
Inhibition of 20-HETE Production Contributes to the Vascular Responses to Nitric Oxide
7
Richard J. Roman Magdalena Alonso-Galicia, Heather A. Drummond, K. Kishta Reddy, John R. Falck and Oxide Inhibition of 20-HETE Production Contributes to the Vascular Responses to Nitric Print ISSN: 0194-911X. Online ISSN: 1524-4563 Copyright © 1997 American Heart Association, Inc. All rights reserved. is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231 Hypertension doi: 10.1161/01.HYP.29.1.320 1997;29:320-325 Hypertension. http://hyper.ahajournals.org/content/29/1/320 the World Wide Web at: The online version of this article, along with updated information and services, is located on http://hyper.ahajournals.org//subscriptions/ is online at: Hypertension Information about subscribing to Subscriptions: http://www.lww.com/reprints Information about reprints can be found online at: Reprints: document. Permissions and Rights Question and Answer information about this process is available in the is located, click Request Permissions in the middle column of the Web page under Services. Further requested the Editorial Office. Once the online version of the published article for which permission is being can be obtained via RightsLink, a service of the Copyright Clearance Center, not Hypertension published in Requests for permissions to reproduce figures, tables, or portions of articles originally Permissions: by guest on January 19, 2014 http://hyper.ahajournals.org/ Downloaded from by guest on January 19, 2014 http://hyper.ahajournals.org/ Downloaded from
-
Upload
musicwritingteam -
Category
Documents
-
view
1 -
download
0
Transcript of Inhibition of 20-HETE Production Contributes to the Vascular Responses to Nitric Oxide

Richard J. RomanMagdalena Alonso-Galicia, Heather A. Drummond, K. Kishta Reddy, John R. Falck and
OxideInhibition of 20-HETE Production Contributes to the Vascular Responses to Nitric
Print ISSN: 0194-911X. Online ISSN: 1524-4563 Copyright © 1997 American Heart Association, Inc. All rights reserved.
is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Hypertension doi: 10.1161/01.HYP.29.1.320
1997;29:320-325Hypertension.
http://hyper.ahajournals.org/content/29/1/320the World Wide Web at:
The online version of this article, along with updated information and services, is located on
http://hyper.ahajournals.org//subscriptions/
is online at: Hypertension Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer information about this process is available in the
is located, click Request Permissions in the middle column of the Web page under Services. Further requestedthe Editorial Office. Once the online version of the published article for which permission is being
can be obtained via RightsLink, a service of the Copyright Clearance Center, notHypertensionpublished in Requests for permissions to reproduce figures, tables, or portions of articles originallyPermissions:
by guest on January 19, 2014http://hyper.ahajournals.org/Downloaded from by guest on January 19, 2014http://hyper.ahajournals.org/Downloaded from

320
Inhibition of 20-HETE Production Contributes to the Vascular Responses to Nitric Oxide
Magdalena Alonso-Galicia, Heather A Drummond, K lshta Reddy, John R Falck, Richard J. Roman
Abstract N~tnc oxide (NO) Inhibits a variety of heme-contam- mg enzymes, mcludmg NO synthase and cytochrome P4501Al and 2Bl The present study exammed whether NO mhlblts the production of 20-hydroxyelcosatetraenolc acid (20-HETE) by cy- tochrome P4504A enzymes and whether blockade of the produc- tion of this substance contributes to the vascular effects of NO Sodium mtroprusslde (SNP, lo-‘, lo-“, and 10m3 mol/L) reduced the productlon of 20-HETE by renal mlcrosomes incubated with arachldomc acid to 71+5%, 29t4%, and 4+2% of control, re- spectlvely (n=S) Slmllar results were obtained with the use of 1-propanamme, 3-(2-hydroxy-2-mtroso-1-propylhydrazmo) (n=3) To determme whether mhlbitlon of 20-HETE contributes to the vasodllatory effects of NO, the effects of dlbromo-dodecenyl- methylsulfinude (DDMS), a selective mhlbltor of the formatlon of 20-HETE, on the response to SNP (lo-’ to 10m3 mol/L) were examined m rat renal artenoles preconstructed with phenylephrme (n=.5) SNP increased vascular diameter m a concentrafion-depen- dent manner to 82t4% of control After DDMS (25 ymol/L), SNP (lo-’ mol/L) increased vascular diameter by only 17+3% The
effects of DDMS on the mean arterial pressure (MAP) and renal blood flow (RBF) responses to mfuslon of an NO donor and a synthase mhibltor were also examined m thlobutabarbltal-anesthe- trzed, Sprague-Dawley rats Infusion of MAHMA NONOate at 1, 3, 5, and 10 nmol/mm reduced MAP by 16+2, 3023,40t5, and 48?5 mm Hg and lowered renal vascular resistance (RVR) by 15+3%, 26?2%, 30?3%, and 34?4% of control After DDMS (10 mg/kg, n=7 rats), the MAP and RVR responses to I-hexamme, 6-(2-hydroxy- 1 -methyl-2-mtrohydrazmo)N-methyl (MAHMA NONOate) averaged only 20% of those seen durmg control In other expenments, MAP Increased by 3224% and RBF fell to 56+5% of control after admmlstratlon of N-mtro-L-argmme (L-NArg) (10 mg/kg IV) After DDMS (10 mg/kg, n=7 rats), MAP Increased by only 19+4% and RBF fell by only 724% after L-NArg These results indicate that NO mhlblts cytochrome P4504A enzymes and that mhlbltlon of the production of 20-HETE contributes to the vasodllatory effects of NO (Hypertension. 1997;29[part 2]:320-325.)
Key Words l mtnc oxide l vasculature l enzymes
R ecent studies have mdlcated that the effects of many renal vasodllators are dependent on the re- lease of NO from the endothehum Blockade of
NO synthesis increases arterial pressure, decreases RBF, and potentiates tubuloglomerular feedback responses. 1 These results indicate that tonic release of NO plays an important modulatory role m the regulation of both renal and penpheral vascular tone. It 1s generally assumed that the vasodllatory effects of NO are mediated by cGMP sec- ondary to shmulatlon of guanylyl cyclase 2.3 This conclu- slon 1s based on the observations that endothehum-depen- dent vasodilators and NO donors increase cGMP m vascular tissue and that methylene blue and other mhlbltors of guanylyl cyclase m many vessels can elm-nnate the va- sodilatory response However, this generalized scheme for NO-induced vasodllatlon has been questioned recently be- cause there are an mcreasmg number of reports that NO donors and/or endothelmm-dependent dilators can produce vasodllatlon and/or membrane hyperpolarlzatlon m some vessels m the presence of guanylyl cyclase mhibltors 4-6
In the renal rmcrocn-culatlon, there have been no direct pharmacological or biochemical studies to examme the mechanisms by which NO or endothehum-dependent dl- lators alter renal vascular tone 1 It 1s not known whether
From the Department of Physiology, Medical College of Wlscon- sm, Milwaukee (M A Xi, H A D , R J R ), and the Department of Molecular Genetics, Unlverslty of Texas Southwestern Medlcal Cen- ter, Dallas (K K R , J R F )
Correspondence to Dr Richard J Roman, Department of Physlol- ogy, Medical College of Wisconsin, 8701 Watertown Plank Rd, Md- waukee, WI 53226-0509 E-mad rroman@post its mew edu
0 1997 American Heart Assoclatlon, Inc
the renal vasodllatory response to NO 1s dependent on the generation of cGMP or if a “cGMP-independent” wg- nalmg pathway also contributes to this response. In this regard, it should be noted that NO has recently been re- ported to inhibit NO synthase7,8 and P450 enzymes of the 1A and 2B 19 and 3C’O families by forming iron-mtrosyl complexes at the catalytic heme bmdmg site m these en- zymes There is also a large body of evidence attributing the toxic effects of nitrates on drug metabolism m the liver to the formatlon of NO from nitrates and the sub- sequent mhlbltlon of P450 enzymes. 1~12 In light of the recent studies from our laboratory indicating that renal vascular smooth muscle cells metabolize AA through a P4504A-dependent pathway to the potent vasoconstrlctor 20-HETE,l3.14 and that this system plays a central role m the regulation of renal vascular tone,ls-‘7 it seems likely that NO might mhlblt the formation of 20-HETE m renal vascular smooth muscle and that this rmght contribute to the vasodllatory effects of NO Therefore, the purpose of the present study was to determme whether NO mhlblts enzymes of the P4504A farmly and whether blockade of the production of 20-HETE contrlbutes to the effects of NO on renal vascular tone, both m vlvo and m vitro.
General Methods
Experiments were performed on lo- to 12-week-old male Sprague-Dawley rats purchased from Harlan Sprague Dawley Laboratones (Indlanapohs, Ind) The rats were housed m an an- imal care faclhty at the Medlcal College of Wlsconsm, which 1s approved by the American Assoclatlon for the Accreditation of Laboratory Ammal Care, and had free access to food and water
by guest on January 19, 2014http://hyper.ahajournals.org/Downloaded from

Alonso-Galicza et al 20-HETE, NO, and Renal Vasculature 321
Selected Abbreviations and Acronyms AA = arach1domc acid
DDMS = dlbromo-dodecenyl-methylsulfim1de D1HETEs = dlhydroxyelcosatetraenolc acids
EETs = elcosatnenolc acids 20-HETE = 20-hydroxyelcosatetraenolc acid
HPLC = high-performance 11qu1d chromatography
L-NArg = N-nltro-L-arglrnne MAHMA NONate = 1-hexamine, 6-(2-hydroxy-l-methyl-
2-nitrosohydrazlno)N-methyl MAP = mean artenal pressure
NO = nitric oxide 17-ODYA = 17-octadecynolc acid
PAPA NONOate = 1-propanamine, 3-(2-hydroxy-2- nitroso-l-propylhydraz1no)
RBF = renal blood flow SNP = sodium rutroprusslde
All protocols mvolvlng animals received approval by the Animal Care Committee of the Medical College of Wlsconsln
Renal Metabolism of AA Adult male rats were anesthetized with an lntraperltoneal m-
Jection of pentobarbltal (50 mg/kg) The ludneys were removed and placed 1n ice-cold 0 9% NaCl solution The renal cortex was homogemzed 1n a 10 mmol/L potassmm phosphate buffer (pH 7 7) containing 250 mmol/L sucrose, 1 mmol/L EDTA, and 10 mmol/L magnesium chloride, and mlcrosomes were prepared by differential centrifugation as we have previously described l8 P4504A enzyme activity was assayed by incubating renal cortical microsomes (0 5 mg) for 30 minutes at 37°C with [ l-“C] AA (0.1 &I, 50 pmol/L, Amersham Carp) 1n 1 mL of a 100 mmol/L potassium phosphate buffer (pH 7 4) containing 5 mmol/L MgCla, 1 mmol/L EDTA, 1 mmol/L NADPH, and an NADPH-regenerating system (10 mmol/L 1soc1trate and ISOCI- trate dehydrogenase, 0 4 unit/ml) under control condltlons and after addition of the various compounds to be tested The reac- t1ons were terminated by acidification to pH 4 through the use of 0 1 moliL formic acid and followed by extraction of the tissue and media with ethyl acetate Metabolltes were separated through the use of a 25 cmx2 mm ID (Supelco Inc) C18-reverse-phase HPLC column and a linear elut1on gradient ranging from aceto- nitnle/water/acetm acid (50/50/O 1) to acetonltrde/acetlc acid (100/O 1) over a 40-minute period The radioactive products were monitored with the use of a radioactive flow detector (model 120, Radlomatlc Instrument Co)
Protocol 1: Effects of NO Donors on AA Metabolism Renal cortical mtcrosomes from male rats were incubated with
[ I-‘4C] AA for 30 minutes in the presence of NADPH under control condltlons and after addition of various concentrations of SNP (lo-‘, lo-“, and 10d3 mol/L) or PAPA NONOate (10e5, 10m4, and lo-’ mol/L) (Cayman Chemical Co), a non-cyamde- releasing NO donor, to the mcubatlon The reactions were ter- minated by acldlficatlon to pH 4 through the use of 0 1 mol/L formic acid and followed by a 11p1d extraction of the tissue and media Metabolltes were then separated by reverse-phase HPLC as described above
Protocol 2: Role of 20-HETE in the Response to NO Donors in Isolated, Perfused Renal Arterioles
Experiments were performed on isolated, perfused renal arte- rioles (< 100 pm) mlcrodlssected from the kidneys of adult male rats Rats were anestheuzed with sodium pentobarbltal (50 mg/kg IP) The left kidney was removed and placed m ice-cold physlologlcal salme solution (PSS) containing (1n mmol/L) 119 NaCl, 4 7 KCl, 1 6 CaC12, 1 2 MgSO,,, 1 2 NaH2P04, 12 NaHCO,, 10 glucose, and 0 03 EDTA, pH 7 4 The kidney was
hemldlssected, and small interlobular arterioles with 50- to lOO- pm ID were removed by mlcrod1ssect1on. The vessels were mounted on glass micropipettes in a water-jacketed perfusion chamber containing PSS that was equilibrated with a 95% 02- 5% CO* gas mixture and maintained at 37°C Indomethacm (5 pmol/L, Sigma Chemical Co) and ba1cale1n (0 5 pmol/L, Blomol) were added to the bath to block the endogenous metabolism of AA through the cyclooxygenase and hpoxygenase pathways, as we have previously described 19 Vessels were secured to the pi- pettes with 10-O silk suture, and side branches were tied off The inflow pipette was connected to a pressurized reservoir to allow for control of lntralummal perfusion pressure, which was monl- tored with the use of a transducer (Cobe). After mounting, the vessels were stretched to the 1n v1vo length with the use of an eyepiece micrometer, the outflow cannula was clamped off, and 1ntralumlnal pressure was set to 90 mm Hg during the experiment
After a 30-minute equlhbratlon period, the vessels were pre- constricted with phenylephrme (1 bmol/L, Sigma Chemical Co), and a cumulative concentration-response curve for SNP (lo-’ to 1O-3 mol/L, Sigma Chemical Co) was obtained Vascular dlam- eters were measured 1 minute after the addition of SNP to the bath with the use of a video system composed of a stereom1cro- scope (Carl Zeiss, Inc), a CCTV video camera (KP-130AU, Hi- tachi), a videocassette recorder (AG-7300, Panasonic), a video monitor (CVM-1271, Sony), and a video measuring system (VIA-loo, Boeckeler Instrument Co) The bath solution was then exchanged for freshly prepared PSS containing 5 pmol/L indo- methacm, 0 5 pmol/L balcaleln, and 25 pmol/L DDMS (n=5 rats) or vehicle (n=5 rats) Thirty minutes after blockade of the production of 20-HETE by DDMS, an experimental cumulative concentrat1onresponse curve for SNP was obtained after again preconstnctlng the vessels with phenylephrlne (1 pmol/L)
To determine whether the effects of DDMS were specific to NO donors and not caused by a generalized 1nhlb1tory action, we studied the effects of DDMS on the vasodllatory responses to the NO-independent dilators dibutyryl-cGMP and adenoslne In these studies, cumulative concentration-response curves for N2,2’-0-dlbutyrylguanoslne 3’*5’cycl1c monophosphate (dlbu- tyryl-cGMP) (Sigma Chemical Co, n=3 vessels, 3 rats) and aden- osme (Sigma Chemical Co; n=3 vessels, 3 rats) were obtained 1n vessels preconstncted with phenylephrme under control con- ditions and after 30 minutes of blockade of the production of 20- HETE by DDMS (25 pmol/L)
Finally, cumulative concentration-response curves for SNP also were obtained 1n phenylephnne-preconstrlcted renal micro- vessels under control conditions and after addition of 100 nmol/L of 20-HETE to the bath to determine whether preventing the fall 1n 20-HETE levels would block the vasodllatory response to SNP (n=6 vessels, 5 rats).
Protocol 3: Influence of 20-HETE on the Renal and Peripheral Vascular Responses to NO Donors
Adult male rats were anesthetized with ketamlne (30 mg/kg) and thlobutabarbltal (Inachn) (100 mglkg) and maintained at 37°C Cannulas were placed 1n a femoral artery and vein for the measurement of systemic blood pressure and intravenous 1nfu- slons. The rats received an intravenous mfus1on of 0 9% NaCl solution contalmng 3% albumm at a rate of 1 2 mL/h throughout the experiment After surgery and a 30-minute equ111bratlon pe- riod, MAP and RBF (measured with an electromagnetic flow- meter) were recorded during a control period and 1n response to 2-minute intravenous infusion of the very short-acting NO donor MAHMA NONOate (Cayman Chemical Co) at doses of 1,3,5, and 10 nmol/mm This donor was selected because 1t does not accumulate 1n the systemic clrculatlon and blood pressure and RBF rapidly return to control (within 15 seconds) after the in- fusion 1s stopped Five minutes, however, was allowed between each dose to allow for full equ111bratlon of the preparation before testing the effects of the next dose After the control responses were recorded, a 2 mg IV bolus inJection of DDMS (6 mglkg) was given followed by a maintenance infusion at a rate of I 2
by guest on January 19, 2014http://hyper.ahajournals.org/Downloaded from
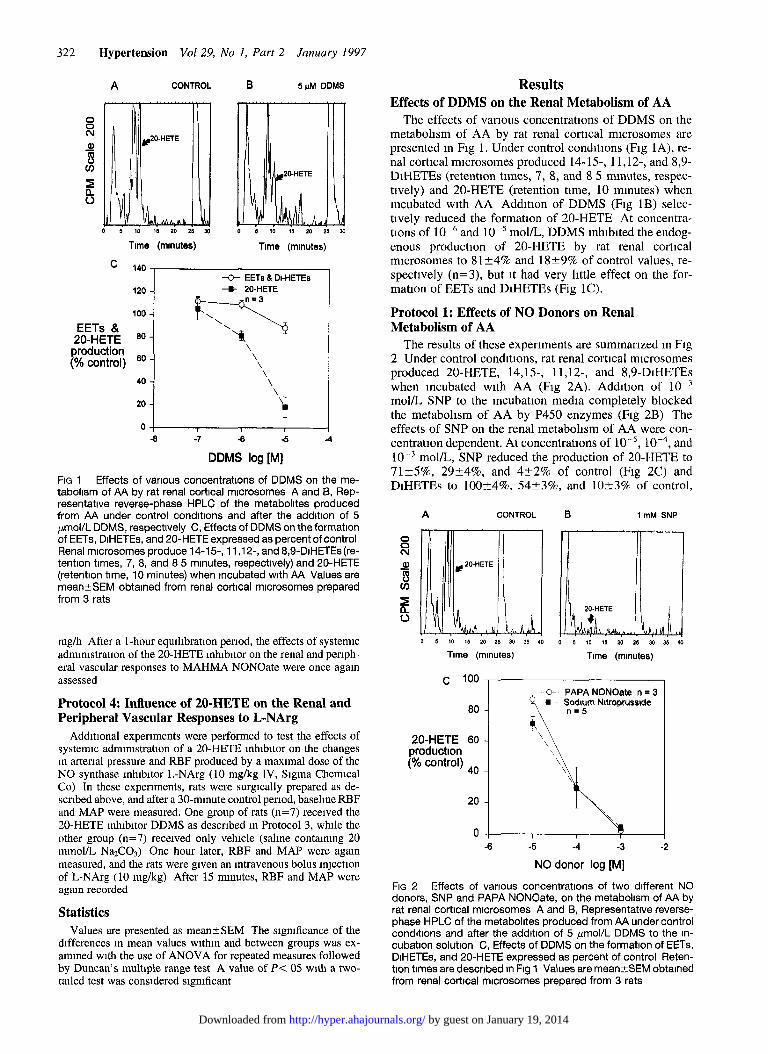
322 Hypertension Vol29, No 1, Part 2 January 1997
A CONTROL B 5pM DDMS
Time (mmutes) Time (minutes)
C 140 -C+ EETs 8 DI-HETEs
120 - + 20-HETE
EETs & looI w PO-HETE so
production e. \ (% control)
\ \ \
DDMS log [M]
FIG 1 Effects of various concentrations of DDMS on the me- taboltsm of AA by rat renal cortical mlcrosomes A and 6, Rep- resentatlve reverse-phase HPLC of the metabolites produced from AA under control conditions and after the addition of 5 pmol/L DDMS, respectively C, Effects of DDMS on the formation of EETs, DiHETEs, and 20-HETE expressed as percent of control Renal microsomes produce 14-l 5, l l ,12-, and 8,9-DIHETEs (re- tention times, 7, 8, and 8 5 minutes, respectively) and 20-HETE (retention time, 10 minutes) when Incubated with AA Values are mean?SEM obtained from renal cortlcal microsomes prepared from 3 rats
mg/h After a l-hour equilibration period, the effects of systemic admnustration of the 20-HETE mhlbltor on the renal and penph- era1 vascular responses to MAHMA NONOate were once again assessed
Protocol 4: Influence of 20-HETE on the Renal and Peripheral Vascular Responses to L-NArg
Additional experiments were performed to test the effects of systemic admnustration of a 20-HETE mtubitor on the changes m arterial pressure and RBF produced by a maximal dose of the NO synthase mlubltor L-NArg (10 mg/kg IV, Sigma Chemical Co) In these experiments, rats were surgically prepared as de- scribed above, and after a 30-minute control period, baseline RBF and MAP were measured. One group of rats (n=7) received the 20-HETE mlubitor DDMS as described m Protocol 3, while the other group (n=7) received only vehicle (saline contammg 20 mmol/L Na2C03) One hour later, RBF and MAP were again measured, and the rats were given an intravenous bolus 1nJection of L-NArg (10 mgikg) After 15 minutes, RBF and MAP were agam recorded
Statistics Values are presented as mean+SEM The significance of the
differences m mean values within and between groups was ex- amined with the use of ANOVA for repeated measures followed by Duncan’s multiple range test A value of P< 05 with a two- tailed test was considered significant
Results Effects of DDMS on the Renal Metabolism of AA
The effects of various concentrations of DDMS on the metabolism of AA by rat renal cortical microsomes are presented m Fig 1, Under control conditions (Fig 1 A), re- nal cortical n-ucrosomes produced 14-15-, 11,12-, and 8,9- DiHETEs (retention times, 7, 8, and 8 5 minutes, respec- tively) and 20-HETE (retention time, 10 minutes) when incubated with AA Adchtlon of DDMS (Fig 1B) selec- tively reduced the formation of 20-HETE At concentra- tions of 10m6 and 10d5 mol/L, DDMS mhiblted the endog- enous productlon of 20-HETE by rat renal cortical rmcrosomes to 8124% and 1829% of control values, re- spectively (n=3), but it had very little effect on the for- mation of EETs and DiHETEs (Fig 1C).
Protocol 1: Effects of NO Donors on Renal Metabolism of AA
The results of these experiments are summanzed m Fig 2 Under control conditions, rat renal cortical rmcrosomes produced 20-HETE, 14,15-, 11,12-, and 8,9-DiHETEs when incubated with AA (Fig 2A). Addition of 10m3 mol/L SNP to the mcubation media completely blocked the metabolism of AA by P450 enzymes (Fig 2B) The effects of SNP on the renal metabolism of AA were con- centration dependent. At concentrations of 10-5, 10m4, and lo-’ mom, SNP reduced the production of 20-HETE to 71?5%, 29?4%, and 422% of control (Fig 2C) and DiHETEs to loo+-4%, 54?3%, and 1023% of control,
CONTROL B 1 mM SNP
Time (minutes) Time (minutes)
PO-HETE 80 -
-6 -5 -4 -3
NO donor log [M]
-2
FIG 2 Effects of various concentrations of two different NO donors, SNP and PAPA NONOate, on the metabolism of AA by rat renal cortical microsomes A and B, Representative reverse- phase HPLC of the metabolltes produced from AA under control conditions and after the addition of 5 pmol/L DDMS to the in- cubation solution C, Effects of DDMS on the formation of EETs, DIHETEs, and 20-HETE expressed as percent of control Reten- tion times are described n Fig 1 Values are meanrSEM obtained from renal cortical mlcrosomes prepared from 3 rats
by guest on January 19, 2014http://hyper.ahajournals.org/Downloaded from
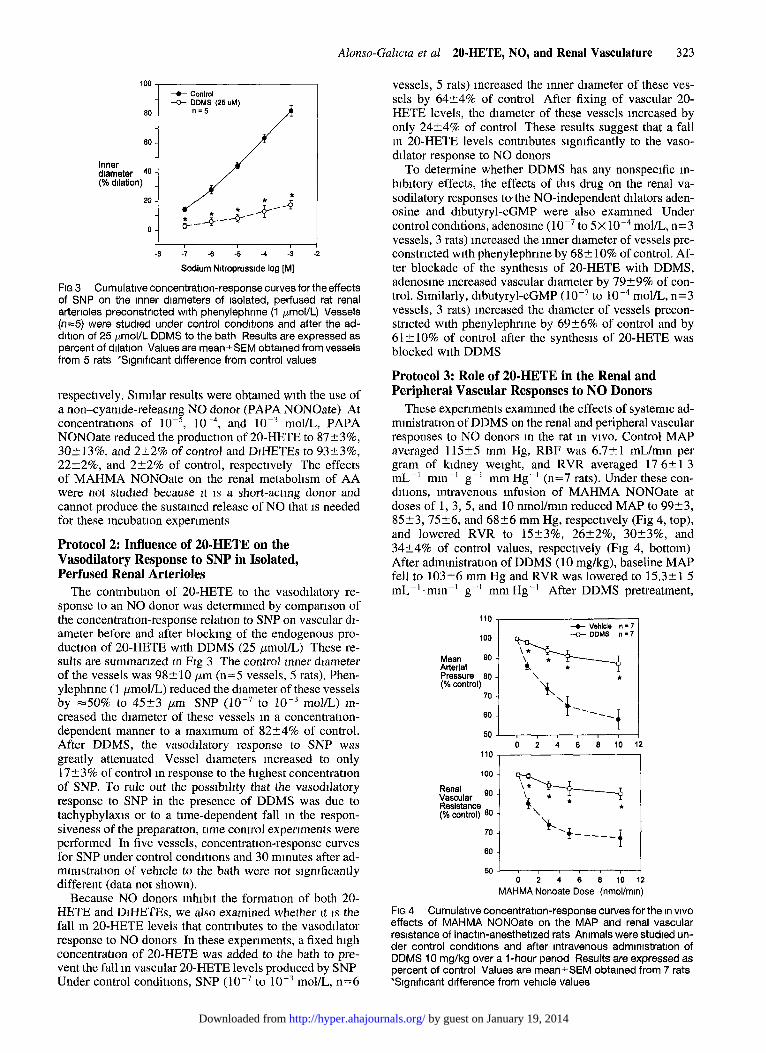
Alonso-Galzcta et al 20.HETE, NO, and Renal Vasculature 323
Inner diameter 4o (% dilation)
0-I I I -6 -7 -6 -5 4 -3 -2
Sodium NWoprusslde log [M]
FIG 3 Cumulative concentration-response curves forthe effects of SNP on the inner diameters of isolated, perfused rat renal arterioles preconstncted with phenylephnne (1 pmol/L) Vessels (n=5) were studied under control condltlons and after the ad- dltlon of 25 pmol/L DDMS to the bath Results are expressed as percent of dllatlon Values are mean+SEM obtained from vessels from 5 rats *Slgnlfcant difference from control values
respectively. Slmllar results were obtained with the use of a non-cyamde-releasmg NO donor (PAPA NONOate) At concentrations of 10m5, 10e4, and lo-’ mol/L, PAPA NONOate reduced the production of 20-HETE to 87+3%, 30?13%, and 2?2% of control and DIHETEs to 93?3%, 22?2%, and 2?2% of control, respectively The effects of MAHMA NONOate on the renal metabohsm of AA were not studled because It 1s a short-actmg donor and cannot produce the sustained release of NO that is needed for these mcubatlon experiments
Protocol 2: Influence of 20-HETE on the Vasodilatory Response to SNP in Isolated, Perfused Renal Arterioles
The contrlbutlon of 20-HETE to the vasodllatory re- sponse to an NO donor was determined by comparison of the concentratton-response relation to SNP on vascular dl- ameter before and after blocking of the endogenous pro- duction of 20-HETE with DDMS (25 pmol/L) These re- sults are summarized m Ag 3 The control mner diameter of the vessels was 98210 pm (n=5 vessels, 5 rats). Phen- ylephrme (I pmol/L) reduced the diameter of these vessels by =50% to 4523 pm SNP (lO-7 to 10m3 mol/L) m- creased the diameter of these vessels m a concentratlon- dependent manner to a maxlmum of 82*4% of control. After DDMS, the vasodllatory response to SNP was greatly attenuated Vessel diameters increased to only 1723% of control m response to the highest concentration of SNP. To rule out the posslblhty that the vasodllatory response to SNP in the presence of DDMS was due to tachyphylaxls or to a time-dependent fall m the respon- siveness of the preparation, time control expenments were performed In five vessels, concentration-response curves for SNP under control condltlons and 30 minutes after ad- mmlstratlon of vehicle to the bath were not significantly different (data not shown).
Because NO donors inhibit the formation of both 20- HETE and DlHETEs, we also examined whether It 1s the fall m 20-HETE levels that contributes to the vasodllator response to NO donors In these experiments, a fixed high concentration of 20-HETE was added to the bath to pre- vent the fall m vascular 20-HETE levels produced by SNP Under control conditions, SNP (10m7 to lo-? mol/L, n=6
vessels, 5 rats) increased the inner diameter of these ves- sels by 64+-4% of control After fixing of vascular 20- HETE levels, the diameter of these vessels increased by only 24?4% of control These results suggest that a fall m 20-HETE levels contributes significantly to the vaso- dilator response to NO donors
To determine whether DDMS has any nonspecific m- hlbltory effects, the effects of this drug on the renal va- sodilatory responses to/the NO-independent dilators aden- osine and dlbutyryl-cGMP were also exammed Under control conditions, adenosme ( lO-7 to 5~ 10v4 mol/L, n=3 vessels, 3 rats) increased the inner diameter of vessels pre- constricted with phenylephrme by 682 10% of control. Af- ter blockade of the synthesis of 20-HETE with DDMS, adenosme increased vascular diameter by 7929% of con- trol. Similarly, dlbutyryl-cGMP (10m7 to 10m4 mol/L, n=3 vessels, 3 rats) increased the diameter of vessels precon- strlcted with phenylephrme by 6926% of control and by 612 10% of control after the synthesis of 20-HETE was blocked with DDMS
Protocol 3: Role of 20-HETE in the Renal and Peripheral Vascular Responses to NO Donors
These experiments exammed the effects of systemic ad- mmistratlon of DDMS on the renal and peripheral vascular responses to NO donors m the rat m VIVO. Control MAP averaged 115t5 mm Hg, RBF was 6.721 mL/mm per gram of kidney weight, and RVR averaged 17 6? 1 3 mL-’ mn-’ g-’ mm Hg-’ (n=7 rats). Under these con- dltlons, Intravenous infusion of MAHMA NONOate at doses of 1, 3,5, and 10 nmol/mm reduced MAP to 99?3, 85-+3, 75&6, and 6826 mm Hg, respectively (Fig 4, top), and lowered RVR to 15+3%, 26?2%, 30+3%, and 3424% of control values, respectively (Fig 4, bottom) After admmistratlon of DDMS (10 mg/kg), baseline MAP fell to 10326 mm Hg and RVR was lowered to 15.3+1 5 mL-’ <mu-’ g-’ mm Hg- I After DDMS pretreatment,
t Vehlde n-7 +DDMS n=7
Mean so Arterial Pressure 80 (% control)
70
60 -1
11.--, -4 !
Resistance b-------4
* (% control) @3 -
70 - ‘k
‘+-----I 60
50 I 0 2 4 6 8 10 12
MAHMA Nonoate Dose (ntnollmm)
FIG 4 Cumulative concentration-response curves for the In VIVO
effects of MAHMA NONOate on the MAP and renal vascular resistance of Inactln-anesthetized rats Animals were studied un- der control conditions and after intravenous administration of DDMS 10 mg/kg over a 1 -hour period Results are expressed as percent of control Values are mean+SEM obtained from 7 rats *Slgnlcant difference from vehicle values
by guest on January 19, 2014http://hyper.ahajournals.org/Downloaded from

324 Hypertension Vol29, No 1, Part 2 January 1997
MAHMA NONOate at doses of 1, 3, 5, and 10 nmol/mm lowered MAP to 10026, 9727, 9427, and 9256 mm Hg, respectively, and RVR fell by only 1 + l%, 6+2%, 7?4%, and 1023% of control, respectively
Protocol 4: Influence of 20-HETE on the Renal and Peripheral Vascular Responses to L-NArg
The effects of DDMS on the MAP and RVR responses to a maximal pressor dose of L-NArg (10 mg/kg IV) were also examined in vehicle (n=7 rats) and DDMS-treated rats (10 mg/kg, n=7 rats). In the vehicle-treated rats, L-NArg increased MAP from 105?2 to 137?4 mm Hg and lowered RBF from 6 1 ?O 7 to 3.220 3 mL/mm per gram of kidney weight (5625% of control). In contrast, m the DDMS-treated animals, MAP increased only from 104&4 to 122&6 mm Hg after admimstratton of L-NArg Similarly, RBF m the DDMS-treated rats fell only from 6.320 7 to 5.0tO 4 mL/mm per gram of kidney weight after L-NArg admmrstration.
Discussion A large number of studies suggest that tonic release of
NO plays an important modulatory role m the regulatron of both renal and peripheral vascular tone It is generally believed that the vasodilatory effects of NO are mediated by cGMP secondary to the strmulatron of guanylyl cyclase m vascular smooth muscle (VSM) cells and the production of cGMP. However, an increasing number of studies have indicated that NO donors or endothelmm-dependent vasodtlators can produce vasodrlation and/or hyper- polarization of VSM in many vascular beds in the pres- ence of guanylyl cyclase inhibitors.4.6 The mechanism of these cGMP-independent effects of NO remains to be determined
In the present study we explored the possibihty that NO may mhibit the formation of 20-HETE and that this may contribute to the vasodilatory effects of NO This hypoth- esis is based on the recent observations that NO inhibits NO synthase and P450 enzymes of the 1A and 2B19 and 3ClO families and from studies mdicatmg that renal VSM cells metabolize AA through a P4504A-dependent path- way to 20-HETEl3J4 and that this system plays a central role m the regulation of renal vascular tone. 15-17 The pres- ent finding that two different NO donors (SNP and PAPA NONOate) blocked the formation of 20-HETE m a con- centration-dependent manner m rat renal cortical micro- somes provides direct evidence to support this possibthty
To study the role that mhibrtton of the endogenous production of 20-HETE plays m the vasodilatory re- sponse to NO, we had to find a way to selecttvely block the system both m vrvo and m vitro In previous studies we have reported that miconazole 1s a highly selectrve mhtbitor of renal epoxygenase activity but has little effect on the formation of 20-HETE.20 17-ODYA, on the other hand, mhrbtts the formatron of both EETs and 20-HETE m the kidney in vtvo when administered directly into the renal mterstitmm; however, rt is extensively protein bound and does not inhibit renal P450 activity when given intravascularly 2O
In the present study we evaluated the potential useful- ness of a new compound (DDMS) that was synthesized by Dr Falck to be a selective, mechanum-based mhtbttor of P450 enzymes of the 4A family that catalyze w-hydroxy- lation of medium and long-chain fatty acids DDMS is a lauric acid analogue, which is the preferred substrate for
the P4504A enzymes, with a dibromo modification on the terminal carbon designed to bmd to the active site of the enzyme. In addmon, the molecule has a methyl sulfimide modification on the Cl carbon to enhance its solubillty m aqueous solutions and to increase the half-life by blocking P-oxidation of the molecule The effects of DDMS on the metabolism of AA m the kidney were evaluated by mcu- bating renal cortical microsomes with 14C-AA m the presence of NADPH We demonstrated that DDMS at a concentration of 10 pmol/L selecttvely mhibited the w-hydroxylation of AA to 20-HETE by rat renal corttcal mtcrosomes, but it had only a slight effect on the formation of DiHETEs At higher concentrations (100 kmol/L), how- ever, DDMS, like 17-ODYA, did inhibit the formation of EETs and DiHETEs by renal cortical mtcrosomes (data not shown) Thus, it appears that DDMS is a first-gener- ation, selective inhibitor of 20-HETE production, with an = lo-fold selecttvtty to Inhibit renal w-hydroxylation over epoxygenase activity.
In the present study, admmtstration of DDMS at a con- centration stmtlar to that which inhibited the formation of 20-HETE by renal mrcrosomes in vitro greatly attenuated the vasodilatory response to SNP m renal arterioles pre- constricted with phenylephrme We have also observed similar effects using 17-ODYA (data not shown) These observations support the hypothesis that mhibmon of the formation of 20-HETE m renal arterioles contrtbutes to the vasodilatory response to NO, however, the mechanism of this mteractron remains to be established.
In previous studies we have reported that 20-HETE is an endogenously formed constrictor of preglomerular re- nal and cerebral artenesl3.14 that normally mhibits the opening of the large-conductance Ca2+-activated K+ chan- nels m VSM 21.22 Inactivation of this channel results m membrane depolarrzatron and promotes vasoconstnction by mcreasmg calcmm influx through voltage-sensitive Ca2+ channels When NO bmds to P4504A enzymes in VSM, it inhibits the endogenous production of 20-HETE. The subsequent fall m 20-HETE levels leads to activation of the large-conductance Ca2’-activated K+ channels and membrane hyperpolarization. Hyperpolarization of renal VSM cells would be expected to decrease calcmm influx through voltage-sensitive Ca2+ channels and promote va- sodilatron. When the endogenous synthesis of 20-HETE is blocked with DDMS, or mtracellular levels of 20-HETE are fixed by exogenous addition of the compound, NO cannot lower 20-HETE levels and promote vasodrlation by hyperpolanzmg the membrane through activation of Ca2+-activated Kf channels. Thus, the response to NO do- nors 1s attenuated by 70% The residual vasodilatory re- sponse to NO donors m vessels treated with DDMS or 20-HETE may represent the cGMP-dependent component of the vasodilatory response to NO It 1s important to note that the vasodilatory response to NO-independent dilators adenosme and dibutyryl-cGMP m renal vessels was not significantly altered by DDMS. These results suggest that the mhibitory effects of DDMS on renal vasodilatory re- sponses are specific to NO-dependent dilators
Experiments were also performed to determine the pos- sible role of 20-HETE m the renal and peripheral vasodi- latory effects of NO donors and synthase mhtbitors m rats m vtvo In these experiments, the dose of DDMS was cho- sen on the basis of prehmmary experiments mdtcatmg that mfusron of DDMS (10 mg/kg) over a l-hour period (either into the renal artery or mtravenously) produced the same
by guest on January 19, 2014http://hyper.ahajournals.org/Downloaded from

Alonso-Galma et al 20-HETE, NO, and Renal Vasculature 325
degree of renal vasodllatlon and blockade of the autoreg- ulatlon of RBF as a dose of 17-ODYA,*O which reduces renal 20-HETE production by 70%. In addition, Brand- Schleber et al*3 have recently reported that a similar dose of DDMS effectively reduces the urinary excretion of 20- HETE, an index of renal 20-HETE production We found that DDMS inhibited the renal vasodilatory response to graded infusions of an NO donor by =80% It also mhib- lted the rise in renal vascular resistance produced by a maximal vasoconstrlctor dose of an inhibitor of NO syn- thase by ~50% These findings are consistent with the results of the m vitro studies on isolated renal arterioles and together suggest that inhibition of the production of 20-HETE may contribute to the vasodllatory effects of NO m the renal mlcrocirculation
Ii was also surpnsmg that systemic admmlstratlon of DDMS greatly attenuated the fall m blood pressure pro- duced by intravenous infusion of MAHMA NONOate and the rise m systemic blood pressure produced by L-NArg. These observations suggest that NO may tonically inhibit the formation of 20-HETE m the rat m vivo in vascular beds other than the kidney and the brain, in which we have established a prominent role for this substance m the reg- ulation of vascular tone.13.14J Clearly, more work is needed to map the vascular beds that produce 20-HETE and the relative contribution of 20-HETE and NO to the regulation of basal vascular tone.
In summary, the present results suggest that NO inhibits renal cytochrome P4504A enzymes and that inhibition of the endogenous production of 20-HETE contributes to the effects of NO on renal and peripheral vascular tone.
Acknowledgments This work was supported by grants from the Natlonal Institutes
of Health HL-29587, HL-36279, and GM 31278 The authors wish to thank Lisa Henderson for excellent technical assistance with the P450 assays
References 1 Navar LG, Inscho EW, MaJ’d DSA, Irnig JD, Harrison-Bernard LM,
Mltchell KM Paracrme regulation of the renal microcIrculation Physrol Rev 1996,76 425-536
2 Loscalzo J, Welch G Nitric oxide and Its role m the carchovascular system Prog Cardtovasc Dzs 1995,38 87-104
3 Bachman S, Mundel P Nltr~c oxide m the kidney synthesis, local- ization and funcuon Am J Kidney DLS 1994,24 112-129
4 Cohen RA The tole of mtnc oxide and other endothehal denved vasoactive substances m vascular &ease Prog Cardrovasc Dls 1995,38 105-128
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
Cohen RA, Vanhoutte PM Endothehnm-dependent hyperpolanza- bon beyond mtrlc oxide and cychc GMP Circulanon 1995.92 3337-3349 Bolontma VM, NaJ’bl S, Palacmo JJ, Pagan0 PJ, Cohen RA Nitric oxide directly activates calcmm-dependent potassium channels m vascular smooth muscle Nature 1994,368 850-853 Hurshman AR, Marletta MA, Nitric oxide complexes of inducible mtnc oxide synthase spectral characterization and effects on cata- lytic actlvlty Blochemzstry 1995,34 5627-5634 Grlscavage JM, Habbs AJ, Ignarro LJ Negative modulation of mtrlc oxide synthase by mtnc oxide and mtroso compounds Adv Phar- macol 1995,34 215-234 Wmk DA, Osawa Y, Darbyshme JF, Jones CR, Eshenaus SC, Nims RW InhIbition of cytochrome P450 by mtrlc oxide and a mtric oxide. releasmg agent Arch Blochem Blophys 1993,300 115-123 Khatsenko OG, Gross SS, Ritlcmd AB, Vane JR Nitric oxide IS a mediator of the decrease m cytochrome P450-dependent metabohsm caused by lmmunostimulants Proc Nat1 Acad Scr USA 1993, 90 11147-11151 Duthu GS, Schertze HG Effect of nitrate on rabbit liver mixed func- honal oxidase activity. Drug Metab Dlspos 1979.7 263-269 Schertze HG, Duthu GS Nitrate binding to rabbit liver mlcrosomes and effects of ammopynclme demethy.&on Btochem Pharmacol 1979,28.873-879 Harder DR, Gebremedhm D, Narayanant J, Jefcoat C, Falck JR, Campbell WB, Roman RJ Formation and a&on of a P4504A me- tabohte of arachidomc acid m cat cerebral mlcrovessels Am J Phys- zol 1994,266 H1840-H1845 Irmg JD, Zou AP, Stec DE, Harder DR. Falck JR, Roman RJ FormatIon and a&on of 20-HETE on rat renal arterioles Am J Phys- 101 1996,270 R217-R227 Inug JD, Zou AP, Ortiz de Montellano PR, SUI Z, Roman RJ Cy- tochrome P450 mhibitors alter afferent artenolar resoonses to elevations m pressure Am J Physlol 1994,266 H1879-Hi885 Zou AP, Irnig J, Kaldunslu M, Ortlz de Montellano PR, SUI 2, Roman RJ Inhibition of renal vascular production of 20-hydroxyeicosatetra- encnc acid impairs autoregulation of renal blood flow Am J Physlol 1994,266 F275-F282 Zou AP, Inug J, Ortiz de Montellano PR, Sm Z, Falck JR, Roman RJ Endogenous P450 metabohtes of arachidomc acid modulate tu- buloelomerular feedback Am J Phwol 1994.267 F934-F941 Ma ?H, Schwartzman ML, Roman RJ Renai metabolism of arach- idomc acid m Dahl rats Am J Physrol 1994,267 R579-R589 Ma YH, Gebremedhm D, Schwartzman ML, Clark JE, Harder DR, Masters BSS, Roman RJ ZO-HETE is an endogenous vasoconstnctor of doe. renal arcuate arteries CUC Res 1993.72 126-136 Zou AP, Ma YH, Orhz de Montellano PR,.Clark JE, Masters BS, Roman RJ Effects of 17-ODYA, a smade-substrate inhibItor of P450 fatty acid w-hydroxylation on renal function m rats .I Phar- macol Exp Ther 1994,268 474-481 Zou AP, Fleming JT, Falck JR, Jacobs ER, Gebremedhm D, Harder DR. Roman RJ 20-hvdroxvelcosatetraenmc acid 1s an endoeenous mhlbltor of the large conductance Ca++-activated K+-channel m renal arterioles. Am J Physrol 1996,270 R228-R237 Harder DR. Camobell !$B. Roman RJ Role of cvtochrome P450 enzymes m’the co&o1 of vascular tone J Vast Res” 1995,32 79-92 Brand-Sclueber E, Taluzawa H, Falck JR, Lamado-Schwartzman M, Nasjletti A Increased dietary salt attenuates a renal vasoconstnctor mechanism mediated by 20-HETE Hypertensron 1996,28 542 Abstract
by guest on January 19, 2014http://hyper.ahajournals.org/Downloaded from




















