
lipid-free biomass formation deriving from turnover of the
intra-cellular reserve lipid (in h�1). The lipid-free biomass
formation, thus, is the result of the utilization of two carbon
sources, the extra-cellular lipid substrate as well as the reserve
lipids as follows: dXf =dt ¼ mXf ðSÞXf þ mXf ðLÞXf , where
mXf(S) and mXf(L) are the specific rates of lipid-free biomass
formation [89, 107].
The equation describing the specific formation rate of
lipid-free biomass from the extra-cellular lipid pool, inspired
by the models proposed by Aggelis and Sourdis [89] and
Galiotou-Panayotou et al. [121], is illustrated by the follow-
ing formula: mXf ðSÞ ¼ mXf ðSÞmaxðS=S0Þ, where mXf(S) is the
specific rate of lipid-free biomass formation from the extra-
cellular lipid, and S0 is the initial concentration of the limiting
substrate (in g/L) that is a Blackman-type equation. It may be
assumed that this rate presents its maximum value at the
beginning of the fermentation and it decreases linearly with
the decrease of the carbon substrate. A Verhust-typemodel in
which biomass formation is governed by the culture density
[m ¼ mmax 1�ðX=XmaxÞð Þ], is also applied in Papanikolaou
et al. [106]. Of course, the above mentioned types of models
(Blackman- or Verhust-type) can also perfectly simulate the
microbial growth in fermentations in which carbon is not
the limiting factor [90, 93], while, in general, Michaelian-
type expressions of specific growth rate (m) seem to be more
suitable for description of models in the steady-stages of
single-stage continuous cultures [122]. Moreover, the
specific formation rate of lipid-free material performed
through the degradation of intra-cellular lipid is presented
by the following formula: mXf ðLÞ ¼ mXf ðLÞmaxðL=LmaxÞ [89]. It
should be stressed that in general the optimized values of
mXf ðLÞmaxwere significantly lower compared with those of
mXf ðSÞmaxsuggesting that lipid turnover was a process less
rapid than that the consumption of extra-cellular aliphatic
chains [107]. Likewise, consumption of both lipid pools
(extra- and intra-cellular lipid) resulted in the formation of
lipid-free biomass that was characterized from its yield per
lipid (extra-cellular – YXf/S and intra-cellular – YXf/L) con-
sumed [89, 107]. Storage lipid turnover was accompanied by
high YXf/L values, sometimes higher than YXf/S. This can be
explained by the fact that transport of extra-cellular aliphatic
chains inside the cell, a phenomenon energetically costly, is
missing during reserve lipid degradation period, while reserve
lipid consumption is used almost exclusively for the synthesis
of lipid-free material. In contrast, at the first growth steps,
when substrate fat was presented in significant amounts, the
microorganism grew with high specific rates, but part of the
energy is consumed in the incorporation of fatty acids and
the formation of storage lipids [107].
As far as the evolution of intra-cellular lipids through
the ex novo process is concerned, it is consisted of the
result of two inverted processes: that of cellular lipid
formation from the extra-cellular pool, and the one of cellular
lipid uptake for lipid-free biomass formation: dL=dt ¼qLXf�mXf ðLÞXf 1
�YXf =L
� �, where qL is the specific formation
rate of accumulated fat (in g/g/h). Taking into consideration
that in the case of the ex novo lipid accumulation process, fat
is produced during primary metabolic growth simultaneously
with lipid-free material, the expression of qL is of the type
qL ¼ amXf ðSÞ and specifically qL ¼ qLmaxðS=S0Þ [89, 107]. It
should also be stated that in one case, simulation of intra-
cellular lipid (and extra-cellular citric acid) production was
performed during growth of Y. lipolytica ACA-DC 50109 on
mixtures of hydrophobic substrates (a low-cost industrial
derivative of tallow composed of fully saturated free-fatty
acids called ‘‘stearin’’) with commercial glucose in nitro-
gen-limited flask experiments, and at that study taking into
consideration that nitrogen had been already exhausted from
the medium, production of lipid was considered as non-
growth coupled process with qL ¼ a [90].
In Table 1, a representation of the various predicted
parameter values that have been obtained in the literature
are presented. As it can be observed, the maximum concen-
tration of lipids inside the cells (Lmax, g/L) largely depends on
the microbial strain used. Furthermore, parameter mXf ðSÞmax
presents similar values regardless of the carbon source (fat or
sugar) and the microorganism (yeast or mold) used.
Likewise, KN values present similarities regardless of the
non-growth associatedmetabolite produced (de novo derived
intra-cellular SCO or extra-cellular citric acid), while YXf/S
and YL/S values present differences related with the carbon
1056 S. Papanikolaou and G. Aggelis Eur. J. Lipid Sci. Technol. 2011, 113, 1052–1073
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

substrate used. Finally, qLmax values are influenced by de novo
or ex novo lipid accumulation process with higher values
reported during growth on fatty media, compared with that
obtained on sugar-based ones. As previously stated growth of
oleaginous microorganisms on fatty substrates was accom-
panied by amXf ðSÞmaxvalue higher that the respectivemXf ðLÞmax
indicating that lipid turnover was a less rapid process com-
pared with the fat uptake from the extra-cellular medium.
2.2 Yeast lipid suitable for biodiesel production
Current industrialization and decrease of petroleum stock
have raised the worldwide need for energy generation deriv-
ing from various alternative and renewable resources (e.g.,
biodiesel, bio-hydrogen, and/or bio-ethanol) with biodiesel
being considered as one of the most important renewable
energy sources due to its economic and environmental
benefits [123]. Biodiesel is prepared through trans-esterifi-
cation of vegetable oils or animal fats with short chain alco-
hols (principally methanol and to lesser extent ethanol or
butanol). Specifically, according to the ‘‘US Standard
Specification for biodiesel’’ (ASTM 6751-02), biodiesel is
defined as ‘‘a fuel that is composed of mono-alkyl esters of
long-chain fatty acids deriving from vegetable oils or animal
fats’’ [124]. This definition is also acceptable in the European
Union specification concerning biodiesel (EN 14214).
Stricter regulations define biodiesel as FAMEs, but current
considerations could likely extend this definition also to fatty
acid ethyl-esters, which can be obtained using bio-ethanol as
alcohol donor (see also: Lois [124]; Adamczak et al. [13]).
Synthesis of biodiesel is performed principally by chemi-
cal catalysis, but it can also be performed via enzyme-cata-
lyzed methods, that although are still hampered by the high
costs of the biocatalyst, significant progress has recently been
made leading to the first industrial enzymatic biodiesel pro-
duction [13]. Although the production cost of the enzyme-
catalyzed and produced biodiesel is around one order of
magnitude higher than the conventional chemical production
[125], enzymatic production of FAMEs is considerably
attractive taking into consideration that starting materials
like low or negative cost waste frying oils, oils with high water
content, etc., for which conventional chemical trans-esteri-
fication can hardly be applied, can perfectly be used as sub-
strates for the enzymes [13]. Moreover, despite the
continuous need for biodiesel production, the last years lack
of oil feedstock has created problems related with the pro-
duction of conventional or so-called ‘‘1st generation biodie-
sel,’’ and for this reason alternative fatty media were used
(e.g., utilization of waste lipids). Specifically in Europe, it has
been realized that the use of conventional plant oils would
rapidly increase their current prices (a situation that has
already been realized in the case of rapeseed oil), allocate
vast areas of cultivable land from food to biodiesel production
and jeopardize the biodiversity in these areas [57, 126]. In
addition, if the need for biodiesel is to be fulfilled by rapeseed
oil it is estimated that the current total volume of European
rapeseed production is not sufficient [126].On the other hand,
the production cost of SCO remains always much higher than
that of plant oils [21]. A representative value of yeast SCO
produced on 2008 was around 3.0 US $ per kg (excluding cost
of feedstock used for SCO production) while the ones of
rapeseed oil, soybean oil, and sunflower oil are 1.4–1.5, 1.2–
1.3, and 1.8–1.9 US $ per kg, respectively [21]. However, the
previously described situation with the increment of biodiesel
needs has resulted in the fact that between 2007 and 2008
there has been a twofold increase of the price of conventional
plant commodity oils [21]. Therefore, the necessity of dis-
covery of novel (non-conventional) sources of oils, which
could be subsequently converted into biodiesel is of crucial
importance, with the oleaginous microorganisms being con-
sidered as potential candidates for the production of this
‘‘2nd generation biodiesel’’ [23]. It should also be taken into
considerations that the economics of SCO bio-processes can
be further ameliorated by using wastes as substrates because
most of them have a negative value and their discharge causes
environmental problems (nevertheless, in this case a well-
established network providing the waste material into the
fermentation plant is of extreme importance for the economic
viability of the process – see: Ratledge andCohen [21]). Then
the produced biomass rich in lipid can be directly trans-
esterified to yield the biodiesel [57, 127], thus avoiding the
oil extraction step, which is one of the most costly steps of the
SCO production procedure [16]. Autotrophic algae present
also advantages with the principal ones being the utilization of
sunlight and CO2 sequestration for SCO production amend-
able for biodiesel production (the so-called ‘‘3rd generation’’
biodiesel) factors that are considered to be as very crucial that
make these microorganisms as the most appropriate for an
environmentally friendly production of biodiesel [21, 32].
However, the small growth rates, the decreased quantity of
CO2 found into the atmosphere that results in carbon-limited
culture conditions, the difficulty to carry out high cell density
cultures and the current increased cost of photo-bioreactors,
pose problems that need to be solved in the future [32], and
therefore the cost of autotrophically produced algal SCO is
significantly higher than the previous categories, being in the
range of 5.6–21.0 US $ per kg, without taking into consider-
ation the cost of oil extraction [21].
The oleaginous yeasts, with their unicellular form and
their higher specific growth rate compared with molds and
algae, can be considered as appropriate organisms that could
be used for the production of this 2nd generation biodiesel
[23]. Moreover, yeasts can be cultured in various carbon
sources including waste materials presenting simultaneously
rapid and significant biomass and oil production. Likewise,
the potential of several oleaginous yeasts to present
efficient growth and lipid accumulation under nitrogen-
excess conditions provided that another nutrient (e.g., phos-
phorus) is limited [29], can be exploited in industrial level for
the valorization of agro-industrial residues and surpluses that
Eur. J. Lipid Sci. Technol. 2011, 113, 1052–1073 The biotechnology of oleaginous yeasts 1057
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

are carbon- and nitrogen-rich, like media composed of N-
acetylglucosamine [30] (it is known that this compound is the
base-unit of chitin, whilst various chitin-based residues are
currently generated in enormous quantities from crustacean-
fabricating facilities and these wastes are disposed by either
burning or land filling, both of which are harmful to the
environment).
It is evident that considering the production of yeast lipid
that will be subsequently converted into biodiesel, only the
process of de novo lipid accumulation is concerned, since
cultivation in fatty materials is principally performed in order
to add value into the fatty material converted, and, therefore,
ex novo lipid accumulation is principally performed in order
to produce specialty ‘‘tailor-made’’ lipids, like substitutes of
cocoa butter of other high-added value exotic fats. In the next
chapter the influence of cultivation parameters on yeast lipid
accumulation will be discussed.
2.2.1 Factors influencing the accumulation of lipid byoleaginous microorganisms
In the process of de novo accumulation of storage lipid the
concentration of the limiting nutrient (nitrogen) frequently
determines the quantity of the biomass produced, whilst the
concentration of the carbon source (e.g., glucose) found in
excess in the growth environment largely determines the
amount of accumulated lipid. Therefore, the molar ratio
C/N plays a key-role in determining the oil content and
biomass density of the oleaginous microorganisms [17,
18]. Generally, it is considered that the process of lipid
accumulation is induced at molar ratio C/N > 20. In some
cases cultures in media in which very high initial C/N ratios
were imposed (e.g., higher than 70) resulted in decreased fat
accumulation, suggesting that optimum initial C/N molar
ratios are required for the conversion conducted (case of
R. toruloides yeast – see Moreton [15]). In contrast, in other
cases (e.g., culture of L. starkeyi on glucose), in trials per-
formed with constant initial glucose and decreasing initial
nitrogen concentrations, accumulation of storage lipid inside
the cells constantly increased even though in some cases high
initial C/N ratios (around 150 moles/moles) were used, for
the range of initial nitrogen concentrations tested [24]. In
similar types of experiments performed withR. glutinisNRRL
Y 1091, the production of total microbial lipids constantly
increased with increase of the C/N molar ration imposed, for
the range of the initial nitrogen concentrations tested [128].
Likewise, in similar trials (utilization of M. isabellina fungus
growing at constant initial nitrogen and increasing initial
glucose concentrations into the medium), for the range of
glucose concentrations tested, despite high initial sugar
quantities (e.g., 100 g/L) and initial C/N ratios (e.g., initial
C/N � 340 moles/moles), constantly increasing SCO
quantities were produced, while also other lipid-free storage
materials were accumulated (Fig. 1 – Papanikolaou et al.
[54]). Moreover, in some cases of lipid accumulating micro-
organisms (e.g., strains belonging to C. echinulata,
M. ramanniana, etc.), cultivations elaborated in high initial
C/N ratio media can be accomplished with significant
quantities of sugar remaining unconsumed into the culture
medium without tendency of finally being consumed [57, 64,
67, 129]. Considering that, in general, both intra-cellular
NADþ- and NADPþ-isocitrate dehydrogenases during
lipid-accumulating conditions show a very limited activity
[55, 130, 131], the key-enzyme regulating the intra-cellular
carbon flow is the one NADPþ-malic enzyme [131], the low
activity of which could down-regulate and cease the uptake of
sugar during the period of accumulation of lipid constituting
the limiting step of the process [21].
Recent investigations have indicated that the process of
storage lipid accumulation is critically influenced by the
specific sugar uptake rate (qS) inside the cells or the fungal
mycelia; in cultures of the oleaginous mold C. echinulata on
nitrogen-limited glucose-based tomato waste hydrolysate
media or starch-based media, decreased qS values resulted
in increased total biomass (X, in g/L) and total biomass yield
(YX/S, in g/g) values, with concomitant lower lipid production
[64, 67]. It appears that in high qS values reported, carbon
flow was channeled more rapidly and in higher quantities
inside themycelia, suggesting higher intra-cellular C/Nmolar
ratio compared with cultures presenting low qS values. For
Figure 1. Cultures of the oleaginous fungus M. isabellina on glu-
cose used as the sole carbon source at increasing initial glucose
concentrations (Glc0 concentrations 45, 50, 60, 70, and 100 g/L)
and constant nitrogen concentration (ammonium sulfate and yeast
extract adjusted at 0.5 g/L of each). Representation of lipid-free
material (Xf, g/L), lipid (L, g/L) and lipid in dry weight (g per 10 g of
total biomass) when the maximum concentration of lipid was
obtained (data from Papanikolaou et al. [54]).
1058 S. Papanikolaou and G. Aggelis Eur. J. Lipid Sci. Technol. 2011, 113, 1052–1073
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

this reason, the carbon flow in the later case seemed of having
been directed principally toward the synthesis of lipid-free
material rather than the storage lipid formation [67].
Furthermore, in the case of C. echinulata, the presence of
proteins in the culture medium seemed to critically influence
the uptake rate of sugar, and, hence, the process of lipid
accumulation, since organic nitrogen of tomato waste hydro-
lysate clearly enhanced glucose uptake and storage lipid pro-
duction, while cultures in the above waste in which proteins
had been removed was accompanied by drastically lower
uptake rate of glucose and production of lipid [64].
Apparently, this is the main reason for which although in
various cases trials have been performed with sugars that
present very high similarity in biochemical level (e.g., glucose,
maltose, lactose, soluble starch, etc.) non-negligible differ-
ences in lipid accumulation have been observed [48, 67, 132].
The nitrogen source has been also reported to be of
importance for the process of de novo lipid accumulation
in yeasts and molds; for instance, in the case of C. albidus
strain CBS 4715, addition of inorganic nitrogen sources
(ammonium sulphate or ammonium chloride) favored the
accumulation of lipid the yeast cells in comparison with
organic nitrogen source (e.g., urea, L-arginine, etc.) [132].
In contrast, in other cases addition of organic nitrogen sour-
ces into the medium has substantially increased the quantity
of SCO produced [15, 133]. In R. toruloides CBS 14 addition
of asparagine instead of ammonium chloride increased the
lipid content in dry yeast mass from 18 to 51%, w/w [133].
Further study of this strain in flask or bioreactor trials showed
the same trend with a wide range of organic N sources
used [15].
The pH and the incubation temperature have been
reported as factors that equally have importance in relation
with the process of SCO accumulation. Specifically, the
incubation temperature critically influenced lipid accumu-
lation in the yeasts C. albidus, A. curvatum, R. minuta, and
Y. lipolytica [16, 105, 132, 134]. For the case of Y. lipolytica
(ex novo lipid accumulation with low-cost saturated free-fatty
acids used as substrate), temperatures of 19 and 398C did not
allow high growth, whilst significant growth was observed
within the range of 24–338C (Xmax ¼ 7.5–8.7 g/L,
mmax ¼ 0.26 h�1). However, fat accumulation was favored
only at T ¼ 288C [105]. Similar trend has been observed for
R. minuta [134], while the optimum temperature for biomass
and lipid production by C. albidus was 208C [132]. Finally,
during continuous cultures of A. curvatum on whey at con-
stant dilution rate (D ¼ 0.04 h�1) and variations in the incu-
bation temperature (from 30 to 358C) at the steady-states
achieved microbial growth and SCO production were clearly
favored when T ¼ 308C [33]. Likewise, it should be noted
that besides the quantity of fat accumulated into the cells or
the mycelia, the incubation temperature has also significant
effect upon the composition of fatty acids of total cellular
lipids produced. For instance, Ferrante et al. [135] have
demonstrated that in C. lipolytica growing on glucose, the
activity of the enzyme catalyzing the transformation of oleoyl-
CoA to linoleoyl-CoA (D12-desaturase) at T ¼ 108C was
doubled compared with that at T ¼ 258C, with correspond-
ing significant rise of the concentration of intra-cellularD9;12C18 : 2 fatty acid in the later case. Similar observations
have been done by Granger et al. [128] during growth of
R. glutinis on glucose. In similar batch bioreactor experiments
in which growth of R. glutinis on glucose was performed, rise
of the incubation temperature resulted in the increment of
total saturated fatty acids (TSFAs), and principally the ones
of medium aliphatic chain (C12:0, C14:0) [134]. On the
other hand, continuous cultures of A. curvatum at constant
D and variations in the incubation temperature did not reveal
noticeable differentiations in the composition of intra-cellular
fatty acids produced [33]. Finally, in contrast with the results
reported by Ferrante et al. [135] and Granger et al. [128], in
batch shake-flask experiments performed by the yeast
C. curvatus ATCC 20509 on N-acetyl-glucosamine utilized
as the sole substrate, decrease of the incubation temperature
(e.g., T ¼ 228C) was accompanied by synthesis of a fat that
was remarkably enriched in saturated fatty acids (principally
C16:0 and C18:0), in comparison with growth in higher
temperatures (e.g., T ¼ 26–308C) [30].
As far as the pH of the medium is concerned, for the case
of Y. lipolytica growing on stearin, medium pH was con-
sidered as crucial factor for SCO production, since cultures
were performed in initial pH values of 5.0–7.0, and substan-
tial growth was observed only at pH 6.0–6.5 whereas fat
accumulation was favored only at pH ¼ 6.0 [105]. On the
other hand, continuous cultures of A. curvatum growing on
whey at constant D and variations of the pH of the medium
resulted in significant and almost unaffected from the pH
accumulation of fat for a pH range between 3.5 and 5.5 [33,
42]. Apparently continuous fermentation at pH ¼ 3.5 pro-
vides considerable advantages since this operation can be
feasible with a pasteurized whey feed in industrial scale [33].
Various yeast strains have been tested in relation with the
potential growth and SCO production on the sugars deriving
from hemicelluloses hydrolysis [23, 25, 48]. It is known that
lignocelluloses biomass hydrolysis results in generation of
various by-products that may have affect on down-stream
SCO biotransformation. Therefore, such types of products
like acetic acid, syringaldehyde, vanillin, furfural, etc. were
added into a glucose-based medium and the production and
study of SCO by R. toruloides Y4 was assessed in the presence
of these inhibitors [136]; it was found that some products like
hydroxymethylfurfural (HMF) and acetic acid had a very
slight inhibitory effect upon growth (e.g., addition of
100 mM of acetic acid or 16 mM of HMF did almost not
at all negatively influence biomass production). In contrast,
addition of other compounds like furfural or vanillin in
quantities ranging between 8 and 12 mM, almost completely
ceased microbial growth [136]. The addition of these inhibi-
tors influenced also the fatty acid composition of total
microbial lipids produced [136].
Eur. J. Lipid Sci. Technol. 2011, 113, 1052–1073 The biotechnology of oleaginous yeasts 1059
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

A significant number of investigators have studied the
synthesis of microbial lipid in nitrogen-limited batch, fed-
batch (flask or bioreactor) and single-stage continuous or
continuous with biomass recycling cultures. In most of the
cases, renewable-or waste-type carbon sources have been
used and in almost all cases, SCO produced can potentially
be used for 2nd generation biodiesel synthesis. In some
reports, investigations deal with SCO production in single-
stage or cell-recycling continuous yeast cultures [33, 44, 47,
49, 137, 138]. It has been demonstrated that lipid accumu-
lation was strongly depended on the dilution rate imposed
[17, 52, 137] as well as of the molar ratio C/N of the growth
medium [17, 88]. D of less than 0.06 h�1 were normally
required for optimum conversions [17, 49], since the
microbial cells need to remain within the chemostat for at
least 12–24 h in order to consume the available nitrogen and
then convert the remaining sugar to oil and ‘‘fatten’’ [17, 47].
Moreover, in a restricted number of reports fed-batch exper-
iments have been performed, remarkably high biomass pro-
duction (�100 g/L) containing various lipid quantities has
been observed [23]. Finally, concerning strains of the non-
conventional yeast Y. lipolytica, cultivation of this microor-
ganism in nitrogen-limited glucose- (or glycerol-) based
media resulted in equivocal results; in highly aerated batch
bioreactors, quantities of lipids were accumulated inside the
yeast cells, whereas nitrogen limitation led also to citric acid
biosynthesis [72]. In contrast, in highly aerated chemostat
cultures and low dilution rates imposed (e.g., D < 0.04 h�1),
lipids were produced in high quantities (e.g., >25% w/w)
inside the yeast cells [47, 138], whereas in at least one case, a
strain of the above-mentioned species produced simul-
taneously SCO and citric acid in shake-flask nitrogen-limited
cultures [70]. It should finally be stressed that in a recent
development, the well-studied lipid- and citric acid-producing
Y. lipolytica strain ACA-DC 50109, was subjected to genetic
manipulation and was revealed capable to secrete significant
inulinase quantities (it was the transformant Z31), that was
revealed capable to simultaneously break-down inulin-based
compounds and produce significant SCO quantities in shake-
flask experiments, e.g., L up to 7.4 g/L with lipid in dry weight
of more than 50% w/w, on nitrogen-limited media containing
the extract of Jerusalem artichoke tubers [78]. The most
representative examples of de novo SCO production by ole-
aginous yeasts are depicted in Table 2.
2.3 Production of cocoa butter substitutes byoleaginous yeasts
One major industrial application referred to yeast lipid pro-
duction is that of the synthesis of microbial substitutes of
cocoa butter [15, 17, 18, 20, 22]. Cocoa butter is commonly
used in the food technology and principally in chocolate
fabrication process, whereas it is also used in various cosme-
tology applications. It is principally produced in some of the
African and Central American countries such as Ivory Coast,
Nigeria, Jamaıca, etc. This lipid is mainly composed of TAGs
of the type P–O–S and S–O–S (P: palmitic acid, O: oleic acid,
S: stearic acid). Oleic acid, hence, is always found esterified
in the position sn-2 of glycerol. Cocoa butter contains
55-67% w/w saturated fatty acid while its composition is
dependent of the plant variety and the culture conditions.
An average fatty acid profile of this fat is: C16:0 23–30%w/w;D9C18:1 30–37%w/w; C18:0 32–37%w/w; D9;12C18:2 2–4%
w/w [17, 22, 140].
The production of cocoa butter substitutes (CBSs), econ-
omically viable during the years 1980–1990 (at that time the
price of the cocoa butter was >8.0 US $ per kg), is depend-
able of the price of this fat. During the years 1990–1994, a
significant fall of cocoa butter price (<2.5 US $ per kg) has
constituted an enormous disadvantage for the production of
various substitutes of this fat [17, 20]. However, the years
after 2000, there has been increment again of its price, due to
the prevalence of harmful insects and viruses that have been
reported to create several problems on cocoa butter pro-
duction [20, 30, 39]. Currently, the price of cocoa butter
is around 5.0 US $ per Kg, though the tendency of this price,
is to present (a remarkably significant) increase in the near
future [22, 30]. An ‘‘extreme’’ scenario recently presented in
both the written and the electronic international press
indicates that cocoa butter risks to disappear the next years
due to general failure of the cultivation techniques of the
cocoa plant, and, thus, the utilization and application of the
various CBSs will be generalized for the food industry.
Numerous approaches have been conducted, in order to
produce lipids having composition similarities with those of
the cocoa butter. The first strategy performed referred to the
preparation of mixtures of different fats of exotic plants (e.g.,
illipe butter, mango fat, kokum butter, sal fat) with fractions
of palm oil [140–142] in order to create fatty materials with
composition and technological properties relatively close to
that of cocoa butter. It is noted however, that already the price
of some of these exotic fats is remarkably high [22]. In
parallel, different biotechnological approaches, either enzy-
matic- or fermentative-ones have been already carried out
for the production of CBSs (for reviews see: Ratledge [17];
Lipp and Anklam [140]; Papanikolaou and Aggelis [22]).
Concerning the utilization of enzymes, it is, in fact, one of the
first (and of primordial importance) axes of lipid biotechnol-
ogy that has been developed in the late 80s. This approach
was related with the utilization of hydrolytic enzymes (prin-
cipally lipases) in media presenting a feeble water concen-
tration in order to synthesize specified TAGs by reactions of
trans- and/or inter-esterification of various plant lipids (e.g.,
trans-esterification of palm oil from stearic acid resulting
in the synthesis of TAGs of the types P–O–S and S–O–S)
[3, 5, 6]. The last years, reactions of the above type have been
ameliorated, optimized, and carried out in larger scale, with,
principally, utilization of various low-cost fatty materials
(e.g., lard, pomace olive oil, etc.) as substrates in order to
create various CBSs [8–11].
1060 S. Papanikolaou and G. Aggelis Eur. J. Lipid Sci. Technol. 2011, 113, 1052–1073
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

Table 2. Lipid production through de novo fatty acid biosynthesis pathwayduring growth of oleaginous yeasts on various carbon sources and
fermentation configurations
Microorganism Culture mode Carbon source Xa) (g/L) Lipid (% w/w) Reference
Candida sp. 107 Single-stage continuous Glucose 18.1 37.3 Gill et al. [137]
C. curvatus ’’ Glucose 13.5 29.0 Evans and Ratledge [49]
’’ ’’ Sucrose 16.0 28.0 ’’
’’ ’’ Lactose 18.0 31.0 ’’
’’ ’’ Xylose 15.0 37.0 ’’
Cryptococcus albidus Shake flasks Xylose 7.3 33.0 Hansson and Dostalek [132]
’’ ’’ Glucose 8.2 40.1 ’’
’’ ’’ Maltose 8.2 37.7 ’’
’’ ’’ Lactose 6.5 26.3 ’’
’’ ’’ Glycerol (pure) 1.4 43.8 ’’
Rhodosporidium toruloides ’’ Glucose 8.0 42.5 Moreton [15]
’’ ’’ Fructose 7.0 27.0 ’’
’’ ’’ Glycerol (pure) 5.8 34.4 ’’
’’ Batch bioreactor Glucose 12.5 42.9 ’’
’’ ’’ Fructose 8.7 39.8 ’’
’’ ’’ Xylose 8.3 42.2 ’’
A. curvatumb) Single-stage continuous Whey 24.1 37.4 Davies [33]
’’ ’’ ’’ 20.8 44.3 ’’
’’ ’’ ’’ 21.2 43.4 ’’
C. curvatus Batch bioreactor ’’ 21.6 36.0 Ykema et al. [44]
’’ Continuous-recycling ’’ 85.0 35.0 ’’
C. curvata Single-stage continuous Lactose 20.0 40.0 Brown et al. [52]
A. curvatum Batch bioreactor Glucose n.r.c) 35.5 Hassan et al. [38]
’’ Single-stage continuous ’’ 14.5 45.6 ’’
C. curvatus Batch bioreactor Prickly pear juice 10.9 45.8 Hassan et al. [74]
’’ ’’ ’’ 11.1 43.2 Hassan et al. [75]
’’ Fed-batch bioreactor Glycerol (pure) 118.0 25.0 Meesters et al. [45]
’’ Fed-batch air-lift bioreactor ’’ 91.0 32.0 Meesters et al. [46]
Y. lipolytica Single-stage continuous Glucose 9.2 25.0 Aggelis and Komaitis[138]
’’ Single-stage continuous Glycerol (raw) 8.1 43.0 Papanikolaou and Aggelis [47]
R. toruloides Fed-batch bioreactor Glucose 106.5 68.1 Li et al. [23]
’’ Shake flasks Glucose & sewage sludge 9.4 68.0 Angerbauer et al. [24]
Lipomyces starkeyi ’’ Glucose & xylose 20.5 61.5 Zhao et al. [25]
Trichosporon fermentans ’’ Glucose 24.1 56.6 Zhu et al. [27]
’’ ’’ Sucrose 19.5 62.6 ’’
’’ ’’ Xylose 17.1 57.8 ’’
’’ ’’ Lactose 16.9 49.6 ’’
’’ ’’ Fructose 21.5 40.7 ’’
’’ ’’ Molasses 36.4 35.3 ’’
Y. lipolytica ’’ Glycerol (raw) 6.5 30.7 Andre et al. [70]
R. toruloides ’’ Glucose 15.0 62.0 Hu et al. [136]
T. fermentans ’’ Rice straw hydrolysate 28.6 40.1 Huang et al. [48]
’’ ’’ Mannose 22.7 50.4 ’’
’’ ’’ Galactose 23.6 59 ’’
’’ ’’ Cellobiose 15.8 65.6 ’’
Rhodotorula mucilaginosa Shake-flasks Starch hydrolysate 21.8 53.0 Li et al. [76]
C. curvatus Fed-batch bioreactor Glycerol (raw) 32.9 52.9 Liang et al. [139]
Y. lipolyticac) Fed-batch bioreactor Glycerol (pure) 4.7 23.1 Makri et al. [72]
R. toruloides Shake-flasks Glucose 19.9 63.3 Wu et al. [29]
’’ Fed-batch bioreactor Jerusalem artichoke extract 39.6 56.5 Zhao et al. [79]
Rhodotorula mucilaginosa ’’ Inulin hydrolysate 14.4 49.0 Zhao et al. [77]
’’ Fed-batch bioreactor Jerusalem artichoke hydrolysate 19.5 52.1 ’’
Y. lipolyticad) Shake-flasks Inulin 13.3 48.3 Zhao et al. [78]
’’ ’’ Jerusalem artichoke extract 14.6 50.6 ’’
Trichosporon capitatum ’’ Molasses 17.0 35.2 Wu et al. [59]
’’ ’’ Glucose 15.4 43.0 ’’
a) X is the dry cell weight produced (in g/L).b) Culture at a constant dilution rate (¼ 0.04 h�1) and various pH values of the medium.c) During these cultures, nitrogen limitation led also to remarkable production of citric acid into the culture medium.d) Genetically engineered transformant Z31 capable of consuming inulin-based media.
Eur. J. Lipid Sci. Technol. 2011, 113, 1052–1073 The biotechnology of oleaginous yeasts 1061
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

Numerous investigators have utilized oleaginous micro-
organisms, and principally yeasts, that can be considered as
‘‘perfect’’ candidates for this purpose [18, 22]. The fact that
thesemicroorganisms stock their lipids principally in the form
of TAGs esterified in the sn-2 position by unsaturated fatty
acids, has favored this approach [16, 17]. However, the main
drawback to alleviate was how to increase the C18:0 and,
generally, the TSAF content inside the yeast cells [20, 22,
140], whilst in most instances the oleaginous yeasts accumu-
late unsaturated fatty acids to more than 65% w/w, in their
total lipids. Many strategies concerning the increase of TSFA
content in the yeasts have been realized so far and they will be
presented in the next paragraphs.
2.3.1 Strategies dealing with the production of CBSsby oleaginous yeasts
The first of the approaches carried out in order to produce a
microbial analogous of cocoa butter, consisted of a conven-
tional culture of an oleaginous yeast (e.g., R. toruloides,
L. lipofer, or Rhodotorula graminis) on glucose in nitrogen-
limited media followed by a separation of the synthesized
cellular lipid. The unit operation proposed was either crys-
tallization fractional separation process in order to finally
recover the fractions S–U–S (S: saturated fatty acid, U:
unsaturated fatty acid) of the produced SCO [143].
Following this type of operation, TAGs of type P–O–P
and P–O–S have been purified and utilized in the fabrication
of chocolate, replacing cocoa butter. The result was revealed
satisfactory considering both fusion point and organoleptic
properties of the product [143]. The inconvenient of this
approach is the fact that only the 35% w/w, of the produced
fat is presented in the form S–U–S [18].
Another strategy used in order to increase the C18:0
amount of the yeast lipids produced, was based on the prin-
ciple that plants, animals, and microorganisms do not pro-
duce their unsaturated fatty acids directly; firstly, a formation
of a saturated precursor is performed, and then, by virtue of
sequential desaturation reactions, double bonds are intro-
duced firstly in the position D9 and subsequently in the
positions D12 and D15 (it is noted that the desaturation in
the position D15 is unusual in the oil-bearing yeasts, whereas
humans are incapable of de novo introducing double bonds
after the 9th carbon in the aliphatic chain) [15, 20, 50]. The
acylated groups desaturation pathway is illustrated in
Figure 2 [17]. Consequently the desaturation activity could
be restricted using various desaturase inhibitors. Sterculic
and malvalic acid are cyclopropene fatty acids found in the
seed-oil of different plants of the families Malvaceae and
Sterculiaceae like sterculia oil and to lesser extent to kapok
oil [144]. Being fatty acids of 17 or 18 carbons with a cyclo-
propenic group in the positionD9, sterculic and malvalic acid
have been reported to inhibit the desaturation activity in
various plants and animals. In fact, these cyclopropene fatty
acids are structural analogous of the natural substrates for the
D9 desaturase enzymes, which carry out the conversion:
C18:0 ! D9C18 : 1. These substances have been equally
utilized in order to modify the accumulated lipid produced
by the oleaginous yeasts [15, 50, 145]. Numerous strains
have been tested (Candida sp. 107, Trichosporon cutaneum,
L. starkeyi,R. toruloides etc). Although in some cases relatively
significant quantities of the cyclopropenic inhibitor have been
added into the culture medium (e.g., up to 4.0 mL/L of
sterculia oil, that indicates presence of more than 2.0 g/L
of cyclopropene fatty acids), cell growth and lipid accumu-
lation have not been altered (total biomass always ranging
between 6.4 and 12.3 g/L containing fat >25% w/w, inside
the yeast cells) [50]. Some results of the production of bio-
mass and lipid and the composition of cellular lipid produced
when cyclopropen fatty acids are into the medium are
depicted in Table 3.
The addition of sterculia oil (containing around 50%w/w,
of sterculic acid) into the culture medium resulted in the
synthesis of a yeast lipid in which the concentration of the
fatty acid C18:0 increased from 3–5% up to �40% w/w [15].
However, this inhibitor having no effect upon the D12 desa-
turation (D9C18 : 1 ! D9;12C18 : 2), the yeast oil presented
relatively high linoleic acid amounts (around 14–24%w/w, of
total lipid). This fatty acid, however, is found in minimal
amounts into the cocoa butter (ranging between 2 and 5%w/w,
of total lipid). In order to produce, hence, with this strategy a
cocoa butter-like yeast lipid, a second inhibition in the D12
position had to be performed. A synthetic D12 cyclopropene
inhibitor, namely ‘‘cis-methylen-octadecenoıc acid’’ was
used, and the obtained result is depicted in Table 4 [15, 145].
The utilization of desaturase inhibitors remarkably
increases the saturation of the SCO produced, having as a
result the synthesis of lipids presenting composition sim-
ilarities with the cocoa butter. However, this strategy presents
two fundamental drawbacks, namely the increased coast of
Triacylglycerols
Acetyl-CoA + Malonyl-CoA
Palmitoyl-CoA Elongase
Stearoyl-CoA ∆9-desaturase
Oleoyl-CoA Transferase
Oleoyl-phosphorolipid
Linoleoyl-phosphorolipid
∆12-desaturase
Figure 2. Pathways of desaturation of acylated groups in the olea-
ginous yeasts (from Ratledge [17], adapted).
1062 S. Papanikolaou and G. Aggelis Eur. J. Lipid Sci. Technol. 2011, 113, 1052–1073
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

the inhibitors used and the fact that these products may
provoke mutagenesis and cancerogenesis [17]. As far as
the second disadvantage is concerned, this is of major import-
ance, taking into consideration the current trends of biotech-
nology, in which eco-friendly and ‘‘healthy’’ approaches
should be considered in order for the mass production of
biotechnological products [22]. By taking into consideration
that addition of other compounds could potentially interfere
with lipid desaturation [15], in order to alleviate this pre-
viously mentioned serious disadvantage, addition into the
culture medium of other natural compounds that are com-
pletely non-toxic could potentially be envisaged. One case
refers to the addition into the culture medium of the essential
oil of the plant Citrus sinensis cv New Hall –Citrus aurantium (a
natural and ‘‘healthy’’ compound), the addition of which,
even in very small concentrations (e.g., 0.3 mL/L) could
result in significant rise in the TSFA content of the yeast
Y. lipolytica [146]. Although in the culture conditions tested
no virtual accumulation of lipid inside the yeast cell occurred,
the fact that TSFAs significantly increased from 24% to 35–
43% w/w, indicates that addition of this compound into the
culture medium in conditions enhancing lipid accumulation,
could result in the synthesis of microbial CBSs [146].
Genetic manipulation strategies based on the destruction
of the gene encoding for D9 dehydrogenase, which is the
responsible enzyme for the biotransformation C18:0 !D9C18 : 1, have also been performed [35, 36, 38, 75, 147,
148]. In most of the cases D9-defective mutants of
Table 3. Effect of the utilization of sterculia oil upon biomass and lipid production and fatty acid composition of the cellular lipids produced by
oleaginous yeasts growing in shake-flask nitrogen-limited experiments with glucose utilized as the sole substrate
Fatty acid composition (% w/w)
Sterculia oil (mL/L) Xa) (g/L) Lb) (g/L) Lipid (% w/w) C16:0 C18:0 C18:1 C18:2
Candida sp. 107 0.0 9.9 3.1 31 28 5 33 27
0.2 6.4 1.5 25 28 33 10 23
2.0 7.1 2.7 37 25 44 9 15
Tricosporon cutaneum 0.0 9.9 2.8 28 29 7 50 13
0.2 8.4 2.5 30 32 24 30 14
2.0 7.6 2.4 32 35 23 25 13
R. toruloides 0.0 12.3 3.7 30 16 4 42 29
0.2 10.1 3.6 36 15 23 24 15
2.0 10.3 3.3 32 13 41 18 15
Cultivation was performed for 4 days (data from Moreton [50]).a) X is the dry cell weight produced (in g/L).b) L is the total cellular lipid produced (in g/L).
Table 4. Effect of the utilization ofD9 andD12 inhibitors upon biomass and lipid production and fatty acid the composition of the cellular lipids
by R. toruloides and A. curvatum
Fatty acid composition (% w/w)
Xa) (g/L) Lipid (% w/w) C16:0 C18:0 C18:1 C18:2
R. toruloides
Control 7.1 24 28 7 39 18
Sterculia oil (0.3 mL/L) 7.6 23 15 48 19 9
D12 inhibitor (0.4 ml/L) 7.8 20 36 8 45 2
Sterculia oil and D12 inhibitor (each 0.3 mL/L) 8.0 34 20 47 22 5
A. curvatum
Control 6.7 49 32 16 44 8
Sterculia oil (0.1 mL/L) 8.6 55 27 26 35 8
Sterculia oil (0.4 mL/L) and D12 inhibitor (4 mL/L) 8.0 52 41 25 28 T
Cocoa butter 23–30 32–37 30–37 2–4
Shake-flask nitrogen-limited experiments with glucose utilized as the sole substrate were done. Cultivation was performed for 4 days (data
from Moreton and Clode [145]).
T < 0.5% w/w.a) X is the dry cell weight produced (in g/L).
Eur. J. Lipid Sci. Technol. 2011, 113, 1052–1073 The biotechnology of oleaginous yeasts 1063
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

A. curvatum have been constructed; cellular suspensions of
the wild strain were treated with various mutagenic factors
like N-methyl-N-nitro-N-nitrosoguanidine (MNNG), ethyl-
methanesulfonate or UV irradiation [35, 36, 38, 75, 141,
147, 148], and then have been grown in a medium supple-
mentary in oleic acid. The colonies were transferred in a
medium without supplementary oleic acid. In general, the
formed colonies in the two types of substrates have been
screened extensively, and small numbers of colonies of aux-
otroph mutants in the unsaturated fatty acids (Ufa mutants)
have been obtained [36, 141], since also fatty acid synthetase
(FAS) mutants have been created [35, 147]. In most of the
cases, the addition of a D9C18 : 1-donor (even in small con-
centrations) is obligatory, since the Ufa mutants may not be
capable at all of synthesizing the above-mentioned fatty acid.
The cellular lipids isolated from the Ufa mutants grown in a
medium supplementary in oleic acid, have been analyzed and
compared with these of the wild strain A. curvatum. The
result is illustrated in Table 5 [141]. A correct use of this
fatty acid with a sugar as co-substrate, results in the synthesis
of a cellular CBS by the various Ufa mutants [141]. For
instance, in the case of the Ufa-25, batch-bioreactor culti-
vations were performed in whey-permeate enriched with a
small quantity of D9C18 : 1 donor (namely rapeseed oil added
in a rate of 0.08 g/g of lactose into the medium), and after
120 h of cultivation a lipid quantity of around 6.0 g/L cor-
responding to lipid in dry weight 41.0% w/w, was achieved,
while SCO produced presented a composition in fatty acids
similar with that of cocoa butter (C18:0–40% w/w, C16:0–
25% w/w, D9C18:1–20% w/w, D9;12C18:2–10% w/w – Ykema
et al. [149]). In a similar manner, the Ufa mutant M3 was
cultivated in batch and single-stage continuous nitrogen-lim-
ited cultures with glucose utilized as substrate (initial or inlet
glucose concentration at 30 g/L, initial C/N � 40 g/g), and
oleic acid and Tween 80 added in small quantities into the
medium (0.2 and 0.5 g/L, respectively), and significant bio-
mass production (biomass yield on glucose consumed
�0.5 g/g) was observed, in both batch and continuous cul-
tures. For the chemostat experiments low D values (i.e.,
ranging between 0.040 and 0.068 h�1) were required for
high SCO production, and maximum lipid in dry weight
to �40–46% w/w, with dry cell mass of �15 g/L for several
steady-states was achieved [38]. The fatty acid composition
of cellular lipid produced was close to that of cocoa butter
[38]. In order to avoid oleic acid addition, revertants of the
Ufa mutants have been chosen, with partially restored D9
desaturase activity [36, 141]. The obtained result is presented
in Table 6.
In another genetic approach aiming at the production of
CBS-SCOs, the D9 fatty acid desaturase gene of C. curvatus
ATCC 20509 was cloned and characterized [148]. This gene
presented a high G þ C content (of 61%) and displayed a
codon usage different in comparison with the one of the non-
oleaginous yeast Saccharomyces cerevisiae, but similar to that
of the basidiomycete Schizophyllum commune [148].
Moreover, the expression of the above gene was studied in
the presence of several fatty acids found into the growth
medium. Repression of desaturase mRNA signals was found
if fatty acids with a double bond at the D9 position were
presented, while fatty acids with a double bond found at
another position (e.g., D10 or D6) or saturated fatty acids
Table 5. Composition of cellular lipids of A. curvatumwild type (WT
– ATCC 20509) strain and auxotroph mutant Ufa-33 (data from Smit
et al. [141]).
Cellular
fatty acids
(% w/w)
WT Ufa 33
Without
supplement
Supplement
C18:1
Supplement
C18:1
C16:0 28 17 22
C18:0 14 11 36
C18:1 44 55 24
C18:2 2 9 10
TSFAsa) 44 31 62
a) TSFAs is the percentage of saturated fatty acids (w/w) in the
microbial lipids produced.
Table 6. Production of lipid by mutant revertants cultivated on diluted whey-permeate in batch laboratory-scale bioreactor experiments and
comparisons with data obtained by A. curvatum wild-type (WT – ATCC 20509) strain and cocoa butter
Xa) (g/L) Lb) (g/L) Lipid (% w/w)
Fatty acid composition (% w/w)
C16:0 C18:0 C18:1 C18:2
WT 4.4 1.2 27.2 28 15 43 8
R22-72 3.0 1.4 46.6 26 36 20 9
R25-75 3.5 1.6 45.7 29 30 28 8
Cocoa butter 23–30 32–37 30–37 2–4
Initial lactose concentration at 10 g/L, initial molar C/N ratio 30 moles/moles, and fermentation time imposed until lactose exhaustion from
the culture medium (data from Ykema et al. [36]).a) X is the dry cell weight produced (in g/L).b) L is the total cellular lipid produced (in g/L).
1064 S. Papanikolaou and G. Aggelis Eur. J. Lipid Sci. Technol. 2011, 113, 1052–1073
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

had no effect upon the transcription of the cloned gene [148].
It is evident that the genetic manipulation of the oleaginous
strains has presented very satisfactory results considering
both quantity of accumulated fats and cellular lipid compo-
sition, since the cellular stearic acid amount increases signifi-
cantly. However, not any process scale-up has been
performed, while, in addition, the wild cultures present
higher productivities compared with the mutants.
In order to obtain by a metabolic manipulation what the
genetic engineers have realized with the aid of mutations,
Davies et al. [37] have performed cultures of A. curvatum in
conditions of limited oxygen tension during the phase of lipid
accumulation. The idea was based on the fact that all of the
reactions of desaturation are oxygen dependant [18]. A
critical limitation of the oxygenation in the fermentation
medium could, hence, decrease the conversion percentage
of the reaction C18:0 ! D9C18:1. The obtained results (fer-
mentations performed in an air-lift bioreactor of 20 L
capacity) are presented in Table 7 [37]. It may be seen that
although the percentage of the fatty acid C18:0 into the
reserve lipids was not very high, the sum of saturated fatty
acids (C16:0, C18:0, C20:0, and C24:0) produced by
A. curvatum was in most of the cases higher than 50% w/
w, of total lipids. This fact represents, indeed, an interesting
result in a double coincidence; firstly because it has been
demonstrated with a completely eco-friendly method that the
oleaginous yeasts have in some circumstances the tendency to
accumulate lipids containing significant quantities of satu-
rated fatty acids. The second reason is that not any expensive
or dangerous method was applied [17]. Oxygen tension
decrease had almost not any negative effect upon cell growth
(�0.4 g of cells formed per 1 g of lactose consumed) and lipid
accumulation (around 37% w/w, of lipids in dry weight). In
addition, this approach is the only one having been extrapo-
lated in semi-industrial (500-L bubble column reactors –
Davies et al. [37]) and finally to industrial (13-m3 reactors
– Davies [42]) scale. In a similar manner, decrease in the
aeration rate during growth of oleaginousmolds of the species
M. circinelloides growing on small-chain organic acids,
resulted in increase of the TSFA content of the lipids pro-
duced, with concomitant increment of biosynthesis of sym-
metric TAGs of the type S–U–S, and therefore a CBS was
synthesized [51].
A simple method that has been applied in earlier or more
recent studies in relation with the production of SCOs pre-
senting somehow increased TSFA content is based on the
‘‘potentiality’’ itself of the oleaginous microorganism to store
relatively saturated fatty acids. For instance, in various cases
strains of A. curvatum (C. curvata – C. curvatus, i.e., strains D,
ATCC 20509, etc.) were capable to produce intra-cellular
lipids with somehow increased quantities of C16:0, resem-
bling, therefore, to palm oil [38, 49]. By performing simple
fermentation techniques like, i.e., the change in the incu-
bation temperature, it is possible to increase the TSFA con-
tent of lipids produced. In such a case, Wu et al. [30] have
performed growth of C. curvatus ATCC 20509 on N-acetyl-
glucosamine, and by shifting the incubation temperature
from T ¼ 308C to T ¼ 228C, the concentration of cellular
TSFAs remarkably increased (from�44 to�54%w/w) while
decrease of the temperature did not have remarkable effect
upon biomass and SCO produced. At T ¼ 228C, biomass of
�18 g/L containing fat at 50% w/w, was produced,
whereas the respective values at T ¼ 308C were 19.1 g/L
and 45%w/w, while interestingly enough, lipid was produced
as a primary anabolic activity despite the presence of nitrogen
into the medium [30]. Pan et al. [41] have performed an
extensive screening of soil-deriving yeasts, and have realized
cultures of the most promising strains on xylose and glycerol.
In some cases, SCO presenting composition close to cocoa
butter was synthesized, due to the spontaneous ability of
yeasts to produce lipids rich in the fatty acids C16:0 and
C18:0 [41]. Likewise, R. toruloides strain Y4 has been
revealed capable of producing significant lipid quantities
during growth on glucose-based media under phosphate-
[29] or sulphate- [150] limited conditions. The various
nutrient limitations applied induced differences (in some
Table 7. Effect of oxygen uptake rate (OUR) in the production and composition of cellular lipids byA. curvatum growing onwhey-permeate in
batch 20-L air-lift bioreactors
OUR (mmol/L/h) Xa) (g/L) Lipid (% w/w)
Fatty acid composition (% w/w)
C16:0 C18:0 C18:1 C18:2 C18:3 C20:0 C24:0
8–10 16.1 39.0 14 16 55 10 1 1 3
7.7 13.6 37.6 16 17 55 5 T 1 3
6.1 7.2 42.4 11 21 52 7 T 2 4
4.3 9.8 40.7 18 25 45 5 T 1 4
2.3 13.2 41.4 23 23 42 3 1 1 4
Initial lactose into the whey varying from 25 to 40 g/L (data from Davies et al. [37]).
T < 0.6% w/w.a) X is the dry cell weight produced (in g/L).
Eur. J. Lipid Sci. Technol. 2011, 113, 1052–1073 The biotechnology of oleaginous yeasts 1065
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

cases remarkable) in the fatty acid composition of the SCO
produced. Therefore, with increasing phosphate limitation
imposed, lipids remarkably enriched in the fatty acid D9C18:1
and less rich in the fatty acid C18:0 were produced [29].
Thus, at initial C/P ratio 72 moles/moles, the cellular con-
centrations of D9C18:1 and C18:0 were 50 and 19% w/w,
while at initial C/P ratio 9552 moles/moles, the respective
values were 62 and 6% w/w. On the other hand, with increas-
ing sulphate limitation imposed, the concentration of TSFAs
remarkably increased, and thus a CBS was created; when the
initial C/S molar ratio imposed was 255 moles/moles, the
concentration of TSFAs was�43% w/w, while the respective
value of TSFAs was �63% w/w (a value very close to cocoa
butter) when the initial C/S molar ratio was 18 310 moles/
moles [150]. In all cases, the TSFAs of R. toruloides were
C16:0 and to lesser extent C18:0 [29, 150].
The last strategy related with the production of a CBS by
oleaginous microorganisms, was based on the yeasts capacity
to grow on fat substrates rich in C18:0, C16:0, and D9C18:1
or their fatty derivatives (e.g., FAMEs of the above-men-
tioned fatty acids) alone or on mixtures with sugars or polyols
[39, 40, 90, 105, 110, 113, 116]. The hydrophobic materials
used as substrates were pure fatty acids like stearic acid [110],
methyl-, ethyl-, butyl-, or vinyl-esters of stearic and palmitic
acid [113] or mixtures of stearin (a negligible-cost industrial
derivative of tallow composed of saturated free-fatty acids
and mainly of C16:0 and C18:0) and hydrolyzed oleic rape-
seed oil (composed mainly of D9C18:1) [39, 105, 107, 108].
The microorganisms Candida sp., C. tropicalis,
C. guillermondii, Torulopsis sp., T. versatilis, Trichosporon sp.
[113], R. toruloides [110, 116], and Y. lipolytica [39, 40, 90,
105, 107, 108] have been used for this purpose. Inmany cases
the microorganisms possessed active desaturase systems (i.e.,
D9 and D12 dehydrogenases) [110, 113]. Even if the initial
fatty substrates used was globally saturated (utilization of
pure stearic acid or mixtures of stearic and palmitic acid
esters as substrates), the cellular lipids contained D9C18:1
(22–44% w/w) and D9;12C18:2 (4–8% w/w) in significant
quantities (Table 8, entries A–F – Matsuo et al. [113];
Gierhart [110]). It is interesting to indicate that in this
strategy relatively large-scale stirred tank bioreactors (100-
L) were utilized [113].
The case of Y. lipolytica strain ACA-DC 50109 [former
registration LGAM S(7)1] was relatively more complicated.
The response of this microorganism was studied on stearin
and this fat permitted an efficient cell growth and significant
lipid accumulation inside the yeast cells (�55%w/w, of lipids
in dry matter, lipid up to�8.0 g/L – Papanikolaou et al. [105,
106]). The microorganism assimilated for growth and main-
tenance the lower aliphatic chain fatty acids (C12:0 and
C14:0) and accumulated as reserve lipid mainly the fatty
acid C18:0. TSFAs at almost at 100% w/w, principally
composed of C18:0 (i.e., average cellular fatty acid compo-
sition at C18:0 ¼ 80–83% w/w, C16:0 ¼ 14–16% w/w,
and traces of C12:0, C14:0, and D9C18:1) were stored, indi-
cating preferential accumulation of C18:0 as storage cellular
fatty acid and negligible D9 desaturation activity. Therefore,
the obtained result was rather unexpected considering
the predominance of unsaturated fatty acids into the
reserve lipids of the yeasts [17]. An oleic acid donor thus,
Table 8. Utilization of pure fatty acids, their derivatives (fatty esters) or low-cost industrial fats in order to produce microbial substitutes of
cocoa butter
A B C D E F G H I Cocoa butter
La) (g/L) 8.7 15.5 12.3 4.1 10.9 9.8 3.4 2.9 1.9
Lipid (% w/w) 35 61 51 35 29 34 42 36 46
C16:0 26 32 26 13 35 39 17 14 21 23–29
C18:0 26 29 37 49 17 19 63 50 68 32–37
C18:1 39 25 22 26 32 23 12 24 8 30–37
C18:2 5 8 8 4 6 6 4 4 2 2–4
TSFAsb) 52 61 63 62 52 58 80 64 89 55–67
A: Candida guillermondii on 14.0 g/L of methyl-stearate and 6.0 g/L of methyl-palmitate in 150-L bioreactor [113].
B: Trichosporon sp. on 14.0 g/L of methyl-stearate and 6.0 g/L of methyl-palmitate in 150-L bioreactor [113].
C: Torulopsis sp. on 13.0 g/L of vinyl-stearate and 7.0 g/L of vinyl-palmitate in 150-L [113].
D: R. toruloides on 40 g/L of pure stearic acid in shake flasks [110].
E: C. guilliermondii on 14.0 g/L of methyl-stearate and 6.0 g/L of methyl-palmitate in 150-L bioreactor [113].
F: Torulopsis versatilis on 14.0 g/L of methyl-stearate and 6.0 g/L of methyl-palmitate in 150-L bioreactor [113].
G: Y. lipolytica on 7.5 g/L of stearin (industrial tallow derivative composed of saturated free-fatty acids) and 2.5 g/L of hydrolyzed oleic
rapeseed oil (HORO) in shake flasks [39].
H: Y. lipolytica on 5 g/L of stearin and 5 g/L of HORO in shake flasks [39].
I: Y. lipolytica on 3.5 g/L of stearin and 1.5 g/L of HORO in 1.5-L bioreactor [108].a) L is the total cellular lipid produced (in g/L).b) TSFAs is the percentage of saturated fatty acids (w/w) in the lipid.
1066 S. Papanikolaou and G. Aggelis Eur. J. Lipid Sci. Technol. 2011, 113, 1052–1073
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com
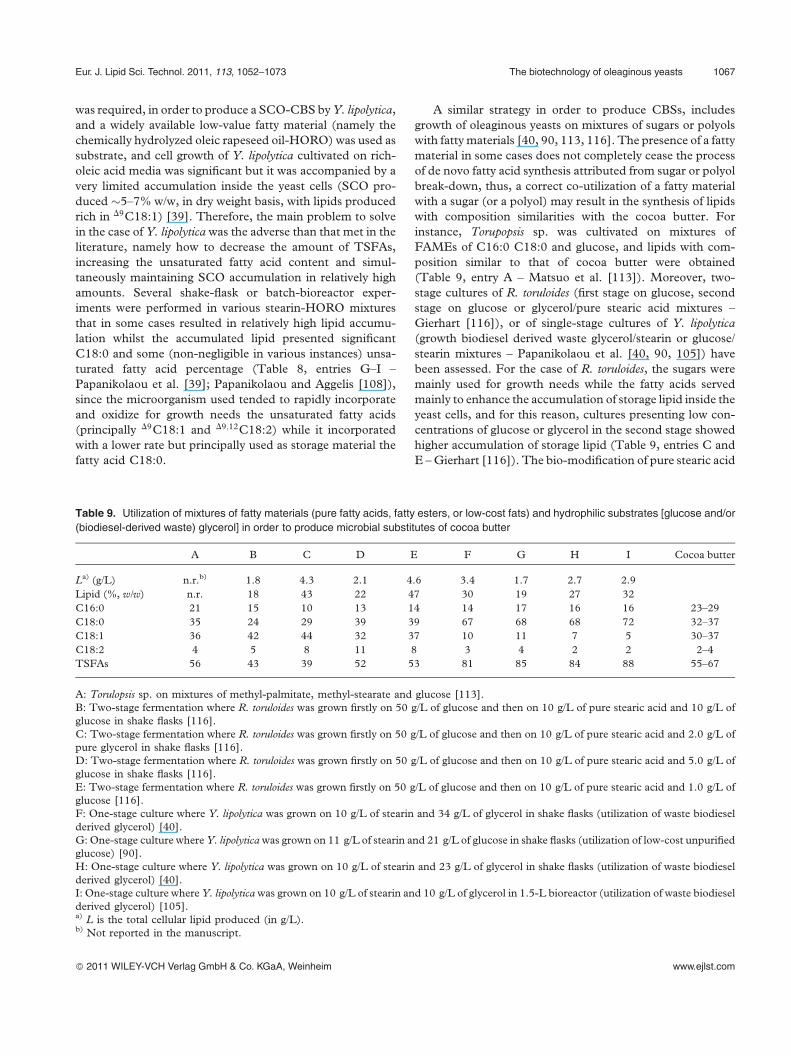
was required, in order to produce a SCO-CBS byY. lipolytica,
and a widely available low-value fatty material (namely the
chemically hydrolyzed oleic rapeseed oil-HORO) was used as
substrate, and cell growth of Y. lipolytica cultivated on rich-
oleic acid media was significant but it was accompanied by a
very limited accumulation inside the yeast cells (SCO pro-
duced �5–7% w/w, in dry weight basis, with lipids produced
rich in D9C18:1) [39]. Therefore, the main problem to solve
in the case of Y. lipolytica was the adverse than that met in the
literature, namely how to decrease the amount of TSFAs,
increasing the unsaturated fatty acid content and simul-
taneously maintaining SCO accumulation in relatively high
amounts. Several shake-flask or batch-bioreactor exper-
iments were performed in various stearin-HORO mixtures
that in some cases resulted in relatively high lipid accumu-
lation whilst the accumulated lipid presented significant
C18:0 and some (non-negligible in various instances) unsa-
turated fatty acid percentage (Table 8, entries G–I –
Papanikolaou et al. [39]; Papanikolaou and Aggelis [108]),
since the microorganism used tended to rapidly incorporate
and oxidize for growth needs the unsaturated fatty acids
(principally D9C18:1 and D9;12C18:2) while it incorporated
with a lower rate but principally used as storage material the
fatty acid C18:0.
A similar strategy in order to produce CBSs, includes
growth of oleaginous yeasts on mixtures of sugars or polyols
with fatty materials [40, 90, 113, 116]. The presence of a fatty
material in some cases does not completely cease the process
of de novo fatty acid synthesis attributed from sugar or polyol
break-down, thus, a correct co-utilization of a fatty material
with a sugar (or a polyol) may result in the synthesis of lipids
with composition similarities with the cocoa butter. For
instance, Torupopsis sp. was cultivated on mixtures of
FAMEs of C16:0 C18:0 and glucose, and lipids with com-
position similar to that of cocoa butter were obtained
(Table 9, entry A – Matsuo et al. [113]). Moreover, two-
stage cultures of R. toruloides (first stage on glucose, second
stage on glucose or glycerol/pure stearic acid mixtures –
Gierhart [116]), or of single-stage cultures of Y. lipolytica
(growth biodiesel derived waste glycerol/stearin or glucose/
stearin mixtures – Papanikolaou et al. [40, 90, 105]) have
been assessed. For the case of R. toruloides, the sugars were
mainly used for growth needs while the fatty acids served
mainly to enhance the accumulation of storage lipid inside the
yeast cells, and for this reason, cultures presenting low con-
centrations of glucose or glycerol in the second stage showed
higher accumulation of storage lipid (Table 9, entries C and
E –Gierhart [116]). The bio-modification of pure stearic acid
Table 9. Utilization of mixtures of fatty materials (pure fatty acids, fatty esters, or low-cost fats) and hydrophilic substrates [glucose and/or
(biodiesel-derived waste) glycerol] in order to produce microbial substitutes of cocoa butter
A B C D E F G H I Cocoa butter
La) (g/L) n.r.b) 1.8 4.3 2.1 4.6 3.4 1.7 2.7 2.9
Lipid (%, w/w) n.r. 18 43 22 47 30 19 27 32
C16:0 21 15 10 13 14 14 17 16 16 23–29
C18:0 35 24 29 39 39 67 68 68 72 32–37
C18:1 36 42 44 32 37 10 11 7 5 30–37
C18:2 4 5 8 11 8 3 4 2 2 2–4
TSFAs 56 43 39 52 53 81 85 84 88 55–67
A: Torulopsis sp. on mixtures of methyl-palmitate, methyl-stearate and glucose [113].
B: Two-stage fermentation where R. toruloides was grown firstly on 50 g/L of glucose and then on 10 g/L of pure stearic acid and 10 g/L of
glucose in shake flasks [116].
C: Two-stage fermentation where R. toruloides was grown firstly on 50 g/L of glucose and then on 10 g/L of pure stearic acid and 2.0 g/L of
pure glycerol in shake flasks [116].
D: Two-stage fermentation where R. toruloides was grown firstly on 50 g/L of glucose and then on 10 g/L of pure stearic acid and 5.0 g/L of
glucose in shake flasks [116].
E: Two-stage fermentation where R. toruloides was grown firstly on 50 g/L of glucose and then on 10 g/L of pure stearic acid and 1.0 g/L of
glucose [116].
F: One-stage culture where Y. lipolytica was grown on 10 g/L of stearin and 34 g/L of glycerol in shake flasks (utilization of waste biodiesel
derived glycerol) [40].
G: One-stage culture where Y. lipolytica was grown on 11 g/L of stearin and 21 g/L of glucose in shake flasks (utilization of low-cost unpurified
glucose) [90].
H: One-stage culture where Y. lipolytica was grown on 10 g/L of stearin and 23 g/L of glycerol in shake flasks (utilization of waste biodiesel
derived glycerol) [40].
I: One-stage culture where Y. lipolytica was grown on 10 g/L of stearin and 10 g/L of glycerol in 1.5-L bioreactor (utilization of waste biodiesel
derived glycerol) [105].a) L is the total cellular lipid produced (in g/L).b) Not reported in the manuscript.
Eur. J. Lipid Sci. Technol. 2011, 113, 1052–1073 The biotechnology of oleaginous yeasts 1067
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

found into the culture medium and the subsequent CBS
production was secured by the active D9 dehydrogenase
activity (Gierhart [110, 116]). Concerning Y. lipolytica,
growth on biodiesel derived glycerol or glucose (alone) in
nitrogen-limited flask cultures resulted in low lipid accumu-
lation, while cellular lipids were mainly composed of unsa-
turated fatty acids [68, 90]. Since growth on stearin alone
resulted in high SCOproduction with cellular lipid composed
mainly of the fatty acid C18:0 and given that some de novo
fatty acid accumulation occurs despite the presence of stearin
into the medium [40], waste glycerol (or glucose) were used
as unsaturated fatty acid precursors so as to produce SCO
with a remarkable content of TSFAs but also containing non-
negligible unsaturated fatty acids (Table 9, entries F–I –
Papanikolaou et al. [40, 90, 105]). Specifically the utilization
of biodiesel derived crude glycerol is of importance since even
though the fatty acid composition of the produced SCO
presented lower similarities with the cocoa butter compared
with the utilization of HORO as D9C18:1 fatty acid (compare
Tables 8 and 9), the current low cost of industrial glycerol
(ranging from 0 to 0.05US $ per kg) indicates the potentiality
of utilization of this co-substrate in the above-mentioned
strategy.
3 Concluding remarks – future perspectives
SCO fermentation attracts much interest the last years.
Amongst microbial lipids, yeast lipids present importance
in both academic and industrial point of view. Due to their
unicellular nature and their potentiality to grow on a plethora
of hydrophilic or hydrophobic substrates, the oleaginous
yeasts are considered as perfect ‘‘tools’’ for studying phenom-
ena of advanced lipid biochemistry and biotechnology [17,
22]. The increasing demand and utilization of 1st generation
biodiesel has notably increased the cost of various food-stuffs,
and this led to the necessity of discovery of non-conventional
sources of oils, that could be subsequently converted into
biodiesel. The oleaginous yeasts, which in most cases are
categorized as GRAS microorganisms and due to their fast
growth rates and their potentiality to grow on a plethora of
hydrophilic substrates, are considered as potential and inter-
esting candidates for the production of this 2nd generation
biodiesel.Moreover, the prospective remarkable increment in
the price of cocoa butter as well as the capability of various
oleaginous yeast strains to produce CBS-SCOs is also of
importance. Utilization of low- or even negative-cost
materials as substrates should be envisaged for both yeast
lipid applications previously mentioned. The removal of a
non-toxic and non-hazardous waste material from the food
industry, that will further be subjected to compost process
costs�0.1–0.3 US $ per kg of waste, therefore valorization of
these wastes with simultaneous production of cocoa butter-
like lipid or 2nd generation biodiesel with the aid of oleagi-
nous yeasts would have much to offer in both economical and
ecological points of view. As far as CBSs are concerned, waste
materials (like cheese–whey) can certainly be considered
again in the future as potential substrates for A. curvatum
strains. The same can be stressed in relation with the utiliz-
ation of stearin or other tallow derivatives (current cost of
tallow is around 0.3–0.4 US $ per kg) or even waste fats of
animal origin as substrates. However, in contrast with the use
of Torulopsis sp., R. toruloides, etc., employment of Y. lipolytica
as microorganism requires a donor of the fatty acid D9C18:1,
that should be cheaper than the rapeseed oil (crude glycerol
could be an option, but also other unsaturated fatty wastes
deriving from food production plants). Our research team is
willing to study in the future the biochemical and physiologi-
cal response of various Y. lipolytica strains (wild or genetically
engineered ones – e.g., strains lacking acyl-CoA oxidases) or
other oleaginous yeast strains (e.g., R. toruloides, L. starkeyi)
during their growth on various renewable substrates so as to
produce metabolites of added-value useful for the food and
chemical industry (SCO, citric acid, lipases, and exotic fats
substitutes).
Financial support concerning the results achieved by our research
team has been kindly provided by: Agricultural University of
Athens; University of Patras; Dracoil SA; Project BIOSIS
(INTERREG III GREECE – ITALY); State Scholarship
Foundation (Athens, Greece); General Secretary of Research and
Technology (Ministry of Development, Greek Government).
The authors have declared no conflict of interest.
References
[1] Buchholz, K., Bornscheuer, U. T., Highlights in biocatal-ysis – historical landmarks and current trends. Eng. Life Sci.2005, 5, 309–323.
[2] Adlercreutz, P., in: Carrea, G., Riva, S. (Eds.), OrganicSynthesis with Enzymes in Non-aqueous Media, Wiley-VCH, Weinheim (Germany) 2008, pp. 3–24.
[3] Metzger, J. O., Bornscheuer, U. T., Lipids as renewableresources: Current state of chemical and biotechnologicalconversion and diversification. Appl. Microbiol. Biotechnol.2006, 71, 13–22.
[4] Bottcher, D., Schmidt, M., Bornscheuer, U. T., in: Lutz,S., Bornscheuer, U. T. (Eds.), Protein Engineering Handbook,Wiley-VCH, Weinheim (Germany) 2009, pp. 753–775.
[5] Bloomer, S., Adlercreutz, P., Mattiasson, B., Triglycerideinteresterification by lipases. 1. Cocoa butter equivalentsfrom a fraction of palm oil. J. Am. Oil Chem. Soc. 1990, 67,519–524.
[6] Chang, M. K., Abraham, G., John, V. Y., Production ofcocoa butter-like fat from interesterification of vegetableoils. J. Am. Oil Chem. Soc. 1990, 67, 832–834.
[7] Soumanou, M. M., Bornscheuer, U. T., Menge, U.,Schmid, R. D., Synthesis of structured triglycerides frompeanut oil with immobilized lipase. J. Am. Oil Chem. Soc.1997, 74, 427–433.
[8] Zhao, H., Lu, Z., Lu, F., Liu, B. X., Zeng, X., Lipase-catalysed acidolysis of lard with caprylic acid to produce
1068 S. Papanikolaou and G. Aggelis Eur. J. Lipid Sci. Technol. 2011, 113, 1052–1073
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

structured lipid. Int. J. Food Sci. Technol. 2006, 41, 1027–1032.
[9] Liu, K. J., Chang, H. M., Liu, K. M., Enzymatic synthesisof cocoa butter analog through interesterification of lard andtristearin in supercritical carbon dioxide by lipase. FoodChem. 2007, 100, 1303–1311.
[10] Ciftci, O. N., Fadiloglu, S., Kowalski, B., Gogus, F.,Synthesis of cocoa butter triacylglycerols using a modelacidolysis system. Gracas y Aceites 2008, 59, 316–320.
[11] Ciftci, O. N., Fadiloglu, S., Kowalski, B., Gogus, F.,Utilization of olive-pomace oil for enzymatic productionof cocoa butter-like fat. J. Am. Oil Chem. Soc. 2009, 86,119–125.
[12] Aggelis, G., Komaitis, M., Pina, M., Graille, J., Specificityof Mucor miehei lipase on methyl ester substrates. Gracas yAceites 1993, 44, 331–334.
[13] Adamczak, M., Bornscheuer, U. T., Bednarski, W., Theapplication of biotechnological methods for the synthesis ofbiodiesel. Eur. J. Lipid Sci. Technol. 2009, 111, 800–813.
[14] Szczesna-Antczak, M., Kubiak, A., Antczak, T., Bielecki,S., Enzymatic biodiesel synthesis – key factors affectingefficiency of the process. Renew. Energy 2009, 34, 1185–1194.
[15] Moreton, R. S., in: Moreton, R. S. (Ed.), Single Cell Oil,Longman Scientific & Technical, Harlow (UK) 1988,pp. 1–32.
[16] Ratledge, C., in: Moreton, R. S. (Ed.), Single Cell Oil,Longman Scientific & Technical, Harlow (UK) 1988,pp. 33–70.
[17] Ratledge, C., in: Kamel, B. S., Kakuda, Y. (Eds.),Technological Advances in Improved and Alternative Sourcesof Lipids, Blackie Academic and Professional, London (UK)1994, pp. 235–291.
[18] Davies, R. J., Holdsworth, J. E., Synthesis of lipids in yeasts:Biochemistry, physiology and production. Adv. Appl. LipidRes. 1992, 1, 119–159.
[19] Certik, M., Shimizu, S., Biosynthesis and regulation ofmicrobial polyunsaturated fatty acid production. J. Biosci.Bioeng. 1999, 87, 1–14.
[20] Ratledge, C., Wynn, J., The biochemistry and molecularbiology of lipid accumulation in oleaginous microorgan-isms. Adv. Appl. Microbiol. 2002, 51, 1–51.
[21] Ratledge, C., Cohen, Z., Microbial and algal lipids: Do theyhave a future for biodiesel or as commodity oils? LipidTechnol. 2008, 20, 155–160.
[22] Papanikolaou, S., Aggelis, G., Yarrowia lipolytica: A modelmicroorganism used for the production of tailor-made lip-ids. Eur. J. Lipid Sci. Technol. 2010, 112, 639–654.
[23] Li, Y. H., Zhao, Z. B., Bai, F. W., High density cultivationof oleaginous yeastRhodosporidium toruloides Y4 in fed-batchculture. Enzyme Microb. Technol. 2007, 41, 312–317.
[24] Angerbauer, C., Siebenhofer, M., Mittelbach, M., Guebitz,G.M., Conversion of sewage sludge into lipids by Lipomycesstarkeyi for biodiesel production. Bioresour. Technol. 2008,99, 3051–3056.
[25] Zhao, X., Kong, X., Hua, Y., Feng, B., Zhao, Z. B.,Medium optimization for lipid production through co-fer-mentation of glucose and xylose by the oleaginous yeastLipomyces starkeyi. Eur. J. Lipid Sci. Technol. 2008, 110,405–412.
[26] Luque, R., Herrero-Davila, L., Campelo, J. M., Clark,J. H., et al., Biofuels: A technological perspective. EnergyEnviron. Sci. 2008, 1, 542–564.
[27] Zhu, L. Y., Zong,M. H., Wu, H., Efficient lipid productionwith Trichosporon fermentans and its use for biodiesel prep-aration. Bioresour. Technol. 2008, 99, 7881–7885.
[28] Papanikolaou, S., Aggelis, G., Biotechnological valorizationof biodiesel derived glycerol waste through production ofsingle cell oil and citric acid by Yarrowia lipolytica. LipidTechnol. 2009, 21, 83–87.
[29] Wu, S., Hu, C., Jin, C., Zhao, X., Zhao, Z. B., Phosphate-limitation mediated lipid production by Rhodosporidium tor-uloides. Bioresour. Technol. 2010, 101, 6124–6129.
[30] Wu, S., Hu, C., Zhao, X., Zhao, Z. B., Production of lipidfrom N-acetylglucosamine by Cryptococcus curvatus. Eur. J.Lipid Sci. Technol. 2010, 112, 727–733.
[31] Vamvakaki, A.-N., Kandarakis, I., Kaminarides, S.,Komaitis, M., Papanikolaou, S., Cheese whey as a renew-able substrate for microbial lipid and biomass production byZygomycetes. Eng. Life Sci. 2010, 10, 348–360.
[32] Chisti, Y., Biodiesel from microalgae. Biotechnol. Adv.2007, 25, 294–306.
[33] Davies, R. J., in: Moreton, R. S. (Ed.), Single Cell Oil,Longman Scientific & Technical, Harlow (UK) 1988,pp. 99–146.
[34] Ratledge, C., Single cell oils – have they a biotechnologicalfuture? Trends Biotechnol. 1993, 11, 278–284.
[35] Ykema, A., Verbree, E. C., Nijkamp, H. J. J., Smit, H.,Isolation and characterization of fatty acid auxotrophs fromthe oleaginous yeast Apiotrichum curvatum. Appl. Microbiol.Biotechnol. 1989, 32, 76–84.
[36] Ykema, A., Verbree, E. C., Verwoert, I. I. G. S., Van derLinden, K. H., et al., Lipid production of revertants of Ufamutants from the oleaginous yeast Apiotrichum curvatum.Appl. Microbiol. Biotechnol. 1990, 33, 176–182.
[37] Davies, R. J., Holdsworth, J. E., Reader, S. L., The effect oflow oxygen uptake rate on the fatty acid profile of theoleaginous yeast Apiotrichum curvatum. Appl. Microbiol.Biotechnol. 1990, 33, 569–573.
[38] Hassan, M., Blanc, P. J., Granger, L. M., Pareilleux, A.,Goma, G., Lipid production by an unsaturated fatty acidauxotroph of the oleaginous yeast Apiotrichum curvatumgrown in single-stage continuous culture. Appl. Microbiol.Biotechnol. 1993, 40, 483–488.
[39] Papanikolaou, S., Chevalot, I., Komaitis, M., Aggelis, G.,Marc, I., Kinetic profile of the cellular lipid composition inan oleaginous Yarrowia lipolytica capable of producing acocoa butter substitute from industrial fats. Antonie vanLeeuwenhoek 2001, 80, 215–224.
[40] Papanikolaou, S., Muniglia, L., Chevalot, I., Aggelis, G.,Marc, I., Accumulation of a cocoa butter-like lipid byYarrowia lipolytica cultivated on agro-industrial residues.Curr. Microbiol. 2003, 46, 124–130.
[41] Pan, L.-X., Yang, D.-F., Shao, L., Li, W., et al., Isolation ofthe oleaginous yeasts from the soil and studies of their lipid-producing capacities. Food Technol. Biotechnol. 2009, 47,215–220.
[42] Davies, R. J., in: Kyle, D. J., Ratledge, C. (Eds.), IndustrialApplications of Single Cell Oils, AOCS Press, Champaign(USA) 1992, pp. 196–218.
Eur. J. Lipid Sci. Technol. 2011, 113, 1052–1073 The biotechnology of oleaginous yeasts 1069
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

[43] Meng, X., Yang, J., Xu, X., Zhang, L., et al., Biodieselproduction from oleaginousmicroorganisms. Renew. Energy2009, 34, 1–5.
[44] Ykema, A., Verbree, E. C., Kater, M. M., Smit, H.,Optimization of lipid production in the oleaginous yeastApiotrichum curvatum in whey permeate. Appl. Microbiol.Biotechnol. 1988, 29, 211–218.
[45] Meesters, P. A. E. P., Huijberts, G.N.M., Eggink, G., Highcell density cultivation of the lipid accumulating yeastCryptococcus curvatus using glycerol as a carbon source.Appl. Microbiol. Biotechnol. 1996, 45, 575–579.
[46] Meesters, P. A. E. P., van der Wal, H., Weusthuis, R.,Eggink, G., Cultivation of the oleaginous yeastCryptococcus curvatus in a new reactor with improved mixingand mass transfer characteristics (SUPER-R). Biotechnol.Tech. 1996, 10, 277–282.
[47] Papanikolaou, S., Aggelis, G., Lipid production byYarrowia lipolytica growing on industrial glycerol in asingle-stage continuous culture. Bioresour. Technol. 2002,82, 43–49.
[48] Huang, C., Zong, M. H., Wu, H., Liu, Q., Microbial oilproduction from rice straw hydrolysate by Trichosporon fer-mentans. Bioresour. Technol. 2009, 100, 4535–4538.
[49] Evans, C. T., Ratledge, C., A comparison of the oleaginousyeast Candida curvata, grown on different carbon sources incontinuous and batch culture. Lipids 1983, 18, 623–629.
[50] Moreton, R. S., Modification of fatty acid composition oflipid accumulating yeasts with cyclopropene fatty acid desa-turase inhibitors. Appl. Microbiol. Biotechnol. 1985, 22, 41–45.
[51] Roux, M. P., Kock, J. L. F., Botha, A., Du Preez, J. C.,et al., Mucor – a source of cocoa butter and gamma-linolenicacid. World J. Microbiol. Biotechnol. 1994, 10, 417–422.
[52] Brown, B. D., Hsu, K. H., Hammond, E. G., Glatz, B. A.,A relationship between growth and lipid accumulation inCandida curvata D. J. Ferm. Bioeng. 1989, 68, 344–352.
[53] Aggelis, G., Stathas, D., Tavoularis, N., Komaitis, M.,Composition of lipids produced by some strains ofCandida species. Production of single-cell oil in a chemostatculture. Folia Microbiol. 1996, 41, 299–302.
[54] Papanikolaou, S., Komaitis, M., Aggelis, G., Single cell oil(SCO) production by Mortierella isabellina grown on high-sugar content media. Bioresour. Technol. 2004, 95, 287–291.
[55] Papanikolaou, S., Sarantou, S., Komaitis, M., Aggelis, G.,Repression of reserve lipid turnover in Cunninghamella echi-nulata and Mortierella isabellina cultivated in multiple-lim-ited media. J. Appl. Microbiol. 2004, 97, 867–874.
[56] Papanikolaou, S., Diamantopoulou, P., Chatzifragkou, A.,Philippoussis, A., Aggelis, G., Suitability of low-cost sugarsas substrates for lipid production by the fungus Thamnidiumelegans. Energy Fuel. 2010, 24, 4078–4086.
[57] Fakas, S., Papanikolaou, S., Batsos, A., Galiotou-Panayotou, M., et al., Evaluating renewable carbon sourcesas substrates for single cell oil production byCunninghamellaechinulata and Mortierella isabellina. Biomass Bioenergy 2009,33, 573–580.
[58] Chatzifragkou, A., Fakas, S., Galiotou-Panayotou, M.,Komaitis, M., et al., Commercial sugars as substrates forlipid accumulation by Cunninghamella echinulata andMortierella isabellina fungi. Eur. J. Lipid Sci. Technol.2010, 112, 1048–1057.
[59] Wu, H., Li, Y., Chen, L., Zong, M. H., Production ofmicrobial oil with high oleic acid content by Trichosporoncapitatum. Appl. Energy 2011, 88, 138–142.
[60] Glatz, B. A., Hammond, E. G., Hsu, K. H., Baehman, L.,et al., in: Ratledge, C., Rattray, J. B. M., Dawson, P. S. S.(Eds.), Biotechnology for the Oils and Fats Industry, AOCSPress, Champaign (USA) 1984, pp. 163–176.
[61] Fakas, S., Papanikolaou, S., Galiotou-Panayotou, M.,Komaitis, M., Aggelis, G., Lipids of Cunninghamellaechinulata with emphasis to g-linolenic acid distributionamong lipid classes. Appl. Microbiol. Biotechnol. 2006, 73,676–683.
[62] Fakas, S., Galiotou-Panayotou, M., Papanikolaou, S.,Komaitis, M., Aggelis, G., Compositional shifts in lipidfractions during lipid turnover in Cunninghamella echinulata.Enzyme Microb. Technol. 2007, 40, 1321–1327.
[63] Fakas, S., Certik, M., Papanikolaou, S., Aggelis, G., et al.,g-Linolenic acid production by Cunninghamella echinulatagrowing on complex organic nitrogen sources. Bioresour.Technol. 2008, 99, 5986–5990.
[64] Fakas, S., Papanikolaou, S., Galiotou-Panayotou, M.,Komaitis, M., Aggelis, G., Organic nitrogen of tomatowaste hydrolysate enhances glucose uptake and lipidaccumulation in Cunninghamella echinulata. J. Appl.Microbiol. 2008, 105, 1062–1070.
[65] Chen, H. C., Chang, C. C., Production of g-linolenic acidby the fungus Cunninghamella echinulata CCRC 31840.Biotechnol. Prog. 1996, 12, 338–341.
[66] Chen, H. C., Liu, T. M., Inoculum effects on the pro-duction of g-linolenic acid by the shake culture ofCunninghamella echinulata CCRC 31840. Enzyme Microb.Technol. 1997, 21, 137–142.
[67] Papanikolaou, S., Galiotou-Panayotou, M., Fakas, S.,Komaitis, M., Aggelis, G., Lipid production by oleaginousMucorales cultivated on renewable carbon sources. Eur. J.Lipid Sci. Technol. 2007, 109, 1060–1070.
[68] Papanikolaou, S., Fakas, S., Fick, M., Chevalot, I., et al.,Biotechnological valorisation of raw glycerol dischargedafter biodiesel (fatty acid methyl-esters) manufacturingprocess: Production of 1,3-propanediol, citric acid andsingle cell oil. Biomass Bioenergy 2008, 32, 60–71.
[69] Fakas, S., Bellou, S., Makri, A., Aggelis, G., in: Aggelis, G.(Ed.), Microbial Conversions of Raw Glycerol, NovaScience Publishers Inc., New York (USA) 2009, pp. 85–99.
[70] Andre, A., Chatzifragkou, A., Diamantopoulou, P., Sarris,D., et al., Biotechnological conversions of biodiesel derivedcrude glycerol by Yarrowia lipolytica strains. Eng. Life Sci.2009, 9, 468–478.
[71] Andre, A., Diamantopoulou, P., Philippoussis, A., Sarris,D., et al., Biotechnological conversions of biodiesel derivedwaste glycerol into added-value compounds by higher fungi:Production of biomass, single cell oil and oxalic acid. Ind.Crops Prod. 2010, 31, 407–416.
[72] Makri, A., Fakas, S., Aggelis, G., Metabolic activities ofbiotechnological interest in Yarrowia lipolytica grown onglycerol in repeated batch cultures. Bioresour. Technol.2010, 101, 2351–2358.
[73] Chatzifragkou, A., Makri, A., Belka, A., Bellou, S., et al.,Biotechnological conversions of biodiesel derived wasteglycerol by yeast and fungal species. Energy 2011, 36,1097–1108.
1070 S. Papanikolaou and G. Aggelis Eur. J. Lipid Sci. Technol. 2011, 113, 1052–1073
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

[74] Hassan, M., Blanc, P. J., Pareilleux, A., Goma, G.,Production of single-cell oil from prickly pear juice fermen-tation by Cryptococcus curvatus grown in batch culture.World J. Microbiol. Biotechnol. 1994, 10, 534–537.
[75] Hassan, M., Blanc, P. J., Pareilleux, A., Goma, G.,Production of cocoa butter equivalents from prickly pearjuice fermentation by an unsaturated fatty acid auxotroph ofCryptococcus curvatus grown in batch culture. Proc. Biochem.1995, 30, 629–634.
[76] Li, M., Liu, G. L., Chi, Z., Chi, Z. M., Single cell oilproduction from hydrolysate of cassava starch by marine-derived yeast Rhodotorula mucilaginosa TJY15a. BiomassBioenergy 2010, 34, 101–107.
[77] Zhao, C. H., Zhang, T., Li, M., Chi, Z. M., Single cell oilproduction from hydrolysates of inulin and extract of tubersof Jerusalem artichoke by Rhodotorula mucilaginosa TJY15a.Proc. Biochem. 2010, 45, 1121–1126.
[78] Zhao, C. H., Cui, W., Liu, X. Y., Chi, Z. M., Madzak, C.,Expression of inulinase gene in the oleaginous yeastYarrowia lipolytica and single cell oil production from inu-lin-containing materials. Metabol. Eng. 2010, 12, 510–517.
[79] Zhao, X., Wu, S., Hu, C., Wang, Q., et al., Lipid pro-duction from Jerusalem artichoke by Rhodosporidium toru-loides Y4. J. Ind. Microbiol. Biotechnol. 2010, 37, 581–585.
[80] Economou, C. N., Aggelis, G., Pavlou, S., Vayenas, D. V.,Modeling of single-cell oil production under nitrogen-lim-ited and substrate inhibition conditions. Biotechnol. Bioeng.2011, 108, 1049–1055.
[81] Aggelis, G., Two alternative pathways for substrate assim-ilation byMucor circinelloides. Folia Microbiol. 1996, 41, 254–256.
[82] Roux, M. P., Kock, J. L. F., Du Preez, J. C., Botha, A., Theinfluence of dissolved oxygen tension on the production ofcocoa butter equivalents and gamma-linolenic acid byMucor circinelloides. Syst. Appl. Microbiol. 1995, 18, 329–334.
[83] Du Preez, J. C., Immelman, M., Kock, J. L. F., Kilian,S. G., The effect of acetic acid concentration on the growthand production of gamma-linolenic acid by Mucor circinel-loides CBS 203.28 in fed-batch culture. World J. Microbiol.Biotechnol. 1997, 13, 81–87.
[84] Fei, Q., Chang, H. N., Shang, L., Choi, J., et al., The effectof volatile fatty acids as a sole carbon source on lipidaccumulation by Cryptococcus albidus for biodiesel pro-duction. Bioresour. Technol. 2011, 102, 2695–2701.
[85] Eroshin, V. K., Krylova, N. I., Efficiency of lipid synthesisby yeast. Biotechnol. Bioeng. 1983, 325, 1693–1700.
[86] Yamauchi, H., Mori, H., Kobayashi, T., Shimizu, S., Massproduction of lipids by Lipomyces starkeyi in microcomputeraided fed-batch culture. J. Ferm. Technol. 1983, 61, 275–280.
[87] Emelyanova, E., Lipid and g-linolenic acid production byMucor inaquisporus. Proc. Biochem. 1997, 3, 173–177.
[88] Ykema, A., Verbree, E. C., Verseveld, H. W., Smit, H.,Mathematical modelling of lipid production by oleaginousyeasts in continuous cultures. Antonie Van Leeuwenhoek1986, 52, 491–506.
[89] Aggelis, G., Sourdis, J., Prediction of lipid accumulation –degradation in oleaginous micro-organisms growingon vegetable oils. Antonie Van Leeuwenhoek 1997, 72,159–165.
[90] Papanikolaou, S., Galiotou-Panayotou, M., Chevalot, I.,Komaitis, M., et al., Influence of glucose and saturated free-fatty acid mixtures on citric acid and lipid production byYarrowia lipolytica. Curr. Microbiol. 2006, 52, 134–142.
[91] Arzumanov, T., Sidorov, I., Shishkanova, N., Finogenova,T., Mathematical modeling of citric acid production byrepeated batch culture. Enzyme Microbiol. Technol. 2000,26, 826–833.
[92] Anastassiadis, S., Aivasidis, A., Wandrey, C., Citric acidproduction by Candida strains under intracellular nitrogenlimitation. Appl. Microbiol. Biotechnol. 2002, 60, 81–87.
[93] Papanikolaou, S., Aggelis, G., Modelling aspects of thebiotechnological valorization of raw glycerol: Productionof citric acid by Yarrowia lipolytica and 1,3-propanediolby Clostridium butyricum. J. Chem. Technol. Biotechnol.2003, 78, 542–547.
[94] Bati, N., Hammond, E. G., Glatz, B. A., Biomodification offats and oils: Trials with Candida lipolytica. J. Am. Oil Chem.Soc. 1984, 61, 1743–1746.
[95] Koritala, S., Hesseltine, C. W., Pryde, E. H., Mounts,T. L., Biochemical modification of fats by microorganisms:A preliminary study. J. Am. Oil Chem. Soc. 1987, 64, 509–513.
[96] Aggelis, G., Komaitis, M., Papanikolaou, S.,Papadopoulos, G., A mathematical model for the studyof lipid accumulation in oleaginous microorganisms. I.Lipid accumulation during growth of Mucor circinelloidesCBS172-27 on a vegetable oil. Gracas y Aceites 1995, 46,169–173.
[97] Aggelis, G., Komaitis, M., Papanikolaou, S.,Papadopoulos, G., A mathematical model for the studyof lipid accumulation in oleaginous microorganisms. II.Study of cellular lipids of Mucor circinelloides during growthon a vegetable oil. Gracas y Aceites 1995, 46, 245–250.
[98] Aggelis, G., Papadiotis, G., Komaitis, M., Microbial fattyacid specificity. Folia Microbiol. 1997, 42, 117–120.
[99] Najjar, A., Robert, S., Guerin, C., Violet-Asther, M.,Carriere, F., Quantitative study of lipase secretion, extra-cellular lipolysis, and lipid storage in the yeast Yarrowialipolytica grown in the presence of olive oil: Analogies withlipolysis in humans. Appl. Microbiol. Biotechnol. 2011, 89,1947–1962 .
[100] Montet, D., Ratomahenina, R., Galzy, P., Pina,M., Graille,J., A study of the influence of the growth media on the fattyacid composition in Candida lipolytica DIDDENS andLODDER. Biotechnol. Lett. 1985, 7, 733–736.
[101] Guo, X., Tomonaga, T., Yanagihara, Y., Ota, Y., Screeningfor yeasts incorporating the exogenous eicosapentaenoicand docosahexaenoic acids from crude fish oil. J. Biosci.Bioeng. 1999, 87, 184–188.
[102] Guo, X., Ota, Y., Incorporation of eicosapentaenoic anddocosahexaenoic acids by a yeast (FO726A). J. Appl.Microbiol. 2000, 89, 107–115.
[103] Fuchtenbusch, B., Wullbrandt, D., Steinbuchel, A.,Production of polyhydroxyalkanoic acids by Ralstonia eutro-pha and Pseudomonas oleovorans from an oil remaining frombiotechnological rhamnose production. Appl. Microbiol.Biotechnol. 2000, 53, 167–172.
[104] Kinoshita, H., Ota, Y., Concentration of docosahexaenoicacid from fish oils using Geotrichum sp. F O347-2. Biosci.Biotechnol. Biochem. 2001, 65, 1022–1026.
Eur. J. Lipid Sci. Technol. 2011, 113, 1052–1073 The biotechnology of oleaginous yeasts 1071
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

[105] Papanikolaou, S., Chevalot, I., Komaitis, M., Marc, I.,Aggelis, G., Single cell oil production by Yarrowia lipolyticagrowing on an industrial derivative of animal fat in batchcultures. Appl. Microbiol. Biotechnol. 2002, 58, 308–312.
[106] Papanikolaou, S., Chevalot, I., Galiotou-Panayotou, M.,Komaitis, M., et al., Industrial derivative of tallow: A prom-ising renewable substrate for microbial lipid, single-cellprotein and lipase production by Yarrowia lipolytica.Electron. J. Biotechnol. 2007, 10, 425–435.
[107] Papanikolaou, S., Aggelis, G., Modeling lipid accumulationand degradation in Yarrowia lipolytica cultivated on indus-trial fats. Curr. Microbiol. 2003, 46, 398–402.
[108] Papanikolaou, S., Aggelis, G., Selective uptake of fatty acidsby the yeast Yarrowia lipolytica. Eur. J. Lipid Sci. Technol.2003, 105, 651–655.
[109] Roux-Van der Merwe, M. P., Badenhorst, J., Britz, T. J.,Fungal treatment of an edible-oil-containing industrialeffluent. World J. Microbiol. Biotechnol. 2005, 21, 947–953.
[110] Gierhart, D. I., US Patent 4485, 173, 1984.
[111] Mlickova, K., Luo, Y., D’Andrea, S., Pec, P., et al., Acyl-CoA oxidase, a key step for lipid accumulation in the yeastYarrowia lipolytica. J. Mol. Catal. B: Enzym. 2004, 28, 81–85.
[112] Mlickova, K., Roux, E., Athenstaedt, K., D’Andrea, S.,et al., Lipid accumulation, lipid body formation, andacyl-coenzyme A oxidases of the yeast Yarrowia lipolytica.Appl. Environ. Microbiol. 2004, 70, 3918–3924.
[113] Matsuo, T., Terashima, M., Hashimoto, Y., Hasida, W.,US Patent 4308 350, 1981.
[114] Thorpe, R. F., Ratledge, C., Fatty acid distribution intriglycerides of yeasts grown on glucose and n-alkanes.J. Gen. Microbiol. 1972, 75, 151–163.
[115] Alvarez, H. M., Kalscheuer, R., Steinbuchel, A.,Accumulation of storage lipids in species of Rhodococcusand Nocardia and effect of inhibitors and polyethylene gly-col. Fett-Lipid 1997, 99, 239–246.
[116] Gierhart, D. I., US Patent 4485, 172, 1984.
[117] Certik,M., Balteszova, L., Sajbidor, J., Lipid formation andg-linolenic acid production by Mucorales fungi grown onsunflower oil. Lett. Appl. Microbiol. 1997, 25, 101–105.
[118] Aoki, H., Miyamoto, N., Furuya, Y., Mankura, M., et al.,Incorporation and accumulation of docosahexaenoic acidfrom the medium by Pichia methanolica HA-32. Biosci.Biotechnol. Biochem. 2002, 66, 2632–2638.
[119] Mantzouridou, F., Tsimidou,M., Roukas, T., Performanceof crude olive pomace oil and soybean oil during carotenoidproduction by Blakeslea trispora in submerged fermentation.J. Agric. Food Chem. 2006, 54, 2575–2581.
[120] Szczesna-Antczak, M., Antczak, T., Piotrowicz-Wasiak,M., Rzyska, M., et al., Relationships between lipases andlipids in mycelia of two Mucor strains. Enzyme Microb.Technol. 2006, 39, 1214–1222.
[121] Galiotou-Panayotou, M., Kalatzi, O., Aggelis, G.,Modelling of simultaneous production of polygalactorunaseand exopolysaccharide by Aureobasidium pullulans ATHUM2915. Antonie Van Leeuwenhoek 1998, 73, 155–162.
[122] Aggelis, G., Fakas, S., Melissis, S., Clonis, Y. D., Growth ofCandida boidinii in a methanol-limited continuous cultureand the formation of methanol-degrading enzymes.J. Biotechnol. 1999, 72, 127–139.
[123] Alper, H., Stephanopoulos, G., Engineering for biofuels:Exploiting innate microbial capacity or importingbiosynthetic potential? Nat. Rev. Microbiol. 2009, 7, 715–723.
[124] Lois, E., Definition of biodiesel. Fuel 2007, 7–8, 1212–1213.
[125] Mu, Y., Xiu, Z. L., Zhang, D. J., A combined bioprocess ofbiodiesel production by lipase with microbial production of1,3-propanediol by Klebsiella pneumoniae. Biochem. Eng. J.2008, 40, 537–541.
[126] Fakas, S., Papanikolaou, S., Galiotou-Panayotou, M.,Komaitis, M., Aggelis, G., in: Pandey, A., Larroche, C.,Soccol, C. R., Dussard, C. G. (Eds.), New Horizons inBiotechnology, AsiaTech Publishers Inc., New Delhi(India) 2009, pp. 38–60.
[127] Vicente, G., Bautista, F. F., Gutierrez, F. J., Rodrıguez, R.,et al., Direct transformation of fungal biomass from sub-merged cultures into biodiesel. Energy Fuel 2010, 24, 3173–3178.
[128] Granger, L. M., Perlot, P., Goma, G., Pareilleux, A.,Kinetics of growth and fatty acid production ofRhodotorula glutinis. Appl. Microbiol. Biotechnol. 1992, 37,13–17.
[129] Hansson, L., Dostalek, M., Effect of culture conditions onmycelian growth and production of g-linolenic acid by thefungus Mortierella ramanniana. Appl. Microbiol. Biotechnol.1988, 28, 240–246.
[130] Boulton, C., Ratledge, C., Correlation of lipid accumu-lation in yeasts with possession of ATP-citrate lyase.J. Gen. Microbiol. 1981, 127, 169–176.
[131] Wynn, J. P., Hamid, A. A., Li, Y., Ratledge, C.,Biochemical events leading to the diversion of carbon intostorage lipids in the oleaginous fungi Mucor circinelloidesand Mortierella alpina. Microbiology UK 2001, 147, 2857–2864.
[132] Hansson, L., Dostalek, M., Influence of cultivation con-ditions on lipid production by Cryptococcus albidus. Appl.Microbiol. Biotechnol. 1986, 24, 12–18.
[133] Evans, C. T., Ratledge, C., Effects of nitrogen source onlipid accumulation in oleaginous yeasts. J. Gen. Microbiol.1984, 130, 1693–1704.
[134] Saxena, V., Sharma, C. D., Bhagat, S. D., Saini, V. S.,Adhikari, D. K., Lipid and fatty acid biosynthesis byRhodotorula minuta. J. Am. Oil Chem. Soc. 1998, 75, 501–505.
[135] Ferrante, G., Ohno, Y., Kates, M., Influence of tempera-ture and growth phase on desaturase activity of the meso-philic yeast Candida lipolytica. Can. J. Biochem. Cell Biol.1983, 161, 171–177.
[136] Hu, C., Zhao, X., Zhao, J., Wu, S., Zhao, Z. B., Effects ofbiomass hydrolysis by-products on oleaginous yeastRhodosporidium toruloides. Bioresour. Technol. 2009, 100,4843–4847.
[137] Gill, C., Hall, M., Ratledge, C., Lipid accumulation in anoleaginous yeast (Candida 107) growing on glucose insingle-stage continuous culture. Appl. Environ. Microbiol.1977, 33, 231–239.
[138] Aggelis, G., Komaitis, M., Enhancement of single-cell oilproduction by Yarrowia lipolytica growing in the presence ofTeuchrium polium L. aqueous extract. Biotechnol. Lett. 1999,21, 747–749.
1072 S. Papanikolaou and G. Aggelis Eur. J. Lipid Sci. Technol. 2011, 113, 1052–1073
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

[139] Liang, Y., Cui, Y., Trushenski, J., Blackburn, J. W.,Converting crude glycerol derived from yellow grease tolipids through yeast fermentation. Bioresour. Technol.2010, 101, 7581–7586.
[140] Lipp, M., Anklam, E., Review of cocoa butter and alterna-tive fats for use in chocolate – Part A. Compositional data.Food Chem. 1998, 62, 73–97.
[141] Smit, H., Ykema, A., Verbree, E. C., Verwoert, I. I. G. S.,Kater, M. M., in: Kyle, D. J., Ratledge, C. (Eds.), IndustrialProduction of Single Cell Oils, AOCS Press, Champaign(USA) 1992, pp. 185–195.
[142] Kaphueakngam, P., Flood, A., Sonwai,1 S., Production ofcocoa butter equivalent from mango seed almond fat andpalm oil mid-fraction. As. J. Food Ag-Ind. 2009, 2, 441–447.
[143] Tatsumi, C., Hashimoto, Y., Terashima, M., Matsuo, T.,US Patent 4032 405, 1979.
[144] Yano, I., Nicholls, B. W., Morris, L. J., James, A. T., Thedistribution of cyclopropane and cyclopropene fatty acids inhigher plants (Malvaceae). Lipids 1972, 7, 30–34.
[145] Moreton, R. S., Clode, M., US Patent 4778 630, 1988.
[146] Papanikolaou, S., Gortzi, O., Margeli, E., Chinou, I., et al.,Effect of Citrus essential oil addition upon growth and
cellular lipids of Yarrowia lipolytica yeast. Eur. J. LipidSci. Technol. 2008, 110, 997–1006.
[147] Hassan, M., Blanc, P. J., Pareilleux, A., Goma, G.,Selection of fatty acid auxotrophs from the oleaginous yeastCryptococcus curvatus and production of cocoa butter equiv-alents in batch culture. Biotechnol. Lett. 1994, 16, 819–824.
[148] Meesters, P. A. E. P., Springer, J., Eggink, G., Cloning andexpression of the D9 fatty acid desaturase gene fromCryptococcus curvatus ATCC 20509 containing histidineboxes and a cytochrome b5 domain. Appl. Microbiol.Biotechnol. 1997, 47, 663–667.
[149] Ykema, A., Kater, M. M., Smit, H., Lipid production inwhey-permeate by an unsaturated fatty acid mutant of theoleaginous yeast Apiotrichum curvatum. Biotechnol. Lett.1989, 11, 477–482.
[150] Wu, S., Zhao, X., Shen, H., Wang, Q., Zhao, Z. B.,Microbial lipid production by Rhodosporidium toruloidesunder sulfate-limited conditions. Bioresour. Technol. 2011,102, 1803–1807.
[151] Papanikolaou, S., Aggelis, G., Lipids of oleaginous yeasts.Part I: Biochemistry of single cell oil production. Eur. J.Lipid. Sci. Technol. 2011, 1031–1051.
Eur. J. Lipid Sci. Technol. 2011, 113, 1052–1073 The biotechnology of oleaginous yeasts 1073
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com
Copyright © 2022 FDOKUMEN




















