
Multipotent mesenchymal stem cells from adult human eye conjunctiva stromal cells
Upload
independentCategory
view
0download
0
Late-Outgrowth Endothelial Cells Attenuate IntimalHyperplasia Contributed by Mesenchymal Stem Cells After
Vascular InjuryChao-Hung Wang, Wen-Jin Cherng, Ning-I Yang, Li-Tang Kuo, Chia-Ming Hsu, Hung-I Yeh,
Yii-Jenq Lan, Chi-Hsiao Yeh, William L. Stanford
Objectives—Mesenchymal stem cells (MSCs) are one of a number of cell types undergoing extensive investigation forcardiac regeneration therapy. It has not yet been determined whether this cell therapy also substantially contributes tovascular remodeling of diseased vessels.
Methods and Results—Human MSCs and a variety of progenitor and vascular cells were used for in vitro and in vivoexperiments. Wire-induced vascular injury mobilized MSCs into the circulation. Compared with human aortic smoothmuscle cells, MSCs exhibited a 2.8-fold increase in the adhesion capacity in vitro (P�0.001) and a 6.3-fold increase invivo (P�0.001). In all animal models, a significant amount of MSCs contributed to intimal hyperplasia after vascularinjury. MSCs were able to differentiate into cells of endothelial or smooth muscle lineage. Coculture experimentsdemonstrated that late-outgrowth endothelial cells (OECs) guided MSCs to differentiate toward an endothelial lineagethrough a paracrine effects. In vivo, cell therapy with OECs significantly attenuated the thickness of the neointimacontributed by MSCs (intima/media ratio, from 3.2�0.4 to 0.4�0.1, P�0.001).
Conclusions—Tissue regeneration therapy with MSCs or cell populations containing MSCs requires a strategy to attenuatethe high potential of MSCs to develop intimal hyperplasia on diseased vessels. (Arterioscler Thromb Vasc Biol.2008;28:54-60.)
Key Words: mesenchymal stem cells � intimal hyperplasia � cell therapy � vascular remodeling
Stem cell therapy has a promising future for the treatmentof a variety of end-stage cardiovascular diseases such
as heart failure, ischemic heart,1–3 and peripheral arterydiseases.4 To date, bone marrow (BM) cells or mobilizedperipheral mononuclear cells are among the most often usedcell populations in clinical settings because of their conve-nience and autologous properties.4,5 Recently, the MAGICcell randomized clinical trial, however, showed that intra-coronary infusion of peripheral blood stem cells mobilizedwith granulocyte colony stimulating factor (G-CSF) aggra-vated restenosis after coronary stenting in myocardial infarc-tion.6 Based on the heterogeneous populations of these cellsand the differentiation capacity of stem cells, uncontrolleddifferentiation is a critical concern when applying these cellsto humans.
Mesenchymal stem cells (MSCs) reside in the BM and canalso be mobilized in response to G-CSF stimulation.7 MSCsfrom the BM are multipotent and have the capacity todifferentiate into cardiomyocytes,8 endothelial cells (ECs),9,10
and smooth muscle cells (SMCs).11 Currently, MSCs are one
of the cell types being used in clinical trials for postmyocar-dial infarction cardiac regeneration therapy.3,12 It has beenshown that an intramyocardial injection of autologous MSCsor intravenous administration of MSCs can increase vasculo-genesis and improve cardiac function after myocardial infarc-tion in animal experiments and clinical trials.3,12 However,abundant evidence suggests that BM-derived circulating pre-cursors can give rise to ECs and SMCs that contribute tovascular repair, remodeling, and lesion formation underphysiological and pathological conditions.13,14 There is a veryhigh possibility that MSCs residing in therapeutic cell popu-lations can adhere to diseased, angioplastic, or stented vesselswith cell administration, causing intimal hyperplasia andeventual restenosis. Herein, we sought to determine thecontribution of MSCs to neointimal formation after vascularinjury, and to investigate possible therapeutic strategies tomodulate this contribution.
MethodsAn expanded Materials and Methods section is available in theonline data supplement #1 at http://atvb.ahajournals.org.
Original received May 3, 2007; final version accepted October 17, 2007.From the Division of Cardiology, Department of Internal Medicine (C.-H.W., W.-J.C., N.-I.Y., L.-T.K., C.-M.H., Y.-J.L., C.-H.Y.), Chang Gung
Memorial Hospital, Chang Gung University College of Medicine, Keelung, Taiwan; the Mackay Memorial Hospital (H.-I.Y.), Mackay Medicine, Nursingand Management College, Taipei Medical University, Taiwan; and the Institute of Biomaterials and Biomedical Engineering (W.L.S.), University ofToronto, Samuel Lunenfeld Research Institute, Mount Sinai Hospital, Toronto, Canada.
Correspondence to Chao-Hung Wang, MD, Division of Cardiology, Department of Internal Medicine, Chang Gung Memorial Hospital, 222, Mai-ChinRoad, Keelung, Taiwan. E-mail [email protected]
© 2007 American Heart Association, Inc.
Arterioscler Thromb Vasc Biol. is available at http://atvb.ahajournals.org DOI: 10.1161/ATVBAHA.107.147256
54 by guest on March 4, 2016http://atvb.ahajournals.org/Downloaded from by guest on March 4, 2016http://atvb.ahajournals.org/Downloaded from by guest on March 4, 2016http://atvb.ahajournals.org/Downloaded from by guest on March 4, 2016http://atvb.ahajournals.org/Downloaded from by guest on March 4, 2016http://atvb.ahajournals.org/Downloaded from by guest on March 4, 2016http://atvb.ahajournals.org/Downloaded from by guest on March 4, 2016http://atvb.ahajournals.org/Downloaded from by guest on March 4, 2016http://atvb.ahajournals.org/Downloaded from by guest on March 4, 2016http://atvb.ahajournals.org/Downloaded from by guest on March 4, 2016http://atvb.ahajournals.org/Downloaded from by guest on March 4, 2016http://atvb.ahajournals.org/Downloaded from by guest on March 4, 2016http://atvb.ahajournals.org/Downloaded from by guest on March 4, 2016http://atvb.ahajournals.org/Downloaded from by guest on March 4, 2016http://atvb.ahajournals.org/Downloaded from

AnimalsMale FVB wild-type, athymic nude (Jackson Laboratory, Bar Har-bor, Me) and eGFP transgenic mice (FVB background) that ubiqui-tously express enhanced green fluorescent (GFP) (Level Biotechnol-ogy)15 were bred and maintained in the Laboratory of AnimalExperiments at Chang Gung Memorial Hospital. The study protocolwas reviewed and approved by the Institutional Animal Care and UseCommittee, Chang Gung Memorial Hospital Faculty of Medicine,and the experiments were conducted according to the Guidelines ofthe American Physiological Society.
Mouse Femoral Wire-Injury ModelFemoral artery injury was induced by inserting a straight spring wire(0.38 mm in diameter, No. C-SF-15-15, Cook) for more than 5 mmtoward the iliac artery.14
Mesenchymal Stem Cells Cultured FromPeripheral BloodTo culture mouse MSCs in peripheral blood, pooled whole bloodwas harvested by cardiac puncture from 20 mice for each experi-ment. MSCs were cultured in 2 groups of mice, including the studygroup (48 hours after the femoral artery wire injury) and a control(sham-operated) group. After Ficoll density centrifugation(Histopaque-1083, density 1.083 g/mL), mononuclear cells (3�106)were plated on 6-cm dishes to culture the MSCs as described above.
Dye-Transfer Assay for Gap JunctionIntercellular CommunicationLucifer yellow (LY; Mr: 457 Da; 0.2%) and rhodamine dextran (RD;Mr:10 kDa) were microinjected into cells. LY can penetrate throughgap junction channels; however, RD is too large to pass through gapjunction channels and therefore served as a tracer dye for cellsoriginally receiving the dye. Transfer of the dye was visualized usinga phase-contrast fluorescence microscope (Axioscope; Carl Zeiss,Jena, Germany), in which LY and RD were respectively detected asgreen and red. DAPI was used to label nuclei (blue). Five minutesafter the donor cell was injected, the presence of dye transfer to othercells was estimated. On average, 25 microinjections were tried foreach experiment.
Results
MSCs Have a High Capacity for Adhering to theMatrix or the Remodeled Vessel Wall After InjuryThe capacity for adhesion was investigated in a variety ofhuman progenitor and vascular cells both in vitro and in vivo.In vitro, compared with other progenitor and vascular cells,MSCs had the strongest adhesion capacity to both fibronectinand collagen (Figure 1A, left and middle panels). Further-more, mixing OECs with MSCs significantly increased theadhesion of OECs to the coated matrix (Figure 1A, right
Figure 1. Adhesive capacity of mesenchymal stem cells (MSCs). A and B, Human MSCs had the strongest adhesive capacity in vitro(A, left and middle panels) and in vivo (n�4 to 5 in each group) (B). *P�0.05, **P�0.01 (fibrinogen), †P�0.05, ‡P�0.01 (collagen), com-pared with smooth muscle cells (SMCs). Mixing MSCs with outgrowth endothelial cells (OEC#s) increased the adherence of OEC#s (A,right panel). EPCs and SPCs, endothelial and smooth muscle progenitor cells, respectively; HAEC, human aortic endothelial cell; MNC,mononuclear cell; OEC#, labeled OEC.
Wang et al MSCs and Intimal Hyperplasia 55
by guest on March 4, 2016http://atvb.ahajournals.org/Downloaded from

panel). In vivo, progenitor or vascular cells were directlyinjected via the abdominal aorta of mice 7 days after the rightfemoral artery wire injury. MSCs had the best adherence tothe injured vessel wall compared with all other types of cells(Figure 1B). No adherence of injected cells was found ineither small arterioles or the uninjured left femoral artery,suggesting that the injected cells did not cause cell embolismand were unable to adhere to intact and healthy vascularsurfaces. The injected MSCs were also unable to adhere to thevessels 1 day after wire injury (data not shown), suggestingthat MSC adhesion required matrix formation on the remod-eled injured vessels, or the adherent platelets covered on theinjured vascular surface interfered with MSC attachment tothe underlying matrix.
Evidence of MSC Mobilization Intothe CirculationBlood G-CSF, VEGF, SDF-1�, and SCF concentrations weremeasured at serial time points in mice after femoral arterywire injury (supplemental Figure IA). The levels of theseblood cytokines, especially G-CSF and VEGF, significantlyincreased within 24 hours. Blood G-CSF concentrationsincreased up to 8-fold 12 hours after vascular injury. Then,standard MSC cultivation was performed on mononuclearcells purified from pooled peripheral blood of mice 48 hoursafter wire injury. From the same amount of peripheral bloodmononuclear cells, colonies of MSCs in the wire-injurygroup could be subcultured and maintained for long-termculture (at least 10 passages), whereas colonies from thecontrols could not be maintained for longer than 2 passages(supplemental Figure IB). MSCs cultured from the wire-injury group had homogeneous morphologies, wereLin�CD45�Sca-1�CD31� (supplemental Figure II), and
were multipotent, as they were capable of differentiatinginto osteocytes (positive for Von Kossa staining, alkalinephosphatase, and bone morphogenetic protein-2) or adipo-cytes (positive for Oil Red O) in response to differentdifferentiation media (supplemental Figure IC), and intoECs and SMCs (described as follows).
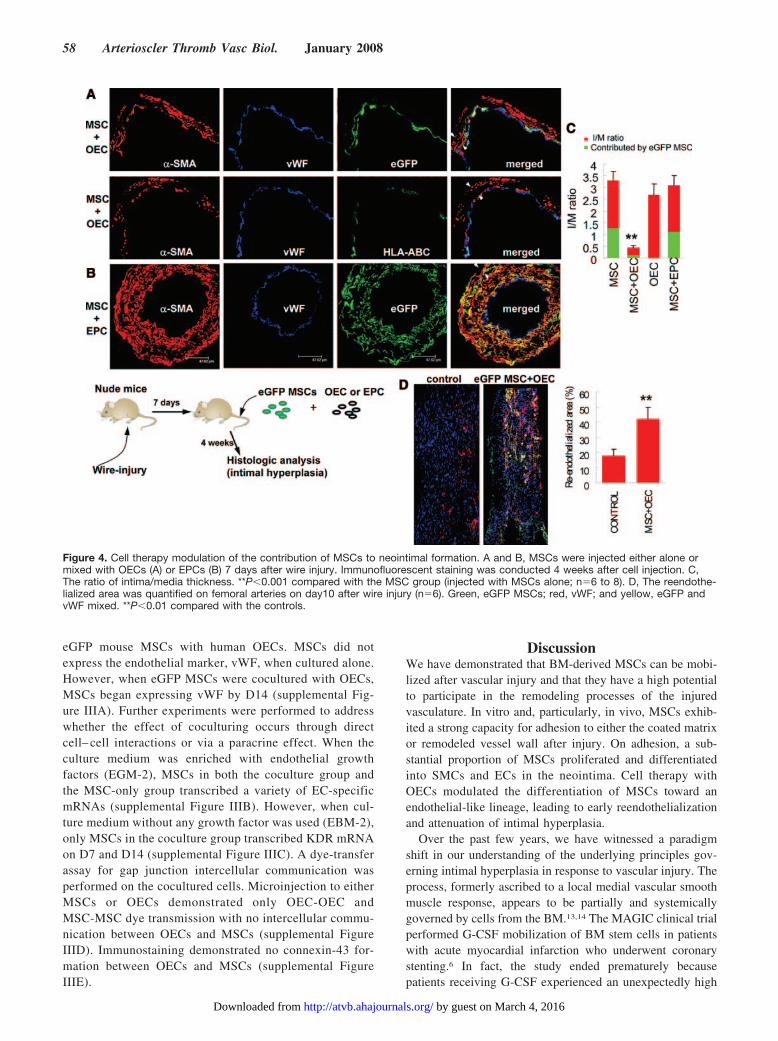
Contribution of MSCs to Intimal HyperplasiaTo investigate whether MSCs contribute to neointimal for-mation in vivo, studies were performed using 2 models, “celltherapy” and “physiological” models. In the cell therapymodel, BM-derived MSCs from eGFP mice were injected viathe tail vein into wild-type littermates 7 days after femoralartery wire injury (Figure 2). One day after the injection,scattered eGFP MSCs were found on the surface of injuredvessels (Figure 2A). Four weeks after cell injection, clustersof eGFP MSCs were identified in the neointima (Figure 2B).Most of the eGFP MSCs were also �-SMA–positive. Onaverage, 31%�15% of cells in the neointima were from theinjected eGFP MSCs.
In the physiological model, eGFP MSCs and radioprotec-tive whole eGFP-negative BM cells were transplanted intothe tibia and the via tail vein, respectively, of wild-typelittermates after 1000 cGy of whole-body irradiation (Figure3A). Two months after MSC BM transplantation, femoralartery wire injury was performed. Four weeks later, theinjured vessels had developed remarkable intimal hyperpla-sia, which contained a significant amount of eGFP-positivecells indicating the contribution of BM MSCs (39%�17%)(Figure 3B, lower panel, D28). In addition, flow cytometryand fluorescent microscopy proved successful engraftment ofeGFP MSCs in the BM (Figure 3A).
Figure 2. Contribution of MSCs to inti-mal hyperplasia. MSCs from eGFPmice were injected via the tail vein 7days after femoral artery injury. A andB, Confocal microscopy demonstratedeGFP MSC adherence to the injuredvascular surface 24 hour after theinjection (A) and a clustered or nodularpattern of eGFP MSC growth as acomponent in neointima 28 days afterthe cell injection (B).
56 Arterioscler Thromb Vasc Biol. January 2008
by guest on March 4, 2016http://atvb.ahajournals.org/Downloaded from

Differentiation of MSCs Into Neointimal CellsIn mice after intra-BM eGFP MSC transplantation, theeGFP� MSCs in neointima differentiated into a few differentcell types. Some of the eGFP� cells were �-SMA�, somewere von Willebrand factor (vWF)� or triple positive, and theothers were pure eGFP� on day 21 after the wire injury(Figure 3B, upper panel, D21). However, when using CD31as a marker of highly differentiated ECs, only cells over thesurface of the neointima were eGFP�CD31� (Figure 3B,middle panel, D21). These findings suggested that mobilizedBM MSCs underwent a differentiation process into eitherSMC- or EC-lineage cells. When injured vessels were inves-tigated at late time points such as 4 weeks after wire injury,immunostaining of vWF and �-SMA was located only on thesurface and the body of the neointima, respectively, suggest-ing that the differentiation processes had ceased (Figure 3B,lower panel, D28).
Modulation of the Contribution of MSCs to theNeointima by Cell TherapyThe findings described above raise the possibility of manip-ulating the differentiation of MSCs, which had adhered to theinjured vascular surface, into the endothelial lineage toachieve early reendothelialization. Thus, we attempted to usecell therapy to modulate MSC differentiation by mixing
eGFP mouse MSCs with human EPCs or OECs. Using thetail vein injection model, cell therapy with OECs significantlyattenuated the contribution of eGFP MSCs and non MSC-derived SMCs to intimal hyperplasia (intima/media ratio,from 3.2�0.4 to 0.4�0.1, P�0.001) (Figure 4A, upper paneland Figure 4C). eGFP cells were either purely �SMA-positive or purely vWF-positive in the neointima 4 weeksafter femoral artery wire injury. A subset of ECs was alsopositive for HLA-ABC, a marker specific for humans (Figure4A, lower panel). These findings suggested that OEC therapycauses early re-endothelialization either by OECs themselvesor by guiding the injected eGFP MSCs to differentiate intoECs. However, EPC therapy or OEC alone was unable tosignificantly attenuate the thickness of intimal hyperplasia(Figure 4B and 4C). To investigate the speed of reendothe-lialization, en face immunostaining was performed on theentire femoral artery 10 days after wire injury. There wasmore extensive endothelial covering of the injured femoralarteries in the “MSC�OEC” group, compared with thecontrols (Figure 4D).
Coculture Experiment: Cell Therapy Guides theFate of MSCsTo clarify the effect of cell therapy on MSC differentia-tion, coculture experiments were performed by coculturing
Figure 3. Differentiation of MSCs into neointimal cells. A, An intra–bone marrow transplantation model was used. Flow cytometry andfluorescent microscopy demonstrated the successful engraftment of the transplanted eGFP MSCs. B, Immunofluorescent staining with�-SMA, vWF, and CD31 on vessels 21 (D21) and 28 days (D28) after wire injury.
Wang et al MSCs and Intimal Hyperplasia 57
by guest on March 4, 2016http://atvb.ahajournals.org/Downloaded from
eGFP mouse MSCs with human OECs. MSCs did notexpress the endothelial marker, vWF, when cultured alone.However, when eGFP MSCs were cocultured with OECs,MSCs began expressing vWF by D14 (supplemental Fig-ure IIIA). Further experiments were performed to addresswhether the effect of coculturing occurs through directcell– cell interactions or via a paracrine effect. When theculture medium was enriched with endothelial growthfactors (EGM-2), MSCs in both the coculture group andthe MSC-only group transcribed a variety of EC-specificmRNAs (supplemental Figure IIIB). However, when cul-ture medium without any growth factor was used (EBM-2),only MSCs in the coculture group transcribed KDR mRNAon D7 and D14 (supplemental Figure IIIC). A dye-transferassay for gap junction intercellular communication wasperformed on the cocultured cells. Microinjection to eitherMSCs or OECs demonstrated only OEC-OEC andMSC-MSC dye transmission with no intercellular commu-nication between OECs and MSCs (supplemental FigureIIID). Immunostaining demonstrated no connexin-43 for-mation between OECs and MSCs (supplemental FigureIIIE).
DiscussionWe have demonstrated that BM-derived MSCs can be mobi-lized after vascular injury and that they have a high potentialto participate in the remodeling processes of the injuredvasculature. In vitro and, particularly, in vivo, MSCs exhib-ited a strong capacity for adhesion to either the coated matrixor remodeled vessel wall after injury. On adhesion, a sub-stantial proportion of MSCs proliferated and differentiatedinto SMCs and ECs in the neointima. Cell therapy withOECs modulated the differentiation of MSCs toward anendothelial-like lineage, leading to early reendothelializationand attenuation of intimal hyperplasia.
Over the past few years, we have witnessed a paradigmshift in our understanding of the underlying principles gov-erning intimal hyperplasia in response to vascular injury. Theprocess, formerly ascribed to a local medial vascular smoothmuscle response, appears to be partially and systemicallygoverned by cells from the BM.13,14 The MAGIC clinical trialperformed G-CSF mobilization of BM stem cells in patientswith acute myocardial infarction who underwent coronarystenting.6 In fact, the study ended prematurely becausepatients receiving G-CSF experienced an unexpectedly high
Figure 4. Cell therapy modulation of the contribution of MSCs to neointimal formation. A and B, MSCs were injected either alone ormixed with OECs (A) or EPCs (B) 7 days after wire injury. Immunofluorescent staining was conducted 4 weeks after cell injection. C,The ratio of intima/media thickness. **P�0.001 compared with the MSC group (injected with MSCs alone; n�6 to 8). D, The reendothe-lialized area was quantified on femoral arteries on day10 after wire injury (n�6). Green, eGFP MSCs; red, vWF; and yellow, eGFP andvWF mixed. **P�0.01 compared with the controls.
58 Arterioscler Thromb Vasc Biol. January 2008
by guest on March 4, 2016http://atvb.ahajournals.org/Downloaded from

rate of restenosis at the stent site. A close correlation betweenthe gain in neointimal volume and improvements in systolicfunction was noted in the cell infusion group, suggesting thatstem-cell therapy accelerated neointimal growth in proportionto the efficacy of cardiac regeneration. Our study highlightsthe possible role of MSCs in this adverse effect.
To date, a variety of cell populations have been applied tostem cell therapeutic trials, including BM-derived mononu-clear cells,4 G-CSF-mobilized peripheral blood stem cells,6
skeletal myoblasts,16 and MSCs.3 We speculate that most ofthese cell populations recruit MSCs as well. In addition, todelete MSCs from the applied cell populations is technicallydifficult and may also influence the potentially beneficialcontribution of MSCs to cardiac regeneration or angiogene-sis. MSCs are highly proliferative and can differentiate intocardiomyocytes,8 ECs,9,10 or SMCs.11 In contrast to theirhematopoietic counterparts, our data demonstrated that MSCshave a strikingly enhanced ability to adhere to matrix-coatedtissue-culture surfaces and to remodeled vessel wall afterinjury, as compared with a variety of other progenitor/vascular cells. This phenomenon is supported by data show-ing that MSCs express several receptors associated withmatrix- and cell-to-cell adhesive interactions.11 Furthermore,our study, for the first time, linked MSCs to the excessiverepair processes of vessels in response to injury.
Human and a few mammalian MSCs were reported to bemobilized into the systemic circulation in response to G-CSFtreatment,7 leading to engraftment at peripheral organ sitesand differentiation according to the niche and repair ofinjured tissues.7,17–19 Evidence showed that stem cells withthe capacity to differentiate into cardiomyocytes are actuallymesenchymal rather than hematopoietic stem cells,8 suggest-ing that MSCs have a higher potential to differentiate intocells with a muscular phenotype, compared with HSCs. Asshown in our study, vascular injury caused an approximately8-fold increase in blood G-CSF concentrations along withmodest elevations in other cytokines such as SCF, SDF-1�,and VEGF, providing an environment potentially optimal forMSC mobilization. Being able to culture MSCs from theperipheral blood of the wire-injured mice lends furthersupport to the evidence for MSC mobilization.
The origins of cells contributing to intimal hyperplasia arediverse including local SMCs, BM-derived vascular progen-itors, stem cells in the adventitia, as well as additional celltypes.20 Because a tremendous amount of medial vascularSMCs underwent apoptosis in the animal model adopted inthis study,21 a significant portion of vascular repair dependson systemically mobilized cells.13 Using the entire BMtransplantation model with an irradiation dose of 1000 cGy,most of the reconstituted BM cells were hematopoietic cells.However, the estimated D0 (the radiation dose that reducessurvival to 37%) of MSCs is 1.3 to 1.4 Gy, and 1 Gy alreadyinduces death in a significant portion of MSCs.22 Previously,Fukuda et al showed that reconstituted BM cells containednon-hematopoietic cells when an irradiation dose of 1050cGy was used.8 Using an intra-BM MSC transplantationmodel, they also demonstrated that MSCs, but not HSCs, inthe BM contributed to the regeneration of myocardial tissueafter myocardial infarction. All of these findings support the
hypothesis that MSCs can participate in the process of postwire-injury vascular remodeling. In line with this, our datashowed that MSCs seeded onto the surface of injured vesselwall 7 days after wire injury and proliferated in a nodular orclustered pattern. These adherent MSCs proliferated anddifferentiated into both SMC- and EC-like cells. Becauseonly a small portion of MSCs were reconstituted in ourintra-BM transplantation model, the proportion of intimalhyperplasia contributed to by MSCs was probably more thanthat estimated by our study.
In an undifferentiated state, MSCs do not express ECsurface markers such as CD31 or CD34.9 However, recentwork has shown that altering culture conditions can renderMSCs capable of differentiating into ECs.9,10 All theseattributes make MSCs an interesting cell phenotype forinvestigation in light of their potential to differentiate intomesoderm-derived ECs23 and their ability to differentiate invivo into ECs.24,25 As shown in our study, although coculturewith OECs may help MSCs express EC phenotypes, VEGF-enriched medium alone can achieve this effect as well, in linewith findings by Oswald et al.26 Because both our dye-transfer assay and connexin-43 immunostaining suggested nodirect communication between OECs and MSCs, the influ-ence of OECs on MSC differentiation is suggested to occurthrough a paracrine effect.
Regarding approaches to attenuate neointimal formation,numerous medical therapeutic strategies, including cell ther-apy, have been investigated. Current concepts support thehypothesis that the earlier that reendothelialization isachieved, the less neointima that forms. Although OECsalone have an inadequate capacity for adhesion, mixing OECswith MSCs is herein suggested to increase the adhesion ofOECs to injured vessel walls via the mediation of MSCsbetween OECs and matrix formed on the injured vessels. Ithas been reported that early EPCs secrete more VEGF than doOECs.27 However, the influence of EPCs on MSC differen-tiation appears to be much less than that of OECs. One of thepossible reasons is that the life-span of early EPCs is muchshorter than that of the late OECs (A brief introduction toearly EPCs and OECs is given in the online supplement#2.).27 Our data suggest that a coinfusion of OECs and MSCsmay help achieve early reendothelialization. These findingscan also explain the discrepancy among reports regarding theadverse effects of stem cell therapy on atherosclerosis,6,28
because there is a wide variation in the amount of endothelialprogenitors in harvested BM cells and in mobilized cells afterG-CSF treatment in different individuals.
In summary, our data clearly demonstrated the potential ofstem cell therapy to contribute to atherosclerosis or poststent-ing restenosis. MSCs, either spontaneously mobilized fromBM or delivered directly by cell therapy, are 2 possiblesources of cells in intimal hyperplasia. The amount of MSCmobilization depends on the effect of cytokines released inresponse to the severity of the vascular injury. However, celltherapy containing an MSC population has a direct impact onneointimal formation. Although the contribution of MSCsand the repair processes carried out by local SMCs aftervascular injury are individualized in different subjects, inter-actions between OECs and MSCs were demonstrated herein
Wang et al MSCs and Intimal Hyperplasia 59
by guest on March 4, 2016http://atvb.ahajournals.org/Downloaded from

to accelerate reendothelialization with beneficial effects onregulating local SMCs and MSCs, leading to remarkableattenuation in intimal hyperplasia. In a clinical setting, it issuggested that before cells are applied to the target organs,cell manipulation should be attempted either by combined-cell therapy or by medical intervention to raise the content ofendothelial progenitors.
AcknowledgmentsWe thank Christine Holmes and Yi-Chun Lin for technical assistancewith the cell culture and microinjection.
Sources of FundingThis work was supported in part by National Science Council ofTaiwan (NSC 95-2314-B-182A-046 and NSC 94-2314-B-182A-192).
DisclosuresNone
References1. Britten MB, Abolmaali ND, Assmus B, Lehmann R, Honold J, Schmitt J,
Vogl TJ, Martin H, Schachinger V, Dimmeler S, Zeiher AM. InfarctRemodeling After Intracoronary Progenitor Cell Treatment in PatientsWith Acute Myocardial Infarction (TOPCARE-AMI): mechanisticinsights from serial contrast-enhanced magnetic resonance imaging.Circulation. 2003;108:2212–2218.
2. Zhang S, Wang D, Estrov Z, Raj S, Willerson JT, Yeh ETH. Both cellfusion and transdifferentiation account for the transformation ofhuman peripheral blood CD34-positive cells into cardiomyocytes invivo. Circulation. 2004;110:3803–3807.
3. Chen Sl, Fang Ww, Ye F, Liu YH, Qian J, Shan Sj, Zhang Jj, ChunhuaRZ, Liao Lm, Lin S, Sun Jp. Effect on left ventricular function ofintracoronary transplantation of autologous bone marrow mesenchymalstem cell in patients with acute myocardial infarction. Am J Cardiol.2004;94:92–95.
4. Tateishi-Yuyama E, Matsubara H, Murohara T, Ikeda U, Shintani S,Masaki H, Amano K, Kishimoto Y, Yoshimoto K, Akashi H, Shimada K,Iwasaka T, Imaizumi T. Therapeutic angiogenesis for patients with limbischaemia by autologous transplantation of bone-marrow cells: a pilotstudy and a randomised controlled trial. Lancet. 2002;360:427–435.
5. Tateno K, Minamino T, Toko H, Akazawa H, Shimizu N, Takeda S,Kunieda T, Miyauchi H, Oyama T, Matsuura K, Nishi Ji, Kobayashi Y,Nagai T, Kuwabara Y, Iwakura Y, Nomura F, Saito Y, Komuro I. Criticalroles of muscle-secreted angiogenic factors in therapeutic neovascular-ization. Circ Res. 2006;98:1194–1202.
6. Kang HJ, Kim HS, Zhang SY, Park KW, Cho HJ, Koo BK, Kim YJ, LeeDS, Sohn DW, Han KS. Effects of intracoronary infusion of peripheralblood stem-cells mobilised with granulocyte-colony stimulating factor onleft ventricular systolic function and restenosis after coronary stenting inmyocardial infarction: the MAGIC cell randomised clinical trial. TheLancet. 2004;363:751–756.
7. Kocher AA, Schuster MD, Szabolcs MJ, Takuma S, Burkhoff D, WangJ, Homma S, Edwards NM, Itescu S. Neovascularization of ischemicmyocardium by human bone-marrow-derived angioblasts prevents car-diomyocyte apoptosis, reduces remodeling and improves cardiacfunction. Nat Med. 2001;7:430–436.
8. Kawada H, Fujita J, Kinjo K, Matsuzaki Y, Tsuma M, Miyatake H,Muguruma Y, Tsuboi K, Itabashi Y, Ikeda Y, Ogawa S, Okano H, HottaT, Ando K, Fukuda K. Nonhematopoietic mesenchymal stem cells can bemobilized and differentiate into cardiomyocytes after myocardialinfarction. Blood. 2004;104:3581–3587.
9. Huss R, Lange C, Weissinger EM, Kolb HJ, Thalmeier K. Evidence ofperipheral blood-derived, plastic-adherent CD34-/low hematopoieticstem cell clones with mesenchymal stem cell characteristics. Stem Cells.2000;18:252–260.
10. Tomanek RJ, Schatteman GC. Angiogenesis: new insights and thera-peutic potential. Anat Rec. 2000;261:126–135.
11. Minguell JJ, Erices A, Conget P. Mesenchymal Stem Cells. Expe BiolMed. 2001;226:507–520.
12. Gnecchi M, He H, Liang OD, Melo LG, Morello F, Mu H, Noiseux N,Zhang L, Pratt RE, Ingwall JS, Dzau VJ. Paracrine action accounts formarked protection of ischemic heart by Akt-modified mesenchymal stemcells. Nat Med. 2005;11:367–368.
13. Sata M, Saiura A, Kunisato A, Tojo A, Okada S, Tokuhisa T, Hirai H,Makuuchi M, Hirata Y, Nagai R. Hematopoietic stem cells differentiateinto vascular cells that participate in the pathogenesis of atherosclerosis.Nat Med. 2002;8:403–409.
14. Wang CH, Ciliberti N, Li SH, Szmitko PE, Weisel RD, Fedak PW, AlOmran M, Cherng WJ, Li RK, Stanford WL, Verma S. Rosiglitazonefacilitates angiogenic progenitor cell differentiation toward endotheliallineage: a new paradigm in glitazone pleiotropy. Circulation. 2004;109:1392–1400.
15. Hsiao YC, Chang HH, Tsai CY, Jong YJ, Horng LS, Lin SF, Tsai TF.Coat color-tagged green mouse with EGFP expressed from the RNApolymerase II promoter. Genesis. 2004;39:122–129.
16. Herreros J, Prosper F, Perez A, Gavira JJ, Garcia-Velloso MJ, Barba J,Sanchez PL, Canizo C, Rabago G, Marti-Climent JM, Hernandez M,Lopez-Holgado N, Gonzalez-Santos JM, Martin-Luengo C, Alegria E.Autologous intramyocardial injection of cultured skeletal muscle-derivedstem cells in patients with non-acute myocardial infarction. Eur Heart J.2003;24:2012–2020.
17. Barbash IM, Chouraqui P, Baron J, Feinberg MS, Etzion S, Tessone A,Miller L, Guetta E, Zipori D, Kedes LH, Kloner RA, Leor J. Systemicdelivery of bone marrow-derived mesenchymal stem cells to theinfarcted myocardium: feasibility, cell migration, and body distri-bution. Circulation. 2003;108:863– 868.
18. Prockop DJ. Marrow stromal cells as stem cells for nonhematopoietictissues. Science. 1997;276:71–74.
19. Bittira B, Shum-Tim D, Al-Khaldi A, Chiu RCJ. Mobilization andhoming of bone marrow stromal cells in myocardial infarction. EurJ Cardiothorac Surg. 2003;24:393–398.
20. Gulati R, Jevremovic D, Peterson TE, Chatterjee S, Shah V, Vile RG,Simari RD. Diverse origin and function of cells with endothelial pheno-type obtained from adult human blood. Circ Res. 2003;93:1023–1025.
21. Sata M, Maejima Y, Adachi F, Fukino K, Saiura A, Sugiura S, Aoyagi T,Imai Y, Kurihara H, Kimura K, Omata M, Makuuchi M, Hirata Y, NagaiR. A mouse model of vascular injury that induces rapid onset of medialcell apoptosis followed by reproducible neointimal hyperplasia. J MolCell Cardiol. 2000;32:2097–2104.
22. Rombouts WJ, Ploemacher RE. Primary murine MSC show highlyefficient homing to the bone marrow but lose homing ability followingculture. Leukemia. 2003;17:160–170.
23. Reyes M, Lund T, Lenvik T, Aguiar D, Koodie L, Verfaillie CM.Purification and ex vivo expansion of postnatal human marrow meso-dermal progenitor cells. Blood. 2001;98:2615–2625.
24. Al-Khaldi A, Eliopoulos N, Martineau D, Lejeune L, Lachapelle K, GalipeauJ. Postnatal bone marrow stromal cells elicit a potent VEGF-dependentneoangiogenic response in vivo. Gene Ther. 2003;10:621–629.
25. Eliopoulos N, Al-Khaldi A, Beausejour CM, Momparler RL, MomparlerLF, Galipeau J. Human cytidine deaminase as an ex vivo drug selectablemarker in gene-modified primary bone marrow stromal cells. Gene Ther.2002;9:452–462.
26. Oswald J, Boxberger S, Jorgensen B, Feldmann S, Ehninger G, BornhauserM, Werner C. Mesenchymal stem cells can be differentiated into endothelialcells in vitro. Stem Cells. 2004;22:377–384.
27. Yoon CH, Hur J, Park KW, Kim JH, Lee CS, Oh IY, Kim TY, Cho HJ, KangHJ, Chae IH, Yang HK, Oh BH, Park YB, Kim HS. Synergistic neovascu-larization by mixed transplantation of early endothelial progenitor cells andlate outgrowth endothelial cells: the role of angiogenic cytokines and matrixmetalloproteinases. Circulation. 2005;112:1618–1627.
28. Ripa RS, Jorgensen E, Wang Y, Thune JJ, Nilsson JC, Sondergaard L,Johnsen HE, Kober L, Grande P, Kastrup J. Stem cell mobilizationinduced by subcutaneous granulocyte-colony stimulating factor toimprove cardiac regeneration after acute ST-elevation myocardialinfarction: result of the double-blind, randomized, placebo-controlledStem Cells in Myocardial Infarction (STEMMI) Trial. Circulation. 2006;113:1983–1992.
60 Arterioscler Thromb Vasc Biol. January 2008
by guest on March 4, 2016http://atvb.ahajournals.org/Downloaded from

Yii-Jenq Lan, Chi-Hsiao Yeh and William L. StanfordChao-Hung Wang, Wen-Jin Cherng, Ning-I Yang, Li-Tang Kuo, Chia-Ming Hsu, Hung-I Yeh,
Mesenchymal Stem Cells After Vascular InjuryLate-Outgrowth Endothelial Cells Attenuate Intimal Hyperplasia Contributed by
Print ISSN: 1079-5642. Online ISSN: 1524-4636 Copyright © 2007 American Heart Association, Inc. All rights reserved.
Greenville Avenue, Dallas, TX 75231is published by the American Heart Association, 7272Arteriosclerosis, Thrombosis, and Vascular Biology
doi: 10.1161/ATVBAHA.107.1472562008;28:54-60; originally published online November 8, 2007;Arterioscler Thromb Vasc Biol.
http://atvb.ahajournals.org/content/28/1/54World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://atvb.ahajournals.org/content/suppl/2007/12/19/ATVBAHA.107.147256.DC1.htmlData Supplement (unedited) at:
http://atvb.ahajournals.org//subscriptions/
at: is onlineArteriosclerosis, Thrombosis, and Vascular Biology Information about subscribing to Subscriptions:
http://www.lww.com/reprints
Information about reprints can be found online at: Reprints:
document. Question and AnswerPermissions and Rightspage under Services. Further information about this process is available in the
which permission is being requested is located, click Request Permissions in the middle column of the WebCopyright Clearance Center, not the Editorial Office. Once the online version of the published article for
can be obtained via RightsLink, a service of theArteriosclerosis, Thrombosis, and Vascular Biologyin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on March 4, 2016http://atvb.ahajournals.org/Downloaded from

Supplements
Supplement #1:
Methods
Animals
Male FVB wild-type, athymic nude (Jackson Laboratory, Bar Harbor, ME) and eGFP
transgenic mice (FVB background) that ubiquitously express enhanced green
fluorescent (GFP) (Level Biotechnology, Taipei, Taiwan)1 were bred and maintained
in the Laboratory of Animal Experiments at Chang Gung Memorial Hospital. The
study protocol was reviewed and approved by the Institutional Animal Care and Use
Committee, Chang Gung Memorial Hospital Faculty of Medicine, and the
experiments were conducted according to the Guidelines of the American
Physiological Society.
Mouse femoral wire-injury model
To test the contribution of MSCs to neointimal formation, a mouse femoral
artery wire-injury model was adopted since this model is best suited for stem cell
studies of post-injury vascular remodeling. Transluminal mechanical injury of the
femoral artery was carried out under a dissecting microscope. Briefly, either the left or
right femoral artery was exposed by blunt dissection. The accompanying femoral
nerve was carefully separated, but the femoral vein was isolated from the artery.2 The
femoral artery and vein were looped together proximally and distally with 6-0 silk
sutures for temporary vascular control during the procedure. A small branch between
the rectus femoris and vastus medialis muscles was isolated, and looped proximally
and ligated distally with 6-0 silk sutures. The vein and connective tissues around the
artery were carefully removed with microsurgical forceps. The exposed muscular
branch artery was dilated by topically applying one drop of 1% lidocaine
hydrochloride. A transverse arteriotomy was performed on this branch. A straight
spring wire (0.38 mm in diameter, no. C-SF-15-15, Cook, Bloomington, IN) was

carefully inserted into the femoral artery toward the iliac artery. The wire was left in
place for 1 min to denude and dilate the artery. Then, the wire was removed, and a silk
suture looped at the proximal portion of the muscular branch artery was secured.
Blood flow to the femoral artery was restored by releasing the sutures placed in the
proximal and distal femoral portions. The skin incision was closed with 5-0 silk
sutures. At different time points, the femoral arteries were excised, fixed in OCT
compound (TissueTek, Tokyo, Japan), and used for the immunofluorescence studies.
Cell culture
Early endothelial progenitor cells (EPCs),2 late-outgrowth endothelial cells (OECs),2
smooth muscle progenitor cells (SPCs),3 and MSCs (mouse)4 were cultured as
previously described. Human aortic endothelial cells (HAECs) and aortic smooth
muscle cells (HASMCs) (Cascade Biologics, U.S.A.) were respectively cultured in
media 200 and 231 with cell growth supplements. Mouse ECs (SVEC4-10) were
purchased from ATCC (CRL-2181) (Rockville, MD) and cultured with DMEM.
MSCs were expanded within MesenCult MSC culture medium (StemCell
Technologies, Vancouver, Canada) and used at passage 4 to 5. For the co-culture
experiments, numbers of MSCs and OECs or EPCs were plated at a 1: 9 ratio.
Immunostaining and RT-PCR were performed at different time points after
co-culturing.
Adherence capacity
In vitro, collagen type I (Rockland) (50 µg/mL) or fibronectin (50 µg/mL) was coated
onto 24-well plates for 2 h at 37 °C. Wells were blocked with 1% BSA in PBS for 15
min, and cells (5 × 105) were added to each well and allowed to attach for 1 h as
previously described.5 In an additional adhesion assay, OECs (5 × 105, labeled with
5-chloromethylflourescein diacetate (CMFDA), green dye, Molecular Probes) were
mixed with MSCs (5 × 105) to test whether the MSCs enhanced the adherence of

OECs to the matrix. Attached cell numbers were randomly counted in four areas of
each well.
In vivo, 1 x 106 cells (either from eGFP mice or from human cells) were injected
via the abdominal aorta into mice (either FVB wild-type or nude mice) 7 days after
femoral artery wire injury. One day after the cell injection, the femoral artery was
harvested to estimate the amount of cell adhesion by immunofluorescence staining.
For human cells, CellTracker Green CMFDA was used for cell tracking.
Mouse bone marrow harvesting
To harvest BM, animals were sacrificed by cervical dislocation, the femurs were
removed, and the BM was flushed into a sterile 15-ml tube using a syringe and a 23-G
needle. Single cell suspensions were prepared by passing the material through a
23-G needle in a small amount of growth medium. Cells were washed, and after the
process of RBC lysis, were counted, and then were ready for use.
Intra-BM transplantation of MSCs
Wild-type FVB mice at 8 weeks of age were lethally irradiated with a total dose of
1000 cGy. After irradiation, the recipient mice received unfractionated BM cells (1 ×
106) (which served as radioprotective cells) from wild-type mice through a tail vein
injection and MSCs (1 × 105) from eGFP mice through a tibial BM injection in each
leg.6 Two months after transplantation, the mice were ready for examination. When
the mice were sacrificed, engraftment of the eGFP MSCs was confirmed by flow
cytometry and culture of BM cells.
Cell therapy model
To investigate the contribution of MSCs to neointimal formation, 1 x 106 eGFP MSCs
were injected into the tail vein of wild-type FVB mice (or nude mice) 7 days after the
femoral artery wire injury. To investigate whether the cell therapy attenuated
MSC-mediated intimal hyperplasia, MSCs (1 x 106) alone or mixed with EPCs or

OECs (1 x 106 for each cell type) were injected via the tail vein of nude mice (1 h
after cells were mixed). Vessels were harvested 28 days after the cell injection.
Confocal immunofluorescence staining
The femoral artery was harvested for immunostaining at different time points after the
cell injection. Frozen sections were stained with the following primary antibodies:
anti-GFP (Chemicon); Cy3-conjugated anti-α-SMA (Sigma, St. Louis, MO);
anti-vWF (DAKO, produced in rabbits); anti-CD31 (BD Pharmingen);
anti-connexin-43 (Chemicon); and anti-HLA-ABC (Biolegend) followed by
incubation with FITC-, PE-, or Alexa Fluor 647-conjugated secondary antibodies.
Slides were mounted using a Prolong Antifade kit (Molecular Probes, Eugene, OR)
and observed under a confocal microscope (Leica TCS SP2 AOBS). Nuclei were
stained with DAPI or Hoechst 33258 (Sigma). For en face denuded arteries,
3-dimensional (3D) confocal images were used to visualize the area of
re-endothelialization. The femoral arteries were harvested 10 days after wire injury (5
days after cell therapy through the abdominal aorta). To measure the
re-endothelialized area, arteries were fixed with 4% paraformaldehyde (PFA) and 10%
sucrose for 2 h. The artery was opened longitudinally, fixed on a slide and then
subjected to immunostaining for eGFP and vWF and 3D confocal microscopic
imaging. After the 3-D imaging was compacted into two dimensions, the areas stained
positive for vWF and the total femoral artery area were measured, and the percentage
areas were calculated.
Immunoassay for chemokines and cytokines
Plasma levels of granulocyte colony-stimulating factor (G-CSF), stroma-derived
factor 1α (SDF-1α), SCF (stem cell factor), and vascular endothelial growth factor
(VEGF) were measured using an enzyme-linked immunosorbent assay (ELISA; R&D
Systems, Minneapolis, MN) at different time points after the surgery.

Mesenchymal stem cells cultured from peripheral blood
To culture mouse MSCs in peripheral blood, pooled whole blood was harvested by
cardiac puncture from 20 mice for each experiment. MSCs were cultured in two
groups of mice, including the study group (48 h after the femoral artery wire injury)
and a control (sham-operated) group. After Ficoll density centrifugation
(Histopaque-1083, density 1.083 g/ml), mononuclear cells (3 × 106) were plated on
6-cm dishes to culture the MSCs as described above. The multipotent capacity for
differentiation of the cultured mobilized MSCs was also tested.
Mesenchymal stem cell differentiation assay
For osteoblast differentiation, cells were stimulated with dexamethasone (10-8 M),
L-ascorbic acid (50 μg/ml), and β-glycerophate (3.5 mM) in 15% alpha-MEM.
Selected culture specimens after 6, 12, and 18 days of growth in the subculture were
stained for mineral deposition by the Von Kossa method. The stained cultures were
examined using phase-contrast optics. For adipocyte differentiation, adipogenesis was
stimulated with dexamethasone (10-6 M), insulin (5 μg/ml), IBMX (50 μM),
indomethacin (60 μM), and rosiglitazone (10 μM) in 15% alpha-MEM. Cells were
fixed on day 14, and adipocytes were visualized by staining with Oil Red O.
Reverse-transcriptase polymerase chain reaction (RT-PCR)
Co-cultured cells were harvested at the indicated time points, and total cellular RNA
was extracted using the RNeasy Mini Kit (Qiagen). RT-PCR was performed with a
OneStep RT-PCR kit (Promega). For each reaction, 1 µg of total RNA served as a
template. For amplification, primer pairs specific for mouse vWF (sense primer
5'-CCG GAA GCG ACC CTC AGA-3' and antisense primer 5'-CGG TCA ATT TTG
CCA AAG ATC T-3'),7 mouse Tie-2 (sense primer 5'-TTG AAG TGA CGA ATG
AGA T-3' and antisense primer 5'-ATT TAG AGC TGT CTG GCT T-3'), mouse
Flk-1 (KDR) (sense primer 5'-AGC TTG GCT CAC AGG CAA CAT CGG-3' and
antisense primer 5'- TGG CCC GCT TAA CGG TCC GTA GG-3'),8 and GFP (sense

primer 5'-AAG TTC ATC TGC ACC ACC G-3' and antisense primer 5'-TGC TCA
GGT AGT GGT TGT-3')1 were used. The bands of the RT-PCR appeared only in
mouse ECs not in HAECs, indicating that the primers used were specific for mice. In
the RT-PCR for Tie-2, 35 cycles of 94 °C for 1 min, 52 °C for 1 min, and 72 °C for 1
min were used. For other genes, 35 cycles of 94 °C for 1 min, 55 °C for 1 min, and 72
°C for 1 min, were used with a final extension at 72 °C for 10 min. The RT-PCR
product of mouse vWF was 130 bp, of Tie-2 was 197 bp, of Flk-1 was 624 bp, and of
GPF was 500 bp.
Dye-transfer assay for gap junction intercellular communication
A cell microinjection assay was performed based on a published procedure with
modifications.9 Briefly, Lucifer yellow (LY; Mr: 457 Da; 0.2%) and rhodamine
dextran (RD; Mr:10 kDa) were microinjected into cells. LY can penetrate through gap
junction channels; however, RD is too large to pass through gap junction channels and
therefore served as a tracer dye for cells originally receiving the dye. Transfer of the
dye was visualized using a phase-contrast fluorescence microscope (Axioscope; Carl
Zeiss, Jena, Germany), in which LY and RD were respectively detected as green and
red. DAPI was used to label nuclei (blue). Five minutes after the donor cell was
injected, the presence of dye transfer to other cells was estimated. On average, 25
microinjections were tried for each experiment.
Statistical analysis
Data were analyzed by one-way analysis of variance. When statistically significant
effects were found, Tukey's test was performed to isolate differences between groups.
Student's t-test was used when appropriate. A p value of < 0.05 was considered
significant. All data are presented as the mean ± SEM in the text and figures.
Supplement #2: Two different types of EPCs can be cultured from adult peripheral

blood, namely early EPCs and OECs (also called late EPCs) according to their
time-dependent appearance. Early EPCs with a spindle shape show peak growth at 2
to 3 weeks and die at 4 weeks, whereas OECs with a cobblestone shape appear late at
2 to 3 weeks, show exponential growth at 4 to 8 weeks, and live up to 12 weeks.10
Early EPCs are a heterogeneous group of cells and secrete high levels of cytokines;
however, OECs are quite homogeneous. They share some endothelial phenotypes and
similarly contribute to neovasculogenesis in vivo. Compared to early EPCs, OECs are
closer to mature ECs in phenotype but show surprising migration, and tube-forming
capabilities.
Supplementary figure legends:
Supplemental Figure I. Evidence of mesenchymal stem cell (MSC) mobilization. A,
Stem cell-mobilizing cytokines significantly increased after wire injury. * p < 0.05, **
p < 0.01 compared to the baseline (0 h) (n = 8-10 in each group). B, Peripheral blood
MSCs were cultured and subcultured to confluence. C, Adipogenic (red) and
osteogenic (brown) potentials of peripheral blood MSCs. Immunostaining was
positive for alkaline phosphatase (ALP, blue) and bone morphogenetic protein-2
(BMP-2, red).
Supplemental Figure II. Phenotypes of mesenchymal stem cells cultured from
peripheral blood. Flow cytometry and confocal immunofluorescent analysis show that
they are Lin-CD45-Sca-1+CD31-.
Supplemental Figure III. Manipulation of mesenchymal stem cell (MSC)
differentiation by late-outgrowth endothelial cells (OECs). A, MSCs from eGFP mice
were co-cultured with human OECs, and immunofluorescent staining was conducted

at serial time points after co-culturing. eGFP, green; α-SMA, blue; vWF, red;
eGFP+α-SMA+, cyan; eGFP +vWF +, yellow. Arrows indicate smooth muscle-like
cells; arrowheads indicate endothelial-like cells. B, RT-PCR for a variety of
endothelial phenotype mRNA was also performed at different time points after
co-culturing in either EGM-2 (enriched with EC growth factors) (B) or EBM-2
(containing no growth factors) (C) (n = 4). mEC, mouse EC. D, dye-transfer assay:
(left panel) OECs, injected dye (*) was transferred to other OECs (green); (middle
and right panels) co-culture of OECs (not labeled) and MSCs (labeled with DAPI,
blue) for 48h; (middle) dye was injected into OECs (*) and transferred to OECs only,
not to MSCs; (right) dye was injected into MSCs (*) and transferred to MSCs only,
not to OECs (n = 25 for each experiment). E, After the co-culturing of human MSCs
(green) and OECs (white) for 14 days, immunostaining revealed that connexin-43 had
only formed between OECs.
References 1. Kawada H, Fujita J, Kinjo K, Matsuzaki Y, Tsuma M, Miyatake H, Muguruma
Y, Tsuboi K, Itabashi Y, Ikeda Y, Ogawa S, Okano H, Hotta T, Ando K, Fukuda K. Nonhematopoietic mesenchymal stem cells can be mobilized and differentiate into cardiomyocytes after myocardial infarction. Blood. 2004;104:3581-3587.
2. Wang CH, Ciliberti N, Li SH, Szmitko PE, Weisel RD, Fedak PW, Al Omran M, Cherng WJ, Li RK, Stanford WL, Verma S. Rosiglitazone facilitates angiogenic progenitor cell differentiation toward endothelial lineage: a new paradigm in glitazone pleiotropy. Circulation. 2004;109:1392-1400.
3. Hsiao YC, Chang HH, Tsai CY, Jong YJ, Horng LS, Lin SF, Tsai TF. Coat color-tagged green mouse with EGFP expressed from the RNA polymerase II promoter. Genesis. 2004;39:122-129.
4. Fukuda D, Sata M, Tanaka K, Nagai R. Potent Inhibitory Effect of Sirolimus on Circulating Vascular Progenitor Cells. Circulation. 2005;111:926-931.

5. Pittenger MF, Mackay AM, Beck SC, Jaiswal RK, Douglas R, Mosca JD, Moorman MA, Simonetti DW, Craig S, Marshak DR. Multilineage Potential of Adult Human Mesenchymal Stem Cells. Science. 1999;284:143-147.
6. Tepper OM, Galiano RD, Capla JM, Kalka C, Gagne PJ, Jacobowitz GR, Levine JP, Gurtner GC. Human endothelial progenitor cells from type II diabetics exhibit impaired proliferation, adhesion, and incorporation into vascular structures. Circulation. 2002;106:2781-2786.
7. Olsen CL, Hsu PP, Glienke J, Rubanyi GM, Brooks AR. Hedgehog-interacting protein is highly expressed in endothelial cells but down-regulated during angiogenesis and in several human tumors. BMC Cancer. 2004;4:43.
8. Tsao PN, Li H, Wei SC, Ko ML, Chou HC, Hsieh WS, Hsieh FJ. Expression of angiogenic factors and their receptors in postnatal mouse developing lung. J Formos Med Assoc. 2004;103:137-143.
9. Blackman CF, Andrews PW, Ubeda A, Wang X, House DE, Trillo MA, Pimentel ME. Physiological levels of melatonin enhance gap junction communication in primary cultures of mouse hepatocytes. Cell Biol Toxicol. 2001;17:1-9.
10. Hur J, Yoon CH, Kim HS, Choi JH, Kang HJ, Hwang KK, Oh BH, Lee MM, Park YB. Characterization of two types of endothelial progenitor cells and their different contributions to neovasculogenesis. Arterioscler Thromb Vasc Biol. 2004;24:288-293.

A
B C
adipogenesis
0
100
200
300
400
500
600
0 6 12 24 48
Time points after wire-injury
pg/m
l
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
0.4
0.45
0.5
0 6 12 24 48
Time points after wire-injury
ng/m
l
0
0.5
1
1.5
2
2.5
3
0 6 12 24 48
Time points after wire-injury
pg/m
l
0
20
40
60
80
100
120
140
160
180
200
0 6 12 24 48
Time points after wire-injury
pg/m
l
G-CSF SDF-1α VEGF SCF
**
** **** **
****
*
****
*
ALP
BMP-2
osteogenesis
supplementary Fig. I
(h) (h) (h) (h)
Peripheral blood MSCs

CD317%
CD455%
Lineage1.5%
Sca-1 C-kit CD45
Sca-156%
Supplement Fig. II

supplementary Fig. III
A Co-culture D7 D14
CEmMSC
mMSC+
OEC mMSC
D0 D7 D14 D7 D14KDR
GFP
α-SMA: MSC vWF: OECConnexin-43 DAPI: nucleus
D Dye-transfer assay
**
OEC alone Co-culture Co-cultureinject OEC inject
MSCLucifer/RhoDAPI(MSC)
*
Co-culture D14
Copyright © 2022 FDOKUMEN




















