
Leader switching occurs during the rescue of defective RNAs by heterologous strains of the...
11
Journal of General Virology (2000), 81, 791–801. Printed in Great Britain ................................................................................................................................................................................................................................................................................... Leader switching occurs during the rescue of defective RNAs by heterologous strains of the coronavirus infectious bronchitis virus Kathleen Stirrups,† Kathleen Shaw, Sharon Evans, Kevin Dalton,‡ David Cavanagh and Paul Britton Division of Molecular Biology, Institute for Animal Health, Compton Laboratory, Compton, Newbury, Berkshire RG20 7NN, UK A defective RNA (D-RNA), CD-61, derived from the Beaudette strain of the avian coronavirus infectious bronchitis virus (IBV), was rescued (replicated and packaged) using four heterologous strains of IBV as helper virus. Sequence analysis of the genomic RNA from the four heterologous IBV strains (M41, H120, HV10 and D207) identified nucleotide differences of up to 17 % within the leader sequence and up to 4–3% within the whole of the adjacent 5H untranslated region (UTR). Analysis of the 5H ends of the rescued D-RNAs showed that the Beaudette leader sequence, present on the initial CD-61, had been replaced with the corresponding leader sequence from the helper IBV strain but the adjacent 5H UTR sequence of the rescued D-RNAs corresponded to the original CD-61 Beaudette sequence. These results demonstrated that the phenomenon of leader switching previously identified for the coronaviruses murine hepatitis virus and bovine coronavirus (BCoV) also occurred during the replication of IBV D-RNAs. Three predicted stem–loop structures were identified within the 5H UTR of IBV. Stem–loop I showed a high degree of covariance amongst the IBV strains providing phylogenetic evidence that this structure exists and is potentially involved in replication, supporting previous observations that a BCoV stem–loop homologue was essential for replication of BCoV defective interfering RNAs. Introduction Avian infectious bronchitis virus (IBV) is a member of the genus Coronavirus (order Nidovirales, family Coronaviridae). IBV primarily grows in the epithelial cells of the respiratory tract causing respiratory disease and retarded growth with reduced egg production, making it the most important poultry respiratory pathogen. Coronaviruses are enveloped RNA viruses with un- segmented single-stranded, positive-sense RNA genomes of 27–32 kb that are 5« capped and 3« polyadenylated (Siddell, 1995 ; de Vries et al., 1997 ; Lai & Cavanagh, 1997). The 5« two- thirds of coronavirus genomes encode the polymerase gene Author for correspondence : Paul Britton. Fax ›44 1635 577263. e-mail paul.britton!bbsrc.ac.uk † Present address : University of Cambridge, Department of Haematology, Division of Transfusion Medicine, Long Road, Cambridge CB2 2PT, UK. ‡ Present address : Departments of Pathology and Cell Biology (BML 342), Yale University School of Medicine, 310 Cedar St, New Haven, CT 06510, USA. producing two polyproteins, Pol1a and Pol1ab, the latter resulting from a fi1 frameshift. During coronavirus replication a3«-coterminal nested set of subgenomic mRNAs is syn- thesized that encodes other virus proteins. Each subgenomic mRNA has a short non-translated leader sequence (60–90 nt), derived from the 5« end of the genome by a discontinuous process. IBV has a genome of 27 608 nt (Boursnell et al., 1987) which produces six subgenomic mRNAs, each containing a 64 nt leader sequence. Two models have been proposed for the discontinuous mechanism for the addition of coronavirus leader sequences, leader-primed transcription and discontinuous transcription during negative-strand synthesis (Sawicki & Sawicki, 1990 ; van der Most & Spaan, 1995; Brian & Spaan, 1997; Lai & Cavanagh, 1997). Both models require the presence of a consensus sequence, which we previously termed the transcription- associated sequence (TAS ; Hiscox et al., 1995), on the genomic sequence, required for leader sequence acquisition during synthesis of subgenomic mRNAs. Following high multiplicity passage of coronaviruses, defective RNAs (D-RNAs) or in some cases defective in- terfering RNAs (DI-RNAs), have been found to be produced ; they lack large internal parts of the genome, have complete 5« 0001-6708 # 2000 SGM HJB
Transcript of Leader switching occurs during the rescue of defective RNAs by heterologous strains of the...

Journal of General Virology (2000), 81, 791–801. Printed in Great Britain. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Leader switching occurs during the rescue of defective RNAsby heterologous strains of the coronavirus infectiousbronchitis virus
Kathleen Stirrups,† Kathleen Shaw, Sharon Evans, Kevin Dalton,‡ David Cavanagh and Paul Britton
Division of Molecular Biology, Institute for Animal Health, Compton Laboratory, Compton, Newbury, Berkshire RG20 7NN, UK
A defective RNA (D-RNA), CD-61, derived from the Beaudette strain of the avian coronavirusinfectious bronchitis virus (IBV), was rescued (replicated and packaged) using four heterologousstrains of IBV as helper virus. Sequence analysis of the genomic RNA from the four heterologousIBV strains (M41, H120, HV10 and D207) identified nucleotide differences of up to 17% within theleader sequence and up to 4±3% within the whole of the adjacent 5H untranslated region (UTR).Analysis of the 5H ends of the rescued D-RNAs showed that the Beaudette leader sequence, presenton the initial CD-61, had been replaced with the corresponding leader sequence from the helperIBV strain but the adjacent 5H UTR sequence of the rescued D-RNAs corresponded to the originalCD-61 Beaudette sequence. These results demonstrated that the phenomenon of leader switchingpreviously identified for the coronaviruses murine hepatitis virus and bovine coronavirus (BCoV)also occurred during the replication of IBV D-RNAs. Three predicted stem–loop structures wereidentified within the 5H UTR of IBV. Stem–loop I showed a high degree of covariance amongst theIBV strains providing phylogenetic evidence that this structure exists and is potentially involved inreplication, supporting previous observations that a BCoV stem–loop homologue was essential forreplication of BCoV defective interfering RNAs.
IntroductionAvian infectious bronchitis virus (IBV) is a member of the
genus Coronavirus (order Nidovirales, family Coronaviridae). IBVprimarily grows in the epithelial cells of the respiratory tractcausing respiratory disease and retarded growth with reducedegg production, making it the most important poultryrespiratory pathogen.
Coronaviruses are enveloped RNA viruses with un-segmented single-stranded, positive-sense RNA genomes of27–32 kb that are 5« capped and 3« polyadenylated (Siddell,1995 ; de Vries et al., 1997 ; Lai & Cavanagh, 1997). The 5« two-thirds of coronavirus genomes encode the polymerase gene
Author for correspondence: Paul Britton.
Fax 44 1635 577263. e-mail paul.britton!bbsrc.ac.uk
† Present address: University of Cambridge, Department of
Haematology, Division of Transfusion Medicine, Long Road, Cambridge
CB2 2PT, UK.
‡ Present address: Departments of Pathology and Cell Biology (BML
342), Yale University School of Medicine, 310 Cedar St, New Haven,
CT 06510, USA.
producing two polyproteins, Pol1a and Pol1ab, the latterresulting from a ®1 frameshift. During coronavirus replicationa 3«-coterminal nested set of subgenomic mRNAs is syn-thesized that encodes other virus proteins. Each subgenomicmRNA has a short non-translated leader sequence (60–90 nt),derived from the 5« end of the genome by a discontinuousprocess. IBV has a genome of 27608 nt (Boursnell et al., 1987)which produces six subgenomic mRNAs, each containing a64 nt leader sequence.
Two models have been proposed for the discontinuousmechanism for the addition of coronavirus leader sequences,leader-primed transcription and discontinuous transcription duringnegative-strand synthesis (Sawicki & Sawicki, 1990 ; van derMost & Spaan, 1995 ; Brian & Spaan, 1997 ; Lai & Cavanagh,1997). Both models require the presence of a consensussequence, which we previously termed the transcription-associated sequence (TAS; Hiscox et al., 1995), on the genomicsequence, required for leader sequence acquisition duringsynthesis of subgenomic mRNAs.
Following high multiplicity passage of coronaviruses,defective RNAs (D-RNAs) or in some cases defective in-terfering RNAs (DI-RNAs), have been found to be produced ;they lack large internal parts of the genome, have complete 5«
0001-6708 # 2000 SGM HJB

K. Stirrups and othersK. Stirrups and others
and 3« UTRs and require helper virus for replication. Theinternal deletions result in the fusion of different regions of thepolymerase gene and varying amounts of the N gene (Makinoet al., 1988b, 1990 ; van der Most et al., 1991 ; Chang et al.,1994 ; Pe! nzes et al., 1994 ; Mendez et al., 1996).
Makino et al. (1986) demonstrated that leader sequencesfrom different murine hepatitis virus (MHV) strains can beexchanged during a mixed infection, indicating that there issome interaction of the MHV polymerase between theheterologous RNAs during replication. Initial work by Makinoet al. (1988a) to determine whether leader sequences can beswitched between helper virus and an MHV DI-RNA failed todemonstrate this event using naturally occurring MHV JHM-derived DI-RNAs rescued with MHV A59. All the naturallyoccurring MHV JHM-derived DI-RNAs had a 9 nt deletion,UUUAUAAAC, downstream of the leader sequence. Re-placement of the 9 nt sequence in an MHV JHM-derived DI-RNA, DIssE, resulted in a modified DI-RNA, DE5-w3, that wasable to leader switch following rescue with MHV A59 (Makino& Lai, 1989). This phenomenon of an MHV DI-RNA acquiringthe leader sequence of the helper virus was termed leaderswitching and was believed to be the result of a high-frequencyrecombination event (Makino & Lai, 1989). The secondcoronavirus system from which D-RNAs were isolatedinvolved the Mebus strain of bovine coronavirus (BCoV;Hofmann et al., 1990). A synthetic BCoV DI-RNA, derivedfrom a naturally occurring Mebus DI-RNA, was modified bythe introduction of specific point mutations within the leadersequence to determine their potential effect(s) on replication ofthe DI-RNA (Chang et al., 1994). All the mutations appeared toaffect replication only minimally, following rescue withhomologous Mebus helper virus. However, analysis of theleader sequences on the rescued D-RNAs showed that theyhad all reverted to the wild-type, Mebus, leader sequence(Chang et al., 1994).
In a recent review (Brian & Spaan, 1997) the authorsproposed that an AU-rich region, immediately downstream ofthe leader junction sequence at the 5« ends of BCoV and MHVDI-RNAs, is involved in leader switching. The authors alsoindicated that a similar AU-rich region is also presentdownstream of the leader junction sequence on DI-RNAs ofthe porcine coronavirus transmissible gastroenteritis virus(TGEV; Mendez et al., 1996) but not on D-RNAs of IBV(Pe! nzes et al., 1994). These observations led the authors to raisethe question as to whether leader switching could occur on IBVD-RNAs. At that time the phenomenon of leader switchinghad not been demonstrated for DI-RNAs from TGEV or D-RNAs from IBV.
Methods+ Virus and cells. IBV strains Beaudette, M41, HV10 and H120(Darbyshire et al., 1979) and D207 (Davelaar et al., 1984) were adapted forgrowth in primary chick kidney (CK) cells and stocks made in 11-day-oldembryonated domestic fowl eggs at 37 °C [harvested from allantoic fluid
at 20 h post-infection (p.i.)] or in CK cells. RNA derived from all five IBVstrains was analysed for the presence of endogenous D-RNAs and foundto be D-RNA free and was therefore used for the rescue of IBV D-RNACD-61 (Pe! nzes et al., 1996). The passage and growth of IBV in CK cellswas as described previously (Pe! nzes et al., 1994).
+ Oligonucleotides. The oligonucleotides used in this study wereobtained from Cruachem or MWG-Biotech and are listed in Table 1.
+ Recombinant DNA techniques. Procedures for recombinantDNA techniques were either standard (Ausubel et al., 1987 ; Sambrook etal., 1989) or according to manufacturers« instructions. RT–PCRs andPCRs were as described previously (Pe! nzes et al., 1994, 1996). Sequencedetermination of plasmid DNA and PCR products was as describedpreviously (Pe! nzes et al., 1994 ; Hiscox et al., 1995).
+ Sequences analysis of the 5« end of IBV genomic RNA. IBVgenomic RNA, from strains M41, HV10, H120 or D207, was extractedfrom virions grown in embryonated eggs (Kottier et al., 1995). A slightlymodified version of the cRACE method (Maruyama et al., 1995) was usedto determine the very 5« ends of the IBV genomic RNAs. Essentially,1 µg of each viral RNA was copied into single-stranded cDNAs (402 nt)using oligonucleotide K5UTR-2 and Superscript II RNase H− reversetranscriptase (200 U; GibcoBRL) at 47 °C for 2 h. The RNA–DNAhybrids were incubated with RNase H (4 U; GibcoBRL) at 37 °C for20 min and the cDNAs purified on GLASSMAX (GibcoBRL) spincolumns. The cDNAs were eluted using 50 µl H
#O at 65 °C and
circularized using T4 RNA ligase (1 U; USB) in 50 mM Tris–HCl (pH7±5), 10 mM MgCl
#, 10 mM DTT, 1 mM ATP and 60 µg}ml BSA at
17 °C for 18 h. The reaction was terminated by the addition of EDTA toa final concentration of 20 mM. The circularized cDNAs were used astemplates for PCRs across the ligated ends using oligonucleotides93}137 and 18A with Pfu DNA polymerase (Stratagene). The PCRproducts (280 bp) were sequenced, using oligonucleotide 93}137, eitherdirectly or following ligation into EcoRV-digested pBluescript SK fromwhich several IBV-derived cDNAs were sequenced. This methodproduced sequence data corresponding to nt 1–151 of the genomicRNAs. To determine sequence data beyond nt 151 each viral RNA wascopied into cDNA by RT–PCR using Pfu DNA polymerase (Stratagene).Different oligonucleotide pairs were utilized depending on the IBV strain,i.e. oligonucleotides 4293}102 and 94}155K5UTR-3 were used forHV10; 5deg-ve43 for H120 ; and 93}10193}102 and 94}155K5UTR-3 for D207. The PCR products were sequenced either directly orfrom cloned DNA from which several IBV-derived cDNAs weresequenced.
+ In vitro transcription and electroporation of primary CKcells. RNA corresponding to IBV D-RNA CD-61 was synthesized invitro from 2 µg NotI-linearized pCD-61 (Pe! nzes et al., 1996) in a totalvolume of 40 µl. This was done using 40 U T7 RNA polymerase(Promega) in 40 mM Tris–HCl (pH 7±5), 6 mM MgCl
#, 2 mM sperm-
idine, 10 mM NaCl, 30 nmol each of ATP, UTP, CTP and GTP, 400 nmolDTT, 40 U RNasin (Promega) at 37 °C for 2 h. CK cells were grown to80–90% confluence in 25 cm# tissue culture flasks (Falcon) and infectedwith 0±5 ml egg-grown IBV helper virus or 1 ml of CK-grown D207virus. At 8 h p.i. the cells were washed twice with PBSa (172 mM NaCl,3 mM KCl, 100 mM Na
#HPO
%, 2 mM KH
#PO
%, pH 7±2), once with
trypsin–versene (0±03% trypsin in 0±5 mM EDTA). The IBV-infected cellsheets were disrupted using 1 ml trypsin–versene for 3–5 min at 37 °C,followed by addition of 0±3 ml 10% newborn calf serum, cooled on ice,centrifuged at 2500 r.p.m. for 3 min and the cells resuspended in 0±3 mlice-cold PBSa. Transcription reactions (40 µl) were mixed with theresuspended IBV-infected cells in 0±4 cm electroporation cuvettes(Invitrogen) for electroporation at 220 V using a BTX Electro Cell
HJC

Leader switching in IBV defective RNAsLeader switching in IBV defective RNAs
Table 1. Oligonucleotides used
Oligonucleotide Sequence* Position† Polarity‡ Purpose
94}155 GAAGGATCCATTAATACGACTCACTATAGGGACTTAAGATAGATATTAATATAT 1–23 RT–PCR43 GGGCCCACTTAAGATAGATATTAATATA 1–22 RT–PCR93}102 GCTGGTCCTCATAGGTGTTC 85–104 RT–PCR93}137 TGACAGGTGGCCAGGTGC 134–151 ® RT–PCR,
sequencingK5UTR-1 CTAGGTAGCCCAAGCAAC 223–240 ® Sequencing18A GGCGGATCCCGGGCGTCGTGGCTGG 255–270 RT–PCRK5UTR-2 CTCAAAAGCCCTTAGAGG 385–402 ® RT–PCR,
sequencingK5UTR-3 GTCTGCTCACTAAACACCAC 492–501 ® Sequencing93}106 GGCAGAAGTTTGACCGTAG 674–692 RT–PCR93}120 CGCGCTAAAGGAGAGACAC 833–851 ® Sequencing5deg-ve CCAYTGRCATGCTGT‡ 874–888 ® RT–PCR95}165 GGATGCTCTGGGCTCATAGTAGAGTGCTCTTTTTC 1040–1074 Sequencing42 CCTCTCCAAGCACTGCTG 1997–2014 ® RT–PCR93}101 GTCTTGCCCGGGTTGGTTTTCTGGAAGTG 2402–2420 ® RT–PCRST4 CTAGCGCAGTTACGCTTCAA 12471–12490 ® Sequencing93}136 GTCCCATTTTAGCCAACATG 12857–12876 ® RT–PCRN1145 AGAGGAACAATGCACAGCTGG 27017–27037 RT–PCR93}100 CAGGATATCGCTCTAACTCTATACTAGCCT 27587–27607 ® RT–PCR
* Underlined regions represent non-IBV sequence.† Refers to the position of the nucleotides in the Beaudette sequence (Boursnell et al., 1987).‡ Degenerate oligonucleotide, in which Y¯C or T and R¯A or G.
Fig. 1. Schematic diagram to outline the rescue of IBV D-RNAs. The CKcells (P0) were infected with helper IBV and at 8 h p.i. they were preparedfor electroporation with in vitro T7-transcribed CD-61 RNA. Theelectroporated cells were incubated at 37 °C for 16 h (to 24 h p.i.).Virus (V1) in the supernatants of the infected cells was used to infect CKcells (P1) and after 20–24 h p.i. virus (V2) in the supernatants was usedto infect CK cells (P2). This was repeated for several passages. Totalcellular RNA was extracted from the infected cells (P1–P7) for eitherNorthern blot analysis or for RT–PCR and sequence analysis.
Manipulator (model ECM-395). The electroporated IBV-infected cells
were placed on ice for 5 min, diluted 1 :10 in CK cell medium (Pe! nzes et
al., 1994), transferred onto tissue culture plates and incubated at 37 °C for
16 h. Virus (V") in 1 ml of the supernatants was used to infect CK cells
and after 20–24 h p.i. virus (V#) from the supernatants was passaged on
CK cells. This was repeated up to five more passages, V$–V
(. RNA was
isolated from cells (P"–P
() infected with each virus (V
"–V
() passage as
outlined in Fig. 1.
+ Isolation and Northern blot analysis of IBV-derived RNA.Total cellular RNA was extracted from the IBV-infected CK cells usingguanidinium isothiocyanate (Pe! nzes et al., 1994). The RNAs wereseparated in denaturing 1% agarose–2±2 M formaldehyde gels(Sambrook et al., 1989) and Northern blotted onto Hybond-C extra0±45 µm nitrocellulose membranes (Amersham). IBV-specific RNAs weredetected by hybridization to a mixture of $#P-labelled PCR fragments(590 bp) generated from the five IBV strains using oligonucleotidesN1145 and 93}100, corresponding to nt 27017–27607 at the 3« end ofthe IBV genome. The PCR fragments were labelled with [$#P]dCTP usingthe random oligonucleotide-primed synthesis method (Feinberg &Vogelstein, 1983).
+ Sequence analysis of IBV D-RNAs. Total cellular RNA extractedfrom P
(-infected CK cells containing the D-RNAs, CD-61, CD61M41,
CD61H120, CD61HV10 and CD61D207 rescued with helper IBVs,Beaudette, M41, H120, HV10 or D207, respectively, was used forRT–PCRs, using oligonucleotide 93}136 for the RT reactions. A varietyof oligonucleotides was used for the PCRs to : (a) distinguish D-RNAsfrom either genomic RNA or subgenomic mRNAs and to confirm theadenosine deletion at nt 749 (A-749) of CD-61 [a characteristic of the D-RNA (Pe! nzes et al., 1994)] and (b) to characterize the leader sequence andadjacent 5« UTR on the rescued D-RNAs. The PCRs for (a) usedoligonucleotides 93}136 and 93}106, which result in a fragment of910 bp spanning domains I and II of D-RNA CD-61 (Pe! nzes et al., 1994).Oligonucleotide 93}120 was used to sequence part of the 910 bp PCRfragment to confirm the absence of A-749. Oligonucleotides 95}165 andST4 were used to sequence the domain I}II junction within the 910 bpPCR fragment. The PCRs for (b) used oligonucleotides 93}136 and94}155, which resulted in a fragment of 1±6 kb extending from within
HJD

K. Stirrups and othersK. Stirrups and others
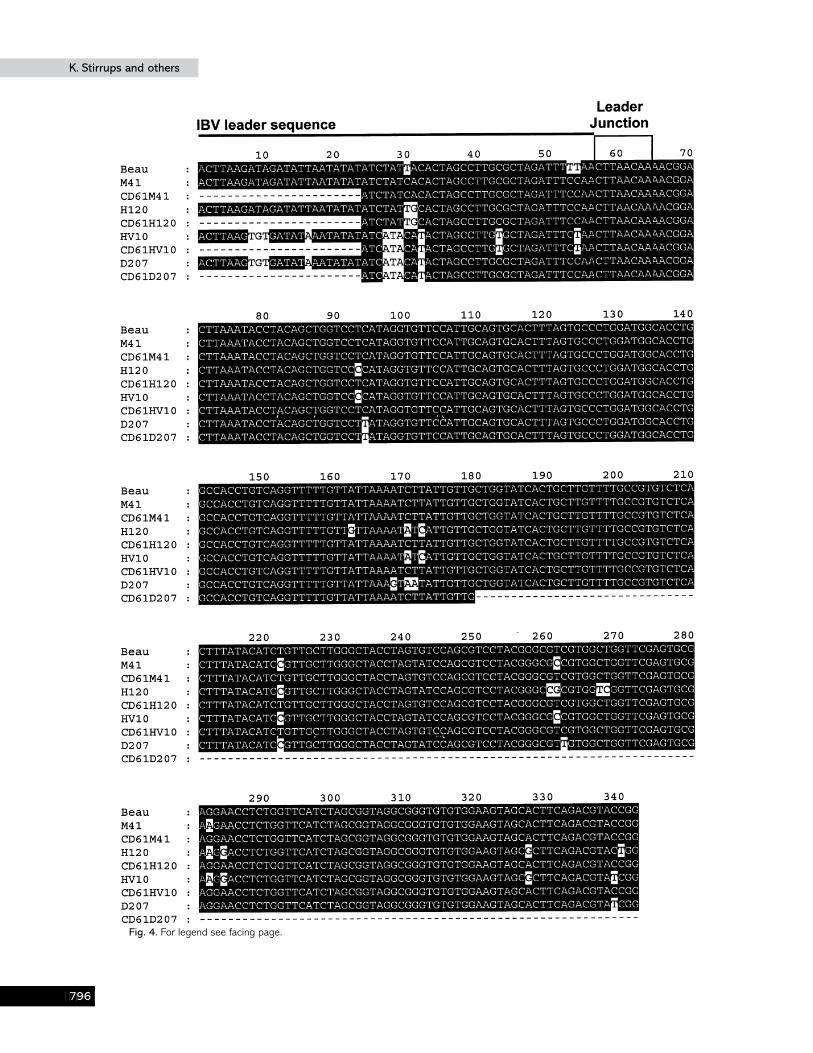
Fig. 2. Alignment of the first 376 nt from cDNA sequences derived from the 5« end of the genomic RNA of five strains of IBV toidentify potential differences between the leader and adjacent 5« UTR sequences. The leader junction sequence, CTTAACAA, isthe point differentiating the leader and adjacent 5« UTR sequences. The figure was generated using GeneDoc in which upper-case letters in the consensus sequence, below the alignments, relate to complete conservation of the nucleotides. The level ofshading relates to the degree of conservation, i.e. 100% conservation black and less than 60 % conservation white. The leadersequence is denoted by a solid black line and the leader junction sequence is boxed.
domain II of CD-61 to nt 23 from the 5« end of D-RNA CD-61.Oligonucleotides 93}137, K5UTR-1, K5UTR-2 and K5UTR-3 were usedto sequence nt 24–342 of the rescued D-RNAs.
+ Computer analysis of sequence data. Sequence data wereassembled using Gap4 of the Staden Sequence Software Programs
(Bonfield et al., 1995). Sequences were aligned using ClustalX version
1.64b (Thompson et al., 1997) and the alignments analysed using
GeneDoc version 2.5 (Nicholas & Nicholas, 1997). Identification of
potential RNA structures was determined using RNAdraw version 1.1
(Matzura & Wennborg, 1996) and MFOLD (Zuker & Stiegler, 1981 ;
HJE
Leader switching in IBV defective RNAsLeader switching in IBV defective RNAs
Jaeger et al., 1989, 1990 ; Zuker, 1989a, b). The MFOLD program usedwas part of the Wisconsin package (GCG version 9.0 UNIX software ;Genetics Computer Group Inc., 1997) (Devereux et al., 1984). Both typesof analyses work on the basis of free-energy minimization.
ResultsSequence analysis of the 5« UTR of IBV genomes
To investigate whether the phenomenon of leaderswitching occurs during the rescue of IBV D-RNAs we decidedto use the IBV Beaudette-derived D-RNA CD-61 for rescuewith four heterologous IBV strains. This system has theadvantage over the MHV and BCoV systems, for whichcoronavirus leader switching had been previously shown,because the IBV D-RNA is unmodified within the 5« UTR.
Three of the heterologous strains, M41, HV10 and H120,belong to the same serotype, Massachusetts, as Beaudettefrom which CD-61 was derived, based on either serology ofthe spike (S) protein (Darbyshire et al., 1979) or S genesequence identity determined from Beaudette (Binns et al.,1985 ; Boursnell et al., 1987), M41 (Binns et al., 1986 ; Niesterset al., 1986) and H120 (Kusters et al., 1989). The fourthheterologous strain, D207, is of a different antigenic serotypeto the Massachusetts strains based on serology of the S protein(Davelaar et al., 1984) and S gene sequence (Kusters et al.,1989). Serotyping of IBV strains is entirely based on the Sprotein}gene and therefore any similarities between the virusesmay not be reflected in nucleotide sequences over otherregions of the genome.
To confirm whether leader switching occurs during therescue of IBV D-RNAs we needed to identify potentialnucleotide differences, within the leader and adjacent 5« UTRsequences, between the IBV strains. The potential substitutionshad to be sufficiently different to act as markers capable ofdistinguishing resultant sequences, derived from any leaderswitching events, from both the helper virus and the D-RNACD-61. Sequences (nt 1–151) of the 5« ends of the genomicRNAs were determined for the four heterologous IBV strainsusing cRACE (Maruyama et al., 1995). Further genomicsequence, following nt 151, was determined from IBV strainsHV10, H120 and D207 by RT–PCR. The genomic sequences(nt 1–376) obtained as described here and from previouslyderived sequence data, for M41 (Pe! nzes, 1995) and Beaudette(Boursnell et al., 1987), were aligned.
Comparison of the aligned sequences identified a variety ofnucleotide substitutions between the IBV sequences (Fig. 2).M41 had 3 (4±7%) and 4 (1±3%), HV10 had 11 (17±2%) and 11(3±5%), H120 had 3 (4±7%) and 19 (6±1%) and D207 had 11(17±2%) and 8 (2±6%) substitutions within the leader andadjacent 5« UTR sequences, respectively, when compared tothe Beaudette sequence. This indicated, except for strain H120,that the proportion of substitutions was higher within theleader sequence than the adjacent 5« UTR. The percentagedifferences of the complete heterologous IBV 5« UTRs (nt
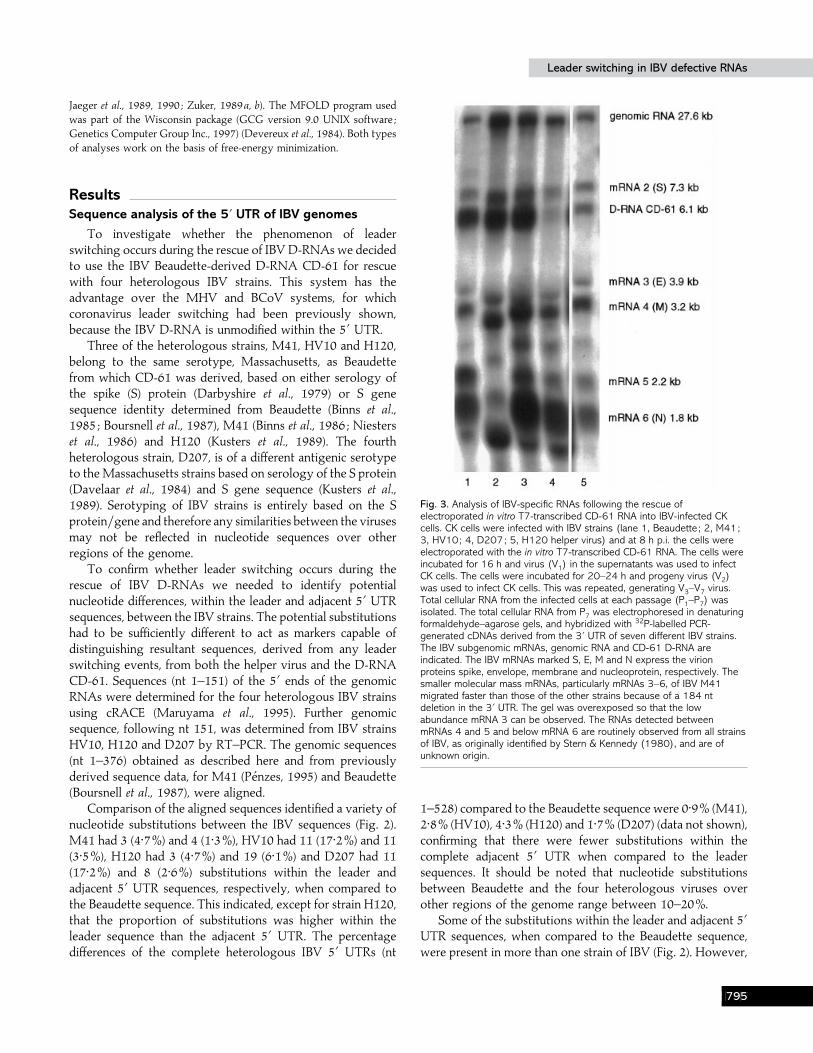
Fig. 3. Analysis of IBV-specific RNAs following the rescue ofelectroporated in vitro T7-transcribed CD-61 RNA into IBV-infected CKcells. CK cells were infected with IBV strains (lane 1, Beaudette ; 2, M41;3, HV10; 4, D207; 5, H120 helper virus) and at 8 h p.i. the cells wereelectroporated with the in vitro T7-transcribed CD-61 RNA. The cells wereincubated for 16 h and virus (V1) in the supernatants was used to infectCK cells. The cells were incubated for 20–24 h and progeny virus (V2)was used to infect CK cells. This was repeated, generating V3–V7 virus.Total cellular RNA from the infected cells at each passage (P1–P7) wasisolated. The total cellular RNA from P7 was electrophoresed in denaturingformaldehyde–agarose gels, and hybridized with 32P-labelled PCR-generated cDNAs derived from the 3« UTR of seven different IBV strains.The IBV subgenomic mRNAs, genomic RNA and CD-61 D-RNA areindicated. The IBV mRNAs marked S, E, M and N express the virionproteins spike, envelope, membrane and nucleoprotein, respectively. Thesmaller molecular mass mRNAs, particularly mRNAs 3–6, of IBV M41migrated faster than those of the other strains because of a 184 ntdeletion in the 3« UTR. The gel was overexposed so that the lowabundance mRNA 3 can be observed. The RNAs detected betweenmRNAs 4 and 5 and below mRNA 6 are routinely observed from all strainsof IBV, as originally identified by Stern & Kennedy (1980), and are ofunknown origin.
1–528) compared to the Beaudette sequence were 0±9% (M41),2±8% (HV10), 4±3% (H120) and 1±7% (D207) (data not shown),confirming that there were fewer substitutions within thecomplete adjacent 5« UTR when compared to the leadersequences. It should be noted that nucleotide substitutionsbetween Beaudette and the four heterologous viruses overother regions of the genome range between 10–20%.
Some of the substitutions within the leader and adjacent 5«UTR sequences, when compared to the Beaudette sequence,were present in more than one strain of IBV (Fig. 2). However,
HJF
K. Stirrups and othersK. Stirrups and others
Fig. 4. For legend see facing page.
HJG

Leader switching in IBV defective RNAsLeader switching in IBV defective RNAs
some of the substitutions were strain-specific, i.e. HV10, H120and D207 had 1, 10 and 4 substitutions, respectively, unique totheir particular sequence. Therefore, the substitutions presenton a particular sequence and their distribution between theleader sequence and the adjacent 5« UTR, when compared tothe Beaudette sequence, were sufficient to differentiate leaderand adjacent 5« UTR sequence combinations.
Rescue of D-RNA CD-61 using heterologous IBVhelper strains
To investigate whether the heterologous IBV strains wouldsupport replication of the Beaudette-derived CD-61 D-RNA,CK cells were infected with the heterologous strains of IBV andelectroporated with in vitro T7-transcribed CD-61 D-RNA.Progeny virus (V
") was serially passaged on CK cells,
generating progeny virus V#–V
((Fig. 1). Total cellular RNA
was extracted from CK cells (P"–P
() previously infected with
V"–V
(virus and examined by Northern blot analysis. An RNA
species, corresponding to the size of D-RNA CD-61, wasidentified in the RNA isolated from P
#–P
(CK cells infected
with the four heterologous strains. The IBV RNAs isolatedfrom P
(-infected cells are shown in Fig. 3 ; however, it should
be noted that the passage number for either initially observingrescue of the D-RNA or detecting the highest amount of D-RNA varied depending on the helper strain used (data notshown). The system was not optimized for the rescue of the D-RNA and was therefore only semi-quantitative. Hence, as canbe seen from Fig. 3, the amounts of CD-61 rescued in the P
(cells varied depending on the heterologous helper virus used,rescue efficiency being least with strain D207. We cannot ruleout the possibility of some incompatibility between thesequences involved in either replication or packaging, of theBeaudette-derived D-RNA, and those present in the helpervirus D207, resulting in a lower efficiency of rescue.
RT–PCR, using oligonucleotides 93}136 and 93}106, wasused to determine whether the 6±1 kb RNA, isolated from P
'CK cells infected with each IBV helper strain, corresponded toD-RNA CD-61. The product of the two oligonucleotidesspans the junction of domains I and II of CD-61 (Pe! nzes et al.,1996), resulting in a 910 bp DNA fragment if derived fromCD-61 or 12±2 kb from genomic RNA. RT–PCR DNAproducts of 910 bp were generated from RNA isolated from P
'CK cells infected with each heterologous helper strain,indicating that CD-61 D-RNA was present in each cell extract,
Fig. 4. Alignment of the first 342 nt from cDNA sequences derived from the 5« ends of the rescued IBV D-RNA CD-61following electroporation of in vitro synthesized T7-transcripts into CK cells infected with different strains of helper IBV.The sequences are presented along with the 5« ends derived from the genomic RNA of the viruses to compare anypotential differences in the input D-RNA, derived from the Beaudette strain, and the sequence following rescue with thedifferent heterologous strains. The first 23 nt of the D-RNA sequences are not presented as these were derived from theoligonucleotide used for the RT–PCR amplifications. The black shading represents complete conservation between the virusand D-RNA sequences and the white regions represent either differences between virus sequences or differences betweenthe input D-RNA sequence and the sequence of the rescued D-RNA. The dashes represent sequence not determined for D-RNA CD61D207. The leader sequence is denoted by a solid line and the leader junction sequence, the point differentiatingthe leader sequence and the adjacent 5« UTR, is boxed.
confirming the Northern blot data. Sequence analysis con-firmed that the 910 bp RT–PCR products had sequencesidentical to the domain I}II junction in D-RNA CD-61. Furtheranalysis confirmed that the A-749 nucleotide deletion, initiallyidentified in IBV D-RNA CD-91 (Pe! nzes et al., 1994) fromwhich CD-61 was derived (Pe! nzes et al., 1996), occurred in theD-RNAs rescued by the heterologous helper viruses. Theseresults confirmed that the 6±1 kb RNA, rescued by all fourheterologous IBV strains, represented D-RNA CD-61, despitethe observed nucleotide substitutions within the leader andadjacent 5« UTR sequences.
Analysis of D-RNA CD-61 leader sequences followingrescue by heterologous helper viruses
RNA was isolated from P'
CK cells and analysed byRT–PCR using oligonucleotides 93}136 and 94}155. Oligo-nucleotide 94}155 corresponded to nt 1–23 of the Beaudettegenome. The 1±59 kb RT–PCR products represented sequencesderived from the rescued D-RNAs from nt 24 in the leadersequence to within domain II.
Sequences representing nt 24–64 of the leader and nt65–342 of the adjacent 5« UTR sequences were determinedfrom the 1±59 kb RT–PCR products. The 318 nt sequences,derived from D-RNAs CD61M41, CD61H120, CD61HV10and CD61D207, rescued with the heterologous IBV strainsM41, H120, HV10 and D207, respectively, were compared tothe corresponding genomic and Beaudette sequences. Theresults showed that all the rescued D-RNAs had leadersequences corresponding to the specific helper virus and notBeaudette-derived CD-61 sequence (Fig. 4). In contrast, nt65–342 of the adjacent 5« UTR sequences corresponded toBeaudette sequence and were therefore derived from CD-61(Fig. 4). The only exception was that D-RNA CD61D207,rescued with D207, had a thymidine residue at nt 94, whichcorresponded to D207 genomic sequence, and not a cytosineresidue corresponding to Beaudette sequence. None of theD207-specific nucleotide substitutions, downstream of nt 94 inthe D207 genomic sequence, were present in the rescuedCD61D207 sequence. Nucleotide 94 is the first substitution inthe adjacent 5« UTR sequence of the D207 genomic sequence30 nt from the leader junction sequence.
Our results demonstrated that leader switching occurredduring rescue of CD-61 by the four heterologous IBV strains.The exact crossover points for leader switching could not be
HJH

K. Stirrups and othersK. Stirrups and others
Fig. 5. Predicted RNA secondary structure of the IBV 5«-terminal region (nt 1–100) based on the Beaudette sequence.Stem–loops I, II and III are identified. The thin arrows indicate the positions of the nucleotide substitutions identified withinthe 5« UTRs of the heterologous IBV strains, M41, HV10, H120 and D207 (indicated in parentheses), used in the rescue ofD-RNA CD-61. The start of the leader junction sequence is indicated with a thick arrow and the consensus sequence isboxed. The dashed line in stem–loop I indicates the extra base paired nucleotides identified in IBV strains D207 and HV10resulting from the nucleotide substitution of a U !A at nt 16.
determined from the available nucleotide substitutions in theIBV strains used. However, IBV strains M41, H120 and D207have a substitution at nt 54 when compared to the Beaudettesequence, indicating that the left-hand boundary of thecrossover region was 2 nt proximal to the leader junctionsequence, CUUAACAA. The right-hand boundary of thecrossover region can be determined for H120 and HV10rescued D-RNAs due to the substitution at nt 93, 28 nt distalto the leader junction sequence. Overall, the observed 3«boundaries of the crossover region are at nt 93 for H120 andHV10, nt 168 for D207 and nt 222 for M41 (Fig. 4) ; however,the actual crossover points could be further upstream.
Identification of potential RNA secondary structures inthe IBV 5« UTRs.
Computer analysis of potential RNA secondary structureswithin the leader and adjacent 5« UTR genomic sequences (nt1–100) of the five IBV strains was carried out. Three stem–loopstructures were predicted with thermodynamic stabilities of®6±2 to ®8±8 kcal}mol for stem–loop I, ®3±94 kcal}mol forstem–loop II and ®3±74 kcal}mol for stem–loop III (1 cal¯4±184 J) (Fig. 5).
Comparison of the predicted secondary structures showedthat the various nucleotide substitutions identified did not alterthe overall predicted structures. Most of the nucleotidesubstitutions occurred within predicted single-strandedregions. However, 7 nt substitutions in D207 and HV10occurred in stem–loop I. Six of the seven nucleotide sub-stitutions showed covariance maintaining the predicted stem–loop structure, providing phylogenetic evidence for thestructure. The seventh nucleotide substitution occurred at nt30 (U!C) to produce a G–C rather than a G–U base pair.This nucleotide substitution also occurred in theM41 sequence,accounting for the increased thermodynamic stability ofstem–loop I for M41 (®8±5 kcal}mol) when compared toBeaudette (®6±2 kcal}mol) and H120 (®6±2 kcal}mol), poss-ibly indicating that this substitution and resultant base pairingmay incur some advantage. Nucleotide substitution at nt 16 (U!A) in HV10 and D207 resulted in a further base pairingwithin stem–loop I (Fig. 5), decreasing the number ofnucleotides within the predicted loop region. This nucleotidesubstitution, possibly together with the nucleotide substitutionat nt 30, counteracts the potential destabilizing effect of thecovariant U–G base pair (nt 9 and 28) resulting from thesenucleotide substitutions in HV10 and D207. The overall result
HJI

Leader switching in IBV defective RNAsLeader switching in IBV defective RNAs
of the nucleotide substitutions in the predicted stem–loop I ofHV10 and D207 was an increased thermodynamic stability,®8±8 kcal}mol, when compared to the predicted structure forBeaudette and H120, and more similar to that found for M41.The leader junction sequence is predicted to occur in a single-stranded region between stem–loops II and III (Fig. 5).Stem–loop III is predicted to occur 4 nt distal to the leaderjunction sequence and probably represents the stem–loopstructure identified in the 5« UTR of BCoV, and subsequentlyfor MHV and TGEV, that has been suggested to contributetowards template switching (Chang et al., 1996).
DiscussionThis study reports the phenomenon of leader switching
during the replication of the IBV Beaudette-derived D-RNACD-61 following rescue with four heterologous helper IBVs.This is the first description of leader switching for IBV and thefirst report of leader switching using a coronavirus D-RNAunmodified in the 5« UTR. Our results also answer the recentquery by Brian & Spaan (1997) as to whether IBV D-RNAscan undergo leader switching.
Coronavirus leader switching was first demonstrated tooccur during the replication of an MHV JHM-derived syntheticDI-RNA modified by the inclusion of a 9 nt sequence, deletedfrom all naturally occurring MHV JHM DI-RNAs, adjacent tothe leader sequence. The second report of a coronavirus D-RNA undergoing leader switching was for BCoV using asynthetic DI-RNA derived from the Mebus strain of BCoV(Chang et al., 1994). Leader switching, referred to as leaderreversion by the authors, was observed to occur followingexperiments designed to investigate the potential role ofvarious nucleotides on the replication of the D-RNA. A seriesof point mutations was introduced into the leader sequence ofthe D-RNA but no potential effect of the mutations could bedetermined because the replicated D-RNAs were found tohave leader sequences identical to the helper virus. The BCoVsystem differed from the MHV system in that homologoushelper virus was used whereas the MHV system usedheterologous helper virus.
Chang et al. (1996) showed that, unlike the acquisition ofthe leader sequence onto coronavirus subgenomic mRNAs,leader switching does not require a complete leader junctionsequence. BCoV DI-RNAs containing either a scrambled or adeleted leader junction sequence were replicated, the modifiedDI-RNAs re-acquiring leader junction sequences as a result ofleader switching. The authors concluded that leader switchinginvolves a recombination event over a region of 24 ntcomprising the last base of the leader junction sequence andthe adjacent 23 nt. Analysis of IBV D-RNAs rescued withheterologous strains indicated that the boundaries of thecrossover region were nt 55 and 95, the latter 31 ntdownstream of the leader junction sequence. Whilst we cannotrule out the possibility that the IBV leader junction sequence is
involved in leader switching it would appear from the work ofChang et al. (1996) that the event may take place downstreamof the IBV leader junction sequence, potentially involving nt64–95.
The BCoV crossover region contains an 8 nt palindromicAU-rich sequence, UUUAUAAA, comprising 8 nt of the 9 ntdeleted from the MHV DI-RNAs that did not undergo leaderswitching (Makino et al., 1988a). No such AU-rich sequence ispresent in any of the five IBV genomic sequences. Absence ofthe AU-rich sequence in the Beaudette sequence led Brian &Spaan (1997) to query whether IBV D-RNAs could undergoleader switching. Interestingly, the five IBV strains have thesequence AAAUACCU (nt 74–81), reminiscent of the pal-indromic sequence, AAAUAUUU, found in the TGEV 5« UTRsequence. It should be noted that the IBV AAAUACCUsequence is not adjacent but 9 nt downstream of the leaderjunction sequence whereas the AU-rich sequence in BCV,MHV and TGEV is adjacent to the leader junction sequence.Deletion of the first 7 nt of the 8 nt AU-rich palindromicsequence in a BCoV D-RNA prevented replication. This is incontrast to the situation in MHV where many of the naturalMHV JHM DI-RNAs lacked the 8 nt AU-rich palindromicsequence.
Three stem–loop structures have been identified within the5« end of the BCoV DI-RNA (Chang et al., 1994, 1996).Deletion mutagenesis of the DI-RNA (Chang et al., 1994)showed that nt 1–8 were not required for replication but largerdeletions, & 12 nt, resulting in loss of stem–loop I, abolishedreplication. Disruption of BCoV DI-RNA stem–loop I by basesubstitution only minimally impaired DI-RNA replication.Analysis of the rescued DI-RNAs showed that they hadacquired helper virus-type leader sequence via leaderswitching. Chang et al. (1994) concluded that stem–loop I iscritical for the cis-acting replication signal associated with theleader sequence. Analysis of the 5« UTR of IBV D-RNA CD-61 and genomic RNAs identified three potential stem–loopstructures (Fig. 5). However, unlike the BCoV DI-RNAstructures, we have no biochemical evidence for the structuresin the IBV 5« UTR. Our phylogenetic evidence for theexistence of stem–loop I in the IBV 5« UTR supports the BCoVdata that this structure plays a major role in replication. IBVleader sequences tend to have more nucleotide substitutionsthan the adjacent 5« UTR, possibly a consequence of somesubstitutions requiring compensatory substitutions to maintainstem–loop I. It would be interesting to know whether MHVDI-RNAs lacking the 9 nt AU-rich sequence are able toreplicate, in the absence of leader switching, following theintroduction of stem–loop I destabilizing mutations.
The demonstration that rescue of IBV D-RNA CD-61 withheterologous virus results in leader switching is similar to theobservation that of Chang et al. (1994) for rescue of BCoV DI-RNAs. However, in our case, rather than reversion to wild-type sequence, as for BCoV, the leader sequence acquired bythe rescued IBV D-RNA was from the heterologous helper
HJJ

K. Stirrups and othersK. Stirrups and others
virus. For the IBV system there was no intrinsic requirementfor altering the leader sequence as no potentially destabilizingnucleotide substitutions had been introduced. We concludethat it is possible that leader switching is part of the replicationmechanism for IBV D-RNAs occurring with both heterologousand homologous helper viruses. We saw no evidence ofBeaudette-type leader sequence on any D-RNA followingrescue by heterologous virus. RT–PCR products derived fromthe rescued D-RNAs, CD61M41 and CD61H120, were clonedand several IBV-derived cDNAs sequenced to confirm thatthere were no minor populations of rescued D-RNAs withBeaudette-type leader sequences.
The results presented in this study together with previousobservations support the hypothesis that in general corona-virus D-RNAs do not appear to replicate by reiterativereplication. IBV D-RNAs leader switch in the absence ofintroduced point mutations indicating that leader sequenceacquisition is part of the replicative process. In this manner it isfeasible that coronavirus D-RNAs replicate in a manneranalogous to the synthesis of coronavirus subgenomicmRNAs, i.e. that they acquire a leader sequence by adiscontinuous process. However, leader acquisition in mRNAsynthesis is TAS dependent whereas leader acquisition duringD-RNA replication appears to be TAS independent. Thediscontinuous addition of a leader sequence during replicationof D-RNAs is an RNA recombination event since it involvesboth donor and acceptor templates. The recombination eventmust, by definition, be extremely efficient at a very early stageand}or there is some positive selection of the chimaeric D-RNA. Otherwise a mixed population of D-RNAs with the twodifferent leader sequences would be found. Therefore, wewould propose that replication of coronavirus D-RNAsinvolves acquisition of the leader sequence from the helpervirus, either from a homologous or heterologous virus, as partof the replication cycle, analogous to the manner thatsubgenomic mRNAs acquire their leader sequence via adiscontinuous mechanism. The acquisition of the leadersequence therefore probably occurs via an enzymatic processinvolving the polymerase. Such a potential activity will onlybe clarified following the development of a reverse geneticssystem for coronaviruses.
This work was supported by the Ministry of Agriculture, Fisheriesand Food, UK, Project code OD1905 and by grant number CT950064 ofthe Fourth RTD Framework Programme of the European Commission. K.Stirrups and K. Dalton were the holders of Research Studentships fromthe Biotechnology and Biological Sciences Research Council (BBSRC). S.Evans was supported by a BBSRC Realizing Our Potential Award(ROPA).
ReferencesAusubel, F. M., Brent, R., Kingston, R. E., Moore, D. D., Seidman, J. G.,Smith, J. A. & Struhl, K. (1987). Current Protocols In Molecular Biology.New York : John Wiley and Sons.
Binns, M. M., Boursnell, M. E. G., Cavanagh, D., Pappain, D. J. C. &Brown, T. D. K. (1985). Cloning and sequencing of the gene encodingthe spike protein of the coronavirus IBV. Journal of General Virology 66,719–726.
Binns, M. M., Boursnell, M. E. G., Tomley, F. M. & Brown, T. D. K.(1986). Comparison of the spike precursor sequences of coronavirus IBVstrains M41 and 6}82 with that of IBV Beaudette. Journal of GeneralVirology 67, 2825–2831.
Bonfield, J. K., Smith, K. F. & Staden, R. (1995). A new DNA sequenceassembly program. Nucleic Acids Research 24, 4992–4999.
Boursnell, M. E. G., Brown, T. D. K., Foulds, I. J., Green, P. F., Tomley,F. M. & Binns, M. M. (1987). Completion of the sequence of the genomeof the coronavirus avian infectious bronchitis virus. Journal of GeneralVirology 68, 57–77.
Brian, D. A. & Spaan, W. J. M. (1997). Recombination and coronavirusdefective interfering RNAs. Seminars in Virology 8, 101–111.
Chang, R. Y., Hofmann, M. A., Sethna, P. B. & Brian, D. A. (1994). Acis-acting function for the coronavirus leader in defective interfering RNAreplication. Journal of Virology 68, 8223–8231.
Chang, R. Y., Krishnan, R. & Brian, D. A. (1996). The UCUAAACpromoter motif is not required for high-frequency leader recombinationin bovine coronavirus defective interfering RNA. Journal of Virology 70,2720–2729.
Darbyshire, J. H., Rowell, J. G., Cook, J. K. A. & Peters, R. W. (1979).Taxonomic studies on strains of avian infectious bronchitis virus usingneutralisation tests in tracheal organ cultures. Archives of Virology 61,227–238.
Davelaar, F. G., Kouwenhoven, B. & Burger, A. G. (1984). Occurrenceand significance of infectious bronchitis virus variant strains in egg andbroiler production in the Netherlands. Veterinary Quarterly 6, 114–120.
Devereux, J., Haeberli, P. & Smithies, O. (1984). A comprehensive setof sequence analysis programs for the VAX. Nucleic Acids Research 12,387–395.
de Vries, A. A. F., Horzinek, M. C., Rottier, P. J. M. & de Groot, R. J.(1997). The genome organisation of the Nidovirales : similarities anddifferences between arteri-, toro- and coronaviruses. Seminars in Virology8, 33–47.
Feinberg, A. P. & Vogelstein, B. (1983). A technique for radiolabelingDNA restriction endonuclease fragments to high specific activity.Analytical Biochemistry 132, 6–13.
Hiscox, J. A., Mawditt, K. L., Cavanagh, D. & Britton, P. (1995).Investigation of the control of coronavirus subgenomic mRNA tran-scription by using T7-generated negative-sense RNA transcripts. Journalof Virology 69, 6219–6227.
Hofmann, M. A., Sethna, P. B. & Brian, D. A. (1990). Bovine corona-virus mRNA replication continues throughout persistent infection in cellculture. Journal of Virology 64, 4108–4114.
Jaeger, J. A., Turner, D. H. & Zuker, M. (1989). Improved predictionsof secondary structures for RNA. Proceedings of the National Academy ofSciences, USA 86, 7706–7710.
Jaeger, J. A., Turner, D. H. & Zuker, M. (1990). Predicting optimal andsuboptimal secondary structure for RNA. Methods in Enzymology 183,281–306.
Kottier, S. A., Cavanagh, D. & Britton, P. (1995). Experimental evidenceof recombination in coronavirus infectious bronchitis virus. Virology 213,569–580.
Kusters, J. G., Niesters, H. G. M., Lenstra, J. A., Horzinek, M. C. & VanDer Zeijst, B. A. M. (1989). Phylogeny of antigenic variants of aviancoronavirus IBV. Virology 169, 217–221.
IAA

Leader switching in IBV defective RNAsLeader switching in IBV defective RNAs
Lai, M. M. & Cavanagh, D. (1997). The molecular biology ofcoronaviruses. Advances in Virus Research 48, 1–100.
Makino, S. & Lai, M. M. C. (1989). High-frequency leader sequenceswitching during coronavirus defective interfering RNA replication.Journal of Virology 63, 5285–5292.
Makino, S., Stohlman, S. A. & Lai, M. M. C. (1986). Leader sequences ofmurine coronavirus mRNAs can be freely reassorted : evidence for therole of free leader RNA in transcription. Proceedings of the NationalAcademy of Sciences, USA 83, 4204–4208.
Makino, S., Shieh, C., Keck, J. G. & Lai, M. M. C. (1988a). Defective-interfering particles of murine coronavirus : mechanism of synthesis ofdefective viral RNAs. Virology 163, 104–111.
Makino, S., Shieh, C.-K., Soe, L. H., Baker, S. C. & Lai, M. M. C.(1988b). Primary structure and translation of a defective interferingRNA of murine coronavirus. Virology 166, 550–560.
Makino, S., Yokomori, K. & Lai, M. M. C. (1990). Analysis of efficientlypackaged defective Interfering RNAs of murine coronavirus : localizationof a possible RNA-packaging signal. Journal of Virology 64, 6045–6053.
Maruyama, I. N., Rakow, T. L. & Maruyama, H. I. (1995). cRACE: asimple method for identification of the 5« end of mRNAs. Nucleic AcidsResearch 23, 3796–3797.
Matzura, O. & Wennborg, A. (1996). RNAdraw: an integrated programfor RNA secondary structure calculation and analysis under 32-bitMicrosoft Windows. Computer Applications in the Biosciences 12, 247–249.
Mendez, A., Smerdou, C., Izeta, A., Gebauer, F. & Enjuanes, L. (1996).Molecular characterization of transmissible gastroenteritis coronavirusdefective interfering genomes : packaging and heterogeneity. Virology217, 495–507.
Nicholas, K. B. & Nicholas, H. B., Jr (1997). GeneDoc : a tool for editingand annotating multiple sequence alignments, 2.4.002 edn : Distributedby author (http :}}www.cris.com}Cketchup}genedoc. html).
Niesters, H. G. M., Lenstra, J. A., Spaan, W. J. M., Zijderveld, A. J.,Bleumink-Pluym, N. M. C., Hong, F., Van Scharrenburg, G. J. M.,Horzinek, M. C. & Van Der Zeijst, B. A. M. (1986). The peplomerprotein sequence of the M41 strain of coronavirus IBV and its comparisonwith Beaudette strains. Virus Research 5, 253–263.
Pe! nzes, Z. (1995). Defective replicating RNAs of coronavirus infectiousbronchitis virus : investigation of replication and genome packaging signals.PhD thesis, University of Hertfordshire, UK.
Pe! nzes, Z., Tibbles, K., Shaw, K., Britton, P., Brown, T. D. K. &Cavanagh, D. (1994). Characterization of a replicating and packageddefective RNA of avian coronavirus infectious bronchitis virus. Virology203, 286–293.
Pe! nzes, Z., Wroe, C., Brown, T. D., Britton, P. & Cavanagh, D. (1996).Replication and packaging of coronavirus infectious bronchitis virusdefective RNAs lacking a long open reading frame. Journal of Virology 70,8660–8668.
Sambrook, J., Fritsch, E. F. & Maniatis, T. (1989). Molecular Cloning : ALaboratory Manual, 2nd edn. Cold Spring Harbor, NY: Cold SpringHarbor Laboratory.
Sawicki, S. G. & Sawicki, D. L. (1990). Coronavirus transcription :subgenomic mouse hepatitis virus replicative intermediates function inRNA synthesis. Journal of Virology 64, 1050–1056.
Siddell, S. G. (1995). The Coronaviridae. In The Coronaviridae, pp. 1–10.Edited by S. G. Siddell. New York : Plenum.
Stern, D. F. & Kennedy, S. I. T. (1980). Coronavirus multiplicationstrategy. I. Identification and characterization of virus-specific RNA.Journal of Virology 34, 665–674.
Thompson, J. D., Gibson, T. J., Plewniak, F., Jeanmougin, F. & Higgins,D. G. (1997). The Clustal X Windows interface : flexible strategies formultiple sequence alignment aided by quality analysis tools. Nucleic AcidsResearch 25, 4876–4882.
van der Most, R. G. & Spaan, W. J. M. (1995). Coronavirus replication,transcription, and RNA recombination. In The Coronaviridae, pp. 11–31.Edited by S. G. Siddell. New York : Plenum.
van der Most, R. G., Bredenbeek, P. J. & Spaan, W. J. M. (1991). Adomain at the 3« end of the polymerase gene is essential for encapsidationof coronavirus defective interfering RNAs. Journal of Virology 65,3219–3226.
Zuker, M. (1989a). Computer prediction of RNA structure. Methods inEnzymology 180, 262–288.
Zuker, M. (1989b). On finding all suboptimal foldings of an RNAmolecule. Science 244, 48–52.
Zuker, M. & Stiegler, P. (1981). Optimal computer folding of largeRNA sequences using thermodynamics and auxiliary information. NucleicAcids Research 9, 133–148.
Received 10 September 1999; Accepted 23 November 1999
IAB




















