
POETICS Cultural classifications under discussion Latent ...

This article was downloaded by: [Università degli Studi di Milano]On: 05 November 2012, At: 05:42Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Critical Reviews in Food Science and NutritionPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/bfsn20
Latent Bioactive Peptides in Milk Proteins: ProteolyticActivation and Significance in Dairy ProcessingM. Gobbetti a , L. Stepaniak b , M. De Angelis d , A. Corsetti c & R. Di Cagno aa Dipartimento di Protezione delle Piante e Microbiologia Applicata, Università degli Studi diBari, Via G. Amendola 165/ A, 70126 Bari, Italyb Department of Food Science, Agricultural University of Norway, 1432-Aas, Norwayc Dipartimento di Scienze degli Alimenti, Sezione di Microbiologia Agro-alimentare,Università di Perugia, S. Costanzo, 06126 Perugia, Italyd Istituto di Science delle Produzion: Alilucuiar. (ISPA), CNR, Viale Einaudi 51, 70125 Bari,ItalyVersion of record first published: 03 Jun 2010.
To cite this article: M. Gobbetti, L. Stepaniak, M. De Angelis, A. Corsetti & R. Di Cagno (2002): Latent Bioactive Peptides inMilk Proteins: Proteolytic Activation and Significance in Dairy Processing, Critical Reviews in Food Science and Nutrition, 42:3,223-239
To link to this article: http://dx.doi.org/10.1080/10408690290825538
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form toanyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae, and drug doses shouldbe independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims,proceedings, demand, or costs or damages whatsoever or howsoever caused arising directly or indirectly inconnection with or arising out of the use of this material.

223
1040-8398/02/$.50© 2002 by CRC Press LLC
Critical Reviews in Food Science and Nutrition, 42(3):223–239 (2002)
Latent Bioactive Peptides in Milk Proteins:Proteolytic Activation and Significance in DairyProcessing
M. Gobbetti,1* L. Stepaniak,2 M. De Angelis,4 A. Corsetti,3 and R. DiCagno1
1Dipartimento di Protezione delle Piante e Microbiologia Applicata, Università degli Studi di Bari, Via G. Amendola 165/A, 70126 Bari, Italy; 2Department of Food Science, Agricultural University of Norway, 1432-Aas, Norway; 3Dipartimentodi Scienze degli Alimenti, Sezione di Microbiologia Agro-alimentare, Università di Perugia, S. Costanzo, 06126Perugia, Italy; 4Istituto di Science delle Produzion: Alilucuiar. (ISPA), CNR, Viale Einaudi 51, 70125 Bari, Italy
Referee: Mr. Dick Fitzgerald, Ph.D., Life Science Department, University of Limerick, Limerick, Ireland
* Corresponding author: M. Gobbetti. Address for correspondence:Dipartimento di Scienze degli Alimenti, Sezione di MicrobiologiaAgro-alimentare, Università di Perugia, S. Costanzo, 06126 Perugia, Italy. e-mail: [email protected]
ABSTRACT: After a brief description of the properties of bioactive peptides, the proteolytic activation of thebioactive sequences from milk protein precursors is discussed. The ability of proteolytic enzymes from varioussources, especially from lactic acid bacteria, to release bioactive peptides and the physiological and biotechnologi-cal significance of these peptides in dairy products are reviewed.
KEY WORDS: bioactive peptides, lactic acid bacteria, proteolysis, ACE-inhibitory activity, dairy products.
ABBREVIATIONS: BP, bioactive peptide; CN, casein; CPPs, caseinophosphopeptides; ACE, Angiotensin-I ConvertingEnzyme; LAB, lactic acid bacteria; PepX, X-prolyl-dipeptidyl aminopeptidase; CEP, cell-envelope proteinase; PEP, prolylendopeptidase.
I. INTRODUCTION
Milk proteins are currently the main source ofa range of biologically active peptides, even thoughother animal and plant proteins contain potentialbioactive sequences. These peptides, which areencrypted within the sequence of the parent pro-teins, can be released by enzymatic proteolysis,for example, during gastrointestinal digestion orduring food processing.
Proteolysis by naturally occurring enzymes inmilk, coagulants and by microbial enzymes, espe-cially from lactic acid bacteria (LAB) used as start-ers, generate bioactive peptides (BPs) during milkfermentation and cheese maturation, thereby en-riching the dairy products. Once produced, BPmay act in the body as regulatory compounds witha hormone-like activity. Concentrates of these pep-
tides are potential health-enhancing nutraceuticalsfor food and pharmaceutical applications.
Besides latent sequences of biologically ac-tive peptides, milk also contains a number ofbiologically active proteins, like lactoferrin andgrowth factors, that promote immuno-defense anddevelopment in the suckling young.1,2
After a brief description of the physiologicaleffects of BP, the proteolytic activation of latent BPand its significance in dairy products are discussed.
II. EFFECTS OF BIOACTIVE PEPTIDES
The biological effects of BPs are listed in Table1. The activity of these hormone-like peptides hasbeen the subject of several recent reviews,1,2,3,4, 5,6
and therefore will only be described briefly here.
Dow
nloa
ded
by [
Uni
vers
ità d
egli
Stud
i di M
ilano
] at
05:
42 0
5 N
ovem
ber
2012

224
TABLE 1Bioactivity of Peptides from Milk Proteins
Adapted from Meisel and Bockelmann 1999.3
FIGURE 1. Bioactive peptides in a sequence of bovine β-casein (A2 variant). Reported shortest peptides for eachcategory are shown. (Compiled from Meisel and Bockelmann, 1999; Asano et al., 1992; Dziuba et al., 1999;Yamamoto and Takano, 1999.3,7,8,9) 1, ACE-inhibitory and/or antihypertensive; 2, opioid agonist; 3, immunomodulatory;4, inhibitory to proline endopeptidase; 5, inhibitory to lactococcal metallo-oligopeptidases and; 6, mineral binding.
Dow
nloa
ded
by [
Uni
vers
ità d
egli
Stud
i di M
ilano
] at
05:
42 0
5 N
ovem
ber
2012

225
The shortest and most physiologically impor-tant BPs contained in the sequence of bovineβ-casein (CN) are shown in Figure 1.
A database containing 527 BPs from milkand other food proteins has been developed tofacilitate searching for new BPs.8
A. Opioid Peptides
Milk-protein derived bioactive peptides mayfunction as regulatory substance defined exorphinsor formones (food hormones). Although this roleneeds further clarification, pharmacological prop-erties similar to opium (morphine) and naloxone-inhibitable properties have been shown.2,10,11 Theα- and β-casomorphins and lactorphins act asopioid agonists, while casoxins act as opioid an-tagonists.10,11,12 Casomorphins may produce anal-gesia, modulate social behavior, influence post-prandial metabolism by stimulating the secretionof insulin and somatostatin, and may influencegastrointestinal absorption of nutrients by pro-longing the gastrointestinal transit time and exert-ing an anti-diarrhoeal effect.11 In vivo physiologi-cal effects occur when casomorphins are absorbedas long-chain precursors and hydrolyzed intosmaller bioactive fragments in the intestinal tis-sue. After crossing the intestinal mucosa, the frag-ments then react with µ- and δ-type receptorslocated throughout the intestinal tract and in thebrain.
B. Phosphopeptides
The CN sequence contains single and multiplephosphoryl residues and several phosphopeptidesare released in vitro and in vivo by enzymatichydrolysis of αs1-, αs2-, and β-CN.12,13 The nega-tively charged side chains of phosphoseryl resi-dues are the anionic binding sites for minerals.Such phosphopeptides withstand proteolysis in thegut, forming soluble complexes with calcium thathinder calcium phosphate precipitation14 and en-hance intestinal absorption of calcium and its re-tention in the body.11 Caseinophosphopeptides(CPPs) added to toothpaste may prevent enameldemineralization and exert an anticariogenic ef-
fect.15 Casein-derived phosphopeptides form orga-nophosphate salt with trace elements such as Fe,Mn, Cu, and Se, which function as carriers and areused in the treatment of rickets.11,16
C. Peptides Inhibitory to Angiotensin-IConverting Enzyme
Among BPs, the ACE-inhibitory peptides havebeen studied the most comprehensively. Informa-tion about biochemical features, physiologicalactivity, and mechanism of inhibition is available.Angiotensin I-converting enzyme (peptidyl-dipep-tidase; EC 3.4.15.1) (ACE) cleaves dipeptidesfrom the carboxy terminal end of the substrate. Itis a multifunctional ectoenzyme that is located indifferent tissues and plays a key physiologicalrole in the regulation of local levels of severalendogenous BP.17,18 ACE has two catalytic sites,one on each lobe of the extracellular portion.19
ACE has been classically associated with the re-nin-angiotensin system, which regulates the pe-ripheral blood pressure, where it catalyzes boththe production of the vasoconstrictor angiotensinII and the inactivation of the vasodilator bradyki-nin. ACE inhibition mainly results in an antihy-pertensive effect but may also influence differentregulatory systems involved in immuno-defenseand nervous system activity.20
Structure-activity correlations among differ-ent peptide inhibitors of ACE indicate that bind-ing to ACE is strongly influenced by the C-termi-nal tripeptide sequence of the substrate. ACEappears to prefer substrates or competitive inhibi-tors which mainly have hydrophobic (aromatic orbranched side chains) amino acid residues at thethree C-terminal positions. However, the struc-ture-activity relationship of ACE-inhibitory pep-tides has not yet been established and very differ-ent antihypertensive sequences have been derivedfrom a large number of food proteins.4
The first competitive inhibitors of ACE werereported from naturally occurring peptides in snakevenom.21,22 Thereafter, many other ACE inhibi-tors were discovered from enzymatic hydroly-sates or the related synthetic peptides of bovineand human caseins, and plant and other food pro-teins.23
Dow
nloa
ded
by [
Uni
vers
ità d
egli
Stud
i di M
ilano
] at
05:
42 0
5 N
ovem
ber
2012

226
D. Immunostimulating and Antimicrobialpeptides
Immunopeptides, obtained from αs1- andβ-CN, and α-lactalbumin stimulate phagocyticactivities of murine and human macrophages, andprotect against Klebsiella pneumoniae infectionin mice.11,24 Depending on the peptide concentra-tion, β-casokinin and β-casomorphin showed bothsuppression and stimulation of lymphocyte pro-liferation.25
Lactoferrin is an iron-binding glycoproteinpresent in most mammalian body fluids; it is acomponent of the host defense against microbialinfection.28 Antimicrobial peptides have beenderived from the whey protein lactoferrin. Diges-tion with pepsin releases peptides from bovineand human lactoferrin which have 100 to 1000times greater bactericidal activity than intactlactoferrin.2,27 Lactoferricin (bovine lactoferrinf17-41) is a pepsin-derived peptide from bovinelactoferrin, which has antimicrobial activity againstGram-negative bacteria and Candida albicans.The peptide is a more potent antimicrobial agentthan undigested lactoferrin because it causes adisruption of the bacterial cell membranes.
E. Antithrombotic Peptides
Similarities between the clotting of bloodand milk and between bovine κ-CN and thehuman fibrinogen γ-chain have been de-scribed.28,29,30 In the final step of blood clotting,thrombin hydrolyses fibrinogen into an insolublefibrin clot. Moreover, the fibrinogen γ-chain isessential for platelet aggregation because its400 to 411 sequence can bind the glycoproteinreceptors onto platelets. The dodecapeptide offibrinogen and the 106 to 110 sequence of theκ-CN and related sequences show functionalhomologies.28,31 The peptide, called casoplatelin,has been produced by trypsin hydrolysis ofκ-CN and shows antithrombotic activity by in-hibiting fibrinogen binding to platelets.29,31,32
Casoplatelins also contains part of the κ-CN106 to 169 sequence, the so-called macropeptidefragment, which inhibits gastrin and then theacid secretion in stomach.10
F. Peptides Inhibitory to HIV-1Proteinase
Infection by the retrovirus immunodeficiencytype-1 virus (HIV-1), the causative agent of theacquired immune deficiency syndrome (AIDS),occurs in a series of steps (replication, integra-tion, maturation) controlled by enzymes (reversetranscriptase, ribonuclease H, integrase, protein-ase), each of which is a potential target for thera-peutic treatment. Blocking the reverse transcriptaseactivity with azidothymidine (AZT) was the firstexample of using an inhibitor to produce an AIDSdrug that targeted a specific retroviral enzyme.33
The virally encoded protease (HIV-PR), an aspar-tic-protease essential for the maturation of viralproducts, is another target in the development ofpeptide analog inhibitors. Peptides produced byhydrolysis of food proteins should also be evalu-ated for this function.4
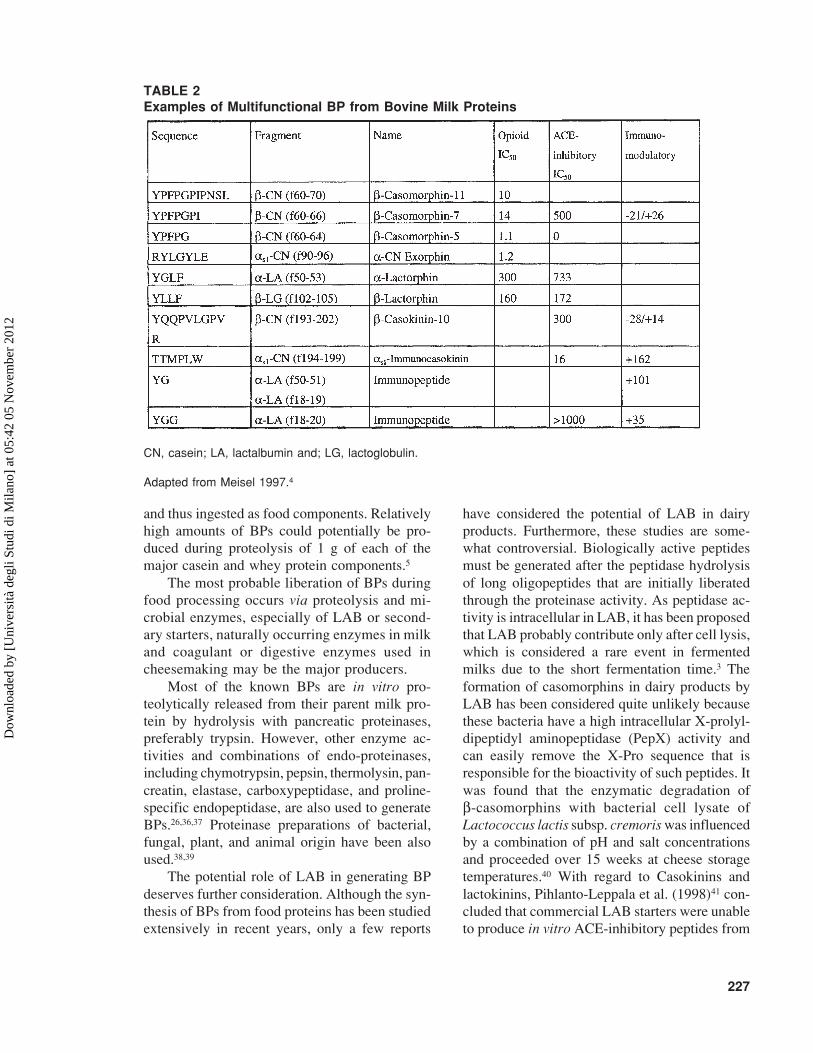
G. Multifunctional Bioactive Peptides
In contrast to endogenous BPs, many milk-derived peptides have multifunctional properties.Regions in the primary structure of caseins con-tain overlapping peptide sequences that exert dif-ferent biological effects. These regions, consid-ered to be strategic zones, are partially protectedfrom further proteolytic breakdown.28 For ex-ample, most β-casomorphins and casokinins areboth ACE-inhibitory and immunostimulatory,4,33,34
and α- and β-lactorphin contain sequences withboth opioid and ACE-inhibitory activities.35 Ex-amples of multifunctional BPs encrypted in bo-vine milk proteins are shown in Table 2.
III. PROTEOLYTIC ACTIVATION OFBIOACTIVE PEPTIDES FROM MILKPROTEINS
The release of BPs from their immediate pre-cursor sequence is a prerequisite for any func-tional role in a living system. In addition to thepossible liberation of BPs during intestinal pro-teolysis, such peptides may already be generatedduring the manufacture of several milk products
Dow
nloa
ded
by [
Uni
vers
ità d
egli
Stud
i di M
ilano
] at
05:
42 0
5 N
ovem
ber
2012
227
and thus ingested as food components. Relativelyhigh amounts of BPs could potentially be pro-duced during proteolysis of 1 g of each of themajor casein and whey protein components.5
The most probable liberation of BPs duringfood processing occurs via proteolysis and mi-crobial enzymes, especially of LAB or second-ary starters, naturally occurring enzymes in milkand coagulant or digestive enzymes used incheesemaking may be the major producers.
Most of the known BPs are in vitro pro-teolytically released from their parent milk pro-tein by hydrolysis with pancreatic proteinases,preferably trypsin. However, other enzyme ac-tivities and combinations of endo-proteinases,including chymotrypsin, pepsin, thermolysin, pan-creatin, elastase, carboxypeptidase, and proline-specific endopeptidase, are also used to generateBPs.26,36,37 Proteinase preparations of bacterial,fungal, plant, and animal origin have been alsoused.38,39
The potential role of LAB in generating BPdeserves further consideration. Although the syn-thesis of BPs from food proteins has been studiedextensively in recent years, only a few reports
have considered the potential of LAB in dairyproducts. Furthermore, these studies are some-what controversial. Biologically active peptidesmust be generated after the peptidase hydrolysisof long oligopeptides that are initially liberatedthrough the proteinase activity. As peptidase ac-tivity is intracellular in LAB, it has been proposedthat LAB probably contribute only after cell lysis,which is considered a rare event in fermentedmilks due to the short fermentation time.3 Theformation of casomorphins in dairy products byLAB has been considered quite unlikely becausethese bacteria have a high intracellular X-prolyl-dipeptidyl aminopeptidase (PepX) activity andcan easily remove the X-Pro sequence that isresponsible for the bioactivity of such peptides. Itwas found that the enzymatic degradation ofβ-casomorphins with bacterial cell lysate ofLactococcus lactis subsp. cremoris was influencedby a combination of pH and salt concentrationsand proceeded over 15 weeks at cheese storagetemperatures.40 With regard to Casokinins andlactokinins, Pihlanto-Leppala et al. (1998)41 con-cluded that commercial LAB starters were unableto produce in vitro ACE-inhibitory peptides from
TABLE 2Examples of Multifunctional BP from Bovine Milk Proteins
CN, casein; LA, lactalbumin and; LG, lactoglobulin.
Adapted from Meisel 1997.4
Dow
nloa
ded
by [
Uni
vers
ità d
egli
Stud
i di M
ilano
] at
05:
42 0
5 N
ovem
ber
2012

228
either casein or whey. It was also reported thatduring milk fermentation probiotic strains (e.g.,Lactobacillus GG) may produce severaloligopeptides that generate BP only after subse-quent digestion by pepsin and trypsin.42
The LAB proteolytic system is very complex.It is composed of extracellularly located serine-proteinase, a transport system specific for di-, tri-,and oligo-peptides (> three residues) and a multi-tude of intracellular peptidases. The specificitiesof the proteolytic enzymes of LAB have beendetermined by several authors.43 Although theearly papers indicated that only small parts ofcaseins were degraded by proteinases, a morerecent study has shown that LAB proteinases(Lactococcus sp. and Lactobacillus sp.) can hy-drolyze more than 40% of the peptide bonds ofthe β-CN resulting in the formation of more than100 different oligopeptides that are then activelydegraded by the complex peptidase system.44,45
αs1-CN may be hydrolyzed with the same capac-ity.43 Because most of the known BP derivedfrom food proteins correspond to milk proteinfragments, several BPs have been identified indairy products. Consequently, LAB could poten-tially generate a large variety of peptides, includ-ing bioactive sequences, and the type of dairyproduct, the technology adopted, and, especially,the strain selection are probably all factors thatmarkedly influence the proteolytic activation ofencrypted BP.
IV. SYNTHESIS AND SIGNIFICANCE OFBIOACTIVE PEPTIDES IN DAIRYPRODUCTS
A. Milk and Milk Hydrolysates
Milk contains an array of bioactive substancesthat extends the maternal benefits to her youngtoo far from the simple nutrition. Some of thesesubstances are fully active through proteins, whileothers are latent until they are proteolytically re-leased. Consequently, the manifestation of latentbioactivities depends on the site of proteolysis(mammary gland or a gastrointestinal site). Inaddition, full expression of bioactivity may re-quire the synergic action of BPs with non-peptide
agents in milk (lipids, glycolipids, and oligosac-charides).46,47 The bioactivities in milk includemodulators of digestive and gastrointestinal func-tions, hemodynamic modulators with potentialeffects on the gastrointestinal tract, hormones andgrowth factors. The proteolytic activity for re-leasing the BPs varies with differentiation or func-tional state, or intramammary infection of themammary gland.47 For example, casomorphinscan be liberated in the mammary gland, trans-ferred to the blood and then reach endogenousopiate receptors.48,49 In this way, casomorphinsmay participate in the endocrine regulation ofpregnancy, for example, by stimulating prolactinrelease.50 The cardiovascular system is anothertarget of casomorphins in pregnant or lactatingmammals. These peptides can exert a positiveinotropic and antiarrhythmic effect and thus mayhave a cardioprotective function.51
The latent bioactivities of milk proteins mayalso be activated by proteolysis of naturally oc-curring microorganisms. Hamel et al. (1985)52
were the first to identify β-casomorphin immu-noreactive material in cow milk that had beenincubated with various bacterial species, includ-ing LAB.
BPs with antithrombotic effects have beenenzymatically produced by trypsin digestion ofthe caseinomacropeptide53 and from β-CN-chymosin hydrolysate in continuous-stirred-tankmembrane reactors. The capacity to produceCPPs from sodium caseinate was compared in arange of commercially available protease prepa-rations of bacterial, fungal, plant, and animalorigin.38 CPPs were enriched from these hydroly-sates by calcium chloride aggregation at pH 7.5followed by ethanol precipitation of the aggre-gates. CPP yield ranged from 3.4 to 16.0% of theoriginal protein. The calcium binding and solu-bilizing abilities of the enriched CPPs rangedfrom 0.40 to 0.61 and 7.4 to 24.0 mg Ca2+ per mgCPP. Hydrolysis of sodium caseinate withBiprotease N100L resulted in the highest yield.The further exploitation of microbially derivedendo proteinases may also result in the produc-tion of more consistent, improved quality andlower cost CPP preparations. CPPs produced invitro may be used to enhance calcium absorptionacross the intestinal mucosa and also to prevent
Dow
nloa
ded
by [
Uni
vers
ità d
egli
Stud
i di M
ilano
] at
05:
42 0
5 N
ovem
ber
2012

229
and treat dental disease due to their anticariogenicproperties.15
ACE inhibitory peptides having different IC50values and corresponding to several α-lactalbu-min and β-lactoglobulin fractions were producedby hydrolysis with combinations of pepsin, trypsin,chymotrypsin, pancreatin, elastase, and carboxy-peptidase.36,37,38
Proteinase K from Tritirachium album pro-duced a hydrolysate from whey proteins, whichwhen administered orally had a greater effect onreducing blood pressure of hypertensive rats thanhydrolyzated obtained by using trypsin, actinaseE, thermolysin, papain, or chymotrypsin. TheseACE inhibitory peptides come from α-lactoglo-bulin, α2-microglobulin, serum albumin andα-casein.39
B. Fermented Milks
The type of starter used is one of the mainfactors that influences the synthesis of antihyper-tensive or opiate BPs in fermented milks; forexample, proteolysis by Lactobacillus helveticusis related to the production of antihypertensivepeptides. Nakamura et al. (1995)54 purified ACE-inhibitory peptides from Calpis, a Japanase softdrink made from skim milk fermented byLb. helveticus and Saccharomyces cerevisiae. Theantihypertensive peptides, Val-Pro-Proαs1 and Ile-Pro-Pro, which originated from both αs1- andβ-CN, have been found in milk inoculated withLb. helveticus55 and in casein hydrolysates pro-duced by Lb. helveticus CP790 extracellular pro-teinase.56 In a placebo-controlled study, the bloodpressure of hypertensive patients decreased sig-nificantly after 4 and 8 weeks of daily ingestion of95 ml of sour milk that contained the twotripeptides;57 the ingested dose of ACE-inhibitorypeptides ranged from 1.2 to 1.6 mg. During long-term feeding of a diet containing Calpis sour milkto spontaneously hypertensive rats, the antihyper-tensive effect not only followed the dose of asingle oral administration but also had an hyper-tensive preventive effect.58
Selected Lactobacillus delbrueckii subsp.bulgaricus and Lactococcus lactis subsp. cremoriswere used to produce two fermented milks that
contained ACE-inhibitory peptides.59 The ACE-inhibitory activity of the fermented milk startedby Lb. delbrueckii subsp. bulgaricus SS1 involvedpeptides such as β-CN f6-14, β-CN f7-14, β-CNf73-82, β-CN f74-82, and β-CN f75-82. The ac-tivity in the fermented milk produced by Lc. lactissubsp. cremoris FT4 had β-CN f7-14 in commonbut differed with fragments such as β-CN f47-52,β-CN f169-175, κ-CN 152 to 160 and κ-CN 155to 160 (Table 3). All of the above CN fragmentshad a higher proportion of hydrophobic residues(> of 60%). In particular, β-CN f73-82 and re-lated intermediates contained the Ile-Pro-Pro trip-eptide (β-CN f74-76) within the whole sequences,as previously mentioned for the Calpis sour milk,and β-CN f7-14 and related intermediate con-tained the internal sequence, Asn-Val-Pro-Gly,which also characterizes part of the sequences ofseveral other antihypertensive peptides isolatedfrom both β-CN and fish proteins.23,55 The β-CNf47-52 is part of another ACE-inhibitory peptidecorresponding to the longer β-CN f43-69 se-quence.55,56 In particular, the Asp-Lys-Ile-His-Pro-Phe sequence has five residues (including the lastfour C-terminal) in common with the octapeptideangiotensin-II (enzyme product of the ACE activ-ity). It should be noted that most drugs used inhypertension therapy are based on compoundsthat may compete for the receptor sites of thevasocostrictor angiotensin-II due to their partialhomology with this product of the ACE activity.The β-CN f169-175 sequence was also identifiedin casein hydrolysate produced by the purifiedextracellular proteinase of Lb. helveticus.60 Thispeptide did not show a strong ACE-inhibitoryactivity (IC50 > 1000 µmol/l). However, the cor-responding hexapeptide, Lys-Val-Leu-Pro-Val-Pro, obtained after being liberated from theC-terminal Gln residue by pancreatic digestion invitro, had a strong ACE-inhibitory activity (IC50= 5 µmol/l) as well as a remarkable antihyperten-sive effect in vivo. The IC50 values of the identi-fied peptide mixture were very low and werecomparable to those of the most active ACE-inhibitory peptides reported in the literature.3,23
When some of these peptides were chemicallysynthesized and used individually, the ACE in-hibitory activity was confirmed but the IC50 val-ues were 50 to 100 times higher. The values,
Dow
nloa
ded
by [
Uni
vers
ità d
egli
Stud
i di M
ilano
] at
05:
42 0
5 N
ovem
ber
2012

230
TABLE 3Sequences, Corresponding Casein Fragments and ACE-Inhibitory Activity of PeptidesContained in Fermented Milks Produced by Lactobacillus delbrueckii subsp.bulgaricus SS1 and Lactococcus lactis subsp. cremoris FT4
a,b Two inhibitory peptide mixtures were identified in the fermented milk produced by Lactobacillusdelbrueckii subsp. bulgaricus SS1 and one inhibitory peptide mixture was identified in the fer-mented milk produced by Lactococcus lactis subsp. cremoris FT4. The inhibitory mixtures corre-sponded to the fractions numbers 15 and 16, and 13, respectively, as isolated by RP-FPLC
c The ACE-inhibitory activity (IC50 mg/l) refers to the three mixtures of peptides.
From Gobbetti et al. 2000.59
b
Dow
nloa
ded
by [
Uni
vers
ità d
egli
Stud
i di M
ilano
] at
05:
42 0
5 N
ovem
ber
2012

231
however, were still within the range found forseveral other antihypertensive peptides. There isevidence that the catalytic sites of ACE may dif-fer in several properties and may have differentconformational requirements, which indicates thatACE inhibitors may selectively inhibit only onecatalytic site. These findings may demonstratethe need for a complex of peptides that haveslightly different conformational features in orderto inhibit ACE activity more completely. Oncethe inhibitory peptides were generated byLb. delbrueckii subsp. bulgaricus and Lc. lactissubsp. cremoris, they were resistant to furtherproteolysis during dairy processing or by trypsin.
Because the resistance to further proteolysisand the low molecular mass, in particular ACE-inhibitory peptides, contained in fermented milksmay resist intestinal digestion and then reach theirtarget sites at the luminal side of the intestinaltract or in peripheral organs.
In agreement with previous reports, a studyconducted on the antihypertensive effects of differ-ent kinds of fermented milks in spontaneouslyhypertensive rats showed that most whey frac-tions of milk fermented by Lb. helveticus andLb. delbrueckii subsp. bulgaricus have the high-est inhibitory activity.55 A fermented milk en-riched with the opioid β-casomorphin 1 to 4 (f60-63) has been produced by using a mutant strain ofLb. helveticus that lacks the PepX.61 As men-tioned previously, such peptidase activity mayeasily alter the X-Pro sequence that is responsiblefor the bioactivity. Antihypertensive compounds,corresponding to polysaccharide-glucopeptidecomplexes, were purified from a cell extract ofLactobacillus casei used in the production of fer-mented milks.62 This effect was confirmed inhuman patients with hypertension and was prob-ably not caused by inhibition of ACE but ratherby enhancement of prostaglandin I2 biosynthesisand the subsequent decrease in peripheral vascu-lar resistance.
UHT milk fermented by the probiotic Lactoba-cillus GG strain and subsequently digested by pep-sin and trypsin enzymes has been found to containBPs.42 The identified peptides, corresponding to frag-ments of αs1- and β-CN, and of α-lactalbumin,have different degrees of immunostimulatory, opioid,and ACE-inhibitory activities. The production of
these BPs could partially explain the probiotic prop-erties attributed to the Lactobacillus GG strain.Yogurt contains peptides that range from ca. 500 to10,000 Da, which, as demonstrated in vitro, mayreduce the risk of colon cancer. By adding peptidesto a cell culture model system, it was shown that thisactivity altered cancer cell proliferation in the co-lon.63
C. Cheeses
Several BPs have been found in cheeses andthis may be due to an intense but not excessiveproteolysis. Ripening conditions and the type ofstarters used variously affect the synthesis of BP.LAB, used as starter microrganisms in cheesemakingor constituting the endogenous lactic acid microf-lora of milk, release many different oligopeptidesfrom cell wall-envelope proteinases (CEP), and pep-tidases hydrolyse such oligopeptides into shorterfragments and amino acids that directly, or as pre-cursors, contribute to the flavor.64,65 Some of thesepeptides are bioactive.
Meisel et al. (1997)66 reported the presence ofACE-inhibitory peptides of low molecular massin several ripened cheeses. The ACE-inhibitoryactivity increases as proteolysis develops, but theACE-inhibition index decreases when the pro-teolysis during cheese maturation exceeds a cer-tain level (the ACE-inhibitory activity detected inmedium-aged Gouda was about double that of thelong-ripened Gouda). An antihypertensive pep-tide derived from αs1-casein was isolated from6-month ripened Parmesan cheese.67 This sequencewas degraded further and was not detectable after15 months of ripening. These results indicate thatsuch BPs liberated during cheese ripening by pro-teolytic enzymes of LAB can be degraded furtherto inactive fragments as a result of further pro-teolysis.66 However, low ACE-inhibition indexwas found in Quarg, an unripened cheese withlimited proteolysis.66
α-Casomorphin 7 was purified from Briecheese and peptides inhibitory to ACE activityhave been isolated from several Italian cheesescharacterized by short (Crescenza and Italico) andmedium (Gorgonzola) ripening.68 β-CN f58-72,which includes the sequence of β-casomorphin-7,
Dow
nloa
ded
by [
Uni
vers
ità d
egli
Stud
i di M
ilano
] at
05:
42 0
5 N
ovem
ber
2012

232
was isolated from Crescenza68 and Cheddarcheeses.69 This sequence has shown antihyperten-sive activity. Most of these Italian cheeses useLb. delbrueckii subsp. bulgaricus as starter. Itwas found by Western blot that Cheddar, Gouda,and Bleu cheeses contain peptides that react withantibodies specific to β-CN f58-70 and β-CN f58-72,70 (Stepaniak, unpublished). The peptides areapparently resistant to total degradation by en-zymes occurring in cheese, including those inmould cheeses. The peptides, which contain se-quences of β-casomorphins, were not degradedby lactococcal metallo-oligopeptidase PepO,lactococcal metallo aminopeptidase PepN, butinhibit these enzymes (see next section). Thesepeptides are also weak inhibitors of ACE(Stepaniak, unpublished).
Proteolytic enzymes of LAB produce CPPsduring ripening of cooked curd cheeses likeComtè.71 Peptides, derived from a cheese slurrythat uses Lc. lactis subsp. lactis as starter culture,have shown anticancer effects.72 Other authorshave, however, shown that β-casomorphins incheeses may be hydrolyzed further by the pro-teolytic enzymes of Lc. lactis subsp. cremoris.The degradation was strongly dependent on thepH and salt concentration of the cheese and wasvery slow during ripening.40
Milk clotting or digestive enzymes have alsobeen studied to determine their capacity to pro-duce BPs. β-CN f193-209, β-CN f194-209, β-CNf69-97, β-CN f141-163 and β-CN f69-84 havebeen produced in vitro by chymosin, trypsin, chy-motrypsin, and CEP from Lc. lactis subsp.cremoris.69,73,74,75,76 These fragments are bitter orcontain sequences of bitter peptides but are alsorelated to a bioactive sequence.3,73,76 β-CN f193-209 and β-CN f194-209 have been found to pos-sess antihypertensive effects.53 These two C ter-minal fragments, released preferentially bychymosin and lactococcal CEP, inhibit ACE andlactococcal endopeptidase77 (Stepaniak, unpub-lished). β-CN f193-209 contains sequences of adouble-function BP that inhibits ACE and is alsoimmunomodulatory.3
ACE inhibitory peptides were also found inenzyme-modified cheese (EMC) obtained by fur-ther hydrolysis of pasteurised cheese homogenizedwith Neutrase and enzyme preparation form
Lb. casei. β-Casomorphin was only found in EMCprepared with Neutrase, which indicates the de-structive action of PepX by LAB onβ-casomorphin content in cheese.78
D. Peptides with Potential PhysiologicalSignificance
Some physiological properties (antihyperten-sive, immunostimulatory, antithrombotic, and HIVinhibitory) attributed to BPs have a common mecha-nism based on the inhibition of target proteolyticenzymes. Protein hydrolysis and/or its inhibitionplay a crucial physiological role in food digestion,blood coagulation and fibrinolysis, blood pressureregulation, hormone neuromodulation, and phago-cytosis. It is also involved in pathological pro-cesses (e.g., emphysema, cancer, and hyperten-sion) and infection (e.g., HIV or invasiveparasites).79 A strict control of proteolysis involvesthe synthesis of zymogens, enzyme specificity, and,especially, specific inhibitors. Inhibitor moleculesexceed 10% of the total protein in human plasma.80
Therefore, it is not surprising that a similarmechanism may influence food ecosystems. Be-sides affecting digestive and food spoilage en-zymes, the presence of inhibitory peptides in foodscan adversely affect intended enzymatic modifi-cations of ingredients during food processing.80
We have seen that proteolytic enzymes from LABproduce BPs in several dairy products. Once lib-erated, BPs or peptides related to BP sequencemay influence the biochemical activity of themicrobial communities. Although in the absenceof a clear in vivo evidence of the link betweenpeptides inhibiting microbial enzymes and bioac-tivity, this fact has probably been underestimatedin dairy processing.
β-CN f58-72, containing the sequence ofβ-casomorphin-7, is released during Cheddar,Jarlsberg, and Crescenza ripening and inhibitsnoncaseolytic oligopeptidase PepO and aminopep-tidase PepN from LAB.68,69 β-Casomorphin-7(β-CN f60-66), which has an opioid effect, inhib-its ACE activity and LAB endo and amino pepti-dases.34,69 β-CN f193-209 and β-CN f194-209produced by chymosin, trypsin, and chymotrypsinand by CEP from Lc. lactis subsp. cremoris pos-
Dow
nloa
ded
by [
Uni
vers
ità d
egli
Stud
i di M
ilano
] at
05:
42 0
5 N
ovem
ber
2012

233
sess antihypertensive effects and inhibit endopep-tidases and aminopeptidases of LAB. After beingreleased, these peptides are not further hydro-lyzed or hydrolyze very slowly by chymosin andCEPs.73,75 These peptides may be actively de-graded by thermolysin and proteinase fromPseudomonas fluorescens.70
LAB peptidases differ in their sensitivity tothe same peptides. Table 4 shows the differentbiochemical features and sensitivities of bacterialpeptidases to β-CN f58-72. This peptide relatedto BP strongly inhibited a metalloendopeptidasefrom Lc. lactis susp. lactis MG1363, but showeda reduced inhibitory activity on metalloaminopep-tidase from the same microrganism.69 Similar toendopeptidase from Lactococcus, thermolisin, ametalloendopeptidase from Bacillus thermo-proteolyticus, is inhibited by phospho-ramidon[N-(α-rhamnopyranosyloxyhydroxyphosphinyl)-Leu-Trp)], but it was not inhibited and it degradedβ-CN f58-72.75 The PepX from Lc. lactis MG1363,which is not a metalloenzyme, was not inhibitedby the above peptides and hydrolyzed β-CN f60-70 and β-CN f60-66.69 The non-caseolyticoligopeptidase PepO of Lb. subsp. casei subsp.casei 2752 was less sensitive to peptides isolatedfrom Italian cheeses, including β-CN f58-72, thanthat of Streptococcus thermophilus 305. Both en-zymes are metalloenzymes but differ in their sen-sitivities to iodoacetic acid and dithiothreitol.68 Incontrast to oligopeptidase PepO from Lactococcus,those from Propionibacterium are almost com-pletely insensitive to peptides from Cheddar andItalian cheeses and are inhibited differently byphosphoramidon and β-CN f58-72.81,82
In conclusion, it can be affirmed that theinhibition of enzymes belonging to the samebiological class is species and strain specificand is related to the type of cheese and ripen-ing period. Changes in specificity and in thesensitivity to inhibitory peptides seem to berelated to minor variations in the amino acidsequences of the enzymes, especially thoseinvolved in the catalytic domain.83 Indeed,peptide analog inhibitors, commonly used inpharmaceutical preparations, specifically in-hibit the target enzyme based on a judiciousselection of the flanking amino acids in thepeptide analog.33
Interestingly, peptides with ACE-inhibitoryactivity isolated from Italian cheeses82 and fromtryptic zein hydrolyzates84 were shown to be in-hibitory to spoilage microbial enzymes, such asthermostable proteinases from psychrotrophic P.fluorescens, which cause bitterness in UHT milk,contribute to age gelation of unconcentrated UHTmilk, and drastically reduce the shelf-life of dairyproducts. The control of these proteolytic enzymesis very important for preventing dairy productspoilage.
V. POTENTIAL USES OF BIOACTIVEPEPTIDES IN FUNCTIONAL FOODS ANDPHARMACEUTICALS
A physiological functional food can be de-fined as a food derived from naturally occurringsubstances that can and should be consumed aspart of the daily diet, and thus provides healthbenefits in reducing the risk of developing dis-ease.85 A nutraceutical is any substance that is afood or a part of a food that provides medical orhealth benefits, including prevention and treat-ment of disease.86 Casein-derived peptides, whichcan be manufactured on an industrial scale, havealready been considered for use as dietary supple-ments and pharmaceutical preparations. CPPs wereobtained from enzymatic hydrolysates by ionexchange chromatography87 or by aggregation ofhydrolysate peptides with bivalent cations in com-bination with ultrafiltration.88 Regarding the pos-sible application of CPPs as mineral carriers, theyhave been proposed for use in dietary productssuch as bread, cake, flour, beverages, chewinggum and in pharmaceutical preparations such astablets, toothpaste, and dental filling material.89
β-Casomorphins have also been produced by ge-netic engineering techniques followed by enzy-matic or chemical cleavage of the microbial fu-sion protein to liberate the required peptide.90
Tryptic hydrolysates of casein, containing ACE-inhibitory peptides, have been suggested for usein functional foods to prevent hypertension. Thisfinding is based on studies with normotensive andmildly hypertensive volunteers who ingested 10 gof a tryptic casein hydrolysate twice a day for4 weeks.91 While ACE-inhibitory peptides de-
Dow
nloa
ded
by [
Uni
vers
ità d
egli
Stud
i di M
ilano
] at
05:
42 0
5 N
ovem
ber
2012

234
TABLE 4Influence of β-Casein f58-72 on the Activity of Characterized Bacterial Peptidases
++ very high inhibition; +, high inhibition; -, low inhibition; —, very low inhibition; =, no inhibition.
From Smacchi and Gobbetti 2000.6
Dow
nloa
ded
by [
Uni
vers
ità d
egli
Stud
i di M
ilano
] at
05:
42 0
5 N
ovem
ber
2012

235
rived from milk products are not as potent as thedrugs commonly used in the treatment of hyper-tension, milk products, such as the Calpis sourmilk, having moderate bioactivity (e.g., ACE-inhibition index ≥ 70, IC50 100 to 500 mmol/l)may be considered as natural functional foods4
that should be consumed as part of the daily diet,and which serve to regulate a particular bodilyprocess when ingested. The di- and tri-peptidescorresponding to the immunomodulating se-quences derived from milk proteins were alsofound to be the active components in dialyzedleukocyte extract from normal donors which wasused in a large multicenter trial to inhibit thedevelopment of infections in patients with pre-AIDS.92 Encouraging results after bi-weekly treat-ment of 93 patients with an AIDS-related syndromeshowed a significantly reduced tendency to progressto a clinically relevant endpoint or to AIDS.
BPs may also provide new dietary methodsfor improving the productive efficiency of live-stock, by using the properties of opioid peptideson intake regulation of ruminants.93
Currently, CPPs and ACE-inhibitory peptidesare the most favored BP for use in foodstuffsformulated to provide specific benefits. Thepresent knowledge on BPs encourages the devel-opment of functional food containing BPs pro-duced by food-grade (GRAS) microorganismsduring fermentation of milk or other materials ofplant or animal origin or a combinations of them.
VI. FUTURE WORK
Difficulties in studying the gastrointestinaltract in vivo limit the knowledge of the bioactivefunctions of milk. The development of dynamicin vitro models capable of studying the interac-tive effect of multiple bioactive components hasbeen postulated.2
More work is needed to develop large-scalefractionation or protein hydrolyzates to obtainproducts enriched with biological peptides of spe-cific function that could be used as nutraceuticaladditives in functional foods. Especially valuableare the short BPs that are resistant to degradationby digestive enzymes and therefore may be easilyabsorbed. Isolation of such peptides is particu-
larly difficult. More research is needed that willprovide a better understanding of the effects ofdifferent processing technologies, especially new,emerging technologies like ultra-high pressuretreatment, on the interaction of BP or bioactiveproteins from milk with other food components.
The exploitation of peptidase inhibitors as trueBPs with potential role in prevention or treatment ofhuman pathologies that involve prolylendopeptidase(PEP) (e.g., Alzheimer’s disease) should be war-ranted in fermented dairy products.94,95,96
In the future transgenic plants may also be apotential source of BPs.
REFERENCES
1. Korhonen, H., Pihlanto Leppala, A., Rantamaki, P.,and Tupasela, T., Impact of processing on bioactiveproteins and peptides, Trends Food Sci. Technol.,1998; 9: 307–319.
2. Schanbacher, F.L., Talhouk, R.S., Murray, F.A.,Gherman, L.I., Willet, L.B., Milk-borne bioactivepeptides, Int. Dairy J., 1998; 8: 393–403.
3. Meisel, H. and Bockelmann, W., Bioactive peptidesencrypted in milks proteins: proteolytic activation andthropho-functional properties, Antonie van Leeuwenhoek,1999; 76: 207–215.
4. Meisel, H., Biochemical properties of bioactive peptidesderived from milk proteins: potential nutraceuticals forfood and pharmaceutical applications, Livestock Prod.Sci., 1997; 50: 125–138.
5. Meisel, H., Overview on milk protein-derived pep-tides, Int. Dairy J., 1998; 8: 363–373.
6. Smacchi, E. and Gobbetti, M., Bioactive peptides indairy products: synthesis and interaction with pro-teolytic enzymes, Food Microbiol., 2000; 17: 129–141.
7. Asano, M., Nio, N., and Ariyoshi, Y., Inhibition ofprolyl endopeptidase by synthetic peptide-fragmentsof human beta-casein, Agric. Biol. Chem., 1991; 553:825–828.
8. Dziuba, J., Minkiewicz, P., Nalecz, D., and Iwaniak,A., Database of biologically active peptide sequences,Nahrung, 1999; 433: 190–195.
9. Yamamoto, N., and Takano, T., Antihypertensive pep-tides derived from milk proteins, Nahrung, 1999; 43:159–164.
10. Meisel, H., Frister, H., and Schlimme, E., Biologi-cally active peptides in milk proteins, Z. Ernharung.,1989; 28: 267–278.
11. Meisel, H. and Schlimme, E., Milk proteins: precur-sors of bioactive peptides, Trends Food Sci. Technol.,1990; 1: 41–43.
12. Schlimme, E., Meisel, H., and Frister, H., Bioactivesequences in milk proteins. In: Milk Proteins: Nutri-
Dow
nloa
ded
by [
Uni
vers
ità d
egli
Stud
i di M
ilano
] at
05:
42 0
5 N
ovem
ber
2012

236
tional, Clinical, Functional and Technological As-pects, Barth, C. A., and Schlimme, E., Eds., Springer,New York, 1988, 143–149.
13. Chiba, H. and Yoshikawa, M., Biologically functionalpeptides from food proteins: new opioid peptides frommilk proteins. In: Protein Tailoring for Food andMedical Uses, Feeney, R. E. and Whitaker, J. R., Eds.,Davis, California, Department of Food Science andTechnology, University of California, 1986, 123–153.
14. Tirelli, A., De Noni, I., and Resmini, P., Bioactivepeptides in milk products, Ital. J. Food Sci., 1997; 2:91–98.
15. Reynolds, E.C., Cain, C.J., Webber, F.L., Black, C.L.,Riley, P.F., Johnson, I.H., and Perich, J.W.,Anticariogenicity of calcium phosphate complexes oftryptic caseinophosphopeptides in the rat, J. DentalRes., 1995; 74: 1272–1279.
16. Kitts, D. D. and Yuan, Y. V., Caseinophosphopeptidesand calcium bioavailability, Trends Food Sci. Technol.,1992; 3: 31–35.
17. Ondetti, M.A. and Cushman, D.W., Enzymes of therenin-angiotensin system and their inhibitors, Annu.Rev. Biochem., 1982; 51: 283–308.
18. Bruneval, P., Hinglais, N., and Alhenc-Gelas, F., An-giotensin I converting enzyme in human intestine andkidney. Ultrastructural immunohistochemical local-ization, Histochemistry, 1986; 86: 73–80.
19. Johnston, C.I., Renin-angiotensin system: a dual tis-sue and hormonal system for cardiovascular control,J. Hypert., 1992; 10: S13–S26.
20. Meisel, H., Casokinins as inhibitors of Angiotensin-Converting-Enzyme. In: New Perspectives in InfantNutrition, Sawatzki, G. and Renner, B., Eds., Thieme,Stuttgart, New York, 1993, 153–159.
21. Ferreira, S.H., Bartet, D.C., and Greene, L.J., Isola-tion of bradykinin-potentiating peptides from Bothropsjararaca venom, Biochemistry, 1970; 9: 2583–2593.
22. Ondetti, M.A., Williams, N.J., Sabo E.F., Pluvec, J.,Weaver, E.R., and Kocy, O., Angiotensin convertingenzyme inhibitors from the venom of Bothropsjararaca, isolation, elucidation of structure and syn-thesis, Biochemistry, 1971; 10: 4033–4039.
23. Yamamoto, N., Antihypertensive peptides derivedfrom food proteins, Biopoly, 1997; 43: 129–134.
24. Migliore-Samour, D., Floc’h, F., and Jollès, P., Bio-logically active casein peptides implicated inimmunomodulation, J. Dairy Res., 1989; 56: 357–362.
25. Kayser, H. and Meisel, H. Stimulation of human pe-ripheral lymphocytes by bioactive peptides derivedfrom bovine milk protein, FEBS Lett., 1996; 383: 18–20.
26. Meisel, H. and Schlimme, E., Bioactive peptides de-rived from milk proteins: ingredients for functionalfoods, Kiel. Milchw. Forsch., 1996; 48: 343–357.
27. Bellamy, W., Takase, M., Wakabayashi, H., Kawase,K., and Tomita, M. Antibacterial spectrum oflactoferricin B, a potent bactericidal peptide derived
from the N-terminal region of bovine lactoferrin, J.Appl. Bacteriol., 1992; 73: 472–479.
28. Fiat, A. M., Milgliore-Samour, D., Jollès, P., Drouet, L.,Sollier, C. B. D., and Caen, J., Biologically active pep-tides from milk proteins with emphasis on two examplesconcerning antithrombotic and immunomodulating ac-tivities, J. Dairy Sci., 1993; 76: 301–310.
29. Jollès, P. and Henschen, A., Comparison between theclotting of blood and milk, Trends Biochem. Sci.,1982; 7: 325–330.
30. Jollès, P., Loucheux-Lefebvre, M.-H., and Henschen,A., Structural relatedness of ε-casein and fibrinogen8-chain, J. Mol. Evol., 1978; 11: 271–277.
31. Jollès, P., Levy-Toledano, S., Fiat, A. M., Soria,C., Gillesen, D., Thomaidis, A., Dunn, F. W., andCaen, J., Analogy between fibrinogen and casein:effect of an undecapeptide isolated from k-caseinon platelet function, Eur. J. Biochem., 1986; 158:379–382.
32. Mazoyer, E., Bal Dit Sollier, C., Drouet, L., Fiat, A.M., Jollès, P., and Caen, J., Active peptides fromhuman and cow’s milk proteins effects on plateletsfunction and vessel wall. In: Foods, Nutrition andImmunity, Vol. 1, Paubert-Braquet, M., Dupont, C.,and Paoletti, R., Eds., Basel, Dyn. Nutr. Res., Karger,1992.
33. Grant, S. K., Meek, T. D., Metcalf, B. W., andPetteway, S. R., Design of peptide analog inhibitors ofproteolytic processes. In: Biologically Active Peptides:Design, Synthesis, and Utilization, Williams, W. V.and Weiner, B. D., Eds., Lancaster, Technomic Pub-lishing Co., 1993, 325–352.
34. Meisel, H. and Schlimme, E., Casokinins as inhibitorsof angiotensin converting enzyme. In: Proceedings ofthe 24th International Dairy Congress, FIL-IDF Eol,Ed., Melbourne, ANCIDF, 1994, 327.
35. Mullally, M.M., Meisel, H., and Fitzgerald, R.J. Syn-thetic peptides corresponding to α-lactalbumin and β-lactoglobulin sequences with angiotensin-I-convertingenzyme inhibitory activity, Biol. Chem. Hoppe-Seyler,1996; 377: 259–260.
36. Pihlanto-Leppala, A., Koskinen P., Piilola K.,Tupasela, T., and Korhonen, H., Angiotensin I-con-verting enzyme inhibitory properties of whey proteindigests: concentration and characterization of activepeptides, J. Dairy Res., 2000; 67: 53–64.
37. Mullally, M.M., Meisel, H., and Fitzgerald, R.J. An-giotensin-I-converting enzyme inhibitory activities ofgastric pancreatic proteinase digests of whey proteins,Int. Dairy J., 1997; 7: 299–303.
38. McDonagh, D. and FitzGerald, R.J., Production ofcaseinophosphopeptides (CPPs) from sodium casein-ate using a range of commercial protease prepara-tions, Int. Dairy J., 1998; 8: 39–45.
39. Abubakar, A., Saito, T., Kitazawa, H., Kawai, Y., andItoh, T., Structural analysis of new antihypertensivepeptides derived from cheese whey by proteinase Kdigestion, J. Dairy Sci., 1994; 81: 3131–3138.
Dow
nloa
ded
by [
Uni
vers
ità d
egli
Stud
i di M
ilano
] at
05:
42 0
5 N
ovem
ber
2012

237
40. Muehlenkamp, M.R., and Warthesen, J.J., β-Casomorphins: analysis in cheese and susceptibilityto proteolytic enzymes from Lactobacillus lactis ssp.cremoris, J. Dairy Sci., 1996; 79: 20–26.
41. Pihlanto-Leppala, A. Rokka, T., and Korhonen, H.,Angiotensin I converting enzyme inhibitory peptidesfrom bovine milk proteins, Int. Dairy. J., 1998; 8:325–331.
42. Rokka, T., Syv‰oja, E. L., Tuominen, J., andKorhonen, H., Release of bioactive peptides by enzy-matic proteolysis of Lactobacillus GG fermented UHTmilk, Milchwissenschaft, 1997; 52: 675–678.
43. Kunji, E. R. S., Mierau, I., Hagting, A., Poolman, B.,and Konings, N., The proteolytic system of lactic acidbacteria, Antonie van Leeuwenhoek, 1996; 70: 187–221.
44. Mierau, I., Kunji, E.R.S., Venema, G., and Kok, J.,Casein and peptide degradation in lactic acid bacteria,Biotech. Genetic Engineering Rev., 1997; 14: 279–301.
45. Juillard, V., Laan, H., Kunji, E.R.S., Jeronimus-Stratingh, C.M., Bruins, A.P., and Konings, W.N.,The extracellular PI-type proteinase of Lactococcuslactis hydrolyses β-casein into more than one hundrenddifferent oligopeptides, J. Bacteriol., 1995; 177: 3472–3478.
46. Nuijens, J. H., van Berkel, P. H. C., and Schanbacher,F. L., Structure and biological actions of lactoferrin, J.Mam. Gland Biol. Neoplasia, 1996; 1: 285–294.
47. Schanbacher, F. L., Talhouck, R. S., and Murray, F.A., Biology and origin of bioactive peptides in milk,Livestock Prod. Sci., 1997; 50: 105–123.
48. Assargard, U., Larsson, C., Norby, U., Linström, L.,and Nyberg, F., Human β-casomorphin-5 containingpeptides in human body fluids. In: β-Casorphins andRelated Peptides: Recent Developments, Brantl, V.and Teschemacher, H., Eds., VCH, Weinheim, 1994,247–254.
49. Koch, G., Lange, E., Link, G., Bodehker, R.H., andTeschemacher, H., Analysis of clinical data and plasmalevels of β-casomorphin-8 immunoreactive materialin pregnant and puerperal women. In: β-Casorphinsand Related Peptides: Recent Developments, Brantl,V., and Teschemacher, H., Eds., VCH, Weinheim,1984, 227–239.
50. Yen, S.S.C., Quigley, M.E., Reid, R.L., Ropert, J.F.,and Cetel, N.S., Neuroendocrinology of opioid pep-tides and their role in the control of gonadotropin andprolactin secretion, Am. J. Obstetics Gynecol., 1985;152: 485–493.
51. Mentz, P., Neubert, K., Liebmann, C., Hoffmann, S.,Schrader, U., and Barth, A., In: β-casomorphin: pos-sible physiological significance, In Proccedings fromthe 1st International Symposium on β-Casorphins andRelated Peptides, Nyberg, F. and Brantl, V., Eds.,Fyris-Tryck AB, Uppsala, 1990, 143–149.
52. Hamel, U., Kielwein, G., and Teschemacher, H., α-Caseimophin immunoreactive materials in cows’ milk
incubated with various bacterial species, J. Dairy Res.,1985; 52: 139–148.
53. Bouhallab, S., Mollè, D., and Lèonil, J., Tryptic hy-drolysis of caseinomacropeptide in membrane reac-tor: preparation of bioactive peptides, Biotechnol. Lett.,1992; 14: 805–810.
54. Nakamura, Y., Yamamoto, N., Sakai, K., Okubo, A.,Yamazaki, S., and Takano, T., Purification and char-acterization of angiotensin I-converting enzyme in-hibitors from sour milk, J. Dairy Sci., 1995; 78: 777–783.
55. Yamamoto, N., Akino, A., and Takano, T., Antihy-pertensive effect of different kinds of fermented milkin spontaneously hypertensive rats. Biosci. Biotech.Biochem., 1994; 58: 776–778.
56. Yamamoto, N., Akino, A., and Takano, T., Antihy-pertensive effect of the peptides derived from caseinby an extracellular proteinase from Lactobacillushelveticus CP790, J. Dairy Sci., 1994; 77: 917–922.
57. Hata, Y., Yamamoto, M., Ohni, H., Nakajima, K.,Nakamura, Y., and Takano, T. A placebo-controlled studyof the effect of sour milk on blood pressure in hypertensivesubjects, Am. J. Clin. Nutr. 1996; 64: 767–771.
58. Nakamura, Y., Masuda, O., and Takano, T. Decreaseof tissue angiotensin-I-converting enzyme activityupon feeding sour milk in spontaneously hypertensiverats, Biosci. Biotech. Biochem., 1996; 60: 488–489.
59. Gobbetti, M., Ferranti, P., Smacchi, E., Goffredi, F.,and Addeo, F., Production of angiotensin-I convertingenzyme (ACE)-inhibitory peptides in fermented milksstarted by Lactobacillus delbrueckii subsp. bulgaricusSS1 and Lactococcus lactis subsp. cremoris FT4. Appl.Environ. Microbiol., 2000; 66: 3898–3904.
60. Maeno, M., Yamamoto, Y., and Takano, T., Identifi-cation of an antihypertensive peptide from casein hy-drolysate by a proteinase from Lactobacillus helveticusCP790, J. Dairy Sci., 1998; 79: 1316–1321.
61. Matar, C. and Goulet, J., β-Casomorphin 4 milk fer-mented by a mutant of Lactobacillus helveticus, Int.Dairy J., 1996; 6: 383–397.
62. Yamamoto, N. Antihypertensive peptides derived fromfood proteins, Biopoly, 1997; 43: 129–134.
63. Ganjam, L. S., Thornton, W. H., Marshall, R. T., andMacDonald, R. S., Antiproliferative effects of yogurtfractions obtained by membrane dialysis on culturedmammalian intestinal cells, J. Dairy Sci., 1997; 80:2325–2329.
64. Fox, P. F., Singh, T., and McSweeney, P. L. H., Pro-teolysis in cheese during ripening. In: Biochemistry ofMilk Products, Andrews, A. T. and Varley, J., Eds., TheRoyal Society of Chemistry, London, 1984, 1–31.
65. Stepaniak, L. and Fox, P. F., Characterization of theprincipal intracellular endopeptidase from Lactococcuslactis subsp. lactis MG 1363, Int. Dairy J., 1995; 5:699–713.
66. Meisel, H., Goepfert, A., and Günther, S., ACE in-hibitory activities in milk products, Milchwissenschaft,1997; 52: 307–311.
Dow
nloa
ded
by [
Uni
vers
ità d
egli
Stud
i di M
ilano
] at
05:
42 0
5 N
ovem
ber
2012

238
67. Addeo, F., Chianese, L., Salzano, A., Sacchi, R.,Cappuccio, U., Ferranti, P., and Malorni, A., Charac-terization of the 12% trichloroacetic acid-insolubleoligopeptides of Parmigiano-Reggiano cheese, J. DairyRes., 1992; 59: 401–411.
68. Smacchi, E. and Gobbetti, M., Peptides from severalItalian cheeses inhibitory to proteolytic enzymes oflactic acid bacteria, Pseudomonas fluorescens ATCC948 and to the angiotensin I-converting enzyme, En-zyme Microb. Technol., 1998; 22: 687–694.
69. Stepaniak, L., Fox, P. F., Sørhaug, T., and Grabska, J.J., Effect of peptides from the sequence 58–72 ofβ-casein on the activity of endopeptidase, aminopep-tidase and X-prolyl-dipeptidyl aminopeptidase fromLactococcus, 1995, Agric. Food Chem., 1995; 43:849–853.
70. Stepaniak, L., and Gobbetti, M., Immunoreactivityand sensitivity of peptides from β-casein, which in-hibit lactococcal peptidases to hydrolysis bythermolisin and proteinase from Pseudomonasfluorescens, Poli. J. Food. Nutr. Sci., 1997; 6: 41–50.
71. Roudot-Algaron, F., Le Bars, D., Kerhoas, L., Einhorn,J., and Gripon, J. C., Phosphopeptides from Comtècheese: nature and origin, J. Food Sci., 1994; 59:544–547.
72. Kim, H. D., Lee, J. H., Shin, Z. I., Nam, H. S., andWoo, H. J., Anticancer effects of hydrophobic pep-tides derived from a cheese slurry, Foods Biotechnol.,1995; 4: 268–272.
73. Fox, P. F., Singh, T. K., and McSweeney, P. L. H.,Biogenesis of flavour compounds in cheese. Chemis-try of Structure-Function Relationships in Cheese,Malir, E. L. and Tunick, M. H., Eds., Plenum Press,New York, 1995, 59–98.
74. Monnet, V., Bockelmann, W., Gripon, J.-C., andTeuber, M., Comparison of cell-wall proteinases fromLactococcus lactis subsp. lactis AC1 and Lactococcuslactis subsp. lactis NCDO 763. I. Specificity towardsβ-casein, Appl. Microbiol. Biotechnol., 1989; 31: 112–118.
75. Stepaniak, L., Gobbetti, M., Sørhaug, T., Fox, P. F.,and Højrup, P., Peptides inhibitory to endopeptidaseand aminopeptidase from Lactococcus lactis ssp. lactisMG 1363, released from bovine sy β-casein bychymosin, trypsin or chymotrypsin, Z. Lebensm. UntersForsch., 1996; 202: 329–333.
76. Visser, S., Proteolytic enzymes and their relation tocheese ripening and flavor: an overview, J. Dairy Sci.,1993; 76: 329–350.
77. Andrews, S.A.T. and Varley, J., Biochemistry of MilkProducts, Royal Society of Chemistry, Cambridge,UK. 1994; Special publication n. 150.
78. Haileselassie, S.S., Lee, B.H., and Gibbs, B.F., Puri-fication and identification of potentially bioactive pep-tides from enzyme-modified cheese, J. Dairy Sci.,1999; 82: 8, 1612–1617.
79. Wenzel, H. R., and Tschesche, H., Reversibleinhibitors of serine proteinases. In: Peptides: Syn-
thesis, Structures and Applications, Gutte, B.,Ed., Academic Press Inc., London, 1995, 321–361.
80. García-Carreño, F. L., Proteinase inhibitors, TrendsFood Sci. Technol. 1996; 7: 197–204.
81. Gobbetti, M. and Stepaniak, L., Peptides from cheeseinhibitory to intracellular endo- and amino-peptidaseactivity of starters: different sensitivity of lactic acidbacteria and Propionibacterium. In: ProceedingsCheese Maturation Congress, Holland College Ed.,Delft, Europel. 1995, 1–9.
82. Gobbetti, M., Stepaniak, L., Fox, P. F., Sørhaug, T.,and Tobiassen, R., Inhibition of endo- and amino-peptidase activities in cytoplasmic fractions ofLactococcus, Lactobacillus and Propionibacterium bypeptides from different cheese, Milchwissenschaft,1995; 50: 565–570.
83. De Vos, W. M. and Siezen, R. J., Engineering pivotalproteins for lactococcal proteolysis. In: Biochemistryof Milk Products, Andrews, A. T., and Varley, J.,Eds., The Royal Soc. Chemistry London, 1994, 56–71.
84. Gobbetti, M., Smacchi, E., Corsetti, A., and Bellucci,M., Inhibition of proteolytic enzymes from Pseudomo-nas fluorescens ATCC 948 and of angiotensin I-con-verting enzyme by peptides from zein, hordein andgluten hydrolyzates. J. Food Protect., 1997; 60, 499–504.
85. Schmidl, M.K., Food products for medical purposes,Trends Food Sci. Technol., 1993; 4: 163–168.
86. De Felice, S.L., The nutritional revolution: its impacton food industry R&D, Trends Food Sci Technol.,1995; 6: 59–61.
87. Kunst, A., Process to isolate phosphopeptides, Euro-pean Patent Application 0 467 199 A., 1992.
88. Brule, G., Roger, L., Fauquant, J., and Piot, M.,Phosphopeptides from casein-based material, U.S.Patent 1982; 4: 358–465.
89. Reynolds, E., Phosphopeptides, PCT Internationalpatent Application WO 87/07615 A1, 1987.
90. Carnie, J., Minter, S., Oliver, S., Perra, F., andMetzlaff, M., Nutritional compositions containig β-casomorphins. UK Patent Application GB 2214810A 1989.
91. Sekyja, S., Kobayashi, Y., Kita, E., Imamura, Y., andToyama, S., Antihypertensive effects of tryptic hy-drolysates of casein on normotensive and hyperten-sive volunteers (in Jpn), J. Japan. Soc. Nutr. FoodSci., 1992; 45: 513–517.
92. Hadden, J.W., Immunotherapy of human immunode-ficiency virus infection, Trends Pharmac. Sci. 1991;12: 107–111.
93. Froetschel, M.A., Bioactive peptides in digesta thatregulate gastrointestinal function and intake, J. Ani-mal Sci., 1996; 7410.: 2500–2508.
94. Steinmetzer, T., Slberring, J., Mrestani-Klaus, C.,Fittkau, S., Barth, A., and Demuth, HU, Peptidylammonium methyl ketones as substrate analog inhibi-
Dow
nloa
ded
by [
Uni
vers
ità d
egli
Stud
i di M
ilano
] at
05:
42 0
5 N
ovem
ber
2012

239
tors of proline-specific peptidases, J. Enzyme Inhibi-tion, 1993; 7: 77–85.
95. Saito, Y., Ohura, S., Kawato, A., Suginami, K., Prolylendopeptidase unhibitors in sake and its by-products,J. Agric. Food Chem., 1997; 45 3: 720–724.
96. Maruyama, S., Miyoshi, S. Kaneko, T., and Tanaka,H., Angiotensin I-converting enzyme inhibitory ac-tivities of synthetic peptides related to the tandemrepeated sequence of a maize endosperm protein, Agric.Biol. Chem., 1989; 53: 1077–1081.
Dow
nloa
ded
by [
Uni
vers
ità d
egli
Stud
i di M
ilano
] at
05:
42 0
5 N
ovem
ber
2012
Copyright © 2022 FDOKUMEN




















