
Intraduodenal infusion of α-ketoglutarate decreases whole body energy expenditure in growing pigs
8
Clinical Nutrition (2006) 25, 489–496 ORIGINAL ARTICLE Intraduodenal infusion of a-ketoglutarate decreases whole body energy expenditure in growing pigs Peter Junghans a , Michael Derno a, , Stefan Pierzynowski b,c , Ulf Hennig a , Paul Eberhard Rudolph d , Wolfgang B. Souffrant a a Research Unit Nutritional Physiology ‘‘Oskar Kellner’’, Research Institute for the Biology of Farm Animals (FBN), Wilhelm-Stahl-Allee 2, D-18196 Dummerstorf, Germany b Department Cell and Organism Biology, Lund University, Helgonava¨gen 3B, 22362 Lund, Sweden c Sea Fisheries Institute, Kollotaja 1, 81-332 Gdynia, Poland d Research Unit Genetics & Biometry, Wilhelm-Stahl-Allee 2, D-18196 Dummerstorf, Germany Received 18 April 2005; accepted 7 November 2005 KEYWORDS a-Ketoglutarate; Administration route; Energy expenditure; Pig model Summary Background & aims: a-Ketoglutarate (AKG) has been suggested to play a particular role as an oxidative fuel for the gut, and thus may have a sparing function for fuels such as glutamate and aspartate. Using the pig model we aimed to quantify how the route of administration (intravenous, i.v.; intragastric, i.g.; intraduodenal, i.d.) affects AKG utilization, whole body energy expenditure (EE) and nutrient oxidation. Methods: Pigs (15 kg) were supplied with a complete nutrient solution (NS) via catheters. To explore the metabolic effects of AKG, 1.0g AKG kgBW 1 d 1 was infused simultaneously with the NS using either the i.d., i.v. or i.g. route. [1- 13 C]AKG (15 mg kgBW 1 ) was infused i.d., i.v. or i.g., respectively, for 3h. AKG utilization (AKG UTIL) was estimated as AKG UTIL ¼ 100– 13 C recovery (% of 13 C dose). 13 C recovery was calculated from the 13 C enrichment in breath CO 2 and the whole-body CO 2 production. Results: AKG infusion and NS via the i.d. route resulted in a reduced AKG UTIL (40.176.7) as compared to the i.v. route (62.972.4, Po0.001) and i.g. route (62.371.6, Po0.001). The total EE was lower with the i.d. route of AKG and NS (745768 kJ kgBW 0.62 d 1 ) as compared to the i.v. route (965754 kJ kgBW 0.62 d 1 , Po0.005) and i.g. route (918743 kJ kgBW 0.62 d 1 , Po0.005). Carbohydrate oxidation was increased with the i.d. route (38.2 g73.4 kgBW 0.62 d 1 ) as compared to the i.v. route (27.872.9 g kgBW 0.62 d 1 , Po0.08) and i.g. route (23.978.5 g kgBW 0.62 d 1 , Po0.05). Fat oxidation was decreased (2.171.9 g kgBW 0.62 d 1 ; Po0.001) with the ARTICLE IN PRESS http://intl.elsevierhealth.com/journals/clnu 0261-5614/$ - see front matter & 2005 Elsevier Ltd and European Society for Clinical Nutrition and Metabolism. All rights reserved. doi:10.1016/j.clnu.2005.11.003 Corresponding author. Tel.: +49 38208 68 684; fax: +49 38208 68 652. E-mail address: [email protected] (M. Derno).
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Intraduodenal infusion of α-ketoglutarate decreases whole body energy expenditure in growing pigs

ARTICLE IN PRESS
Clinical Nutrition (2006) 25, 489–496
0261-5614/$ - sdoi:10.1016/j.c
�CorrespondiE-mail addr
http://intl.elsevierhealth.com/journals/clnu
ORIGINAL ARTICLE
Intraduodenal infusion of a-ketoglutaratedecreases whole body energy expenditurein growing pigs
Peter Junghansa, Michael Dernoa,�, Stefan Pierzynowskib,c, Ulf Henniga,Paul Eberhard Rudolphd, Wolfgang B. Souffranta
aResearch Unit Nutritional Physiology ‘‘Oskar Kellner’’, Research Institute for the Biology of Farm Animals(FBN), Wilhelm-Stahl-Allee 2, D-18196 Dummerstorf, GermanybDepartment Cell and Organism Biology, Lund University, Helgonavagen 3B, 22362 Lund, SwedencSea Fisheries Institute, Kollotaja 1, 81-332 Gdynia, PolanddResearch Unit Genetics & Biometry, Wilhelm-Stahl-Allee 2, D-18196 Dummerstorf, Germany
Received 18 April 2005; accepted 7 November 2005
KEYWORDSa-Ketoglutarate;Administrationroute;Energy expenditure;Pig model
ee front matter & 2005lnu.2005.11.003
ng author. Tel.: +49 382ess: derno@fbn-dumme
SummaryBackground & aims: a-Ketoglutarate (AKG) has been suggested to play a particularrole as an oxidative fuel for the gut, and thus may have a sparing function for fuelssuch as glutamate and aspartate. Using the pig model we aimed to quantify how theroute of administration (intravenous, i.v.; intragastric, i.g.; intraduodenal, i.d.)affects AKG utilization, whole body energy expenditure (EE) and nutrient oxidation.Methods: Pigs (15 kg) were supplied with a complete nutrient solution (NS) viacatheters. To explore the metabolic effects of AKG, 1.0 g AKG kgBW�1 d�1 wasinfused simultaneously with the NS using either the i.d., i.v. or i.g. route. [1-13C]AKG(15mg kgBW�1) was infused i.d., i.v. or i.g., respectively, for 3 h. AKG utilization(AKG UTIL) was estimated as AKG UTIL ¼ 100–13C recovery (% of 13C dose). 13Crecovery was calculated from the 13C enrichment in breath CO2 and the whole-bodyCO2 production.Results: AKG infusion and NS via the i.d. route resulted in a reduced AKG UTIL(40.176.7) as compared to the i.v. route (62.972.4, Po0.001) and i.g. route(62.371.6, Po0.001). The total EE was lower with the i.d. route of AKG and NS(745768kJ kgBW�0.62 d�1) as compared to the i.v. route (965754kJ kgBW�0.62 d�1,Po0.005) and i.g. route (918743kJ kgBW�0.62 d�1, Po0.005). Carbohydrate oxidationwas increased with the i.d. route (38.2g73.4 kgBW�0.62 d�1) as compared to the i.v.route (27.872.9 gkgBW�0.62 d�1, Po0.08) and i.g. route (23.978.5 gkgBW�0.62 d�1,Po0.05). Fat oxidation was decreased (2.171.9g kgBW�0.62 d�1; Po0.001) with the
Elsevier Ltd and European Society for Clinical Nutrition and Metabolism. All rights reserved.
08 68 684; fax: +49 38208 68 652.rstorf.de (M. Derno).

ARTICLE IN PRESS
P. Junghans et al.490
i.d. route as compared to the i.v. route (11.571.4 gkgBW�0.62 d�1, Po0.001) and i.g.route (11.973.1g kgBW�0.62 d�1, Po0.001).Conclusions: The i.d. infusion of AKG in combination with the NS affected the wholebody EE and nutrient oxidation, in comparison to that obtained with the i.v. and i.g.routes. It was concluded that the i.d. administration of AKG markedly controlled thenutrient partitioning in the oxidation processes. Finally, in contrary to theobservations with glutamine or glutamate, a considerable percentage of the AKGinfusion was retained in the body irrespective of the route of administration.& 2005 Elsevier Ltd and European Society for Clinical Nutrition and Metabolism. Allrights reserved.
Introduction
a-Ketoglutarate (AKG) is a key intermediate inoxidative metabolism (the conversion of AKG tosuccinate with the loss of CO2 and formation ofNADH) and nitrogen metabolism (the formation ofglutamate by transamination or scavenging freeammonia). Some studies have reported beneficialeffects of AKG and glutamine in clinical nutrition,particularly with respect to gut integrity. Theavailability of glutamine as an enterocyte fuelsubstrate is essential for the preservation of afunctional barrier to microorganisms.1 The enteraladministration of AKG in the early postoperativephase may improve gut integrity and reduceinfectious complications after trauma and surgeryin man. However, enrichment of a whole protein-based formula with AKG did not improve proteinmetabolism or decrease muscle catabolism aftermajor abdominal surgery.2 Glutamine and AKGsupplementation have prevented the decrease ofmuscle free glutamine concentration and influ-enced protein synthesis after surgery.3 Arginino-AKG-fed rats used as a trauma model (simulated byfemur fracture) show a greater use of arginine formetabolic purposes, including an efficient ureacycle operation.4 AKG treatment early in postnatallife increased bone density in lambs.5 Furthermore,it is known that AKG participates in the non-enzymatic oxidative decarboxylation during hydro-gen peroxide decomposition, and enhances themetabolism of fats which can suppress oxygenradical generation and thus prevent lipid perox-idative damage.6 It has been proposed to includeAKG in nutritional formulas, because AKG couldreplace dietary dispensable amino acids by shunt-ing ammonium back into the dispensable aminoacid pool.7 Nevertheless, knowledge about the roleof para-mitochondrial AKG in the whole bodymetabolism, especially that of the gut, is still notcomplete and remains obscure.8 Therefore, theobjective of these studies presented here isto determine how the route of administration
(intraduodenal, i.d.; intragastric, i.g.; and intra-venous, i.v.) affects AKG utilization, whole bodyenergy expenditure (EE), and nutrient oxidation.
Materials and methods
Animals and surgical procedures
The study protocol was approved by the AnimalCare and Use Committee of the Ministry ofNutrition, Agriculture, Forestry, and Fishery,Schwerin, State Mecklenburg-Vorpommern, Ger-many (permission No. VI 522a-7221.31-1-018/99).Thirty-six castrated male pigs (German Landrace)with initial body weight (BW) of about 15 kg, and 2months of age were used. About 4 weeks afterweaning, the animals were taken from our own pigexperimental station in Dummerstorf and housed inindividual metabolic cages. During 1 week ofadaptation the animals were fed the diet ‘‘Ferkel-start Plus’’ (Composition: wheat, soy bean meal,corn flakes, wheat bran, milk powder, soy bean oil,minerals, and vitamins; contents: 13.4MJ ME,17.5% crude protein, 1.23% lysine, 4.3% crude fat,4.0% crude fibre, 0.8% Ca, and 0.6% P; Trede & vonPein GmbH, Dammfleth, Germany) and kept undercontrolled ambient temperature (22 1C). The pigswere surgically fitted with jugular, artery, gastricand duodenal catheters for the infusion of nutrientsand AKG, and for blood sampling. Briefly, 7 daysprior to the infusion studies, and following a feed-deprivation period (16 h), pigs were sedated by i.m.injection of 4mg kgBW�1 azaperone (Stresnils,Janssen-Cilag, Germany). To prevent excessivesalivation, 0.04mg kgBW�1 atropine (Atropinumsulfuricum solutum 1%, WDT, Garbsen, Germany)were given. Subsequently, after starting isofluraneanesthesia the pigs were implanted with SILASTICs
medical grade silicone tubing (o.d. 3.18, i.d.1.57mm; Silastics, SEDAT, Irigny, France) in theA. carotis communis, V. jugularis externa, and

ARTICLE IN PRESS
a-ketoglutarate and nutrient utilization 491
duodenum (3–4 cm caudal of Papilla duodeni) orstomach lumen (ca. 3–5 cm caudal of cardia, faciesparietalis of corpus ventriculi), respectively, underaseptic conditions.
Prior to final closure, 2ml antibiotic (TNVJungtiersuspensions, Veyx-Pharma GmbH, Schwar-zenborn, Germany) was administered intraabdom-inally. Immediately post-operatively, the pigsreceived an intramuscular injection of 4ml anti-biotic (TNV Jungtiersuspensions, Veyx-PharmaGmbH, Schwarzenborn, Germany) and a long-lasting painkiller (Rimadyls, Pfizer, Karlsruhe,Germany).
The study was performed in a stepwise fashion.After performing surgery on 36 pigs and makingcertain that the animals were recovered, they wererandomly distributed to the 6 different treatmentcombinations (see below).
Experimental procedures
To explore the effects of the AKG administrationroute and its interaction with the administrationroute of a simultaneous infusion of a complete
Table 1 Composition and nutrient contents of the nutri
Energy content of nutrient solution: 3.753MJ l�1
Amino acids (g) dissolved in 750ml of solutionAlanine 12.0Arginine 8.5Aspartic acid 2.6phenylalanine 5.9Glutamic acid 4.2Glycine 5.9Histidine 5.1Isoleucine 4.2Leucine 4.2
Glucose (g) dissolved in 1316ml of solutionGlucose (anhydride) 250
Electrolytes (mmol)Na+ 80.0Ca2+ 5.0Mg2+ 10.0K+ 60.0
Vitamins (mg)Thiamine (B1) 2.5Riboflavin (B2) 3.6Pyridoxine (B6) 4.0Pantothenate 15Ascorbic acid (C) 100Cyanocobalamin (B12) 0.005
nutrient solution (NS) (see Table 1 for contents) onAKG utilization, 6 combinations of administrationroutes were tested: Treatment 1, AKG i.d. and NSi.d.; Treatment 2, AKG i.d. and NS i.v.; Treatment3, AKG i.v. and NS i.v.; Treatment 4, AKG i.v. and NSi.d.; Treatment 5, AKG i.v. and NS i.g.; Treatment6, AKG i.g. and NS i.g. (see also Table 2).
The animals were given an infusion of NSconsisting of KabivenTM emulsion supplementedwith SoluvitTM vitamin solution (both from Frese-nius Kabi, Uppsala, Sweden) (Table 1). The NS wascontinuously administered either i.d., i.v. or i.g.,respectively, at a rate of 75ml h�1 by means ofperistaltic pumps (VEB MLW Labortechnik, Ilmenau,Germany). Typically, a pig of 15 kg BW was dailyinfused with about 1800ml NS. This amountcovered 1.7 fold of the energy requirement formaintenance (750 kJ kgBW�0.62 d�1) for pigs of thatweight.9 The NS infusion was started immediatelyafter surgery (d 1) to permit adaptation to thisnutrient source, whereas the infusion of AKG (1 gAKG kgBW�1 d�1, dissolved in sterile saline E5 gAKG 100ml�1) was started on d 3. Prior andbetween infusions of AKG and NS, the catheterswere kept open by a constant infusion (2ml h�1) of
ent solution.
Lysine 6.8Methionine 4.2Proline 5.1Serine 3.4Threonine 4.2Tryptophan 1.4Tyrosine 0.17Valine 5.5
Lipids (g) emulsified in 500ml of solutionPurified soybean oil 100Purified chicken eggs lecithin 6Glycerol anhydride 11
CO32� 97.0
PO43� 17.5
Cl� 116.0
Nicotinamide 40Biotin 0.06Folic acid 0.4Methyl-parahydroxybenzoate 0.5

ARTICLE IN PRESS
P. Junghans et al.492
physiological saline containing heparin(10,000 IU l�1; Heparin-sodium, B. Braun MelsungenAG, Melsungen, Germany).
On d 3 after surgery, the animals housed inmetabolic cages were moved to open-circuitrespiration chambers (4m3, 24 1C environmentaltemperature, 50% relative humidity).
After 3d of adaptation to the chambers, the 24-hCO2 production and oxygen consumption weremeasured in each pig for a period of 3d underconstant nutrient supply. On d 7 after surgery,15mgkgBW�1 [1-13C]AKG (Chemotrade, Leipzig,Germany) dissolved in 50ml saline solution wasinfused for a period of 3h via one of the cathetersby means of syringe pumps (Perfusor compacts, B/Braun, Melsungen AG, Germany). To calculate theAKG utilization, the gas samples were drawn from therespiration chambers prior to (�24, �1h) and attimed intervals every 30min from time 0 until 5 h,and thereafter at 6, 9, 12 and 24h after the start ofthe [1-13C]AKG infusion. Air samples were transferredinto breath bags (Tesseraux, Burstadt, Germany) forsubsequent measurement of the 13C/12C ratio inbreath CO2 by means of the IRIS infrared 13C isotopeanalyser (WATV GmbH, Bremen, Germany.10 One hourbefore and 3h after starting the [1-13C]AKG infusion,blood samples were taken to derive the AKG carbonincorporation into plasma proteins. Plasma proteinswere isolated from 200ml heparin plasma by adding2ml methanol (�20 1C), vortexing for 1min andcentrifugation (1500gmin�1, 4 1C, 10min), as de-scribed by Wessel and Flugge.11 The supernatant wasremoved and the protein pellet was gently driedunder a N2 stream. The 13C/12C ratio in the plasmaprotein was measured by an element analyser (EA1108, Fisons Instr. Rodano, Milan, Italy) coupledonline to an isotope ratio mass spectrometer system(Delta S, Finnigan MAT, Bremen).
The enzymatic activities of g-glutamyl transpep-tidase (GGT), glutamate dehydrogenase (GLDH),and aspartate aminotransferase (ASAT) and theplasma urea and protein concentrations weredetermined using standard test kits purchased fromBoehringer Mannheim (Mannheim, Germany). Theenzymes were chosen to examine essential meta-bolic functions of AKG, such as, glutathionemetabolism and redox homeostasis,6 AKG-gluta-mate-aspartate interconversion, and NH3 detoxifi-cation via glutamate and aspartate to urea.
Calculations
The AKG utilization AKG UTIL (% of 13C dose) wasestimated as
AKG UTIL ¼ 100� ð13C AKG recovery=0:62Þ. (1)
The 13C AKG recovery (% of 13C dose) wascalculated as the ratio of the 13CO2 production(mmol h�1) to the infusion rate of [1-13C]AKG(mmol h�1)
13C AKG recovery
¼13CO2 production=infusion rate of ½13C�AKG;
ð2Þ
where 13CO2 production ¼ CO2 production(mmol h�1)� 13CO2 enrichment (APE/100).
The cumulative 13C recovery at 24 h was used tocalculate the AKG utilization (Eq. (1)).
Because carbon dioxide from tracer oxidation isnot quantitatively recovered in the breath,12 the13CO2 production has to be corrected for incom-plete recovery of 13CO2 in the exhalation. Thecorrection factor for the specific experimentalconditions was determined by means of infusing13C labelled sodium bicarbonate and determiningthe recovery. It amounted to a mean value of 0.62for Treatments 1, 4 and 6, respectively (Junghanset al., unpublished results).
The 13C incorporation into total plasma proteinPP INCORP (% of 13C dose) was calculated as
PP INCORP ¼ 13C enrichment of plasma protein
� plasma protein pool=13C dose; ð3Þ
where
plasma protein pool ðmol CÞ
¼ concentration of plasma protein
ðmol C� l�1Þ � VprotðlÞ. ð4Þ
The concentration of the plasma protein (molC l�1) was calculated from the concentration ofplasma protein (g protein l�1)� 0.5/12, i.e., thecarbon content in plasma protein was considered tobe about 50%.13 The distribution volume of theplasma protein (Vprot) was approximated at80ml kg BW�1.14
The whole body EE (kJ d�1) was calculated fromthe CO2 production (l d�1), O2 consumption (l d�1)and urinary N excretion (g N d�1), using theequation of Brouwer (1965)15:
EE ¼ 5:16� CO2 productionþ 16:18
� O2 consumption� 5:90� N excretion: ð5Þ
The oxidation rate of carbohydrates (CHOX;g d�1) and fat (FatOX; g d�1) were calculated from
CHOX ¼ 4:57� CO2 production� 3:23
� O2 consumption� 2:60� N excretion;
ð6Þ
ARTICLE IN PRESS
a-ketoglutarate and nutrient utilization 493
FatOX ¼ 1:69� O2 consumption� 1:69
� CO2 production� 2:03� N excretion
ð7Þ
as reported by Simonson and DeFronzo.16 For Nexcretion, an estimated value of 2 g N d�1 wastaken in Eqs. (5)–(7).
Statistical analysis
For the detection of statistically significant differ-ences between the means of the metabolic para-meters shown in Tables 2 and 3 for theexperimental groups, Treatments 1–6, a one wayvariance analysis model was used with Tukey–Kra-mer tests to ensure a multiple test risk of first kinda ¼ 0:05. All computations were performed withthe GLM procedure of SAS software, version 8.2.17
Table 2 AKG utilization, whole-body energy expenditurintraduodenal (i.d.), intravenous (i.v.), and intragastricsolution (NS), respectively.
Administration route
AKG: i.d.NS: i.d. i.v.Treatment: 1 2No. pigs: 6 6
AKG UTIL; % 40.176.7a 38.174.1a
EE, kJ kgBW�0.62 d�1 745768b 794731b
CHOX, g kgBW�0.62 d�1 38.273.4c 26.277.7FatOX, g kgBW�0.62 d�1 2.171.9d 9.270.7
aTreatment 1 and 2 vs. Treatment 3–6, Po0.0001; bTreatment 1 an2, 5 and 6, Po0.05; cTreatment 1 vs. Treatment 3, Po0.06; cTreat2–6, Po0.001.
Table 3 Effect of AKG on blood parameters in pigs after i(i.g.) administration of AKG and a complete nutrient solu
Administration route
AKG: i.d. iNS: i.d. i.v. iTreatment: 1 2 3No. pigs: 6 6 6
Urea, mmol L�1 1.770.5 8.775.4a 1Plasma protein, g L�1 50.077.3 53.572.4 5ASAT, nkat L�1 4397223 319710 3GLDH, nkat L�1 57.0752.7 22.0711.6 1GGT, nkat L�1 10717404d 5757168 8
aTreatment 2 vs. Treatment 1 and 3–6, Po0.05; bTreatment 5Po0.05; dTreatment 1 vs. Treatment 2, 4 and 6, Po0.05.
Results
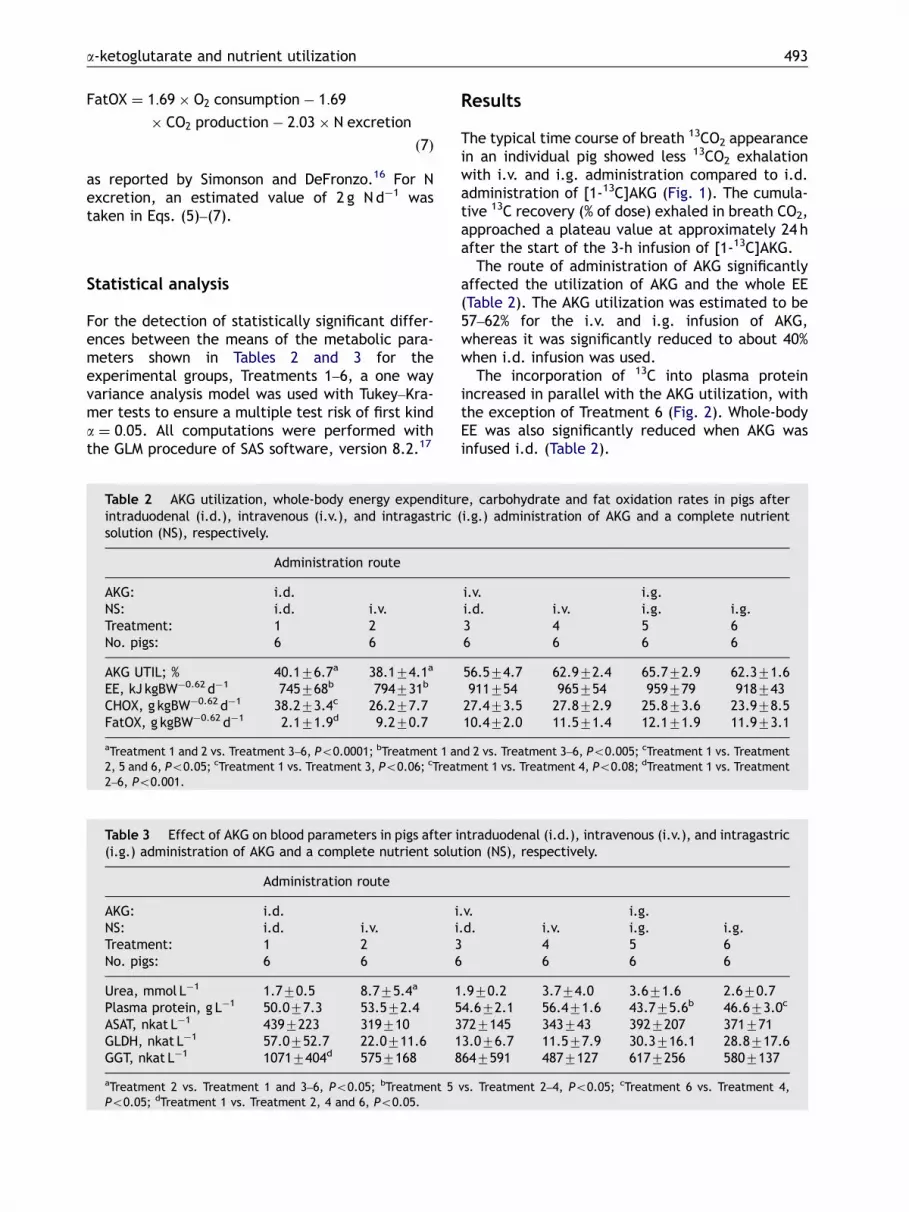
The typical time course of breath 13CO2 appearancein an individual pig showed less 13CO2 exhalationwith i.v. and i.g. administration compared to i.d.administration of [1-13C]AKG (Fig. 1). The cumula-tive 13C recovery (% of dose) exhaled in breath CO2,approached a plateau value at approximately 24 hafter the start of the 3-h infusion of [1-13C]AKG.
The route of administration of AKG significantlyaffected the utilization of AKG and the whole EE(Table 2). The AKG utilization was estimated to be57–62% for the i.v. and i.g. infusion of AKG,whereas it was significantly reduced to about 40%when i.d. infusion was used.
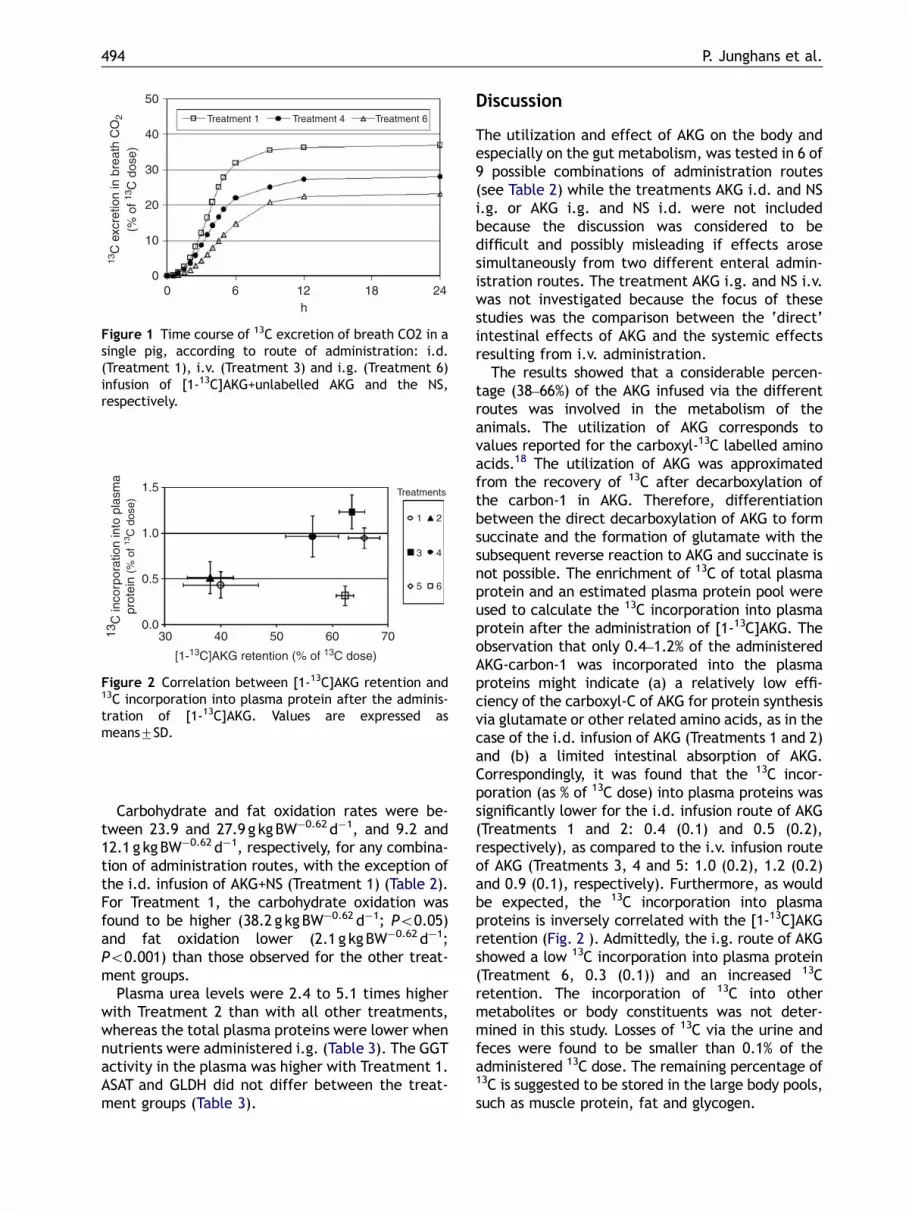
The incorporation of 13C into plasma proteinincreased in parallel with the AKG utilization, withthe exception of Treatment 6 (Fig. 2). Whole-bodyEE was also significantly reduced when AKG wasinfused i.d. (Table 2).
e, carbohydrate and fat oxidation rates in pigs after(i.g.) administration of AKG and a complete nutrient
i.v. i.g.i.d. i.v. i.g. i.g.3 4 5 66 6 6 6
56.574.7 62.972.4 65.772.9 62.371.6911754 965754 959779 91874327.473.5 27.872.9 25.873.6 23.978.510.472.0 11.571.4 12.171.9 11.973.1
d 2 vs. Treatment 3–6, Po0.005; cTreatment 1 vs. Treatmentment 1 vs. Treatment 4, Po0.08; dTreatment 1 vs. Treatment
ntraduodenal (i.d.), intravenous (i.v.), and intragastriction (NS), respectively.
.v. i.g.
.d. i.v. i.g. i.g.4 5 66 6 6
.970.2 3.774.0 3.671.6 2.670.74.672.1 56.471.6 43.775.6b 46.673.0c
727145 343743 3927207 3717713.076.7 11.577.9 30.3716.1 28.8717.6647591 4877127 6177256 5807137
vs. Treatment 2–4, Po0.05; cTreatment 6 vs. Treatment 4,
ARTICLE IN PRESS
50
40
30
20
10
00 6 12 18 24
13C
exc
retio
n in
bre
ath
CO
2
(% o
f 13C
dos
e)
h
Treatment 1 Treatment 4 Treatment 6
Figure 1 Time course of 13C excretion of breath CO2 in asingle pig, according to route of administration: i.d.(Treatment 1), i.v. (Treatment 3) and i.g. (Treatment 6)infusion of [1-13C]AKG+unlabelled AKG and the NS,respectively.
1.5
1.0
0.5
0.0
13C
inco
rpor
atio
n in
to p
lasm
apr
otei
n (%
of 13
C d
ose)
30 40 50 60 70
[1-13C]AKG retention (% of 13C dose)
Treatments
1 2
3 4
5 6
Figure 2 Correlation between [1-13C]AKG retention and13C incorporation into plasma protein after the adminis-tration of [1-13C]AKG. Values are expressed asmeans7SD.
P. Junghans et al.494
Carbohydrate and fat oxidation rates were be-tween 23.9 and 27.9 g kgBW�0.62 d�1, and 9.2 and12.1 g kgBW�0.62 d�1, respectively, for any combina-tion of administration routes, with the exception ofthe i.d. infusion of AKG+NS (Treatment 1) (Table 2).For Treatment 1, the carbohydrate oxidation wasfound to be higher (38.2 g kgBW�0.62 d�1; Po0.05)and fat oxidation lower (2.1 g kgBW�0.62 d�1;Po0.001) than those observed for the other treat-ment groups.
Plasma urea levels were 2.4 to 5.1 times higherwith Treatment 2 than with all other treatments,whereas the total plasma proteins were lower whennutrients were administered i.g. (Table 3). The GGTactivity in the plasma was higher with Treatment 1.ASAT and GLDH did not differ between the treat-ment groups (Table 3).
Discussion
The utilization and effect of AKG on the body andespecially on the gut metabolism, was tested in 6 of9 possible combinations of administration routes(see Table 2) while the treatments AKG i.d. and NSi.g. or AKG i.g. and NS i.d. were not includedbecause the discussion was considered to bedifficult and possibly misleading if effects arosesimultaneously from two different enteral admin-istration routes. The treatment AKG i.g. and NS i.v.was not investigated because the focus of thesestudies was the comparison between the ‘direct’intestinal effects of AKG and the systemic effectsresulting from i.v. administration.
The results showed that a considerable percen-tage (38–66%) of the AKG infused via the differentroutes was involved in the metabolism of theanimals. The utilization of AKG corresponds tovalues reported for the carboxyl-13C labelled aminoacids.18 The utilization of AKG was approximatedfrom the recovery of 13C after decarboxylation ofthe carbon-1 in AKG. Therefore, differentiationbetween the direct decarboxylation of AKG to formsuccinate and the formation of glutamate with thesubsequent reverse reaction to AKG and succinate isnot possible. The enrichment of 13C of total plasmaprotein and an estimated plasma protein pool wereused to calculate the 13C incorporation into plasmaprotein after the administration of [1-13C]AKG. Theobservation that only 0.4–1.2% of the administeredAKG-carbon-1 was incorporated into the plasmaproteins might indicate (a) a relatively low effi-ciency of the carboxyl-C of AKG for protein synthesisvia glutamate or other related amino acids, as in thecase of the i.d. infusion of AKG (Treatments 1 and 2)and (b) a limited intestinal absorption of AKG.Correspondingly, it was found that the 13C incor-poration (as % of 13C dose) into plasma proteins wassignificantly lower for the i.d. infusion route of AKG(Treatments 1 and 2: 0.4 (0.1) and 0.5 (0.2),respectively), as compared to the i.v. infusion routeof AKG (Treatments 3, 4 and 5: 1.0 (0.2), 1.2 (0.2)and 0.9 (0.1), respectively). Furthermore, as wouldbe expected, the 13C incorporation into plasmaproteins is inversely correlated with the [1-13C]AKGretention (Fig. 2 ). Admittedly, the i.g. route of AKGshowed a low 13C incorporation into plasma protein(Treatment 6, 0.3 (0.1)) and an increased 13Cretention. The incorporation of 13C into othermetabolites or body constituents was not deter-mined in this study. Losses of 13C via the urine andfeces were found to be smaller than 0.1% of theadministered 13C dose. The remaining percentage of13C is suggested to be stored in the large body pools,such as muscle protein, fat and glycogen.

ARTICLE IN PRESS
a-ketoglutarate and nutrient utilization 495
These results indicated clearly that the i.d.infusion of AKG might improve the utilization ofnutrients, as measured by indirect calorimetry.Thus, the EE under i.d. AKG infusion was only745–794 kJ kgBW�0.62 d�1, as compared to i.v. ori.g. infusion having 911 to 965 kJ kgBW�0.62 d�1.Conversely, the utilization of AKG was less for thei.d. administration route. This observation mightindicate that the particular function of AKG is anoxidative fuel for the gut (formation of CO2 andNADH), and that its role is to spare fuels such asglutamate and aspartate.8 The observation of thelower utilization of AKG when administered i.d.also corresponded to previous findings, where onlya small percentage (no more than 11%) of the AKGadministered via the intestinal lumen to pigsappeared in the portal circulation.19 Similarly,Reeds et al.20 did not find a portal uptake ofglutamate in fed suckling pigs. Thus, they con-cluded that the gut metabolized virtually all of theenteral glutamate during absorption.
Different metabolic effects of nutrients accord-ing to the enteral administration routes (i.d. vs.i.g.) were also found for carbohydrates. Thus,these effects might result from different signalscaused by the interaction of nutrients with thereceptors in the gastric and intestinal regions andthe release of gastrointestinal hormones, such as,amylin and glucagon-like peptide-1.21
The mode of action of AKG administered i.d. vs.i.g./i.v. is not known. However, from the clinicalpoint of view, the result is certainly important sinceenteral nutrition is routinely recommended forpostoperative patients. Information about theenergy saving and thus anabolic features ofintestinal AKG supply vs. the oral (gastric) and i.v.routes is important when preparing new nutrientformulas and guidelines for their administrationroutes.
The enzymatic activities of GGT, GLDH, and ASATcan reflect essential metabolic functions of AKG, asmentioned above. The observed increase in thecatalytic activities of GGT might suggest anincrease in the intestinal metabolism in responseto the i.d. infusion of AKG (Treatment 1 vs.Treatments 2, 4 and 6).
Tracer studies using 13C-labelled AKG allowed usfor the first time to quantify the utilization of AKG.The combination of the stable isotope approachwith indirect calorimetry opened the possibility to‘‘trace’’ AKG and to assess the impact of AKG onwhole body EE and nutrient oxidation at the sametime.
In summary, AKG infused via different routes maybe utilized by a considerable percentage of 38–66%.I.d. infused AKG is utilized much less than via the
i.v. and i.g. routes. The i.d. infusion of AKG leads toa decrease in the whole body EE which correspondsto the nutrient-sparing effects of AKG by itsfunction as an oxidative fuel.
As the i.d. administration of AKG (Treatments 1and 2) may markedly affect EE and nutrientutilization, in comparison to the other administra-tion routes (i.v., i.g.), therapeutic strategies usedin clinical nutrition should be reassessed.
Bypassing the gastric tract with its specificcharacteristics (acidic pH, chemical and enzymaticconversions, emptying delay, etc.), i.d. adminis-tration could trigger a metabolic effect due to afaster and more efficient targeting of substrates,such as, AKG for the organism/patients.
Implications
The results obtained in this study can be useful inclinical nutrition. The contemporary problem inclinical nutrition is to choose optimal nutrientcompositions and efficient routes of administrationadequate to the metabolic status (malnutrition,rachitic, etc.) of the patient. We believe that theeffect of AKG on and in the gut and whole bodymetabolism observed in this pig model can provideimportant information about nutrient utilizationfor the clinician both in human and veterinarymedicine. Finally, it is demonstrated for the firsttime that the indispensable enterocyte nutrient ofthe glutamine family is not totally metabolized toCO2 after enteral administration.
Acknowledgements
We gratefully acknowledge Dr. Lorrine Svendsenand Dr. Cornelia C. Metges for valuable commentson the manuscript.
References
1. Clark EC, Patel SD, Chadwick PR, et al. Glutamine depriva-tion facilitates tumour necrosis factor induced bacterialtranslocation in Caco-2 cells by depletion of enterocyte fuelsubstrate. Gut 2003;52:224–30.
2. Wiren M, Permert J, Larsson J. Alpha-ketoglutarate-supple-mented enteral nutrition: effects on postoperativenitrogen balance and muscle catabolism. Nutrition 2002;18:725–8.
3. Blomqvist BI, Hammarqvist F, von der Decken A, et al.Glutamine and alpha-ketoglutarate prevent the decrease inmuscle free glutamine concentration and influence proteinsynthesis after total hip replacement. Metabolism 1995;44:1215–22.
4. Jeevanandam M, Begay CK, Holaday NJ, et al. Nutritionaland metabolic effects and significance of mild orotic

ARTICLE IN PRESS
P. Junghans et al.496
aciduria during dietary supplementation with arginine or itsorganic salts after trauma injury in rats. Metabolism1997;46:785–92.
5. Harrison AP, Tygesen MP, Sawa-Wojtanowicz B, et al. Alpha-ketoglutarate treatment early in postnatal life improvesbone density in lambs at slaughter. Bone 2004;35:204–9.
6. Velvizhi S, Dakshayani KB, Subramanian P. Effects of alpha-ketoglutarate on antioxidants and lipid peroxidation pro-ducts in rats treated with ammonium acetate. Nutrition2002;18:747–50.
7. Pierzynowski SG, Sjodin A. A perspective of glutamine andits derivates. J Anim Feed Sci 1998;7:79–91.
8. Lambert BD, Stoll B, Niinikoski H, et al. Net portalabsorption of enterally fed alpha-ketoglutarate is limitedin young pigs. J Nutr 2002;132:3383–6.
9. Gesellschaft fur Ernahrungsphysiologie der Haustiere (GEH).Energie- und Nahrstoffbedarf landwirtschaftlicher Nutz-tiere. Nr. 4: Versorgungsempfehlungen fur Mastschweine.DLG-Verlag, Frankfurt, Main, 1987.
10. Junghans P, Derno M, Gehre M, et al. Calorimetric validationof 13C bicarbonate and doubly labelled water method fordetermining the energy expenditure in goats. Z Ernahrungs-wiss 1997;36:268–72.
11. Wessel D, Flugge UI. A method for the quantitative recoveryof protein in dilute solution in the presence of detergentsand lipids. Anal Biochem 1984;138:141–3.
12. Leijssen DP, Elia M. Recovery of 13CO2 and 14CO2 in humanbicarbonate studies: a critical review with original data.Clin Sci 1996;91:665–77.
13. Hoffmann L, Schiemann R. From calorie to joule. Newproportions in measuring energy metabolism and calculating
the parameters of energetic feed evaluation. Arch Tier-ernahr 1980;30:733–42 [in German].
14. Kaboth W. In: Begemann H, editor. Klinische Hamatologie.Stuttgart: Georg Thieme; 1975.
15. Brouwer E. Report of sub-committee on constants andfactors. In: Blaxter KL, editor. Proceedings of the thirdsymposium on energy metabolism. London, New York; 1965.p. 441–3.
16. Simonson DC, DeFronzo RA. Indirect calorimetry: methodo-logical and interpretative problems. Am J Physiol 1990;258:E399–412.
17. SAS Institute Inc. SAS/STATs user’s guide, version 8. Cary,NC: SAS Institute Inc.; 1999.
18. Junghans P, Beyer M, Saggau E, et al. Estimate of thepostabsorptive protein and fat metabolism in growing pigsafter long-term feeding of diets with soy protein isolate orcasein using a 13C,15N-amino acid approach. In: McCrackenKJ, Unsworth EF, Wylie ARG, editors. Energy metabolism inanimals. Proceedings of the 14th symposium on energymetabolism in farm animals. Wallingford, UK: CAB Interna-tional; 1998. p. 181–4.
19. Buddington RK, Pajor A, Buddington KK, et al. Absorption ofalpha-ketoglutarate by the gastrointestinal tract of pigs.Comp Biochem Physiol A: Mol Integr Physiol 2004;138:215–20.
20. Reeds PJ, Burrin DG, Jahoor F, et al. Enteral glutamateis almost completely metabolized in first pass by thegastrointestinal tract of infant pigs. Am J Physiol 1996;270:E413–8.
21. Feinle C, O’Donovan D, Horowitz M. Carbohydrate andsatiety. Nutr Rev 2002;60:155–69.












![Regulation of Lysine Catabolism through Lysine[mdash]Ketoglutarate Reductase and Saccharopine Dehydrogenase in Arabidopsis](https://static.fdokumen.com/doc/165x107/631cc83693f371de19019c93/regulation-of-lysine-catabolism-through-lysinemdashketoglutarate-reductase-and.jpg)






