
Cytolytic Effects and Apoptosis Induction of Newcastle Disease Virus Strain AF2240 on Anaplastic...
12
ORIGINAL PAPER Cytolytic Effects and Apoptosis Induction of Newcastle Disease Virus Strain AF2240 on Anaplastic Astrocytoma Brain Tumor Cell Line Rola Ali • Aied M. Alabsi • Abdul Manaf Ali • Aini Ideris • Abdul Rahman Omar • Khatijah Yusoff • Riyadh Saif-Ali Accepted: 3 June 2011 / Published online: 14 June 2011 Ó Springer Science+Business Media, LLC 2011 Abstract Newcastle disease virus (NDV) is a member of genus Avulavirus within the family Paramyxoviridae. Interest of using NDV as an anticancer agent has arisen from its ability to kill tumor cells with limited toxicity to normal cells. In this investigation, the cytotolytic properties of NDV strain AF2240 were evaluated on brain tumor cell line, anaplastic astrocytoma (U-87MG), by using MTT assay. Cytological observations were studied using fluorescence microscopy and transmission electron microscopy to show the apoptogenic features of NDV on U-87MG. DNA laddering in agarose gel electrophoresis and terminal deoxyribonucleotide transferase-mediated dUTP-X nick end-labeling staining assay confirmed that the mode of cell death was by apoptosis. However, analysis of the cellular DNA content by flowcytometery showed that there was a loss of treated U-87MG cells in all cell cycle phases (G1, S and G2/M) accompanied with increasing in sub-G1 region (apoptosis peak). Early apoptosis was observed 6 h post- inoculation by annexin-V flow-cytometry method. It could be concluded that NDV strain AF2240 is a potent antitumor agent that induce apoptosis and its cytotoxicity increasing while increasing of time and virus titer. Keywords NDV strain AF2240 Á Cytotoxicity Á Apoptosis Á Flow-cytometry Á Anaplastic astrocytoma Background Brain tumor is formed by abnormal and uncontrolled cell division; it is abnormal growth of tissue found inside the skull. It can be dangerous because of the importance of the brain and the limited amount of space inside the skull. Most primary brain tumors do not metastasize systemically but spread locally through extensions of infiltrating tumor cells in normal brain. Grade III astrocytoma, anaplastic astrocytoma, is commonly spread to surrounding brain tissue and it is the most common primary brain tumor in adults [42]. Surgery; radiation and chemotherapy are the current treatments for brain tumor [18, 24, 26, 40]. New- castle disease virus (NDV) was first isolated in 1926 in Newcastle, England, in domestic chickens where it caused a severe disease known as Newcastle disease. ND is a disease of poultry that affecting the alimentary and respi- ratory tract as well as the central nervous system [9]. Exposure to humans however, results in mild conjunctivi- tis, laryngitis and influenza-like symptoms [11]. The NDV showed promise in clinical trials as an antineoplastic agent. It is one of the nonengineered oncolytic viruses, which has a long history as a broad-spectrum oncolytic agent that can destroy tumor cells and stimulate the immune system [12, 19]. Many preclinical studies indicate that NDV rep- licates in human cancer cells but not in normal cells, some R. Ali Á K. Yusoff Faculty of Biotechnology and Bimolecular Sciences, Universiti Putra Malaysia, Serdang, Malaysia A. Ideris Á A. R. Omar Faculty of Veterinary Medicine, Universiti Putra Malaysia, Serdang, Malaysia A. M. Alabsi Á A. M. Ali (&) Faculty of Agricultures and Biotechnology, Universiti Sultan Zainal Abidin, Kuala Terengganu, Malaysia e-mail: [email protected] A. M. Alabsi e-mail: [email protected] R. Saif-Ali Faculty of Medicine, Universiti Malaya, Kuala Lumpur, Malaysia 123 Neurochem Res (2011) 36:2051–2062 DOI 10.1007/s11064-011-0529-8
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Cytolytic Effects and Apoptosis Induction of Newcastle Disease Virus Strain AF2240 on Anaplastic...

ORIGINAL PAPER
Cytolytic Effects and Apoptosis Induction of Newcastle DiseaseVirus Strain AF2240 on Anaplastic Astrocytoma Brain TumorCell Line
Rola Ali • Aied M. Alabsi • Abdul Manaf Ali •
Aini Ideris • Abdul Rahman Omar •
Khatijah Yusoff • Riyadh Saif-Ali
Accepted: 3 June 2011 / Published online: 14 June 2011
� Springer Science+Business Media, LLC 2011
Abstract Newcastle disease virus (NDV) is a member of
genus Avulavirus within the family Paramyxoviridae.
Interest of using NDV as an anticancer agent has arisen from
its ability to kill tumor cells with limited toxicity to normal
cells. In this investigation, the cytotolytic properties of NDV
strain AF2240 were evaluated on brain tumor cell line,
anaplastic astrocytoma (U-87MG), by using MTT assay.
Cytological observations were studied using fluorescence
microscopy and transmission electron microscopy to
show the apoptogenic features of NDV on U-87MG. DNA
laddering in agarose gel electrophoresis and terminal
deoxyribonucleotide transferase-mediated dUTP-X nick
end-labeling staining assay confirmed that the mode of cell
death was by apoptosis. However, analysis of the cellular
DNA content by flowcytometery showed that there was a
loss of treated U-87MG cells in all cell cycle phases (G1, S
and G2/M) accompanied with increasing in sub-G1 region
(apoptosis peak). Early apoptosis was observed 6 h post-
inoculation by annexin-V flow-cytometry method. It could
be concluded that NDV strain AF2240 is a potent antitumor
agent that induce apoptosis and its cytotoxicity increasing
while increasing of time and virus titer.
Keywords NDV strain AF2240 � Cytotoxicity �Apoptosis � Flow-cytometry � Anaplastic astrocytoma
Background
Brain tumor is formed by abnormal and uncontrolled cell
division; it is abnormal growth of tissue found inside the
skull. It can be dangerous because of the importance of the
brain and the limited amount of space inside the skull.
Most primary brain tumors do not metastasize systemically
but spread locally through extensions of infiltrating tumor
cells in normal brain. Grade III astrocytoma, anaplastic
astrocytoma, is commonly spread to surrounding brain
tissue and it is the most common primary brain tumor in
adults [42]. Surgery; radiation and chemotherapy are the
current treatments for brain tumor [18, 24, 26, 40]. New-
castle disease virus (NDV) was first isolated in 1926 in
Newcastle, England, in domestic chickens where it caused
a severe disease known as Newcastle disease. ND is a
disease of poultry that affecting the alimentary and respi-
ratory tract as well as the central nervous system [9].
Exposure to humans however, results in mild conjunctivi-
tis, laryngitis and influenza-like symptoms [11]. The NDV
showed promise in clinical trials as an antineoplastic agent.
It is one of the nonengineered oncolytic viruses, which has
a long history as a broad-spectrum oncolytic agent that can
destroy tumor cells and stimulate the immune system
[12, 19]. Many preclinical studies indicate that NDV rep-
licates in human cancer cells but not in normal cells, some
R. Ali � K. Yusoff
Faculty of Biotechnology and Bimolecular Sciences,
Universiti Putra Malaysia, Serdang, Malaysia
A. Ideris � A. R. Omar
Faculty of Veterinary Medicine, Universiti Putra Malaysia,
Serdang, Malaysia
A. M. Alabsi � A. M. Ali (&)
Faculty of Agricultures and Biotechnology, Universiti Sultan
Zainal Abidin, Kuala Terengganu, Malaysia
e-mail: [email protected]
A. M. Alabsi
e-mail: [email protected]
R. Saif-Ali
Faculty of Medicine, Universiti Malaya,
Kuala Lumpur, Malaysia
123
Neurochem Res (2011) 36:2051–2062
DOI 10.1007/s11064-011-0529-8

types of NDV are able to directly kill certain types of
cancer cells and NDV–infected cancer cells can cause the
immune system to respond in different ways [34]. Early
success with the viral vaccine in humans was first reported
in the United States in 1980 [32]. Interest in the use of
NDV as an anticancer agent has arisen from the ability of
the virus to selectively kill human tumor cells with limited
toxicity to normal cells. Many strains of NDV (73-T,
MH68, Italian, Ulester, Rokin, PV701 and HUJ) have been
shown to exhibit an oncolytic activity [12, 36]. In addition,
the oncolytic effects of six Malaysian strains of NDV
(AF2240, 01/C, Ijuk, S, F, and V4) have also been studied
on several tumor cell lines [34, 36]. The goal of most
cancer therapy is to reduce the number of tumor cells and
to prevent their further accumulation (decrease the rate of
cell proliferation). Therefore, in this study the effects of
NDV strain AF2240 on the proliferation and the morpho-
logical changes of brain tumor cell line, anaplastic astro-
cytoma (U-87MG), was tested in vitro.
Materials and Methods
Propagation and Purification of NDV Strain AF2240
NDV was propagated in allantoic fluid of 9–11 days-old
embryonated chicken eggs at 37�C for 48 h. The allantoic
fluid was harvested and the presence of virus was con-
firmed by the haemaglutination test [2]. NDV strains
AF2240 purified as previously described [4, 47].
Cells and Cell Culture
Human brain tumor cell line U-87MG was obtained from
American Type Culture Collection (ATCC, VA, USA).
HCN-2 and 3T3, normal cell lines, kindly provided by Prof.
Dr. Abdul Manaf Ali, Faculty of Biotechnology and
Molecular Biology, University Putra Malaysia (UPM) were
used as control. All cell lines were grown as a monolayer in
25 cm2 tissue culture flasks (NunclonTM, Denmark) at 37�C
in an atmosphere of 5% CO2. MEM medium supplemented
with 10% fetal calf serum, 1% antibiotics and three addi-
tional supplements (2 mM Glutamine, 1 mM Sodium
Pyruvate, and 1% nonessential amino acid) was used for
U-87MG. Normal cell lines were grown in DMEM medium.
MTT Cytotoxicity Assay
U-87MG cells were trypsinized and counted using hemo-
cytometer then were seeded in 96-well micro plate at
3 9 105 cells/ml and then incubated at 37�C in 5% CO2 to
allow cells attachment. The medium was removed and
replaced with fresh medium containing test virus at various
concentrations (double dilution) and were incubated at
37�C, 5% CO2 for 72 h. Each concentration was assayed in
triplicates (n = 3). Seventy-two hours later, 20 ll of MTT
(5 mg/ml) solution was added to each well and then the plate
was further incubated for 4 h. All remaining supernatant
were removed and 150 ll of DMSO was added to dissolve
the formed crystal formazan. MTT assay reading was per-
formed using ELISA plate reader (Biotek EL340, USA).
Quantification of Apoptosis Using Propidium Iodide
and Acridine Orange Double staining
Cells were quantified using propidium iodide (PI) and
acridine-orange (AO) double staining according to standard
procedures and examine under fluorescence microscope
(Lieca attached with Q-Floro Software) [17, 30]. U-87MG
cells were seeded in six-well plate and incubated at 37�C in
5% CO2 atmosphere. Twenty-four hours later, the medium
in each well was removed and replaced with the virus at
IC50 concentration dissolved in medium and incubated at
37�C in 5% CO2 atmosphere for 24, 48, and 72 h. After
incubation period, detached cells in the medium were
collected and added back to trypsinised adherent cells. The
cell suspensions were washed with PBS and then incubated
with 5 ll of acridine orange (10 lg/ml) and 5 ll propidium
iodide (10 lg/ml) at a ratio of 1:1 in 1 ml of cells and
recentrifuged at 1,000 rpm/5 min. After centrifuge, super-
natant was removed leaving 50 ll of remaining supernatant
with pellet. The pellet was resuspended and 10 ll of cell
suspension was pipetted on slide before putting on cover
slip. Within 30 min, the slide was analyzed using fluores-
cent microscope (Leica, Germany). Each experiment was
assayed three times (n = 3). Viable, apoptotic and necrotic
cells was quantified in a population of 200 cells. In apop-
totic bodies, blebbing of plasma membrane and conden-
sation of chromatin were seen. In contrast, necrotic cells
fluoresced red after propidium iodide staining with tiny
fraction of chromatin dispersed around the nucleus. This
assay provides a useful quantitative evaluation and was
done three times (n = 3).
Transmission Unltrastructural Effects of NDV Strains
AF2240 on U-87MG Cells (TEM)
U-87MG brain tumor cells were treated with NDV at IC50
concentration and incubated for 24, 48, and 72 h at 37�C.
The cultured cells were harvested using trypsin and cen-
trifuged for 10 min at 1,500 rpm. The pellets were fixed in
4% (v/v) glutaraldehyde in 0.1 M coccadylate buffer (pH
7.4) for 4 h at 4�C. The fixed cells were centrifuged, and
the pellets were blocked in serum which was later fixed
in glutaraldeyde overnight at 4�C. The specimens were
washed in three changes of sodium coccadylate buffer
2052 Neurochem Res (2011) 36:2051–2062
123

(pH 7.4) for 10 min each, postfixed in 1% osmium tetraoxide
at 4�C. The specimens were then washed in three changes of
sodium coccadylate buffer (pH 7.4) for 10 min each and
dehydrated with a graded series of acetone (35, 50, 75, 95,
and 100%). The cells were then infiltrated with acetone and
resin and embedded with 100% resin in beam capsule, and
left to polymerize at 60�C for 48 h. The area of interest in the
embedded cells resin block was chosen using the toulidine
blue staining and later examined using light microscope. The
selected area was cut in ulltrathin sections using ultrami-
crotome. The sections were placed into a grid and stained
with uranyl acetate for 10 min followed by 50% filtered
acetone, and finally stained using lead which was then
washed twice with distilled water. The stained samples were
then viewed under transmission electron microscopy (Phil-
lips, Eindhoven, The Netherlands).
DNA Fragmentation Assay
Cells at a concentration of 5 9 106 cells/ml were seeded
into six-well plate (NunclonTM, Denmark) in 2 ml culture
medium with a concentration of IC50 value of virus. Some
wells were left with no virus to be used as a control. After
the 72 h of incubation, detached cells in the medium were
collected and added back to trypsinised adherent cells. The
cells were spun down at 1,000 rpm for 10 min. The
supernatant was discarded and the pellet was washed with
PBS twice. The DNA extraction from treated and untreated
cells was carried out according to protocol of a kit for
Blood and Cultured Cells from QIAGEN.
TUNEL Assay
The cells treated with concentration of IC50 value of virus
value were grown on Lab-Tek Chamber slides and the
slides were incubated at 37�C in an atmosphere of 5% CO2.
The slides were washed with PBS after 24, 48 and 72 h and
processed in the apoptosis detection assay. Each experi-
ment was assayed three times (n = 3). The TUNEL Assay
was carried out using a kit for Apoptosis Detection from
Promega, USA.
Analysis of Cellular DNA Content Using Propidium
Iodide
Cells at a concentration of 5 9 106 cells/ml of U-87MG
cell line was seeded into six-well plate in 2 ml culture
medium with a concentration of IC50 value of virus and
were incubated at 37�C in an atmosphere of 5% CO2 for
72 h. Some wells were left with no virus to be used as a
control. After the incubation period, the cultured cells were
harvested using trypsin and centrifuged for 10 min at
1,000 rpm at room temperature. Cell pellets were fixed by
adding 500 ll of 80% cold ethanol and kept for at least 2 h
at -20�C. Cells were pelleted at 1,000 rpm for 10 min and
the ethanol was discarded. The cell pellet was washed with
1 ml (PBS/sodium azide) twice. The pellet was resus-
pended with 1 ml of (PBS ? 0.1% triton X-100 ?10 mm
EDTA ? 50 lg/ml RNase ? 2 lg/ml Propidium iodide)
followed by incubated for � to 1 h at 4�C. Finally, samples
were placed in 12 9 75 Falcon tubes and the cell cycle was
analyzed by flow cytometer (Beckman Coulter, USA).
Each experiment was assayed three times (n = 3).
Flow Cytometry (Annexin V/PI Double Staining)
Cells at a concentration of 5 9 106 cells/ml of U-87MG
cell line was seeded into six-well plate in 2 ml culture
medium with a concentration of IC50 value of virus and
were incubated at 37�C in an atmosphere of 5% CO2 for
72 h. Some wells were left with no virus to be used as a
control. After the incubation period, the cultured cells were
harvested using trypsin and centrifuged for 10 min at
1,000 rpm. The early apoptosis detection for treated and
untreated cells was carried out using Annexin V & Apo
2.7-PE kit (Clontech Laboratories, Inc., USA). Each
experiment was assayed three times (n = 3).
Statistical Analysis
Data was expressed as mean ± SD. Statistical analysis was
performed with Student’s t-test for data from MTT cyto-
toxicity assay, AO/PI staining assay, TUNEL and, flow
cytometry. Differences were considered significant at
P \ 0.001.
Results
Cytotolytic Effects of NDV on Normal Cells and Brain
Tumor Cells
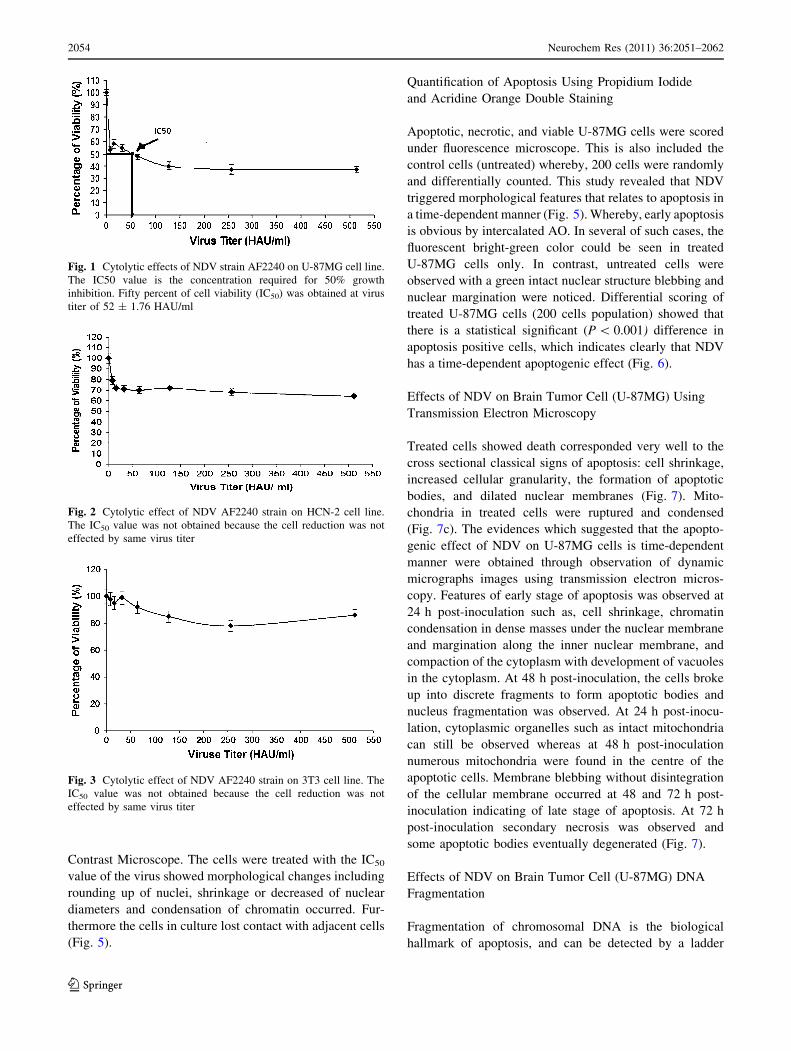
The cytotoxicity of NDV on U87 cells was investigated
using MTT assay after 72 h treatment (Figs. 1, 4). NDV
was clearly found to exert antiproliferative effects toward
U87 brain tumor cells. The IC50 value, which is the con-
centration required for 50% growth inhibition, is deter-
mined to be 52 ± 1.1. Comparatively, NDV showed no
significant cytolytic effect at the same titre used in the
brain cell line toward HCN-2 and 3T3, normal cell lines
(Figs. 2, 3).
Phase Contrast Microscope
The morphological changes of U-87MG cells after treat-
ment with NDV strain AF2240 were observed under Phase
Neurochem Res (2011) 36:2051–2062 2053
123
Contrast Microscope. The cells were treated with the IC50
value of the virus showed morphological changes including
rounding up of nuclei, shrinkage or decreased of nuclear
diameters and condensation of chromatin occurred. Fur-
thermore the cells in culture lost contact with adjacent cells
(Fig. 5).
Quantification of Apoptosis Using Propidium Iodide
and Acridine Orange Double Staining
Apoptotic, necrotic, and viable U-87MG cells were scored
under fluorescence microscope. This is also included the
control cells (untreated) whereby, 200 cells were randomly
and differentially counted. This study revealed that NDV
triggered morphological features that relates to apoptosis in
a time-dependent manner (Fig. 5). Whereby, early apoptosis
is obvious by intercalated AO. In several of such cases, the
fluorescent bright-green color could be seen in treated
U-87MG cells only. In contrast, untreated cells were
observed with a green intact nuclear structure blebbing and
nuclear margination were noticed. Differential scoring of
treated U-87MG cells (200 cells population) showed that
there is a statistical significant (P \ 0.001) difference in
apoptosis positive cells, which indicates clearly that NDV
has a time-dependent apoptogenic effect (Fig. 6).
Effects of NDV on Brain Tumor Cell (U-87MG) Using
Transmission Electron Microscopy
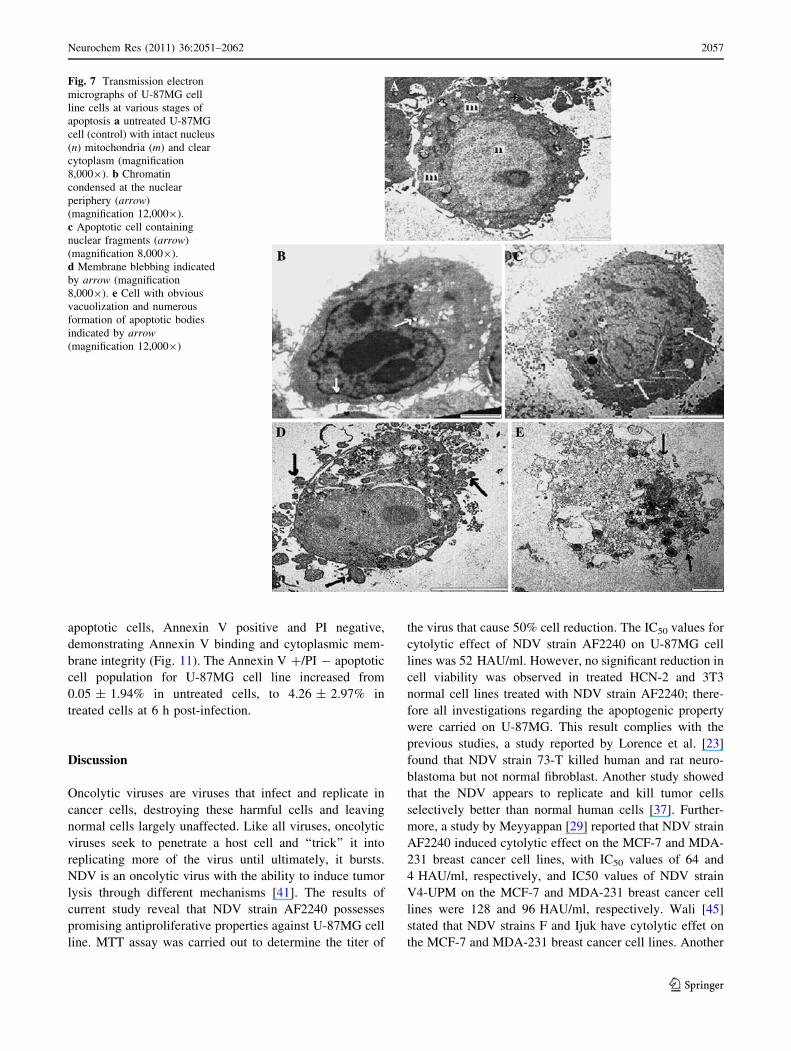
Treated cells showed death corresponded very well to the
cross sectional classical signs of apoptosis: cell shrinkage,
increased cellular granularity, the formation of apoptotic
bodies, and dilated nuclear membranes (Fig. 7). Mito-
chondria in treated cells were ruptured and condensed
(Fig. 7c). The evidences which suggested that the apopto-
genic effect of NDV on U-87MG cells is time-dependent
manner were obtained through observation of dynamic
micrographs images using transmission electron micros-
copy. Features of early stage of apoptosis was observed at
24 h post-inoculation such as, cell shrinkage, chromatin
condensation in dense masses under the nuclear membrane
and margination along the inner nuclear membrane, and
compaction of the cytoplasm with development of vacuoles
in the cytoplasm. At 48 h post-inoculation, the cells broke
up into discrete fragments to form apoptotic bodies and
nucleus fragmentation was observed. At 24 h post-inocu-
lation, cytoplasmic organelles such as intact mitochondria
can still be observed whereas at 48 h post-inoculation
numerous mitochondria were found in the centre of the
apoptotic cells. Membrane blebbing without disintegration
of the cellular membrane occurred at 48 and 72 h post-
inoculation indicating of late stage of apoptosis. At 72 h
post-inoculation secondary necrosis was observed and
some apoptotic bodies eventually degenerated (Fig. 7).
Effects of NDV on Brain Tumor Cell (U-87MG) DNA
Fragmentation
Fragmentation of chromosomal DNA is the biological
hallmark of apoptosis, and can be detected by a ladder
Fig. 1 Cytolytic effects of NDV strain AF2240 on U-87MG cell line.
The IC50 value is the concentration required for 50% growth
inhibition. Fifty percent of cell viability (IC50) was obtained at virus
titer of 52 ± 1.76 HAU/ml
Fig. 2 Cytolytic effect of NDV AF2240 strain on HCN-2 cell line.
The IC50 value was not obtained because the cell reduction was not
effected by same virus titer
Fig. 3 Cytolytic effect of NDV AF2240 strain on 3T3 cell line. The
IC50 value was not obtained because the cell reduction was not
effected by same virus titer
2054 Neurochem Res (2011) 36:2051–2062
123
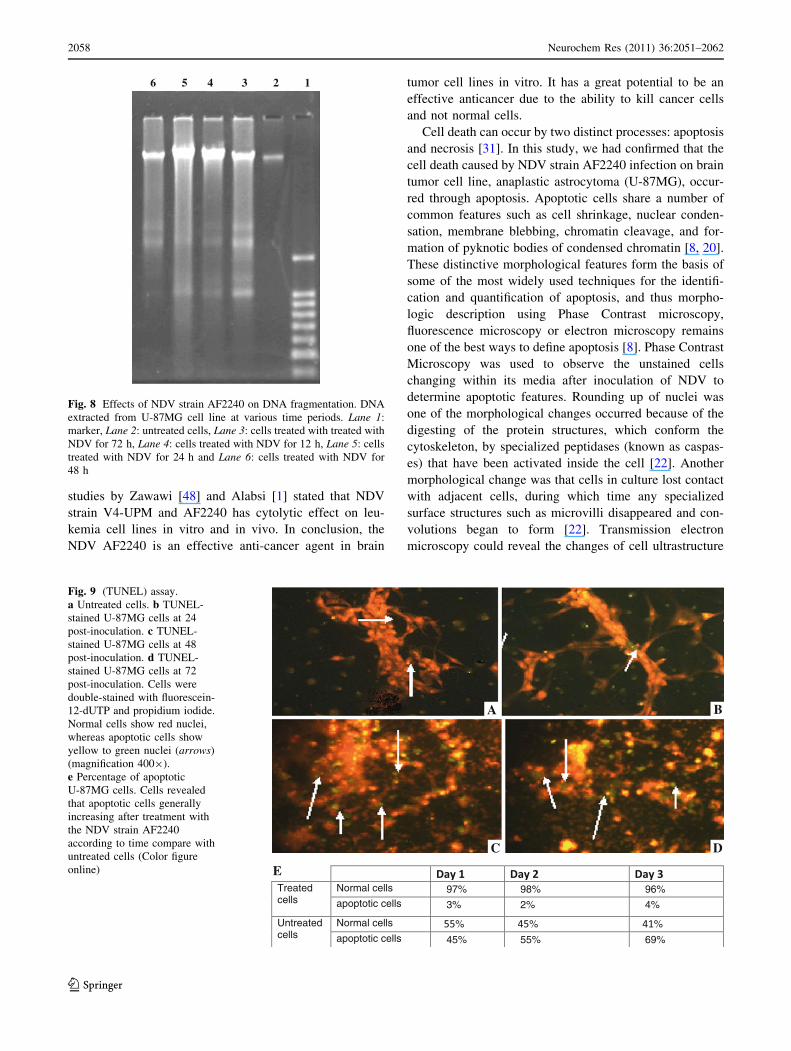
formation on gel electrophoresis. The DNA ladder assay is
generally accepted as specific for apoptosis because it
detects oligonucleosomal cleavage rather than artificial
DNA cleavage or necrosis. The DNA of U-87MG cells
treated for 24, 48 and 72 h with NDV strain AF2240 were
extracted and analyzed by agarose gel electrophoresis
(Fig. 8). Untreated U-87MG cells contained only high-
molecular-weight genomic DNA. Whereas DNA extracted
from U-87MG at 24, 48 and 72 h post-inoculation showed
the characteristic pattern of nucleosomal laddering specific
to apoptosis which was visible as faint bands on the gel
(Fig. 8). NDV strain AF2240 produced DNA fragments of
lower molecular weight consisting of multimers of
180–200 bp on U-87MG cell line.
Apoptotic (U-87MG) Cells Detection by TUNEL Assay
After Treatment of NDV
Apoptotic cells were detected and localization by green
fluorescence (FITC-12-dUTP) on a red background [pro-
pidium iodide (PI)] as observed by fluorescence micros-
copy, as described above. Untreated U-87MG cells were
shown in red because of the staining with propidium
iodide. Red staining is indicating viable cells which contain
intact genomic DNA. Whereas treated U-87MG cells were
shown the TUNEL? nuclei appeared as yellow to green
fluorescence. The treated cells containing multiple DNA
breaks at 24, 48 and 72 h post-inoculation were stained
yellow to green fluorescence indicating apoptosis (Fig. 9).
The analysis of many different slides revealed that the
green staining was generally more pronounced in apoptotic
cells treated with the NDV strain AF2240 according to
time (Fig. 9e).
Effects of NDV on Cell Cycle Distribution
The state of U-87MG cells was monitored by flow
cytometry after propidium iodide staining nuclei. DNA
histogram showed that NDV strain AF2240 did not induce
cell cycle arrest in any specific phase (Fig. 10). In
untreated U-87MG cells, the G1, S, and G2/M populations
represented 51.14, 31.10, and 14.63%, respectively
(Table 1). The treatment resulted in a loss of U-87MG cells
in all three phases of the cycle accompanied with a large
increase in the sub-G1 region (Table 1). NDV induced a
concentration- and time-dependent increase in the propor-
tion of sub-G1 population. Apoptosis peak (sub-G1) was
found in untreated cells with small percentage for about
3.14%. The sub-G1 population in U-87MG cells treated
with NDV strain AF2240 were 10.29 and 19.45% for 24
A
0
0.2
0.4
0.6
0.8
1
1.2
hoursO
D
CONTROL IC 50 IC 25
1
97
3
98
2
96
4
0
10
20
30
40
50
60
70
80
90
100
% c
ell p
op
ula
tio
n
days
2
61
39
57
43
51
49
0
20
40
60
80
100
%ce
ll p
op
lula
tio
n
days
3
55
45
47
53
45
55
0
20
40
60
80
100
%ce
ll p
op
ula
tio
n
0 20 40 60 80 1 2 3
1 2 3 1 2 3
days
non-viable viablenon-viable viable
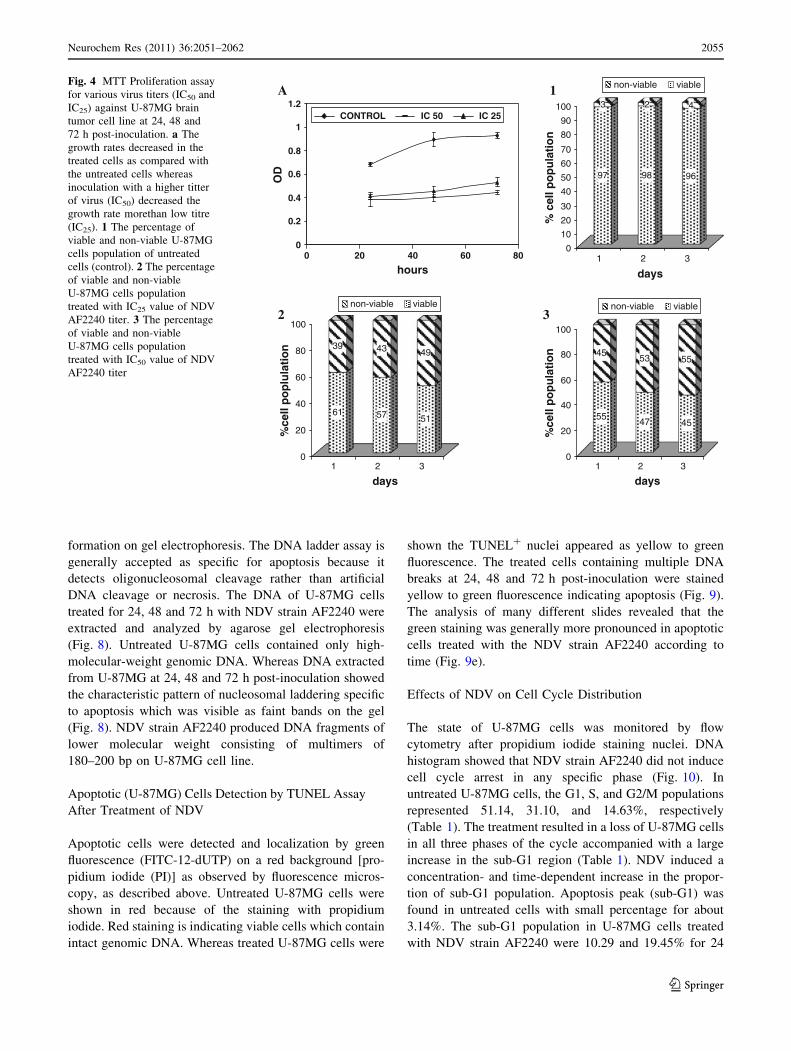
non-viable viableFig. 4 MTT Proliferation assay
for various virus titers (IC50 and
IC25) against U-87MG brain
tumor cell line at 24, 48 and
72 h post-inoculation. a The
growth rates decreased in the
treated cells as compared with
the untreated cells whereas
inoculation with a higher titter
of virus (IC50) decreased the
growth rate morethan low titre
(IC25). 1 The percentage of
viable and non-viable U-87MG
cells population of untreated
cells (control). 2 The percentage
of viable and non-viable
U-87MG cells population
treated with IC25 value of NDV
AF2240 titer. 3 The percentage
of viable and non-viable
U-87MG cells population
treated with IC50 value of NDV
AF2240 titer
Neurochem Res (2011) 36:2051–2062 2055
123

and 48 h post-inoculation, respectively (Fig. 10). This
virus-induced cell cycle perturbation concurred with the
results of the TUNEL assay to suggest that brain tumor
U-87MG cell lines undergo apoptosis more extensively
with increasing in time.
Early Apoptosis Detection on U-87MG Cells After
Treatment with NDV Strain AF2240 by Flow
Cytometry (Annexin V/PI Double Staining)
Apoptotic cells exclude all dyes which are in use for cell
viability assays, such as PI, while necrotic cells do not. In
cells with a damaged cell membrane PI induces a red
fluorescence on the DNA, whilst it is excluded by cells
with a preserved cytoplasm membrane. Hence during the
initial phase of apoptosis, the cells are still able to exclude
PI and therefore do not show any red fluorescence signal,
similar to that of living cells. Figure 11 showed the results
of Annexin V/PI flow cytometry of U-87MG cells after
treatment with IC50 value of NDV strain AF2240. The
lower left quadrant of the cytograms shows the viable cells,
which excluded PI and were negative for Annexin V
binding. The upper right quadrant represents the non-via-
ble, necrotic cells, positive for Annexin V binding and
showing PI uptake. The lower right quadrant represents the
Fig. 5 a Phase contrast microscope examination of untreated
U-87MG cells and b phase contrast microscope examination of
U-87MG cells treated with NDV AF2240 (IC50, 52 HAU/ml) after
72 h. Untreated cells showed normal structure without prominent
apoptosis and necrosis. The virus caused the cells to lose contact with
adjacent cells, rounding up of nuclei and cell membrane. The virus
caused the cells to lose contact with adjacent cells, and cell membrane
blebbing (arrow). (magnification 2009). c Fluorescence microscopy
examination of Untreated U-87MG cells (control) and d fluorescence
microscopy examination of U-87 MG cells treated with NDV AF2240
IC50, 52 HAU/ml after 72 h. Viable cells are uniformly green with
round nucleus (N), The apoptotic cells are green with condensed
chromatin (C), nuclear fragmentation (F), and cell membrane
blebbing (arrow) (magnification 4009) (Color figure online)
0
10
20
30
40
50
60
70
80
90
100
110
control 24 hrs 48 hrs 72 hrs
time (hours)
cells
per
cen
tag
e (%
)
viable
apoptoic
necrotic
Fig. 6 Percentages of viable, apoptotic, and necrotic cells after NDV
strain AF2240 treatment using fluorescence microscopy. U-87MG
cell death via apoptosis increased significantly (*P \ 0.001) in time-
dependent manner. However, no significant (P [ 0.001) difference
was observed in the cell count of necrosis
2056 Neurochem Res (2011) 36:2051–2062
123
apoptotic cells, Annexin V positive and PI negative,
demonstrating Annexin V binding and cytoplasmic mem-
brane integrity (Fig. 11). The Annexin V ?/PI - apoptotic
cell population for U-87MG cell line increased from
0.05 ± 1.94% in untreated cells, to 4.26 ± 2.97% in
treated cells at 6 h post-infection.
Discussion
Oncolytic viruses are viruses that infect and replicate in
cancer cells, destroying these harmful cells and leaving
normal cells largely unaffected. Like all viruses, oncolytic
viruses seek to penetrate a host cell and ‘‘trick’’ it into
replicating more of the virus until ultimately, it bursts.
NDV is an oncolytic virus with the ability to induce tumor
lysis through different mechanisms [41]. The results of
current study reveal that NDV strain AF2240 possesses
promising antiproliferative properties against U-87MG cell
line. MTT assay was carried out to determine the titer of
the virus that cause 50% cell reduction. The IC50 values for
cytolytic effect of NDV strain AF2240 on U-87MG cell
lines was 52 HAU/ml. However, no significant reduction in
cell viability was observed in treated HCN-2 and 3T3
normal cell lines treated with NDV strain AF2240; there-
fore all investigations regarding the apoptogenic property
were carried on U-87MG. This result complies with the
previous studies, a study reported by Lorence et al. [23]
found that NDV strain 73-T killed human and rat neuro-
blastoma but not normal fibroblast. Another study showed
that the NDV appears to replicate and kill tumor cells
selectively better than normal human cells [37]. Further-
more, a study by Meyyappan [29] reported that NDV strain
AF2240 induced cytolytic effect on the MCF-7 and MDA-
231 breast cancer cell lines, with IC50 values of 64 and
4 HAU/ml, respectively, and IC50 values of NDV strain
V4-UPM on the MCF-7 and MDA-231 breast cancer cell
lines were 128 and 96 HAU/ml, respectively. Wali [45]
stated that NDV strains F and Ijuk have cytolytic effet on
the MCF-7 and MDA-231 breast cancer cell lines. Another
Fig. 7 Transmission electron
micrographs of U-87MG cell
line cells at various stages of
apoptosis a untreated U-87MG
cell (control) with intact nucleus
(n) mitochondria (m) and clear
cytoplasm (magnification
8,0009). b Chromatin
condensed at the nuclear
periphery (arrow)
(magnification 12,0009).
c Apoptotic cell containing
nuclear fragments (arrow)
(magnification 8,0009).
d Membrane blebbing indicated
by arrow (magnification
8,0009). e Cell with obvious
vacuolization and numerous
formation of apoptotic bodies
indicated by arrow(magnification 12,0009)
Neurochem Res (2011) 36:2051–2062 2057
123
studies by Zawawi [48] and Alabsi [1] stated that NDV
strain V4-UPM and AF2240 has cytolytic effect on leu-
kemia cell lines in vitro and in vivo. In conclusion, the
NDV AF2240 is an effective anti-cancer agent in brain
tumor cell lines in vitro. It has a great potential to be an
effective anticancer due to the ability to kill cancer cells
and not normal cells.
Cell death can occur by two distinct processes: apoptosis
and necrosis [31]. In this study, we had confirmed that the
cell death caused by NDV strain AF2240 infection on brain
tumor cell line, anaplastic astrocytoma (U-87MG), occur-
red through apoptosis. Apoptotic cells share a number of
common features such as cell shrinkage, nuclear conden-
sation, membrane blebbing, chromatin cleavage, and for-
mation of pyknotic bodies of condensed chromatin [8, 20].
These distinctive morphological features form the basis of
some of the most widely used techniques for the identifi-
cation and quantification of apoptosis, and thus morpho-
logic description using Phase Contrast microscopy,
fluorescence microscopy or electron microscopy remains
one of the best ways to define apoptosis [8]. Phase Contrast
Microscopy was used to observe the unstained cells
changing within its media after inoculation of NDV to
determine apoptotic features. Rounding up of nuclei was
one of the morphological changes occurred because of the
digesting of the protein structures, which conform the
cytoskeleton, by specialized peptidases (known as caspas-
es) that have been activated inside the cell [22]. Another
morphological change was that cells in culture lost contact
with adjacent cells, during which time any specialized
surface structures such as microvilli disappeared and con-
volutions began to form [22]. Transmission electron
microscopy could reveal the changes of cell ultrastructure
6 5 4 3 2 1
Fig. 8 Effects of NDV strain AF2240 on DNA fragmentation. DNA
extracted from U-87MG cell line at various time periods. Lane 1:
marker, Lane 2: untreated cells, Lane 3: cells treated with treated with
NDV for 72 h, Lane 4: cells treated with NDV for 12 h, Lane 5: cells
treated with NDV for 24 h and Lane 6: cells treated with NDV for
48 h
%
E Treated cells
Normal cells 97% 89 96% apoptotic cells 3% %2 4%
Untreated cells
Normal cells % % %apoptotic cells 45% %55 69%
C D
A B
Fig. 9 (TUNEL) assay.
a Untreated cells. b TUNEL-
stained U-87MG cells at 24
post-inoculation. c TUNEL-
stained U-87MG cells at 48
post-inoculation. d TUNEL-
stained U-87MG cells at 72
post-inoculation. Cells were
double-stained with fluorescein-
12-dUTP and propidium iodide.
Normal cells show red nuclei,
whereas apoptotic cells show
yellow to green nuclei (arrows)
(magnification 4009).
e Percentage of apoptotic
U-87MG cells. Cells revealed
that apoptotic cells generally
increasing after treatment with
the NDV strain AF2240
according to time compare with
untreated cells (Color figure
online)
2058 Neurochem Res (2011) 36:2051–2062
123

during the apoptotic process [21]. Intracellular and plasma
membrane structural modifications have been widely rec-
ognized as crucial factors involved in cell injury and death.
Changes in nuclear morphology and in organelle structure
as well as specific phenomena at the cell surface, namely
surface smoothing and surface blebbing, are often consid-
ered as markers associated with cell pathology [25].
Morphological features of apoptosis were also observed
by using fluorescent microscopy (AO/PI method). Chro-
matin condensation, nuclear shrinkage and formation of
apoptotic bodies can easily be observed under fluorescence
microscopy, after appropriate staining of nuclei with DNA-
specific fluorochromes [6]. The AO/PI viability assay is a
fluorometric cell viability assay. AO is a membrane-
permeable cationic dye that binds to nucleic acids of viable
cells and at low concentrations it causes a green fluores-
cence. PI is impermeable to intact membranes but readily
penetrates the membranes of nonviable cells and binds to
DNA or RNA, causing orange fluorescence. When AO and
PI are used simultaneously, viable cells stained green
fluorescence under dark field fluorescence microscopy,
while nonviable cells stained fluorescence orange [27].
The cell viability results showed an obvious decrease of
living cells in treated U-87MG cells. When viewed under
fluorescence microscopy, it was observed that untreated
cells had round intact nuclei and stained green indicating
viable cells whereas the treated cells stained green and
exhibited features of apoptotic cells, which had irregular
Fig. 10 Cell cycle (DNA
content) flow cytometer
histograms of U-87MG brain
tumor cell lines treated with
IC50 value of NDV strain
AF2240 a untreated cells
(control) b cells treated with
NDV after 24 h c cells treated
with NDV after 48 h. Shows
increasing in percentage of sub-
G1 (apoptotic) cell population
(broken DNA)
Table 1 Percentage of U-87MG cells in different cell cycle phase after treatment with virus
U-87MG cells Cell cycle phase
Sub-G1 (%) G1 (%) S (%) G2/M (%)
Untreated cells (control) 3.14 51.14 31.10 14.63
Cells treated after 24 h 10.29 49.63 29.94 10.14
Cells treated after 48 h 19.45 47.78 25.70 8.06
Fig. 11 Contour diagram of Annexin V/PI flow cytometry
a untreated U-87MG and b U-87MG cells at 6 h post-inoculation
of IC50 value of NDV strain AF 2240. The lower left quadrants of
each panel (Rl) show the viable cells, which exclude PI and are
negative for Annexin V binding. The upper right quadrants (R2)
contain the non-viable, necrotic cells, positive for Annexin V binding
and for PI uptake. The lower right quadrants (R3) represent the
apoptotic cells, Annexin V positive and PI negative. The Annexin
V ?/PI - apoptotic cell population for U-87MG cell line increased
from 0.05 ± 1.94% in untreated cells, to 4.26 ± 2.97%
Neurochem Res (2011) 36:2051–2062 2059
123

shaped nucleus, shrinkage, condensed chromatin and cell
membrane blebbing [5]. Quantification of apoptosis with
differential scoring of treated and control cells revealed a
significant (P \ 0.05) difference in the number of apop-
totic cells in a time-dependent manner but with insignifi-
cant difference between numbers of necrosis positive cells
in control and treated cells, concluding that NDV strain
AF2240, did not induce necrotic effects. Fluorescence
microscopy examination confirmed the onset of apoptosis
features. These morphological criteria that implicate
apoptotic cell death were further confirmed by transmission
electron microscopy and transmission electron microscopy.
TEM has been considered a milestone of the research in
the field of apoptosis. By means of TEM the results in this
study showed that untreated brain tumor U-87MG cells
showed no changes and exhibited intact nucleus with clear
cytoplasm while after treatment of U-87MG cells with
NDV strain AF2240, some cells showed typical morpho-
logic changes of apoptosis, including shrinkage and
membrane blebbing. Other changes occur within the
nucleus during apoptotic death such as the appearance of
dense, crescent-shaped chromatin aggregates which line
nuclear membrane. Later, the nucleolus disintegrates;
nuclear membrane develops deep invaginations and, ulti-
mately, the nucleus fragmente into dense granular particles
(apoptotic bodies) which eventually degenerated to ‘‘sec-
ondary necrosis’’ [38]. It is thought that the nuclear chan-
ges are due to activation of endogenous nuclease(s) which
cleaves DNA into oligonucleosomal fragments [35, 38].
In addition the terminal deoxynucleotidyl transferase
(TdT)-mediated dUTP-fluorescein nick-end labeling
(TUNEL) assay was also conducted to confirm the results
of morphological features of the oncolytic effect of NDV
on the U-87MG brain cell line. TUNEL staining is the
standard technique for detection of apoptosis because it
permits visualization of DNA cleavage which can be
visualized and quantities directly by fluorescence micros-
copy [14, 15, 39, 46].
This test showed that NDV strain AF2240 caused a
significant increase in the percentage of apoptotic cells on
U-87MG brain cell line and the percentage of apoptotic
cells increased with increasing of time, in which apoptotic
cells were more abounded at longer durations of treatment.
In the cell populations of control in both cell lines, cells
were having intact DNA or low undetectable levels of
fragmentation but treated cells were TUNEL positive and
apoptotic cells increased correlated with the time of
treatment.
DNA ladder assay provides a sensitive assay for the
detection of DNA fragmentation, but this method is qual-
itative rather than quantitative [44]. In this study we
reported that NDV strain AF2240 stimulate DNA frag-
mentation characteristic of apoptosis in U-87MG cell line
at 24, 48 and 72 h post-inoculation. The DNA ladder assay
can be decreased in samples where the number of apoptotic
cells is low [28] and during the apoptotic process genomic
DNA is cleaved in more consequent steps, first cleavage
into fragments with length 300 kbp is followed by addi-
tional fragmentation into 50 kbp and thereafter to oligo-
nucleosomal DNA fragments [35]. It was also reported by
[6] that DNA laddering is not detectable in adherent cells
but is characteristic of the floating cells regardless of
treatment. Even with enhancement of the fragmented DNA
in the sample, DNA ladder formation is observed only
when the extent of oligonucleosomal cleavage is promi-
nent. Internucleosomal cleavage of DNA is likely to be in
the later phase of apoptotic process [7, 16, 35]. In most cell
types, the biochemical characteristics of apoptotic response
include activation of endogenous calcium and magnesium
dependent endonucleases, leading to fragmentation of the
chromosomal DNA. Initially, the DNA fragments are large
(50–300 kb) but are later digested to oligonucleosomal size
(multimers of 180–200 bp). The formation of this distinct
DNA ladder is considered to be a biochemical hallmark of
apoptosis.
Further confirmation of the mode of cell death was
carried by flow cytometric analysis of cell cycle. It mea-
sures apoptotic changes in cells by staining with DNA dyes
[43]. This method is useful for quantitative estimates of the
fractions of cells in the different phases of the cell cycle
[13]. Untreated and treated U-87MG brain cell line were
evaluated for apoptosis by measuring the amount of
apoptotic cells using of DNA flow cytometry (FCM). The
apoptotic cells with degraded DNA were represented in so-
called ‘‘sub-G1’’ peaks on DNA histograms. Apoptotic
cells, due to a change in membrane permeability, showed
an increased up-take of the vital dye, PI, compared to live
cells [33, 43].
Figures and tables reveal that there was a loss of treated
U-87MG cells in all three phases of the cycle (G1, S and
G2/M) accompanied with a large increase in the sub-G1
region (apoptosis peak) in the fluorescence histograms.
Concluding that NDV strain AF2240 caused an increasing
in the percentage of sub-G1 region which increased with
increasing of time and did not induce specific cell cycle
arrest in specific phase.
Annexin binding assay is a method permits the detection
of the early phases of apoptosis before the loss of cell
membrane integrity [3, 44]. This is based on the phenom-
enon that phosphatidylserine (PS) is exposed at the outer
membrane of the cell during apoptosis and on the ability of
annexin V to bind to PS with high affinity [10]. Phospha-
tidylserine (PS) only exists in the cytoplasm side of
cell plasma membrane, and externalization of PS occurs in
the early stage of apoptosis. Annexin-V could specifi-
cally conjugate to the PS to detect the apoptotic cells [21].
2060 Neurochem Res (2011) 36:2051–2062
123

The analysis of the treated U-87MG cells by annexin V
versus PI, revealed four populations: live cells, apoptotic
cells, late apoptotic cells and permeabilized cells. In this
experiment, we observed increasing in the percentage of
target cells in the annexin positive–PI negative (early
apoptosis) quadrant at 6 h post-inoculation.
These results complies with the other studies [1, 29, 45,
48], which stated that the effectiveness of NDV strains
included NDV strain AF2240 as oncolytic agent was found
on breast cancer cell lines and leukemia cell lines and the
nature of cell death caused by this virus was characterized
as apoptosis. Therefore these results show that NDV strain
AF2240 was capable to induce apoptosis on brain tumor
cells, anaplastic astrocytoma (U-87MG), in vitro.
To put these results together, we believe that the
induction of apoptosis play a role in inhibiting brain tumor
cells when treat with NDV strain AF2240 and that cyto-
toxicity increasing while increasing the titer of the virus.
This discovery may provide a theoretical basis for this type
of treatment. The mechanism of the virus infection is still
unclear. NDV would have a bright future in the treatment
of tumors and further work may lead to relative antitumor
agents to be used in clinical settings.
Acknowledgments This research was funded in part by the
National Cancer Council (MAKNA), Malaysia. The authors also
acknowledge additional support from Universiti Putra Malaysia
(UPM), Serdang, Malaysia.
References
1. Alabsi A (2008) Effects of newcastle disease virus strains
AF2240 and V4-UPM on cytolysis and apoptosis of leukemia cell
lines. Doctor of philosophy Thesis, Universiti Putra Malaysia,
Malaysia
2. Alexander DJ (1988) Newcastle disease diagnosis. In: Alexander
DJ (ed) Newcastle disease. Kluwer Acadimic Publishers, Neth-
erlands, pp 147–160
3. Aubry J, Blaecke A, Lecoanet-Henchoz S, Jeannin P, Herbault N,
Caron G, Moine V, Bonnefoy J (1999) Annexin V used for
measuring apoptosis in the early events of cellular cytotoxicity.
Cytometry 37:197–204
4. Chambers P, Samson ACR (1980) A new structural protein for
Newcastle disease virus. J Gen Virol 50:155–166
5. Chan KM, Rajab NF, Ishak MHA, Ali AM, Yusoff K,
Din LB, Inayat-Hussain SH (2006) Goniothalamin induces apop-
tosis in vascular smooth muscle cells. Chemico-Biol Interact
159:129–140
6. Clarke RG, Lund EK, Johnson IT, Pinder AC (2000) Apoptosis
can be detected in attached colonic adenocarcinoma HT29 cells
using Annexin V binding, but not by TUNEL assay or sub-G0
DNA content. Cytometry 39:141–150
7. Cohen GM, Sun XM, Snowden RT, Dinsdale D, Skilleter DN
(1992) Key morphological features of apoptosis may occur in the
absence of internucleosomal DNA fragmentation. Biochem J
286:331–334
8. Doonan F, Cotter TG (2008) Morphological assessment of
apoptosis. Methods 44:200–204
9. Doyle TM (1927) A hitherto unrecorded disease of fowls due to a
filter passing virus. J Comp Pathol 40:144–169
10. Engeland MV, Nieland LJW, Ramaekers FCS, Schutte B,
Reutelingsperger CPM (1998) Annexin V-Affinity assay: a
review on an apoptosis detection system based on phosphatidyl-
serine exposure. Cytom 31:1–9
11. Fenner F, Bachmann P, Gibbs E, Murphy F, Studdert M, White D
(1987) Veterinary Virol. Academic Press, Orlando
12. Freeman AI, Zakay-Rones Z, Gomori JM, Linetsky E, Rasooly L,
Greenbaum E, Rozenman-Yair S, Panet A, Libson E, Irving CS,
Galun E, Siegal T (2006) Phase I/II trial of intravenous NDV-
HUJ oncolytic virus in recurrent glioblastoma multiforme. Mol
Ther 13:221–228
13. Fried J, Perez AG, Clarkson BD (1976) Flow cytofluorometric
analysis of cell cycle distributions using propidium iodide,
Properties of the method and mathematical analysis of the data.
J Cell Biol 71:172–181
14. Gao N, Keane MJ, Ong T, Ye J, Miller WE, Wallace WE (2001)
Effects of phospholipid surfactant on apoptosis induction by
respirable quartz and kaolin in NR8383 rat pulmonary. Toxicol
Appl Pharmacol 175:217–225
15. Gavrieli Y, Sherman Y, Ben-Sasson SA (1992) Identification of
programmed cell death in situ via specific labeling of nuclear
DNA fragmentation. J Cell Biol 119:493–501
16. Gooch JL, Yee D (1999) Strain-specific differences in formation
of apoptotic DNA ladders in MCF-7 breast cancer cells. Cancer
Lett 144:31–37
17. Guan T, Qin F, Du J, Geng L, Zhang Y, Li M (2007) AICAR inhibits
proliferation and induced S-phase arrest, and promotes apoptosis in
CaSki cells. Acta Pharmacologica Sinica 28:1984–1990
18. Henson JW (2005) Glioblastoma multiforme and anaplastic gli-
oma: a patient guide. http://brain.mgh.harvard.edu/PatientGuide.
htm. Accessed on 7 Feb
19. Kirn D, Martuza RL, Zwiebel J (2001) Replication-selective
virotherapy for cancer: biological principles, risk management
and future directions. Nat Med 7:781–787
20. Lin JC, Ho YS, Lee JJ, Liu CL, Yang TL, Wu CH
(2007) Induction of apoptosis and cell-cycle arrest in human
colon cancer cells by meclizine. Food Chem Toxicol 45:
935–944
21. Lin W, Li D, Chen Q, Lu H (2004) Clinical and experimental
study of oxaliplatin in treating human gastric carcinoma. World J
Gastroenterol 19:2911–2915
22. Liu C, Xu HY, Liu (2001) Induction of caspase-dependent
apoptosis in cultured cells by the avian coronavirus infectious
bronchitis virus. Am Soc Microbiol 75:6402–6409
23. Lorence RM, Richard KW, Katubig BB, Reyes HM, Phuangsab
A, Mitchell BR, Cascino CJ, Walter RJ, Peeple ME (1994)
Complete regression of human neuroblastoma xenographts in
athimic mice after local Newcastle disease virus therapy. Natl
Cancer Inst 86:1228–1233
24. Maher EA, Furnari FB, Bachoo RM, Rowitch DH, Louis DN,
Cavenee WK, DePinho RA (2001) Malignant glioma: genetics
and biology of a grave matter. Genes Dev 15:1311–1333
25. Malorni W, Fais S, Fiorentini C (1998) Morphological aspects of
apoptosis. In: Cossarizza A, Boraschi D (eds) Apoptosis: labo-
ratory manual of experimental methods. Purde University, West
Hafayette.
26. Mangiardi JR, Kane H (2003) A,B,C’s of brain tumors from their
biology to their treatment. http://drshortz.com/brain%20tumor.
htm. Accessed on 27 March
27. Mascotti K, McCullough J, Burger SR (2000) HPC viability
measurement: trypan blue versus acridine orange and propidium
iodide. Transfusion 40:693–696
28. Matalova E, Panova A (2002) Detection of apoptotic DNA ladder
in pig leukocytes and its precision using LM-PCR (Ligation
Neurochem Res (2011) 36:2051–2062 2061
123

Mediated Polymerase Chain Reaction). J Univ Vet Pharm Sci
71:163–168
29. Meyyappan M (2003) Oncolytic effect of Newcastle disease virus
on the MCF-7 and MDA-231 breast cancer cell lines. Master
Thesis, Universiti Putra Malaysia, Malaysia
30. Mishell BB, Shiiqi SM, Henry C (1980) Selected methods. In:
Mishell BB, Shiiqi SM (eds) Cellular immunology. Freeman, San
Francisco, pp 21–22
31. Muneesh T, Vishva MD (1995) Fas- and tumor necrosis factor-
induced apoptosis is inhibited by the poxvirus crmA gene prod-
uct. Am Soc Biochem Mol Biol 270:3255–3260
32. Nelson NJ (1999) Scientific Interest in Newcastle disease virus is
reviving. J Natl Cancer Inst 91:1708–1710
33. Nicoletti I, Migliorati G, Pagliacci MC, Grignani F, Riccardi C
(1991) A rapid and simple method for measuring thymocyte
apoptosis by propidium iodide staining and flow cytometry.
J Immunol Methods 139:271–279
34. Niederhuber JE (2006) Questions and answers about Newcastle
disease virus. http://www.cancer.gov/cancertopics/pdq/cam/NDV/
Patient/page2. Accessed 16 March
35. Oberhammer F, Wilson JW, Dive C, Morris ID, Hickman JA,
Wakeling AE, Walker PR, Sikorska M (1993) Apoptotic death in
epithelial cells: cleavage of DNA to 300 and/or 50 kb fragments
prior to or in the absence of internucleosomal fragmentation.
EMBO J 12:3679–3684
36. Omar A, Aini I, Ali A, Othman F, Yusoff K, Abdullah JM, Wali H,
Zawawi M, Meyyappan N (2003) An overview on the development
of Newcastle disease virus as an anti-cancer therapy. Malays J Med
Sci 10:4–12
37. Reichard KW, Lorence RM, Cascino CJ, Peeples ME, Walter RJ,
Fernando MB, Reyes HM, Greager JA (1992) Newcastle disease
virus selectively kills human tumor cells. J Surg Res 52:448–453
38. Sho E, Sho M, Singh TM, Xu C, Zarins CK, Masuda H (2001)
Blood flow decrease induces apoptosis of endothelial cells in
previously dilated arteries resulting from chronic high blood flow.
Arterioscler Thromb Vasc Biol 21:1139–1145
39. Stahelin BJ, Marti U, Solioz M, Zimmermann H, Reichen J
(1998) False positive staining in the TUNEL assay to detect
apoptosis in liver and intestine is caused by endogenous nucleases
and inhibited by diethyl pyrocarbonate. J Clin Pathol 51:204–208
40. Stupp R, Mason WP, van den Bent MJ, Weller M, Fisher B,
Taphoorn MJB, Belanger K, Brandes AA, Marosi C, Bogdahn U,
Curschmann J, Janzer RC, Ludwin SK, Gorlia T, Allgeier A,
Lacombe D, Cairncross JG, Eisenhauer E, Mirimanoff RO (2005)
Radiotherapy plus concomitant and adjuvant temozolomide for
glioblastoma. N Engl J Med 352:987–996
41. Szeberenyi J, Fabian Z, Torocsik B, Kiss K, Csatary LK (2003)
Newcastle disease virus-induced apoptosis in PC12 pheochro-
mocytoma cells. Am J Therap 10:282–288
42. Tatter SB (2005) The new WHO Classification of Tumors
affecting the Central Nervous System. http://neurosurgery.mgh.
harvard.edu/newwhobt.htm. Accessed on 11 May
43. Telford WG, King LE, Fraker PJ (1994) Rapid quantitation of
apoptosis in pure and heterogeneous cell populations using flow
cytometry. J Immunol Methods 172:1–16
44. Vermes I, Haanen C, Steffens-Nakken H, Reutelingsperger C
(1995) A novel assay for apoptosis flow cytometric detection of
phosphatidylserine early apoptotic cells using fluorescein labeled
expression on Annexin V. J Immunol Methods 184:39–51
45. Wali HSM (2003) The effects of Newcastle disease virus (NDV) on
breast cancer cell lines. Master Thesis, Universiti Putra Malaysia,
Malaysia
46. Yasuhara S, Zhu Y, Matsui T, Tipirneni N, Yasuhara Y, Kaneki
M, Rosenzweig A, Jeevendra Martyn JA (2003) Comparison of
Comet assay, electron microscopy, and flow cytometry for
detection of apoptosis. J Histochem Cytochem 51:873–885
47. Yusoff K, Tan WS, Lau CH, Ng BK, Ibrahim AL (1996) Sequence
of the haemagglutinin-neuraminadase gene of the Newcastle dis-
ease virus oral vaccine strain V4 (UPM). Avian Pathol 25:837–844
48. Zawawi M (2007) Cytolytic effect of Newcastle disease virus
strain V4 (UPM) on leukemic cell lines CEM-SS and HL60.
Master Thesis, Universiti Putra Malaysia, Malaysia
2062 Neurochem Res (2011) 36:2051–2062
123




















