
Isolation of differentially expressed genes in NPM-ALK-positive anaplastic large cell lymphoma
9
Central European Journal of Biology * E-mail: [email protected] Research Article Institute of Agriculture, Lithuanian Research Centre for Agriculture and Forestry, LT58344 Akademija, Kedainiai, Lithuania Rita Armonienė, Kristina Jonavičienė*, Vytautas Ruzgas, Gintaras Brazauskas Isolation of differentially expressed genes in wheat caryopses with contrasting starch granule size 1. Introduction Wheat is one of the most important crop species grown worldwide. More than 200 million hectares of wheat are cultivated annually with a total harvest of almost 600 million tonnes of grain (FAO; http://www.fao.org/). The storage starches produced in cereal endosperm account for over 90% of the world market for starch (the major cereals in terms of tonnage of yield are maize and wheat). Most of wheat harvested is consumed by humans for making a wide range of bread items or used for livestock feeding. However, starch being cheap and renewable raw material is used not only in the agri-food sector, but in many non-food industrial applications, such as manufacturing of protein–based films or starch– based plastics [1] and as a source of energy after conversion to ethanol [2,3]. The synthesis of starch in the endosperm occurs within the amyloplasts and is completed by a series of enzymatic reactions catalyzed by several different classes of enzymes [4]. Starch synthases (SS, E.C. 2.4.1.21) are important factors controlling the rate of starch synthesis, while the granule–bound starch synthases (GBSS: EC 2.4.1.21) are related to the synthesis of amylose. Starch branching enzymes (SBEs, E.C. 2.4.1.18) form the branched structure of the amylopectin molecule, while amylopectin is synthesized by the coordinated actions of SS, starch branching enzymes (SBEs, E.C. 2.4.1.18) and starch debranching enzymes (DBEs, E.C. 3.2.1.41 and E.C. 3.2.1.68). Starch consists of two major glucose polymers (amylose and amylopectin) with the ratio of amylose/amylopectin ranging between 25–28 and 72–75%, respectively. The relative amounts of amylose and amylopectin are responsible for starch physical and chemical properties with strong influences on the functional properties of flour and on its specific uses in the food and manufacturing industries. In wheat, starch granules exhibit a bimodal size distribution, a characteristic unique to members of the grass Triticeae family. The starch granules, designated A– and B–type [5], can be distinguished based on Cent. Eur. J. Biol. • 8(3) • 2013 • 297-305 DOI: 10.2478/s11535-013-0127-z 297 Received 31 July 2012; Accepted 19 November 2012 Keywords: cDNA-AFLP • Starch synthesis genes • Triticum aestivum • Endosperm development • Relative gene expression • Starch A–granules Abstract: In order to identify genes responsible for starch granule initiation during early development of wheat caryopsis, nine winter wheat breeding lines were studied. Two breeding lines, which are the most diverse in A–type granule size (26.85 μm versus 23.65 μm) were chosen for further differential gene expression analysis in developing caryopses at 10 and 15 days post–anthesis (DPA). cDNA–amplified fragment length polymorphism (cDNA-AFLP) analysis resulted in 384 transcript-derived fragments, out of which 18 were identified as being differentially expressed. Six differentially expressed genes, together with the six well–known starch biosynthesis genes, were chosen for semi-quantitative gene expression analysis in developing wheat caryopses at 10 and 15 DPA. This study provides genomic information on 18 genes differentially expressed at early stages of wheat caryopses development and reports on the identification of genes putatively involved in the production of large A–type granules. These genes are targets for further validation on their role in starch granule synthesis control and provide the basis for the development of DNA marker tools in winter wheat breeding for enhanced starch quality. © Versita Sp. z o.o. Author’s copy
-
Upload
univ-lyon1 -
Category
Documents
-
view
3 -
download
0
Transcript of Isolation of differentially expressed genes in NPM-ALK-positive anaplastic large cell lymphoma

Central European Journal of Biology
* E-mail: [email protected]
Research Article
Institute of Agriculture, Lithuanian Research Centre for Agriculture and Forestry, LT58344 Akademija, Kedainiai, Lithuania
Rita Armonienė, Kristina Jonavičienė*, Vytautas Ruzgas, Gintaras Brazauskas
Isolation of differentially expressed genes in wheat caryopses with contrasting starch granule size
1. IntroductionWheat is one of the most important crop species grown worldwide. More than 200 million hectares of wheat are cultivated annually with a total harvest of almost 600 million tonnes of grain (FAO; http://www.fao.org/). The storage starches produced in cereal endosperm account for over 90% of the world market for starch (the major cereals in terms of tonnage of yield are maize and wheat). Most of wheat harvested is consumed by humans for making a wide range of bread items or used for livestock feeding. However, starch being cheap and renewable raw material is used not only in the agri-food sector, but in many non-food industrial applications, such as manufacturing of protein–based films or starch–based plastics [1] and as a source of energy after conversion to ethanol [2,3].
The synthesis of starch in the endosperm occurs within the amyloplasts and is completed by a series of enzymatic reactions catalyzed by several different classes of enzymes [4]. Starch synthases (SS, E.C.
2.4.1.21) are important factors controlling the rate of starch synthesis, while the granule–bound starch synthases (GBSS: EC 2.4.1.21) are related to the synthesis of amylose. Starch branching enzymes (SBEs, E.C. 2.4.1.18) form the branched structure of the amylopectin molecule, while amylopectin is synthesized by the coordinated actions of SS, starch branching enzymes (SBEs, E.C. 2.4.1.18) and starch debranching enzymes (DBEs, E.C. 3.2.1.41 and E.C. 3.2.1.68). Starch consists of two major glucose polymers (amylose and amylopectin) with the ratio of amylose/amylopectin ranging between 25–28 and 72–75%, respectively. The relative amounts of amylose and amylopectin are responsible for starch physical and chemical properties with strong influences on the functional properties of flour and on its specific uses in the food and manufacturing industries.
In wheat, starch granules exhibit a bimodal size distribution, a characteristic unique to members of the grass Triticeae family. The starch granules, designated A– and B–type [5], can be distinguished based on
Cent. Eur. J. Biol. • 8(3) • 2013 • 297-305DOI: 10.2478/s11535-013-0127-z
297
Received 31 July 2012; Accepted 19 November 2012
Keywords: cDNA-AFLP • Starch synthesis genes • Triticum aestivum • Endosperm development • Relative gene expression • Starch A–granules
Abstract: In order to identify genes responsible for starch granule initiation during early development of wheat caryopsis, nine winter wheat breeding lines were studied. Two breeding lines, which are the most diverse in A–type granule size (26.85 μm versus 23.65 μm) were chosen for further differential gene expression analysis in developing caryopses at 10 and 15 days post–anthesis (DPA). cDNA–amplified fragment length polymorphism (cDNA-AFLP) analysis resulted in 384 transcript-derived fragments, out of which 18 were identified as being differentially expressed. Six differentially expressed genes, together with the six well–known starch biosynthesis genes, were chosen for semi-quantitative gene expression analysis in developing wheat caryopses at 10 and 15 DPA. This study provides genomic information on 18 genes differentially expressed at early stages of wheat caryopses development and reports on the identification of genes putatively involved in the production of large A–type granules. These genes are targets for further validation on their role in starch granule synthesis control and provide the basis for the development of DNA marker tools in winter wheat breeding for enhanced starch quality.
© Versita Sp. z o.o.
Author’s co
py

Isolation of differentially expressed genes in wheat caryopses with contrasting starch granule size
size, shape, relative proportion, and the timing of their initiation in the endosperm, a process which, presumably, is under a defined genetic program. C–type starch granules have also been reported in wheat [6,7]. Differences in starch granule composition [8] and their molecular structure [9] have been reported to play an important role in product application [10].
Despite considerable advances in our knowledge of the starch biosynthesis pathway, factors controlling starch granule initiation and granule size remain unclear. Loss of isoamylase activity from barley, rice and potato leads to an increase in granule initiation [11,12], consistent with the idea that isoamylases suppress the sites of new granule initiation, although DBEs are probably not directly involved in initiating granule synthesis [13-15]. Down–regulation of a pullulanase-type DBE (limit dextrinase) inhibitor activity in barley revealed reduced numbers of the small B–type starch granules, as well as reduced amylose relative to amylopectin levels and reduced total starch [16]. This confirms the important role of pullulanase–type DBE during determination of starch granule size. Analysis of SS IV in Arabidopsis thaliana mutants revealed a specific function for this enzyme class in the control of granule numbers being selectively involved in the priming of starch granules [17]. However, the specific factors controlling the size of starch granules in any species are unknown. The objective of this study was (1) to screen a set of advanced winter wheat lines for the starch A–granule size; (2) to identify differentially expressed genes in developing wheat caryopses with a contrasting starch A-granule sizes; and (3) to compare expression patterns of the candidate and well–known starch biosynthesis genes in developing wheat caryopses.
2. Experimental Procedures2.1 Plant materialNine winter wheat breeding lines of contrasting A–type starch granule size were selected from winter wheat collection of Lithuanian Research Centre for Agriculture and Forestry (Table 1). Six seeds of each breeding line were sown into pots (10 dm3) filled with peat substrate in three replications. Plants were established with regular irrigation and fertilization in a greenhouse for one month and then vernalized for two months under semi–natural conditions in the greenhouse by maintaining +4ºC without supplemental lightning. After vernalization plants were grown in the greenhouse at 20/16ºC (day/night) with a photoperiod of 16 h provided with a supplemental lighting if needed. Developing wheat caryopses from the middle part of the ear of three plants per replication were collected on the fifth, tenth and fifteenth days post anthesis (DPA), immediately frozen in liquid nitrogen and stored at –80oC until RNA extraction. The remaining three plants per replication were grown to full maturity and the grains were harvested for starch granule analysis.
2.2 Starch isolation and granule size measurement
Five grams of wheat grains per replication were milled with micro–blender, mixed with 5 ml ddH2O and the resulting paste was kept in plastic tubes for 2 hours prior to protein washing. Proteins were dissociated by kneading the paste with hands using distilled water until all starch was washed out. The outwashes were filtered through nylon screen with a pore size of 75 µm by centrifugation at 1000xg for 10 min. The starch samples were desiccated in a thermostat at
Genotype Origin A-type granule size Mean ± SEa (µm) Homogeneous groupsb
C1 Dream/Pesma 23.65 ± 0.159 A
C4 Zentos/Z-296 24.31 ± 0.155 B
C3 Maverich/Lu9-396 24.43 ± 0.161 B
C2 Flair/Bill 24.96 ± 0.142 C
A5 Pegasos/Residence 25.57 ± 0.169 D
A4 Dirigent/Cortez DH 25.62 ± 0.125 D
A2 Pegasos/Biscay DH 25.97 ± 0.129 DE
A1 Flair/Lut 9-329 DH 26.23 ± 0.189 E
A3 Pegasos/Lut 96-6 26.85 ± 0.164 F
Table 1. A–type granule size of 9 winter wheat genotypes used in the study.
aStandard error bHomogeneous groups denoted by the same letter are not significantly different at P<0.01
298
Author’s co
py

R. Armonienė et al.
35°C. Starch granule size was measured under a light microscope (Nicon Eclipse E800). Samples were prepared by mixing 0.01 g of desiccated starch with 1 ml ddH2O. Five random images were photographed from each slide replication and in each image one hundred starch granules were randomly chosen for starch granule diameter measurements. The granule size diameter was measured by Image Pro-Discovery 4.5 software. Analysis of variance (ANOVA) was used to determine significant (P<0.01) differences between genotypes.
2.3 RNA isolation and cDNA synthesisTotal RNA was isolated using RNAqueousTM Phenol–free total RNA Isolation kit (Ambion, Austin, TX, USA). RNA quality and quantification was checked by running 5 µl of the total RNA on a 1% agarose gel. Poly(A)+RNA was isolated from the total RNA with Dynabeads mRNA Purification Kit (Dynal Biotech ASA, Oslo, Norway). Poly(A)+RNA was reverse transcribed with an anchored oligo–dT18 primer using RevertAidTM H Minus First Strand cDNA Synthesis Kit (Fermentas, Vilnius, Lithuania) according to the manufacturer’s protocol. Twenty microliters of reverse transcription reaction product were immediately used in a second strand cDNA synthesis reaction by adding 30 U of DNA Polymerase I (Fermentas), incubating for 2 h at 15°C. The double–stranded cDNA was purified by adding an equal volume of phenol:chloroform:isoamyl alcohol (25:24:1), washed with ethanol, resuspended in a final volume of 35 µl nuclease–free ddH2O and quantified by running 8 µl of cDNA on 1% agarose gel.
2.4 cDNA-AFLP analysisThe cDNA-AFLP procedure was conducted as described earlier by Jonavičienė et al. [18] with minor modifications. PCR mix in selective amplification contained 10 µl of 20x diluted pre–amplification product, 0.25 µM of AseI selective primer with two selective nucleotides, 1.5 µM of TaqI selective primer with two selective nucleotides, 2 mM MgCl2, 0.2 mM dNTP mix, and 0.4 U Taq polymerase in a 20 µl of total reaction volume. The sequences of primers used for AFLP analysis are indicated in Table 2. PCR amplification was conducted in a Mastercycler gradient thermo cycler (Eppendorf, Hamburg, Germany) using the following program: 12 cycles: 94°C, 30 s; 65°C (–0.7°C/cycle), 30 s; 72°C, 1 min and 24 cycles: 94°C, 30 s; 56°C, 30 s; 72°C, 1 min; followed by a final extension step of 10 min at 72°C. The selective PCR products were fractionated in a 1.5% agarose gel electrophoresis at 100 W for 2 h. The fragments were visualized by using ethidium bromide and UV light.
2.5 Identification and cloning of TDFsThe final PCR products of genotype A3 and genotype C1 at 10 and 15 DPA were individually loaded next to each other on the gel for the side-by-side comparisons. The reproducibility of transcript-derived fragments (TDFs) was tested by repeating PCR amplifications 2 to 3 times for each primer combination used. Only robust and reproducible bands were included in the analysis. TDFs were excised from the agarose gel with a sterile surgical blade, extracted from agarose with a DNA Extraction Kit (Fermentas) and eluted with 30 µl of sterile water. DNA fragments were cloned into the pJET1.2/blunt Cloning Vector using GeneJetTM PCR Cloning Kit (Fermentas) according to the manufacturer’s instructions. Plasmid DNA was isolated with GeneJetTM Plasmid Miniprep Kit (Fermentas) and sent for sequencing at the Sequencing Centre (Institute of Biotechnology, Vilnius, Lithuania).
2.6 Primer design for semi quantitative PCRDNA sequences of TDFs were clustered and assembled with the software ChromasPro 1.5 (Technelysium Pty Ltd, Brisban, Australia). The assembled sequences were blasted against the nr protein database of Genbank with BLASTX. PCR primers were designed using PrimerBlast (http://www.ncbi.nlm.nih.gov). The primer pairs were chosen to amplify fragments of approximately 200 bp in length. The well–known starch biosynthesis genes SSI, SSII, SSIII, SBEI, SBEII and GBSSI were amplified as described by Hurkman et al. [19] while the housekeeping control ADP–ribosylation
No. Primer pairs Sequences (5′→3′)
1 OLIGO46/A13 OLIGO66/T13
5’- GACTGCGTACCTAATAG-3’ 5’- GATGAGTCCTGACCGAAG-3’
2 OLIGO47/A14OLIGO67/T14
5’- GACTGCGTACCTAATAT-3’5’- GATGAGTCCTGACCGAAT-3’
3 OLIGO44/A11OLIGO66/T13
5’- GACTGCGTACCTAATAA-3’ 5’- GATGAGTCCTGACCGAAG-3’
4 OLIGO44/A11OLIGO68/T15
5’- GACTGCGTACCTAATAA-3’5’- GATGAGTCCTGACCGACA-3’
5 OLIGO44/A11OLIGO73/T20
5’- GACTGCGTACCTAATAA-3’5’- GATGAGTCCTGACCGAGC-3’
6 OLIGO46/A13OLIGO74/T21
5’- GACTGCGTACCTAATAG-3’5’- GATGAGTCCTGACCGAGG-3’
7 OLIGO56/A23OLIGO76/T23
5’- GACTGCGTACCTAATTA-3’5’- GATGAGTCCTGACCGATA-3’
8 OLIGO-A121 OLIGO64/T11
5’- GACTGCGTACCTAATACA -3’5’- GATGAGTCCTGACCGAAA-3’
9 OLIGO-A123OLIGO76/T23
5’- GACTGCGTACCTAATACC-3’5’- GATGAGTCCTGACCGATA-3’
10 OLIGO-A124OLIGO64/T11
5’- GACTGCGTACCTAATACG-3’5’- GATGAGTCCTGACCGAAA-3’
Table 2. cDNA–AFLP primer sequences used for differential gene expression.
299
Author’s co
py

Isolation of differentially expressed genes in wheat caryopses with contrasting starch granule size
factor gene (Ta2991) was amplified using the primers developed by Paolacci et al. [20].
2.7 Semi quantitative PCRGene–specific PCR amplification was performed in a Dream Taq buffer (includes 20 mM MgCl2) (Fermentas, Lithuania) in a total volume of 10 µl. The PCR solution contained 10 mM dNTP mix, 10 µM of each forward and reverse primer, 0.25 U Dream Taq polymerase (Fermentas) and 20 ng of cDNA. The PCR amplification was conducted in a Mastercycler gradient thermo cycler (Eppendorf, Hamburg, Germany) as follows: initial denaturation of 2 min at 95ºC, followed by 30 cycles of 30 s at 95ºC, 30 s at 55oC and the 1 min elongation step at 72ºC, then followed by 5 min at 72°C for a final extension. The gene–specific PCR products were analyzed in 1.5% agarose gels with ethidium bromide following standard procedures. The gene expression rate of the target gene versus the reference gene Ta2991 was determined with gel image analysis software GelQuant
(MiniBIS Pro, Israel). Target gene expression of each wheat genotype at each time point was normalized to Ta2291 gene expression level to obtain relative gene expression.
3. Results3.1 Size distribution of starch granulesNine LRCAF–bred winter wheat breeding lines were analyzed for A–type starch granule size. The mean size of granules ranged from 23.65 µm to 26.85 µm in diameter for wheat lines C1 and A3, respectively (Table 1). Analysis of variance (ANOVA) was performed for a total of 13,500 measured granules in order to test the significance of variation among accessions. Most of the lines had little variation in granule size and were grouped into six homogeneous groups significantly different at P<0.01. The most contrasting wheat lines C1 and A3 were chosen for further gene expression analysis.
Table 3. Functional annotation of the 12 TDFs revealing significant (E<1e-10) sequence similarities in BLASTX search against the non-redundant (nr) protein database of GenBank along with the TDF size and accession numbers in GenBank protein database.
a Expect (E) value describes the probability of the alignment occurring by chance.
TDF number
Derived from
genotype
Size (bp)
Top BLASTX hit in GenBank E valuea Homologous protein Function Species
TDF01 C1 425 gi׀231540׀ 7E-173 endosperm-specific beta-amylase1
affection of the enzymatic activity levels and thermostabilities of b-amylases
S. cereale
TDF02 A3 772 gi׀38637314׀ 0.0 GTP-binding proteinregulation of a wide range of processes in the cell, including growth
O. sativa
TDF03 C1 855 gi׀60686892׀ 1E-139delta 1-pyrroline-5-carboxylate synthetase
a key role in proline biosynthesis T. aestivum
TDF04 A3 843 gi׀397632׀ 0.0 translation initiation factor 4A
acting in the initiation phase of eukaryotic translation T. aestivum
TDF05 C1 801 gi׀76556513׀ 3E-171 transposase-related protein w-gary1
transposase-related protein with unknown function T. aestivum
TDF06 A3 1491 gi׀357164562׀ 0.0 polyadenilate-binding protein 2-like
involvment in post-transcriptional gene expression processes B. distachyon
TDF07 A3 840 gi׀56606827׀ 0.0 calreticulin–like protein
Ca2+-storage protein in the endoplasmic reticulum T. aestivum
TDF08 A3 1005 gi׀226528152׀ 2E-176 calnexin precursorinteraction with newly synthesized glycoproteins in the endoplasmic reticulum
Z. mays
TDF09 C1 801 gi׀357118112׀ 3E-70 predicted protein unknown function B. distachyon
TDF12 C1 192 gi׀22296816׀ 5E-34protein phosphatase 2A regulatory A subunit
regulation of auxin transport L. perenne
TDF14 A3 582 gi׀224589269׀ 2E-127 Serpin 3 (SER3) inhibition of the function of SERine proteases T. aestivum
TDF18 A3 678 gi׀164472656׀ 6E-149calcium-dependent protein kinase 3-like isoform (CDPK)
control of plant development through the important roles in the regulation of Ca2+
B. distachyon
300
Author’s co
py
R. Armonienė et al.
3.2 Isolation of differentially expressed fragments
cDNA-AFLP analysis was performed using 29 primer pair combinations for the genotypes A3 and C1 wheat caryopses collected at 10 and 15 DPA and revealed 384 transcript-derived fragments (TDFs) with an average of 13 fragments per primer pair. The size of these fragments ranged from approx. 200 to 2000 bp. Of these, 18 TDFs (4.7%) were identified as being differentially expressed (presence/absence) in genotypes A3 or C1. The expression pattern included 12 (66.7%) transcript-derived fragments (TDFs) of genotype A3, whereas 6 (33.3%) TDFs were present in genotype C1. Eighteen TDFs were extracted from agarose gels, cloned and sequenced. A total of twelve TDFs revealed significant (E value<1e-10) sequence similarities in a BLASTX search against the non-redundant (nr) protein database of GenBank (Table 3). These sequences were deposited to the GenBank EST database with accession numbers of KC175594 to KC175605.
3.3 Expression patterns of starch biosynthesis genes
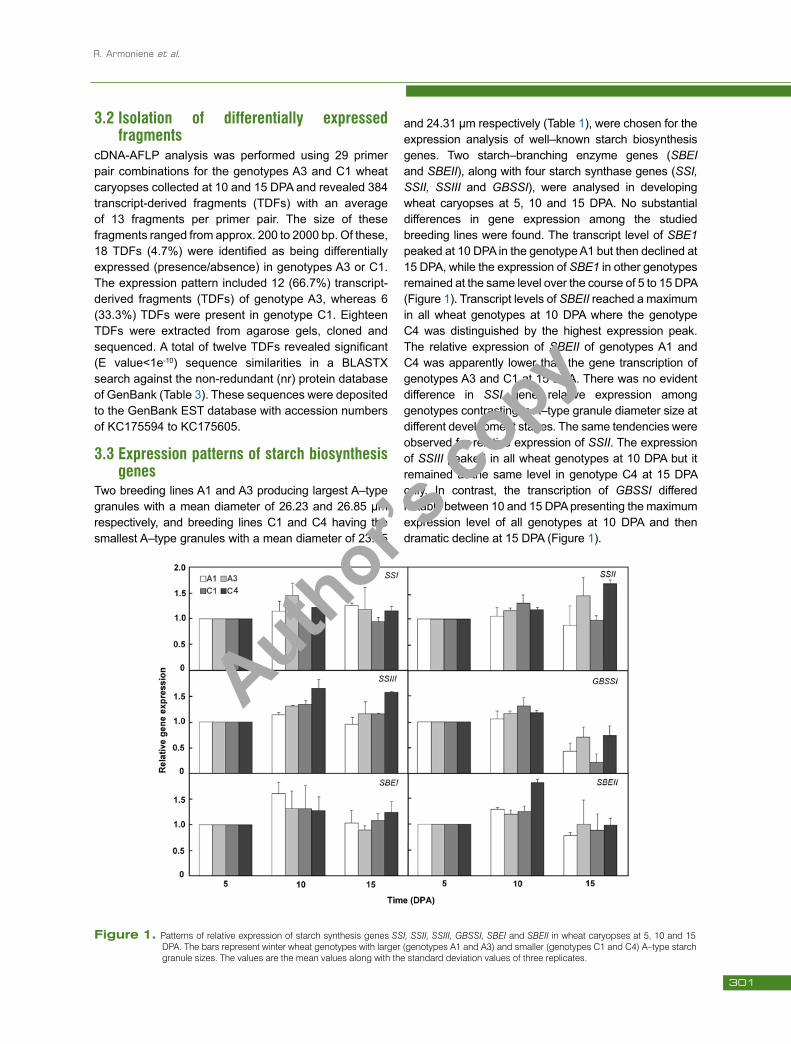
Two breeding lines A1 and A3 producing largest A–type granules with a mean diameter of 26.23 and 26.85 µm respectively, and breeding lines C1 and C4 having the smallest A–type granules with a mean diameter of 23.65
and 24.31 µm respectively (Table 1), were chosen for the expression analysis of well–known starch biosynthesis genes. Two starch–branching enzyme genes (SBEI and SBEII), along with four starch synthase genes (SSI, SSII, SSIII and GBSSI), were analysed in developing wheat caryopses at 5, 10 and 15 DPA. No substantial differences in gene expression among the studied breeding lines were found. The transcript level of SBE1 peaked at 10 DPA in the genotype A1 but then declined at 15 DPA, while the expression of SBE1 in other genotypes remained at the same level over the course of 5 to 15 DPA (Figure 1). Transcript levels of SBEII reached a maximum in all wheat genotypes at 10 DPA where the genotype C4 was distinguished by the highest expression peak. The relative expression of SBEII of genotypes A1 and C4 was apparently lower than the gene transcription of genotypes A3 and C1 at 15 DPA. There was no evident difference in SSI gene relative expression among genotypes contrasting in A–type granule diameter size at different development stages. The same tendencies were observed for relative expression of SSII. The expression of SSIII peaked in all wheat genotypes at 10 DPA but it remained at the same level in genotype C4 at 15 DPA only. In contrast, the transcription of GBSSI differed notably between 10 and 15 DPA presenting the maximum expression level of all genotypes at 10 DPA and then dramatic decline at 15 DPA (Figure 1).
Figure 1. Patterns of relative expression of starch synthesis genes SSI, SSII, SSIII, GBSSI, SBEI and SBEII in wheat caryopses at 5, 10 and 15 DPA. The bars represent winter wheat genotypes with larger (genotypes A1 and A3) and smaller (genotypes C1 and C4) A–type starch granule sizes. The values are the mean values along with the standard deviation values of three replicates.
301
Author’s co
py
Isolation of differentially expressed genes in wheat caryopses with contrasting starch granule size
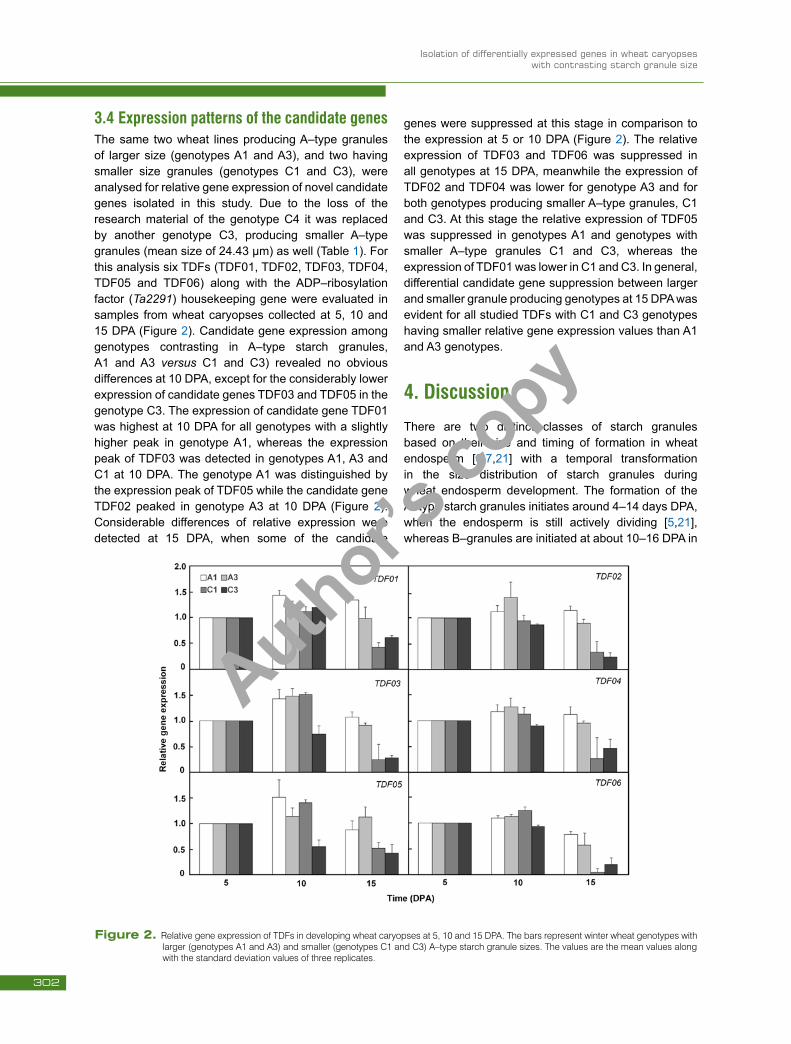
3.4 Expression patterns of the candidate genes The same two wheat lines producing A–type granules of larger size (genotypes A1 and A3), and two having smaller size granules (genotypes C1 and C3), were analysed for relative gene expression of novel candidate genes isolated in this study. Due to the loss of the research material of the genotype C4 it was replaced by another genotype C3, producing smaller A–type granules (mean size of 24.43 µm) as well (Table 1). For this analysis six TDFs (TDF01, TDF02, TDF03, TDF04, TDF05 and TDF06) along with the ADP–ribosylation factor (Ta2291) housekeeping gene were evaluated in samples from wheat caryopses collected at 5, 10 and 15 DPA (Figure 2). Candidate gene expression among genotypes contrasting in A–type starch granules, A1 and A3 versus C1 and C3) revealed no obvious differences at 10 DPA, except for the considerably lower expression of candidate genes TDF03 and TDF05 in the genotype C3. The expression of candidate gene TDF01 was highest at 10 DPA for all genotypes with a slightly higher peak in genotype A1, whereas the expression peak of TDF03 was detected in genotypes A1, A3 and C1 at 10 DPA. The genotype A1 was distinguished by the expression peak of TDF05 while the candidate gene TDF02 peaked in genotype A3 at 10 DPA (Figure 2). Considerable differences of relative expression were detected at 15 DPA, when some of the candidate
genes were suppressed at this stage in comparison to the expression at 5 or 10 DPA (Figure 2). The relative expression of TDF03 and TDF06 was suppressed in all genotypes at 15 DPA, meanwhile the expression of TDF02 and TDF04 was lower for genotype A3 and for both genotypes producing smaller A–type granules, C1 and C3. At this stage the relative expression of TDF05 was suppressed in genotypes A1 and genotypes with smaller A–type granules C1 and C3, whereas the expression of TDF01 was lower in C1 and C3. In general, differential candidate gene suppression between larger and smaller granule producing genotypes at 15 DPA was evident for all studied TDFs with C1 and C3 genotypes having smaller relative gene expression values than A1 and A3 genotypes.
4. DiscussionThere are two distinct classes of starch granules based on their size and timing of formation in wheat endosperm [6,7,21] with a temporal transformation in the size distribution of starch granules during wheat endosperm development. The formation of the A–type starch granules initiates around 4–14 days DPA, when the endosperm is still actively dividing [5,21], whereas B–granules are initiated at about 10–16 DPA in
Figure 2. Relative gene expression of TDFs in developing wheat caryopses at 5, 10 and 15 DPA. The bars represent winter wheat genotypes with larger (genotypes A1 and A3) and smaller (genotypes C1 and C3) A–type starch granule sizes. The values are the mean values along with the standard deviation values of three replicates.
302
Author’s co
py

R. Armonienė et al.
stromules [22]. As formation of A- and B–type granules are competing processes, the early to middle caryopsis developmental stages (10 and 15 DPA) spanning initiation periods of both A– and B–type granules were chosen for isolation of genes involved in granule size determination.
Grain weight is determined by the productivity of starch yield, and the yield management should reflect differences in source–sink relations for starch deposition during grain–filling rates [23]. Sink activity, including the activity of key enzymes involved in carbohydrate metabolism, is physiological restrain in accumulation of carbohydrate reserves during grain filling [24]. Starch synthases SS and GBSS play important role in regulating starch synthesis in grain endosperm and in determining individual grain weight in wheat [25,26]. The first evidence suggesting the possible involvement of starch synthases in the priming of starch biosynthesis came from an analysis of A. thaliana [17]. Further studies [27] have shown that the simultaneous elimination of SSIV and SSIII completely prevents the synthesis of starch in A. thaliana leaves. These results raised the question of the SSIII role in the priming of starch synthesis. In the absence of SSIV, SSIII seemed to be responsible for the initiation of the single granule per chloroplast and also it could interact with other starch synthases and branching enzymes in a phosphorylation–dependent manner [27]. However, studies using A. thaliana provide us knowledge about starch biosynthesis in leaves [28,29], however, the initiation of starch synthesis in wheat grain still remains unclear. The present study aimed at finding a possible involvement of starch synthases in the determination of different A–type granule sizes. As shown in previous studies, which focused on temporal or spatial patterns of starch synthesis gene expression [30-32], the expression of our studied starch biosynthesis genes markedly increased at the early stage of grain filling (from 5 to 10 DPA) and remained at the same level at 15 DPA. Even though it was confirmed that changes in starch synthases (SS and GBSS) activity is closely related to the starch granule size distribution during grain filling [7] no obvious evidence for differences in relative gene expression between genotypes contrasting in A–type granule size was found (Figure 1). These findings support the model of these genes playing important role mainly in starch biosynthesis but not in the determination of A–type granule size at the early stages of caryopses development.
Twelve genes identified in this study to be putatively involved in starch granule biosynthesis showed homology to known proteins described in the model plant Brachypodium distachyon and other Poaceae species such as wheat, rye, rice, maize and perennial ryegrass
(Table 3). These genes are known to play a role in the regulation of gene expression and signal transduction in plant development. One of our identified TDFs – TDF01 was predicted to encode the endosperm–specific beta–amylase 1, which was shown to be involved in both the synthesis and degradation of starch [33-35]. The changes in transcriptional expression of isoamylases were previously reported to correlate with the ratio of B– to A–type granules in terms of their volume and number [31]. Moreover, Burton et al. [11] and Bustos et al. [12] both proposed that isoamylases could be a major determinant of granule initiation because of the reduction in granule size and the increase in their number in the isa1 (isoamylase 1) mutants of barley and potato, respectively. Furthermore, A. thaliana mutants lacking all four forms of isoamylase were shown to be devoid of starch granules [14]. The guanosine triphosphate (GTP)-binding proteins, in our study encoded by TDF02, were recently detected in the amyloplast membrane–bound protein of wheat endosperm at 10 DPA [36], thereby playing the role in signal–transduction cascades of starch accumulation. The increase in relative expression of TDF04, encoding translation initiation factor 4A, is consistent with the earlier work of Stamova et al. [31], who detected a correlation between the percentage in number of A–granules and expression profiles of genes encoding several transcription factors. The polyadenilate–binding proteins participate in post–transcriptional mRNA processing [37] while five genes encoding calcium dependent protein kinases (CDPKs) were up–regulated in the endosperm stage in rice [38]. The temporal patterns of expression of these candidate genes were found to be consistent with their role in starch biosynthesis, with higher peaks at 15 DPA in the genotypes A1 and A3, containing larger A–type starch granules, rather in genotypes C1 and C4 producing smaller granules (Figure 2). The lower expression level of candidate genes in genotypes producing smaller granules might be associated with the onset of B–type granule development at 15 DPA, as it has been reported earlier [7,31]. The results of our study suggest a putative role for the identified candidate genes in the determination of starch granule size. Among eighteen starch biosynthesis genes, identified in the wheat genotypes differentiating in A–type starch granules size, five of them (TDF10, TDF11, TDF15, TDF16, TDF17) showed no homology to known proteins in other species. These findings suggest identification of the novel genes involved in mediation of starch biosynthesis and need further validation.
In conclusion, the studied genes were identified as the putative targets for starch A–type granule development regulation in winter wheat. However, the
303
Author’s co
py

Isolation of differentially expressed genes in wheat caryopses with contrasting starch granule size
molecular mechanism of the granule initiation remains to be clarified. The genomic information obtained from the identified differentially expressed genes provides clues for the development of functional markers for selection of genotypes producing larger A–type starch granule in winter wheat. Moreover, this genomic data could be used further to validate the described starch and as yet unknown biosynthesis genes, thus providing a first step towards winter wheat cultivars with enhanced starch content.
AcknowledgementsThis work was sponsored via the long–term research programme ‘Genetics and directional transformation of genotypes in agricultural and forest plants’ implemented by the Lithuanian Research Centre for Agriculture and Forestry.
[1] Renard D., Della Valle G., Popineau Y., Plant biopolymer science: food and non-food applications, The Royal Society of Chemistry, Cambridge, 2002
[2] Röper H., Renewable raw materials in Europe – industrial utilization of starch and sugar, Starch/Stärke, 2002, 54, 89-99
[3] Smith A.M., Prospects for increasing starch and sucrose yields for bioethanol production, Plant J., 2008, 54, 546-558
[4] Ball S.G., Morell M.K., From bacterial glycogen to starch: understanding the biogenesis of the plant starch granule, Ann. Rev. Plant Biol., 2003, 54, 207-235
[5] Wilson J.D., Bechtel D.B., Todd T.C., Seib P.A., Measurement of wheat starch granule size distribution using image analysis and laser diffraction technology, Cereal Chem., 2006, 83, 259-268
[6] Bechtel D.B., Wilson J.D., Amyloplast formation and starch granule development in hard red winter wheat, Cereal Chem., 2003, 80, 175-183
[7] Zhang C., Jiang D., Liu F., Cai J., Dai T., Cao W., Starch granules size distribution in superior and inferior grains of wheat is related to enzyme activities and their gene expressions during grain filling, J. Cereal Sci. 2010, 51, 226-233
[8] Shinde S.V., Nelson J.E., Huber K.C., Soft wheat starch pasting behavior in relation to A- and B-type granule content and composition, Cereal Chem., 2003, 80, 91-98
[9] Jane J., Yoo S., Wong K., Duvick S.A., Gardner C.A., Structures of amylopectin and starch granules – how are they synthesized?, J. Appl. Glycosci., 2003, 50, 167–172
[10] Park S.H., Chung O.K., Seib P.A., Effects of varying weight ratios of large and small wheat starch granules on experimental straight-dough bread, Cereal Chem., 2005, 82, 166-172
[11] Burton R.A., Jenner H., Carrangis L., Fahy B., Fincher G.B., Hylton C., et al., Starch granule initiation and growth are altered in barley mutants that lack isoamylase activity, Plant J., 2002, 31, 97-112
[12] Bustos R., Fahy B., Hylton C.M., Seale R., Nebane N.M., Edwards A., et al., Starch granule initiation is controlled by a heteromultimeric isoamylase in potato tubers, Proc. Natl. Acad. Sci. USA, 2004, 101, 2215-2220
[13] Delatte T., Trevisan M., Parker M.L., Zeeman S.C., Arabidopsis mutants Atisa1 and Atisa2 have identical phenotypes and lack the same multimeric isoamylase, which influences the branch point distribution of amylopectin during starch synthesis, Plant J., 2005, 41, 815-830
[14] Streb S., Delatte T., Umhang M., Eicke S., Schorderet M., Reinhardt D., et al., Starch granule biosynthesis in Arabidopsis is abolished by removal of all debranching enzymes but restored by the subsequent removal of an endoamylase, Plant Cell, 2008, 20, 3448-3466
[15] Wattebled F., Dong Y., Dumez S., Delvallé D., Planchot V., Berbezy P., et al., Mutants of Arabidopsis lacking a chloroplastic isoamylase accumulate phytoglycogen and an abnormal form of amylopectin, Plant Physiol., 2005, 138, 184-195
[16] Stahl Y., Coates S., Bryce J.H., Morris P.C., Antisense downregulation of the barley limit dextrinase inhibitor modulates starch granule size distribution, starch composition and amylopectin structure, Plant J., 2004, 39, 599-611
[17] Roldán I., Wattebled F., Mercedes Lucas M., Delvallé D., Planchot V., Jimenéz S., et al., The phenotype of soluble starch synthase IV defective mutants of Arabidopsis thaliana suggests a novel function of elongation enzymes in the control of starch granule formation, Plant J., 2007, 49, 492-504
References
304
Author’s co
py

R. Armonienė et al.
[18] Jonavičienė K., Studer B., Asp T., Jensen L.B., Paplauskienė V., Lazauskas S., et al., Identification of genes involved in a water stress response in timothy and mapping of orthologous loci in perennial ryegrass, Biol. Plantarum, 2012, 56, 473-483
[19] Hurkman W.J., McCue K.F., Altenbach S.B., Korn A., Tanaka C.K., Kothari K.M., et al., Effect of temperature on expression of genes encoding enzymes for starch biosynthesis in developing wheat endosperm, Plant Sci., 2003, 164, 873-881
[20] Paolacci A.R., Tanzarella O.A., Porceddu E., Ciaffi M., Identification and validation of reference genes for quantitative RT-PCR normalization in wheat, BMC Mol. Biol., 2009, 10, 11
[21] Parker M.L., The relationship between A-type and B-type starch granules in the developing endosperm of wheat, J. Cereal Sci., 1985, 3, 271-278
[22] Langeveld S.M.J., van Wijk R., Stuurman N., Kijne J.W., de Pater S., B-type granule containing protrusions and interconnections between amyloplasts in developing wheat endosperm revealed by transmission electron microscopy and GFP expression, J. Exp. Bot. 2000, 51, 1357-1361
[23] Jenner C.F., Ugalde T.D., Aspinall D., The physiology of starch and protein deposition in the endosperm of wheat, Funct. Plant Biol., 1991, 18, 211-226
[24] Kubo A., Fujita N., Harada K., Matsuda T., Satoh H., Nakamura Y., The starch-debranching enzymes isoamylase and pullulanase are both involved in amylopectin biosynthesis in rice endosperm, Plant Physiol., 1999, 121, 399-410
[25] Jiang D., Cao W., Dai T., Jing Q., Activities of key enzymes for starch synthesis in relation to growth of superior and inferior grains on winter wheat (Triticum aestivum L.) spike, Plant Growth Regul., 2003, 41, 247-257
[26] Yang J., Zhang J., Wang Z., Xu G., Zhu Q., Activities of key enzymes in sucrose-to-starch conversion in wheat grains subjected to water deficit during grain filling, Plant Physiol., 2004, 135, 1621-1629
[27] Szydlowski N., Ragel P., Raynaud S., Lucas M.M., Roldan I., Montero M., et al., Starch granule initiation in Arabidopsis requires the presence of either class IV or class III starch synthases, Plant Cell, 2009, 21, 2443-2457
[28] Zeeman S.C., Kossmann J., Smith A.M., Starch: its metabolism, evolution, and biotechnological modification in plants, Annu. Rev. Plant Biol., 2010, 61, 209-234
[29] D’Hulst C., Mérida Á., The priming of storage glucan synthesis from bacteria to plants: current knowledge and new developments, New Phytol., 2010, 188, 13-21
[30] Shewry P.R., Underwood C., Wan Y., Lovegrove A., Bhandari D., Toole G., et al., Storage product synthesis and accumulation in developing grains of wheat, J. Cereal Sci. 2009, 50, 106-112
[31] Stamova B.S., Laudencia-Chingcuanco D., Beckles D.M., Transcriptomic analysis of starch biosynthesis in the developing grain of hexaploid wheat, Int. J. Plant Genom., 2009, 2009, 23
[32] Wang Z., Li W., Qi J., Shi P., Yin Y., Starch accumulation, activities of key enzyme and gene expression in starch synthesis of wheat endosperm with different starch contents, J. Food Sci. Technol., 2011, DOI: 10.1007/s13197-011-0520-z
[33] James M.G., Robertson D.S., Myers A.M., Characterization of the maize gene sugary1, a determinant of starch composition in kernels, Plant Cell, 1995, 7, 417-429
[34] Nakamura Y., Umemoto T., Takahata Y., Komae K., Amano E., Satoh H., Changes in structure of starch and enzyme activities affected by sugary mutations in developing rice endosperm. Possible role of starch debranching enzyme (R-enzyme) in amylopectin biosynthesis, Physiol. Plantarum, 1996, 97, 491-498
[35] Wattebled F., Planchot V., Dong Y., Szydlowski N., Pontoire B., Devin A., et al., Further evidence for the mandatory nature of polysaccharide debranching for the aggregation of semicrystalline starch and for overlapping functions of debranching enzymes in Arabidopsis leaves, Plant Physiol., 2008, 148, 1309-1323
[36] Balmer Y., Vensel W.H., DuPont F.M., Buchanan B.B., Hurkman W.J., Proteome of amyloplasts isolated from developing wheat endosperm presents evidence of broad metabolic capability, J. Exp. Bot., 2006, 57, 1591-1602
[37] Wahle E., Ruegsegger U., 3’-End processing of pre-mRNA in eukaryotes, FEMS Microbiol. Rev., 1999, 23, 277-295
[38] Ye S., Wang L., Xie W., Wan B., Li X., Lin Y., Expression profile of calcium-dependent protein kinase (CDPKs) genes during the whole lifespan and under phytohormone treatment conditions in rice (Oryza sativa L. ssp. indica), Plant Mol. Biol., 2009, 70, 311-325
305
Author’s co
py




















