
Lipids of oleaginous yeasts. Part I: Biochemistry of single cell oil production
21
Review Article Lipids of oleaginous yeasts. Part I: Biochemistry of single cell oil production Seraphim Papanikolaou 1 and George Aggelis 2 1 Laboratory of Food Microbiology and Biotechnology, Department of Food Science and Technology, Agricultural University of Athens, Athens, Greece 2 Unit of Microbiology, Department of Biology, Division of Genetics, Cell and Development Biology, University of Patras, Patras, Greece In the first part of this review, the biochemistry of lipid accumulation in the oleaginous microorganisms is depicted. Lipid biosynthesis form sugars and related substrates is a secondary anabolic activity, conducted after essential nutrient (usually nitrogen) depletion in the medium. Due to this exhaustion, the carbon flow is directed towards the accumulation of intracellular citric acid that is used as acetyl-CoA donor in the cytoplasm. Acetyl-CoA generates cellular fatty acids and subsequently triacylglycerols. Lipid accumulation from hydrophobic substrates is a growth associated process, being independent from nitrogen exhaustion in the medium. Medium fatty acids are incorporated with various incorporation rates and are either dissimilated for growth needs or become ‘‘substrate’’ for intracellular biotransformations. ‘‘New’’ fatty acid profiles (in both extra- and intracellular lipids) that did not previously exist in the medium are likely to be produced. Oleaginous microorganisms consume their own storage lipids when their metabolic abilities cannot be saturated by the extracellular carbon source. Reserve lipid breakdown is independent from the type of the carbon source used for lipid accumulation. In most cases it is accompanied by lipid-free biomass production. Lipid mobilization is a specific process, as preferential degradation of the neutral lipid fractions is observed. Keywords: Biodiesel / Lipid accumulation – degradation / Lipid biotechnology / Oleaginous microorganisms / Single cell oil Received: January 10, 2011 / Revised: March 19, 2011 / Accepted: April 12, 2011 DOI: 10.1002/ejlt.201100014 1 Introduction The utilization of oleaginous microorganisms (these that can accumulate lipids to more than 20% w/w, in their dry cell mass) as sources of oils and fats (the so-called single cell oils, SCOs) in large-scale operations presents a great industrial interest. Although the production price of SCOs is higher compared with the traditional utilization of common oils and fats due to the obligatory maintenance of aseptic conditions [1], several alternative options for large-scale production of SCOs exist, since the price of various naturally occurring lipids and fats of the plant and animal kingdom can tremen- dously vary (from 0.3 to up to 100 US $ per kg—see: Ratledge and Wynn [2]). Therefore identification of microorganisms capable of producing in increased quantities lipids with struc- ture and composition similar to that of high-value fats, and subsequent large-scale production can present an enormous financial interest [2]. Furthermore, various oleaginous micro- organisms have the potentiality to present remarkable growth and production of SCO on wastes and by-products of the argo-industrial sector [3–8]; thus, valorization of these resi- dues together with production of potentially high-added value lipid could increase the viability of the process being simultaneously beneficial for the environment. Moreover, the continuously increasing demand and utilization of the so- called ‘‘1st generation biodiesel’’ (fatty acid methyl-esters— FAMEs deriving from trans-esterification of principally plant oils) that has occurred the last 15 years has increased the price Correspondence: Dr. Seraphim Papanikolaou, Assistant Professor in Food Bioprocesses, Laboratory of Food Microbiology and Biotechnology, Department of Food Science and Technology, Agricultural University of Athens, 75 Iera Odos, 11855 Athens, Greece E-mail: [email protected] Fax: þ30-210-5294700 Abbreviations: Aox, acyl-CoA oxidases; DOT, dissolved oxygen tension (in %, v/v); Fru, fructose (in g/L); Glc, glucose (in g/L); Glol, glycerol (in g/L); L, total intra-cellular lipid (in g/L); PUFAs, polyunsaturated fatty acids; S, substrate fat (in g/L); SCO, single cell oil; TAGs, triacylglycerols; X, total biomass (in g/L); Xf, lipid-free biomass (in g/L) Eur. J. Lipid Sci. Technol. 2011, 113, 1031–1051 1031 ß 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com
Transcript of Lipids of oleaginous yeasts. Part I: Biochemistry of single cell oil production

Review Article
Lipids of oleaginous yeasts. Part I: Biochemistry of singlecell oil production
Seraphim Papanikolaou1 and George Aggelis2
1 Laboratory of Food Microbiology and Biotechnology, Department of Food Science and Technology,
Agricultural University of Athens, Athens, Greece2 Unit of Microbiology, Department of Biology, Division of Genetics, Cell and Development Biology, University
of Patras, Patras, Greece
In the first part of this review, the biochemistry of lipid accumulation in the oleaginous microorganisms is
depicted. Lipid biosynthesis form sugars and related substrates is a secondary anabolic activity,
conducted after essential nutrient (usually nitrogen) depletion in the medium. Due to this
exhaustion, the carbon flow is directed towards the accumulation of intracellular citric acid that is
used as acetyl-CoA donor in the cytoplasm. Acetyl-CoA generates cellular fatty acids and subsequently
triacylglycerols. Lipid accumulation from hydrophobic substrates is a growth associated process, being
independent from nitrogen exhaustion in the medium.Medium fatty acids are incorporated with various
incorporation rates and are either dissimilated for growth needs or become ‘‘substrate’’ for intracellular
biotransformations. ‘‘New’’ fatty acid profiles (in both extra- and intracellular lipids) that did not
previously exist in the medium are likely to be produced. Oleaginous microorganisms consume their
own storage lipids when their metabolic abilities cannot be saturated by the extracellular carbon source.
Reserve lipid breakdown is independent from the type of the carbon source used for lipid accumulation.
In most cases it is accompanied by lipid-free biomass production. Lipid mobilization is a specific process,
as preferential degradation of the neutral lipid fractions is observed.
Keywords: Biodiesel / Lipid accumulation – degradation / Lipid biotechnology / Oleaginous microorganisms / Single
cell oil
Received: January 10, 2011 / Revised: March 19, 2011 / Accepted: April 12, 2011
DOI: 10.1002/ejlt.201100014
1 Introduction
The utilization of oleaginous microorganisms (these that can
accumulate lipids to more than 20% w/w, in their dry cell
mass) as sources of oils and fats (the so-called single cell oils,
SCOs) in large-scale operations presents a great industrial
interest. Although the production price of SCOs is higher
compared with the traditional utilization of common oils and
fats due to the obligatory maintenance of aseptic conditions
[1], several alternative options for large-scale production of
SCOs exist, since the price of various naturally occurring
lipids and fats of the plant and animal kingdom can tremen-
dously vary (from 0.3 to up to 100US $ per kg—see: Ratledge
and Wynn [2]). Therefore identification of microorganisms
capable of producing in increased quantities lipids with struc-
ture and composition similar to that of high-value fats, and
subsequent large-scale production can present an enormous
financial interest [2]. Furthermore, various oleaginousmicro-
organisms have the potentiality to present remarkable growth
and production of SCO on wastes and by-products of the
argo-industrial sector [3–8]; thus, valorization of these resi-
dues together with production of potentially high-added
value lipid could increase the viability of the process being
simultaneously beneficial for the environment.Moreover, the
continuously increasing demand and utilization of the so-
called ‘‘1st generation biodiesel’’ (fatty acid methyl-esters—
FAMEs deriving from trans-esterification of principally plant
oils) that has occurred the last 15 years has increased the price
Correspondence: Dr. Seraphim Papanikolaou, Assistant Professor in
Food Bioprocesses, Laboratory of Food Microbiology and Biotechnology,
Department of Food Science and Technology, Agricultural University of
Athens, 75 Iera Odos, 11855 Athens, Greece
E-mail: [email protected]
Fax: þ30-210-5294700
Abbreviations: Aox, acyl-CoA oxidases; DOT, dissolved oxygen tension
(in %, v/v); Fru, fructose (in g/L); Glc, glucose (in g/L); Glol, glycerol (in
g/L); L, total intra-cellular lipid (in g/L); PUFAs, polyunsaturated fatty acids;
S, substrate fat (in g/L); SCO, single cell oil; TAGs, triacylglycerols; X, total
biomass (in g/L); Xf, lipid-free biomass (in g/L)
Eur. J. Lipid Sci. Technol. 2011, 113, 1031–1051 1031
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

of various foodstuffs, and this situation has led to the neces-
sity of discovery of non-conventional sources of oils, that
could be subsequently converted into biodiesel. The oleagi-
nous microorganisms (yeasts, molds, and algae) are con-
sidered as potential candidates for the production of this
lipid that would result in the generation of the so-called
‘‘2nd generation’’ biodiesel [7, 9–16]. Finally, specifically
the oleaginous yeasts, with their unicellular form, are con-
sidered as the more appropriate organisms that have been
used as tools in the comprehension of numerous phenomena
of lipid biochemistry [2, 4, 17–20].
A very large variety of substrates has been used as carbon
sources for the oleaginous microorganisms such as analytical-
grade or industrially-derived (therefore, of low-cost) sugars
or sugar-enriched wastes or residues, polysaccharides,
N-acetylglucosamine, hydrolysates of various products or
by-products, vegetable oils, pure free fatty acids, FAMEs,
fatty byproducts or wastes, n-alkanes, ethanol, glycerol, and
organic acids [4, 7, 8, 10, 13–18, 21–30].
The microbial lipids are mainly composed of triacylgly-
cerols (TAGs) [1, 2, 27, 31, 32]. Other components pre-
sented in non-negligible quantities are free fatty acids
[25, 26, 31–34], other neutral lipids (such as monoacylgly-
cerols, diacylglycerols, and steryl-esters), sterols and polar
fractions (e.g. phospholipids, sphingolipids, glycolipids)
[5, 27, 31, 32, 35]. As a general remark it should be stressed
that when growth is carried out on various hydrophobic
substances utilized as substrates (the so-called ‘‘ex novo’’
lipid accumulation), the microbial lipid produced contains
lower quantities of TAGs compared with growth elaborated
on sugar-based substrates (the so-called ‘‘de novo’’ lipid
accumulation) [5, 25, 31, 32, 36–40]. Additionally, the oil
accumulated by the oleaginous fungi is more unsaturated
than that of the yeasts [2, 18–20, 28–30, 41]. This is the
main reason for which the oleaginous molds have been prin-
cipally used in order to produce lipids rich in polyunsaturated
fatty acids (PUFAs) of medical and dietetical interest like
g-linolenic acid (D6,9,12C18:3) [5, 6, 28–32, 42–49], dihomo-
g-linolenic acid, arachidonic acid, docosahexaenoic acid,
eicosapentanoic acid, etc (for reviews see: Ratledge [4];
Certik and Shimizu [50]; Ratledge and Wynn [2];
Sakuradani and Shimizu [51]; Sakuradani et al. [52]). In
contrast, few oleaginous yeast strains have been revealed
capable to synthesize such types of ‘‘rare’’ PUFAs
[2, 18, 20, 30]. Nevertheless, the last years, the utilization
of genetic engineering has resulted in the construction of
genetically modified yeast strains (principally belonging to
the species Yarrowia lipolytica), that are capable of (super)-
expressing various desaturases (e.g. D5-, D6-, D8-desaturate),
elongases and acyl-transferases, therefore, being, capable to
synthesize the above mentioned ‘‘rare’’ PUFAs and to trans-
fer these newly synthesized molecules into TAGs [53–56].
In the present review article, we will be interested in the
biochemical events related with the lipid accumulation in the
oleaginous yeasts when various carbon sources (hydrophilic
or hydrophobic ones) are utilized as substrates. Also the
events related with storage lipid degradation (turnover) in
the oleaginous microorganisms will be assessed and compre-
hensively discussed.
2 The biochemistry of lipid accumulation anddegradation in the oleaginousmicroorganisms
2.1 Lipid production from fermentation of sugars andrelated substrates used as carbon source
2.1.1 Substrates used
De novo accumulation of cellular lipids is an anabolic bio-
chemical process in which, by virtue of quasi-inverted b-
oxidation reaction series, acetyl-CoA issued by the intermedi-
ate cellular metabolism, generates cellular fatty acids. Fatty
acids are then esterified with glycerol generating structural
(phospholipids, sphingolipids, etc) and reserve (mainly
TAGs) lipids [2, 3, 7, 18–20, 23].
With the notable exception of cellulose and methanol a
very high number of carbon sources have been considered as
substrates for the de novo lipid biosynthesis from oleaginous
microorganisms. Concerning the conversion of cellulose into
SCO, few papers have recently appeared in the international
literature [57, 58]. Sugar-based media such as simple sugars
(e.g. glucose and fructose), lactose, sucrose, whey, glucose-
enriched wastes, molasses, etc [5, 6, 10, 16, 31, 32, 46, 47,
59–68] have been used. Xylose-based media have been
recently considered as substrates of noticeable importance
due to the abundance of xylose as substrate, deriving after
chemical hydrolysis of various lignocellulosic materials [11,
28, 69, 70]. In addition, the utilization of sugar-based sub-
strates more complicated compared with glucose, such as
polysaccharides (e.g. starch and pectin), has been studied.
Although the above substrates are similarly metabolized, in
some cases the results that have been achieved, in terms of
both lipid and fatty acid composition of the SCO produced,
presented notable differences [45, 48, 71].
The stoichiometry of glucose (and similar sugars such
as lactose, fructose, etc) metabolism indicates that about
1.1 moles of acetyl-CoA are generated from 100 g of glucose
(�0.56 moles) catabolized [18, 28, 30]. As far as xylose is
concerned, this compound can be either metabolized through
the phosphoketolase reaction, which is the most efficient
pathway yielding around 1.2 moles of acetyl-CoA per
100 g of xylose (�0.66 moles) utilized, or the pentose phos-
phate pathway, where around 1.0 mole of acetyl-CoA is
formed per 100 g of xylose utilized [18, 28]. Therefore, if
all the acetyl-CoA produced is channeled towards lipid syn-
thesis, the maximum theoretical yield of SCO produced per
glucose consumed is around 0.32 g/g [18]. This value is
somewhat higher concerning the fermentation of xylose
(around 0.34 g/g), assuming that oleaginous microorganisms
1032 S. Papanikolaou and G. Aggelis Eur. J. Lipid Sci. Technol. 2011, 113, 1031–1051
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

utilize exclusively the phosphoketolase pathway for xylose
assimilation. With reference to glycerol, the maximum theor-
etical yield of SCO is around 0.30 g/g [18]. However, even
under ideal conditions for SCO production (e.g. highly aer-
ated chemostat cultures) lipid yield on glucose consumed can
rarely be higher than 0.22 g/g [1, 2, 7], whereas in other
reports this threshold is considered to be in the level of
0.20 g/g or even lower [28, 46, 47]. However, in
Thamnidium elegans grown on sucrose in shake flasks, the
conversion yield of lipid produced per sugar consumed was
�0.24 g/g, while utilization of other sugars (glucose or fruc-
tose) equally resulted in exceptional conversion yields, i.e.
>0.20 g/g [65]. Maximum conversion yields of the same
magnitude compared with growth of T. elegans on sucrose
(�0.23 g/g) have been reported forCunninghamella echinulata
cultivated on xylose in shake-flask experiments [28]. The
conversion threshold of glycerol into SCO is around
0.10 � 0.02 g/g, in general lower than that obtained on glu-
cose, xylose or other sugars, regardless of the oleaginous
strain used [28, 29, 49, 72–78], presumably due to the
relatively poor regulation of the enzymes involved in the
primary metabolic steps of glycerol assimilation (e.g. glycerol
kinase, 3-P-glycerol dehydrogenase) [7, 76]. In contrast, the
respective value is slightly higher (e.g.�0.15 g/g) for the case
of algae like Schizochytrium limacinum growing heterotroph-
ically on glycerol [79–81], while only in two cases so far
conversion of glycerol into SCO has been performed with
a conversion yield higher than 0.20 g/g; it was either the case
of Aspergillus sp. strains that stored remarkable amounts of
intra-cellular lipids and also produced quantities of extra-
cellular oxalic acid cultured in shake flasks [82], or the case of
the fungus T. elegans that produced indeed high SCO
quantities (from 5.9 to 11.6 g/L corresponding to lipid in
dry weight ranging between 60 and 70% w/w), with conver-
sion yield ranging between 0.16 and 0.22 g/g, equally in
shake-flask experiments [83].
Ethanol has been considered as a potential substrate for
the de novo lipid biosynthesis of the oleaginous microorgan-
isms by a number of investigators [18, 59, 84–86], given that
it is considered as a very proper one since no residual carbon
arises from its uses in fermentation processes [18]. Taking
into consideration that ethanol is by far the more reduced
substrate by any other considered yet for the process of de
novo lipid accumulation, the final stoichiometric balance for
SCO synthesis from ethanol could result in a theoretical yield
of 0.54 g of lipid per 1 g of ethanol consumed [18].
Nevertheless such high conversion yields have never been
achieved in the literature with the conversion threshold of
ethanol into SCO being around 0.31 g/g [18, 29, 84].
Coming back to glycerol, even though this substrate
presents a slightly lower theoretical conversion yield com-
pared with glucose, this carbon source is of substantial and
increasing importance due to its appearance into the market
volume in continuously growing quantities due to application
into an industrial scale of the bio-diesel production process.
In general, the utilization of glycerol as a microbial substrate,
refers mainly to the production of 1,3-propanediol by bac-
teria (for review see: Papanikolaou [87]), whilst to the best of
our knowledge, only some investigations have been con-
ducted so far with (bio-diesel derived waste) glycerol utilized
as substrate (or co-substrate) by microorganisms in order to
produce SCO [26, 28, 29, 49, 72–83]. In the above-cited
papers trials were performed with Cryptococcus curvatus, S.
limacinum,Y. lipolytica,Mortierella isabellina,C. echinulata and
T. elegans strains. Moreover, in a recent investigation,
Chatzifragkou et al. [83] have performed a relatively exten-
sive screening of yeast and Zygomycetes strains (species
Candida boidinii, C. curvata, C. oleophila, C. pulcherrima, C.
echinulata, M. isabellina, M. ramanniana, Mucor sp., Pichia
membranifaciens, Rhodotorula sp., T. elegans, Y. lipolytica,
Zygosaccharomyces rouxii, Zygorhynchus moelleri) cultivated
on bio-diesel derived glycerol in conditions favoring the pro-
duction of SCO (nitrogen-limited conditions) and they have
found that although yeast strains in general produced higher
quantities of biomass compared with the molds, the pro-
duction of lipid was increased in the fungal strains. The more
promising SCO producer amongst all strains tested was the
Zygomycete T. elegans [83].
Citric acid [88], acetic acid [89–91], or other low-mol-
ecular weight organic acids [90, 92, 93] have been equally
considered as substrates for SCO production. Specially, as far
as acetic acid is concerned, it is considered as a remarkable
pollutant, generated either in the process water of Uranium
bleaching or as effluent issued from the Fischer–Tropsch
reaction [90, 93]; the investigations, thus, concerning its
biotransformation in SCO are very interesting in both econ-
omical and ecological terms.
2.1.2 Biochemistry of de novo lipid accumulation
All microorganisms are capable to synthesize lipids, though
only the oleaginous strains may accumulate inside their cells
significant lipid quantities (i.e. >20% w/w, on dry cell basis).
In a series of investigations, it has been demonstrated that the
oleaginous microorganisms do not possess a hyperactive sys-
tem of fatty acid biosynthesis, but they are capable of pro-
ducing in significant quantities, acetyl-CoA, the basic unit of
fatty acid biosynthesis [3, 20]. The biochemistry, thus, of de
novo lipid biosynthesis, may be divided in two distinct par-
ties: the intermediate cellular metabolism and the biosyn-
thesis of TAGs.
The net product of glycolysis is pyruvic acid, which passes
through the mitochondrial membrane to the mitochondrion
matrix. Pyruvate-dehydrogenase catalyzes the formation of
acetyl-CoA from pyruvic acid, and acetyl-CoA either enters
inside the Krebs cycle, or is transported again into the cyto-
plasm in order to enhance biosynthesis of cellular fatty acids
[2, 3, 18, 20]. Since the mitochondrial membrane is not
permeable by acetyl-CoA, the transformation of this com-
pound to acetyl-carnitine (catalyzed by carnitine-acyl-trans-
Eur. J. Lipid Sci. Technol. 2011, 113, 1031–1051 The biochemistry of oleaginous yeasts 1033
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

ferase) should be necessary for the transport of this unit into
the cytosol. Though, the capital role carnitine-acyl-transfer-
ase is exactly the opposite, namely the transport of the acetyl-
CoA, issued by b-oxidation, inside the mitochondrion
matrix. Minimal amounts of acetylcarnitine may pass
through the mitochondrion matrix in order to enter the
cytosol, and this is the case of de novo lipid biosynthesis of
the non-oleaginous microorganisms [2, 4, 20]. In the olea-
ginous microorganisms, acetyl-CoA that constitutes the pre-
cursor of intracellular biosynthesis of fatty acids derives from
breakdown of citric acid that under some circumstances has
been previously accumulated inside the mitochondria and
then is transported into the cytosol (for reviews see: Ratledge
[18, 20]; Ratledge and Wynn [2]; Fakas et al. [30]).
The key step for lipid accumulation in the oleaginous
microorganisms is the change of intracellular concentration
of various metabolites, conducted after exhaustion of some
nutrients into the culture medium [2, 7, 18, 20]. In most of
the performed studies, the essential nutrient the depletion of
which induced the accumulation of reserve lipid is that of
nitrogen, whereas the biochemistry of de novo lipid accumu-
lation of lipid has been completely elucidated only when
extracellular nitrogen in the limiting factor of microbial
growth [1–4, 7, 8, 30]. Nitrogen exhaustion provokes a rapid
decrease of the concentration of intracellular AMP (adeno-
sine monophosphate), since, by virtue of AMP-desaminase,
the microorganism cleaves AMP in IMP (inosine monophos-
phate) and NH4þ ions. The NH4
þ ions constitute a comp-
lementary nitrogen source, necessary for synthesis of cell
material after the extracellular nitrogen limitation [94].
The excessive decrease of intra-cellular AMP concentration
alters the Krebs cycle function; NADþ- (and in various cases
also NADPþ-) isocitrate dehydrogenase, enzyme responsible
for the transformation of isocitric to a-ketoglutaric acid,
losses its activity, since it is allosterically activated by intra-
cellular AMP [47, 95–97]. Thus, iso-citric acid is accumu-
lated inside the mitochondrion. This acid is found in
equilibrium with citrate (reaction catalyzed by isocitrate
acotinase). When the intra-mitochondrial citric acid concen-
tration reaches a critical value, citrate enters the cytoplasm in
exchange with malate [98]. Finally, citric acid is cleaved by
the ATP-citrate lyase (ATP-CL), the enzyme-key of lipid
accumulation process in the oil-bearing microorganisms, in
acetyl-CoA and oxaloacetate [94, 96, 99] (for reviews see:
Ratledge [18, 20]; Ratledge andWynn [2]). Acetyl-CoA, by a
quasi-inverted b-oxidation process, will generate the cellular
fatty acids (for reviews see: Ratledge and Wynn [2];
Papanikolaou and Aggelis [7]; Fakas et al. [30]). NADPH,
indispensable fatty acid biosynthesis, is provided by the inter-
mediate cellular metabolism, in which the importance of
malic enzyme has been considered as crucial for various
oil-bearing microorganisms, specifically in the cases in which
NADPH is produced exclusively by virtue of the reaction
catalyzed by the above mentioned enzyme under nitrogen-
limited conditions [2, 97]. Schematically, the intermediate
cellular metabolism of the oleaginous microorganisms in
which lipid accumulation is performed after nitrogen exhaus-
tion from the medium, is presented in Fig. 1 (adapted by
Davies and Holdsworth [3]).
ATP-CL is an enzymatic complex that is considered to be
the most important factor to account for the oleaginicity of
the various microorganisms, being absent in the non-oleagi-
nous microbial cells [18]. The reaction catalyzed through the
use of ATP-CL is depicted as follows [18, 20]:
Net product of the action of ATP-CL, therefore, is acetyl-
CoA, that will be further converted into intra-cellular fatty
acids. If ATP-CL enzymatic complex does not exist, nitrogen
exhaustion leads in the accumulation of citric acid inside the
cytoplasm. In this case, citric acid either will be excreted into
the culture medium (case of citric acid production by
Aspergillus niger and Candida sp. strains—see: Ratledge
[20]; Papanikolaou and Aggelis [7]) or will provoke the
inhibition of the 6-phosphoro-fructokinase, having as result
intracellular accumulation of polysaccharides based on
Nitrogen limitationNitrogen limitation
Function of AMPFunction of AMP--desaminasedesaminase
Degradation of intraDegradation of intra--cellular AMPcellular AMP
Deactivation of NADDeactivation of NAD-- (and potentially NADP(and potentially NADP--) dependent) dependentdehydrogenasedehydrogenase
Citric acid accumulation inside the mitochondrionCitric acid accumulation inside the mitochondrion
Citric acid transport outside the Citric acid transport outside the mitochondrion, in exchange withmitochondrion, in exchange with malatemalate
Citric acid cleavage, formation of acetylCitric acid cleavage, formation of acetyl--CoACoA,,reaction catalyzed by ATPreaction catalyzed by ATP--citrate citrate lyaselyase
Condensation reactions of acetylCondensation reactions of acetyl--CoACoA for for acylacyl--CoACoA biosynthesis biosynthesis ––Onset of fatty acids accumulationOnset of fatty acids accumulation
Figure 1. Consecutive steps leading to the de novo lipid biosynth-
esis in oleaginous microorganisms growing under nitrogen-limited
conditions (from Davies and Holdsworth [3], adapted).
1034 S. Papanikolaou and G. Aggelis Eur. J. Lipid Sci. Technol. 2011, 113, 1031–1051
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

6-phosporo-glucose (case of Aureobasidium pullulans—see:
Galiotou-Panayotou et al. [100]). Due to the significant
biochemical similarity between the intracellular de novo lipid
accumulation and the extracellular secretion and production
of citric acid, the last years, the yeasts have been divided and
classified by various authors as either lipid-accumulating or
citric acid-producing ones [49,101–103]. The pattern of inter-
mediate metabolism and de novo lipid biosynthesis is pre-
sented in Fig. 2.
The first reaction of fatty acid biosynthesis after acetyl-
CoA generation, is catalyzed by a biotin-dependent acetyl-
CoA carboxylase [18]. Its action is described as follows:
Acetyl-CoAþHCO3 þ ATP ! Malonyl-CoAþ Pi
This reaction is considered as the restricting step for fatty acid
biosynthesis. It is activated by the presence of citric acid in the
oleaginous strain Candida sp. 107, but not in the non-ole-
aginous strain Candida utilis [104]. In fact, in the absence of
citrate, the abovementioned enzyme exists asmultifunctional
inactive protein with a molecular size of 2.4 � 105 daltons,
while the presence of citrate induces enzyme monomers
aggregation into an active macro-structural protein with a
size of 7–9 � 106 daltons [18]. Following the generation of
malonyl-CoA, the biosynthesis of fatty acids is performed
with the aid of the multi-enzymatic complex of fatty acid
synthetase (FAS). The effectuated reaction series can be
summarized as follows [3, 30]:
Acetyl-CoAþ 7malonyl-CoAþ 14NADPH
! Palmitoyl-CoAþ 7CO2 þ 14NADPþ 7 CoASH
þ 6H2O
Generally, the multi-enzymatic complexes of FAS and
ATP-CL are inhibited by the presence of exogenous long
aliphatic chains (e.g. fatty acids, n-alkanes, etc) found into the
culture medium [34, 105]. Whatever the origin of the intra-
cellular aliphatic chains, after the biosynthesis of fatty-CoA
esters, an esterification with glycerol takes place in order for
the reserve lipids to be stocked in the form of TAGs [18, 20].
This synthesis is conducted mainly by the so-called pathway
of a-glycerol phosphate acylation (for reviews see: Ratledge
[18]; Davies and Holdsworth [3]; Athenstaedt and Daum
[106]; Mullner and Daum [107]; Fakas et al. [30]). In this
metabolic pathway, free-fatty acids are activated to coenzyme
A and are subsequently used for the acylation of the glycerol
backbone to synthesize TAGs. In the first step of TAGs
assembly, glycerol-3-phosphate (G-3-P) is acylated by G-
3-P acyltranferase (GAT) at the sn-1 position to yield 1-
acyl-G-3-P (lysophospatidic acid-LPA), which is then further
acylated by lysophosphatidic acid acyltransferase (also named
1-acyl-G-3-P acyltransferase-AGAT) in the sn-2 position to
yield phosphatidic acid (PA). This is followed by dephos-
phorylation of PA by phosphatidic acid phosphohydrolase
(PAP) to release diacylglycerol (DAG). In the final step DAG
is acylated either by diacylglycerol acyltransferase or phos-
pholipid diacylglycerol acyltransferase to produce TAGs (for
reviews see: Ratledge [18]; Davies and Holdsworth [3];
Athenstaedt and Daum [106]; Mullner and Daum [107];
Fakas et al. [30]). The steps of TAG assembly through the
a-glycerol phosphate acylation pathway, a pathway that is
very commonly used in the oleaginous microorganisms, are
depicted in Fig. 3. Alternatively, PA can be synthesized
through the dihydroxyacetone-phosphate (DHAP) pathway
as follows (studies performed principally on the non-
oleaginous yeast Saccharomyces cerevisiae—for reviews see:
Athenstaedt and Daum [106]; Mullner and Daum [107]):
DHAP is acylated at the sn-1 position by the enzyme DHAP
acyltransferase (DHAPAT). The product, 1-acyl-DHAP, is
reduced by 1-acyl-DHAP reductase (ADR) to yield 1-acyl-
G-3-P, which is further acylated to yield PA, reaction cata-
lyzed by AGAT. Finally, as far as the structure of the TAGs
produced is concerned, although their final composition
could theoretically be a random substitution of acyl-CoA
groups into glycerol, in the case of the oleaginous microor-
ganisms that have been examined, the glycerol sn-2 position
in most of the cases is occupied by unsaturated fatty acids
(therefore, vegetable-type TAGs are produced—see: Thorpe
and Ratledge [36]; Ratledge [18, 20]; Guo and Ota [108]).
Glycerol
3-P-Glycerol
Pyruvate abc
Pyruvate CH3COSCoA
Oxaloacetate
MalateMDm
dMalate
PD
CSCitrate
Iso-citrate Ac
a-ketoglutarate ICDH
Krebs
ICL
Citrate
Secretion
ACL
CH3COSCoA
Malonyl-SCoA
FAS
TAGs
Oxaloacetate
MDc
MENADPH-CO2
NADPH
NADP
GK
3-P-Dihydroxyacetone
ATPADP
Glycolysis
Hexose, pentose Glyconeogenesis
DHAkGlucose
Biomass
ATP ADP
NAD NADH
Biomass
ATP
ADP
NAD
NADH
CYTOSOL
MITOCHONDION
Figure 2. Intermediate metabolism in the oleaginous microorgan-
isms. (a–c) Systems of pyruvate transport from cytoplasm to mito-
chondrion and inversely for the malate. (d) System of citrate and
malate transport between cytoplasm and mitochondrion. Enzymes:
Ac, acotinase; ACC, acetyl-CoA carboxylase; ACL, ATP-citrate
lyase; FAS, fatty acid synthetase; ICDH, iso-citrate dehydrogenase;
MDc, malate dehydrogenase (cytoplasmic); MDm, malate dehydro-
genase (mitochondrial); PD, pyruvate dehydrogenase; PFK, phos-
pho-fructo-kinase; PK, pyruvate kinase (from Ratledge [18];
Papanikolaou and Aggelis [7], adapted).
Eur. J. Lipid Sci. Technol. 2011, 113, 1031–1051 The biochemistry of oleaginous yeasts 1035
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

In batch experiments, lipid accumulation involving de
novo biosynthesis pathway in the oleaginous microorganisms
is essentially a secondary anabolic activity, since, according to
the biochemical steps previously illustrated, it is enhanced
after the nitrogen depletion of the culture medium due to
‘‘metabolic overflow’’ at carbon excess conditions (for
reviews see: Ratledge [4]; Ratledge and Wynn [2]; Dyal
and Narine [41]; Fakas et al. [30]; Papanikolaou and
Aggelis [8]). However, it should be stressed in this point that
the limitation of nitrogen is not the sole factor of crucial
importance for accumulation of storage lipid during growth
of oleaginous microorganisms on sugars or similarly metab-
olized compounds. For instance, in recent investigations
it has been demonstrated that the oleaginous yeast
Rhodosporidium toruloides can present efficient production
of biomass and accumulation of storage lipid in nitrogen-rich
media, provided that phosphorus or sulphate is the limiting
factor of cell growth [14, 109]. Interestingly enough, the
cultivation on sulphate- or phosphate-limited nitrogen-rich
media induced remarkable changes in the total fatty acid
composition of storage lipids produced, with (noticeable)
increment of the cellular saturated fatty acid content of R.
toruloides when sulphate-limited conditions were imposed
[109]. In the later case, thus, with a simple fermentation
technique and based on this property of R. toruloides, a
cocoa-butter substitute was produced (for more details see
part II [155] of the current review-article). In any case, the
property of microorganisms to store significant quantities of
SCO in nitrogen-rich and phosphate- or sulphate-limited
media, can be exploited in industrial level for the valorization
of sugar-rich andnitrogen-rich agro-industrial residues [14, 109].
In general, growth of oleaginous microorganisms on sugar (or
similarly metabolized compounds) based nitrogen- (or sul-
phate- or phosphate-) limited media can be divided into two
distinct phases: (a) phase in which all nutrients are available
for themicrobial growth (the balanced growth phase). During
this phase, cell growth (including total biomass—X and lipid-
free biomass—Xf production) is carried out, and substrate
(sugar) is consumed with relatively high assimilation rates. In
this phase, cellular lipids (L) that include quantities of TAGs
are produced but, in general, they contain polar fractions (e.g.
sphingolipids, phospholipids, etc.) corresponding to mem-
brane lipids [5, 31]. In this phase, total lipid in dry matter
corresponds to a value of 5–10% w/w; (b) lipid accumulation
phase. In this phase, neutral lipids (mostly TAGs) are accu-
mulated inside the microbial cells or fungal mycelia, while
substrate (sugar) uptake could potentially present somehow
lower uptake rate compared with the balanced growth phase.
It should also be stressed that in several cases (e.g. exper-
iments performed with the oleaginousmoldsM. isabellina and
T. elegans or also with other oleaginous microorganisms),
during nitrogen-limiting phase besides lipid, also lipid-free
biomass is synthesized. This lipid-free biomass increment
despite nitrogen-limited conditions into the medium
indicates partial cell growth (and cell proliferation) and prin-
cipally accumulation of non-lipid storage materials like intra-
cellular polysaccharides [28, 29, 46]. A pattern of lipid
accumulation kinetics in an oleaginous microorganism
during growth on sugar-based nitrogen-limited media is pre-
sented in Fig. 4 (data from Papanikolaou et al. [65]). It is
easily understood that the onset of lipid accumulation is given
after exhaustion of assimilable nitrogen into the culture
medium. Furthermore, it can be seen that after complete
exhaustion of sugar from the fermentation medium, some
quantities of previously stored lipid are degraded (lipid turn-
over) in favor of lipid-free material formation.
In general, de novo synthesized yeast lipid is composed
of C16 and C18 fatty acids. Palmitic acid (C16:0) constitutes
the 15–25% w/w, of total lipids, whilst palmitoleic (D9C16:1)
is, in general, presented in percentages inferior than 5% w/w
[4, 7, 20, 24]. Likewise, stearic acid (C18:0) is generally a
minor component of the yeast lipid (5–8% w/w). Oleic acid
(D9C18:1) is the principal fatty acid accumulated inside the
yeast cells (amounts sometimes higher than 70% w/w),
while linoleic (D9,12C18:2) is found in the second position
RCOSCoA +3-GLYCEROL-PHOSPHATE
CoA
Glycerol-3-phosphate acyl transferase
LYSOPHOSPHATIDIC ACID
RCOSCoA
CoA
Lysophosphatidic acidacyl transferase
PHOSPHATIDIC ACID
Pi
H2O
Phosphatidic acid phosphohydrolase
DIACYLGLYCEROL
RCOSCoA
CoA
Diacylglycerol acyltransferase or Phospholipid diacylglycerol acyltransferase
TRIACYLGLYCEROL
Figure 3. Formation of intracellular triacylglycerols via the pathway
of a-glycerol phosphate acylation (from Ratledge [18, 4], adapted).
1036 S. Papanikolaou and G. Aggelis Eur. J. Lipid Sci. Technol. 2011, 113, 1031–1051
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

(15–25% w/w) [4, 20]. More unsaturated fatty acids (e.g. a-
linolenic acid—D9,12,15C18:3) are not frequently synthesized
into the yeast lipid reserves [22]. It may be assumed, there-
fore, the yeast lipid produced is, in general, composed of
unsaturated fatty acids. Only in rare cases (e.g. case of
Lipomyces starkeyi DSM 70295 growing on glucose-enriched
sewage sludge—see: Angerbauer et al. [10]) the quantity of
saturated fatty acids accumulated by the oleaginous yeasts
(principally C16:0 and to lesser extent C18:0) is higher than
50% w/w, of total lipids. In fact, the above event (the poten-
tiality of producing through de novo fatty acid accumulation
process globally saturated microbial lipids) is considered to
be the limiting step for the synthesis of microbial analogous of
expensive exotic fats (e.g. cocoa-butter) [3, 8, 20, 110].
Various strategies have been performed in order to alleviate
the above disadvantage. The fatty acid profiles of several
oleaginous yeasts when de novo lipid accumulation is per-
formed are depicted in Table 1.
2.2 Lipid production from fermentation ofhydrophobic substrates used as carbon source
2.2.1 Substrates used
A somehow restricted number of yeast strains have been
recorded to be capable of growing on fats and at the same
time accumulate significant lipid quantities. These yeasts
belong to the genera Torulopsis (T. versatilis, Torulopsis sp.),
Candida [C. tropicalis, C. guilliermondii, Yarrowia (C.) lipoly-
tica], Trichosporon, Geortichum and to the species Pichia meth-
anolica, Apiotrichum curvatun and Rhodosporidium toruloides
[21, 25, 33, 37, 38, 40, 117–128]. It is evident that the
number ofmicroorganisms that are capable to consume soaps
and free-fatty acids is higher, since culture on these materials
is done regardless of the lipolytic capacity of the microorgan-
ism used. In contrast, the microorganisms that are able to
proceed with TAG or fatty-esters break-down, should obli-
gatory possess an active lipase system into their enzymatic
arsenal [8, 17, 122, 129, 130]. As far as the yeast Y. lipolytica
is concerned, in various reports in the past years it was
considered as a non-oleaginous microorganism, since it
had been assumed as ineffectual of accumulating significant
lipid quantities from sugars or similarly metabolized com-
pounds during submerged growth in nitrogen-limited media
[20]. However, the capacity of at least some Y. lipolytica
strains to accumulate high lipid quantities (up to 60% w/
w, in dry weight) when various fats or oils were used as sole
carbon and energy source is out of question indicating the
oleaginicity of this microorganism [25, 33, 37, 40, 122, 124,
125, 128, 131] (for reviews see: Beopoulos et al. [27];
Papanikolaou and Aggelis [8]; Sabirova et al. [132]). It
should also be noticed that in at least one case a strain of
Y. lipolytica produced significant intra-cellular fat quantities
(23–43%w/w, lipid in dry weight) when cultivated on glucose
or glycerol (de novo fatty acid accumulation) in highly aer-
ated bioreactor experiments [64, 74, 76], whereas on the
other hand, small production of lipids occurred in shake-flask
experiments, in which the nitrogen limitation imposed led to
remarkable production of extra-cellular citric acid [49, 133].
The fatty materials utilized as substrate from the oleagi-
nous strains may be vegetable oils [33, 37, 119, 122, 129,
130, 134], fatty esters (methyl-, ethyl-, butyl-, or vinyl-esters
of fatty acids) [21], soap-stocks [120], pure free-fatty acids
[117, 118, 126, 127], industrial fats composed of free-fatty
acids of animal or vegetable origin [25, 40, 124, 125, 133]
and crude fish oils [38, 39, 108]. In the case of the growth of
0
2
4
6
8
10
12
0
5
10
15
20
25
30
0 50 100 150 200 250 300
X (g/L)
L (g/L)
Fru (g/L)
Bio
mas
s (X
), Li
pids
(L) [
g/L]
Fru
ctos
e (F
ru) [
g/L]
Time [h]
0
20
40
60
80
100
120
140
0
10
20
30
40
50
60
70
80
0 50 100 150 200 250 300
Ammonium ions (mg/L)
Lipid in dry weight (%, w/w)
Am
mon
ium
ion
s (m
g/L)
Lipi
d in
dry
wei
ght
(%, w
/w)
Time [h]
(a)
(b)
Figure 4. Kinetics of the oleaginous fungus Thamnidium elegans
growing on fructose in shake-flask nitrogen-limited experiments
(initial fructose at 30 g/L). Representation of biomass production
(X, g/L), lipid production (L, g/L), fructose consumption (Fru, g/L)
(a), ammonium ions assimilation and lipid in dry weight evolution (b)
(data from Papanikolaou et al. [65]).
Eur. J. Lipid Sci. Technol. 2011, 113, 1031–1051 The biochemistry of oleaginous yeasts 1037
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

various microorganisms on media in which hydrophobic
materials are found as substrate (or co-substrate), the
quantity of TAGs accumulated into the total accumulated
lipids, can be substantially lower compared with the lipids
produced through the de novo lipid accumulation pathway;
for instance, in the case of strains of the molds Amylomyces
rouxii and Cunninghamella blakesleeana and the yeast C. lip-
olytica cultured on soybean oil utilized as the sole substrate,
intra-cellular lipid produced contained significant free-fatty
acids quantities ranging between 30 and 83% w/w, of total
intra-lipid produced, while TAGs represented in various
cases a marginal compound of SCO produced [37].
Likewise, in the case of the oleaginous yeast Candida sp.
107, fractionation of the lipids produced from this microor-
ganism grown on various n-alkanes showed that, although the
TAGs still constituted the major SCO fraction, their relative
proportion was less than when glucose was utilized as the sole
growth substrate, while loss in TAGs was compensated for by
corresponding increase in the quantity of phospholipids [36].
Yeast lipid containing TAGs but also considerable quantities
of free-fatty acids has been reported during growth of C.
lipolytica 1094 on corn oil utilized as the sole substrate
[33]. Likewise, Aoki et al. [123] have studied the effect of
the addition of various hydrophobic materials (e.g. TAGs,
fatty acid ethyl-esters or free-fatty acids) used as glucose co-
substrates upon the biochemical behavior of the yeast P.
methanolica, and it was revealed that the intra-cellular lipid
compounds were almost of the same chemical structure with
the fatty material that was added into the medium. It was also
interesting to state that in the case of the addition of free-fatty
acids, fat accumulation ten-fold increased compared with the
addition of methyl-esters or TAGs (lipid accumulation
around 20% w/w, in dry matter against 2.5–2.8% w/w,
respectively), while in the case of added free-fatty acids as
co-substrate, SCO produced contained around 93% w/w,
free-fatty acids into the total microbial lipids stored [123].
Growth of Geotrichum sp. and C. guilliermondii on crude fish
oils resulted in the accumulation of TAGs in quantities 30–
60% w/w, into the total SCO produced (substantially lower
TAG quantities compared with de novo lipid accumulation
from sugars—see i.e.: Ratledge [18, 20]), while polar lipids or
non-identified fatty compounds were also found in noticeable
concentrations [38, 108]. Finally, growth of Y. lipolytica
ACA-DC 50109 on industrial fats composed of free-fatty
Table 1. Fatty acid composition of lipid produced by various yeast strains growing on sugars (or similarly-metabolized like glycerol,
molasses, etc) substrates in culture conditions favoring the accumulation of microbial lipid
Strain Lipid (% w/w) C16:0 C16:1 C18:0 C18:1 C18:2 C18:3 Reference
Candida sp. 107a) 37.1 37 1 14 36 7 T. Gill et al. [104]
Candida sp. 107 n.r. 28 n.r. 8 41 17 17 Davies [22]
Candida sp. 40.3 23 13 3 54 5 2 Aggelis et al. [24]
Rhodotorula gracilis 41.0 21 T. 13 51 11 3 Choi et al. [111]
Candida curvatab) 29.1 36 T. 14 40 7 T. Evans and Ratledge [59]
Candida curvatab) 28.0 37 T. 10 44 6 T. Evans and Ratledge [59]
Apiotrichum curvarumb) 31.0 34 T. 10 43 7 2 Hassan et al. [113]
Cryptococcus curvatusb) 38.0 24 T. 10 46 9 6 Hassan et al. [114]
Cryptococcus curvatusb) 25.0 18 T. 16 50 16 T. Meesters et al. [72]
Cryptococcus curvatusb) 50.0 31 - 22 42 1 n.r. Wu et al. [15]
Cruptococcus albidus 46.3 14 T. 9 53 18 2 Hansson and Dostalek [112]
Cruptococcus albidus n.r 20 n.r 11 59 6 6 Davies [22]
Yarrowia lipolytica 43.2 15 2 11 47 21 3 Papanikolaou and Aggelis [74]
Yarrowia lipolytica 30.7 12 11 9 57 11 T. Andre et al. [75]
Yarrowia lipolytica a) 22.3 13 17 6 55 7 n.r. Makri et al. [76]
Rhodosporidium toruloides 67.5 20 1 15 47 13 3 Li et al. [9]
Rhodosporidium toruloides 65.2 34 T. 13 48 1 T. Hu et al. [115]
Rhodosporidium toruloides 62.1 26 2 5 62 3 T. Wu et al. [14]
Rhodosporidium toruloides 55.6 43 T. 16 35 2 T. Wu et al. [109]
Lipomyces starkeyi 68.0 56 2 14 26 T. T. Angerbauer et al. [10]
Lipomyces starkeyi 61.5 37 4 6 49 1 T. Zhao et al. [11]
Rhodotorula mucilaginosa 48.6 22 2 9 55 11 T. Zhao et al. [116]
Trichosporon capitatum 37.6 12 1 2 74 9 n.r. Wu et al. [16]
Rhodotorula sp. 22.0 22 1 7 56 12 n.r. Chatzifragkou et al. [83]
Candida oleophila 15.3 13 3 7 66 11 n.r. Chatzifragkou et al. [83]
T. <0.5% w/w; n.r.: not reported.a) Representation of the neutral fraction of microbial lipids produced.b) Cryptococcus curvatus was formely Candida curvata and then Apiotrichum curvatum; thus these microorganisms in fact are the same species.
1038 S. Papanikolaou and G. Aggelis Eur. J. Lipid Sci. Technol. 2011, 113, 1031–1051
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

acids (a fully saturated derivative of tallow called ‘‘stearin’’, or
on mixtures of stearin with fully hydrolyzed oleic rapeseed
oil), resulted in SCO production in which TAGs were the
principal lipid fraction of the accumulated fat (45–55% w/w,
of total lipids), while free-fatty acids were produced in sig-
nificant amounts as well (about 30–40% w/w, of total lipids)
[25, 40]. Likewise, fundamental biochemical differences exist
between de novo lipid accumulation from nitrogen-limited
sugar-based substrates and ex novo lipid production from fats
or other hydrophobic compounds utilized as sole carbon and
energy source. These differences will be depicted in the
following chapter.
2.2.2 The biochemistry of ex novo lipid accumulation
The free-fatty acids (existed as initial substrate or produced
after lipase-catalyzed hydrolysis of the TAGs/fatty esters) are
incorporated, with the aid of active transport, inside the
microbial cell. The incorporated fatty acids are either dis-
similated for growth needs or become a substrate for endo-
cellular bio-transformations (synthesis of ‘‘new’’ fatty acid
profiles which did not exist previously in the substrate) [25,
38–40, 122, 125, 130]. The dissimilated free-fatty acids will
be degraded, by virtue of the process of b-oxidation, into
smaller chain acyl-CoAs and acetyl-CoA, reactions catalyzed
by the various acyl-CoA oxidases (Aox) (for reviews see:
Fickers et al. [135]; Beopoulos et al. [27]), providing, thus,
firstly the necessary energy for cell growth and maintenance
(channel of acetyl-CoA inside the Krebs cycle), and secondly
the formation of organic substances (intermediate metab-
olites) which constitute the precursors for the synthesis of
cellular materials [4].
The degradation of hydrophobic materials has been
studied in enzymatic and molecular level in significant details
by strains of the non-conventional yeast Y. lipolytica [35, 118,
126, 127, 136–140]. This yeast when cultivated on TAG-
type substrates has been reported to secrete an extra-cellular
lipase called Lip2p, encoded by the LIP2 gene [137]. This
gene encoded for the biosynthesis of a precursor pre-mature
protein with Lys-Arg cleavage site. The secreted lipase was
reported to be a 301-amino-acid glycosylated polypeptide,
which belongs to the TAG hydrolase family (EC 3.1.1.3).
The Lip2p precursor protein was processed by the KEX2-like
endoprotease encoded by the geneXPR6, whereas deletion of
the above gene resulted in the secretion of an active but fewer
stable pro-enzyme [27, 135, 137]. Simultaneously other
intra-cellular lipases (Lip7p and Lip8p) may also be secreted
into the culture medium, that present different fatty acid
specificities, with maximum activity being displayed againstD9C18:1, C6:0 (capronic acid) and C10:0 (capric acid) fatty
acids (for reviews see: Fickers et al. [135]; Beopoulos et al.
[27]). Then, the released free-fatty acids, produced after
lipase-catalyzed hydrolysis of the TAGs would be incorpor-
ated inside the yeast cells. It is interesting to state that for the
case ofY. lipolytica yeast, the various individual substrate fatty
acids are incorporated inside the microbial cell with different
rates [25, 40, 125], before being subjected to degradation
performed by the various intra-cellular Aox. In fact, it has
been revealed that the aforementioned biochemical process is
a multi-step reaction requiring different enzymatic activities
of five acyl-CoA oxidase isozymes (Aox1p through Aox5p),
encoded by the POX1 through POX5 genes [118, 126, 127,
135, 136, 139, 140]. Aox3p is specific for short chain acyl-
CoAs, Aox2p preferentially oxidizes long-chain acyl-CoAs
while Aox1p, Aox4p and Aox5p do not appear of being
sensitive in the chain length of the aliphatic acyl-CoA chain
[27, 135, 138, 139]. Moreover, the physiological function of
the above-mentioned oxidases has been investigated by gene
disruption [136]; mutations in Aox4 and Aox5 resulted in an
increase in total Aox activity. The growth of mutant Y. lip-
olytica strains was analyzed and in the presence of POX1 gene
only, strains did not grow on fatty acids, whereas POX4 alone
elicited partial growth, while the growth of the double POX2-
POX3-deleted mutant was normal on media containing pureD9C18:1 as the sole carbon source [136, 139]. b-Oxidation
contributes one mole of NADH and one mole of FADH2 for
every 1 mole of acetyl-CoA generated, before the entering of
acetyl-CoA inside the Krebs cycle [4, 20], and is depicted in
Fig. 5.
Principal biochemical differences exist between de novo
and ex novo lipid biosynthesis; in the later case, lipid accumu-
lation occurs simultaneously with cell growth, being entirely
independent from nitrogen exhaustion from the culture
medium [25, 40, 122, 129–131, 134]. When fats or other
hydrophobic materials are utilized as the sole carbon and
energy source, accumulation of reserve lipid is a growth-
coupled process [25, 40, 129, 131] in which lipid is accu-
mulated simultaneously with lipid-free material formation in
the presence of assimilable nitrogen into the culture medium.
A pattern of lipid accumulation kinetics in an oleaginous
microorganism during growth on fat-based media (is pre-
sented in Fig. 6 (data from Papanikolaou et al. [131]).
Moreover, in order to further demonstrate the independence
RCH2- CH 2 - CH 2- CO - SCoA RCH2- CH = CH- CO - SCoA
FAD FADH
RCH2- CHOH - CH2- CO - SCoA
+ H2O
NAD
NADH
RCH2- CO - CH2- CO - SCoACH3- CO - SCoA
RCH2- COSCoA
New cycle of reactions
+
+ CoA-SH
Figure 5. b-Oxidation process of a fatty acid with even number of
carbon atoms (from Ratledge [4], adapted).
Eur. J. Lipid Sci. Technol. 2011, 113, 1031–1051 The biochemistry of oleaginous yeasts 1039
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com
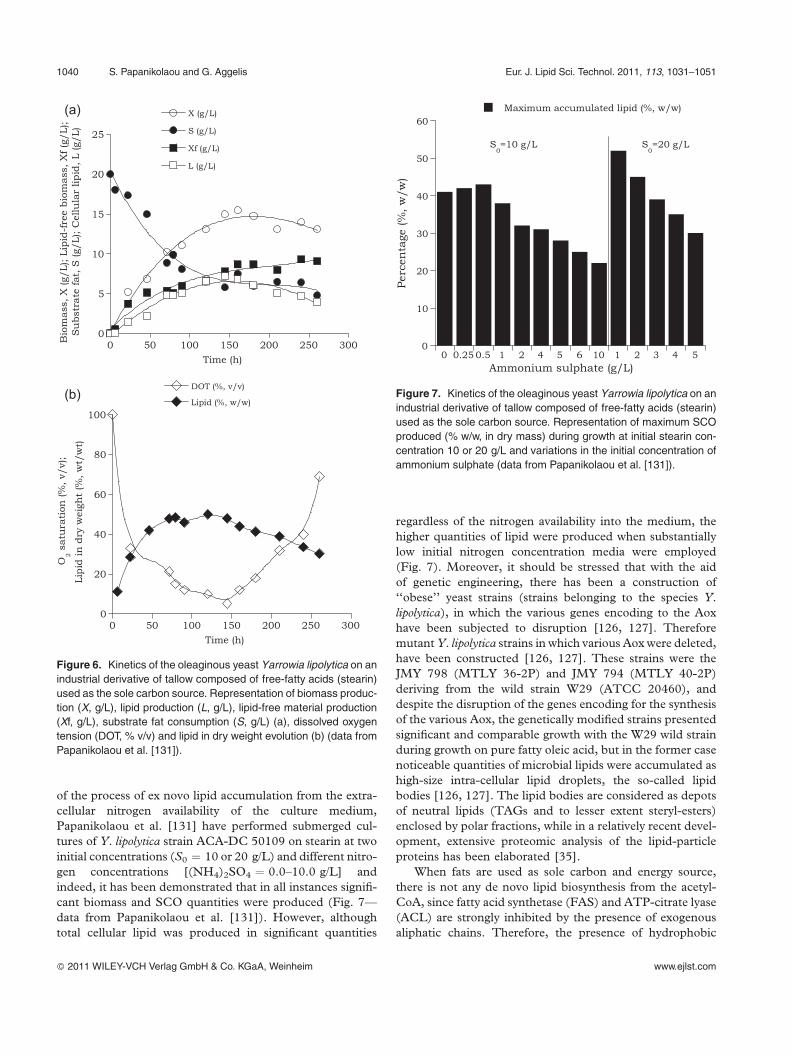
of the process of ex novo lipid accumulation from the extra-
cellular nitrogen availability of the culture medium,
Papanikolaou et al. [131] have performed submerged cul-
tures of Y. lipolytica strain ACA-DC 50109 on stearin at two
initial concentrations (S0 ¼ 10 or 20 g/L) and different nitro-
gen concentrations [(NH4)2SO4 ¼ 0.0–10.0 g/L] and
indeed, it has been demonstrated that in all instances signifi-
cant biomass and SCO quantities were produced (Fig. 7—
data from Papanikolaou et al. [131]). However, although
total cellular lipid was produced in significant quantities
regardless of the nitrogen availability into the medium, the
higher quantities of lipid were produced when substantially
low initial nitrogen concentration media were employed
(Fig. 7). Moreover, it should be stressed that with the aid
of genetic engineering, there has been a construction of
‘‘obese’’ yeast strains (strains belonging to the species Y.
lipolytica), in which the various genes encoding to the Aox
have been subjected to disruption [126, 127]. Therefore
mutantY. lipolytica strains in which various Aox were deleted,
have been constructed [126, 127]. These strains were the
JMY 798 (MTLY 36-2P) and JMY 794 (MTLY 40-2P)
deriving from the wild strain W29 (ATCC 20460), and
despite the disruption of the genes encoding for the synthesis
of the various Aox, the genetically modified strains presented
significant and comparable growth with the W29 wild strain
during growth on pure fatty oleic acid, but in the former case
noticeable quantities of microbial lipids were accumulated as
high-size intra-cellular lipid droplets, the so-called lipid
bodies [126, 127]. The lipid bodies are considered as depots
of neutral lipids (TAGs and to lesser extent steryl-esters)
enclosed by polar fractions, while in a relatively recent devel-
opment, extensive proteomic analysis of the lipid-particle
proteins has been elaborated [35].
When fats are used as sole carbon and energy source,
there is not any de novo lipid biosynthesis from the acetyl-
CoA, since fatty acid synthetase (FAS) and ATP-citrate lyase
(ACL) are strongly inhibited by the presence of exogenous
aliphatic chains. Therefore, the presence of hydrophobic
0
5
10
15
20
25
0 50 100 150 200 250 300
X (g/L)
S (g/L)
Xf (g/L)
L (g/L)
Bio
mas
s, X
(g/L
); Li
pid-
free
bio
mas
s, X
f (g/
L);
Su
bstr
ate
fat,
S (g
/L);
Cel
lula
r lip
id, L
(g/L
)
Time (h)
0
20
40
60
80
100
0 50 100 150 200 250 300
DOT (%, v/v)
Lipid (%, w/w)
O2 s
atu
rati
on (%
, v/v
);
Lipi
d in
dry
wei
ght
(%, w
t/w
t)
Time (h)
(a)
(b)
Figure 6. Kinetics of the oleaginous yeast Yarrowia lipolytica on an
industrial derivative of tallow composed of free-fatty acids (stearin)
used as the sole carbon source. Representation of biomass produc-
tion (X, g/L), lipid production (L, g/L), lipid-free material production
(Xf, g/L), substrate fat consumption (S, g/L) (a), dissolved oxygen
tension (DOT, % v/v) and lipid in dry weight evolution (b) (data from
Papanikolaou et al. [131]).
0
10
20
30
40
50
60
0 0.25 0.5 1 2 4 5 6 10 1 2 3 4 5
Maximum accumulated lipid (%, w/w)
Perc
enta
ge (%
, w/w
)
Ammonium sulphate (g/L)
S0
SL/g 01=0=20 g/L
Figure 7. Kinetics of the oleaginous yeast Yarrowia lipolytica on an
industrial derivative of tallow composed of free-fatty acids (stearin)
used as the sole carbon source. Representation of maximum SCO
produced (% w/w, in dry mass) during growth at initial stearin con-
centration 10 or 20 g/L and variations in the initial concentration of
ammonium sulphate (data from Papanikolaou et al. [131]).
1040 S. Papanikolaou and G. Aggelis Eur. J. Lipid Sci. Technol. 2011, 113, 1031–1051
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

materials in the culture medium should be generally incom-
patible with de novo biogenesis of cellular lipids even if
glucose or other hydrophilic materials presented into the
medium and were co-metabolized with the exogenous fatty
acids [34, 105]. For instance, Meyer and Schweizer [105]
proceeded with growth of S. cerevisiae and C. lipolytica strains
on mixtures of glucose and pentadecanoic acid. It has been
reported that, although the extra-cellular concentration of
pentadecanoıc acid was indeed low (about 0.3% w/v), the
fatty acid synthetase activity of the strains was completely
repressed, having as a result, only the presence of odd fatty
acids into the cellular lipids. However, in more recent inves-
tigations it has been demonstrated that some de novo bio-
synthesis of fatty acids from glucose or glycerol could
potentially occur, in spite of the presence of significant
exogenous quantities of fatty materials; for instance, the
biochemical behavior of P. methanolica HA-32 was studied
in media composed of glucose (at 50 g/L) and fish oil (or its
derivatives ethyl-esters or free-fatty acids) (at 30 g/L). The
fish oil (or its derivatives) contained significant quantities of
docosahexanoic acid (DHA—D6,9,12,15,17,19C22:6), a fatty
acid that through the de novo lipid accumulation from glu-
cose can never be synthesized in P. methanolica [123].
Although the extra-cellular presence of TAGs or free-fatty
acids into the culture medium resulted in an intra-cellular
fatty acid profile similar to that of the substrate fat, and,
therefore, significant DHA quantities were detected into
the SCO produced presumably through its direct incorpora-
tion from the fatty substrate, the presence of fatty acid ethyl-
esters resulted in the synthesis of a cellular lipid containing
very restricted quantities of DHA, and presenting similarities
with the one that had been synthesized when glucose was
used as the sole carbon source [123]. Therefore, the yeast P.
methanolica could perform de novo fatty acid biosynthesis
from glucose, in spite of the presence of exogenous aliphatic
onto the culture medium [123]. Likewise, in investigations
performed with the yeast Y. lipolytica ACA-DC 50109 in
submerged cultures, in which growth was supported by the
simultaneous use of stearin (mixtures of free-fatty acids and
mainly of C16:0 and C18:0) and industrial glycerol or glu-
cose, extensive studies of the intra-cellular lipid profile of
microbial lipid produced, equally suggested some de novo
synthesis of intra-cellular fatty acids, in spite of the presence
of long-chain fatty acids found into the culturemedium [26, 102];
when Y. lipolytica ACA-DC 50109 had been cultured on
stearin utilized as the sole substrate, no dehydrogenation
or elongation reactions were conducted in cellular level
[40], while the microbial lipid produced was almost com-
pletely saturated and was composed mainly of C18:0 fatty
acid [40, 131]. Given that growth ofY. lipolytica on glucose or
glycerol used as the sole substrate is accompanied by syn-
thesis of a lipid that is globally unsaturated [49, 74, 102, 133],
enrichment of the reserve lipid with unsaturated fatty acids
during growth on glucose/stearin or glycerol/stearin mixtures
(principally D9C18:1 and D9,12C18:2) that occurred, indicates
de novo fatty acid biosynthesis despite the presence of long
aliphatic chains into the medium (see relevant results in
Table 2).
The oleaginous yeasts growing on various hydrophobic
materials (e.g. oils, fats, free-fatty acids, FAMEs), accumu-
late and at the same time modify the fatty acid composition of
the employed fatty substance utilized as substrate [21, 25, 39,
40, 108, 117, 119, 122, 125]. The phenomena that are
controlling the fatty acid composition of the cellular lipids
are the specific rate of incorporation of substrate aliphatic
chains inside themicrobial cells and the intra-cellular changes
of fatty acids defined by the enzymatic capabilities of the
microorganism [25, 122]. In addition, the (typo- and
stereo-) specificity of the microbial lipases versus the sub-
strate fat (in the case in which TAGs are used as carbon
source), can equally potentially lead to a biomodification of
the substrate fatty acid composition as a function of the
bioconversion time [33, 122, 125]. For all of the above cited
reasons, the fatty acid profile of the accumulated reserve lipid
can potentially be significantly distinguishable comparedwith
that of the initial fat substrate employed. Various oleaginous
yeasts, possessing active systems of intra-cellular desatura-
tions (desaturases D9 and D12), can potentially be implicated
in the accumulation of a more unsaturated fat compared with
the substrate [21, 117, 120, 141]. In other cases (growth on
various fish oils) yeast strains may consume more rapidly
from the substrate the saturated (C14:0 and C16:0) and
oligounsaturated (D9C18:1) fatty acids, storing into their cells
the polyunsaturated fatty acids (e.g. DHA) fatty acids, result-
ing equally in the synthesis of a SCO more unsaturated
compared with the substrate fat [39]. On the other hand,
various yeasts present the tendency to dissimilate for growth
and maintenance the unsaturated and lower aliphatic chain
fatty acids, and accumulate the saturated ones, resulting,
thus, in the synthesis of cellular lipids more saturated com-
pared with the substrate [25, 40, 122, 125]. Likewise, in other
cases, the accumulated lipid composition is almost the same
with that of the substrate [33, 37, 119], whereas, finally, it is
interesting to indicate that in some cases the fatty acid com-
position of the intra-cellular free-fatty acids can present
remarkable differences in comparison with the stored
TAGs [128]. The fatty acid profiles of the initial substrate
fat used and the accumulated lipid for fermentations per-
formed by oleaginous microorganisms is presented in
Table 3. Likewise, in various cases growth of oleaginous
microorganisms on fatty mixtures may result in selective
uptake of the various individual fatty acids from the culture
medium, and during fermentation the extra-cellular fat also
may present significant compositional differences compared
with the initial substrate fat [37, 125, 142]. The fatty acid
profiles of the initial lipid substrate and the remaining lipid
fraction after fat fermentations performed by eukaryotic
microorganisms are presented in Table 4.
Eur. J. Lipid Sci. Technol. 2011, 113, 1031–1051 The biochemistry of oleaginous yeasts 1041
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

2.3 Degradation of reserve lipid in the oleaginousmicroorganisms
After exhaustion or decrease in the uptake rate of the carbon
source in the culture medium the oleaginous microorgan-
isms, as a general rule, consume their own lipid reserves.
Several types of studies have been performed for this purpose
using oleaginous yeasts, molds and bacteria and under var-
ious environmental conditions, breakdown (turnover) of the
previously produced lipophilic materials was conducted, with
this process being performed simultaneously with biosyn-
thesis of new lipid-free biomass [5, 6, 25, 37, 40, 47, 48,
124, 125, 129–135, 143, 144]. Likewise, a significant part of
work has been performed in relation the mobilization of
neutral lipids in non-oleaginous yeasts (inmost cases research
performed with S. cerevisiae—see i.e.: Athenstaedt et al.
[145]; for review see: Mullner and Daum [107]).
Utilization of TAGs and steryl-esters (STEs) (the main lipid
compounds found in both oleaginous and non-oleaginous
microorganisms—see: Ratledge [20]; Mullner and Daum
[107]) from the lipid particles requires the action of TAG
lipase(s) (EC 3.1.1.3) and STE hydrolase(s) (EC 3.1.1.13).
An intra-cellular lipase’s and hydrolase’s system is expressed
either constitutively or inducibly [146, 147], being respon-
sible for the cleavage of the esters and the generation of fatty
acids that will be subsequently catabolized [34, 146, 147].
Moreover, in the case of plant or mammalian cells, in which
storage lipid mobilization is also routinely observed, some
proteins associated with the phospholipid monolayer of lipid
particles, such as perilipins (in the case of mammalian cells)
and oleosins (in the case of the plants), were assumed to be
involved in the mobilization of the neutral lipid core of the
particle by serving as docking and/or activating proteins for
TAG lipases and STE hydrolases [107]. Analysis of the most
abundant yeast lipid particle proteins by mass spectrometry
led to the identification of several polypeptides with unknown
function [145], but in any case, no one of these polypeptides,
was homologous to perilipins and oleosins [107].
The presence of lipases and STE hydrolases is a prereq-
uisite in order for onset of storage lipid turnover to be given
[107, 146, 147], while carbon starvation conditions are also
required [25, 32, 124, 131, 134, 143, 144, 146]. In a series of
investigations it has been demonstrated that the phenomenon
of cellular lipid degradation in the oleaginous microorgan-
isms was independent of the culture ‘‘pre-history’’ (meaning,
in fact, independent of the type of the carbon source assimi-
lated by the microorganism in order to proceed with reserve
lipid accumulation), since such turnover has been observed
Table 2. Composition of lipid produced by Yarrowia lipolytica ACA-DC 5010 during growth on various carbon sources and fermentation
configurations
Fatty acid composition of stearin (% w/w)
C16:0 C18:0 C18:1 C18:2
25 52 2 T.
Fatty acid composition of microbial lipid produced (% w/w)
S0 (g/L) Glol0 (g/L) Glc0 (g/L) C16:0 C18:0 C18:1 C18:2 Reference
10 – – 13 83 3 T. Papanikolaou et al. [25] a)
– 30 – 15 10 45 20 Papanikolaou et al. [133] a)
– – 30.0 11 6 53 10 Papanikolaou et al. [102] a)
– 50 – 15 11 47 21 Papanikolaou and Aggelis [74] b)
10 11 – 16 72 7 2 Papanikolaou et al. [40] c)
10 10 – 15 76 5 2 Papanikolaou et al. [26] a)
10 23 – 16 68 7 2 Papanikolaou et al. [26] a)
10 34 – 14 67 10 3 Papanikolaou et al. [26] a)
11 – 21 15 68 7 3 Papanikolaou et al. [102] a)
11 – 35 15 68 8 3 Papanikolaou et al. [102] a)
9 – 29 17 56 18 8 Papanikolaou et al. [102] a)
T. <0.5% w/w. Representation of cellular fatty acid composition in the fermentation time in which the maximum concentration of microbial
lipid had been achieved. S0, initial concentration of fatty material (stearin, that is a fully saturated tallow derivative composed of free-fatty
acids); Glol0, initial concentration of glycerol (utilization of bio-diesel derived waste glycerol as substrate); Glc0, initial concentration of
glucose (utilization of commercial-glucose as substrate). Fatty acids and glucose (or glycerol) were simultaneously consumed by the
microorganism.a) Batch flask culture.b) Single-stage continuous culture.c) Batch bioreactor culture.
1042 S. Papanikolaou and G. Aggelis Eur. J. Lipid Sci. Technol. 2011, 113, 1031–1051
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com
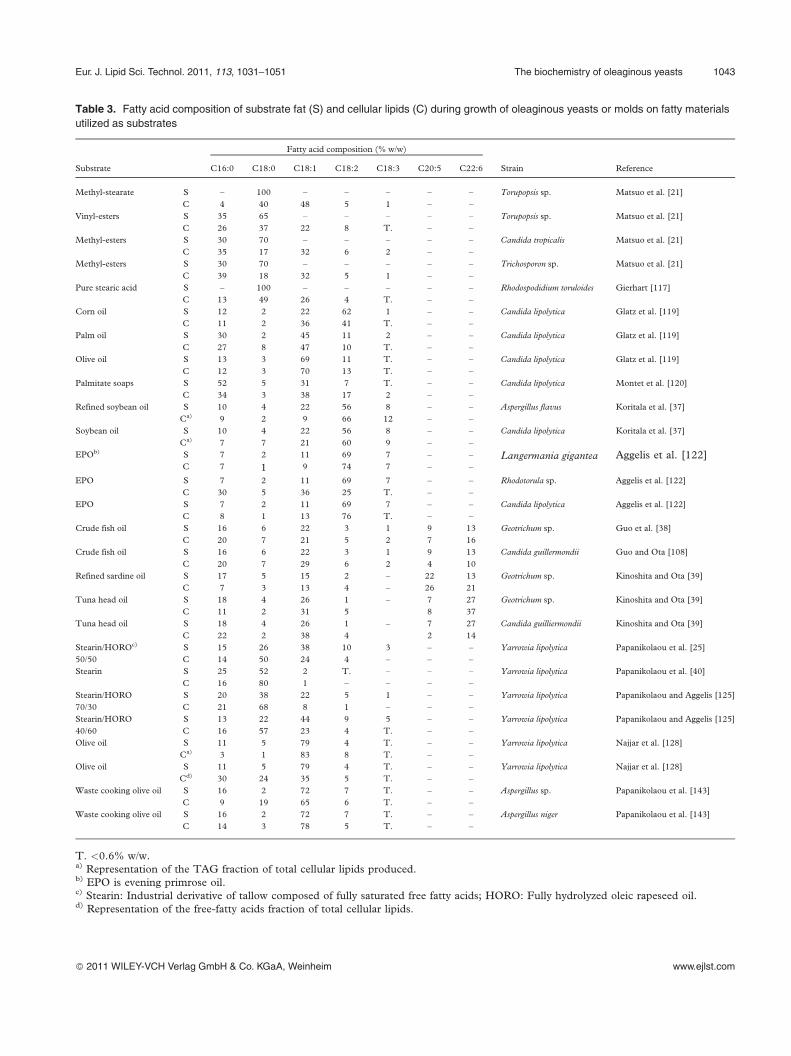
Table 3. Fatty acid composition of substrate fat (S) and cellular lipids (C) during growth of oleaginous yeasts or molds on fatty materials
utilized as substrates
Fatty acid composition (% w/w)
Substrate C16:0 C18:0 C18:1 C18:2 C18:3 C20:5 C22:6 Strain Reference
Methyl-stearate S – 100 – – – – – Torupopsis sp. Matsuo et al. [21]
C 4 40 48 5 1 – –
Vinyl-esters S 35 65 – – – – – Torupopsis sp. Matsuo et al. [21]
C 26 37 22 8 T. – –
Methyl-esters S 30 70 – – – – – Candida tropicalis Matsuo et al. [21]
C 35 17 32 6 2 – –
Methyl-esters S 30 70 – – – – – Trichosporon sp. Matsuo et al. [21]
C 39 18 32 5 1 – –
Pure stearic acid S – 100 – – – – – Rhodospodidium toruloides Gierhart [117]
C 13 49 26 4 T. – –
Corn oil S 12 2 22 62 1 – – Candida lipolytica Glatz et al. [119]
C 11 2 36 41 T. – –
Palm oil S 30 2 45 11 2 – – Candida lipolytica Glatz et al. [119]
C 27 8 47 10 T. – –
Olive oil S 13 3 69 11 T. – – Candida lipolytica Glatz et al. [119]
C 12 3 70 13 T. – –
Palmitate soaps S 52 5 31 7 T. – – Candida lipolytica Montet et al. [120]
C 34 3 38 17 2 – –
Refined soybean oil S 10 4 22 56 8 – – Aspergillus flavus Koritala et al. [37]
Ca) 9 2 9 66 12 – –
Soybean oil S 10 4 22 56 8 – – Candida lipolytica Koritala et al. [37]
Ca) 7 7 21 60 9 – –
EPOb) S 7 2 11 69 7 – – Langermania gigantea Aggelis et al. [122]C 7 1 9 74 7 – –
EPO S 7 2 11 69 7 – – Rhodotorula sp. Aggelis et al. [122]
C 30 5 36 25 T. – –
EPO S 7 2 11 69 7 – – Candida lipolytica Aggelis et al. [122]
C 8 1 13 76 T. – –
Crude fish oil S 16 6 22 3 1 9 13 Geotrichum sp. Guo et al. [38]
C 20 7 21 5 2 7 16
Crude fish oil S 16 6 22 3 1 9 13 Candida guillermondii Guo and Ota [108]
C 20 7 29 6 2 4 10
Refined sardine oil S 17 5 15 2 – 22 13 Geotrichum sp. Kinoshita and Ota [39]
C 7 3 13 4 – 26 21
Tuna head oil S 18 4 26 1 – 7 27 Geotrichum sp. Kinoshita and Ota [39]
C 11 2 31 5 8 37
Tuna head oil S 18 4 26 1 – 7 27 Candida guilliermondii Kinoshita and Ota [39]
C 22 2 38 4 2 14
Stearin/HOROc) S 15 26 38 10 3 – – Yarrowia lipolytica Papanikolaou et al. [25]
50/50 C 14 50 24 4 – – –
Stearin S 25 52 2 T. – – – Yarrowia lipolytica Papanikolaou et al. [40]
C 16 80 1 – – – –
Stearin/HORO S 20 38 22 5 1 – – Yarrowia lipolytica Papanikolaou and Aggelis [125]
70/30 C 21 68 8 1 – – –
Stearin/HORO S 13 22 44 9 5 – – Yarrowia lipolytica Papanikolaou and Aggelis [125]
40/60 C 16 57 23 4 T. – –
Olive oil S 11 5 79 4 T. – – Yarrowia lipolytica Najjar et al. [128]
Ca) 3 1 83 8 T. – –
Olive oil S 11 5 79 4 T. – – Yarrowia lipolytica Najjar et al. [128]
Cd) 30 24 35 5 T. – –
Waste cooking olive oil S 16 2 72 7 T. – – Aspergillus sp. Papanikolaou et al. [143]
C 9 19 65 6 T. – –
Waste cooking olive oil S 16 2 72 7 T. – – Aspergillus niger Papanikolaou et al. [143]
C 14 3 78 5 T. – –
T. <0.6% w/w.a) Representation of the TAG fraction of total cellular lipids produced.b) EPO is evening primrose oil.c) Stearin: Industrial derivative of tallow composed of fully saturated free fatty acids; HORO: Fully hydrolyzed oleic rapeseed oil.d) Representation of the free-fatty acids fraction of total cellular lipids.
Eur. J. Lipid Sci. Technol. 2011, 113, 1031–1051 The biochemistry of oleaginous yeasts 1043
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

even though SCO accumulation previously occurred through
de novo [30, 32, 67, 144, 148] or ex novomechanism [25, 40,
124, 129, 131, 134, 143]. In any case, the released cellular
fatty acids will be catabolized via the process of b-oxidation
and the produced acetyl-CoAwill be further converted via the
Krebs cycle and the anaplerotic by-pass of glyoxylic acid [40,
67, 97, 148]; in the case in which de novo lipid accumulation
had been previously performed, during lipid turnover period,
sugar (that could potentially exist into the culture medium),
is no longer assimilated, while the function of Krebs cycle
through the utilization of NADþ- (and potentially NADPþ)-
isocitrate dehydrogenase had already been suppressed due to
extra-cellular nitrogen limitation [47, 99]. In general, the
activity of glyoxylic acid by-pass enzymes (carnitine acetyl-
transferase and iso-citrate lyase) increases considerably in
cells growing on C2 compounds (e.g. ethanol) or on sub-
strates leading to C2 unit’s formation (i.e. previously accu-
mulated TAGs, extra-cellular TAGs, hydrocarbons or free
fatty acids), while this activity is indeed reduced during
growth on glucose or other sugars [67, 142, 148, 149].
Therefore, even though microbial growth had been per-
formed on sugars or similarly metabolized compounds,
SCO turnover should be principally performed through the
glyoxylic acid by-pass pathway, while cellular nitrogen
(obtainable via AMP-desaminase) should secure the biosyn-
thesis of new lipid-free material [47]. On the other hand,
degradation of accumulated lipid when ex novo lipid
accumulation had been previously performed is conducted
indisputable principally through the glyoxylic acid anapler-
otic by-pass pathway; in fact, reserve lipid turnover generally
Table 4. Fatty acid composition of initial (S) and remaining (SR) substrate fat during growth of oleaginous yeasts or molds on fatty materials
utilized as substrates
Fatty acid composition (% w/w)
Substrate C16:0 C18:0 C18:1 C18:2 C18:3 C20:5 C22:6 Strain Reference
Refined soybean oil S 10 4 22 56 8 – – Candida lipolytica Koritala et al. [37]
SRa) 12 5 23 51 7 – –
SRb) 8 4 24 58 6 – –
Refined soybean oil S 10 4 22 56 8 – – Amylomyces rouxii Koritala et al. [37]
SRa) 12 4 22 55 8 – –
SRb) 4 1 24 59 8 – –
Refined soybean oil S 10 4 22 56 8 – – Cunninghamella
blakesleeana
Koritala et al. [37]
SRa) 18 7 24 46 6 – –
SRb) 5 2 24 61 8 – –
EPOc) S 7 2 11 69 7 – – Langermania gigantea Aggelis et al. [122]
SR 9 2 10 72 7 – –
EPO S 7 2 11 69 7 – – Rhodotorula sp. Aggelis et al. [122]
SR 7 2 10 72 8 – –
EPO S 7 2 11 69 7 – – Candida lipolytica Aggelis et al. [122]
SR 7 2 11 71 9 – –
Refined sardine oil S 17 5 15 2 – 22 13 Geotrichum sp. Kinoshita and Ota [39]
SR 5 4 10 2 – 37 19
Stearin/HOROd) S 20 38 22 5 1 – – Yarrowia lipolytica Papanikolaou and Aggelis [125]
70/30 SR 22 74 4 T. – – –
Stearin/HORO S 16 33 29 7 3 – – Yarrowia lipolytica Papanikolaou and Aggelis [125]
60/40 SR 15 76 8 T. – – –
Stearin/HORO S 13 22 44 9 5 – – Yarrowia lipolytica Papanikolaou and Aggelis [125]
40/60 SR 17 71 11 T. T. – –
Sunflower oil S 5 5 34 53 n.r. – – Yarrowia lipolytica Kamzolova et al. [142]
SR 5 24 27 35 n.r. – –
Waste cooking olive oil S 16 2 72 7 T. – – Aspergillus niger Papanikolaou et al. [143]
SR 5 4 88 3 T. – –
Waste cooking olive oil S 16 2 72 7 T. – – Penicillium expansum Papanikolaou et al. [143]
SR 11 2 83 4 T. – –
T. <0.5% w/w; n.r.: not reported.a) Free-fatty acid fraction of the residual oil.b) TAG fraction of the residual oil.c) EPO is evening primrose oil.d) Stearin: Industrial derivative of tallow composed of fully saturated free-fatty acids; HORO: Fully hydrolyzed oleic rapeseed oil.
1044 S. Papanikolaou and G. Aggelis Eur. J. Lipid Sci. Technol. 2011, 113, 1031–1051
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

occurs when the extra-cellular flow rate of aliphatic chains is
considerably decreased. This fact can be either due to the
apparent absence of substrate fat from the culture medium
[134, 143] or due to the presence of fat that is substantially
rich in the fatty acid C18:0 and cannot be easily catabolized
by the yeast strains due to discrimination against this fatty
acid [25, 40, 121, 124, 125]. In the case of ex novo lipid
accumulation, culture conditions could be still favorable for
microbial growth since extra-cellular nitrogen may be pre-
sented into the medium even during reserve lipid degradation
[40]. Additionally, biosynthesis of carnitine acetyl-transferase
and iso-citrate lyase had been previously induced due to the
cultivation on fatty materials [142, 143, 150]. Moreover, in
recent investigations it has been demonstrated that lack of
some nutritional components such as iron and magnesium
from the fermentation medium, can partially or even com-
pletely suppress the degradation of microbial lipids [47]. On
the other hand, utilization of multiple-limited media did not
negatively influence the process of storage lipid accumu-
lation, and therefore utilization of such types of media
presents obvious interest for the process of SCO fermentation
in industrial scale [47].
Microbial lipid turnover has been extensively studied inC.
echinulata and Y. lipolytica. It has been demonstrated that C.
echinulata tended to selectively degrade its stored TAGs
instead of the other neutral lipid fractions (e.g. diacylgly-
cerols, free-fatty acids, STE etc) [32]. Moreover, the intra-
cellular saturated and monounsaturated fatty acids were rap-
idly and selectively consumed, while the polyunsaturated
ones (e.g. D9,12C18:2, D6,9,12C18:3) were not. Therefore,
after accomplishment of SCO turnover, the remaining cel-
lular lipid was drastically more unsaturated compared with
the originally accumulated lipid [32]. On the contrary, Y.
lipolytica tended to dissimilate for growth and energy require-
ments the unsaturated fatty acids regardless of their concen-
trations into the stored lipid reserves, and thus, after storage
lipid turnover, the remaining cellular lipid was almost
completely saturated [125]. Mobilization of microbial
lipophilic compounds has also been studied in oleaginous
bacteria; in this case the tested strains had previously
accumulated both TAGs and poly-hydroxy-alkanoates
(PHAs), and as in the case of oleaginous yeasts and molds
turnover of storage materials occurred due to shifting from
carbon excess to carbon limited conditions [146]. In this case,
it has been revealed that simultaneous consumption of
TAGs and PHAs at the stationary growth phase was
performed [146].
Reserve lipid turnover was in general accompanied by
generation of lipid-free material (Xf) providing evidence that
accumulated lipids act as precursor in the biosynthesis of cell
materials. Holdsworth and Ratledge [144] have proceeded
with a two-stage fermentation of the oleaginous yeast C.
curvata growing on glucose. In the first stage, lipid accumu-
lation was enhanced (growth on glucose, initial molar ratio
C/N ¼ 60), while, in the second stage, no extra-cellular car-
bon source was employed. The microorganism used its
accumulated lipids; in this case, the obtained yield YXf/L
(g of lipid-free biomass formed per 1 g of cellular lipid
consumed) was about 1.9 g/g [144] that can be considered
as somehow elevated, since the maximum theoretical
yield of conversion of TAGs into lipid-free biomass is around
1.7 g/g [30, 32, 144]. The same yield presented drastically
lower values (0.6–0.8 g/g) during growth of Y. lipolytica
ACA-DC 50109 on industrial fats composed of mixtures
of saturated and unsaturated fatty acids [25, 40, 124], whilst
growth of Mucor circinelloides on vegetable oils was accom-
panied by a yield YXf/L 0.63–0.66 g/g [134]. Finally, growth
of the mold C. echinulata on glucose enriched tomato
waste hydrolysate was accompanied by cellular lipid degra-
dation with yield YXf/L value 0.95 g/g [32], with the respect-
ive value for M. isabellina cultured on glucose being around
1.0 � 0.1 g/g [47] and on starch being slightly lower (0.7–
0.8 g/g) [48].
3 Concluding remarks—future perspectives
The production of SCOs presents continuous expansion the
last years. Amongst microbial lipids, yeast lipids present a
high importance in both academic and industrial point of
view. Due to their unicellular nature and their potentiality to
grow on a plethora of hydrophilic or hydrophobic substrates,
the oleaginous yeasts are considered as perfect ‘‘tools’’ for
studying phenomena of advanced lipid biochemistry and
biotechnology [4, 8, 132, 135]. Interestingly, the last years
there have been works indicating interesting analogies
between lipid metabolism in the yeast cells and the gastro-
intestinal tract and vascular system [128, 151].
Fundamental differences in biochemical level exist
between de novo lipid accumulation from hydrophilic sub-
strates and ex novo lipid accumulation from hydrophobic
substrates. In the former case, lipid production is a secondary
anabolic activity occurring after nitrogen had been depleted
from the growth medium. On the other hand, ex novo lipid
production is a growth associated process occurring simul-
taneously with cell growth, being entirely independent from
nitrogen exhaustion from the culture medium. The major
industrial applications related with the de novo accumulation
of storage lipid in yeasts refer to the production of ‘‘non-
specified’’ lipids that will be further converted into FAMEs
(2nd generation bio-diesel), since the conventional utilization
of edible oils as starting materials for the synthesis of bio-
diesel has resulted in significant increase in their price the last
years [1]. Moreover, through the de novo lipid accumulation,
it is possible to produce yeast lipids that have a fatty acid
composition similar to that of cocoa-butter [3, 4, 19, 20]. The
prospective (significant) rise in the price of cocoa-butter
indicates again the potentiality of the utilization of oleaginous
yeasts to produce cocoa butter-like cellular lipids. Needless
to indicate that for both the above mentioned potentialities,
the utilization of low- or even negative-cost hydrophilic
Eur. J. Lipid Sci. Technol. 2011, 113, 1031–1051 The biochemistry of oleaginous yeasts 1045
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

materials as substrates should be envisaged (hemicelluloses
hydrolysates, bio-diesel derived waste glycerol, whey, sugar-
based wastes, etc.), in order for an economically viable bio-
process to be performed.
The process of lipid production from fatty materials is
critically influenced by the fatty acid composition of the fat
used as substrate. ‘‘New’’ fatty acid profiles (in both extra-
and intra-cellular level) that did not previously exist in the
substrate fat may be produced during fat fermentation led by
the oleaginous yeasts. An important application of the ex
novo lipid production refers to the valorization of fatty wastes
and the production of lipid-rich microbial biomass (contain-
ing in several cases valuable PUFAs) that will further be used
as food or feed supplement [33, 38, 40, 108, 119, 120, 124,
131, 143]. On the other hand, themost important application
of the ex novo lipid accumulation process in the oleaginous
microorganisms refers to the ‘‘improvement’’ and the ‘‘up-
grade’’ of the fatty materials utilized as substrates in order to
produce ‘‘tailor-made’’ lipids of high-added value. Such cases
refer to the valorization of low-cost saturated fatty acids so as
to produce intra-cellular substitutes of cocoa-butter or other
high-value exotic fats like shea butter, sal fat etc (for review
see: Papanikolaou and Aggelis [8]) or the valorization of low-
or negative-cost unsaturated fatty substances in order to
produce lipids containing themedically important g-linolenic
acid [129, 130, 152].
The number of research investigations on the topic of
SCO fermentation presents a significant expansion the last
years. Research teams work on the identification and selec-
tion of ‘‘new’’ naturally occurring oleaginous strains capable
of storing huge lipid quantities inside their cells or mycelia.
Optimization of culture conditions related with the maximi-
zation of SCO production is being currently performed.
These studies present interest for future experiments.
Likewise, studies that concern advanced lipid biochemistry,
identification of limiting steps related with the accumulation
of storage lipid inside the cells or mycelia of the oleaginous
microorganisms and extensive analysis of microbial lipid
produced are also performed, while, equally, these studies
present interest for future experimental work. The same is
stressed in relation with investigations dealing with cloning of
specific genes coding for lipid-accumulating key enzymes
[153], kinetic modeling [29, 102, 124, 129, 130] and meta-
bolic flux analysis and finally proteomic—genomic analysis
[154] for several oleaginous microorganisms.
Financial support concerning the results achieved by our research
team has been kindly provided by: Agricultural University of
Athens; University of Patras; Dracoil SA; Project BIOSIS
(INTERREG III GREECE, ITALY); State Scholarship
Foundation (Athens, Greece); General Secretary of Research and
Technology (Ministry of Development, Greek Government).
The authors have declared no conflict of interest.
References
[1] Ratledge, C., Cohen, Z., Microbial and algal lipids: Do theyhave a future for biodiesel or as commodity oils? LipidTechnol. 2008, 20, 155–160.
[2] Ratledge, C., Wynn, J., The biochemistry and molecularbiology of lipid accumulation in oleaginous microorgan-isms. Adv. Appl. Microbiol. 2002, 51, 1–51.
[3] Davies, R. J., Holdsworth, J. E., Synthesis of lipids in yeasts:Biochemistry, physiology and production. Adv. Appl. LipidRes. 1992, 1, 119–159.
[4] Ratledge, C., Microbial lipids, in: Kleinkauf, H., Dohren,H. (Eds.), Biotechnology, 2nd Edn., Vol. 7, Wiley-VCH,Weinheim (Germany) 1997, pp. 135–197.
[5] Fakas, S., Certik, M., Papanikolaou, S., Aggelis, G., et al.,g-Linolenic acid production by Cunninghamella echinulatagrowing on complex organic nitrogen sources. Bioresour.Technol. 2008, 99, 5986–5990.
[6] Fakas, S., Papanikolaou, S., Galiotou-Panayotou, M.,Komaitis, M., Aggelis, G., Organic nitrogen of tomatowaste hydrolysate enhances glucose uptake and lipidaccumulation in Cunninghamella echinulata. J. Appl.Microbiol. 2008, 105, 1062–1070.
[7] Papanikolaou, S., Aggelis, G., Biotechnological valorizationof biodiesel derived glycerol waste through production ofsingle cell oil and citric acid by Yarrowia lipolytica. LipidTechnol. 2009, 21, 83–87.
[8] Papanikolaou, S., Aggelis, G., Yarrowia lipolytica: A modelmicroorganism used for the production of tailor-made lip-ids. Eur. J. Lipid Sci. Technol. 2010, 112, 639–654.
[9] Li, Y. H., Zhao, Z. B., Bai, F. W., High density cultivationof oleaginous yeastRhodosporidium toruloides Y4 in fed-batchculture. Enzyme Microb. Technol. 2007, 41, 312–317.
[10] Angerbauer, C., Siebenhofer, M., Mittelbach, M., Guebitz,G.M., Conversion of sewage sludge into lipids by Lipomycesstarkeyi for biodiesel production. Bioresour. Technol. 2008,99, 3051–3056.
[11] Zhao, X., Kong, X., Hua, Y., Feng, B., Zhao, Z. B.,Medium optimization for lipid production through co-fer-mentation of glucose and xylose by the oleaginous yeastLipomyces starkeyi. Eur. J. Lipid Sci. Technol. 2008, 110,405–412.
[12] Zhu, L. Y., Zong,M. H., Wu, H., Efficient lipid productionwith Trichosporon fermentans and its use for biodiesel prep-aration. Bioresour. Technol. 2008, 99, 7881–7885.
[13] Subramaniam, R., Dufreche, S., Zappi, M., Bajpai, R.,Microbial lipids from renewable resources: Productionand characterization. J. Ind. Microbiol. Biotechnol. 2010,37, 1271–1287.
[14] Wu, S., Hu, C., Jin, C., Zhao, X., Zhao, Z. B., Phosphate-limitation mediated lipid production by Rhodosporidium tor-uloides. Bioresour. Technol. 2010, 101, 6124–6129.
[15] Wu, S., Hu, C., Zhao, X., Zhao, Z. B., Production of lipidfrom N-acetylglucosamine by Cryptococcus curvatus. Eur. J.Lipid Sci. Technol. 2010, 112, 727–733.
[16] Wu, H., Li, Y., Chen, L., Zong, M. H., Production ofmicrobial oil with high oleic acid content by Trichosporoncapitatum. Appl. Energy 2011, 88, 138–142.
[17] Ratledge, C., Boulton, C. A., Fats and Oils, in: Moo-Young, M., Blanch, H. N., Drew, S., Wang, D. I. C.(Eds.), Comprehensive Biotechnology, the Principles,
1046 S. Papanikolaou and G. Aggelis Eur. J. Lipid Sci. Technol. 2011, 113, 1031–1051
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

Applications and Regulations of Biotechnology in Industry,Agriculture and Medicine, Pergamon Press, New York(USA) 1985, pp. 983–1003.
[18] Ratledge, C., Biochemistry, stoichiometry, substrates andeconomics, in: Moreton, R. S. (Ed.), Single Cell Oil,Longman Scientific & Technical, Harlow (UK) 1988,pp. 33–70.
[19] Ratledge, C., Single cell oils – have they a biotechnologicalfuture? Trends Biotechnol. 1993, 11, 278–284.
[20] Ratledge, C., Yeasts, moulds, algae and bacteria as sourcesof lipids, in: Kamel, B. S., Kakuda, Y. (Eds.), TechnologicalAdvances in Improved and Alternative Sources of Lipids,Blackie Academic and Professional, London (UK) 1994,pp. 235–291.
[21] Matsuo, T., Terashima, M., Hashimoto, Y., Hasida, W.,US Patent 4308 350, 1981.
[22] Davies, R. J., Yeast oil from cheese whey – process develop-ment, in: Moreton, R. S. (Ed.), Single Cell Oil, LongmanScientific & Technical, Harlow (UK) 1988, pp. 99–146.
[23] Moreton, R. S., Physiology of lipid accumulating yeasts, in:Moreton, R. S. (Ed.), Single Cell Oil, Longman Scientific &Technical, Harlow (UK) 1988, pp. 1–32.
[24] Aggelis, G., Stathas, D., Tavoularis, N., Komaitis, M.,Composition of lipids produced by some strains ofCandida species. Production of single-cell oil in a chemostatculture. Folia Microbiol. 1996, 41, 299–302.
[25] Papanikolaou, S., Chevalot, I., Komaitis, M., Aggelis, G.,Marc, I., Kinetic profile of the cellular lipid composition inan oleaginous Yarrowia lipolytica capable of producing acocoa-butter substitute from industrial fats. Antonie VanLeeuwenhoek 2001, 80, 215–224.
[26] Papanikolaou, S., Muniglia, L., Chevalot, I., Aggelis, G.,Marc, I., Accumulation of a cocoa-butter-like lipid byYarrowia lipolytica cultivated on agro-industrial residues.Curr. Microbiol. 2003, 46, 124–130.
[27] Beopoulos, A., Cescut, J., Haddouche, R., Uribelarrea,J. L., et al., Yarrowia lipolytica as a model for bio-oil pro-duction. Prog. Lipid Res. 2009, 48, 375–387.
[28] Fakas, S., Papanikolaou, S., Batsos, A., Galiotou-Panayotou, M., et al., Evaluating renewable carbon sourcesas substrates for single cell oil production byCunninghamellaechinulata and Mortierella isabellina. Biomass Bioenergy 2009,33, 573–580.
[29] Fakas, S., Bellou, S., Makri, A., Aggelis, G., Single cell oiland gamma-linolenic acid production by Thamnidium ele-gans grown on raw glycerol, in: Aggelis, G. (Ed.), MicrobialConversions of Raw Glycerol, Nova Science Publishers Inc.,New York (USA) 2009, pp. 85–99.
[30] Fakas, S., Papanikolaou, S., Galiotou-Panayotou, M.,Komaitis, M., Aggelis, G., Biochemistry and biotechnologyof single cell oil, in: Pandey, A., Larroche, C., Soccol, C. R.,Dussard, C. G. (Eds.), New Horizons in Biotechnology,AsiaTech Publishers Inc., New Delhi (India) 2009,pp. 38–60.
[31] Fakas, S., Papanikolaou, S., Galiotou-Panayotou, M.,Komaitis, M., Aggelis, G., Lipids of Cunninghamellaechinulata with emphasis to g-linolenic acid distributionamong lipid classes. Appl. Microbiol. Biotechnol. 2006, 73,676–683.
[32] Fakas, S., Galiotou-Panayotou, M., Papanikolaou, S.,Komaitis, M., Aggelis, G., Compositional shifts in lipid
fractions during lipid turnover in Cunninghamella echinulata.Enzyme Microb. Technol. 2007, 40, 1321–1327.
[33] Bati, N., Hammond, E. G., Glatz, B. A., Biomodification offats and oils: Trials with Candida lipolytica. J. Am. Oil Chem.Soc. 1984, 61, 1743–1746.
[34] Alvarez, H. M., Kalscheuer, R., Steinbuchel, A.,Accumulation of storage lipids in species of Rhodococcusand Nocardia and effect of inhibitors and polyethylene gly-col. Fett-Lipid 1997, 99, 239–246.
[35] Athenstaedt, K., Jolivet, P., Boulard, C., Zivy, M., et al.,Lipid particle composition of the yeast Yarrowia lipolyticadepends on the carbon source. Proteomics 2006, 6, 1450–1459.
[36] Thorpe, R. F., Ratledge, C., Fatty acid distribution intriglycerides of yeasts grown on glucose and n-alkanes.J. Gen. Microbiol. 1972, 75, 151–163.
[37] Koritala, S., Hesseltine, C. W., Pryde, E. H., Mounts,T. L., Biochemical modification of fats by microorganisms:A preliminary study. J. Am. Oil Chem. Soc. 1987, 64, 509–513.
[38] Guo, X., Tomonaga, T., Yanagihara, Y., Ota, Y., Screeningfor yeasts incorporating the exogenous eicosapentaenoicand docosahexaenoic acids from crude fish oil. J. Biosci.Bioeng. 1999, 87, 184–188.
[39] Kinoshita, H., Ota, Y., Concentration of docosahexaenoicacid from fish oils using Geotrichum sp. FO347-2. Biosci.Biotechnol. Biochem. 2001, 65, 1022–1026.
[40] Papanikolaou, S., Chevalot, I., Komaitis, M., Marc, I.,Aggelis, G., Single cell oil production by Yarrowia lipolyticagrowing on an industrial derivative of animal fat in batchcultures. Appl. Microbiol. Biotechnol. 2002, 58, 308–312.
[41] Dyal, S. D., Narine, S. S., Implications for the use ofMortierella fungi in the industrial production of essentialfatty acids. Food Res. Int. 2005, 38, 445–467.
[42] Aggelis, G., Ratomahenina, R., Arnaud, A., Galzy, P., et al.,Etude de l’influence des conditions de culture sur la teneuren acide gamma linolenique de souches de Mucor.Oleagineux 1988, 43, 311–317.
[43] Aggelis, G., Pina, M., Graille, J., Localisation de l’acidegamma linolenique dans les myceliums et les spores chezdeux Mucorales. Oleagineux 1990, 45, 229–232.
[44] Aggelis, G., Komaitis, M., Dimitroulias, G., Pina, M.,Graille, J., Possibilite de production d’acide gamma linole-nique par culture de Mucor circinelloides CBS 172-27sur quelques huiles vegetales. Oleagineux 1991, 46, 208–212.
[45] Chen, H. C., Chang, C. C., Production of g-linolenic acidby the fungus Cunninghamella echinulata CCRC 31840.Biotechnol. Prog. 1996, 12, 338–341.
[46] Papanikolaou, S., Komaitis, M., Aggelis, G., Single celloil (SCO) production by Mortierella isabellina grown onhigh-sugar content media. Bioresour. Technol. 2004, 95,287–291.
[47] Papanikolaou, S., Sarantou, S., Komaitis, M., Aggelis, G.,Repression of reserve lipid turnover in Cunninghamellaechinulata and Mortierella isabellina cultivated in multiple-limited media. J. Appl. Microbiol. 2004, 97, 867–874.
[48] Papanikolaou, S., Galiotou-Panayotou, M., Fakas, S.,Komaitis, M., Aggelis, G., Lipid production by oleaginousMucorales cultivated on renewable carbon sources. Eur. J.Lipid Sci. Technol. 2007, 109, 1060–1070.
Eur. J. Lipid Sci. Technol. 2011, 113, 1031–1051 The biochemistry of oleaginous yeasts 1047
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

[49] Papanikolaou, S., Fakas, S., Fick, M., Chevalot, I., et al.,Biotechnological valorisation of raw glycerol dischargedafter bio-diesel (fatty acid methyl-esters) manufacturingprocess: Production of 1,3-propanediol, citric acid andsingle cell oil. Biomass Bioenergy 2008, 32, 60–71.
[50] Certik, M., Shimizu, S., Biosynthesis and regulation ofmicrobial poly-unsaturated fatty acid production.J. Biosci. Bioeng. 1999, 87, 1–14.
[51] Sakuradani, E., Shimizu, S., Single cell oil production byMortierella alpina. J. Biotechnol. 2009, 114, 31–36.
[52] Sakuradani, E., Ando, A., Ogawa, J., Shimizu, S., Improvedproduction of various polyunsaturated fatty acids throughfilamentous fungus Mortierella alpina breeding. Appl.Microbiol. Biotechnol. 2009, 84, 1–10.
[53] Damude, H. G., Zhang, H., Farrall, L., Ripp, K. G., et al.,Identification of bifunctional delta 12/omega 3 fatty aciddesaturases for improving the ratio of omega 3 to omega 6fatty acids in microbes and plants. Proc. Natl. Acad. Sci.USA 2006, 103, 9446–9451.
[54] Damude, H. G., Pollak, D.M.W., Xue, Z., Zhu, Q. Q., USPatent WO2007136646 A2, 2007.
[55] Damude, H. G., Zhu, Q. Q., US Patent WO2007127381A2, 2007.
[56] Chang, L., Chen, D., Chen, Y., Nicaud, J. M., et al.,Production of functional g-linolenic acid (GLA) by expres-sion of fungal D12 and D6 desaturase genes in the oleagi-nous yeast Yarrowia lipolytica, in: Ching, J. F. (Ed.),Biocatalysis and Agricultural Biotechnology, CRC Press,Taylor & Francis Group, Boca Raton (USA) 2009,pp. 164–179.
[57] Peng, X.-W., Chen, H.-Z., Microbial oil accumulation andcellulase secretion of the endophytic fungi from oleaginousplants. Ann. Microbiol. 2007, 57, 239–242.
[58] Hui, L., Wan, C., Hai-Tao, D., Xue-Jiao, C., et al., Directmicrobial conversion of wheat straw into lipid by a cellulo-lytic fungus of Aspergillus oryzae A-4 in solid-state fermen-tation. Bioresour. Technol. 2010, 101, 7556–7562.
[59] Evans, C. T., Ratledge, C., A comparison of the oleaginousyeast Candida curvata, grown on different carbon sources incontinuous and batch culture. Lipids 1983, 18, 623–629.
[60] Ykema, A., Verbree, E. C., Kater, M. M., Smit, H.,Optimization of lipid production in the oleaginous yeastApiotrichum curvatum in whey permeate. Appl. Microbiol.Biotechnol. 1988, 29, 211–218.
[61] Ykema, A., Verbree, E. C., Nijkamp, H. J. J., Smit, H.,Isolation and characterization of fatty acid auxotrophs fromthe oleaginous yeast Apiotrichum curvatum. Appl. Microbiol.Biotechnol. 1989, 32, 76–84.
[62] Ykema, A., Verbree, E. C., Verwoert, I. I. G. S., Van derLinden, K. H., et al., Lipid production of revertants of Ufamutants from the oleaginous yeast Apiotrichum curvatum.Appl. Microbiol. Biotechnol. 1990, 33, 176–182.
[63] Brown, B. D., Hsu, K. H., Hammond, E. G., Glatz, B. A.,A relationship between growth and lipid accumulation inCandida curvata D. J. Ferm. Bioeng. 1989, 68, 344–352.
[64] Aggelis, G., Komaitis, M., Enhancement of single-cell oilproduction by Yarrowia lipolytica growing in the presence ofTeuchrium polium L. aqueous extract. Biotechnol. Lett. 1999,21, 747–749.
[65] Papanikolaou, S., Diamantopoulou, P., Chatzifragkou, A.,Philippoussis, A., Aggelis, G., Suitability of low-cost sugars
as substrates for lipid production by the fungus Thamnidiumelegans. Energy Fuel. 2010, 24, 4078–4086.
[66] Chatzifragkou, A., Fakas, S., Galiotou-Panayotou, M.,Komaitis, M., et al., Commercial sugars as substrates forlipid accumulation by Cunninghamella echinulata andMortierella isabellina fungi. Eur. J. Lipid Sci. Technol.2010, 112, 1048–1057.
[67] Vamvakaki, A.-N., Kandarakis, I., Kaminarides, S.,Komaitis, M., Papanikolaou, S., Cheese whey as a renew-able substrate for microbial lipid and biomass production byZygomycetes. Eng. Life Sci. 2010, 10, 348–360.
[68] Minkevich, I. G., Dedyukhina, E. G., Chistyakova, T. I.,The effect of lipid content on the elemental compositionand energy capacity of yeast biomass. Appl. Microbiol.Biotechnol. 2010, 88, 799–806.
[69] Huang, C., Zong, M. H., Wu, H., Liu, Q., Microbial oilproduction from rice straw hydrolysate by Trichosporon fer-mentans. Bioresour. Technol. 2009, 100, 4535–4538.
[70] Pan, L.-X., Yang, D.-F., Shao, L., Li, W., et al., Isolation ofthe oleaginous yeasts from the soil and studies of their lipid-producing capacities. Food Technol. Biotechnol. 2009, 47,215–220.
[71] Sajbidor, J., Certik, M., Dobronova, S., Influence of differ-ent carbon sources on growth, lipid content and fatty acidcomposition in four strains belonging to Mucorales.Biotechnol. Lett. 1988, 10, 347–350.
[72] Meesters, P. A. E. P., Huijberts, G.N.M., Eggink, G., Highcell density cultivation of the lipid accumulating yeastCryptococcus curvatus using glycerol as a carbon source.Appl. Microbiol. Biotechnol. 1996, 45, 575–579.
[73] Meesters, P. A. E. P., van der Wal, H., Weusthuis, R.,Eggink, G., Cultivation of the oleaginous yeastCryptococcus curvatus in a new reactor with improved mixingand mass transfer characteristics (SUPER-R). Biotechnol.Tech. 1996, 10, 277–282.
[74] Papanikolaou, S., Aggelis, G., Lipid production byYarrowia lipolytica growing on industrial glycerol in asingle-stage continuous culture. Bioresour. Technol. 2002,82, 43–49.
[75] Andre, A., Chatzifragkou, A., Diamantopoulou, P., Sarris,D., et al., Biotechnological conversions of bio-diesel derivedcrude glycerol by Yarrowia lipolytica strains. Eng. Life Sci.2009, 9, 468–478.
[76] Makri, A., Fakas, S., Aggelis, G., Metabolic activities ofbiotechnological interest in Yarrowia lipolytica grown onglycerol in repeated batch cultures. Bioresour. Technol.2010, 101, 2351–2358.
[77] Liang, Y., Cui, Y., Trushenski, J., Blackburn, J. W.,Converting crude glycerol derived from yellow grease tolipids through yeast fermentation. Bioresour. Technol.2010, 101, 7581–7586.
[78] Rymowicz, W., Fatykhova, A. R., Kamzolova, S. V.,Rywinska, A., Morgunov, I. G., Citric acid production fromglycerol-containing waste of biodiesel industry by Yarrrowialipolytica in batch, repeated batch, and cell recycle regimes.Appl. Microbiol. Biotechnol. 2010, 87, 971–979.
[79] Chi, Z., Pyle, D. J., Wen, Z., Frear, C., Chen, S.,A laboratory study of producing docosahexanoic acid frombiodiesel-waste glycerol by microalgal fermentation. Proc.Biochem. 2007, 42, 1537–1545.
[80] Pyle, D. J., Garcia, R. A., Wen, Z., Producing docosahex-aenoic acid (DHA)-rich algae from biodiesel-derived crude
1048 S. Papanikolaou and G. Aggelis Eur. J. Lipid Sci. Technol. 2011, 113, 1031–1051
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

glycerol: Effects of impurities on DHA production and algalbiomass composition. J. Agric. Food Chem. 2008, 56, 3933–3939.
[81] Wen, Z., Pyle, D. J., Athalye, S. K., Production of omega-3polyunsaturated fatty acids from biodiesel-derived crudeglycerol by microalgal and fungal fermentation, in:Aggelis, G. (Ed.), Microbial Conversions of Raw Glycerol,Nova Science Publishers Inc., New York (USA) 2009,pp. 41–63.
[82] Andre, A., Diamantopoulou, P., Philippoussis, A., Sarris,D., et al., Biotechnological conversions of bio-diesel derivedwaste glycerol into added-value compounds by higher fungi:Production of biomass, single cell oil and oxalic acid. Ind.Crops Prod. 2010, 31, 407–416.
[83] Chatzifragkou, A., Makri, A., Belka, A., Bellou, S., et al.,Biotechnological conversions of biodiesel derived wasteglycerol by yeast and fungal species. Energy 2011, 36,1097–1108.
[84] Eroshin, V. K., Krylova, N. I., Efficiency of lipid synthesisby yeast. Biotechnol. Bioeng. 1983, 325, 1693–1700.
[85] Yamauchi, H., Mori, H., Kobayashi, T., Shimizu, S., Massproduction of lipids by Lipomyces starkeyi in microcomputeraided fed-batch culture. J. Ferm. Technol. 1983, 61, 275–280.
[86] Emelyanova, E., Lipid and g-linolenic acid production byMucor inaquisporus. Proc. Biochem. 1997, 3, 173–177.
[87] Papanikolaou, S., Microbial conversion of glycerol into 1,3-propanediol: Glycerol assimilation, biochemical eventsrelated with 1,3-propanediol biosynthesis and biochemicalengineering of the process, in: Aggelis, G. (Ed.), MicrobialConversions of Raw Glycerol, Nova Science Publishers Inc.,New York (USA) 2009, pp. 137–168.
[88] Aggelis, G., Two alternative pathways for substrate assim-ilation byMucor circinelloides. Folia Microbiol. 1996, 41, 254–256.
[89] Roux, M. P., Kock, J. L. F., Du Preez, J. C., Botha, A., Theinfluence of dissolved oxygen tension on the production ofcocoa butter equivalents and gamma-linolenic acid byMucor circinelloides. System. Appl. Microbiol. 1995, 18,329–334.
[90] Botha, A., Strauss, T., Kock, J. L. F., Pohl, C. H., Coetzee,D. J., Carbon source utilization and g-linolenic productionby Mucoralean fungi. System. Appl. Microbiol. 1997, 20,165–170.
[91] Immelman,M., Du Preez, J. C., Kilian, S. G., Effect of C:Nratio on gamma-linolenic acid production by Mucor circi-nelloides grown on acetic acid. System Appl Microbiol. 1997,20, 158–164.
[92] Fei, Q., Chang, H. N., Shang, L., Choi, J., et al., The effectof volatile fatty acids as a sole carbon source on lipidaccumulation by Cryptococcus albidus for biodiesel pro-duction. Bioresour. Technol. 2011, 102, 2695–2701.
[93] Du Preez, J. C., Immelman, M., Kock, J. L. F., Kilian,S. G., The effect of acetic acid concentration on the growthand production of gamma-linolenic acid by Mucor circinel-loides CBS 203.28 in fed-batch culture. World J. Microbiol.Biotechnol. 1997, 13, 81–87.
[94] Evans, C. T., Ratledge, C., The role of the mitochondrialNADþ isocitrate dehydrogenase in lipid accumulation bythe oleaginous yeast Rhodosporidium toruloides CBS 14. Can.J. Microbiol. 1985, 31, 479–484.
[95] Boulton, C., Ratledge, C., Regulatory studies on citratesynthase in Candida 107, an oleaginous yeast. J. Gen.Microbiol. 1980, 121, 441–447.
[96] Boulton, C., Ratledge, C., Correlation of lipid accumu-lation in yeasts with possession of ATP-citrate lyase.J. Gen. Microbiol. 1981, 127, 169–176.
[97] Wynn, J. P., Hamid, A. A., Li, Y., Ratledge, C.,Biochemical events leading to the diversion of carbon intostorage lipids in the oleaginous fungi Mucor circinelloidesand Mortierella alpina. Microbiology UK 2001, 147, 2857–2864.
[98] Evans, C. T., Scragg, A. H., Ratledge, C., A comparativestudy of citrate efflux from mitochondria of oleaginous andnon-oleaginous microorganisms. Eur. J. Biochem. 1983,130, 195–204.
[99] Botham, P., Ratledge, C., A biochemical explanation forlipid accumulation in Candida 107 and other oleaginousmicroorganisms. J. Gen. Microbiol. 1979, 114, 361–375.
[100] Galiotou-Panayotou, M., Kalatzi, O., Aggelis, G.,Modelling of simultaneous production of polygalactorunaseand exopolysaccharide by Aureobasidium pullulansATHUM 2915. Antonie Van Leeuwenhoek 1998, 73, 155–162.
[101] Anastassiadis, S., Aivasidis, A., Wandrey, C., Citric acidproduction by Candida strains under intracellular nitrogenlimitation. Appl. Microbiol. Biotechnol. 2002, 60, 81–87.
[102] Papanikolaou, S., Galiotou-Panayotou, M., Chevalot, I.,Komaitis, M., et al., Influence of glucose and saturated free-fatty acid mixtures on citric acid and lipid production byYarrowia lipolytica. Curr. Microbiol. 2006, 52, 134–142.
[103] Papanikolaou, S., Chatzifragkou, A., Fakas, S., Galiotou-Panayotou, M., et al., Biosynthesis of lipids and organicacids by Yarrowia lipolytica strains cultivated on glucose.Eur. J. Lipid Sci. Technol. 2009, 111, 1221–1232.
[104] Gill, C., Hall, M., Ratledge, C., Lipid accumulation in anoleaginous yeast (Candida 107) growing on glucose insingle-stage continuous culture. Appl. Environ. Microbiol.1977, 33, 231–239.
[105] Meyer, K. H., Schweizer, E., Control of fatty-acid synthe-tase levels by exogenous long-chain fatty acids in the yeastsCandida lipolytica and Saccharomyces cerevisiae. Eur. J.Biochem. 1976, 65, 317–324.
[106] Athenstaedt, K., Daum, G., Phosphatidic acid, a key inter-mediate in lipid metabolism. Eur. J. Biochem. 1999, 266, 1–16.
[107] Mullner, H., Daum, G., Dynamics of neutral lipid storagein yeast. Acta Biochim. Polonica 2004, 51, 323–347.
[108] Guo, X., Ota, Y., Incorporation of eicosapentaenoic anddocosahexaenoic acids by a yeast (FO726A). J. Appl.Microbiol. 2000, 89, 107–115.
[109] Wu, S., Zhao, X., Shen, H., Wang, Q., Zhao, Z. B.,Microbial lipid production by Rhodosporidium toruloidesunder sulfate-limited conditions. Bioresour. Technol. 2011,102, 1803–1807.
[110] Smit, H., Ykema, A., Verbree, E. C., Verwoert, I. I. G. S.,Kater, M. M., Producstion of cocoa butter equivalents byyeast mutants, in: Kyle, D. J., Ratledge, C. (Eds.), IndustrialProduction of Single Cell Oils, AOCS Press, Champaign(USA) 1992, pp. 185–195.
[111] Choi, S. Y., Ryu, D. Y., Rhee, J. S., Effects of growth rateand oxygen on lipid synthesis and fatty acid composition of
Eur. J. Lipid Sci. Technol. 2011, 113, 1031–1051 The biochemistry of oleaginous yeasts 1049
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

Rhodotorula gracilis. Biotechnol. Bioeng. 1982, 14, 1165–1172.
[112] Hansson, L., Dostalek, M., Influence of cultivation con-ditions on lipid production by Cryptococcus albidus. Appl.Microbiol. Biotechnol. 1986, 24, 12–18.
[113] Hassan, M., Blanc, P. J., Granger, L. M., Pareilleux, A.,Goma, G., Lipid production by an unsaturated fatty acidauxotroph of the oleaginous yeast Apiotrichum curvatumgrown in single-stage continuous culture. Appl. Microbiol.Biotechnol. 1993, 40, 483–488.
[114] Hassan, M., Blanc, P. J., Pareilleux, A., Goma, G.,Selection of fatty acid auxotrophs from the oleaginousyeast Cryptococcus curvatus and production of cocoa-butterequivalents in batch mode. Biotechnol. Lett. 1994, 16, 819–824.
[115] Hu, C., Zhao, X., Zhao, J., Wu, S., Zhao, Z. B., Effects ofbiomass hydrolysis by-products on oleaginous yeastRhodosporidium toruloides. Bioresour. Technol. 2009, 100,4843–4847.
[116] Zhao, C. H., Zhang, T., Li, M., Chi, Z. M., Single cell oilproduction from hydrolysates of inulin and extract of tubersof Jerusalem artichoke by Rhodotorula mucilaginosa TJY15a.Proc. Biochem. 2010, 45, 1121–1126.
[117] Gierhart, D. I., US Patent 4485 173, 1984.
[118] Beopoulos, A., Mrozova, Z., Thevenieau, F., Le Dall,M. T., et al., Control of lipid accumulation in the yeastYarrowia lipolytica. Appl. Environ. Microbiol. 2008, 74,7779–7789.
[119] Glatz, B. A., Hammond, E. G., Hsu, K. H., Baehman, L.,et al., Production and modification of fats and oils byyeast fermentation, in: Ratledge, C., Rattray, J. B. M.,Dawson, P. S. S. (Eds.), Biotechnology for the Oils andFats Industry, AOCS Press, Champaign (USA) 1984,pp. 163–176.
[120] Montet, D., Ratomahenina, R., Galzy, P., Pina,M., Graille,J., A study of the influence of the growth media on the fattyacid composition in Candida lipolytica DIDDENS andLODDER. Biotechnol. Lett. 1985, 7, 733–736.
[121] Lee, I. M., Hammond, E. G., Cornette, J. L., Glatz, B. A.,Triacylglycerol assembly from binary mixtures of fattyacids by Apiotrichum curvatum. Lipids 1993, 28, 1055–1061.
[122] Aggelis, G., Papadiotis, G., Komaitis, M., Microbial fattyacid specificity. Folia Microbiol. 1997, 42, 117–120.
[123] Aoki, H., Miyamoto, N., Furuya, Y., Mankura, M., et al.,Incorporation and accumulation of docosahexaenoic acidfrom the medium by Pichia methanolica HA-32. Biosci.Biotechnol. Biochem. 2002, 66, 2632–2638.
[124] Papanikolaou, S., Aggelis, G., Modeling lipid accumulationand degradation in Yarrowia lipolytica cultivated on indus-trial fats. Curr. Microbiol. 2003, 46, 398–402.
[125] Papanikolaou, S., Aggelis, G., Selective uptake of fatty acidsby the yeast Yarrowia lipolytica. Eur. J. Lipid Sci. Technol.2003, 105, 651–655.
[126] Mlickova, K., Luo, Y., D’Andrea, S., Pec, P., et al., Acyl-CoA oxidase, a key step for lipid accumulation in the yeastYarrowia lipolytica. J. Mol. Catal. B: Enzym. 2004, 28, 81–85.
[127] Mlickova, K., Roux, E., Athenstaedt, K., D’Andrea, S.,et al., Lipid accumulation, lipid body formation, andacyl-coenzyme A oxidases of the yeast Yarrowia lipolytica.Appl. Environ. Microbiol. 2004, 70, 3918–3924.
[128] Najjar, A., Robert, S., Guerin, C., Violet-Asther, M.,Carriere, F., Quantitative study of lipase secretion, extra-cellular lipolysis, and lipid storage in the yeast Yarrowialipolytica grown in the presence of olive oil: Analogies withlipolysis in humans. Appl. Microbiol. Biotechnol. 2011, 89,1947–1962.
[129] Aggelis, G., Komaitis, M., Papanikolaou, S., Papadopoulos,G., Amathematicalmodel for the study of lipid accumulationin oleaginous microorganisms. I. Lipid accumulation duringgrowth of Mucor circinelloides CBS 172-27 on a vegetable oil.Gracas y Aceites 1995, 46, 169–173.
[130] Aggelis, G., Komaitis, M., Papanikolaou, S.,Papadopoulos, G., A mathematical model for the studyof lipid accumulation in oleaginous microorganisms. II.Study of cellular lipids of Mucor circinelloides during growthon a vegetable oil. Gracas y Aceites 1995, 46, 245–250.
[131] Papanikolaou, S., Chevalot, I., Galiotou-Panayotou, M.,Komaitis, M., et al., Industrial derivative of tallow: A prom-ising renewable substrate for microbial lipid, single-cellprotein and lipase production by Yarrowia lipolytica.Electron. J. Biotechnol. 2007, 10, 425–435.
[132] Sabirova, J. S., Haddouche, R., Van Bogaert, I. N., Mulaa,F., et al., The ‘‘LipoYeasts’’ project: Using the oleaginousyeast Yarrowia lipolytica in combination with specific bac-terial genes for the bioconversion of lipids, fats and oils intohigh-value products. Microb. Biotechnol. 2010, 21, 47–54.
[133] Papanikolaou, S., Muniglia, L., Chevalot, I., Aggelis, G.,Marc, I., Yarrowia lipolytica as a potential producer of citricacid from raw glycerol. J. Appl. Microbiol. 2002, 92, 737–744.
[134] Aggelis, G., Sourdis, J., Prediction of lipid accumulation –degradation in oleaginous micro-organisms growing on veg-etable oils. Antonie Van Leeuwenhoek 1997, 72, 159–165.
[135] Fickers, P., Benetti, P. H., Wache, Y., Marty, A., et al.,Hydrophobic substrate utilisation by the yeast Yarrowialipolytica, and its potential applications. FEMS Yeast Res.2005, 5, 527–543.
[136] Wang, H.-J., Le Dall, M. T., Wache, Y., Laroche, C., et al.,Evaluation of acyl coenzyme A oxidase (Aox) isozyme func-tion in the n-alkane-assimilating yeast Yarrowia lipolytica.J. Bacteriol. 1999, 181, 5140–5148.
[137] Pignede, G., Wang, H.-J., Fudalej, F., Seman, M., et al.,Autocloning and amplification of LIP2 in Yarrowia lipoly-tica. Appl. Environ. Microbiol. 2000, 66, 3283–3289.
[138] Mauersberger, S., Wang, H.-J., Gaillardin, C., Barth, G.,Nicaud, J. M., Insertional mutagenesis in the n-alkaneassimilating yeast Yarrowia lipolytica: Generation of taggedmutations in genes involved in hydrophobic substrate util-ization. J. Bacteriol. 2001, 183, 5102–5109.
[139] Luo, Y. S., Nicaud, J. M., Van Veldhoven, P., Chardot, T.,The acyl-CoA oxidases from the yeast Yarrowia lipolytica:Characterization of Aox2p. Arch. Biochem. Biophys. 2002,407, 32–38.
[140] Fickers, P., Fudalej, F., Nicaud, J. M., Destain, J., Thonart,P., Selection of new over-producing derivatives for theimprovement of extracellular lipase production by thenon-conventional yeast Yarrowia lipolytica. J. Biotechnol.2005, 115, 379–386.
[141] Gierhart, D. I., US Patent 4485 172, 1984.
[142] Kamzolova, S. V., Finogenova, T., Morgunov, I. G.,Microbiological production of citric and isocitric acids fromsunflower oil. Food Technol. Biotechnol. 2008, 46, 51–59.
1050 S. Papanikolaou and G. Aggelis Eur. J. Lipid Sci. Technol. 2011, 113, 1031–1051
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

[143] Papanikolaou, S., Dimou, A., Fakas, S., Diamantopoulou,P., et al., Biotechnological conversion of waste cooking oliveoil into lipid-rich biomass using Aspergillus and Penicilliumstrains. J. Appl. Microbiol. 2011, 110, 1138–1150.
[144] Holdsworth, J. E., Ratledge, C., Lipid turnover in oleagi-nous yeasts. J. Gen. Microbiol. 1988, 134, 339–346.
[145] Athenstaedt, K., Zweytick, D., Jandrositz, A., Kohlwein,S. D., Daum, G., Identification and characterization ofmajor lipid particle proteins of the yeast Saccharomycescerevisiae. J. Bacteriol. 1999, 181, 6441–6448.
[146] Alvarez, H., Kalscheuer, R., Steinbuchel, A., Accumulationand mobilization of storage lipids by Rhodococcus opacusPD630 and Rhodococcus ruber NCIMB 40126. Appl.Microbiol. Biotechnol. 2000, 54, 218–223.
[147] Waltermann, M., Luftmann, H., Baumeister, D.,Kalscheuer, R., Steinbuchel, A., Rhodococcus opacus strainPD630 as a new source of high-value single-cell oil?Isolation and characterization of triacylglycerols and otherstorage lipids. Microbiology UK 2000, 146, 1143–1149.
[148] Holdsworth, J. E., Veenhuis, M., Ratledge, C., Enzymeactivities in oleaginous yeasts accumulating and utilizingexogenous or endogenous lipids. J. Gen. Microbiol. 1988,134, 2907–2915.
[149] Kamzolova, S. V., Yusupova, A. I., Dedyukhina,E. G., Chistyakova, T. I., et al., Succinic acid synthesisby ethanol-grown yeasts. Food Technol. Biotechnol. 2009, 47,144–152.
[150] Wynn, J. P., Ratledge, C., Evidence that the rate-limitingstep for the biosynthesis of arachidonic acid in Mortierellaalpina is at the level of the 18:3 to 20:3 elongase.Microbiology UK 2000, 146, 2325–2331.
[151] Pembroke, T., The yeast Yarrowia lipolytica as a model forobesity. Nat. Inst. Health. Sci. Res. Bull. 2006, 3, 11–13.
[152] Roux-Van der Merwe, M. P., Badenhorst, J., Britz, T. J.,Fungal treatment of an edible-oil-containing industrialeffluent. World J. Microbiol. Biotechnol 2005, 21, 947–953.
[153] Tang, W., Zhang, S., Wang, Q., Tan, H., Zhao, Z. B., Theisocitrate dehydrogenase gene of oleaginous yeast Lipomycesstarkeyi is linked to lipid accumulation. Can. J. Microbiol.2009, 55, 1062–1069.
[154] Liu, H., Zhao, X., Wang, F., Jiang, X., et al., The proteomeanalysis of oleaginous yeast Lipomyces starkeyi. FEMS YeastRes. 2011, 11, 42–51.
[155] Papanikolaou, S., Aggelis, G., Lipids of oleaginousyeasts. Part II: Technology and potential applications.Eur. J. Lipid. Sci. Technol. 2011, 1052–1073.
Eur. J. Lipid Sci. Technol. 2011, 113, 1031–1051 The biochemistry of oleaginous yeasts 1051
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com




















