
Late Weichselian Marine 14C Reservoir Ages at the Western Coast of Norway
11
Late Weichselian Marine 14 C Reservoir Ages at the Western Coast of Norway Stein Bondevik Department of Geology, University of Bergen, Alle ´gaten 41, N-5007 Bergen, Norway Hilary H. Birks Botanical Institute, University of Bergen, Alle ´gaten 41, N-5007 Bergen, Norway Steinar Gulliksen Radiological Dating Laboratory, NTNU, Sem Sælandsvei 5, N-7034 Trondheim, Norway and Jan Mangerud Department of Geology, University of Bergen, Alle ´gaten 41, N-5007, Bergen, Norway Received October 28, 1998 A shallow marine Late Weichselian deposit on the outer coast of western Norway contains both terrestrial plant material and articu- lated marine shells. We have 14 C dated both types of material from eight different stratigraphic levels covering the time interval 12,300 to 11,100 14 C yr B.P. The difference between 14 C-dated terrestrial plant material and marine shell material (the marine reservoir age) ranges from 200 to 525 yr, with a weighted average of 380 6 32 yr. This is almost identical to the present reservoir age of 379 6 20 yr for southern Norway. In the mid-Younger Dryas (YD) interval the res- ervoir age in the North Atlantic (55°N– 65°N) was 700 – 800 yr, considerably greater than the present reservoir age and the age we have measured for the Bølling–Allerød interval. The reason for this increase during the YD is probably a combination of reduced inflow of surface waters to the North Atlantic and more extensive sea-ice cover. Evidence from marine cores show that the southeastern Nor- wegian Sea experienced rapid fluctuations in the inflow of warm Atlantic surface water during the period 12,300 to 11,000 yr B.P. However, the reservoir age apparently did not increase during these colder periods (Older Dryas I and II). The reason is probably that, in contrast to the YD, these colder periods did not last long enough and/or were of too limited extent to alter the reservoir age of the ocean. A comparison of the obtained 14 C dates with the varve 14 C chronology from Lake Suigetsu indicates an age of 12,770 cal yr B.P. for the AL/YD boundary. © 1999 University of Washington. Key Words: Late Weichselian; shallow marine deposit; 14 C reservoir age; western Norway. INTRODUCTION Radiocarbon dating is now the most important tool for establishing precise time frames for Late Weichselian (Wis- consin) and Holocene deposits. A precise correlation between climate records or other events identified in marine, terrestrial, and ice-core successions is a priority objective for ongoing and future research (e.g., Lowe and NASP Members, 1995; Hafli- dason et al., 1995). Such correlations rely mainly on the radiocarbon method and require a correction of marine 14 C dates due to the reservoir age of the ocean. The reservoir age is closely related to ocean circulation (Stuiver et al., 1983). Today the inflow of warm Atlantic surface water to the North Sea and Norwegian Sea results in a reservoir age of ca. 400 –500 yr in the North Atlantic. Changes in inflow of Atlantic water through time may have influenced the reservoir age in this area (Mangerud and Gulliksen, 1975). Foraminifera and shell fragments within the Vedde Ash (mid- Younger Dryas) in North Atlantic cores are 700 – 800 14 C yr older than terrestrial plant material within the same Ash bed in lakes (Koc ¸ Karpuz and Jansen, 1992; Bard et al., 1994; Austin et al., 1995; Haflidason et al., 1995). Therefore, it has been concluded that the reservoir age was greater by 300 – 400 yr during the Younger Dryas. Ash beds are synchronous time markers and are ideal for comparing 14 C ages for contemporaneous marine and terrestrial environments. However, in the Norwegian Sea and North Atlantic only the Vedde Ash bed (ca. 10,300 yr B.P.) and the Saksunarvatn Ash bed (ca. 9000 yr B.P.) (Birks et al., 1996) are available for this purpose during the Late Weichselian time frame. To overcome these limitations, we have dated paired samples of marine molluscs and terrestrial plant material in cores from a shallow marine deposit at Kulturmyra at the westernmost coast of Norway (Fig. 1). Quaternary Research 52, 104 –114 (1999) Article ID qres.1999.2048, available online at http://www.idealibrary.com on 0033-5894/99 $30.00 Copyright © 1999 by the University of Washington. All rights of reproduction in any form reserved. 104
Transcript of Late Weichselian Marine 14C Reservoir Ages at the Western Coast of Norway

Late Weichselian Marine 14C Reservoir Ages
wle1mfasechiocwAHccaocf
r
e
Quaternary Research52, 104–114 (1999)Article ID qres.1999.2048, available online at http://www.idealibrary.com on
0CA
at the Western Coast of Norway
Stein Bondevik
Department of Geology, University of Bergen, Alle´gaten 41, N-5007 Bergen, Norway
Hilary H. Birks
Botanical Institute, University of Bergen, Alle´gaten 41, N-5007 Bergen, Norway
Steinar Gulliksen
Radiological Dating Laboratory, NTNU, Sem Sælandsvei 5, N-7034 Trondheim, Norway
and
Jan Mangerud
Department of Geology, University of Bergen, Alle´gaten 41, N-5007, Bergen, Norway
Received October 28, 1998
fois
consin) and Holocene deposits. A precise correlation betweenc trial,a andf afli-d herd
tion( ics s in ar gesi edt 975).F id-Yo d inle enc 00 yrd
al forc striale orthA theSa timef aireds al inc thew
A shallow marine Late Weichselian deposit on the outer coast ofestern Norway contains both terrestrial plant material and articu-
ated marine shells. We have 14C dated both types of material fromight different stratigraphic levels covering the time interval 12,300 to1,100 14C yr B.P. The difference between 14C-dated terrestrial plantaterial and marine shell material (the marine reservoir age) ranges
rom 200 to 525 yr, with a weighted average of 380 6 32 yr. This islmost identical to the present reservoir age of 379 6 20 yr forouthern Norway. In the mid-Younger Dryas (YD) interval the res-rvoir age in the North Atlantic (55°N–65°N) was 700–800 yr,onsiderably greater than the present reservoir age and the age weave measured for the Bølling–Allerød interval. The reason for this
ncrease during the YD is probably a combination of reduced inflowf surface waters to the North Atlantic and more extensive sea-iceover. Evidence from marine cores show that the southeastern Nor-egian Sea experienced rapid fluctuations in the inflow of warmtlantic surface water during the period 12,300 to 11,000 yr B.P.owever, the reservoir age apparently did not increase during these
older periods (Older Dryas I and II). The reason is probably that, inontrast to the YD, these colder periods did not last long enoughnd/or were of too limited extent to alter the reservoir age of thecean. A comparison of the obtained 14C dates with the varve 14Chronology from Lake Suigetsu indicates an age of 12,770 cal yr B.P.or the AL/YD boundary. © 1999 University of Washington.
Key Words: Late Weichselian; shallow marine deposit; 14Ceservoir age; western Norway.
INTRODUCTION
Radiocarbon dating is now the most important toolstablishing precise time frames for Late Weichselian (W
033-5894/99 $30.00opyright © 1999 by the University of Washington.ll rights of reproduction in any form reserved.
104
r-
limate records or other events identified in marine, terresnd ice-core successions is a priority objective for ongoing
uture research (e.g., Lowe and NASP Members, 1995; Hason et al., 1995). Such correlations rely mainly on tadiocarbon method and require a correction of marine14Cates due to the reservoir age of the ocean.The reservoir age is closely related to ocean circula
Stuiver et al., 1983). Today the inflow of warm Atlanturface water to the North Sea and Norwegian Sea resulteservoir age of ca. 400–500 yr in the North Atlantic. Chann inflow of Atlantic water through time may have influenche reservoir age in this area (Mangerud and Gulliksen, 1oraminifera and shell fragments within the Vedde Ash (mounger Dryas) in North Atlantic cores are 700–80014C yrlder than terrestrial plant material within the same Ash be
akes (KocKarpuz and Jansen, 1992; Bardet al.,1994; Austint al., 1995; Haflidasonet al., 1995). Therefore, it has beoncluded that the reservoir age was greater by 300–4uring the Younger Dryas.Ash beds are synchronous time markers and are ide
omparing14C ages for contemporaneous marine and terrenvironments. However, in the Norwegian Sea and Ntlantic only the Vedde Ash bed (ca. 10,300 yr B.P.) andaksunarvatn Ash bed (ca. 9000 yr B.P.) (Birkset al., 1996)re available for this purpose during the Late Weichselian
rame. To overcome these limitations, we have dated pamples of marine molluscs and terrestrial plant materiores from a shallow marine deposit at Kulturmyra atesternmost coast of Norway (Fig. 1).

001 apifl tota iss d ti rvoa rvaw anc
asil steN boa aftd sl yr
w hore.T erial( them basinfl mosss on int y thes
mi-n theN has at TheN s am ea,a aterm thes . AtB f thes e
sea coresm
LATE WEICHSELIAN MARINE 14C RESERVOIR AGES 105
The Kulturmyra sequence includes the time interval 12,31,100 yr B.P. (14,500–12,800 cal yr B.P.), a period with ructuations of the inflow of warm Atlantic surface watershe Norwegian Sea (Lehman and Keigwin, 1992; Koc¸ Karpuznd Jansen, 1992; Haflidasonet al.,1995). The purpose of thtudy was to measure the marine reservoir age directly anf these rapid fluctuations had any influence on the resege of the Norwegian Sea. Our main result is that the resege for the Bølling–Allerød interval was 3806 32 yr on theestern coast of Norway, close to the present value,onsiderably less than the Younger Dryas reservoir age.
SETTING
The studied site, Kulturmyra, is a 7- to 9-m-deep bocated on the strandflat at the outermost coast of weorway (Figs. 1 and 2). Today the surface of the basin is at 2.5 m altitude. Between 13,000 and 11,000 yr B.P.,eglaciation, the threshold of the basin was 2–5 m below
evel (Svendsen and Mangerud, 1990). At that time Kulturm
FIG. 1. Map of the Norwegian Sea region showing the present surfaentioned in the text (HM 79-6/4; Koc¸ Karpuz and Jansen, 1992; and Tr
–d
estir
oir
d
nrng
ereaa
as a shallow marine basin located ca. 300 m from the she proximity to the shore enabled terrestrial plant matTPM) derived from the surroundings to be deposited inarine sediments, together with articulated shells on the
oor. The dated plant fragments are mostly leaves andtems very unlikely to have been reworked before depositihe basin. We assume that the TPM has approximatelame real ages as the molluscs.Today Atlantic Water and Norwegian Coastal Water do
ate along the Norwegian coast. Atlantic Water entersorwegian Sea in the Faeroe–Shetland area (Fig. 1); it
emperature of about 9°C and a salinity above 35‰.orwegian Coastal Water, with salinities below 35‰, iixture of Atlantic water, brackish water from the Baltic Snd runoff from the Norwegian mainland. The Coastal Wixes with deeper Atlantic Water in such a way that
alinity along the Norwegian coast increases northwardud, a permanent hydrographic station ca. 100 km north otudied site, summer temperatures in water,50 m deep ar
circulation, the location of Kulturmyra, and the location of the two deep.1; Lehman and Keigwin, 1992).
ceoll 3

1 ityb nse1
h ad ugm lest foe idl dm BeS ofv -i t 1m s oT enfi asp neo Ms eins bol nlya rt o
t d(
werep im.T trom-e salaU por-ta r andP
n EC1 ea-s cm-d aF
rt oft ing-u rine,a
. 3).T ndl tent
arine andl ., fora
BONDEVIK ET AL.106
1–12°C, and winter temperatures are 5–6°C. The salinetween 33.0 and 33.5‰ year around (Aure and Øste993).
METHODS
A piston corer provided 2-m-long core sections witiameter of 11 cm. To make certain we obtained enoaterial for dating, three parallel cores were collected
han 1 m apart. The cores were X-ray radiographedxamination of sedimentary structures and as an a
ocate articulated shells. The cores were easily correlateatching sedimentary structures and the Vedde Ashample intervals ranged from 1 to 6 cm and consistedolume of 100 to more than 500 cm3 of sediment, consumng three halves of the cores. This material was sieved am after soaking in water overnight. The componentPM were selected under a stereo-microscope and ided. Only material of undoubted terrestrial origin wicked for radiocarbon dating. The fragments were cleaf silt and other contamination and air-dried. All TPamples were more than 12 mg dry weight before bubmitted for preparation in the Trondheim radiocaraboratory (Table 1). From the marine material (MM) orticulated shells were used for dating. In the lower pa
FIG. 2. Kulturmyra is located on Bergsøya at the outer Norwegianacustrine deposits and supports bog vegetation. Kulturmyra was a shallfter deglaciation now lies at ca. 20 m altitude on Bergsøya.
isn,
hsrtobyd.a
25fti-
d
gn
f
he core a cod bone (Gadus morhua) was found and dateTable 1, Fig. 3).
Both TPM and MM samples were prepared and targetsroduced at the Radiological Dating Laboratory in Trondhehe 14C content was measured by accelerator mass spectry (AMS) analysis at the T. Svedberg Laboratory, Uppniversity, Sweden (TUa numbers, Table 1). The outer
ions (.23%) of the shell samples were dissolved. All14C agesre reported as conventional radiocarbon ages (Stuiveolach, 1977).The total organic carbon content was determined with a
2 LECO carbon analyzer. Magnetic susceptibility was mured with a Bartington MS2 instrument using a 12-iameter ring sensor.d18O and d13C were measured withinnigan MAT 251 mass spectrometer.
RESULTS
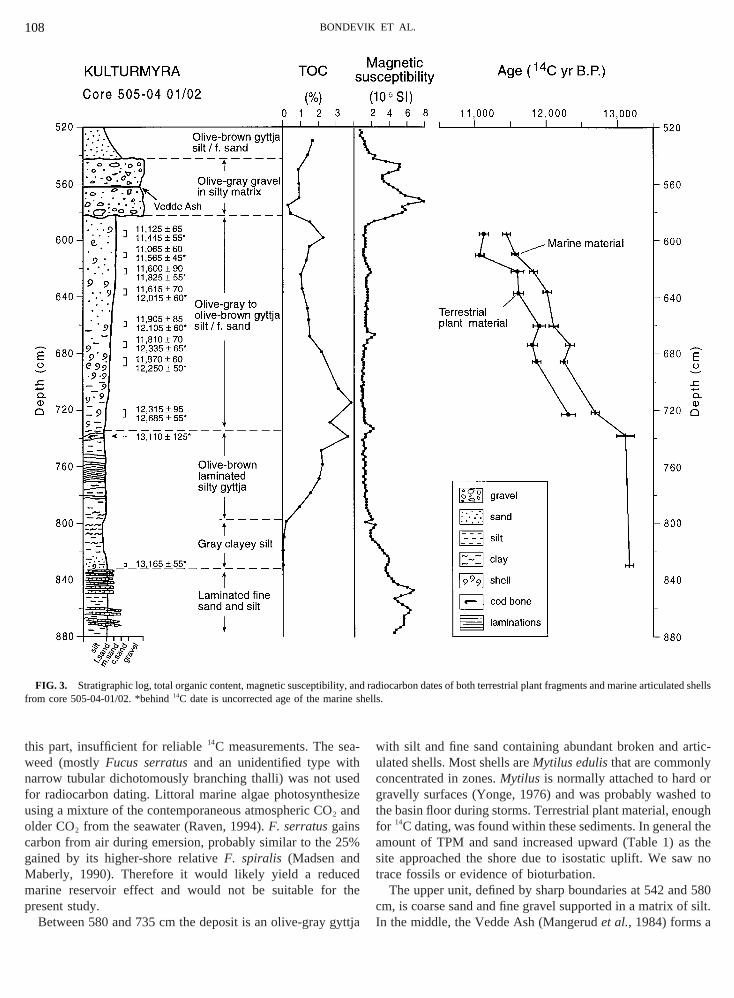
The Late Weichselian deposits are found in the lower pahe stratigraphy of the basin. They are part of a shallowpward sequence from glacialmarine, through shallow mand into lacustrine sediments.The studied sequence can be divided into four units (Fig
he lower, from 797 to 880 cm, is a gray silty clay with saaminae having a very low total organic carbon (TOC) con
st close to the open Norwegian Sea. Today the basin is filled in with mmarine basin ca. 12,700–10,000 yr B.P. The shoreline of 12,700 yr B.Pmed just
coaow

( np teM ana t1 oir
a f thea
owns ofs d in
TABLE 1
T
T
T
T
T
T
T
T
T
T
T
T
T
T
T
T
TT
LATE WEICHSELIAN MARINE 14C RESERVOIR AGES 107
Fig. 3). These sediments are interpreted as having beeosited during and immediately after deglaciation. Articulaacoma calcareaat the transition between the laminated snd the overlying silty clay at 828–829 cm was dated3,1656 55 yr B.P. (TUa-1902, not corrected for reserv
Radiocarbon Ages of Terrestrial Plant Material and
Laboratorynumber
Depth(cm)
Sed. vol.(cm3) Material dated
Ua-1896 593–597 150 TPM: Leaves ofSalix herbacea, S. polaris, DryaSalix budscale and capsule. Seeds ofD. octopeCarex trigonous,Carex nigratype,Comarum pTerrestrial mosses (Racomitrium, PolytrichumsDrepanocladus uncinatus, Fissidens cristatupalustre, A. turgidum, Distichium capillaceum
Ua-1891 593–597 MM:Litorina littorea
Ua-1897 609–611 100 TPM: Leaves ofSalix herbacea, S. polaris, Dryaviviparum. Salixtwig, bud, budscale, capsuleCochlearia,Gramineae,Carex lenticular,SileneTerrestrial mosses (Racomitrium, Polytrichum,Fissidens osmundoides, Aulacomnium turgid)
Ua-1892 608–610 MM: ArticulatedMytilus edulis
Ua-1898 620–623 351 TPM: Leaves ofSalix herbacea, S. polaris, Dryaviviparum, Saxifraga oppositifolia, Salixvegetaof Dryas, Cochlearia,Gramineae,Saxifraga opJuncus.Terrestrial mosses (Racomitrium, PolyPohlia, Dicranum).
Ua-1893 620–623 MM: ArticulatedMytilus edulis
Ua-1885 635–639 435 TPM: Leaves ofSalix herbacea, Dryas, Salixbud,viviparumbulbil, seeds ofSaxifraga cespitosaintermedia, S. saginoidestype,Juncus,GramineTerrestrial mosses (Racomitrium, Polytrichum,Drepanocladus uncinatus, Dicranum, AulacoRhytidiadelphus squarrosus, Tortulacf. norveg
Ua-1886 635–637 MM: ArticulatedMytilus edulis
Ua-1899 659–661 200 TPM: Leaves ofSalix herbacea, S. polaris, Dryaviviparum, Salixtwig, budscales, Seeds ofDryaCarex trigonous. Terrestrial mosses (RacomitriuBryum, Drepanocladus uncinatus, Dicranum).
Ua-1894 659–660 MM: ArticulatedMytilus edulis
Ua-1887 672–675 335 TPM: Leaves ofSalix herbacea, Dryas, PolygonSalix bud, budscale, capsule, Seeds ofDryas,GCarex trigonous,Sagina intermedia.Terrestrial(Racomitrium, Polytrichum, Bryum, Dicranum).
Ua-1888 672–675 MM: ArticulatedMytilus edulis
Ua-1900 683–688 475 TPM: Leaves ofSalix herbacea, S. polaris, Dryaviviparum. Salixfemale flower. Seeds ofDryasCochlearia.Terrestrial mosses (Racomitrium, PBryum, Pohlia, Fissidens).
Ua-1895 683–688 MM: ArticulatedMytilus edulis
Ua-1889 720–726 580 TPM: Leaves ofSalix polaris, Dryas, Salixbud, bOxyria fruit. Terrestrial mosses (Racomitrium,juniperinum, Polytrichumsp.,Bryum, AulacomTortula cf. norvegica).
Ua-1890 721–722 MM: ArticulatedMytilus edulis
Ua-1901 738 Jawbone from Cod (Gadus morhua L.)Ua-1902 828–829 MM: ArticulatedMacoma calcarea
* Assumed value, not measured.
de-ddo
ge, Table 1) and gives a minimum age for delgaciation orea.The next unit, between 735 and 797 cm, is an olive-br
ilty gyttja with higher TOC values and many fine laminaeeaweed. Only a few terrestrial plant fragments were foun
rine Material from Core 505-04-01/02, Kulturmyra
Dry weight(mg)
Sample size(mg C)
14C age(yr B.P.)
Reservoir ageMM–TPM
d13C(‰ PDB)
ctopetala,, Cochlearia,stre.Bryum,ulacomnium
18.0 1.4 11,1256 65 224.5
1.9 11,4456 55 3206 85 1.3
olygonumeds ofaulis.um,
35.5 2.1 11,0656 60 226.5
2.0 11,5656 45 5006 75 1.9
olygonumparts, seedsitifolia,um, Bryum,
14.2 1.4 11,6006 90 225.8
2.1 11,8256 55 2256 105 20.4
ygonumginaarex.
um,ium turgidum,
13.0 1.6 11,6156 70 226.5
1.8 12,0156 60 4006 92 0.8
olygonumramineae,Polytrichum,
13.7 1.5 11,9056 85 223.5
3.1 12,1056 60 2006 104 0.7
viviparum.ineae,sses
12.8 1.7 11,8106 70 226.0
0.9 12,3356 65 5256 96 2.6
olygonummineae,
trichum,
19.7 2.6 11,8706 60 223.9
2.1 12,2506 50 3806 78 1.2
cales,trichum
turgidum,
6.5 1.2 12,3156 95 227.8
2.2 12,6856 55 3706 110 1.58
2.3 13,1106 125 15.0*0.5 13,1656 55 0.2
Mean and standard deviation 3656 116Weighted mean 3806 32
Ma
s otalaalu
pp.,s, A).
s, P. Se
acBryum.
s, Ptivepos
trich
Pol, Saae,CBrymn
ica).
s, Ps,Gm,
umrammo
s, P,Graoly
udsPolynium
t a-w thn sef sizuoc 5%g dM edm thp
ytt
w rtic-u yc org ed tot ughf l thea s thes w not
580c f silt.I
laf
BONDEVIK ET AL.108
his part, insufficient for reliable14C measurements. The seeed (mostlyFucus serratusand an unidentified type wiarrow tubular dichotomously branching thalli) was not u
or radiocarbon dating. Littoral marine algae photosynthesing a mixture of the contemporaneous atmospheric CO2 andlder CO2 from the seawater (Raven, 1994).F. serratusgainsarbon from air during emersion, probably similar to the 2ained by its higher-shore relativeF. spiralis (Madsen anaberly, 1990). Therefore it would likely yield a reducarine reservoir effect and would not be suitable forresent study.Between 580 and 735 cm the deposit is an olive-gray g
FIG. 3. Stratigraphic log, total organic content, magnetic susceptibilitrom core 505-04-01/02. *behind14C date is uncorrected age of the marin
de
e
ja
ith silt and fine sand containing abundant broken and alated shells. Most shells areMytilus edulisthat are commonloncentrated in zones.Mytilus is normally attached to hardravelly surfaces (Yonge, 1976) and was probably wash
he basin floor during storms. Terrestrial plant material, enoor 14C dating, was found within these sediments. In generamount of TPM and sand increased upward (Table 1) aite approached the shore due to isostatic uplift. We sarace fossils or evidence of bioturbation.
The upper unit, defined by sharp boundaries at 542 andm, is coarse sand and fine gravel supported in a matrix on the middle, the Vedde Ash (Mangerudet al.,1984) forms a
nd radiocarbon dates of both terrestrial plant fragments and marine articuted shellshells.
y, ae s

distinct 2-cm-thick bed. The lower boundary was interpreted tob ld.A mi ent posi thi 95
res ef but tht nM ew htea
toL M( ent 80f iths tchi ageo them1 en1f ,0a inew itt lieb ala ref ao mn agf r.
R
d tM ths n ot ad ThT fram s tl oulh gb asw olTo nr ort
to the basin floor from the rocky sides of the basin. Onlya havea enceb e isc rangew
V
le 1),a 00 yrB hicht mon-tD andh eN obli-gna tad,V hera at oft un-t mert n ofr entm eanJ ) fort 5 kmn
theY asa cticdT asa tot ast ofG
soc pidi d ice-r sea( 2;H ca.1 (ODI TheS Nor-w
uth-w aus,1 notr ationa andB atec f Kul-t the
LATE WEICHSELIAN MARINE 14C RESERVOIR AGES 109
e the Allerød/Younger Dryas (AL/YD) boundary in the fie14C date of 11,1256 65 yr B.P. (TUa-1896, Table 1) fro
mmediately below the lower contact confirms that it represhe AL/YD boundary. The coarse sediment likely was deted by local sea ice, which transported material fromntensely frost-weathered YD shore (Blikra and Longva, 19
The eight pairs of14C dates for the TPM and the MM ahown relative to depth in Figure 3. The MM14C dates arrom 200 to 525 yr older than the TPM dates (Table 1),here is no obvious correlation of age as a function of dephe studied sequence. The mean of the difference betweeM and TPM dates is 3656 116 yr. If the differences areighed according to their standard deviations, a weigverage of 3806 32 yr is obtained.The radiocarbon ages were matched (Pearson, 1986)
ake Suigetsu14C chronology built from AMS dates of TPKitagawa and van der Plicht, 1998) by varying the sedimation rate and and using the weighted reservoir age of 3or the marine dates (Fig. 4). The best fit was obtained wedimentation rate of 0.105 cm/year. According to this mang, the AL/YD boundary at 580 cm in Kulturmyra has anf 12,770 cal yr B.P. In contrast, the AL/YD boundary inarine varved deposits in the Cariaco basin (Hughenet al.,998), shown by gray-scale measurements, occur betwe3,050 and 12,900 cal yr B.P. When matching the14C dates
rom the Cariaco basin with the Suigetsu data between 15nd 12,000 cal yr B.P., we found that the best fit is obtahen 128 yr is subtracted from the Cariaco time scale. W
his adjustment, the AL/YD boundary in the Cariaco basinetween 12,770 and 12,940 cal yr B.P., very close to the cge boundary we found for Kulturmyra. In the GRIP ice co
rom the Greenland ice sheet, the AL/YD boundary has anf 12,705 ice-core yr B.P (J. P. Steffensen, personal comication). However, the establishment of a precise cal yr
or the AL/YD boundary is beyond the scope of this pape
DISCUSSION
eal Age Difference between Samples in a Pair
Our study depends on the assumption that the TPM anM had no age difference when deposited together in
hallow marine basin. The TPM derives from the vegetatiohe hillside surrounding the basin and was transportedeposited in the basin, together with the marine shells.PM consists of tiny leaves, seeds, and delicate mossents that are very unlikely to have been in transit acros
and surface for a long time before deposition. If so, they wave been decomposed and destroyed before reachinasin. We assume that the TPM, when deposited in the bas probably formed that year and was no more than 2 yrhe dated MM consists of seven samples ofMytilus edulisandne sample ofLittorina littorea. Both species normally live oocky surfaces, and so the shells were most likely transp
ts-
e).
tinthe
d
the
-yra-
ca.
00dhsyrsgeu-e
heen
ndeg-hedthein,d.
ed
rticulated shells were dated, and they probably did notny significant age when deposited. The real age differetween the TPM and the MM within the same samplonsidered to be less than 30–50 yr, the possible ageithin a 3- to 5-cm-thick sample (Table 1).
egetation and Climate
The plant assemblages, based on the macrofossils (Tabre similar through the sequence between 12,300 and 11,1.P. (14,500–12,770 cal yr B.P.). The vegetation from w
hey originated was treeless and dominated by arctic andane taxa. The prostrate shrubsSalix herbacea, S. polaris,andryas octopetalawere associated with sedges, grasses,erbs such asPolygonum viviparumthat are common in thorwegian mountains today. Among the mosses are theate montane taxaAulacomnium turgidumand Tortula cf.orvegica. A. turgidumandSalix herbaceanow occur as lows 430 m altitude on the coastal mountains of Møre (Sågsøy, ca. 25 km south of Kulturmyra), but at much higltitudes inland. The Kulturmyra assemblage resembles th
he low- and mid-alpine vegetation in the Norwegian moains today. This correlates with a cool climate and sumemperatures of 5–7.5°C (see Birks (1994) for a discussioegional Allerød climate in western Norway). The presean July temperature at Kulturmyra is ca. 13°C, and the m
anuary temperature is ca. 2.5°C (values from Aune (1993he nearest climate stations at Ålesund and Vigra, ca. 2orth of Kulturmyra).Mytilus edulis is present throughout the core below
D/AL boundary. The presence ofM. edulishas been usedn indicator of warmer-than-present conditions in the Aruring the Holocene climatic optimum (Hjortet al., 1995).odayM. edulis is not found in higher Arctic waters, suchround Svalbard (Hjortet al., 1995), but it penetrates far in
he southeastern Barents Sea and along the western coreenland (Peacock, 1989).Two colder periods, Older Dryas I and II (OD II is al
alled the inter-Allerød cold period), are evident from a rancrease in polar fauna, sea-ice diatom assemblage, anafted debris in cores from the southeastern NorwegianLehman and Keigwin, 1992; Koc¸ Karpuz and Jansen, 199aflidasonet al., 1995). The events have been dated to2,300–12,000 yr B.P. (OD I) and 11,800–11,500 yr B.P.
I) (the ages were corrected for a reservoir age of 440 yr).candinavian ice sheet advanced during OD I along theegian coast (Mangerud, 1970, 1977).Indications of two colder periods in pollen records in soestern Norway are inferred to represent OD I and II (P988). However, macrofossil studies in that area haveevealed any small oscillatory changes in the local vegett that time (Birks, 1993, and unpublished data; van Dinterirks, 1996, and unpublished data). Similarly, no climhanges were detected at Kråkenes to the southwest ourmyra for this time, either in the local vegetation or in

a dv hab nsT tha set maa het seb to ts w-a tiof semb thi thet hel ged onap rvas d.
D ns
tea bo1 ity
r llym rf wasi s notr edd andc renti ela-ts1
arec -m d att andK fam gure5 lciticl e pre-p
da ,1
er Plicht,1 he boundarb
BONDEVIK ET AL.110
quatic ecosystem (Birkset al.,in press). However, the Allerøegetation along the western Norwegian coast may noteen sensitive to small, brief (ca. 200 yr) climatic oscillatiohe vegetation was well developed and was analogous tobove the tree line in Norway today. Betula trees were ab
heir nearest occurrences being in southern Sweden, Dennd the British Isles (van Dinter and Birks, 1996). Ot
hermophilous plant taxa were also absent. Allerød aslages of aquatic animals and diatoms at Kråkenes Lakeouth of Kulturmyra consisted of taxa typical of cool, lolpine climates, and quantitative temperature reconstruc
rom pollen, chironomid, cladocera, and coleoptera aslages from the Allerød at Kråkenes are all consistent with
nterpretation (Birkset al., in press). This suggests thaterrestrial vegetation was in equilibrium with the climate. Tack of evidence at Kulturmyra for local vegetation chanuring the studied time period is consistent with this regiicture. The samples are widely spaced at ca. 150-yr inteo short fluctuations (;200 yr) may not have been sample
o the Marine Samples Represent Open Ocean Conditio
Today the difference in salinity between the coastal waround Kulturmyra and the open Norwegian Sea is a–2‰. Mytilus edulis tolerates an extremely wide salin
FIG. 4. Radiocarbon ages with 1s error from Kulturmyra core 505-04-0998). A reservoir age of 380 yr was subtracted from the marine ages.etween AL/YD is at 580 cm and shows a calendar age of 12,770 yr B
ve.at
nt,rk,
rm-he
ns-
is
slls,
?
rsut
ange, from 4 to 5‰ to above 35‰ (inner Baltic to fuarine conditions). (Bayneet al.,1976). To check if the wate
rom the area where the shells precipitated the carbonatenfluenced by any fresh water, and for that reason doeecord thetrue 14C reservoir age of the ocean, we perform18O measurements on the shells. Isotopic ratios of oxygenarbon can be related to salinity because of the diffesotopic composition of ocean and fresh water. A linear rionship has been demonstrated betweend18O in M. edulisandalinity in the seawater where the shells calcified (Ingramet al.,996; Mook, 1971).Thed18O values obtained from the shells in Kulturmyra
ompared in Figure 5 withd18O values from planktonic forainifera in the Norwegian Sea and North Sea that calcifie
he same time (Koc¸ Karpuz and Jansen, 1992; Lehmaneigwin, 1992). The mussel shell (M. edulis) is composed on outer calcitic layer and an inner aragonitic layer. Thed18Oeasurements of the shells from Kulturmyra shown in Fiare from the inner aragonitic layer, because the outer ca
ayer had already been dissolved when the samples werared for the14C measurements.Aragonite is enriched ind18O relative to calcite precipitate
t the same temperature (Tarutaniet al., 1969; Grossman984). The average difference betweend18O from Mytilus and
2 curve-matched to the Lake Suigetsu chronology (Kitagawa and van dt fit was obtained with a constant sedimentation rate of 0.105 cm/yr. Ty
1/0Bes.P.

f ou2 em -t set
as 1).B avd ordva er LaW . Td eas relat hat urs edt rap
during spring and summer and slow or absent during the colderw frac-t thef d bea day.T withv nedr
inr thei n thes s int havei
V
thed ne-o era pen-d rates)ra At-l tingm 900.Ws t thei oms thec ivingh ta e be-t ownh ver,d t thec ange8 aver e fors 0)e sti se-q e dif-f strials egianc oft
thei ea.H dis-t in,1 ftedd yrB ryas( ns.
i win1 terNe ocf tot hellc orwg out2 r tht inpuo .
LATE WEICHSELIAN MARINE 14C RESERVOIR AGES 111
oraminifera from the Norwegian Sea (HM 79-6/4) is ab.25‰ d18O (Fig. 5). Because thed18O measurements of thussels are from aragonite and thed18O values of the plank
onic foraminifera are from calcite, the real difference is cloo 3‰ d18O (Tarutaniet al., 1969).
The present salinity (S)–d18O relationship for the North Sehows a S:d18O ratio of 1:;0.3 (Israelson and Buchardt, 199ecause of cold climate and retreating glaciers in Scandinuring the AL/BØ, fresh water must have been much mepleted ind18O than today. A change to more negatived18Oalues in fresh water would increase the S:d18O ratio (Rohlingnd Bigg, 1998). A ratio of S:d18O 5 1:0.5–0.6 would be moreasonable for the North Sea/Norwegian Sea during the
eichselian (E. Jansen, personal communication, 1998)eviation of d18O from foraminifera in the Norwegian Suggests that the salinity at Kulturmyra, using the linearion S:d18O 5 1:0.6, would have been about 4–6‰ less that in the open ocean. However, surface temperatures dummers were probably higher in coastal waters comparhe open ocean, as they are today. Growth in shells is
FIG. 5. d18O isotopes fromM. edulisfrom Kulturmyra compared tod18Osotopes from planktonic foraminifera from Troll 3.1 (Lehman and Keig992) and HM 79-6/4 (Koc¸ Karpuz and Jansen, 1992) in the southeasorwegian Sea. (See Fig. 1 for location). Note that thed18O values onM.duliswere made on aragonite that is enriched ind18O relative to calcite. Tompare thed18O values from Kulturmyra with thed18O values from theoraminifera, thed18O curve from Kulturmyra should be shifted 0.5–0.9‰he left indicating a difference of about 3‰. Considering that the salcified under higher temperatures than the foraminifera in the open Nian Sea, the difference ind18O corresponds to a salinity difference of ab–3‰ between the two water masses. This difference is only 1–2‰ large
oday and shows that the shells calcified in marine water with very littlef fresh water. Radiocarbon ages are not corrected for a reservoir age
t
r
iae
tehe
-ningtoid
inter months (Seed, 1976). If we assume a 2°C higherionation temperature for the shells in Kulturmyra thanorams in the Norwegian Sea, the salinity difference woulbout 2–3‰, only 1–2‰ larger than the difference is tohis shows that the shells calcified in salty, marine waterery little input of any fresh water, indicating that the obtaieservoir age is representative of the open ocean.
The d18O andd13C values of the dated shells are shownelation to the reservoir ages (Fig. 6). There is no trend insotope ratios versus the reservoir ages. The variations itable isotopes of the shells, probably reflecting variationemperature and salinity of the waters, do not seem tonfluenced the reservoir ages.
ariation in Reservoir Age Present and 10,000–12,30014CYears B.P.
The most appropriate definition of a reservoir age isifference between14C ages of samples grown contemporausly in the atmosphere and in the actual reservoir (Stuivetl., 1986). The marine reservoir age is time and space deent due to reservoir parameters (size, carbon exchangeestricting its response to variations in the14C/12C ratio of thetmosphere. An estimate of reservoir ages for the North
antic was made by Mangerud and Gulliksen (1975) by dauseum samples of molluscs collected about AD 1eighted mean reservoir ages of 4306 20 and 4706 30 yr for
outhern and northern Norway, respectively, indicated thanfluence of fresh water (from the Baltic Sea and run-off frouthern Norway) on the Atlantic water decreased inoastal current northward along the Norwegian coast gigher reservoir ages. Reliable atmospheric14C data were novailable; thus, the estimate was based on the differenc
ween the radiocarbon age of the samples and their knistoric age before AD 1950, that is about 50 yr. Howeendrochronological calibration data have revealed thaorresponding terrestrial radiocarbon ages are in the r0–100 yr B.P. (Stuiver and Becker, 1993), and we hecalculated the present (i.e., ca. AD 1900) reservoir agouthern Norway as 3796 20 yr, close to Olsson’s (198stimate of 3706 25 yr. More important, it is also almo
dentical to the reservoir age found for the Kulturmyrauence, calculated as the weighted mean radiocarbon ag
erence between marine shells and contemporary terreamples. It shows that the reservoir ages along the Norwoast during Bølling–Allerød were quite similar to thoseoday (Fig. 7).
The last deglaciation was marked by rapid changes innflow of Atlantic surface waters into the Norwegian Sigh-resolution marine records show three periods with
inct increases of polar foraminifera (Lehman and Keigw992), diatoms (Koc¸ Karpuz and Jansen, 1992), and ice-raetritus (Haflidasonet al.,1995) between 12,500 and 10,000.P. The most prominent of these is the Younger D
11,000–10,000 yr B.P.) return to almost glacial conditio
,n
se-
ant

14C ages within or close to the Vedde Ash in marine succes-so lan(1V ab hil ashsc t thm tingt olda thii facw a-icc dW
altn urit hei senC an7 urvm Lak
S rvoira nce( heoJ1 f theo se oft
s thef ringt astw ore,t gianc al top wardI xten-s Koce inm in-c onal
ledc m1 evet ellm trenb onst r as
sen,1 ntic(o ervoirav hem
BONDEVIK ET AL.112
ions in the North Atlantic are 700–800 yr older than14C agesf terrestrial plant material within the same ash bed onKoc Karpuz and Jansen, 1992; Bardet al.,1994; Austinet al.,995; Haflidasonet al., 1995). Determining a14C age for theedde Ash on the shelf and in the deep Sea is problemecause of bioturbation, depth of the dated molluscs w
iving, reworking of older sediments, and ice rafting ofhards (Austinet al., 1995). Nevertheless, Bardet al. (1994)oncluded, after a correction of the bioturbation bias, thaean14C age is still about 11,000–11,100 yr B.P., indica
hat the reservoir age of the sea surface was 300–400 yrt that time in the North Atlantic than today. The reason for
s probably a combination of reduced convection of suraters to the North Atlantic and an increase of the seover during the Younger Dryas (Bardet al.,1994; Stocker anright, 1996).Even though the southeastern Norwegian Sea was
ately influenced by cold and temperate water masses dhe period 12,000–11,000 yr B.P., the data presentedmply that the reservoir age was not higher than at preonsidering the limited number of paired samples (Figs. 3, Table 1), there is no obvious trend in the data set. By catching the dated sequence to the chronology from
FIG. 6. d13C and d18O values in relation to their reservoir ages. Filircles show values obtained from measurements on pure aragonite fro4C-dated shells. The open circles are from shells located at the same lhe core as the dated shells and are measurements from the entire shight thus be a mixture of both calcite and aragonite. There is noetween reservoir ages and isotope ratios indicating that small variati
emperature and salinity do not seem to have influenced the reservoiignificantly.
d
ticle
e
ersee
er-ngret.dee
uigetsu we obtained a good fit using the weighted resege of 380 yr for the marine14C ages throughout the sequeFig. 4). The rapid climate shifts of the OD I and OD II in tcean did not last more than 200–400 yr (Koc¸ Karpuz andansen, 1992; Lehman and Keigwin, 1992; Haflidasonet al.,995), possibly not long enough to alter the reservoir age ocean significantly, or these variations were missed becau
he sample density.Numerical models suggest that the cover of sea ice i
actor that contributes most to higher reservoir ages duhe YD (Stocker and Wright, 1996). The Norwegian coas partly ice-free during the late-glacial period. Theref
he reservoir ages we have found for the western Norweoast between 11,000 and 12,300 yr B.P., almost identicresent values, may not apply to areas farther west to
celand and Greenland where the ocean was more eively covered by sea ice during Late Weichselian time (t al., 1993). New radiocarbon ages of the Vedde Asharine cores outside Iceland confirm this picture, with
reasing ages toward the west (H. Haflidason, pers
thel inanddin
ges
FIG. 7. Reservoir ages from historic shells (Mangerud and Gullik975) from southern Norway, from the Vedde Ash Bed in the North AtlaBardet al.,1994, Austinet al.,1995, Haflidasonet al.,1995) and Kulturmyran the outermost coast of western Norway (this study). The weighted resge from Kulturmyra between 14,500 and 12,800 cal yr B.P. is 3806 32 yr,ery close to the present value of 3796 20 yrs. The reservoir age for tid-Younger Dryas Ash zone, the Vedde Ash Bed, is 700–800 yr.

communication, 1998). This might suggest that the mariner giac thN
99w Unv t thQ rinF onst s.E s aJ reaa scb rsitP HigR Rs thN
A 90.
A va
A 95ryas
B t, JNoras
B I.pp
B cia
B rns
B rt, Garv
B ctioea
B ngrwy,
G amo-
H 99ropSe
H andedu
from eastern Svalbard and the Holocene marine climatic optimum.Polar
H n, J.,st
I rd ofrnia.
I n and
K ology
K f therapid
K nstruc-egianws
L lantic
L lantic-
M er asiralis
M tions
M shells,k
M ecent
M 984).tionsntic.
M arbonl-
O rn
P ation
P entalwest
P eriod
R318.
R itical
LATE WEICHSELIAN MARINE 14C RESERVOIR AGES 113
eservoir ages for the Younger Dryas along the Norweoast are slightly lower than the 700 – 800 yr found fororwegian Sea–North Sea.
ACKNOWLEDGMENTS
Most of this paper was written during the spring and summer of 1hile the first author visited the Quaternary Research Center at theersity of Washington. He is most grateful to both staff and faculty auaternary Research Center. Dagfinn Bondevik helped with the corank Hague did the magnetic susceptibility measurements, Stig M
he TOC analysis, and Rune Søraas the13C/12C and18O/16O measurementystein Jansen helped with the interpretation of the oxygen isotopeohn Raven with information about seaweed photosynthesis. S.B. gppreciates financial support toward a PC from Hans and Helga Reuequest of 1997. The work is a contribution to the Strategic Univerogram at the University of Bergen, “Rapid Sedimentation andesolution Stratigraphy: Processes and Application,” funded by theearch Council of Norway. H.H.B. was supported during this work byorwegian Research Council.
REFERENCES
une, B. (1993). “Air Temperature Normals, Normal Period 1961–19Report 02/93 Klima. Den norske meteorologiske institutt. Oslo.
ure, J., and Østensen, Ø. (1993). Hydrographic normals og long-termations in Norwegian coastal waters.Fisken og Havet6, 1–75.
ustin, W. E. N., Bard, E., Hunt, J. B., Kroon, D., and Peacock, J. D. (19The 14C age of the Icelandic Vedde Ash: Implications for Younger Dmarine reservoir age corrections.Radiocarbon37, 53–62.
ard, E., Arnold, M., Mangerud, J., Paterne, M., Labeyrie, L., DupraMelieres, M.-A., Sønstegaard, E., and Duplessy, J.-C. (1994). TheAtlantic atmosphere-sea surface14C gradient during the Younger Dryclimatic event.Earth and Planetary Science Letters126,275–287.
ayne, B. L., Widdows, J., and Thompson, R. J. (1976). Physiology:In“Marine Mussels: Their Ecology and Physiology” (B. L. Bayne, Ed.),411. Cambridge Univ. Press. Cambridge, UK.
irks, H. H. (1993). The importance of plant macrofossils in late-glaclimatic reconstructions: An example from western Norway.QuaternaryScience Reviews12, 719–726.
irks, H. H. (1994). Late-glacial vegetational ecotones and climatic pattewestern Norway.Vegetational History and Archaeobotany3, 107–119.
irks, H. H., Gulliksen, S., Haflidason H., Mangerud, J., and Possne(1996). New radiocarbon dates from the Vedde Ash and the SaksunAsh from western Norway.Quaternary Research45, 119–127.
irks, H. H., Battarbee, R. W., and Birks, H. J. B. (in press). A reconstruof the aquatic ecosystem of Kråkenes Lake during the late glacial andHolocene.Journal of Paleolimnology.
likra, L. H., and Longva, O. (1995). Frost-shattered debris facies of YouDryas age in the coastal sedimentary successions in western NoPalaeoenvironmental implications.Palaeogeography, PalaeoclimatologPalaeoecology118,89–110.
rossman, E. L. (1984). Stable isotope fractionation in live benthic fornifera from the southern California borderland.Palaeogeography, Palaeclimatology, Palaeoecology47, 301–327.
aflidason, H., Sejrup, H. P., Klitgaard Kristensen, D., and Johnsen, S. (1Coupled response of the late glacial climatic shifts of northwest Eureflected in Greenland ice cores: Evidence from the northern NorthGeology23, 1059–1062.
jort, C., Mangerud, J., Adrielsson, L., Bondevik, S., Landvik, J. Y.,Salvigsen, O. (1995). Radiocarbon dated common mussels Mytilus
ne
8i-
eg,en
ndtlyh’syhe-e
”
ri-
).
.,th
.
l
in
.atn
nrly
eray:
i-
5).ea.
lis
Research14, 239–243.
ughen, K. A., Overpeck, J. T., Lehman, S. J., Kashgarian, M., Southoand Peterson, L. C. (1998). A new14C calibration data set for the ladeglaciation based on marine varves.Radiocarbon40, 483–494.
ngram, B. L., Ingle, J. C., and Conrad, M. E. (1996). Stable isotope recolate Holocene salinity and river discharge in San Francisco Bay, CalifoEarth and Planetary Science Letters141,237–247.
sraelson, C., and Buchardt, B. (1991). The isotope composition of oxygecarbon in some fossil and recent bivalve shells from east Greenland.LUN-DQUA Report33, 117–123.
itagawa, H., and van der Plicht, J. (1998). A 40,000-year varve chronfrom Lake Suigetsu, Japan: Extension of the14C calibration curve.Radio-carbon40, 505–515.
oc Karpuz, N., and Jansen, E. (1992). A high-resolution diatom record olast deglaciation from the SE Norwegian Sea: Documentation ofclimatic changes.Paleoceanography7, 499–520.
oc, N., Jansen, E., and Haflidason, H. (1993). Paleoceanographic recotions of surface ocean conditions in the Greenland, Iceland and NorwSeas through the last 14 ka based on diatoms.Quaternary Science Revie12, 115–140.
ehman, S. J., and Keigwin, L. D. (1992). Sudden changes in North Atcirculation during the last deglaciation.Nature356,757–762.
owe, J. J., and NASP Members (1995). Palaeoclimate of the North AtSeaboards during the last glacial/interglacial transition.Quaternary International 28, 51–61.
adsen, T., and Maberly, S. C. (1990). A comparison of air and watenvironments for photosynthesis by the intertidal algea Fucus Sp(Phaeophyta).Journal of Phycology26, 24 –30.
angerud, J. (1970). Late Weichselian Vegetation and Ice-Front Oscillain the Bergen District, western Norway.Norsk Geografisk Tidsskrift24,121–148.
angerud, J. (1977). Late Weichselian marine sediments containingforaminifera, and pollen, at Ågotnes, western Norway.Norsk GeologisTidsskrift57, 23–54.
angerud, J., and Gulliksen, S. (1975). Apparent radiocarbon ages of Rmarine shells from Norway, Spitsbergen, and Arctic Canada.QuaternaryResearch5, 263–273.
angerud, J., Lie, S. E., Furnes, H., Kristiansen, I. L., and Lømo, L. (1A Younger Dryas ash bed in Western Norway, and its possible correlawith tephra in cores from the Norwegian Sea and the North AtlaQuaternary Research21, 85–104.
ook, W. G. (1971). Paleotemperatures and chlorinities from stable cand oxygen isotopes in shell carbonate.Palaeogeography, Palaeoclimatoogy, Palaeoecology9, 245–263.
lsson, I. U. (1980). Content of14C in marine mammals from NortheEurope.Radiocarbon22, 662–675.
aus, A. (1988). Late Weichselian vegetation, climate, and floral migrat Sandvikvatn, North Rogaland, south-western Norway.Boreas 17,133–139.
eacock, J. D. (1989). Marine molluscs and Late Quaternary environmstudies with particular reference to the Late-Glacial period in northEurope: A review.Quaternary Science Reviews8, 179–192.
earson, G. W. (1986). Precise calendrical dating of known growth-psamples using a ‘curve fitting’ technique.Radiocarbon28, 292–299.
aven, J. A. (1994). Photosynthesis in Aquatic Plants.In “Ecophysiology ofPhotosynthesis” (E. D. Schulze and M. M. Caldwell, Eds.), pp. 299–Springer-Verlag, Berlin.
ohling, E. J., and Bigg, G. R. (1998). Paleosalinity and delta 18O: A crassessment.Journal of Geophysical Research103,1307–1318.

Seed, R. (1976).In “Marine Mussels: Their Ecology and Physiology” (B. L.
S latio
S f th
S ibra
S
S n-1
S n s
tigraphy on the outer coast of Sunnmøre, western Norway.Norsk Geologisk
T oly-ationta
V andacialo-
Y rniv.
BONDEVIK ET AL.114
Bayne, Ed.), pp. 411. Cambridge Univ. Press. Cambridge, UK.
tocker, T. F., and Wright, D. G. (1996). Rapid changes in ocean circuand atmospheric radiocarbon.Paleoceanography11, 773–795.
tuiver, M., and Becker, B. (1993). High-precision decadal calibration oradiocarbon time scale, AD 1950–6000 BC.Radiocarbon35, 35–65.
tuiver, M., Pearson, G. W., and Braziunas, T. F. (1986). Radiocarbon age calof marine samples back to 9000 cal yr BP.Radiocarbon28, 980–1021.
tuiver, M., and Polach, H. A. (1977). Reporting of14C data.Radiocarbon19,355–363.
tuiver, M., Quay, P. D., and Ostlund, H. G. (1983). Abyssal water carbodistribution and the age of the world oceans.Science219,849–851.
vendsen, J. I., and Mangerud, J. (1990). Sea-level changes and polle
n
e
tion
4
tra-
Tidsskrift70, 111–134.
arutani, T., Clayton, R. N., and Mayeda, T. K. (1969). The effect of pmorphism and magnesium substitution on oxygen isotope fractionbetween calcium carbonate and water.Geochimica et Cosmochimica Ac33, 987–996.
an Dinter, M., and Birks, H. H. (1996). Distinguishing fossil Betula nanaB. pubescens using their wingless fruits: Implications for the late-glvegetational history of western Norway.Vegetational History and Archaebotany5, 229–240.
onge, C. M. (1976). The ‘mussel’ form and habit.In “Marine Mussels: TheiEcology and Physiology” (B. L. Bayne, Ed.), pp. 1–12. Cambridge UPress. Cambridge, UK.




















