
Indolocarbazole antitumour compounds by combinatorial biosynthesis
9
Available online at www.sciencedirect.com Indolocarbazole antitumour compounds by combinatorial biosynthesis Jose ´ A Salas and Carmen Me ´ ndez The indolocarbazoles constitute a family of natural products with potential therapeutic applications in the treatment of cancer and neurodegenerative disorders. Members of this family are either potent stabilizers of topoisomerase I-DNA covalent complex or potent inhibitors of protein kinases. Rebeccamycin, staurosporine, AT2433 and K252a are members of this family, which are produced by different actinomycete strains, and whose biosynthesis gene clusters have been isolated and characterized. A number of indolocarbazole derivatives have been generated by applying combinatorial biosynthesis technologies to these clusters either in the producer strain or in the heterologous hosts after expression of part or the entire gene cluster. Combinatorial biosynthesis is therefore providing a new approach for increasing structural diversity in this family of natural products. Addresses Departamento de Biologı´a Funcional and Instituto Universitario de Oncologı´a del Principado de Asturias (I.U.O.P.A), Universidad de Oviedo, 33006 Oviedo, Spain Corresponding author: Salas, Jose ´A([email protected]) Current Opinion in Chemical Biology 2009, 13:152–160 This review comes from a themed issue on Molecular Diversity Edited by Christian Hertweck and Hiroyuki Osada Available online 27th February 2009 1367-5931/$ – see front matter # 2009 Elsevier Ltd. All rights reserved. DOI 10.1016/j.cbpa.2009.02.003 Introduction The indolocarbazole alkaloids constitute an important class of natural products, which have been isolated from actinomycetes, cyanobacteria, fungi, slime moulds and marine invertebrates [1 ]. After the isolation of the first indolocarbazole in 1977, this family of compounds has attracted the attention of many researchers from different disciplines because of the variety of chemical structures and the interesting biological activities showed by this family of compounds. Structurally (Figure 1a), the mem- bers of this family are characterized by a core consisting of either an ‘open’ bisindolylmaleimide (e.g. arcyriarubin A), or a ‘closed’ indolo[2,3-a]carbazole (e.g. rebeccamy- cin, staurosporine). Most of the latter compounds are, in fact, derivatives of the indolo[2,3-a]pyrrolo[3,4-c]carba- zole ring system, to which a sugar residue is often attached, but they differ in the way the sugar is attached to the indolocarbazole moiety: through one (as in rebec- camycin and AT2433) or two (as in staurosporine and K-252a) N-glycosidic bonds. Indolocarbazoles display a wide range of biological activi- ties, including antibacterial, antifungal, antiviral, hypo- tensive, antitumour or neuroprotective properties, but their greatest interest is based on their antitumour proper- ties [2,3], and great efforts are made to generate indolo- carbazole derivatives with improved properties for the treatment of cancer. Some indolocarbazole derivatives have entered into clinical trials for cancer and also for the treatment of neurodegenerative disorders, and dia- betes-associated pathologies [4 ]. When considering the mode of action, indolocarbazoles can be subdivided into two different subgroups. The first subgroup is represented by rebeccamycin that is a potent stabilizer of topoisomerase I-DNA covalent complex. The second one, represented by staurosporine, includes potent inhibi- tors of protein kinases A, C and K. This suggests that the structural differences between these two indolocarba- zoles are essential for target selectivity. A number of indolocarbazole derivatives have been pro- duced by chemical synthesis or semi-synthesis [5,6]. However, in the past few years the isolation and charac- terization of biosynthesis gene clusters for several indo- locarbazoles produced by actinomycetes offers a promising alternative for increasing structural diversity in this family of compounds through the use of combi- natorial biosynthesis technology. This article revises the gene clusters characterized, the enzymatic characteriz- ation of different steps in indolocarbazole biosynthesis and how combinatorial biosynthesis is being used as a tool for diversifying and expanding the chemical space in this family of natural products. Biosynthetic origin of indolocarbazoles Early studies on the biosynthetic origin of indolocarbazoles shed light on the biosynthetic pathways through the identi- fication of metabolic precursors and some biosynthesis intermediates. Furthermore, the flexibility of the biosyn- thetic machinery was tested for accepting alternative pre- cursors, and several new analogues were generated. The biosyntheses of staurosporine and rebeccamycin were studied by feeding isotope-labeled precursors to their producer organisms: Lentzea albida (formerly Streptomyces staurosporeus) staurosporine producer and Lechevalieria aerocolonigenes (formerly Saccharothrix aerocolonigenes) rebeccamycin producer [7–10]. It was established that Current Opinion in Chemical Biology 2009, 13:152–160 www.sciencedirect.com
Transcript of Indolocarbazole antitumour compounds by combinatorial biosynthesis

Available online at www.sciencedirect.com
Indolocarbazole antitumour compounds by combinatorialbiosynthesisJose A Salas and Carmen Mendez
The indolocarbazoles constitute a family of natural products
with potential therapeutic applications in the treatment of
cancer and neurodegenerative disorders. Members of this
family are either potent stabilizers of topoisomerase I-DNA
covalent complex or potent inhibitors of protein kinases.
Rebeccamycin, staurosporine, AT2433 and K252a are
members of this family, which are produced by different
actinomycete strains, and whose biosynthesis gene clusters
have been isolated and characterized. A number of
indolocarbazole derivatives have been generated by applying
combinatorial biosynthesis technologies to these clusters
either in the producer strain or in the heterologous hosts after
expression of part or the entire gene cluster. Combinatorial
biosynthesis is therefore providing a new approach for
increasing structural diversity in this family of natural products.
Addresses
Departamento de Biologıa Funcional and Instituto Universitario de
Oncologıa del Principado de Asturias (I.U.O.P.A), Universidad de
Oviedo, 33006 Oviedo, Spain
Corresponding author: Salas, Jose A ([email protected])
Current Opinion in Chemical Biology 2009, 13:152–160
This review comes from a themed issue on
Molecular Diversity
Edited by Christian Hertweck and Hiroyuki Osada
Available online 27th February 2009
1367-5931/$ – see front matter
# 2009 Elsevier Ltd. All rights reserved.
DOI 10.1016/j.cbpa.2009.02.003
IntroductionThe indolocarbazole alkaloids constitute an important
class of natural products, which have been isolated from
actinomycetes, cyanobacteria, fungi, slime moulds and
marine invertebrates [1�]. After the isolation of the first
indolocarbazole in 1977, this family of compounds has
attracted the attention of many researchers from different
disciplines because of the variety of chemical structures
and the interesting biological activities showed by this
family of compounds. Structurally (Figure 1a), the mem-
bers of this family are characterized by a core consisting of
either an ‘open’ bisindolylmaleimide (e.g. arcyriarubin
A), or a ‘closed’ indolo[2,3-a]carbazole (e.g. rebeccamy-
cin, staurosporine). Most of the latter compounds are, in
fact, derivatives of the indolo[2,3-a]pyrrolo[3,4-c]carba-
zole ring system, to which a sugar residue is often
Current Opinion in Chemical Biology 2009, 13:152–160
attached, but they differ in the way the sugar is attached
to the indolocarbazole moiety: through one (as in rebec-
camycin and AT2433) or two (as in staurosporine and
K-252a) N-glycosidic bonds.
Indolocarbazoles display a wide range of biological activi-
ties, including antibacterial, antifungal, antiviral, hypo-
tensive, antitumour or neuroprotective properties, but
their greatest interest is based on their antitumour proper-
ties [2,3], and great efforts are made to generate indolo-
carbazole derivatives with improved properties for the
treatment of cancer. Some indolocarbazole derivatives
have entered into clinical trials for cancer and also for
the treatment of neurodegenerative disorders, and dia-
betes-associated pathologies [4�]. When considering the
mode of action, indolocarbazoles can be subdivided into
two different subgroups. The first subgroup is
represented by rebeccamycin that is a potent stabilizer
of topoisomerase I-DNA covalent complex. The second
one, represented by staurosporine, includes potent inhibi-
tors of protein kinases A, C and K. This suggests that the
structural differences between these two indolocarba-
zoles are essential for target selectivity.
A number of indolocarbazole derivatives have been pro-
duced by chemical synthesis or semi-synthesis [5,6].
However, in the past few years the isolation and charac-
terization of biosynthesis gene clusters for several indo-
locarbazoles produced by actinomycetes offers a
promising alternative for increasing structural diversity
in this family of compounds through the use of combi-
natorial biosynthesis technology. This article revises the
gene clusters characterized, the enzymatic characteriz-
ation of different steps in indolocarbazole biosynthesis
and how combinatorial biosynthesis is being used as a tool
for diversifying and expanding the chemical space in this
family of natural products.
Biosynthetic origin of indolocarbazolesEarly studies on the biosynthetic origin of indolocarbazoles
shed light on the biosynthetic pathways through the identi-
fication of metabolic precursors and some biosynthesis
intermediates. Furthermore, the flexibility of the biosyn-
thetic machinery was tested for accepting alternative pre-
cursors, and several new analogues were generated.
The biosyntheses of staurosporine and rebeccamycin were
studied by feeding isotope-labeled precursors to their
producer organisms: Lentzea albida (formerly Streptomycesstaurosporeus) staurosporine producer and Lechevalieriaaerocolonigenes (formerly Saccharothrix aerocolonigenes)rebeccamycin producer [7–10]. It was established that
www.sciencedirect.com
Indolocarbazole antitumour compounds by combinatorial biosynthesis Salas and Mendez 153
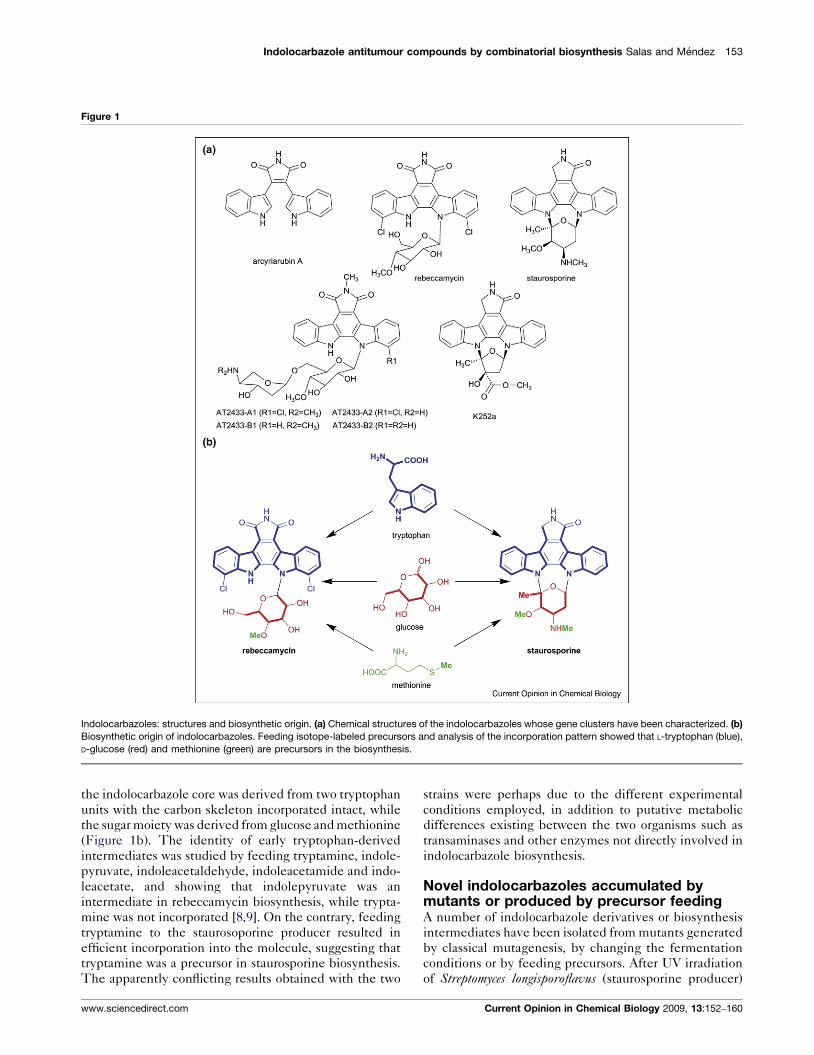
Figure 1
Indolocarbazoles: structures and biosynthetic origin. (a) Chemical structures of the indolocarbazoles whose gene clusters have been characterized. (b)Biosynthetic origin of indolocarbazoles. Feeding isotope-labeled precursors and analysis of the incorporation pattern showed that L-tryptophan (blue),
D-glucose (red) and methionine (green) are precursors in the biosynthesis.
the indolocarbazole core was derived from two tryptophan
units with the carbon skeleton incorporated intact, while
the sugar moiety was derived from glucose and methionine
(Figure 1b). The identity of early tryptophan-derived
intermediates was studied by feeding tryptamine, indole-
pyruvate, indoleacetaldehyde, indoleacetamide and indo-
leacetate, and showing that indolepyruvate was an
intermediate in rebeccamycin biosynthesis, while trypta-
mine was not incorporated [8,9]. On the contrary, feeding
tryptamine to the staurosoporine producer resulted in
efficient incorporation into the molecule, suggesting that
tryptamine was a precursor in staurosporine biosynthesis.
The apparently conflicting results obtained with the two
www.sciencedirect.com
strains were perhaps due to the different experimental
conditions employed, in addition to putative metabolic
differences existing between the two organisms such as
transaminases and other enzymes not directly involved in
indolocarbazole biosynthesis.
Novel indolocarbazoles accumulated bymutants or produced by precursor feedingA number of indolocarbazole derivatives or biosynthesis
intermediates have been isolated from mutants generated
by classical mutagenesis, by changing the fermentation
conditions or by feeding precursors. After UV irradiation
of Streptomyces longisporoflavus (staurosporine producer)
Current Opinion in Chemical Biology 2009, 13:152–160

154 Molecular Diversity
mutants were isolated that accumulated K-252c and 30-O-
demethyl-staurosporine [11,12]. The latter compound
was converted into staurosporine by feeding experiments
and hence it was proposed as the last intermediate in
staurosporine biosynthesis [13]. Addition of potassium
bromide (but not fluoride or iodide salts) to cultures of
L. aerocolonigenes resulted in production of bromorebec-
camycin with concomitant suppression of rebeccamycin
accumulation [14]. On the contrary, addition of 6-fluoro-
tryptophan, 5-fluoro-tryptophan, or 4-fluoro-tryptophan
produced fluoroindolocarbazoles [15,16]. Attempts to per-
form precursor-directed biosynthesis with the staurospor-
ine producer using different indole derivatives (indole-3-
acetonitrile, indole-3-acetic acid, indole-3-acetamide, 5-
fluorotryptophan and 6-fluorotryptophan, 5-hydroxytryp-
tophan, indole-3-pyruvate, indole-3-propionic acid, tryp-
tamine, 5-fluorotryptamine and 6-fluorotryptamine and 5-
hydroxytryptamine) were unsuccessful.
Isolation of indolocarbazole gene clustersThe first report on the cloning of an indolocarbazole
biosynthesis gene was based on an indirect observation.
In the course of some bioconversion studies, it was found
that the aglycone J-104303 (identical to the rebeccamycin
aglycone but possessing hydroxyl groups instead of the
chlorine atoms) was converted into its N-glucosylated
derivative by a rebeccamycin producer strain. From a
gene library of genomic DNA of this producer strain, a
clone was isolated in Streptomyces lividans capable of
carrying out this conversion and, upon sequencing of
the DNA insert, found to contain a gene (designated as
ngt gene) coding for a glycosyltransferase [17�]. These
results suggested that ngt was probably involved in rebec-
camycin biosynthesis in this organism, although no defi-
nite proof was reported at that time. Using the ngt gene as
a probe, the rebeccamycin gene cluster was identified,
characterized and heterologously expressed in Strepto-myces albus [18�]. The same cluster was later on reported
by two different laboratories [19,20�] and expressed in
Escherichia coli, although at low level [20�]. A number of
biosynthetic intermediates in rebeccamycin biosynthesis
were identified either by co-expressing different sets of
genes of the cluster [18�] or by generating insertional
inactivation mutants [19] and further analysis of the
accumulated products. Three other indolocarbazole gene
clusters have been also reported: staurosporine [21�],AT2433 [22] and K-252a [23], together with that of the
closely related violacein [24,25]. The genetic organization
of these four indolocarbazole gene clusters is quite similar
and all of them contain genes required for the biosyn-
thesis of the aglycone, biosynthesis and transfer of the
deoxysugar and regulation (Figure 2a). Interestingly, the
AT2433 cluster is unique in containing an extra gene
AtmA, absent in the other clusters, that has been pro-
posed to participate in the biosynthesis of the aglycon
(see below). Furthermore, the rebeccamycin, AT2433
and K252a contain two genes each coding for transmem-
Current Opinion in Chemical Biology 2009, 13:152–160
brane proteins possibly involved in secretion, but these
genes are missing in the staurosporine cluster.
Pathways for indolocarbazole biosynthesisand characterization of enzymes involvedOn the basis of similarities of the deduced gene products
with proteins in databases, on the chemical structures of
products accumulated by different mutants or produced
by recombinant strains co-expressing different sets of
genes and on in vitro testing of enzyme activities, path-
ways for the biosynthesis of these indolocarbazoles have
been proposed (Figure 2b). Conversion of L-tryptophan to
7-chloro-L-tryptophan is the first step in rebeccamycin
and AT2433 biosynthesis and serves to supply a unique
monomer precursor. This reaction is catalyzed in the
rebeccamycin pathway by a two-component enzyme,
RebF (flavin reductase)-RebH (halogenase), enzymes
that have been characterized in vitro [26–28]. In the
case of the AT2433 cluster, the flavin reductase gene is
absent [22]. Staurosporine and K-252a lack halogen atoms
and therefore these clusters do not contain these two-
component enzymes. A second pair of enzymes is then
responsible for the formation of the chromopyrrolic acid
intermediate through the fusion of two tryptophans or
halogenated tryptophan moieties. This process is cata-
lyzed by the tandem action of two enzymes: an L-trypto-
phan oxidase (RebO, StaO and AtmO in rebeccamycin,
staurosporine and AT2433 biosyntheses, respectively)
and a heme protein dimer with chromopyrrolic acid
synthase activity (RebD, StaD and AtmD for the same
pathways) [29,30�,31,32]. Interestingly, AT2433 is the
only naturally occurring indolocarbazole to have an asym-
metrically halogenated indolocarbazole core. It has been
proposed that AtmO and AtmD might have unique sub-
strate specificities, leading to the final formation of an
asymmetrically halogenated aglycone [22]. In addition, an
extra protein AtmA has been postulated to participate in
the introduction of the nitrogen atom at the pyrrole
moiety [22].
The ‘open’ chromopyrrolic acid intermediate must be
now converted into a ‘closed’ indolocarbazole aglycone
either containing an imide (rebeccamycin-like as in arcyr-
iaflavin A) or an amide (staurosporine-like as in K252c)
function at the pyrrole moiety. This requires an enzyme
pair that is present in all four clusters (RebC/RebP, StaC/
StaP, AtmC/AtmP and InkE/InkP). RebP (or its homol-
ogue StaP) are cytochrome P450s that appear to catalyze
the oxidation of chromopyrrolic acid to a set of three
aglycones (K252c, arcyriaflavin A and 7-hydroxy-K252c),
differing in the oxidation state in the pyrrole-derived ring
[33]. A catalytic mechanism for the StaP enzyme has been
proposed on the basis of the analysis of the crystal
structure of StaP with chromopyrrolic acid [34]. The
second enzyme, RebC (or its homologue StaC) is a
flavin-dependent hydroxylase, whose structure has been
recently elucidated alone as well as in the presence of
www.sciencedirect.com

Indolocarbazole antitumour compounds by combinatorial biosynthesis Salas and Mendez 155
Figure 2
Biosynthesis of indolocarbazoles. (a) Genetic organization of the gene cluster for rebecamycin (reb), staurosporine (sta), K252a (ink) and AT2433
(atm). (b) Proposed pathways for indolocarbazoles biosynthesis. R Cl in rebeccamycin and R H or Cl for intermediates during AT2433-A1
biosynthesis.
chromopyrrolic acid and K252c [35]. The P450 enzymes
(RebP or StaP), in the presence of StaC convert the three
aglycones into a predominant K252c product, while in the
presence of RebC basically arcyriaflavin A is formed [36].
www.sciencedirect.com
It has been proposed that the role of RebC (and that
of StaC) is to sequester a reactive intermediate produced
by its partner protein (RebP or StaP) and to react with
it enzymatically, preventing its conversion to several
Current Opinion in Chemical Biology 2009, 13:152–160

156 Molecular Diversity
degradation products that includes, at low levels, the
desired product. After the formation of the aglycone,
sugar transfer occurs catalyzed by a glycosyltransferase.
Rebeccamycin and staurosporine contain an hexose sugar,
D-glucose and L-ristosamine, respectively, while K-252a
contains a dihydrostreptose and AT2433 an aminodideox-
ypentose and a D-glucose moieties. Genes for the bio-
synthesis of these sugars are part of the gene clusters, with
the exception of rebeccamycin that can incorporate D-
glucose from primary metabolism. Candidate N-glycosyl-
transferase genes for sugar transfer have been found in all
four clusters. In the case of staurosporine and K-252a, a
second C–N bond must be established between the sugar
and the aglycone, a reaction catalyzed by a P450 mono-
oxygenase (StaN for staurosporine and InkC for K-252a).
As final steps in the biosynthesis, several methylation
steps decorate the final indolocarbazole structure acting
on the sugars (in all four cases) or in the imide function (in
the case of AT2433).
Figure 3
Indolocarbazoles produced by combinatorial biosynthesis with modification
Current Opinion in Chemical Biology 2009, 13:152–160
Combinatorial biosynthesis for novelindolocarbazoles: modifications in theaglycone moietyThe availability of the genes for the biosynthesis of the
four indolocarbazoles mentioned above, together with the
relative substrate flexibility of secondary metabolism
enzymes, has made possible the generation of a good
number of novel indolocarbazole derivatives (Figures 3
and 4). One of the more efficient strategies was the use of
combinatorial biosynthesis approaches to create meta-
bolic pathways in another actinomycete host such as S.albus by co-expression of different indolocarbazole bio-
synthesis genes either from a pathway or in combination
with genes from other indolocarbazole pathways, thus
rendering 24 novel indolocarbazoles (Figure 3) [37��].This allowed the dissection and reconstitution of the
rebeccamycin pathway and the understanding of how
this indolocarbazole is synthesized. These studies
showed that rebeccamycin derivatives can be produced
s in the aglycone moiety.
www.sciencedirect.com

Indolocarbazole antitumour compounds by combinatorial biosynthesis Salas and Mendez 157
Figure 4
Indolocarbazoles produced by combinatorial biosynthesis with modifications in the deoxysugar moiety.
www.sciencedirect.com Current Opinion in Chemical Biology 2009, 13:152–160

158 Molecular Diversity
in the absence of the RebH halogenase (Figure 3, com-
pounds 1, 10, 19–21), indicating that a full pathway for
dideschloro-rebeccamycin can be reconstituted and also
showing that the rest of the enzymes of the pathway were
able to use non-halogenated intermediates [37��]. Rebec-
camycin derivatives with chlorine atoms attached to
different positions of the tryptophan moiety were pro-
duced by co-expressing genes involved in the biosyn-
thesis of the rebeccamycin aglycone together with two
genes coding for halogenases acting on different positions
of the tryptophan moiety (Figure 3, compounds 2, 3, 9, 11,
18). These were the PyrH 5-chloro-tryptophan halogen-
ase involved in pyrroindomycin biosynthesis and the
ThaI 6-chloro-tryptophan halogenase involved in thieno-
lodin biosynthesis. These experiments showed that halo-
gen atoms can be positioned in different parts of the
indolocarbazole ring. Another interesting feature for
creating indolocarbazole structural diversity is the pivotal
role of RebC vs. StaC. These two enzymes are respon-
sible for the oxidation state of the indolocarbazole ring:
RebC leading to the formation of an imide function and
StaC to an amide function. This implies that when co-
expressing indolocarbazole genes, just by exchanging
these two genes in any gene construct we can produce
rebeccamycin-like (Figure 3, compounds 14–18, 20–24;
Figure 4, compounds 42, 47, 48, 52) or staurosporine-like
derivatives (Figure 3, compounds 10–13; Figure 4, com-
pounds 25–30) ([37��], AP Salas et al., unpublished data).
Combinatorial biosynthesis for novelindolocarbazoles: modifications in the sugarmoietyIncreasing evidence suggest the existence of some flexi-
bility in secondary metabolite glycosyltransferases
regarding the sugar donor [38�,39,40]. Taking advantage
of this flexibility a number of novel glycosylated deriva-
tives from staurosporine have been generated. The com-
plete staurosporine pathway was reconstituted in S. albususing a two-plasmid system [41��]. The first plasmid
(‘aglycone plasmid’) contained the genes required for
staurosporine aglycone formation together with a gene
coding for the staurosporine glycosyltransferase StaG and
a second gene coding for the cytochrome P450 enzyme
StaN. The latter has been shown to be responsible for the
establishment of the second C–N linkage between the
sugar and the indolocarbazole moiety [42,43]. The second
plasmid (‘sugar plasmid’) contained the genes necessary
for the biosynthesis of the staurosporine sugar moiety
with or without two genes coding for an O-methyltrans-
ferase and a N-methyltransferase that modify the sugar
moiety. Coexistence of these two plasmids in S. albusresulted in the production of staurosporine and several
glycosylated derivatives with differences in the sugar
moiety (Figure 4, compounds 31, 36–38). On the basis
of this two-plasmid system, the replacement of the ‘sugar
plasmid’ by a family of ‘sugar cassette plasmids’, each one
directing the biosynthesis of different L-deoxysugars and
Current Opinion in Chemical Biology 2009, 13:152–160
D-deoxysugars [44] lead to the formation of novel glyco-
sylated indolocarbazoles containing L-rhamnose, L-oli-
vose, L-digitoxose or D-olivose attached either through
single (in all the cases) (Figure 4, compounds 32–35) or
double (in the case of the former three sugars but not for
D-olivose) C–N bonds (Figure 4, compounds 39–41)
[41��]. Recently, similar rebeccamycin derivatives have
been produced by replacing the staC by the rebC gene in
the ‘aglycone plasmid’ (AP Salas et al., unpublished data)
(Figure 4, compounds 43–46, 49–51). Through biocon-
version experiments it has been also shown that the RebG
rebeccamycin glycosyltransferase can glucosylate a set of
indolocarbazole surrogates, and evidence for a remarkable
lack of regioselectivity of this transferase in the presence
of asymmetric substrates has been provided (Figure 4,
compounds 53–57)[45]. Interestingly, the RebM O-meth-
yltransferase can modify in vitro many of the glycosylated
compounds produced by RebG (Figure 4) [45]. The RebM
O-methyltransferase also shows an interesting peculiarity
regarding the methyl group donor. RebM is capable of
using a non-natural cofactor analogue of S-adenosyl-L-
methionine (a novel synthetic cofactor bearing a pendant
50-amino acid N-mustard) as methyl group donor to gen-
erate a number of new rebeccamycin analogues [46��].This could be another way to increase the molecular
biodiversity in indolocarbazoles for drug discovery.
Concluding remarksAs an alternative to chemical synthesis, the introduction of
combinatorial biosynthesis technology and its application
to indolocarbazole biosynthesis gene clusters may
represent a promising way of expanding the structural
diversity of this family of biologically active natural pro-
ducts. The identification of the genes responsible for the
biosynthesis of four indolocarbazoles (rebeccamycin, staur-
osporine, AT2433 and K252a) provides genetical tools for
such purpose. More than 50 different indolocarbazole
derivatives have been produced in a rational way through
the expression of selected genes in a convenient bacterial
host. Two interesting features of their biosynthetic path-
ways make them interesting targets for combinatorial
biosynthesis. Firstly, indolocarbazole pathways are rela-
tively simple mainly in comparison to pathways for other
more complex natural products and, secondly, some of the
indolocarbazole biosynthetic enzymes have some degree
of substrate flexibility, being capable of recognizing and
processing different intermediates thus making possible
the generation of novel derivatives. Applying combinator-
ial biosynthesis to this group of compounds will contribute
to further expanding the chemical space of indolocarba-
zoles and thus to provide potentially active derivatives as
inhibitors of topoisomerases or protein kinases.
AcknowledgementsResearch in the authors’ laboratory has been supported by grants from theSpanish Ministry of Science and Innovation (BFU2006-00404 to JAS andBIO2005-04115 to CM), the Red Tematica de Investigacion Cooperativa de
www.sciencedirect.com

Indolocarbazole antitumour compounds by combinatorial biosynthesis Salas and Mendez 159
Centros de Cancer to JAS (Ministry of Health, Spain; ISCIII-RETIC RD06/0020/0026) and from the UE FP6 (ActinoGen; Integrated project no005224).
References and recommended readingPapers of particular interest, published within the period of review,have been highlighted as:
� of special interest�� of outstanding interest
1.�
Sanchez C, Mendez C, Salas JA: Indolocarbazole naturalproducts: occurrence, biosynthesis, and biological activity.Nat Prod Rep 2006, 23:1007-1045.
This review provides a survey of 257 indolocarbazoles isolated fromdifferent biological sources or generated by combinatorial biosynthesisand describes recent developments in molecular genetics, enzymologyand metabolic engineering, with an emphasis on the development ofanalogues that have entered clinical trials for its future use against canceror other diseases.
2. Akinaga S, Sugiyama K, Akiyama T: UCN-01 (7-hydroxystaurosporine) and other indolocarbazolecompounds: a new generation of anti-cancer agents for thenew century? Anti-Cancer Drug Design 2000, 15:43-52.
3. Bailly C: Topoisomerase I poisons and suppressors asanticancer drugs. Curr Med Chem 2001, 7:39-58.
4.�
Butler MS: Natural products to drugs: natural product derivedcompounds in clinical trials. Nat Prod Rep 2005, 22:162-195.
A comprehensive description on natural products or derived compoundsthat are being evaluated in clinical trials or in registration.
5. Knolker HJ, Reddy KR: Isolation and synthesis of biologicallyactive carbazole alkaloids. Chem Rev 2002, 102:4303-4427.
6. Prudhomme M: Biological targets of antitumorindolocarbazoles bearing a sugar moiety. Curr Med ChemAnticancer Agents 2004, 4:509-521.
7. Pearce CJ, Doyle TW, Forenza S, Lam KS, Schroeder DR: Thebiosynthetic origins of rebeccamycin. J Nat Prod 1988,51:937-940.
8. Lam KS, Forenza S, Doyle TW, Pearce CJ: Identification ofindolepyruvic acid as an intermediate of rebeccamycinbiosynthesis. J Ind Microbiol 1990, 6:291-294.
9. Yang SW, Cordell GA: Origin of nitrogen in the indolocarbazoleunit of staurosporine. J Nat Prod 1997, 60:788-790.
10. Yang SW, Lin LJ, Cordell GA, Wang P, Corley DG: O- and N-Methylation in the biosynthesis of staurosporine. J Nat Prod1999, 62:1551-1553.
11. Goeke K, Hoehn P, Ghisalba O: Production of the staurosporineaglycone K-252c with a blocked mutant of the staurosporineproducer strain Streptomyces longisporoflavus and bybiotransformation of staurosporine with Streptomycesmediocidicus ATCC 13279. J Antibiot (Tokyo) 1995, 48:428-430.
12. Hoehn P, Ghisalba O, Moerker T, Peter HH: 30-Demethoxy-30-hydroxystaurosporine, a novel staurosporine analogueproduced by a blocked mutant. J Antibiot (Tokyo) 1995,48:300-305.
13. Weidner S, Kittelmann M, Goeke K, Ghisalba O, Zahner H: 30-Demethoxy-30-hydroxystaurosporine-O-methyltransferasefrom Streptomyces longisporoflavus catalyzing the last step inthe biosynthesis of staurosporine. J Antibiot (Tokyo) 1998,51:679-682.
14. Lam KS, Schroeder DR, Veitch JM, Matson JA, Forenza S: Isolationof a bromo analog of rebeccamycin from Saccharothrixaerocolonigenes. J Antibiot (Tokyo) 1991, 44:934-939.
15. Lam KS, Schroeder DR, Veitch JM, Colson KL, Matson JA,Rose WC, Doyle TW, Forenza S: Production, isolation andstructure determination of novel fluoroindolocarbazoles fromSaccharothrix aerocolonigenes ATCC 39243. J Antibiot (Tokyo)2001, 54:1-9.
16. Long BH, Rose WC, Vyas DM, Matson JA, Forenza S: Discoveryof antitumor indolocarbazoles: rebeccamycin, NSC 655649,
www.sciencedirect.com
and fluoroindolocarbazoles. Curr Med Chem AntiCancer Agents2002, 2:255-266.
17.�
Ohuchi T, Ikeda-Araki A, Watanabe-Sakamoto A, Kojiri K,Nagashima M, Okanishi M, Suda H: Cloning and expression of agene encoding N-glycosyltransferase (ngt) from SaccharothrixaerocolonigenesATCC39243.JAntibiot (Tokyo)2000,53:393-403.
This paper reports the cloning of the first indolocarbazole biosynthesisgene and experimental evidence for the formation of an N-glucosylatedderivative by biotransformation.
18.�
Sanchez C, Butovich IA, Brana AF, Rohr J, Mendez C, Salas JA:The biosynthetic gene cluster for the antitumor rebeccamycin:characterization and generation of indolocarbazolederivatives. Chem Biol 2002, 9:519-531.
In this paper, the cloning, sequencing and heterologous expression of thefirst indolocarbazole cluster characterized is reported together with theformation of some biosynthesis intermediates by expression of differentsets of genes of the cluster in S. albus.
19. Onaka H, Taniguchi S, Igarashi Y, Furumai T: Characterization ofthe biosynthetic gene cluster of rebeccamycin fromLechevalieria aerocolonigenes ATCC 39243. Biosci BiotechnolBiochem 2003, 67:127-138.
20.�
Hyun CG, Bililign T, Liao J, Thorson JS: The biosynthesis ofindolocarbazoles in a heterologous Escherichia coli host.Chembiochem 2003, 4:114-117.
The first report on the heterologous expression of an indolocarbazolegene cluster in Escherichia coli.
21.�
Onaka H, Taniguchi S, Igarashi Y, Furumai T: Cloning of thestaurosporine biosynthetic gene cluster from Streptomycessp. TP-A0274 and its heterologous expression inStreptomyces lividans. J Antibiot (Tokyo) 2002, 55:1063-1071.
It describes the cloning, sequencing and heterologous expression of thestaurosporine cluster and a number of insertional inactivation mutantsaccumulating specific biosynthetic intermediates.
22. Gao Q, Zhang C, Blanchard S, Thorson JS: Decipheringindolocarbazole and enediyne aminodideoxypentosebiosynthesis through comparative genomics: insights fromthe AT2433 biosynthetic locus. Chem Biol 2006, 13:733-743.
23. Kim SY, Park JS, Chae CS, Hyun CG, Choi BW, Shin J, Oh KB:Genetic organization of the biosynthetic gene cluster for theindolocarbazole K-252a in Nonomuraea longicatena JCM11136. Appl Microbiol Biotechnol 2007, 75:1119-1126.
24. August PR, Grossman TH, Minor C, Draper MP, MacNeil IA,Pemberton JM, Call KM, Holt D, Osburne MS: Sequence analysisand functional characterization of the violacein biosyntheticpathway from Chromobacterium violaceum. J Mol MicrobiolBiotechnol 2000, 2:513-519.
25. Sanchez C, Brana AF, Mendez C, Salas JA: Reevaluation of theviolacein biosynthetic pathway and its relationship toindolocarbazole biosynthesis. Chembiochem 2006, 7:1231-1240.
26. Yeh E, Garneau S, Walsh CT: Robust in vitro activity of RebF andRebH, a two-component reductase/halogenase, generating 7-chlorotryptophan during rebeccamycin biosynthesis. Proc NatlAcad Sci U S A 2005, 102:3960-3965.
27. Yeh E, Blasiak LC, Koglin A, Drennan CL, Walsh CT: Chlorinationby a long-lived intermediate in the mechanism of flavin-dependent halogenases. Biochemistry 2007, 46:1284-1292.
28. Yeh E, Cole LJ, Barr EW, Bollinger JM Jr, Ballou DP, Walsh CT:Flavin redox chemistry precedes substrate chlorination duringthe reaction of the flavin-dependent halogenase RebH.Biochemistry 2006, 45:7904-7912.
29. Howard-Jones AR, Walsh CT: Enzymatic generation of thechromopyrrolic acid scaffold of rebeccamycin by the tandemaction of RebO and RebD. Biochemistry 2005, 44:15652-21563.
30.�
Nishizawa T, Aldrich CC, Sherman DH: Molecular analysis of therebeccamycin L-amino acid oxidase from Lechevalieriaaerocolonigenes ATCC 39243. J Bacteriol 2005, 187:2084-2092.
The last two papers report the first in vitro characterization of enzymesinvolved in early steps in the biosynthesis of rebeccamycin.
31. Nishizawa T, Gruschow S, Jayamaha DH, Nishizawa-Harada C,Sherman DH: Enzymatic assembly of the bis-indole core ofrebeccamycin. J Am Chem Soc 2006, 128:724-725.
Current Opinion in Chemical Biology 2009, 13:152–160

160 Molecular Diversity
32. Asamizu S, Kato Y, Igarashi Y, Furumai T, Onaka H: Directformation of chromopyrrolic acid from indole-3-pyruvic acidby StaD, a novel hemoprotein in indolocarbazole biosynthesis.Tetrahedron Lett 2006, 47:473-475.
33. Howard-Jones AR, Walsh CT: Nonenzymatic oxidative stepsaccompanying action of the cytochrome P450 enzymes StaPand RebP in the biosynthesis of staurosporine andrebeccamycin. J Am Chem Soc 2007, 129:11016-11107.
34. Makino M, Sugimoto H, Shiro Y, Asamizu S, Onaka H, Nagano S:Crystal structures and catalytic mechanism of cytochromeP450 StaP that produces the indolocarbazole skeleton. ProcNatl Acad Sci U S A 2007, 104:11591-11596.
35. Ryan KS, Howard-Jones AR, Hamill MJ, Elliott SJ, Walsh CT,Drennan CL: Crystallographic trapping in the rebeccamycinbiosynthetic enzyme RebC. Proc Natl Acad Sci U S A 2007,104:15311-21536.
36. Howard-Jones AR, Walsh CT: Staurosporine and rebeccamycinaglycones are assembled by the oxidative action of StaP,StaC, and RebC on chromopyrrolic acid. J Am Chem Soc 2006,128:12289-12298.
37.��
Sanchez C, Zhu L, Brana AF, Salas AP, Rohr J, Mendez C,Salas JA: Combinatorial biosynthesis of antitumorindolocarbazole compounds. Proc Natl Acad Sci U S A 2005,102:461-466.
A good example of the use of combinatorial biosynthesis to increasestructural diversity in natural products with the generation of more than 30different indolocarbazoles in the heterologous host Streptomyces albus.
38.�
Salas JA, Mendez C: Engineering the glycosylation of naturalproducts in actinomycetes. Trends Microbiol 2007, 15:219-232.
In this paper, the ‘state-of-the-art’ on the use of combinatorial biosynth-esis to alter the glycosylation pattern in natural products produced byactinomycetes is reviewed.
39. Luzhetskyy A, Mendez C, Salas JA, Bechthold A:Glycosyltransferases, important tools for drug design.Curr Top Med Chem 2008, 8:680-709.
Current Opinion in Chemical Biology 2009, 13:152–160
40. Mendez C, Luzhetskyy A, Bechthold A, Salas JA: Deoxysugars inbioactive natural products: development of novel derivativesby altering the sugar pattern. Curr Top Med Chem 2008,8:710-724.
41.��
Salas AP, Zhu L, Sanchez C, Brana AF, Rohr J, Mendez C,Salas JA: Deciphering the late steps in the biosynthesis of theanti-tumour indolocarbazole staurosporine: sugar donorsubstrate flexibility of the StaG glycosyltransferase. MolMicrobiol 2005, 58:17-27.
In this paper, a number of novel glycosylated staurosporine derivativesare generated in an heterologous host by combining two plasmids eachone coding for different parts of the indolocarbazole molecule.
42. Onaka H, Asamizu S, Igarashi Y, Yoshida R, Furumai T:Cytochrome P450 homolog is responsible for C-N bondformation between aglycone and deoxysugar in thestaurosporine biosynthesis of Streptomyces sp. TP-A0274.Biosci Biotechnol Biochem 2005, 69:1753-1759.
43. Sanchez C, Mendez C, Salas JA: Engineering biosyntheticpathways to generate antitumor indolocarbazole derivatives.J Ind Microbiol Biotechnol 2006, 33:560-568.
44. Rodrıguez L, Aguirrezabalaga I, Allende N, Brana AF, Mendez C,Salas JA: Engineering deoxysugar biosynthetic pathways fromantibiotic-producing microorganisms. A tool to produce novelglycosylated bioactive compounds. Chem Biol 2002, 9:721-729.
45. Zhang C, Albermann C, Fu X, Peters NR, Chisholm JD, Zhang G,Gilbert EJ, Wang PG, Van Vranken DL, Thorson JS: RebG- andRebM-catalyzed indolocarbazole diversification.Chembiochem 2006, 7:795-804.
46.��
Zhang C, Weller RL, Thorson JS, Rajski SR: Natural productdiversification using a non-natural cofactor analogue of S-adenosyl-L-methionine. J Am Chem Soc 2006, 128:2760-2761.
Interesting description on the ability of the RebM O-methyltransferaseto use a non-natural cofactor analogue of S-adenosyl-L-methionine asmethyl group donor to generate a number of new rebeccamycinanalogues.
www.sciencedirect.com




















