
Comparative study of the cytolytic activity of myotoxic phospholipases A 2 on mouse endothelial...
14
Comparative study of the cytolytic activity of myotoxic phospholipases A 2 on mouse endothelial (tEnd) and skeletal muscle (C2C12) cells in vitro Bruno Lomonte a, *, Yamileth Angulo a, b , Stefano Rufini c , Wonhwa Cho d , Jose´ Roberto Giglio e , Motonori Ohno f , Jose´ Julia´n Daniele g , Patricia Geoghegan h , Jose´ Maria Gutie´ rrez a a Instituto Clodomiro Picado, Facultad de Microbiologı´a, San Jose ´, Costa Rica b Departamento de Bioquı´mica, Escuela de Medicina, Universidad de Costa Rica, San Jose´, Costa Rica c Dipartimento di Biologı´a, Universita ´ degli Studi di Roma, Roma, Italy d Department of Chemistry, University of Illinois at Chicago, Chicago, IL, U.S.A. e Departamento de Bioquı´mica, Faculdade de Medicina de Ribeira ˜o Preto, Universidade de Sa ˜o Paulo, Sa ˜o Paulo, Brazil f Department of Chemistry, Faculty of Science, Kyushu University, Kyushu, Japan g Departamento de Quı´mica Biolo ´gica, Facultad de Ciencias Quı´micas, Universidad Nacional de Co ´rdoba, Co ´rdoba, Argentina h Departamento Contralor, Instituto Nacional de Microbiologı´a Carlos Malbra ´n, Buenos Aires, Argentina Received 4 February 1998; accepted 15 May 1998 Abstract A rapid in vitro cytolytic eect of some myotoxic phospholipases A 2 (PLA 2 s) isolated from the venoms of Viperidae snakes has been previously described. This study was undertaken to investigate if cytolytic activity is a common property of the myotoxic proteins from this group. Murine endothelial cells (tEnd) and skeletal muscle myotubes (C2C12) were utilized as targets. The release of lactic dehydrogenase was quantified as a measure of cell damage, 3 h after exposure of cells to the dierent PLA 2 s, including representatives from the genera Bothrops, Agkistrodon, Trimeresurus, Crotalus (family Viperidae), and Notechis (family Elapidae). All of the group II myotoxic PLA 2 s tested Toxicon 37 (1999) 145–158 0041-0101/98/$ - see front matter # 1998 Elsevier Science Ltd. All rights reserved. PII: S0041-0101(98)00171-8 PERGAMON * Author to whom correspondence should be addressed.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Comparative study of the cytolytic activity of myotoxic phospholipases A 2 on mouse endothelial...

Comparative study of the cytolytic activity ofmyotoxic phospholipases A2 on mouse
endothelial (tEnd) and skeletal muscle (C2C12)cells in vitro
Bruno Lomonte a, *, Yamileth Anguloa, b, Stefano Ru®ni c,Wonhwa Chod, Jose Roberto Giglio e, Motonori Ohno f,
Jose Julia n Daniele g, Patricia Geogheganh,Jose Maria Gutie rrez a
aInstituto Clodomiro Picado, Facultad de MicrobiologõÂa, San JoseÂ, Costa RicabDepartamento de BioquõÂmica, Escuela de Medicina, Universidad de Costa Rica, San JoseÂ, Costa Rica
cDipartimento di BiologõÂa, Universita degli Studi di Roma, Roma, ItalydDepartment of Chemistry, University of Illinois at Chicago, Chicago, IL, U.S.A.
eDepartamento de BioquõÂmica, Faculdade de Medicina de RibeiraÄo Preto, Universidade de SaÄo Paulo, SaÄo
Paulo, BrazilfDepartment of Chemistry, Faculty of Science, Kyushu University, Kyushu, Japan
gDepartamento de QuõÂmica BioloÂgica, Facultad de Ciencias QuõÂmicas, Universidad Nacional de CoÂrdoba,
CoÂrdoba, ArgentinahDepartamento Contralor, Instituto Nacional de MicrobiologõÂa Carlos MalbraÂn, Buenos Aires, Argentina
Received 4 February 1998; accepted 15 May 1998
Abstract
A rapid in vitro cytolytic e�ect of some myotoxic phospholipases A2 (PLA2s) isolatedfrom the venoms of Viperidae snakes has been previously described. This study wasundertaken to investigate if cytolytic activity is a common property of the myotoxicproteins from this group. Murine endothelial cells (tEnd) and skeletal muscle myotubes
(C2C12) were utilized as targets. The release of lactic dehydrogenase was quanti®ed as ameasure of cell damage, 3 h after exposure of cells to the di�erent PLA2s, includingrepresentatives from the genera Bothrops, Agkistrodon, Trimeresurus, Crotalus (family
Viperidae), and Notechis (family Elapidae). All of the group II myotoxic PLA2s tested
Toxicon 37 (1999) 145±158
0041-0101/98/$ - see front matter # 1998 Elsevier Science Ltd. All rights reserved.
PII: S0041 -0101 (98)00171 -8
PERGAMON
* Author to whom correspondence should be addressed.

displayed rapid cytolytic activity when tested in the micromolar range of concentrations (8±32 mM). In contrast, the group I myotoxic PLA2 notexin was devoid of this activity.
Aspartate-49 and lysine-49 PLA2 group II variants showed a comparable cytolytic e�ect.Skeletal muscle myotubes, obtained after fusion and di�erentiation of C2C12 myoblasts,were signi®cantly more susceptible to the cytolytic action of myotoxins than endothelial
cells, previously reported to be more susceptible than undi�erentiated myoblasts under thesame assay conditions. Cytolytic activity appears to be a common characteristic of group IImyotoxic PLA2s of the Viperidae. Bee venom PLA2, a group III enzyme of known
myotoxicity, also displayed cytotoxic activity on C2C12 myotubes, being devoid of activityon endothelial cells. These results suggest that in vitro di�erentiated skeletal musclemyotubes may represent a suitable model target for the study of myotoxic PLA2s of the
structural group II found in snake venoms. # 1998 Elsevier Science Ltd. All rightsreserved.
1. Introduction
Extracellular phospholipases A2 (EC 3.1.1.4; PLA2s) constitute a family ofproteins with a common enzymatic activity and architecture, but with a diversespectrum of pharmacological e�ects. The PLA2s of snake venoms have acquired aconsiderable heterogeneity of toxic activities during evolution, includingneurotoxic, myotoxic, cytotoxic, anticoagulant, and in¯ammatogenic e�ects(Rosenberg, 1990; Ogawa et al., 1995). These actions are not always related to theprotein's PLA2 activity, and dissociations between catalytic and toxic activities ofvenom PLA2s have been frequently documented. For example, several Lys-49PLA2 myotoxin isoforms (Liu et al., 1990; Francis et al., 1991; Cintra et al., 1993)found in crotalid venoms lack, or have extremely low catalytic activity, and yetdisplay myotoxicity comparable or even higher than that of their enzymatically-active Asp-49 PLA2 counterparts (Homsi-Brandeburgo et al., 1988; Lomonte andGutie rrez, 1989; Kihara et al., 1992). Thus, catalytic activity of PLA2s may becentral to some, but not all of the toxic e�ects induced by this family of proteins.
Myotoxic PLA2s are among the main factors responsible for the drastic necrosisof skeletal muscle observed in victims of envenoming by snakes (Mebs andOwnby, 1990; Harris, 1991; Gutie rrez and Lomonte, 1997). Current evidenceindicates that these myotoxins bind to the plasma membrane of muscle ®bers(Brenes et al., 1987; Dixon and Harris, 1996) and alter their permeability(Gutie rrez et al., 1984a,b; Bultro n et al., 1993a; Incerpi et al., 1995; Ru®ni et al.,1996), although a speci®c receptor has not been identi®ed yet, and the mode bywhich their rapid toxic e�ect is exerted is largely unknown.
As an approach to the characterization of the mechanisms involved inmyotoxicity, cultured cells have been evaluated as targets for myotoxic PLA2s.Secreted forms of PLA2s have been classi®ed into three groups, on the basis oftheir primary structures: group I includes mammalian pancreatic and Elapidaesnake venom PLA2s; group II is formed by the mammalian non-pancreatic, aswell as the Viperidae snake venom PLA2s; and ®nally, PLA2s from bee venom
B. Lomonte et al. / Toxicon 37 (1999) 145±158146

and lizards are classi®ed as group III (Dennis, 1994). Some myotoxins of thegroup II PLA2s cause a rapid cytolytic e�ect upon cell lines of skeletal muscleorigin, as well as upon a variety of other cell types in vitro (Incerpi et al., 1995;Krizaj et al., 1991; Bruse s et al., 1993; Bultro n et al., 1993b; Bieber et al., 1994;Lomonte et al., 1994a,b). However, the relationship between myotoxicity in vivoand cytotoxicity in vitro caused by this group of PLA2s has not been fullyexamined, and more work is needed to clarify if these two e�ects aremanifestations of the same toxic mechanism. In the present study, we havescreened and compared the cytolytic activity of several myotoxic PLA2s, in orderto investigate (a) if this activity constitutes a common property among myotoxins,and (b) if it can be correlated to the structural characteristics of these proteins.
Previous studies with myotoxin II from Bothrops asper, which showed a broadtarget speci®city, described that an endothelial cell line (tEnd) was slightly moresusceptible than skeletal muscle myoblasts to cytolysis (Lomonte et al., 1994a,b).Other reports demonstrated that di�erentiated myotubes are highly susceptibletargets for the cytolytic activity of related group II PLA2 myotoxins (Bruse s et al.,1993; Incerpi et al., 1995; Bieber et al., 1994). Therefore, in this work, we haveutilized both the tEnd endothelial cell line, and a skeletal muscle myoblast linewhich di�erentiates into myotubes (C2C12), for the comparative study of thecytotoxic activity of myotoxins.
2. Materials and methods
2.1. Phospholipases A2 and toxins
All the PLA2s and toxins utilized in this study were puri®ed by at least one ofthe authors, with the exception of bee (Apis mellifera) venom PLA2, which waspurchased from Sigma (St. Louis, MI, U.S.A.), notexin from Notechis scutatusscutatus (Karlsson et al., 1972), kindly provided by Dr B. Persson (University ofUppsala, Sweden) and B. jararacussu bothropstoxin II, kindly provided by DrA.C.O. Cintra (Universidade de SaÄ o Paulo, Brazil).
Myotoxins I (Gutie rrez et al., 1984b), II (Lomonte and Gutie rrez, 1989), III(Kaiser et al., 1990), and IV (Dõ az et al., 1994) from the venom of Bothrops asper,as well as B. nummifer myotoxin (Gutie rrez et al., 1986), B. godmani myotoxins Iand II (Dõ az et al., 1992), B. schlegelii myotoxin I (Angulo et al., 1997), and B.neuwiedii myotoxin I (Geoghegan et al., in preparation), were puri®ed by twocycles of ion-exchange chromatography on carboxymethyl-Sephadex C-25,essentially as described by Lomonte and Gutie rrez (1989). B. jararacussubothropstoxin II (originally denoted fraction SIII±SPIV) was puri®ed by acombination of gel ®ltration on Sephadex G-75 and ion-exchange chromatographyon SP-Sephadex C-25 (Homsi-Brandeburgo et al., 1988). Agkistrodon piscivoruspiscivorus basic D49 and K49 PLA2s were puri®ed by gel ®ltrationchromatography using a Sephadex G-50 column followed by ion exchange
B. Lomonte et al. / Toxicon 37 (1999) 145±158 147

chromatography using a SP-Sepharose column (Pharmacia) as described(Maraganore et al., 1984). Trimeresurus ¯avoviridis basic protein I was puri®ed bySephadex G-100 gel ®ltration followed by CM-cellulose and DEAE-cellulosechromatographies (Yoshizumi et al., 1990). Crotalus durissus terri®cus crotamineand crotoxin were puri®ed by gel ®ltration on Sephadex G-75 in 0.05 Mammonium formate at pH 3.5 (Laure, 1975). The crotoxin A (crotapotin) and B(phospholipase A2) subunits, were puri®ed as described by Hendon and Fraenkel-Conrat (1971). The recombinant human class II PLA2 was expressed inbaculovirus-infected Sf9 cells and puri®ed by ion exchange chromatography usinga Mono S column (Pharmacia), as described by Snitko et al. (1997).
2.2. Cell cultures
Two cell types were utilized in this study. tEnd (Bussolino et al., 1991) is amurine endothelial cell line of capillary origin, transformed with polyoma virus.C2C12 (ATCC CRL-1772) is a murine myoblast cell line which can fuse anddi�erentiate into myotubes. Both cell types were routinely grown in Dulbecco'sModi®ed Eagle's Medium (DMEM; Sigma D-5796) supplemented with 10% fetalcalf serum (FCS; Sigma F-2442), 2 mM glutamine, 1 mM pyruvic acid, penicillin(100 U/ml), streptomycin (0.1 mg/ml), and amphotericin B (0.25 mg/ml), in ahumi®ed atmosphere with 7% CO2, at 378C. Cells were harvested fromsubcon¯uent monolayers grown in 25 cm2 bottles after their detachment bytrypsin (1500 U/ml) containing 5.3 mM EDTA, for 5 min at 378C. Theresuspended cells were seeded in 96-well microplates, at an approximate initialdensity of 1±4 � 104 cells/well, in the same medium. After reaching nearcon¯uence, usually in 3±5 days, tEnd endothelial cells were utilized directly in thecytotoxicity assay described below. In the case of C2C12 myoblasts, after reachingnear con¯uence, growth medium was replaced by di�erentiation medium, whichconsisted of DMEM supplemented with 1% FCS (Ebisui et al., 1995). After 4±6additional days of culture, when a vast proportion of long multinucleatedmyotubes was observed, cells were utilized in the cytotoxicity assays, as describedbelow.
2.3. Cytotoxicity assays
Cytotoxic activity was determined as previously described (Lomonte et al.,1994b). In brief, the di�erent toxins/PLA2s were diluted in assay medium, whichconsisted of DMEM supplemented with 1% FCS, and then added to the cellcultures growing in the 96-well plates (after aspirating their medium) in a totalvolume of 150 ml/well. Controls for 0% and 100% toxicity consisted of assaymedium, and 0.1% Triton X-100 in assay medium, respectively. After 3 h ofincubation, an aliquot of 100 ml of supernatant was collected for determination ofthe lactic dehydrogenase (LDH) activity released from damaged cells, using acolorimetric end-point assay (Sigma No. 500). In some experiments performed
B. Lomonte et al. / Toxicon 37 (1999) 145±158148

with myotubes, creatine kinase (CK) activity was determined in cell supernatantsusing a colorimetric end-point assay (Sigma No. 520). In addition, cells wereobserved microscopically at the end of the incubation in order to evaluatemorphological alterations.
3. Results
Initial dose±response screenings of the cytotoxic e�ect caused by di�erentmyotoxin isoforms (I, III, and IV) from B. asper venom on tEnd cells, showedsimilar activities to that of B. asper myotoxin II (data not shown), which had beenpreviously characterized in this assay system (Lomonte et al., 1994a,b). On thisbasis, two toxin concentrations were selected for the comparative analysis of allthe samples, which were assayed simultaneously. Results obtained with thedi�erent myotoxins/PLA2s on tEnd cells, are summarized in Fig. 1. At theconcentration of 20 mg/well (133 mg/ml, approximately 8 mM for the PLA2
Fig. 1. Comparison of the cytotoxic activity of di�erent phospholipases A2 and myotoxins, on
endothelial cells (tEnd) in vitro. (A) 20 mg/well (08 mM for phospholipases A2); (B) 80 mg/well(032 mM). Cytotoxicity was estimated by the release of lactic dehydrogenase to the supernatant after
3 h. Each bar represents mean2SD of two determinations. All samples were assayed simultaneously
on two independent experiments. n.t. = not tested.
B. Lomonte et al. / Toxicon 37 (1999) 145±158 149

myotoxins), the most active proteins were the four myotoxin isoforms from B.
asper (I±IV), together with B. nummifer and B. neuwiedii myotoxins (Fig. 1). At
the concentration of 80 mg/well (533 mg/ml, approximately 32 mM) all of the class
II myotoxic PLA2s from Bothrops, Trimeresurus, and Agkistrodon venoms were
clearly cytolytic (Fig. 1). In the case of crotoxin, its puri®ed subunit B was
cytolytic, while both the subunit A and the heterodimeric complex were devoid
of activity, even at 80 mg/well. The e�ects of notexin, bee venom PLA2, and
crotamine were not signi®cant, even at 80 mg/well, in comparison to negative
controls. The recombinant human class II PLA2, which was tested only at
20 mg/well due to limited availability, was also devoid of cytolytic activity
(Fig. 1). Microscopically, endothelial cells damaged by myotoxic PLA2s
appeared similar to those exposed to detergent, with an apparent release of their
cytoplasmic contents into the medium, but remained attached to their
substratum.
The e�ect of myotoxins was also studied on the C2C12 muscle cells which fused
into long multinucleated myotubes a few days after lowering the FCS
concentration to 1% (Fig. 2). On some occasions, a spontaneous contractile
activity of control myotubes was observed under the microscope. Supernatants
obtained after lysis of myotubes by detergent contained signi®cant amounts of CK
activity. However, instability of CK activity in the supernatants, as well as
interference of Triton X-100 in its determination were observed (data not shown).
Therefore, all subsequent cytotoxicity experiments with myotubes were performed
using the release of LDH as an estimator of cell damage.
Myotubes were ®rst tested in time-course experiments using B. asper myotoxin
II, in order to evaluate the kinetics of the cytolytic action using this target. In
similarity with earlier studies with endothelial cells (Lomonte et al., 1994b),
myotoxin II caused a rapid release of LDH from myotubes, which reached its
peak by 1 h of exposure (Fig. 3). The cytotoxic e�ect of the di�erent proteins on
C2C12 myotubes is summarized in Fig. 4. These cells were more susceptible than
endothelial cells to the action of myotoxins, since high cytolytic activities were
generally observed already at the concentration of 20 mg/well (Fig. 4). The higher
susceptibility of myotubes was also observed in a comparative dose±response
curve performed with B. asper myotoxin II on both cell types (Fig. 5). In
similarity with the results obtained with tEnd endothelial cells, all of the class II
myotoxic PLA2s from Bothrops, Trimeresurus, and Agkistrodon venoms tested,
together with the subunit B of crotoxin, were clearly cytotoxic to C2C12
myotubes (Fig. 4). In addition, a signi®cant cytolytic e�ect of whole crotoxin, and
a strong activity of bee venom PLA2 were observed. On the other hand, notexin,
crotamine, and the subunit A of crotoxin, were devoid of cytolytic activity (Fig. 4).
Microscopically, exposure of C2C12 cultures to cytolytic myotoxins caused a
complete disappearance of myotubes, leaving only myoblasts in the monolayer,
although they also appeared damaged but still adhering to their substratum
(Fig. 2).
B. Lomonte et al. / Toxicon 37 (1999) 145±158150
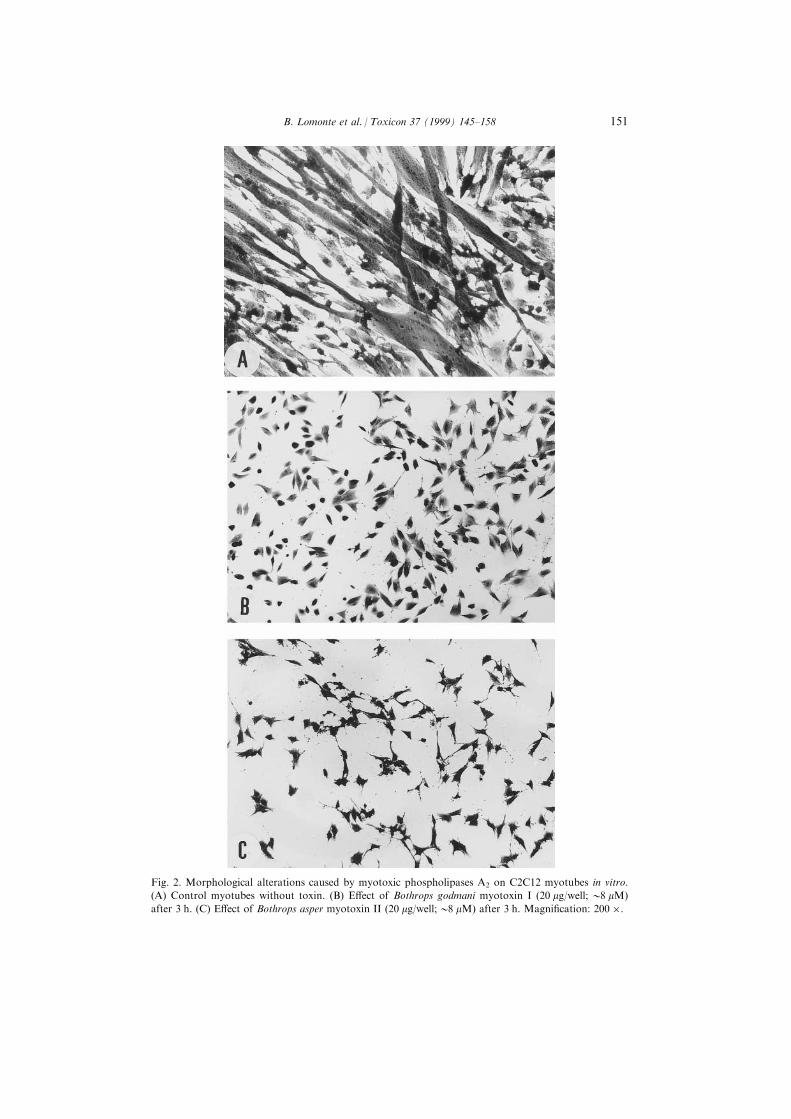
Fig. 2. Morphological alterations caused by myotoxic phospholipases A2 on C2C12 myotubes in vitro.
(A) Control myotubes without toxin. (B) E�ect of Bothrops godmani myotoxin I (20 mg/well; 08 mM)
after 3 h. (C) E�ect of Bothrops asper myotoxin II (20 mg/well;08 mM) after 3 h. Magni®cation: 200 �.
B. Lomonte et al. / Toxicon 37 (1999) 145±158 151
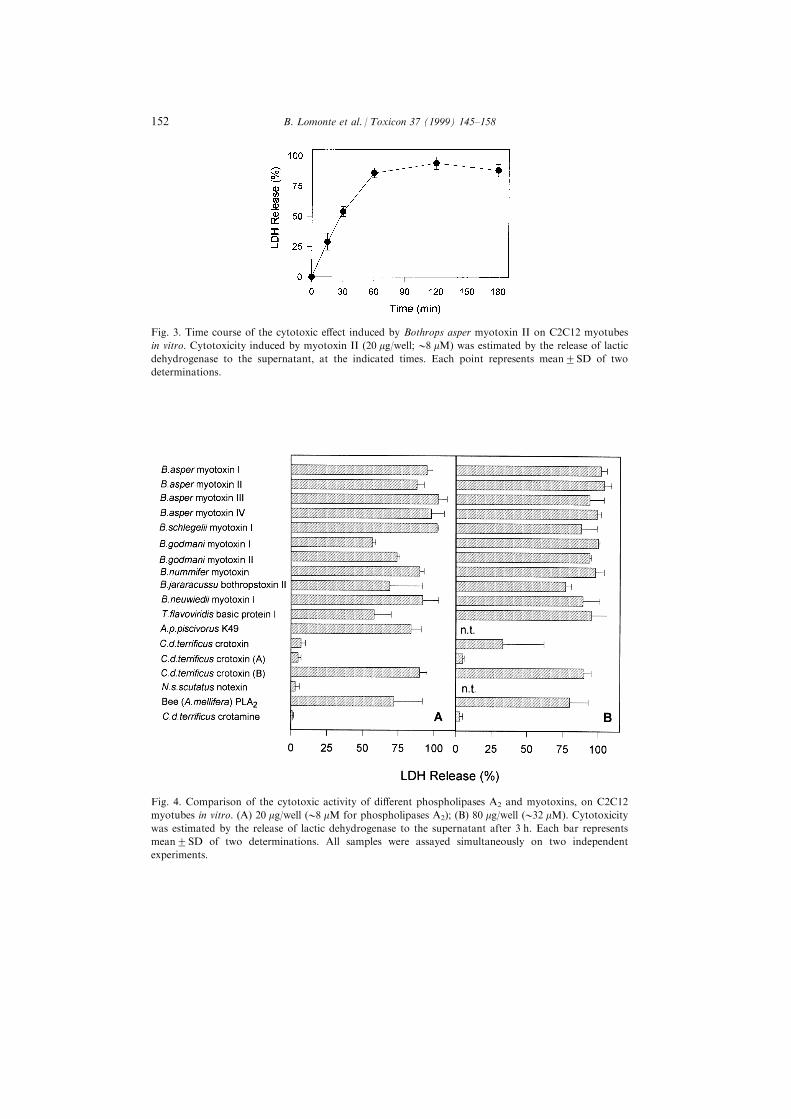
Fig. 3. Time course of the cytotoxic e�ect induced by Bothrops asper myotoxin II on C2C12 myotubes
in vitro. Cytotoxicity induced by myotoxin II (20 mg/well;08 mM) was estimated by the release of lactic
dehydrogenase to the supernatant, at the indicated times. Each point represents mean2SD of two
determinations.
Fig. 4. Comparison of the cytotoxic activity of di�erent phospholipases A2 and myotoxins, on C2C12
myotubes in vitro. (A) 20 mg/well (08 mM for phospholipases A2); (B) 80 mg/well (032 mM). Cytotoxicity
was estimated by the release of lactic dehydrogenase to the supernatant after 3 h. Each bar represents
mean2SD of two determinations. All samples were assayed simultaneously on two independent
experiments.
B. Lomonte et al. / Toxicon 37 (1999) 145±158152

4. Discussion
Initial reports of the rapid in vitro cytolytic e�ect of some myotoxic PLA2sisolated from snake venoms (Krizaj et al., 1991; Bruse s et al., 1993; Bultro n et al.,1993b; Bieber et al., 1994; Lomonte et al., 1994a,b; Incerpi et al., 1995; Fletcher etal., 1996), resembling the fast damage to muscle ®bers occurring in vivo (Gutie rrezet al., 1984a; Lomonte et al., 1994c), opened the possibility to utilize cultured cellsas a model for the study of the myotoxic activity of this group of proteins. Theestablishment of a valid in vitro model for myotoxicity would facilitate thedetailed characterization of molecular events involved in the muscle damagecaused by myotoxins. However, it has been pointed out that in vitro cytotoxicityand in vivo myotoxicity may be two related, but not necessarily identicalphenomena induced by myotoxins, which should be analyzed and interpreted withprudence (Fletcher et al., 1997; Gutie rrez and Lomonte, 1997). To this end, it wasnecessary to investigate if cytolysis is a common e�ect induced by myotoxicPLA2s, since only few isolated examples have been described in the literature.
A cytotoxicity assay previously developed for the study of B. asper myotoxin IIand its neutralization (Lomonte et al., 1994b) was utilized for the simultaneousscreening of all the samples, with tEnd endothelial cells or C2C12 myoblasts/myotubes as targets. Results showed that the latter cells were signi®cantly moresusceptible to the action of myotoxic PLA2s. This was evident in (a) the generalcomparison of the activity of di�erent myotoxins on both cell types, (b) thecomparative dose±response curve performed with B. asper myotoxin II, and (c)the microscopic observation of morphological cell alterations, in which myotubeswere rapidly lysed, disappearing from the monolayers. Thus, our present ®ndingscon®rm the higher susceptibility of di�erentiated myotubes in comparison to othercell types, in agreement with earlier descriptions (Bruse s et al., 1993; Incerpi et al.,1995).
Fig. 5. Comparison of the cytotoxic activity of Bothrops asper myotoxin II on C2C12 myotubes (®lled
circles) or endothelial cells (empty circles) in vitro. Cytotoxicity was estimated by the release of lactic
dehydrogenase to the supernatant after 3 h. Each point represents mean2SD of two determinations.
B. Lomonte et al. / Toxicon 37 (1999) 145±158 153

Under our experimental conditions, all of the fourteen class II myotoxic PLA2stested, including representatives of Bothrops, Trimeresurus, Agkistrodon, andCrotalus, showed a fast cytolytic activity in vitro. This e�ect was more pronouncedon myotubes than on endothelial cells. Thus, results suggest that cytotoxicity is acommon activity among the class II PLA2 myotoxins. Moreover, theseobservations favor the concept of a close relationship between the in vitrocytotoxic and in vivo myotoxic mechanisms exerted by this particular structuralgroup of proteins. The reported cytotoxic activity of ammodytin L (Krizaj et al.,1991; Incerpi et al., 1995; Ru®ni et al., 1996), a group II myotoxic PLA2 fromVipera ammodytes (Krizaj et al., 1991), is also in agreement with this conclusion.
Among the class II myotoxic PLA2s studied, no major di�erences wereobserved in the cytolytic activity induced by either Asp-49 or Lys-49 variants,indicating that this e�ect is independent of the intrinsic in vitro enzymatic activityof these proteins. This result might be expected, considering that both types ofPLA2s exert comparable myotoxicity in vivo (Kihara et al., 1992; DõÂ az et al.,1991). On the other hand, the human class II PLA2, for which no toxic activitieshave been described, was devoid of cytotoxic e�ect upon endothelial cells (up to8 mM), in contrast to the snake venom class II myotoxic PLA2s. This is inagreement with the divergent evolutive processes that these two related types ofproteins have undergone (Davidson and Dennis, 1990; Ogawa et al., 1995), andsuggests that cytotoxic activity probably appeared after the divergence of thecommon ancestor of class II PLA2s of reptiles and mammals.
Results obtained with crotoxin from C. durissus terri®cus showed a clearcytolytic activity of its subunit B (the enzymatically-active component), but not ofits subunit A. This is in agreement with the reported myotoxic activities of thetwo isolated components in vivo (Gopalakrishnakone et al., 1984; Kouyoumdjanet al., 1986). Interestingly, intact crotoxin was not cytolytic to endothelial cells,but showed a signi®cant e�ect on myotubes. While the isolated subunit B was aspotent as other class II myotoxic PLA2s, intact crotoxin was signi®cantly lesscytotoxic, indicating that subunit A decreases the cytotoxic activity of subunit B.This is in contrast to other pharmacological e�ects of crotoxin in vitro, such asblockade of neuromuscular transmission, in which subunit A plays an enhancingrole (Bon, 1997). It has been proposed that subunit A acts as a `chaperone',precluding the binding of subunit B to low a�nity non-speci®c sites, and that thecomplex dissociates upon binding to high a�nity sites at the presynaptic terminal(Bon, 1997). The lower cytolytic e�ect of crotoxin in our in vitro model mayre¯ect a low dissociation of subunits A and B, perhaps due to a relatively lowdensity of high a�nity binding sites on the membrane of di�erentiating myotubes.Our observation might be related to the particular myotoxic behavior of crotoxinin vivo, which, in contrast to the other group II myotoxic PLA2s studied here,causes systemic rather than local myonecrosis (Azevedo-Marques et al., 1985).
In contrast to the group II myotoxic PLA2s, the `small' myotoxin crotaminefrom C. durissus terri®cus was devoid of cytolytic e�ect in our model, inagreement with previous reports studying this protein (Fletcher et al., 1996) andthe closely related myotoxin a (Bruse s et al., 1993). Although classi®ed as a
B. Lomonte et al. / Toxicon 37 (1999) 145±158154

myotoxin, crotamine does not act by disrupting the integrity of muscle cell plasmamembrane. Instead, it induces a sodium in¯ux and contracture (Chang and Tseng,1978), together with vacuolation and calcium release from sarcoplasmic reticulum(Cameron and Tu, 1978; Furukawa et al., 1994). Such pharmacological action isnot expected to cause rapid plasma membrane rupture nor LDH release.
Myotoxic PLA2s from group I (notexin) and III (bee venom PLA2) werestudied in the present model. Notexin was devoid of cytolytic e�ect even ondi�erentiated myotubes, in agreement with earlier studies performed with skeletalmuscle myoblasts and endothelial cells (Harris, 1991; Lomonte et al., 1994a).Therefore, notexin must utilize a di�erent molecular mechanism for the inductionof myonecrosis, compared to the class II myotoxic PLA2s from viperids. Thiscould also be the case of other related class I myotoxic PLA2s from elapids, whichare generally also characterized by a potent neurotoxic activity. Accordingly, amyotoxic PLA2 from Naja naja atra was devoid of cytolytic activity on primarycultures of human skeletal muscle (Fletcher et al., 1996). However, at least oneexample of a class I PLA2 with cytolytic activity has been described in the case ofnigexin from Naja nigricollis (Chwetzo� et al., 1989). Regarding the bee venomPLA2, recently con®rmed to be myotoxic (Ownby et al., 1997), it is interesting tonote that the use of di�erentiated myotubes allowed detection of its cytolyticactivity, which was not evident in our endothelial cell assay, or in previous reportsusing mammary tumor (Martikainen et al., 1993) or melanoma (Daniele et al.,1997) cell lines. Thus, in the case of bee venom PLA2, an agreement betweenmyotoxicity and rapid cytotoxicity on cultured myoblasts was observed.
It is tempting to speculate on the possible reasons for the higher susceptibilityof di�erentiated myotubes, in comparison to other cells types tested in the presentand previous studies (Bruse s et al., 1993; Lomonte et al., 1994a; Incerpi et al.,1995). Cell binding studies performed with crotoxin have shown that this PLA2
may interact with membrane protein receptors or membrane lipids with markedlydi�erent a�nities, the former being much higher (Bon, 1997). If this holds true inthe case of other PLA2s as well, then, at the relatively high concentrationsrequired to rapidly lyse di�erent cells in culture (micromolar range), the class IIPLA2s could be acting through the recognition of relatively ubiquitous membranelipids (or other low a�nity acceptor sites), explaining the observed broad cellspeci®city in their cytolytic action (Lomonte et al., 1994a). On the other hand,fusion and di�erentiation of myoblasts into myotubes may induce the expressionof a high a�nity receptor site for myotoxic PLA2s, enhancing the susceptibility tolower myotoxin concentrations. A proteic membrane PLA2 receptor, named M-type receptor, has been characterized in skeletal muscle cells (Lambeau et al.,1997), although its possible functional role in the action of class II myotoxicPLA2s has not been clearly de®ned. As an alternative view, due to their particularstructural and physiological characteristics, myotubes may have a lower ability tocope with perturbations of membrane homeostasis in comparison to other celltypes, as suggested by Lomonte et al. (1994a), thereby being more susceptible tothe action of myotoxins. Both hypotheses would be compatible with the observedselectivity of myotoxic PLA2s for mature skeletal muscle ®bers in vivo.
B. Lomonte et al. / Toxicon 37 (1999) 145±158 155

Undoubtedly, identi®cation of the cellular receptor(s) for the class II myotoxicPLA2s is needed for a precise understanding of their mode of action.
In conclusion, the present results indicate that, while rapid in vitro cytolysis isnot an activity exerted by all structural types of myotoxins (i.e. the `small' non-enzymatic myotoxins, or some class I myotoxic/neurotoxic PLA2s), it is acommon activity of the structural group II of myotoxic PLA2s from the Viperidaefamily studied so far. In this regard, the use of in vitro di�erentiated skeletalmuscle cells appears as a promising model for further detailed analyses on themyotoxic actions of these proteins.
Acknowledgements
We thank Drs A. C. O. Cintra (University of SaÄ o Paulo, Brazil) and BertilPersson (Uppsala University, Sweden) for kindly providing bothropstoxin-II andnotexin, respectively, as well as Javier Nu nÄ ez, Ricardo BolanÄ os, and CarlosAlberto Vieira for their excellent technical assistance. These studies weresupported by Vicerrectorõ a de Investigacio n, University of Costa Rica (# 741-96-265), International Foundation for Science (F/1388-3F and F/2766-1), Fundac° aÄ ode Amparo a Pesquisa do Estado de SaÄ o Paulo (Proc. 1996/5085-7) and ConselhoNacional de Desenvolvimento Cientõ ®co e Tecnolo gico (Proc. 420009/97-3).
References
Angulo, Y., Chaves, E., Alape, A., Rucavado, A., Gutie rrez, J. M., Lomonte, B., 1997. Isolation and
characterization of a myotoxic phospholipase A2 from the venom of the arboreal snake Bothriechis
(Bothrops) schlegelii from Costa Rica. Archs Biochem. Biophys. 339, 260±267.
Azevedo-Marques, M. M., Cupo, P., Coimbra, T. M., Hering, S. E., Rossi, M. A., Laure, C. J., 1985.
Myonecrosis, myoglobinuria and acute renal failure induced by South American rattlesnake
(Crotalus durissus terri®cus) envenomation in Brazil. Toxicon 23, 631±636.
Bieber, A. L., Ziolkowski, C., d'Avis, P. A., 1994. Rattlesnake toxins alter development of muscle cells in
culture. Ann. N.Y. Acad. Sci. 710, 126±145.
Bon, C. (1997) Multicomponent neurotoxic phospholipases A2. In Venom phospholipase A2 enzymes: struc-
ture, function, and mechanism, ed. R. M. Kini, pp. 267±285. John Wiley and Sons, England.
Brenes, F., Gutie rrez, J. M., Lomonte, B., 1987. Immunohistochemical demonstration of the binding of
Bothrops asper myotoxin to skeletal muscle sarcolemma. Toxicon 25, 574±577.
Bruse s, J. L., Capaso, J., Katz, E., Pilar, G., 1993. Speci®c in vitro biological activity of snake venom myo-
toxins. J. Neurochem. 60, 1030±1042.
Bultro n, E., Gutie rrez, J. M., Thelestam, M., 1993a. E�ects of Bothrops asper (terciopelo) myotoxin III, a
basic phospholipase A2, on liposomes and mouse gastrocnemius muscle. Toxicon 31, 217±222.
Bultro n, E., Thelestam, M., Gutie rrez, J. M., 1993b. E�ects on cultured mammalian cells of myotoxin III,
a phospholipase A2 isolated from Bothrops asper (terciopelo) venom. Biochim. Biophys. Acta 1179, 253±
259.
Bussolino, F., De Rossi, M., Sica, A., Colotta, F., Wang, J. M., Bocchietto, E., Padura, I. M., Bosia, A.,
Dejana, E., Mantovani, A., 1991. Murine endothelioma cell lines transformed by polyoma middle T
oncogene as target for and producers of cytokines. J. Immunol. 147, 2122±2129.
B. Lomonte et al. / Toxicon 37 (1999) 145±158156

Cameron, D. L., Tu, A. T., 1978. Chemical and functional homology of myotoxin a from prairie rattle-
snake venom and crotamine from South American rattlesnake venom. Biochim. Biophys. Acta 532,
147±154.
Chang, C., Tseng, K., 1978. E�ect of crotamine, a toxin of South American rattlesnake venom, on the
sodium channel of murine skeletal muscle. Br. J. Pharmacol. 63, 551±559.
Chwetzo�, S., Tsunasawa, S., Sakiyama, F., Me nez, A., 1989. Nigexine, a phospholipase A2 from cobra
venom with cytotoxic properties not related to esterase activity. J. Biol. Chem. 264, 13289±13297.
Cintra, A. C. O., Marangoni, S., Oliveira, B., Giglio, J. R., 1993. Bothropstoxin-I: amino acid sequence
and function. J. Prot. Chem. 12, 57±64.
Daniele, J. J., Bianco, I. D., Delgado, C., Briones Carrillo, D., Fidelio, G. D., 1997. A new phospholipase
A2 isoform isolated from Bothrops neuwiedii (yarara chica) venom with novel kinetic and chromato-
graphic properties. Toxicon 35, 1205±1215.
Davidson, F., Dennis, E. A., 1990. Evolutionary relationships and implications for the regulation of phos-
pholipase A2 from snake venom to human secreted forms. J. Mol. Evol. 31, 228±238.
Dennis, E. A., 1994. Diversity of group types, regulation and function of phospholipase A2. J. Biol. Chem.
269, 13057±13060.
Dõ az, C., Gutie rrez, J. M., Lomonte, B., Gene , J. A., 1991. The e�ect of myotoxins isolated from Bothrops
snake venoms on multilamellar liposomes: relationship to phospholipase A2, anticoagulant and myo-
toxic activities. Biochim. Biophys. Acta 1070, 455±460.
Dõ az, C., Gutie rrez, J. M., Lomonte, B., 1992. Isolation and characterization of basic myotoxic phospho-
lipases A2 from Bothrops godmani (Godman's pit viper) snake venom. Archs Biochem. Biophys. 298,
135±142.
Dõ az, C., Lomonte, B., Zamudio, F., Gutie rrez, J. M., 1994. Puri®cation and characterization of myotoxin
IV, a phospholipase A2 variant, from Bothrops asper snake venom. Natural Toxins 3, 26±31.
Dixon, R. W., Harris, J. B., 1996. Myotoxic activity of the toxic phospholipase, notexin, from the venom
of the Australian tiger snake. J. Neuropathol. Exp. Neurol. 55, 1230±1237.
Ebisui, C., Tsujinaka, T., Morimoto, T., Kan, K., Ijima, S., Yano, M., Kominami, E., Tanaka, K.,
Monden, M., 1995. Interleukin-6 induces proteolysis by activating intracellular proteases (cathepsins
B and L, proteasome) in C2C12 myotubes. Clin. Sci. 89, 431±439.
Fletcher, J. E., Hubert, M., Wieland, S. J., Gong, Q. H., Jiang, M. S., 1996. Similarities and di�erences in
mechanisms of cardiotoxins, mellitin and other myotoxins. Toxicon 34, 1301±1311.
Fletcher, J. E., Selistre de Araujo, H. S. and Ownby, C. L. (1997) Molecular events in the myotoxic action
of phospholipases. In Venom phospholipase A2 enzymes: structure, function, and mechanism, ed. R. M.
Kini, pp. 455±497. John Wiley and Sons, England.
Francis, B., Gutie rrez, J. M., Lomonte, B., Kaiser, I. I., 1991. Myotoxin II from Bothrops asper (tercio-
pelo) venom is a lysine-49 phospholipase A2. Archs Biochem. Biophys. 284, 352±359.
Furukawa, K. I., Funayama, K., Ohkura, M., Oshima, Y., Tu, A. T., Ohizumi, Y., 1994. Ca2+ release
induced by myotoxin a, a radio-labeled probe having novel Ca2+ release properties in sarcoplasmic reti-
culum. Br. J. Pharmacol. 113, 233±239.
Gopalakrishnakone, P., Dempster, D. W., Hagwood, B. J., Elder, H. Y., 1984. Cellular and mitochondrial
changes induced in the structure of murine skeletal muscle by crotoxin, a neurotoxic phosholipase A2
complex. Toxicon 22, 85±98.
Gutie rrez, J. M. and Lomonte, B. (1997) Phospholipase A2 myotoxins from Bothrops snake venoms. In
Venom phospholipase A2 enzymes: structure, function, and mechanism, ed. R. M. Kini, pp. 321±352.
John Wiley and Sons, England.
Gutie rrez, J. M., Ownby, C. L., Odell, G. V., 1984a. Pathogenesis of myonecrosis induced by crude venom
and a myotoxin of Bothrops asper. Exp. Molec. Pathol. 40, 367±379.
Gutie rrez, J. M., Ownby, C. L., Odell, G. V., 1984b. Isolation of a myotoxin from Bothrops asper venom:
partial characterization and action on skeletal muscle. Toxicon 22, 115±128.
Gutie rrez, J. M., Lomonte, B., Cerdas, L., 1986. Isolation and partial characterization of a myotoxin from
the venom of the snake Bothrops nummifer. Toxicon 24, 885±894.
Harris, J. B. (1991) Phospholipases in snake venoms and their e�ects on nerve and muscle. In Snake
Toxins, ed. A. L. Harvey, pp. 91±129. Pergamon Press, New York.
B. Lomonte et al. / Toxicon 37 (1999) 145±158 157

Hendon, R. A., Fraenkel-Conrat, H., 1971. Biological roles of the two-components of crotoxin. Proc.
Natl. Acad. Sci. U.S.A. 68, 1560±1563.
Homsi-Brandeburgo, M. I., Queiroz, L. S., Santo-Neto, H., Rodrigues-Simioni, L., Giglio, J. R., 1988.
Fractionation of Bothrops jararacussu snake venom: partial chemical characterization and biological ac-
tivity of bothropstoxin. Toxicon 26, 615±627.
Incerpi, S., de Vito, P., Luly, P., Ru®ni, S., 1995. E�ect of ammodytin L from Vipera ammodytes on L-6
cells from rat skeletal muscle. Biochim. Biophys. Acta 1268, 137±142.
Kaiser, I. I., Gutie rrez, J. M., Plummer, D., Aird, S. D., Odell, G. V., 1990. The amino acid sequence of a
myotoxic phospholipase from the venom of Bothrops asper. Archs Biochem. Biophys. 278, 319±325.
Karlsson, E., Eaker, D., Ryde n, L., 1972. Puri®cation of a presynaptic neurotoxin from the venom of the
Australian tiger snake, Notechis scutatus scutatus. Toxicon 10, 405±413.
Kihara, H., Uchikawa, R., Hattori, S., Ohno, M., 1992. Myotoxicity and physiological e�ects of three
Trimeresurus ¯avoviridis phospholipases A2. Biochem. Int. 28, 895±903.
Kouyoumdjan, J. A., Harris, J. B., Johnson, M. A., 1986. Muscle necrosis caused by the sub-units of cro-
toxin. Toxicon 24, 575±583.
Krizaj, I., Bieber, A. L., Ritonja, A., Gubensek, F., 1991. The primary structure of ammodytin L, a myo-
toxic phospholipase A2 homologue from Vipera ammodytes venom. Eur. J. Biochem. 202, 1165±1168.
Lambeau, G., Cupillard, L. and Lazdunski, M. (1997) Membrane receptors for venom phospholipases A2.
In Venom phospholipase A2 enzymes: structure, function, and mechanism, ed. R. M. Kini, pp. 389±412.
John Wiley and Sons, England.
Laure, C. J., 1975. Die primaÈ rstruktur des crotamins. Hoppe-Seyler's Z. Physiol. Chem. 356, 213±215.
Liu, S. Y., Yoshizumi, K., Oda, N., Ohno, M., Tokunaga, F., Iwanaga, S., Kihara, H., 1990. Puri®cation
and amino acid sequence of basic protein II, a Lysine-49 phospholipase A2 with low activity, from
Trimeresurus ¯avoviridis venom. J. Biochem. 107, 400±408.
Lomonte, B., Gutie rrez, J. M., 1989. A new muscle damaging toxin, myotoxin II, from the venom of the
snake Bothrops asper (terciopelo). Toxicon 27, 725±733.
Lomonte, B., Tarkowski, A., Hanson, L. AÊ ., 1994a. Broad cytolytic speci®city of myotoxin II, a lysine-49
phospholipase A2 of Bothrops asper snake venom. Toxicon 32, 1359±1369.
Lomonte, B., Tarkowski, A., Bagge, U., Hanson, L. AÊ ., 1994b. Neutralization of the cytolytic and myo-
toxic activities of phospholipases A2 from Bothrops asper snake venom by glycosaminoglycans of the
heparin/heparan sulfate family. Biochem. Pharmacol. 47, 1509±1518.
Lomonte, B., Lundgren, J., Johansson, B., Bagge, U., 1994c. The dynamics of local tissue damage induced
by Bothrops asper snake venom and myotoxin II on the mouse cremaster muscle: an intravital and elec-
tron microscopic study. Toxicon 32, 41±55.
Maraganore, J. M., Merutka, G., Cho, W., Welches, W., Ke zdy, F. J., Heinrikson, R. L., 1984. A new
class of phospholipases A2 with lysine in place of aspartate 49. J. Biol. Chem. 259, 13839±13843.
Martikainen, P., Nyman, K., Nevalainen, T. J., 1993. Toxic e�ects of human pancreatic and snake and bee
venom phospholipases A2 on MCF-7 cells in culture. Toxicon 31, 835±843.
Mebs, D., Ownby, C. L., 1990. Myotoxic components of snake venoms: their biochemical and biological
activities. Pharmac. Ther. 48, 223±236.
Ogawa, T., Kitajima, M., Nakashima, K., Sakaki, Y., Ohno, M., 1995. Molecular evolution of group II
phospholipases A2. J. Mol. Evol. 41, 867±877.
Ownby, C. L., Powell, J. R., Jiang, M. S., Fletcher, J. E., 1997. Mellitin and phospholipase A2 from bee
(Apis mellifera) venom cause necrosis of murine skeletal muscle in vivo. Toxicon 35, 67±80.
Rosenberg, P. (1990) Phospholipases. In Handbook of Toxinology, eds. W. T. Shier and D. Mebs, pp. 67±
277. Marcel Dekker, New York.
Ru®ni, S., Cesaroni, M. P., Balestro, N., Luly, P., 1996. Proliferative e�ect of ammodytin L from the
venom of Vipera ammodytes on 208F rat ®broblasts in culture. Biochem. J. 320, 467±472.
Snitko, Y., Yoon, E. T., Cho, W., 1997. High speci®city of human secretory class II phospholipase A2 for
phosphatidic acid. Biochem. J. 321, 737±741.
Yoshizumi, K., Liu, S. Y., Miyata, T., Saita, S., Ohno, M., Iwanaga, S., Kihara, H., 1990. Puri®cation and
amino acid sequence of basic protein I, a lysine-49 phospholipase A2 with low activity, from the venom
of Trimeresurus ¯avoviridis (Habu snake). Toxicon 28, 43±54.
B. Lomonte et al. / Toxicon 37 (1999) 145±158158




















