
Histological Studies on Early Oogenesis in Barfin Flounder ...
Upload
independentCategory
view
3download
0
Increased Expression of T Cell Chemokines andTheir Receptors in Chronic HepatitisC: Relationship With the Histological Activity ofLiver DiseaseArantxa Apolinario, Ph.D., Pedro L. Majano, Ph.D., Eduardo Alvarez-Pe´rez, M.D., Alicia Saez, Ph.D.,Carlos Lozano, M.D., Javier Vargas, M.D., and Carmelo Garcı´a-Monzon, M.D.Hepatology Unit, Serology and Biochemistry Service, and Pathology Service, University Hospital SantaCristina, Madrid, Spain
OBJECTIVES: Although chemokines seem to be important incertain inflammatory disorders, little is known about the roleof these proteins in chronic hepatitis C.
METHODS: Expression of selected CXC and CC chemokinesand their receptors was assessed by immunohistochemistryand flow cytometry in chronic hepatitis C. Tissue samplesfrom normal liver and that of sustained responders were alsoevaluated. A comparative analysis between the histologicalgrading and the intrahepatic expression level of chemokineswas performed.
RESULTS: The majority of liver-derived T lymphocytes ex-pressed CXCR3 and CCR5 chemokine receptors, represent-ing high enrichment over levels of CXCR3� and CCR5� Tcells in blood from chronic hepatitis C. An intense intrahe-patic expression of their respective ligands, the CXC che-mokine Mig, and RANTES, was detected in the same pa-tients studied, being restricted to the sinusoidal endotheliumand to hepatocytes, respectively. A statistically significantassociation between the intrahepatic chemokine expressionlevel and the inflammatory activity of chronic hepatitis Cwas found. Of note was the marked expression of bothCXCR3 and its ligand Mig on endothelial cells from portalneovessels in chronic hepatitis C.
CONCLUSIONS: Intrahepatic chemokine signaling couldplay a key role regulating significant pathological eventsduring chronic hepatitis C, opening new avenues for thera-peutic interventions based on chemokine activities. (Am JGastroenterol 2002;97:2861–2870. © 2002 by Am. Coll. ofGastroenterology)
INTRODUCTION
The histological hallmark of chronic hepatitis C (CHC) is aninfiltration of lymphoid cells in the portal tracts that disruptsthe limiting plate, invading the surrounding parenchyma andleading to periportal or “piecemeal” necrosis (1). T lym-phocytes are regarded as the predominant cell population
infiltrating the areas of liver damage in CHC (2, 3), andthere is increasing evidence to suggest that extension of theinflammatory process from portal tracts into the periportaland lobular areas could be crucial in the development ofprogressive liver injury with potential prognostic implica-tions (4, 5). It is conceivable, therefore, that a greater un-derstanding of the mechanisms regulating T cell migrationand recruitment to the liver parenchyma in CHC can provideinsights into the pathogenesis of this viral liver disease.
The recruitment and migration of inflammatory cells de-pends on the local production of chemotactic cytokineswithin inflamed tissues (6). These proinflammatory media-tors, called chemokines (CK), belong to a superfamily oflow molecular weight cytokines that can be classified intofour groups, of which the largest subgroups are the CXC andCC families. In the CXC CK, one amino acid separates thefirst two cysteine residues, whereas in the CC CK, the firsttwo cysteine residues are adjacent to each other (7). TheCXC family can be further subdivided into CXC CK, con-taining ELR (the three amino acid residues [glutamic-leucine-arginine] that immediately precede the first cysteineamino acid in the CXC chemokine family), which are im-portant in neutrophil chemotaxis and angiogenesis (8), andthe non-ELR CXC CK, such as monokine induced by IFN-�(Mig) and interferon-�–inducible protein (IP-10) betweenothers, which are chemotactic for T lymphocytes and haveangiostatic activity (9). On the other hand, the CC CK,including MCP-1 and RANTES, show chemotactic activitypreferentially for T cells (10). Finally, CK exert their func-tions by binding to specific cell surface receptors on targetcells. Five human CXC CK receptors and nine human CCCK receptors have so far been identified (11).
Although intrahepatic up-regulation of CK expression hasbeen observed in CHC (12–14), there is little informationregarding the relative contribution of specific CK/CK re-ceptor interactions (CK pairs) to the development of rele-vant pathological events in the course of CHC, such ashepatic infiltration by activated T cells (15) and portal
THE AMERICAN JOURNAL OF GASTROENTEROLOGY Vol. 97, No. 11, 2002© 2002 by Am. Coll. of Gastroenterology ISSN 0002-9270/02/$22.00Published by Elsevier Science Inc. PII S0002-9270(02)05471-0

angiogenesis (16). Considering that Mig and RANTES sig-naling attract memory and activated T cells, but not restingT cells (17, 18), and that CXCR3-binding mediates angio-static activity (19), in the present study we aimed to com-pare the expression pattern of a selected CXC CK (Mig) andits receptor (CXCR3) as well as a CC CK (RANTES) and itsreceptor (CCR5) in peripheral blood and liver tissue frompatients with histopathologically distinct grades of CHC,and to determine the relationship between both the intrahe-patic inflammation and portal angiogenesis and the localexpression of these CK pairs.
MATERIALS AND METHODS
Study PopulationA total of 43 consecutive adult patients (median age 37 yr,range 22–59) that were referred to our outpatient clinicbetween January, 1999, and February, 2000, with persis-tenly elevated serum ALT levels as well as the presence ofboth antibodies to hepatitis C virus (HCV) and HCV RNAwere included in this study. Blood samples from each pa-tient for quantification and typing of HCV RNA were ob-tained when a diagnostic liver biopsy was carried out. De-mographic, biochemical, and virological characteristics ofthese patients are summarized in Table 1. We further studiedfive of these HCV patients who achieved a sustained bio-chemical and virological response after combined antiviraltherapy and underwent a liver biopsy in that period (6months after the end of treatment). Finally, we wanted tocompare pretreatment data on the five sustained responders(SR) and a number of matched nonresponders (NR) toantiviral therapy. Written informed consent was obtainedfrom each patient, and approval for the study was granted bythe institution’s Ethical Committee.
Virological MarkersQuantification of the amount of the HCV RNA was per-formed by Amplicor HCV Monitor (Roche Diagnostic Sys-tems, Branchburg, NJ), based on a simplified RNA extrac-tion and a single, combined reverse transcription andamplification reaction, mediated by termus thermophilusDNA polymerase (20). Genotype was tested by Line ProbeAssay (Inno-Lipa HCV II, Innogenetics, Hanover, Germa-ny).
Antibodies for Immunohistochemistry and FlowCytometryThe primary antibodies (Abs) used for immunohistochem-istry were CXCR3 mouse antihuman monoclonal IgG1(PharMingen, San Diego, CA), CCR5 mouse antihumanmonoclonal IgG1 (PharMingen), Mig goat antihuman poly-clonal IgG (R&D Systems, Abingdon, UK), RANTES goatantihuman polyclonal IgG (Santa Cruz Biotechnology,Santa Cruz, CA), and TEA 1/3 mouse antihuman monoclo-nal IgG1 recognizing the molecule ve-cadherin on endothe-lial cells, described elsewhere (21). Secondary Abs for im-munohistochemistry were peroxidase-conjugated rabbitantigoat (Dakopatts, Copenhagen, Denmark), rabbit anti-mouse Igs (Dakopatts), and alkaline phosphatase antialka-line phosphatase mouse monoclonal (Dakopatts). TheP3X63 mouse IgG1 myeloma supernatant was used as neg-ative control in all immunostaining experiments. The Absused for flow cytometry were phycoerythrin (PE)–conju-gated CD3 mouse antihuman monoclonal IgG1 (total Tlymphocytes) (Becton Dickinson, San Diego, CA), CXCR3mouse antihuman monoclonal IgG1 (PharMingen), andCCR5 mouse antihuman monoclonal IgG1 (PharMingen).
Liver Tissue StudiesLiver biopsy samples from all patients were obtained usinga Menghini needle by a percutaneous route. Additionally,seven tissue samples from histologically normal livers werealso assessed as controls for immunohistochemical analysis.After patients had provided written consent, these livertissue samples were obtained during abdominal surgeryfrom patients with noncomplicated cholecystectomy (n � 4)or morbid obesity (n � 3); all had normal serum amino-transferase levels and were negative for anti-HCV antibod-ies.
LIVER HISTOLOGY. Biopsy specimens from all patientswere evaluated under code by the same pathologist, estab-lishing the histological diagnosis. For an accurate compar-ative data analysis between intrahepatic inflammation andthe local expression of CK, the grade of inflammation wasscored on a scale of 0–4, using the classification of chronicviral hepatitis proposed by Scheuer (1). In addition, weclassified histological activity of CHC as mild or severe,defining mild CHC as �2, and severe CHC as �2.
IMMUNOHISTOCHEMICAL STAINING OF LIVER BI-OPSY SECTIONS. A total of 27 liver biopsy samples frompatients with CHC included in this study were assessedusing an indirect immunoperoxidase staining technique. Inaddition, the following experiments of double immunostain-ing in selected liver tissue sections were also carried out: 1)immunoperoxidase and double immunoenzymatic staining,using standard procedures as detailed elsewhere (16); and 2)evaluation of immunoperoxidase-stained liver biopsy sec-tions, as follows. Each section was semiquantitatively eval-uated by two independent observers using a Nikon lightmicroscope (Nikon Labophot, Tokyo, Japan) without the
Table 1. Demographic, Biochemical, and Virological Features ofChronic Hepatitis C Patients Studied
Number of patients 43Age (yr) 37 (22–59)Sex (male/female) 24/19ALT (U/L) 195 (49–351)HCV genotype
1b 27 (63%)non-1b 16 (37%)
HCV viremia (copies/ml) 738,309 (163,252–2,237,541)
Data are given as median (range).
2862 Apolinario et al. AJG – Vol. 97, No. 11, 2002

knowledge of either clinical or histological diagnosis. Fourdistinct items were recorded on separate sheets as follows:0 � no staining; �1 � positive staining in �30% of cellsper high power field (�250); �2 � positive staining in�30% but in �70% of cells per high power field; and � 3 �positive staining of �70% of cells per high power field,according to a previously validated scoring system (22).Finally, data were averaged to median values, giving anumerical score for each liver biopsy specimen, and thenused for statistical analysis.
ISOLATION OF LYMPHOCYTES FROM LIVER BI-OPSY SAMPLES. A total of 16 fresh liver biopsy sampleswere immediately collected in a sterile tube containingRPMI-1640 (Flow, Irvine, UK) supplemented with 50IU/ml penicilin-streptomycin, 1% L-glutamine (Flow), and10% fetal calf serum (Gibco, Middlesex, UK), which isreferred to as complete medium. All samples were hand-delivered to the laboratory. Then, the collagenase isolationmethod was carried out as previously described (16). Thenumber of mononuclear cells recovered from these liverbiopsy samples ranged from 1 � 105 to 6 � 105 cells, andtheir viability was invariably �90%.
Isolation of Peripheral Blood LymphocytesThese cells were obtained from heparinized venous blooddrawn at the time of liver biopsy of the same 16 patientswho were selected for isolation of liver-infiltrating lympho-cytes. Peripheral blood lymphocytes were separated by Fi-coll-Hypaque (Pharmacia, Uppsala, Sweden) density gradi-ent at 1500 rpm for 30 min at room temperature and washedtwice in complete medium.
Flow CytometryTwo-color staining experiments were carried out using amethod previously described (16). Stained mononuclearcells were analyzed with a FACscan cytofluorometer (Bec-ton Dickinson). Lymphocytes were gated on the basis ofboth forward and side scatter parameters to exclude debrisand dead cells and by gating on CD3� cells. The percent-ages of positive T cells were calculated using LYSYS IIsoftware (Becton Dickinson).
Statistical AnalysisTo determine the statistical significance of variations in theimmunohistochemical scoring of the molecules studied, wefirst calculated the mean values of staining scores from eachone of the histological groups evaluated; these values werethen compared by the Mann-Whitney U test for nonpara-metric data. Student’s t test was used to assess the statisticalsignificance of the differences observed between the periph-eral blood and intrahepatic lymphocyte subsets; p valuesbelow 0.05 were considered as significant.
RESULTS
Histopathological EvaluationA low grade of intrahepatic inflammation (�2) defining amild histological form of CHC was found in 25 patients(58%), whereas a high inflammatory grade (�2) was seen inthe liver tissue from 18 patients (42%). Liver biopsy sam-ples from five patients with CHC who achieved a sustainedbiochemical and virological response after antiviral therapyshowed nonspecific reactive changes with a grading score of0.
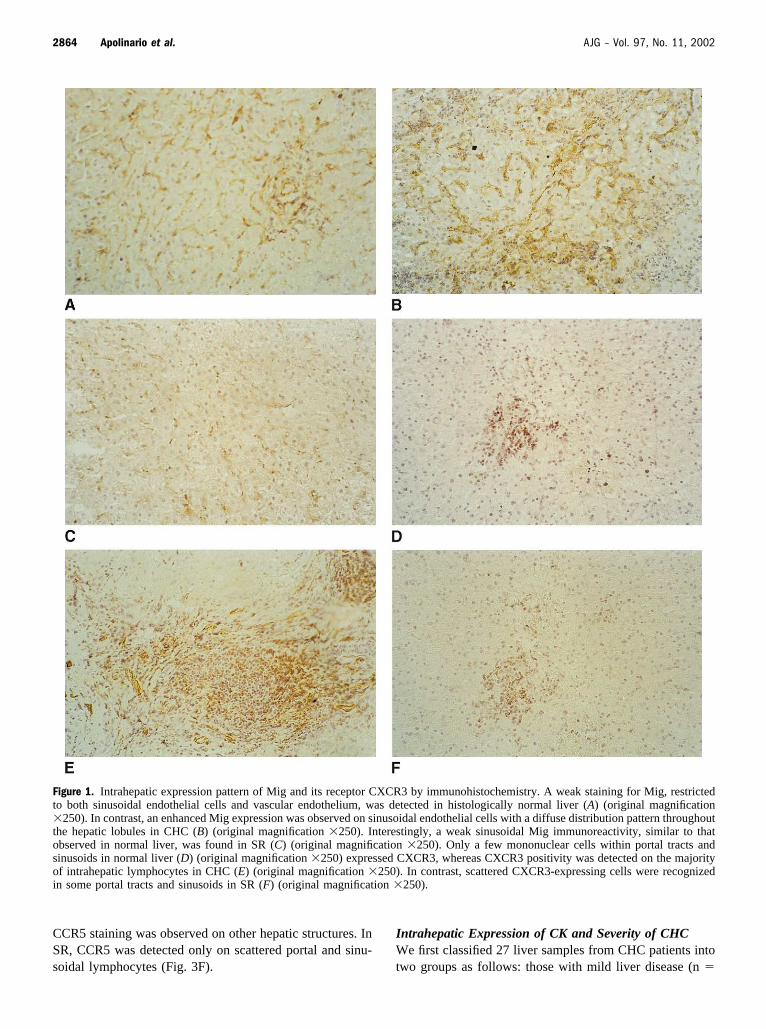
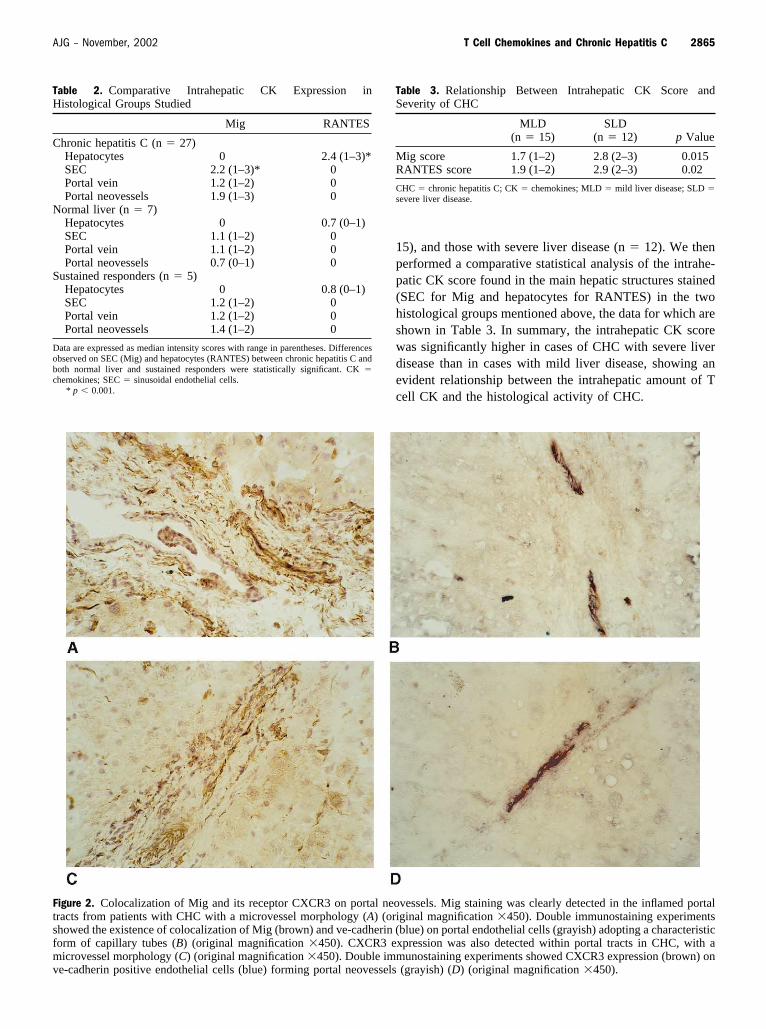
Intrahepatic Expression Pattern of T Cell CK and TheirReceptorsAn intense expression of the CXC chemokine Mig wasfound in all liver tissue samples from patients with CHC.Positive Mig staining was observed mainly on sinusoidalendothelial cells (SEC), with a diffuse distribution patternthroughout the hepatic lobules. Although the intrahepaticexpression pattern of Mig was similar in normal liver (Fig.1A), the staining intensity was significantly enhanced inCHC (Fig. 1B and Table 2), being more intense in thoseacinar areas that were more heavily infiltrated by lymphoidcells. A weak sinusoidal Mig immunoreactivity, similar tothat observed in normal liver, was found in the liver tissuefrom SR (Fig. 1C). Besides a weak expression on portalvascular endothelium, Mig staining was clearly detected inthe inflamed portal tracts from patients with CHC with amicrovessel morphology (Fig. 2A). Double immunostainingexperiments showed that Mig colocalized with ve-cadherin–expressing portal microvessels (Fig. 2B), ve-cadherin beinga selective endothelial cell marker (21). On the other hand,only a few mononuclear cells in portal tracts and sinusoidsin normal liver (Fig. 1D) expressed CXCR3, whereasCXCR3 positivity was detected on the majority of liver-infiltrating lymphocytes in all cases of CHC studied (Fig.1E). In contrast, scattered CXCR3-expressing cells wererecognized in some portal tracts and sinusoids in sustainedresponders (Fig. 1F). Notably, CXCR3 expression was alsodetected within enlarged portal tracts in CHC, adopting acharacteristic microvessel morphology (Fig. 2C). We fur-ther carried out double immunostaining experiments, show-ing CXCR3 expression on ve-cadherin positive endothelialcells forming portal neovessels (Fig. 2D).
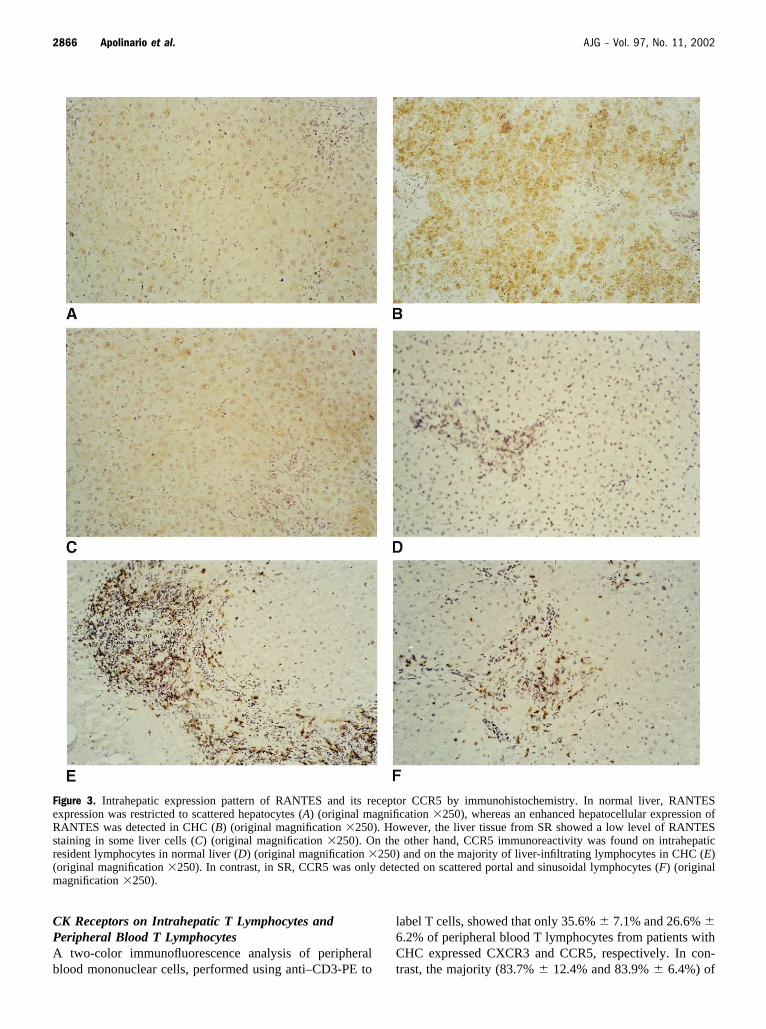
In normal liver, RANTES expression was restricted toscattered hepatocytes (Fig. 3A), whereas an increased intra-hepatic expression of RANTES was detected in CHC, thisdifference being statistically significant (Table 2). Hepato-cellular RANTES expression was more intense in thoseperiportal and lobular areas where lymphocyte infiltrationwas more evident (Fig. 3B). However, the liver tissue fromSR showed a low level of RANTES staining in some livercells (Fig. 3C). CCR5 immunoreactivity was found on in-trahepatic resident lymphocytes in normal liver (Fig. 3D)and on liver-infiltrating lymphocytes in CHC (Fig. 3E). No
2863AJG – November, 2002 T Cell Chemokines and Chronic Hepatitis C
CCR5 staining was observed on other hepatic structures. InSR, CCR5 was detected only on scattered portal and sinu-soidal lymphocytes (Fig. 3F).
Intrahepatic Expression of CK and Severity of CHCWe first classified 27 liver samples from CHC patients intotwo groups as follows: those with mild liver disease (n �
Figure 1. Intrahepatic expression pattern of Mig and its receptor CXCR3 by immunohistochemistry. A weak staining for Mig, restrictedto both sinusoidal endothelial cells and vascular endothelium, was detected in histologically normal liver (A) (original magnification�250). In contrast, an enhanced Mig expression was observed on sinusoidal endothelial cells with a diffuse distribution pattern throughoutthe hepatic lobules in CHC (B) (original magnification �250). Interestingly, a weak sinusoidal Mig immunoreactivity, similar to thatobserved in normal liver, was found in SR (C) (original magnification �250). Only a few mononuclear cells within portal tracts andsinusoids in normal liver (D) (original magnification �250) expressed CXCR3, whereas CXCR3 positivity was detected on the majorityof intrahepatic lymphocytes in CHC (E) (original magnification �250). In contrast, scattered CXCR3-expressing cells were recognizedin some portal tracts and sinusoids in SR (F) (original magnification �250).
2864 Apolinario et al. AJG – Vol. 97, No. 11, 2002
15), and those with severe liver disease (n � 12). We thenperformed a comparative statistical analysis of the intrahe-patic CK score found in the main hepatic structures stained(SEC for Mig and hepatocytes for RANTES) in the twohistological groups mentioned above, the data for which areshown in Table 3. In summary, the intrahepatic CK scorewas significantly higher in cases of CHC with severe liverdisease than in cases with mild liver disease, showing anevident relationship between the intrahepatic amount of Tcell CK and the histological activity of CHC.
Figure 2. Colocalization of Mig and its receptor CXCR3 on portal neovessels. Mig staining was clearly detected in the inflamed portaltracts from patients with CHC with a microvessel morphology (A) (original magnification �450). Double immunostaining experimentsshowed the existence of colocalization of Mig (brown) and ve-cadherin (blue) on portal endothelial cells (grayish) adopting a characteristicform of capillary tubes (B) (original magnification �450). CXCR3 expression was also detected within portal tracts in CHC, with amicrovessel morphology (C) (original magnification �450). Double immunostaining experiments showed CXCR3 expression (brown) onve-cadherin positive endothelial cells (blue) forming portal neovessels (grayish) (D) (original magnification �450).
Table 2. Comparative Intrahepatic CK Expression inHistological Groups Studied
Mig RANTES
Chronic hepatitis C (n � 27)Hepatocytes 0 2.4 (1–3)*SEC 2.2 (1–3)* 0Portal vein 1.2 (1–2) 0Portal neovessels 1.9 (1–3) 0
Normal liver (n � 7)Hepatocytes 0 0.7 (0–1)SEC 1.1 (1–2) 0Portal vein 1.1 (1–2) 0Portal neovessels 0.7 (0–1) 0
Sustained responders (n � 5)Hepatocytes 0 0.8 (0–1)SEC 1.2 (1–2) 0Portal vein 1.2 (1–2) 0Portal neovessels 1.4 (1–2) 0
Data are expressed as median intensity scores with range in parentheses. Differencesobserved on SEC (Mig) and hepatocytes (RANTES) between chronic hepatitis C andboth normal liver and sustained responders were statistically significant. CK �chemokines; SEC � sinusoidal endothelial cells.
* p � 0.001.
Table 3. Relationship Between Intrahepatic CK Score andSeverity of CHC
MLD(n � 15)
SLD(n � 12) p Value
Mig score 1.7 (1–2) 2.8 (2–3) 0.015RANTES score 1.9 (1–2) 2.9 (2–3) 0.02
CHC � chronic hepatitis C; CK � chemokines; MLD � mild liver disease; SLD �severe liver disease.
2865AJG – November, 2002 T Cell Chemokines and Chronic Hepatitis C
CK Receptors on Intrahepatic T Lymphocytes andPeripheral Blood T LymphocytesA two-color immunofluorescence analysis of peripheralblood mononuclear cells, performed using anti–CD3-PE to
label T cells, showed that only 35.6% � 7.1% and 26.6% �6.2% of peripheral blood T lymphocytes from patients withCHC expressed CXCR3 and CCR5, respectively. In con-trast, the majority (83.7% � 12.4% and 83.9% � 6.4%) of
Figure 3. Intrahepatic expression pattern of RANTES and its receptor CCR5 by immunohistochemistry. In normal liver, RANTESexpression was restricted to scattered hepatocytes (A) (original magnification �250), whereas an enhanced hepatocellular expression ofRANTES was detected in CHC (B) (original magnification �250). However, the liver tissue from SR showed a low level of RANTESstaining in some liver cells (C) (original magnification �250). On the other hand, CCR5 immunoreactivity was found on intrahepaticresident lymphocytes in normal liver (D) (original magnification �250) and on the majority of liver-infiltrating lymphocytes in CHC (E)(original magnification �250). In contrast, in SR, CCR5 was only detected on scattered portal and sinusoidal lymphocytes (F) (originalmagnification �250).
2866 Apolinario et al. AJG – Vol. 97, No. 11, 2002

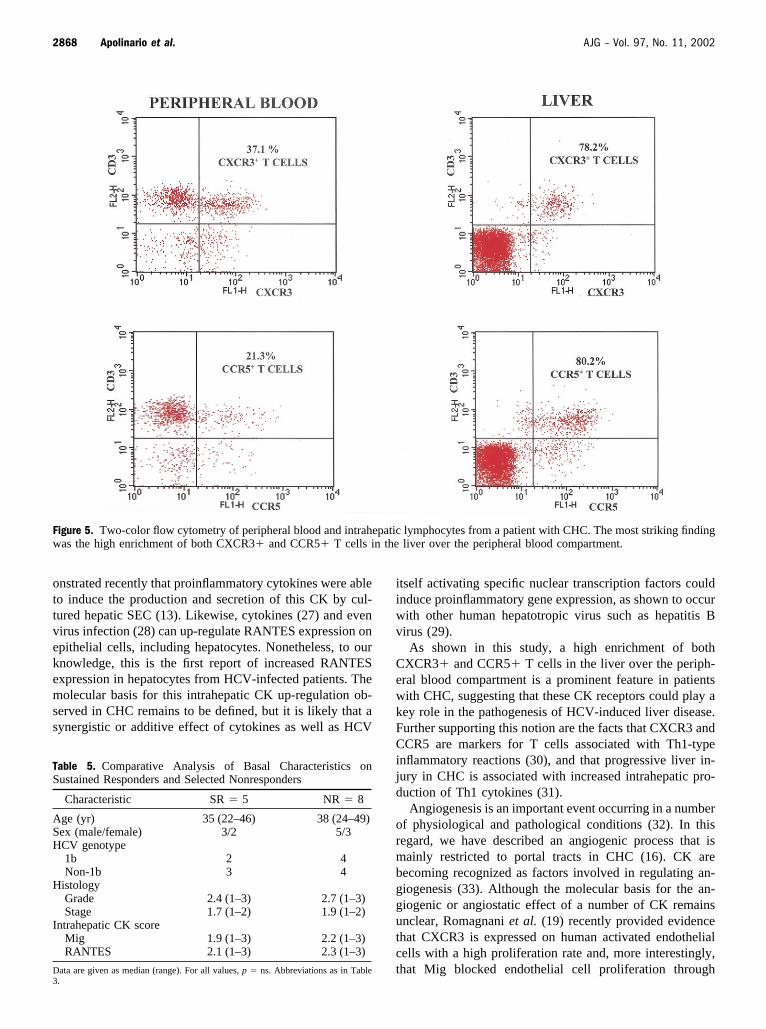
intrahepatic T lymphocytes from the same CHC patientsexpressed the T cell CK receptors CXCR3 and CCR5,respectively. Differences found between the peripheralblood and intrahepatic compartments were statistically sig-nificant (Table 4 and Fig. 4). The most striking finding wasthe high enrichment of both CXCR3� and CCR5� T cellsin the liver over the peripheral blood compartment (Fig. 5).
Relationship Between CK Expression and TherapeuticResponseTo test this hypothesis, we compared pretreatment intrahe-patic CK values of the five SR and of the eight NR matchedfor sex, age, histology, and genotype. No significant differ-ences in the basal intrahepatic CK scores (such for Mig asfor RANTES) between SR and NR were observed (medianMig score: SR 1.9 vs NR 2.2, p � ns; median RANTESscore: SR 2.1 vs NR 2.3, p � ns) (Table 5).
DISCUSSION
An increasing amount of experimental evidence now sug-gests that the liver pathology in HCV-infected patients is adirect result of the intrahepatic immune response to this
virus (23, 24). To this end, selective recruitment to the livertissue of effector T cells capable of producing cytokines,such as IFN-� and TNF-� (Th1 cytokines), is key to achiev-ing an effective antiviral response (25). Given that CK pairsplay a fundamental role in T cell recruitment (8), and thathepatic infiltration by memory and activated T lymphocytesfeatures chronic HCV infection (15), attention has focusedon identifying CK pairs in the liver tissue of these patients.In this regard, our study show evidence that CK pairs, suchas Mig/CXCR3 and RANTES/CCR5, seem to be importantin regulating some relevant pathological events in the courseof CHC, such as hepatic inflammation and portal angiogen-esis. Thus, we found a significantly higher intrahepaticexpression of Mig and RANTES in CHC than in normalliver samples and in those of sustained responders (Table 2).Interestingly, both CXCR3 and CCR5, the respective recep-tors for Mig and RANTES, were expressed on the majorityof intrahepatic T lymphocytes from patients with CHC. It istherefore conceivable that intrahepatic CK signaling duringchronic HCV infection may occur, thus contributing to therecruitment from the vasculature into the liver of primed Tcells.
Although an intense expression of Mig and RANTES inthe inflamed liver from CHC patients has previously beendescribed by different authors (13, 14), little is known aboutits relationship with the severity of this viral liver disease.To address this issue, we evaluated the expression level ofthese two chemoattractants in CHC liver biopsies with low(mild) and high (severe) histological grades of inflamma-tion, showing for the first time that the intrahepatic expres-sion level of both Mig and RANTES is positively related tothe severity of hepatic inflammation in CHC (Table 3). Thissuggests that intrahepatic production of T cell CK may playa decisive role regulating migration of effector T cells frominflamed portal areas and further recruitment into periportaland lobular ones, thereby promoting the progression toadvanced forms of liver disease.
Predicting the virological response to combined therapyis a crucial point in the management of patients with CHC.In this regard, viral load and HCV genotyping have beenshown to be relevant to the outcome of anti-HCV therapy(26); however, given the high proportion of patients whostill do not respond to antiviral therapy, the search for otherfactors predictive of response is needed. In this study, weexplored whether the intrahepatic CK levels might influencethe type of response to antiviral therapy, but we did notobserve any significant differences in the basal intrahepaticCK scores in a selected population of SR and NR (Table 5).However, this study design does not allow us to draw anydefinite conclusions on this matter, and future studies on alarge population of unselected CHC patients, with a moreappropriate design for assessing potential factors predictiveof response, are warranted.
Based on our immunohistochemical results, a potentialsource for Mig and RANTES production are SEC andhepatocytes, respectively. With regard to Mig, it was dem-
Figure 4. Bar graph depicting expression of CXCR3 and CCR5 onperipheral blood T lymphocytes (PBTL) and intrahepatic T lym-phocytes (IHTL) in chronic hepatitis C (CHC). The majority ofIHTL expressed CK receptors compared with a small population ofPBTL.
Table 4. Expression of CXCR3 and CCR5 on Peripheral BloodT Lymphocytes (PBTL) and Intrahepatic T Lymphocytes(IHTL) in Chronic Hepatitis C (CHC)
% Cells,PBTL
% Cells,IHTL
pValue
CXCR3�/CD3� 35.6 � 7.1 83.7 � 12.4 0.03CCR5�/CD3� 26.6 � 7.2 83.9 � 6.4 0.002
Data are expressed as mean � SD.
2867AJG – November, 2002 T Cell Chemokines and Chronic Hepatitis C
onstrated recently that proinflammatory cytokines were ableto induce the production and secretion of this CK by cul-tured hepatic SEC (13). Likewise, cytokines (27) and evenvirus infection (28) can up-regulate RANTES expression onepithelial cells, including hepatocytes. Nonetheless, to ourknowledge, this is the first report of increased RANTESexpression in hepatocytes from HCV-infected patients. Themolecular basis for this intrahepatic CK up-regulation ob-served in CHC remains to be defined, but it is likely that asynergistic or additive effect of cytokines as well as HCV
itself activating specific nuclear transcription factors couldinduce proinflammatory gene expression, as shown to occurwith other human hepatotropic virus such as hepatitis Bvirus (29).
As shown in this study, a high enrichment of bothCXCR3� and CCR5� T cells in the liver over the periph-eral blood compartment is a prominent feature in patientswith CHC, suggesting that these CK receptors could play akey role in the pathogenesis of HCV-induced liver disease.Further supporting this notion are the facts that CXCR3 andCCR5 are markers for T cells associated with Th1-typeinflammatory reactions (30), and that progressive liver in-jury in CHC is associated with increased intrahepatic pro-duction of Th1 cytokines (31).
Angiogenesis is an important event occurring in a numberof physiological and pathological conditions (32). In thisregard, we have described an angiogenic process that ismainly restricted to portal tracts in CHC (16). CK arebecoming recognized as factors involved in regulating an-giogenesis (33). Although the molecular basis for the an-giogenic or angiostatic effect of a number of CK remainsunclear, Romagnani et al. (19) recently provided evidencethat CXCR3 is expressed on human activated endothelialcells with a high proliferation rate and, more interestingly,that Mig blocked endothelial cell proliferation through
Figure 5. Two-color flow cytometry of peripheral blood and intrahepatic lymphocytes from a patient with CHC. The most striking findingwas the high enrichment of both CXCR3� and CCR5� T cells in the liver over the peripheral blood compartment.
Table 5. Comparative Analysis of Basal Characteristics onSustained Responders and Selected Nonresponders
Characteristic SR � 5 NR � 8
Age (yr) 35 (22–46) 38 (24–49)Sex (male/female) 3/2 5/3HCV genotype
1b 2 4Non-1b 3 4
HistologyGrade 2.4 (1–3) 2.7 (1–3)Stage 1.7 (1–2) 1.9 (1–2)
Intrahepatic CK scoreMig 1.9 (1–3) 2.2 (1–3)RANTES 2.1 (1–3) 2.3 (1–3)
Data are given as median (range). For all values, p � ns. Abbreviations as in Table3.
2868 Apolinario et al. AJG – Vol. 97, No. 11, 2002

CXCR3 binding. A novel finding of our study was thecolocalization of CXCR3 and its specific ligand Mig onneovessels within inflamed portal tracts in CHC, suggestingthat an angiostatic effect mediated by Mig/CXCR3 interac-tions may be operative in vivo during CHC. Whether thepotential angiostatic effect of some CXC CK, such as Migbut also IP-10 (34), may modulate the portal angiogenicprocess taking place during CHC deserves further investi-gation.
In conclusion, the data presented here provide evidencethat intrahepatic CK signaling may play a key role in re-cruiting effector T cells into the liver parenchyma as well asin regulating the portal angiogenic process that occurs inCHC. The results of this study open up the possibility ofnew therapeutic interventions based on CK activities,searching for an anti-inflammatory effect (blocking CK re-ceptors) or for an angiostatic effect (enhancing endothelialCXCR3 binding).
ACKNOWLEDGMENTS
This work was supported by grants to C.G-M from Institutode Salud Carlos III (FIS 01/0152) and from European Co-mission (QLRT-2000-02114). An unrestricted educationalgrant from Schering Plough Biotech is also acknowledged.The authors thank Rocha Barroso for secretarial help.
Reprint requests and correspondence: Carmelo Garcıa-Monzon,M.D., Hepatology Unit, University Hospital Santa Cristina, Mae-stro Vives, 2, 28009-Madrid, Spain.
Received Jan. 2, 2002; accepted June 14, 2002.
REFERENCES
1. Scheuer PJ. Classification of chronic viral hepatitis: A need forreassessment. J Hepatol 1991;13:372–4.
2. Gonzalez-Peralta RP, Davis GL, Lau JYN. Pathogeneticmechanisms of hepatocellular damage in chronic hepatitis Cvirus infection. J Hepatol 1994;21:255–9.
3. Moreno-Otero R, Garcıa-Buey L, Mateos FG, et al. Pathogen-esis of chronic viral hepatitis: Lessons from immunohisto-chemistry. Viral Hepatitis Rev 1996;2:61–79.
4. Di Bisceglie AM, Goodman ZD, Ishak KG, et al. Long-termclinical and histopathological follow-up of chronic post-trans-fusion hepatitis. Hepatology 1991;14:969–74.
5. Brunt EM. Grading and staging the histopathological lesionsof chronic hepatitis: The Knodell histology activity index andbeyond. Hepatology 2000;31:241–6.
6. Baggiolini M, Dewald B, Moser B. Human chemokines: Anupdate. Annu Rev Immunol 1997;15:675–705.
7. Rollins BJ. Chemokines. Blood 1997;90:909–28.8. Kunkel SLJ. Through the looking glass: The diverse in vivo
activities of chemokines. J Clin Invest 1999;104:1333–4.9. Bone-Larson C, Simpson KJ, Colleti LM, et al. The role of
chemokines in the immunopathology of the liver. ImmunolRev 2000;177:8–20.
10. Mackay CR. Chemokine receptors and T-cell chemotaxis. JExp Med 1996;184:799–804.
11. Murdoch C, Finn A. Chemokine receptors and their role in
inflammation and infectious diseases. Blood 2000;95:3032–43.
12. Narumi S, Tominaga Y, Tamaru M, et al. Expression ofIFN-inducible protein-10 in chronic hepatitis. J Immunol1997;158:5536–44.
13. Shields PL, Morland CM, Salmon M, et al. Chemokine andchemokine receptor interactions provide a mechanism for se-lective T cell recruitment to specific liver compartments withinhepatitis C-infected liver. J Immunol 1999;163:6236–43.
14. Kusano F, Tanaka Y, Marumo F, et al. Expression of C-Cchemokines is associated with portal and periportal inflamma-tion in the liver of patients with chronic hepatitis C. Lab Invest2000;80:415–22.
15. Chang KM, Rehermann B, Chisari FV. Immunopathology ofhepatitis C. Springer Semin Immunopathol 1997;19:57–68.
16. Garcıa-Monzon C, Sanchez-Madrid F, Garcıa-Buey L, et al.Vascular adhesion molecule expression in viral chronichepatitis: Evidence of neoangiogenesis in portal tracts. Gas-troenterology 1995;108:231–41.
17. Liao F, Rabin RL, Yannelli JR, et al. Human Mig chemokine:Biochemical and functional characterization. J Exp Med 1995;182:1301–14.
18. Wu L, Paxton WA, Kassam N, et al. CCR5 levels and expres-sion pattern correlate with infectability by macrophage-tropicHIV-1 in vitro. J Exp Med 1997;185:1681–91.
19. Romagnani P, Annunziato F, Lasagni L, et al. Cell cycle-dependent expression of CXC chemokine receptor 3 by endo-thelial cells mediates angiostatic activity. J Clin Invest 2001;107:53–63.
20. Young KK, Resnick R, Myers TH. Detection of hepatitis Cvirus RNA by a combined reverse-transcription-polymerasechain reaction assay. J Clin Microbiol 1993;31:882–6.
21. Leach L, Clark P, Lampugnani MG, et al. Immunoelectroncharacterization of the inter-endothelial junctions of humanterm placenta. J Cell Sci 1993;104:1073–81.
22. Garcıa-Monzon C, Martın-Perez E, Lo Iacono O, et al. Char-acterization of pathogenic and prognostic factors of nonalco-holic steatohepatitis associated with obesity. J Hepatol 2000;33:716–24.
23. Koziel MJ, Duddley D, Wong JT, et al. Intrahepatic cytotoxicT lymphocytes specific for hepatitis C virus in persons withchronic hepatitis. J Immunol 1992;149:3339–44.
24. Tseng CT, Miskovsky E, Houghton M, et al. Characterizationof liver T-cell receptor ��2b T cells obtained from individualschronically infected with hepatitis C virus (HCV): Evidencefor these cells playing a role in the liver pathology associatedwith HCV infections. Hepatology 2001;33:1312–20.
25. Cerny A, Chisari FV. Pathogenesis of chronic hepatitis C:Immunological features of hepatic injury and viral persistence.Hepatology 1999;30:595–601.
26. Lauer GM, Walker BD. Hepatitis C virus infection. N EnglJ Med 2001;345:41–52.
27. Wang JH, Devalia JL, Xia C, et al. Expression of RANTES byhuman bronchial epithelial cells in vitro and in vivo and theeffect of corticoids. Am J Respir Cell Mol Biol 1996;14:27–35.
28. Lin YL, Liu CC, Chuang JI, et al. Involvement of oxidativestress, NF-IL-6, and RANTES expression in Dengue-2-virus-infected human liver cells. Virology 2000;276:114–26.
29. Majano PL, Garcıa-Monzon C, Lopez-Cabrera M, et al. In-ducible nitric oxide synthase expression in chronic viralhepatitis: Evidence for a virus-induced gene upregulation.J Clin Invest 1998;101:1343–52.
30. Qin S, Rottman JB, Myers P, et al. The chemokine receptorsCXCR3 and CCR5 mark subsets of T cells associated withcertain inflammatory reactions. J Clin Invest 1998;101:746–54.
2869AJG – November, 2002 T Cell Chemokines and Chronic Hepatitis C

31. Napoli J, Bishop GA, McGuinness PH, et al. Progressive liverinjury in chronic hepatitis C infection correlates with increasedintrahepatic expression of Th1-associated cytokines. Hepatol-ogy 1996;24:759–66.
32. Folkman J. Angiogenesis in cancer, vascular, rheumatoid andother disease. Nat Med 1995;1:27–31.
33. Luster AD. Chemokines. Chemotactic cytokines that mediateinflammation. N Engl J Med 1998;338:436–45.
34. Luster AD, Greenberg SM, Leder P. The IP-10 chemokinebinds to a specific cell suface heparan sulfate shared withplatelet factor 4 and inhibits endothelial cell proliferation. JExp Med 1995;182:219–32.
2870 Apolinario et al. AJG – Vol. 97, No. 11, 2002
Copyright © 2022 FDOKUMEN




















