
Induction of cytolytic function in resting peripheral blood CD8+/ Leu-7+ T cells through IL2/p 75...
12
CELLULAR IMMUNOLOGY 130,29 l-302 (1990) Induction of Cytolytic Function in Resting Peripheral Blood CD8+/ Leu-7+ T Cells through IL2/p 75 IL2-Receptor Interaction: A Study in the Allogeneic Human Bone Marrow Transplantation Model’ LUCILA MADARIAGA,* ELISABETHLEROY,* NATHALIE MOIR&* MATTHIEU ROULEAU,* ZOHAIR MISHAL,~ HENRI ROCHANT,$ JEAN-PAUL VERNANT,$ BERNARD CHARPENTIER,* MOHAMED BEN ARIBIA,* AND ANNA SENIK* *Laboratoire d’lmmunologie Cellulaire et de Transplantation et tService de Cytofluoromhrie, I.R.S.C., B. P. 8, 94801 VillejuiJ; France: and $Service d’H&natologie, Hipital Henri Mondor, Crheil, France Received January 23, 1990; accepted May 22, 1990 CD8+/Leu-7+ T cells which circulate in increased proportions in the blood of long-term sur- viving BMT patients are for the most part high-density resting lymphocytes lacking ILZR-LU (~55) expression. We show that they can be induced by IL2 to manifest cytolytic function after 24-48 hr stimulation by using rather high concentrations of IL2 (at least 50 U/ml). This function was much more readily induced in high-density CD8+/Leu-7+ T cells than in high-density CD8+/Leu-7- T cells and occurred in the presence of minimal cell proliferation. Other cytokines involved in primary CTL differentiation (IFN-y, IL4 and IL6) were without effect suggesting that CD8+/Leu-7+ T cells are, in the BMT model, in vivo preactivated CTL ready to differentiate into cytolytic effecters under the sole IL2 stimulus. TU27 Mab directed at IL2R-8 (~75) subunit almost completely prevented ILZ-induced cytolytic function of CD8+ T cells while 33B3.1 Mab directed at IL2R-a (~55) subunit was ineffective, suggesting that the signal for this function has its origin in ILZR-fl chains constitutively expressed by these cells. o 1990 Academic PKSS, IIIC. INTRODUCTION In normal individuals, CD8+/Leu-7+ (CD57+) T cells constitute approximately 20% of CD8+ T cells and are in vivo primed CTL, essentially responsible for mitogen and anti-CD3-induced cytotoxicity manifested by freshly isolated PBL in a 4-hr cyto- toxicity assay ( 1,2). In this respect, they can be considered competent lytic cells that do not require differentiation to be activated. Attempts to stimulate such cells re- vealed that they proliferate very little in response to mitogenic lectins (3) and to anti- CD3 Mab (2). However, when stimulated with anti-CD3 + PMA2 or PHA + SRBC, they proliferate vigorously suggesting that appropriately stimulated accessory path- ways are necessary for their full activation (2). After allogeneic bone marrow transplantation (BMT), an abnormal expansion of CD8+/Leu-7+ T lymphocytes often persists in the blood of healthy long-term surviv- ’ This work was supported by grants from ARC and from INSERM N” 50.88.38. ’ Abbreviations used: BMT, bone marrow transplantation; IL2R, interleukin 2 receptor; PHA, phyto- haemagglutinin; PMA, phorbol myristate acetate; SRBC, sheep red blood cells. 291 0008-8749190 $3.00 Copyright 0 1990 by Academic Press, Inc. All rights ofreproduction in any form reserved.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Induction of cytolytic function in resting peripheral blood CD8+/ Leu-7+ T cells through IL2/p 75...

CELLULAR IMMUNOLOGY 130,29 l-302 (1990)
Induction of Cytolytic Function in Resting Peripheral Blood CD8+/ Leu-7+ T Cells through IL2/p 75 IL2-Receptor Interaction: A Study
in the Allogeneic Human Bone Marrow Transplantation Model’
LUCILA MADARIAGA,* ELISABETH LEROY,* NATHALIE MOIR&* MATTHIEU ROULEAU,* ZOHAIR MISHAL,~ HENRI ROCHANT,$
JEAN-PAUL VERNANT,$ BERNARD CHARPENTIER,* MOHAMED BEN ARIBIA,* AND ANNA SENIK*
*Laboratoire d’lmmunologie Cellulaire et de Transplantation et tService de Cytofluoromhrie, I.R.S.C., B. P. 8, 94801 VillejuiJ; France: and $Service d’H&natologie, Hipital Henri Mondor, Crheil, France
Received January 23, 1990; accepted May 22, 1990
CD8+/Leu-7+ T cells which circulate in increased proportions in the blood of long-term sur- viving BMT patients are for the most part high-density resting lymphocytes lacking ILZR-LU (~55) expression. We show that they can be induced by IL2 to manifest cytolytic function after 24-48 hr stimulation by using rather high concentrations of IL2 (at least 50 U/ml). This function was much more readily induced in high-density CD8+/Leu-7+ T cells than in high-density CD8+/Leu-7- T cells and occurred in the presence of minimal cell proliferation. Other cytokines involved in primary CTL differentiation (IFN-y, IL4 and IL6) were without effect suggesting that CD8+/Leu-7+ T cells are, in the BMT model, in vivo preactivated CTL ready to differentiate into cytolytic effecters under the sole IL2 stimulus. TU27 Mab directed at IL2R-8 (~75) subunit almost completely prevented ILZ-induced cytolytic function of CD8+ T cells while 33B3.1 Mab directed at IL2R-a (~55) subunit was ineffective, suggesting that the signal for this function has its origin in ILZR-fl chains constitutively expressed by these cells. o 1990 Academic PKSS, IIIC.
INTRODUCTION
In normal individuals, CD8+/Leu-7+ (CD57+) T cells constitute approximately 20% of CD8+ T cells and are in vivo primed CTL, essentially responsible for mitogen and anti-CD3-induced cytotoxicity manifested by freshly isolated PBL in a 4-hr cyto- toxicity assay ( 1,2). In this respect, they can be considered competent lytic cells that do not require differentiation to be activated. Attempts to stimulate such cells re- vealed that they proliferate very little in response to mitogenic lectins (3) and to anti- CD3 Mab (2). However, when stimulated with anti-CD3 + PMA2 or PHA + SRBC, they proliferate vigorously suggesting that appropriately stimulated accessory path- ways are necessary for their full activation (2).
After allogeneic bone marrow transplantation (BMT), an abnormal expansion of CD8+/Leu-7+ T lymphocytes often persists in the blood of healthy long-term surviv-
’ This work was supported by grants from ARC and from INSERM N” 50.88.38. ’ Abbreviations used: BMT, bone marrow transplantation; IL2R, interleukin 2 receptor; PHA, phyto-
haemagglutinin; PMA, phorbol myristate acetate; SRBC, sheep red blood cells.
291
0008-8749190 $3.00 Copyright 0 1990 by Academic Press, Inc. All rights ofreproduction in any form reserved.

292 MADARIAGA ET AL.
ing patients, reaching up to 60% of CD8+ T cells (4). Like their normal counterparts (5) they can exert a potent suppressor function in PWM-induced antibody produc- tion by normal B cells (4). They are in an apparent resting state as judged by their small size and by the lack, on their surface, of activation antigens such as Tat or HLA-DR. In addition, when taken directly from the blood, they mediate lectin-de- pendent lysis only to a small extent unless they have been stimulated for approxi- mately 2 days with PWM in the presence of accessory cells (Leroy et al., in press3). These data suggested that most CD8+/Leu-7+ T cells from long-term surviving BMT patients are primed CTL awaiting further cellular activation.
Given the fact that resting primed CTL can be induced by IL2 alone to express cytolytic activity in the absence of added antigen (6), and that freshly isolated periph- eral blood CD8+ T cells exhibit appreciable lytic activity that can be further enhanced by IL2 (7), we assumed herein that CD8+/Leu-7+ T cells from long-term surviving BMT patients would also be activated by IL2. Such was indeed the case, as revealed in preliminary studies, and we further concentrated (1) on the comparison between CD8+/Leu-7+ and CD8+/Leu-7- T cells and (2) on the molecular form of IL2R in- volved in this process. In this study we used anti-Tat and TU27 Mab which block IL2 binding to, respectively, the IL2R-a (~55) subunit (8) and the IL2R-P (~75) sub- unit (9). It is noteworthy that TU27 Mab reacts in cytofluorometry not only with NK cells but also with peripheral blood CD8+ T cells and TCR-y/G bearing cells ( 10, 11) indicating the constitutive expression of p75 IL2R on these cells. Accordingly TU27 Mab will almost completely inhibit ILZactivation of NK cells ( 12) as well as the rapid induction of cytolytic activity that a brief exposure to IL2 can provoke in peripheral blood CD8+ T cells and TCR--r/G+ bearing cells (10). We confirm and extend these results by showing that patients’ CD8+/Leu-7+ T cells, almost completely devoid of CD56’-contaminating cells, will develop a more potent cytolytic potential than CD8+/Leu-7- T cells when exposed to IL2 and that the cytotoxicity signal is transmit- ted through isolated p75 IL2R in the presence of minimal cell proliferation.
MATERIALS AND METHODS
Patients
Patients came from the Bone Marrow Transplantation Unit of Hopital Henri Mondor (Service d’HCmatologie, Creteil, France), where they had received allogeneic bone marrow transplantation for hematological malignancies. They were selected for this study on the basis of (1) a long-term graft (between 18 to 80 months post- transplantation) and (2) a large expansion on CD8+/Leu-7+ circulating T lympho- cytes. To determine this, the proportions of those cells were routinely estimated by double IF analysis. In the 20 selected patients, the proportions of CD8+ cells and CD8+/Leu-7+ cells among T lymphocytes were, respectively, 47.6 + 11.3% and 27.0 k 8.8% which yielded a proportion of 57% CD8+/Leu-7+ cells within the CD8+ T cell subset (versus N 16% for 7 normal bone marrow donors). All the patients were in good health except two patients who suffered from chronic GVHD.
‘Leroy, E., Madariaga, L., Ben Aribia, M. H., Mishal, Z., Thedorou, I., Rochant, H., Vemant, J. P., Senik, A., Exp. Haematol., in press.

IL2-INDUCED ACTIVATION OF CD8+/Leu-7+ CTL 293
Monoclonal Antibodies
To eliminate undesired cell subsets from PBL by C lysis, we used the following Mab: anti-HNKl (CD57) (subclone G8) was donated by Dr. M. Lipinski, (I.G.R. Villejuif, France); anti-CD4 was purchased from Immunotech (Luminy, Marseille, France); anti-NKH 1 Mab which defines a 220-kD pan-NK cell antigen (CD56) was kindly given by Dr. T. Hercend (I.G.R. Villejuif, France). 33B3.1 Mab4 which recog- nizes the IL2 binding site of the p55 IL2 receptor (CD25), and 18E64 Mab which recognizes another epitope of this receptor were a gift from Dr. D. Olive (Unite IN- SERM 119, Marseille, France). Phycoerythrin (PE)-conjugated anti-NKH 1 was pur- chased from Coulter Clone (Margency, France). FITC-conjugated and PE-conju- gated anti-CD25, anti-CD3, anti-CD8, and anti-CD57 Mab were purchased from Immunotech. For IF analysis, TU27 Mab directed at the P-subunit of IL2R (9) was used on whole PBL at 1: 1000 final dilution, followed by FITC-conjugated F(ab’)2 anti-mouse Ig. Control staining was done with irrelevant IgGl Mab. In double IF analysis, electronic gates were set on CD8+ bright T cells detected by using PE-conju- gated anti-CD8 Mab. For inhibition studies TU27 Mab and anti-Tat Mab (kindly given by Dr. T. Waldmann (NIH, Bethesda, MD) were added to the cells 30 min before IL2. Control ascites were UN2 Mab which are directed against sheep red blood cells (kindly provided by Dr. M. D. Scharff, Albert Einstein College of Medi- cine, NY).
Fractionation of Patients’ Lymphocytes
Mononuclear cells were isolated by Ficoll-Isopaque (d = 1.078) gradient centrifu- gation and were fractionated by rosetting with sheep red blood cells treated with 2- aminoethylisothiouronium (Sigma Chemical). Rosetting (E+) cells were recovered after lysis of the sheep red blood cells in Gey’s solution. Fractionation of E+ lympho- cytes according to their density was routinely performed on discontinuous Percoll gradients (Pharmacia Fine Chemicals, Bois d’Arcy, France) as described by Timonen et al. (13). Four different concentrations of Percoll in culture medium were used (43.5,48, 5 1, and 62%). Fractions were recovered at each interface from the top to the bottom of the gradients and were numbered 1 to 4. The highest density fractions F3 and F4 (approximately 30 and 50%, respectively, of CD8’/Leu-7+ T cells) were used in our assays. To eliminate CD4+ T cells and NKH l+/CD56+ cells by C lysis, 1 X lo7 high-density cells were incubated for one-half hour at room temperature in 1 ml RPM1 1640 medium containing 5% fetal calf serum, a l/20 final dilution of anti- CD4 Mab, and a l/200 final dilution of anti-NKH 1 Mab. After centrifugation, the cells were resuspended in l/6 diluted rabbit complement and incubated for 60 min at 37°C. The cells were then washed and submitted to a second treatment including 10 pg/ml 33B3.1 and l/200 diluted NKH 1 Mab to eliminate cells expressing IL2R- (Y chain and CD56+ antigen, respectively. Dead cells were eliminated by centrifuga- tion over a 38% Percoll density gradient (550g for 10 min). Because after lysis with anti-CD4 and complement, more than 90% of the remaining cells were CD8+ and less than 3% CD4+, they will be referred to as CD8+ cells. Less than 1% of them were Tat+ or NKH l+/CD56+.
4 We thank Dr. K. Sugamura for the kind gift of TU27 Mab, Dr. H. Bazin for the kind gift of 33B3.1 Mab, and Dr. J. Y. Perrot for expert assistance with the flow cytometry.

294 MADARIAGA ET AL.
For immunofluorescence sorting of Leu-7-positive or negative cells, we used a FACS 440 (Becton-Dickinson, Mountain View, CA). Fluorescent cells (Leu-7+) above background were separated under sterile conditions from nonfluorescent cells. Both sorted populations were more than 95% pure as assessed by reanalysis.
In some experiments, CD8+/Leu-7+ T cells were positively selected by panning as described elsewhere ( 14). In brief, high-density CD8 T cells were treated with anti- HNKl Mab and washed. These cells were then incubated for 2 hr at 4°C on petri dishes coated with 100 pg/ml affinity-purified goat anti-mouse IgM antibodies (Jack- son Immunoresearch Laboratories, PA). The CDS+/Leu-7+ T cells bound to the pan- ning plates were detached by pipetting and were finally 90% pure.
Cell Cultures
Lymphocytes (5 X lo4 or 1 X 105) were cultured in triplicate in the wells of 96-well round-bottomed microtiter plates (Nunc) in 0.2-ml RPM1 1640 supplemented with 10% human AB serum and antibiotics. Unless otherwise stated, all the cultures were set up in the presence of4000 rad-irradiated autologous monocytes (4 X 104) obtained from E-negative cells of the patients or from their bone marrow donors. Usually 50 U/ml recombinant IL2 was added to the cells for a 2- or 3-day stimulation period. Cell-mediated cytotoxicity was then measured in a “Cr release assay and cell prolifer- ation was determined by measuring the amount of [3H]thymidine (1 PCi, 25 Ci/ mmol, CEA, Gif sur Yvette, France) incorporated in stimulated cells after a 6-hr pulse.
Target Cells for “Cr Release Assay
PHA-induced lymphoblasts were obtained after a 3-day stimulation with 1 pg/ml purified PHA (HA 16, Wellcome Laboratories, England). These cells, as well as K562 cells, were washed four times, labeled with 100 &i 51Cr (CEA) for 3 hr at 37°C and further washed three times before being used in the 4-hr 5’Cr release assay. A lytic unit was defined by the number of cells mediating 30% lysis.
Recombinant Human Cytokines
r IL2 (10’ U/mg protein, 1 U = 6.5 p&l) was a gift from Dr. D. Lando (Roussel Uclaf, Romainville, France). r IL6 (8.96 X lo* U/mg protein) was a gift from Dr. W. Fiers (Laboratorium voor Moleculaire Biologie, Gent, Belgique). r IL4 was given by Dr. J. Banchereau (Unicet, Dardilly, France).
RESULTS
Comparison between the Cytolytic Activities Induced by IL2 in High-Density CD8+/ Leu- 7’ T Cells and CD8+/Leu- 7- T Cells from BMT Patients
In preliminary studies, it was determined that most (380%) CD8+/Leu-7+ T cells from long-term BMT patients were recovered in the high-density fractions of Percoll gradients with minimal CD56+ cell contamination (i.e., NK cells and MHC-unre- stricted CTL). Further elimination by C lysis of the few residual CD56+ cells (< l- 2%) was an important prerequisite for this study because a substantial portion of CD56’ MHC-unrestricted CTL has been shown to coexpress the CD8 and Leu-7

IL2-INDUCED ACTIVATION OF CD8+/Leu-7* CTL 295
60
.‘“- E 0 40 E P ::
s
20
A I I
.
I
y” l
71
I&
8 I 0
I “0 R I I
- IL2 + IL2 - IL2 + IL2
CDS+/ Led- CDs’/ Leu-7+
80
60
40
20
C
403 2O:l 10:1
Effector:target cell ratio
51
FIG. 1. Comparison between CDS+/Leu-7+ T cells and CD8+/Leu-7- T cells in their ability to express IL2-induced cytotoxicity against PHA-blasts. (A) I X 10’ FACS-purified CD8+/Leu-7+ T cells and CD8+/ Leu-7- T cells from the same BMT patients were cultivated for 3 days with (0) or without (0) 50 U/ml IL2 in the presence of 4 X 10“ autologous irradiated monocytes. 5’Cr-labeled PHA-blasts (3 X 103) were then added to the cultures which yielded an approximate 20: 1 effector:target cell ratio as estimated from the number of alive cells. Percentage of lysis in accessory cells and IL2 was < 1%. (B) 1 X 10’ whole high- density CD8+ T cells from UPN AL 232 (m) and from UPN AU 027 (Cl) were stimulated with IL2 under the same conditions as in A. They were then pooled, washed, and tested for cytotoxicity against PHA- blasts at various effector: target cells ratios. The same cell populations depleted of Leu-7+ cells were (0) for UPN AL 232 and (0) for UPN AU 027.
antigen and to respond directly to IL2 by enhanced cytolytic capacity against K562 cells ( 15). After all the purification steps carried out to isolate CDS+/Leu-7+ T cells and CD8+/Leu-7- T cells, both cell populations did not significantly lyse K562 cells either before or after IL2 treatment, as routinely controlled (not shown). To examine the reactivity of CD8+/Leu-7+ T cells to IL2, as regards cytolytic activity, they were cultivated for 3 days in the presence of irradiated autologous adherent cells and 50 U/ml IL2. PHA-blasts were then used as indicator target cells in lectin-dependent cytotoxicity. In this system, the “lethal hit” step delivered by IL2-activated cells to these targets is triggered by the trace amounts of PHA carried over by lymphoblasts, as verified in control experiments using various target cells preincubated or not with PHA (not shown). Figure 1A illustrates several paired determinations in which FACS-purified CD8+/Leu-7+ and CD8+/Leu-7- T cells from the same individuals were compared for IL2-induced cytolytic activity. It is seen that both subpopulations reacted to IL2 but that higher levels of cytotoxicity were displayed by CD8+/Leu-7+ T cells than by CD8+/Leu-7- T cells. This difference was highly significant as assessed by the Wilcoxon test for paired data (P < 0.03).
In other experiments ILZstimulated whole high-density CD8+ T cells were assayed for cytolytic function. They were compared with CDV T cells treated with anti-Leu-7

296 MADARIAGA ET AL.
0
10 50 100 250 500
IL2 U/ml
FIG. 2. Dose-response and kinetics of IL2 stimulation. (A) IL2 concentrations were applied for 4 hr n , 10 hr n , 24 hr q and 48 hr 0 (including the cytotoxicity test) on high-density CDS+ T cells from UPN 45 under the same conditions as in Fig. 1 B. The experiment was performed twice with similar results. Bars are +SD.
Mab + C. Figure 1 B illustrates two typical experiments showing that the cytotoxicity exerted by Leu-7-depleted CD8+ T cells was reduced by about 70% when compared to that of whole CD8+ T ceils. The number of lytic units obtained with the cells of five different BMT patients was calculated: 7 18 + 268 (SE) lytic units/lo7 cells (from 333 to 2 105) were found in whole IL2-stimulated CD8+ T cells versus 155 + 124 lytic units/lo7 cells (from < 10 to 300) in Leu-7-depleted cells. Thus, the large proportion of patient’s CD8+ T cells that coexpress the Leu-7 antigen (~56%) are responsible for the major part of IL2-induced cytolytic function.
Study of Dose-Response and Kinetics of ILZ-Induced Cytolytic Function High-density CD8+ T cells from BMT patients were stimulated with increasing
concentrations of IL2 and the resulting cytolytic function was examined at different times. As depicted in Fig. 2, no significant lytic activity was detected up to 10 hr following IL2 application at doses ranging from 10 to 500 U/ml IL2 (i.e., from 65 pM to 3.25 nM); it was however, highly significant after 24 hr and maximum after 48 hr. Although lo/ml IL2 were able to induce some cytolytic function in the cells, maximum lytic activity was reached at loo-250 U/ml IL2 (650 PM- 1.6 WV), a con- centration which is far above what is necessary to saturate high-affinity receptors. The delay required to activate the cytolytic function of patients’ CD8+/Leu-7+ T cells suggested that they had to differentiate from their resting precursors. In this respect, they differ from CD8+/Leu-7+ T cells of normal individuals which, according to Phil- lips et al. are fully differentiated CTL exerting the bulk of LDCC displayed by periph- eral blood T cells without previous stimulation (1) and which, as stated below, can be further activated by a 4-hr exposure to IL2.
IL2-Induced Cytolytic Function Occurs in the Presence of Limited Cell Proliferation of CD8’/Leu- 7+ T Cells Induction by exogeneous IL2 of cytolytic function in the sole context of accessory
cells was achieved with very limited proliferation of patients’ CDS+/Leu-7+ T cells.

ILZ-INDUCED ACTIVATION OF CD8+/Leu-7+ CTL 297
t
CD8+ ILeu-7+
15
10
5
L--l
CDS+ ILeu-7-
anti-CD3 - - + +
IL2 - + _ -
FIG. 3. Proliferative response of CD8+/Leu-7+ T cells and CD8+/Leu-7- T cells stimulated with anti- CD3 in the presence or absence ofIL2.5 X 1 O4 FACS-purified CD8+ cell subsets of UPN 74 were stimulated for 3 days with anti-CD3 (5 &ml) in the presence or absence of 50 U/ml IL2.40% autologous 4000 rad- irradiated monocytes were used as accessory cells. cpm in the latter, tested in the presence of IL2 or Mab, were negligible. Values are cpm f SD. This experiment is representative of four performed.
Thus, after a 3-day exposure to 50 U/ml IL2,5 X lo4 CDV/Leu-7+ T cells incorpo- rated 1356 f 1024 cpm versus 279 f 133 cpm in nonstimulated controls (n = 10). [3H]thymidine incorporation in IL2-stimulated CD8+/Leu-7- T cells was also mar- ginal, although slightly higher (2464 f 2 162 cpm, y1= 10). Interestingly the combina- tion of anti-CD3 and IL2 induced a substantial proliferative response in CDV/Leu- 7+ T cells (Fig. 3). Note that anti-CD3 was by itself ineffective in inducing the prolifer- ation of CD8+/Leu-7+ T cells in conditions which were perfectly suitable for CD8+/ Leu-7- T cells, as previously shown (2). These results suggest that IL2 can deliver to the cytolytic precursors contained in the CD8+/Leu-7+ T cell population a differenti- ation signal but that another cosignal, here derived from the CD3 structure, is re- quired to promote cell division.
Ability of Cytokines Other than IL2 to Induce the Cytolytic Activity ofpatients’ CD8’/ Leu- 7+ T Cells
The role of IL2 in the primary (16) or secondary (6) proliferation and differentia- tion of primed CTL from resting precursors is well established. Other cytokines such

298 MADARIAGA ET AL.
TABLE 1
Respective Capabilities of IL2, IL4, and IL6 to Recall the Cytolytic Potential of Patients CD8+/Leu-7+ T Lymphocytes
Stimulation Induced cytotoxicity
(% specific lysis)
Experiment I: UPN 245 None IL2 (50 U/ml) IL4 (100 U/ml)
Experiment II: UPN 22 1 None IL2 (100 U/ml) IL4 ( 150 U/ml) IL2 (100 U/ml) and IL4 (150 U/ml)
Experiment III: UPN 74 None IL2 (50 U/ml) IL6 (2000 U/ml) IL2 (50 U/ml) and IL6 (2000 U/ml) IFN-7 (1000 U/ml) IL2 (50 U/ml) and IFN-7 (1000 U/ml)
6.2 f 3.6 41.8 f 5.2
7.9 f 2.8
IO.9 f 3.2 79.7 + 6.7 17.1 f 2.4 66.0 f 1.5
15.3 f 1.8 39.1 f 2.8 15.9 f 3.3 33.6 f 3.7 15.4+0.5 34.0 + 1.5
Note. High-density CD8+/Leu-7+ T lymphocytes positively selected by panning were stimulated for 48 hr in the presence of the indicated cytokines. PHA-blasts (3 X 104) were directly added to the cultures at the end of the stimulation period. Values are means ?Z SD of triplicate determinations.
as EN--, ( 17), IL4 ( 18), and IL6 ( 19) have recently been shown to act as potent helper factors for inducing cytolytic activity in in vitro primed CTL. This prompted us to test these cytokines in our system using CD8+/Leu-7+ T cells positively selected by panning to obtain sufficient cells. In these experiments, the cells were cultivated for 48 hr in the absence of adherent cells to avoid the influence of monocyte products. Table 1 shows that IL2 was the only factor capable of inducing cytolytic function in patients’ CD8+/Leu-7+ T cells. The combination of IL2 with either IL4, IL6, or IF’N- y did not result in any form of synergy.
Expression of ~7.5 IL2R on Patients’ CD8’ T Cells and Efect of Anti-IL2R Mab on IL2-Induced Cytolytic Activity
Double immunofluorescence analysis of patients’ PBL T cells revealed that CD8+ T cells were Tat-negative and that the most intense fluorescence staining with TU27 (5-15% of the cells) was located on CD56+ and dimly stained CD8+ cell populations (not shown). Little but significant TU27 staining was nevertheless observed on the CD8+ T cell population (CD8+ bright) as previously shown for the cells of normal individuals (10, 11). Thus, in the PBL of two selected patients only 1 and 2% TU27+ cells were, respectively, detected within the CD8+ (bright) T cell population. The contribution of p75 IL2R, detected by TU27 Mab, in IL2-induced cytolytic function, is illustrated in Fig. 4A. The presence of TU27 Mab during the 48-hr IL2 stimulation (50 U/ml) profoundly inhibited the induction of cytolytic activity in CD8+ T cells,
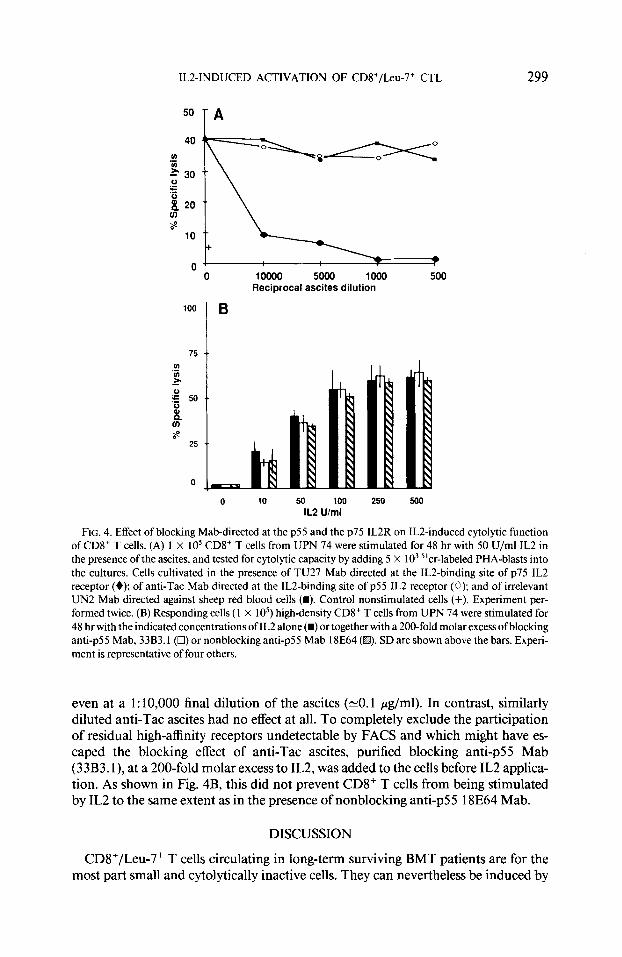
ILZ-INDUCED ACTIVATION OF CD8+/Leu-7+ CTL 299
0 10000 5000 1000 500 Reciprocal ascites dilution
100 ] B
0 10 50 100 250 so0
IL2 U/ml
FIG. 4. Effect of blocking Mab-directed at the ~55 and the ~75 IL2R on ILZ-induced cytolytic function of CD8+ T cells. (A) 1 X IO5 CD8+ T cells from UPN 74 were stimulated for 48 hr with 50 U/ml IL2 in the presence of the ascites, and tested for cytolytic capacity by adding 5 X IO3 “cr-labeled PHA-blasts into the cultures. Cells cultivated in the presence of TU27 Mab directed at the ILZ-binding site of ~75 IL2 receptor (+); of anti-Tat Mab directed at the lL2-binding site of ~55 IL2 receptor (0); and of irrelevant UN2 Mab directed against sheep red blood cells (m). Control nonstimulated cells (+). Experiment per- formed twice. (B) Responding cells (1 X 105) high-density CD8+ T cells from UPN 74 were stimulated for 48 hr with the indicated concentrations of IL2 alone(m) or together with a 200-fold molar excess ofblocking anti-p55 Mab, 33B3.1(0) or nonblocking anti-p55 Mab 18E64 (I@. SD are shown above the bars. Experi- ment is representative of four others.
even at a 1: 10,000 final dilution of the ascites (-0.1 pg/ml). In contrast, similarly diluted anti-Tat ascites had no effect at all. To completely exclude the participation of residual high-affinity receptors undetectable by FACS and which might have es- caped the blocking effect of anti-Tat ascites, purified blocking anti-p55 Mab (33B3. l), at a 200-fold molar excess to IL2, was added to the cells before IL2 applica- tion. As shown in Fig. 4B, this did not prevent CD8+ T cells from being stimulated by IL2 to the same extent as in the presence of nonblocking anti-p55 18E64 Mab.
DISCUSSION
CD8+/Leu-7+ T cells circulating in long-term surviving BMT patients are for the most part small and cytolytically inactive cells. They can nevertheless be induced by

300 MADARIAGA ET AL.
IL2 to manifest strong cytolytic activity in LDCC assays. The full cytolytic status of these cells is acquired after 24-hr incubation with relatively high concentrations of IL2 (650 pA4 to 1.6 nM) in the context of very limited cell proliferation. It is well established that IL2 alone is a sufficient factor to reactivate in vitro-generated memory CTL (6) and to enhance the basal cytolytic activity of in vivo primed, competent CTL (7). In contrast, other differentiation factors including IFN-r ( 17), IL4 ( 18), and IL6 ( 19) are involved together with IL2, in the primary differentiation of CTL precursors into functional effecters. The fact that none of the latter three cytokines could replace IL2 or synergize with it may reflect the peculiar differentiation state of CD8+/Leu- 7+ CTL circulating in BMT patients. It seems likely that the CD8+/Leu-7+ T cell population contains in vivo preactivated CTL (pre-CTL) apparently poised in a rest- ing state and awaiting further activation. The delayed kinetics of IL2-induced cyto- toxicity of patients’ CD8+/Leu-7+ T cells, when compared to that of CD8+/Leu-7+ T cells isolated from normal individuals would support this interpretation. In fact we observed that the cytolytic function of normal CD8+/Leu-7+ T cells, isolated from the low-density fractions of Percoll gradients, could be strongly enhanced by IL2 when it was present during the 4-hr cytotoxicity assay (not shown).
Patients’ CD8+/Leu-7+ T cells were found to display more IL2-induced cytolytic function than CD8+/Leu-7- T cells. This is in keeping with the observation that CD8+/Leu-7+ T cells are responsible for the major part of lectin-induced cytotoxicity and of anti-CD3-induced cytotoxicity spontaneously mediated by freshly isolated normal peripheral blood lymphocytes (1,2). However, substantial cytolytic function could also be induced in high-density CD8+/Leu-7- T cells of some BMT patients after 48-hr incubation with IL2, as well as in low-density CD8+/Leu-7- T cells from normal individuals after overnight stimulation with IL2 (not shown). The Leu-7 anti- gens is not a specific marker of cells exhibiting cytolytic function, and the function of Leu-7 molecules is unknown. Therefore the reason why Leu-7 is preferentially expressed on circulating CTL or pre-CTL is still an open question. Interestingly, a considerable overlapping (280%) exists between CD8+ T cells carrying the Leu-7 antigen and those carrying the CD 11 b antigen in normal individuals (5) and in BMT patients as well (20). CD8+/CD 11 b+ T cells in the latter are also strong inhibitors of PWM-induced antibody production by B cells (2 1). CD1 1 b is the (Y subunit of the glycoprotein complex CD 11 b/CD 18 (Mac- 1) which plays a central role in polymor- phonuclear leukocyte adhesion (22) and which belongs to the superfamily of leuko- cyte differentiation molecules designated integrins (23). One can conceive that the Mac- 1 receptor might take part in the cellular interactions involved in the physiologi- cal activation and cytolytic function of CD8+/Leu-7+ T cells, and hence contribute to the advantage of CD8+/Leu-7+ T cells over CD8+/Leu-7- T cells (most of the latter being CD 11 b-negative).
The molecular basis for the ability of CD8+ T cells to directly respond to IL2 was investigated by using Mab specifically directed at the IL2 binding site of either ~55 IL2R (anti-Tat and 33B3.1) or ~75 IL2R (TU27 Mab). While the former were with- out effect, the latter profoundly inhibited the induction of cytotoxicity in patients’ CD8+ T cells exposed to IL2, as has been shown for normal peripheral blood CD8+ T cells and for TCR-y/G cells (lo), suggesting the predominant participation of iso- lated ~75 IL2R constitutively expressed in these cells. The reactivity pattern of TU27 Mab with the patients’ PBL was similar to that obtained with normal PBL (10, 1 I), the main fluorescence intensity being localized on NK (CD56+) cells, and the CD8+

ILZ-INDUCED ACTIVATION OF CD8+/Leu-7+ CTL 301
T cells being faintly stained. While -60% of CD8+ (bright) T cells coexpressed the Leu-7 antigen, only < 1 to 2% of them were scored as positive for TU27. This suggests that only a minority of CD8+/Leu-7+ T cells constitutively express ~75 IL2R, at least at a level sufficient to be detected by flow cytometry analysis. It is, however, not excluded that CD8+ T cells expressing TU27 antigen at a lower level (undetectable by FACS),, might also be activated and contribute to the cytotoxicity. On the other hand, the fact that CD8+/Leu-7+ T cells reacted more readily to IL2 stimulation than CD8+/Leu-7- T cells would indicate that the former express more TU27 molecules than the latter. Experiments are in progress to examine this issue by using three-color immunofluorescence analysis.
Although the constitutive ~75 IL2R detected by TU27 Mab on NK cells are re- sponsible for mediating the signals for cell cytotoxicity and for cell proliferation as well (12, 24), similar receptors expressed on CD8+/Leu-7+ T cells appear primarily to transmit a signal for enhanced cytolytic function. Thus, despite the presence of monocytes which allow a small proportion of purified resting CD4+ and CD8+ T cells to enter cell cycle when exposed to IL2, (25) CD8+/Leu-7+ T cells proliferated to a very limited extent in response to IL2. However, introducing soluble anti-CD3 Mab (which by itself was ineffective) together with IL2 at the initiation of the cultures, resulted in significant cell proliferation, suggesting that a CD3-derived cosignal is able to synergyze with IL2 to drive these cells into cell cycle. We are currently investigating whether the ~75 IL2R constitutively expressed by CD8+/Leu-7+ T cells are in fact initially responsible for cell proliferation. It is noteworthy that CD8+/Leu-7+ T cells displayed a very low proliferative response to anti-CD3 Mab in conditions which were perfectly suitable for the proliferation of CD8+/Leu-7- T cells, i.e., in the pres- ence of monocytes. This suggests that triggering of the CD3/TCR complex with spe- cific Mab is not sufficient to activate an autocrine proliferation pathway in CD8+/ Leu-7+ T cells. By using PHA and SRBC stimuli, Ruthlein et al. have indicated that the CD2 receptor is, in contrast, a major pathway for cell proliferation and for cyto- lytic function of these cells (2). Our present results further suggest that exogeneous IL2 may play an important role in the biology of these cells, being able not only to directly trigger their cytolytic function but also to supplement incomplete CD3-de- rived signal for cell proliferation.
The in vivo role and the antigenic specificity of BMT patient’s CD8+/Leu-7+ T cells are largely unknown. The suppressor function they exert on in vitro PWM-induced antibody synthesis by normal B cells is not at all correlated with in vivo immunodefi- ciency, even when they circulate at very increased cell numbers (4). We have recently suggested (Leroy et al., in press) that part at least of the suppression observed in the in vitro assays that make use of lectins might be due to lectin-induced cytotoxicity mediated by activated CD8+/Leu-7+ cytotoxic T cells against surrounding cells. It is possible that both cytotoxic and suppressor cells may in fact coexist within the CD8+/ Leu-7+ T cells population; the balance between these two kinds of cells depending on the clinical situation. It is noteworthy that in patients suffering from acquired im- mune deficiency syndrome (AIDS), increased proportions of circulating CD8+/Leu- 7+ T cells are also found (2627). Alveolar CD8+/Leu-7+ T cells of such patients have recently been shown to exert potent suppression on CTL effector cells, through a noncytolytic mechanism (28). The cytolytic potential of these cells was, however, not tested in LDCC assays. It would be of great interest to know whether the suppression function of these cells can also be enhanced by IL2. It would also be interesting to

302 MADARIAGA ET AL.
know whether CDV/Leu-7+ T cells which are increased in other clinical situations including kidney (29) and heart (30) transplantations, as well as cytomegalovirus in- fection (3 1 ), are in essence CTL, and whether they display the same exquisite suscep- tibility to IL2 activation as CD8+/Leu-7+ T cells of BMT patients.
REFERENCES
1. Philipps, J. H., and Lanier, L. L., J. Immunol. 136, 1579, 1986. 2. Riithlein, J., James, S. P., and Strober, W., J. Immunol. 141,3791, 1988. 3. Abo, T., and Balch, C. M., J. Immunol. 129, 1758, 1982. 4. Leroy, E., Calve, C. F., Divine, M., Gourdin, M. F., Baujean, F., Aribia, M. H., Mishal, Z., Vemant,
J. P., Farcet, J. P., and Senik, A., J. Immunol. 137,2180, 1986. 5. Clement, L. T., Grossi, C. E., and Gartland, L. G., J. Immunol. 133,246 1, 1984. 6. Lefrancois, L., Klein, J. R., Paetkau, V., and Bevan, M. J., J. Immunol. 132, 1845, 1984. 7. Perez, P., Hoffman, R. W., Titus, J. A., and Segal, D. M., J. Exp. Med. 163, 166, 1986. 8. Leonard, W. J., Depper, J. M., Uchiyama, T., Smith, K. A., Waldmann, T. A., and Greene, W. C.,
Nature (London) 300,267, 1982. 9. Takeshita, T., Goto, Y., Taka, K., Nagata, K., Asao, H., and Sugamura, K., J. Exp. Med. 169, 1323.
10. Yagita, H., Nakata, M., Azuma, A., Nitta, T., Takeshita, T., Sugamura, K., and Okumura, K., J. Exp. Med. 170,1445, 1989.
Il. Ohashi, Y., Takeshita, T., Nagata, K., and Sugamura, K., J. Immunol., 143,3548, 1989. 12. Phillips, J. H., Takeshita, T., Sugamura, K., and Lanier, L. L., J. Exp. Med. 170,29 I, 1989. 13. Timonen, T., Reynolds, C. W., Ortaldo, J. B., and Herberman, R. B., J. Immunol. Methods. 51,269,
1982. 14. Wysocki, L. J., and Sato, V. L., Proc. Nat/ Acad. Sci. USA. 75,2844, 1978. 15. Lanier, L. L., Mele, A., Cwirla, S., Federspiel, N., and Phillips, J. H., Fed. Proc. 45,2823, 1986. 16. Erard, F., Corthesy, P., Nabholtz, M., Lowenthal, J. W., Zaech, P., Plaetink, G., and MacDonald,
H. R., J. Immunol. 134,1644, 1985. 17. Maraskovsky, E., Chen, W. F., and Shortman, K., J. Immunol. 143, 12 10, 1989. 18. Widmer, M. B., and Brabstein, K. H., Nature (London) 326,795, 1987. 19. Okada, M., Kitahara, M., Kishimoto, S., Matsuda, T., Hirano, T., and Kishimoto, T., J. Immunol.
141,1543, 1988. 20. Velardi, A., Terenzi, A., Cucciaioni, S., Millo, R., Grossi, C. E., Grignani, F., and Martelli, M. F.,
Blood71, 1196, 1988. 2 1. Klingemann, H. G., Lum, L. G., and Storb, R., Transplantation 44,38 1, 1987. 22. Sanchez-Madrid, F., Nagy, J. A., Robins, E., Simon, P., and Springer, T. A., J. Exp. Med. 158, 1785,
1983. 23. Hynes, R. O., Cell48,549, 1987. 24. Siegel, J. P., Sharon, M., Smith, P. L., and Leonard, W. H., Science 238,75, 1987. 25. Ben Aribia, M. H., Leroy, E., Lantz, O., Metivier, D., Autran, B., Charpentier, B., Hercend, T., and
Senik, A., J. Immunol. 139,443, 1987. 26. Gupta, S., J. C/in. Immunol. 6,502, 1986. 27. Plaeger-Marshall, S. C., Spina, A., Giorgi, J. V., Mitsuyasu, R., Wolfe, P., Gottlieb, M., and Beall, B.,
J. Clin. Immunol. 7, 16, 1987. 28. Joly, P., Guillon, J. M., Mayaud, C., Plata, F., Theodorou, I., Denis, M., Debre, P., and Autran, B., J.
Immunol. 143,2 193, 1989. 29. Fregona, I. R., Guttman, D., and Jean, R., Transplantation 35,25, 1985. 30. Maher, P., O’Toole, C. M., Wreghitt, T. G., Spiegelhalter, D. J., and English, T. A., Clin. Exp. Immu-
nol. 62,515, 1985. 31. Wursch, A. M., Gratama, J. W., Middledorp, J. M., Nissen, C., Gratwohl, A., Speck, B., Jansen, J.,
d’Amoro, J., The, T. H., and Degas& G. C., Clin. Exp. Immunol. 62,278, 1985.




















