
Increased density and synapto-protective effect of adenosine A2A receptors upon sub-chronic...
7
INCREASED DENSITY AND SYNAPTO-PROTECTIVE EFFECT OF ADENOSINE A 2A RECEPTORS UPON SUB-CHRONIC RESTRAINT STRESS G. M. A. CUNHA, a,b1 P. M. CANAS, a1 C. R. OLIVEIRA a AND R. A. CUNHA a * a Center for Neuroscience of Coimbra, Institute of Biochemistry, Fac- ulty of Medicine, University of Coimbra, 3004-504 Coimbra, Portugal b Department of Physiology and Pharmacology, Federal University of Ceará, 60430-270, Fortaleza, Brazil Abstract—Stress initially causes adaptive changes in the brain and can lead to neurodegeneration if continuously present. Noxious brain conditions trigger the release of adenosine that can control brain function and neurodegeneration through in- hibitory A 1 and facilitatory A 2A receptors. We tested the effect of restraint stress on the density of adenosine receptors and their effect on the outcome of stress, focusing in a known affected region, the hippocampus. Sub-chronic restraint stress (6 h/day for 7 days) caused a parallel decrease of the density of A 1 receptors (15–20%) and an increase (near 250%) of A 2A receptor density in rat hippocampal nerve terminals. This indicates that sub-chronic stress unbalances adenosine receptors, up-reg- ulating A 2A and down-regulating A 1 receptors. Sub-chronic stress did not cause hippocampal neurodegeneration but decreased the immunoreactivity (immunohistochemistry and Western blot) of a synaptic marker, synaptophysin. The blockade of A 2A receptors with 7-(2-phenylethyl)-5-amino-2-(2- furyl)-pyrazolo-[4,3-e]-1,2,4-triazolo[1,5-c]pyrimidine (0.05 mg/ kg, daily i.p. injection) attenuated the loss of synaptophysin immunoreactivity observed in the hippocampus of rats sub- jected to sub-chronic restraint stress. This ability of A 2A recep- tor antagonists to prevent the earliest stress-induced synap- tic modifications provides a neurochemical and morpholog- ical correlate for the interest of A 2A receptor antagonists to attenuate the burden of chronic stress. © 2006 IBRO. Pub- lished by Elsevier Ltd. All rights reserved. Key words: stress, adenosine, A 1 receptor, A 2A receptor, hippocampus, nerve terminals. Long lasting stress increases the activity of the hypo- thalamic–pituitary–adrenal axis (HPA) and causes cogni- tive impairment both in humans as well as in animal mod- els, mainly as a result of morphological and functional changes occurring in hippocampal circuits (reviewed in Kim and Diamond, 2002; McEwen, 1999). Although hip- pocampal atrophy and neuron loss is observed in humans and primates subject to stress (Uno et al., 1989; e.g. Sheline et al., 1996), the stress-induced cognitive defects in rodents correlate better with an atrophy and cyto-archi- tectural changes of CA3 apical dendrites (e.g. Magariños and McEwen, 1995; Magariños et al., 1997; Sousa et al., 2000; Stewart et al., 2005). Chronic stress also triggers a loss of synapses (Sandi et al., 2003; Sousa et al., 2000) typified by the loss of synaptic markers, like synaptophysin (Thome et al., 2001; but see Rosenbrock et al., 2005). Adenosine is a brain neuromodulator that increases its extracellular concentration in noxious brain conditions (re- viewed in Fredholm et al., 2005), namely during stress (e.g. Scaccianoce et al., 1989). Adenosine can either fa- cilitate or inhibit neuronal activity by activation of metabo- tropic A 1 receptors (inhibitory) or facilitatory A 2A receptors (reviewed in Fredholm et al., 2005), both of which are predominantly located in synapses in the limbic and neo- cortex (Rebola et al., 2003, 2005). Furthermore, it is well established that the activation of A 1 receptors confers neuroprotection against acute noxious brain stimuli (de Mendonça et al., 2000), whereas the blockade of A 2A receptors affords a robust neuroprotection in chronic nox- ious conditions in the adult brain (Cunha, 2005). Interest- ingly, A 2A receptor antagonists have been proposed as potential anti-depressants (El Yacoubi et al., 2003). This is in accordance with the ability of adenosine to control cor- ticotrophin and cortisol/corticosterone release (Chau et al., 1999; Geiger and Glavin, 1985; Jegou et al., 2003; Scac- cianoce et al., 1989) and to modulate hippocampal excit- ability (Fredholm et al., 2005), a key region in the control of HPA (Herman and Cullinan, 1997). However, the setup of adenosine receptors is dynamic and is known to be mod- ified by chronic brain insults (reviewed in Cunha, 2005). Given that glucocorticoids modify the expression of A 1 receptors (Gerwins and Fredholm, 1991; Svenningsson and Fredholm, 1997; Ren and Stiles, 1999) and chronic stress modifies the A 1 receptor-mediated anti-nociceptive response (Silva-Torres et al., 2003), we investigated the consequences of restraint stress on the set up of both A 1 and A 2A receptors in the hippocampus to test if the manip- ulation of any particular adenosine receptors would be potentially beneficial to prevent the early modifications caused by restraint stress in the hippocampus. EXPERIMENTAL PROCEDURES Male Wistar rats (6 – 8 weeks old, 140 –160 g, obtained from Charles River, Barcelona, Spain) were used throughout this study 1 Contributed equally to the experiments. *Corresponding author. Tel: 351-239-820-190; fax: 351-239-822-776. E-mail address: [email protected]; URL: http://cnc.cj.uc.pt/lab_lef (R. A. Cunha). Abbreviations: ANOVA, analysis of variance; B max , maximal number of binding sites; DPCPX, 1,3-dipropyl-8-cyclopentylxanthine; HPA, hypo- thalamic–pituitary–adrenal; K D , dissociation constant; PBS, phosphate buffer saline; SCH 58261, 7-(2-phenylethyl)-5-amino-2-(2-furyl)-pyra- zolo-[4,3-e]-1,2,4-triazolo[1,5-c]pyrimidine; TBS, Trizma buffer saline; TBS-T, Tris-buffered saline, pH 7.6 containing 0.1% Tween 20; XAC, 8-{4-[(2-aminoethyl)amino]carbonylmethyl-oxyphenyl}xanthine. Neuroscience 141 (2006) 1775–1781 0306-4522/06$30.000.00 © 2006 IBRO. Published by Elsevier Ltd. All rights reserved. doi:10.1016/j.neuroscience.2006.05.024 1775
Transcript of Increased density and synapto-protective effect of adenosine A2A receptors upon sub-chronic...

IOR
GAa
ub
C
AaNchrerfrdsusdWbfkijttial
Kh
LttecK1
*ECAbtbzT8
Neuroscience 141 (2006) 1775–1781
0d
NCREASED DENSITY AND SYNAPTO-PROTECTIVE EFFECTF ADENOSINE A2A RECEPTORS UPON SUB-CHRONIC
ESTRAINT STRESSpaSita2lt(
ev(ct(pcenMriipit1caHaiGrasrcaupc
M
. M. A. CUNHA,a,b1 P. M. CANAS,a1 C. R. OLIVEIRAa
ND R. A. CUNHAa*
Center for Neuroscience of Coimbra, Institute of Biochemistry, Fac-lty of Medicine, University of Coimbra, 3004-504 Coimbra, Portugal
Department of Physiology and Pharmacology, Federal University ofeará, 60430-270, Fortaleza, Brazil
bstract—Stress initially causes adaptive changes in the brainnd can lead to neurodegeneration if continuously present.oxious brain conditions trigger the release of adenosine thatan control brain function and neurodegeneration through in-ibitory A1 and facilitatory A2A receptors. We tested the effect ofestraint stress on the density of adenosine receptors and theirffect on the outcome of stress, focusing in a known affectedegion, the hippocampus. Sub-chronic restraint stress (6 h/dayor 7 days) caused a parallel decrease of the density of A1
eceptors (15–20%) and an increase (near 250%) of A2A receptorensity in rat hippocampal nerve terminals. This indicates thatub-chronic stress unbalances adenosine receptors, up-reg-lating A2A and down-regulating A1 receptors. Sub-chronictress did not cause hippocampal neurodegeneration butecreased the immunoreactivity (immunohistochemistry andestern blot) of a synaptic marker, synaptophysin. The
lockade of A2A receptors with 7-(2-phenylethyl)-5-amino-2-(2-uryl)-pyrazolo-[4,3-e]-1,2,4-triazolo[1,5-c]pyrimidine (0.05 mg/g, daily i.p. injection) attenuated the loss of synaptophysinmmunoreactivity observed in the hippocampus of rats sub-ected to sub-chronic restraint stress. This ability of A2A recep-or antagonists to prevent the earliest stress-induced synap-ic modifications provides a neurochemical and morpholog-cal correlate for the interest of A2A receptor antagonists tottenuate the burden of chronic stress. © 2006 IBRO. Pub-ished by Elsevier Ltd. All rights reserved.
ey words: stress, adenosine, A1 receptor, A2A receptor,ippocampus, nerve terminals.
ong lasting stress increases the activity of the hypo-halamic–pituitary–adrenal axis (HPA) and causes cogni-ive impairment both in humans as well as in animal mod-ls, mainly as a result of morphological and functionalhanges occurring in hippocampal circuits (reviewed inim and Diamond, 2002; McEwen, 1999). Although hip-
Contributed equally to the experiments.Corresponding author. Tel: �351-239-820-190; fax: �351-239-822-776.-mail address: [email protected]; URL: http://cnc.cj.uc.pt/lab_lef (R. A.unha).bbreviations: ANOVA, analysis of variance; Bmax, maximal number ofinding sites; DPCPX, 1,3-dipropyl-8-cyclopentylxanthine; HPA, hypo-halamic–pituitary–adrenal; KD, dissociation constant; PBS, phosphateuffer saline; SCH 58261, 7-(2-phenylethyl)-5-amino-2-(2-furyl)-pyra-olo-[4,3-e]-1,2,4-triazolo[1,5-c]pyrimidine; TBS, Trizma buffer saline;
CBS-T, Tris-buffered saline, pH 7.6 containing 0.1% Tween 20; XAC,-{4-[(2-aminoethyl)amino]carbonylmethyl-oxyphenyl}xanthine.
306-4522/06$30.00�0.00 © 2006 IBRO. Published by Elsevier Ltd. All rights reseroi:10.1016/j.neuroscience.2006.05.024
1775
ocampal atrophy and neuron loss is observed in humansnd primates subject to stress (Uno et al., 1989; e.g.heline et al., 1996), the stress-induced cognitive defects
n rodents correlate better with an atrophy and cyto-archi-ectural changes of CA3 apical dendrites (e.g. Magariñosnd McEwen, 1995; Magariños et al., 1997; Sousa et al.,000; Stewart et al., 2005). Chronic stress also triggers a
oss of synapses (Sandi et al., 2003; Sousa et al., 2000)ypified by the loss of synaptic markers, like synaptophysinThome et al., 2001; but see Rosenbrock et al., 2005).
Adenosine is a brain neuromodulator that increases itsxtracellular concentration in noxious brain conditions (re-iewed in Fredholm et al., 2005), namely during stresse.g. Scaccianoce et al., 1989). Adenosine can either fa-ilitate or inhibit neuronal activity by activation of metabo-ropic A1 receptors (inhibitory) or facilitatory A2A receptorsreviewed in Fredholm et al., 2005), both of which areredominantly located in synapses in the limbic and neo-ortex (Rebola et al., 2003, 2005). Furthermore, it is wellstablished that the activation of A1 receptors conferseuroprotection against acute noxious brain stimuli (deendonça et al., 2000), whereas the blockade of A2A
eceptors affords a robust neuroprotection in chronic nox-ous conditions in the adult brain (Cunha, 2005). Interest-ngly, A2A receptor antagonists have been proposed asotential anti-depressants (El Yacoubi et al., 2003). This is
n accordance with the ability of adenosine to control cor-icotrophin and cortisol/corticosterone release (Chau et al.,999; Geiger and Glavin, 1985; Jegou et al., 2003; Scac-ianoce et al., 1989) and to modulate hippocampal excit-bility (Fredholm et al., 2005), a key region in the control ofPA (Herman and Cullinan, 1997). However, the setup ofdenosine receptors is dynamic and is known to be mod-
fied by chronic brain insults (reviewed in Cunha, 2005).iven that glucocorticoids modify the expression of A1
eceptors (Gerwins and Fredholm, 1991; Svenningssonnd Fredholm, 1997; Ren and Stiles, 1999) and chronictress modifies the A1 receptor-mediated anti-nociceptiveesponse (Silva-Torres et al., 2003), we investigated theonsequences of restraint stress on the set up of both A1
nd A2A receptors in the hippocampus to test if the manip-lation of any particular adenosine receptors would beotentially beneficial to prevent the early modificationsaused by restraint stress in the hippocampus.
EXPERIMENTAL PROCEDURES
ale Wistar rats (6–8 weeks old, 140–160 g, obtained from
harles River, Barcelona, Spain) were used throughout this studyved.

aentcb
R
Twipparcaas1witdqbrt
t2bndpthfpip
aggcuwpflsparCw
Pn
Tat2nTac
rKHt(sp4tbW
B
H2tfitawoduo(docAiAEtva23
dbSbaRtdth4acocb2
nR(a
W
Trdt
G. M. A. Cunha et al. / Neuroscience 141 (2006) 1775–17811776
nd were handled in accordance with the EU guidelines for use ofxperimental animals (86/609/EEC), with care to minimize theumber of animals used and their suffering. The rats were anes-hetized under halothane atmosphere before being killed by de-apitation or deeply anesthetized with pentobarbital (100 mg/kg)efore being dissected.
estraint stress model
he stress model used consisted of a physical restraint of the rats,hich was carried out in a new environment. Thus, the rats were
ndividually placed in a room adjacent to their colony in an inde-endent plastic compartment and immobilized in a 25�7 cmlastic bottle, with a plastic taper on the outside and a 1 cm holet one end for breathing. After the termination of each dailyestraint stress session, the rats were returned to their homeages, where they were left undisturbed under constant temper-ture (22�2 °C) and light cycles (12-h light/dark cycle) with freeccess to food and water. The schedule of sub-chronic restrainttress consisted in a daily 6 h immobilization period (between0:00 AM and 4:00 PM) during seven consecutive days. The ratsere weighed daily and killed 24 h after the last session of
mmobilization. Control age-matched rats were handled as theirested littermates except that they were not immobilized. Theecision to carry out the evaluation of the neurochemical conse-uences of sub-chronic restraint stress 24 h after the last immo-ilization period was dictated by the need of isolating the effects ofepeated sub-chronic stress from these of acute stress caused byhe last period of immobilization.
When rats were treated with the selective A2A receptor an-agonist 7-(2-phenylethyl)-5-amino-2-(2-furyl)-pyrazolo-[4,3-e]-1,,4-triazolo[1,5-c]pyrimidine (SCH 58261) (generously providedy Scott Weiss, Vernalis, UK), this drug was injected intraperito-eally (300–350 �l) at a dose of 0.05 mg/kg (dissolved in 10%imethylsulphoxide in saline) 30 min before each immobilizationeriod. We chose to administer SCH 58261 intraperitoneally sincehis route of administration of this particular dose of SCH 58261as previously been shown to afford neuroprotection against dif-erent types of neuronal injuries (reviewed in Cunha, 2005) withouteripheral effects (Ongini, 1997). The control rats were injected
ntraperitoneally with 300–350 �l saline with 10% dimethylsul-hoxide.
The rats were anesthetized under halothane atmosphere andblood sample was collected by cardiac puncture to measure
lucose concentration by the glucose oxidase method using alucometer (Elite, Bayer, Lisbon, Portugal). The plasma levels oforticosterone were determined by fluorimetry (Mattingly, 1962),sing the same blood sample. Briefly, 1 ml of plasma was mixedith 7.5 ml dichloromethane, shaken, centrifuged to separate thehases and the plasma layer removed. At time 0, 2.5 ml of theuorescent derivatization reagent (seven volumes of concentratedulfuric acid with three volumes of ethanol) was added to thelasma layer. After shaking for 2 s, the supernatant was removednd transferred to cuvettes for reading at 12 min. The fluorometriceadings were taken at 530 nm emission and 470 nm excitation.orticosterone (1.25 mg/ml, from Sigma-Aldrich, Sintra, Portugal)as used to estimate the standard curve.
reparation of membranes from hippocampalerve terminals
he rats were then killed by decapitation after the halothanenesthesia. Membranes from Percoll-purified hippocampal synap-osomes were prepared as previously described (Rebola et al.,003). Briefly, the two hippocampi from one rat were homoge-ized at 4 °C in sucrose solution (0.32 M) containing 50 mMris–HCl, 2 mM EGTA and 1 mM dithiothreitol, pH 7.6, centrifugedt 3000�g for 10 min at 4 °C. The supernatants were collected,
entrifuged at 14,000�g for 20 min at 4 °C and the pellet was desuspended in 1 ml of a 45% (v/v) Percoll solution made up in arebs solution (composition 140 mM NaCl, 5 mM KCl, 25 mMEPES, 1 mM EDTA, 10 mM glucose, pH 7.4). After centrifuga-
ion at 14,000�g for 2 min at 4 °C, the top layer was removedsynaptosomal fraction), washed in 1 ml Krebs solution and re-uspended in 10 ml of incubation buffer (25 mM Tris, 2 mM MgCl2,H 7.4). This mixture was centrifuged at 14,000�g for 20 min at°C and the pellet corresponded to the membranes of nerve
erminals. This pellet was either resuspended in the incubationuffer for binding studies or in SDS-PAGE buffer (see below) forestern blot analysis.
inding studies
ippocampal nerve terminal membranes were first incubated withU/ml adenosine deaminase (Sigma-Aldrich) for 30 min at 37 °C,
o remove endogenous adenosine. The mixture was then centri-uged at 14,000�g for 10 min at 4 °C and the pellets resuspendedn the incubation buffer. Binding of the selective A1 receptor an-agonist, 3H-1,3-dipropyl-8-cyclopentylxanthine (DPCPX) (specificctivity of 109.0 Ci/mmol; from DuPont NEN, Anagene, Portugal)as for 2 h at 37 °C, with 92–126 �g of protein in a final volumef 300 �l in the incubation solution containing 2 U/ml adenosineeaminase, as previously described (Rebola et al., 2003). A sat-ration curve was constructed using five different concentrationsf 3H-DPCPX (0.6, 1, 3, 6, and 20 nM), as previously carried outRebola et al., 2003; Coelho et al., 2006). To evaluate the bindingensity of A2A receptors, we used 3H-SCH 58261, which is thenly A2A receptor ligand that has been shown to selectively bindortical A2A receptors (Lopes et al., 2004) and devoid of effects on1 receptors, such as ZM 241385 (see Lopes et al., 1999). Bind-
ng of 3H-SCH 58261 (specific activity of 77 Ci/mmol; prepared bymersham, Buckinghamshire, UK and generously offered by Dr.nnio Ongini, Shering-Plough, Milan, Italy) was for 1 h at room
emperature (23–25 °C) with 162–214 �g of protein in a finalolume of 300 �l in the incubation solution containing 4 U/mldenosine deaminase, as previously described (Rebola et al.,005). A single supra-maximal and selective concentration ofH-SCH 58261 (Lopes et al., 2004) was used to evaluate theensity of A2A receptors as previously done (Duarte et al., 2006)ecause of the limited availability of this custom-made compound.pecific binding was determined by subtraction of the non-specificinding, which was measured in the presence of 1 �M 8-{4-[(2-minoethyl)amino]carbonylmethyl-oxyphenyl}xanthine (XAC) (fromesearch Biochemical Inc., Sigma-Aldrich), a mixed A1/A2 recep-
or antagonist. Each binding assay data point was performed inuplicate. The binding reactions were stopped by vacuum filtrationhrough glass fiber filters (GF/C filters) using a 24 well Brandelarvester. The filters were then placed in scintillation vials andml of scintillation liquid (Ready Safe, Pharmacia-Portugal) was
dded. Radioactivity was determined after at least 12 h with aounting efficiency of 55–60%. To ensure an error lower than 5%f counts, the samples were counted for 10 min. The proteinoncentration was determined using the Bio-Rad protein assayased on Bradford dye-binding procedure (see Rebola et al.,003, 2005).
The specific binding from saturation experiments was fitted byon-linear regression to a one binding site equation using theaphson-Newton method, performed with a commercial software
GraphPad Inplot) to determine the binding parameters (dissoci-tion constant, KD, and maximal number of binding sites, Bmax).
estern blot analysis
he analysis of adenosine A1 receptor immunoreactivity was car-ied out as previously described (Rebola et al., 2003). Briefly, afteretermining the amount of protein, each sample was diluted withwo volumes of SDS-PAGE buffer containing 8 M urea, 100 mM
ithiothreitol, 2% (w/v) sodium dodecyl sulfate and 375 mM Tris–
HasudbsbaBawbaCdThtuq
iaabtaib
I
Thcprps14csiwa�cle
wctmbtscd
wsrwprw
(Urm(aL
icwwtt1mt4iwdcrfpmesd(ltcwC
S
Doe(
Tpss(ae1mpdicwiro
G. M. A. Cunha et al. / Neuroscience 141 (2006) 1775–1781 1777
Cl pH 6.8 and incubated for 2 h at 37 °C. These diluted samplesnd the pre-stained molecular weight markers (Amersham) wereeparated by SDS-PAGE (10% with a 4% concentrating gel)nder reducing conditions and electro-transferred to polyvinyli-ene difluoride membranes (0.45 �m, from Amersham). Afterlocking for 2 h at room temperature with 5% milk in Tris-bufferedaline, pH 7.6 containing 0.1% Tween 20 (TBS-T), the mem-ranes were incubated overnight at 4 °C with either a rabbitnti-adenosine A1 receptor antibody (1:750 dilution from Affinityioreagents, Golden, CA, USA) or a mouse anti-synaptophysinntibody (1:1000 dilution from Sigma-Aldrich). After four 10 minashing periods with TBS-T containing 0.5% milk, the mem-ranes were incubated with the alkaline phosphatase-conjugatednti-goat or anti-mouse secondary antibody (1:2000 dilution, fromalbiochem, Darmstadt, Germany) in TBS-T containing 1% milkuring 90 min at room temperature. After five 10 min-washes inBS-T with 0.5% milk, the membranes were incubated with En-anced Chemi-Fluorescence (Amersham) during five minutes andhen analyzed with a VersaDoc 3000 (Biorad, Hercules, CA, USA)sing the Quantity One software (Biorad) for the densitometricuantification of the bands.
The membranes were then re-probed and tested for �-tubulinmmunoreactivity to confirm that similar amounts of protein werepplied to the gels. Briefly, the membranes were incubated for 1 ht room temperature with a 0.1 M glycine buffer (pH 7.2) thenlocked as previously described before incubation with an anti-�-ubulin antibody (1:1000 from Zymed Laboratories, Alfredo Cav-lheiro, Lisbon, Portugal). The membranes were then washed and
ncubated with alkaline phosphatase-conjugated secondary anti-ody as described above.
mmunohistochemistry
he rats were anesthetized with pentobarbital (100 mg/kg), theeart was exposed and, after clamping the descending aorta, aatheter was inserted in the ascending aorta. The animal was thenerfused with saline with 4% sucrose (200 ml) while opening theight atria to allow the outflow of the perfusate. Rats were thenerfused with 200 ml of 4% paraformaldehyde in saline with 4%ucrose. After its fixation, the brain was removed, maintained for2 h in the same paraformaldehyde solution and subsequently for8 h in a phosphate buffer saline (PBS) solution with 30% su-rose. The brain was then frozen in dry ice and 20 �m coronalections were prepared using a cryostat. The sections were storedn PBS with 0.01% sodium azide until mounting in slides coatedith 2% gelatin with chromium and potassium sulfate. After dryingt room temperature, the mounted sections were stored at20 °C. Consecutive sections from each animal were used to
arry out the three histochemical procedures described below. Ateast four groups of sections from each animal were used andach group was processed independently in different days.
The general neuronal morphology in hippocampal sectionsas evaluated by using a Cresyl Violet staining of Nissl bodies,arried out as previously described (Lopes et al., 2003). Briefly,he sections were dried at room temperature and stained for 10in with Cresyl Violet solution (Sigma-Aldrich; 0.5% in acetateuffer). The sections were then washed twice with acetate buffer,wice in 100% ethanol before being cleared with xylene. Nissltaining was evaluated qualitatively by inspection of the pyramidalell layer of both CA1 and CA3 regions by at least two indepen-ent researchers.
To evaluate the presence of degenerated neurons, sectionsere stained with FluoroJade-C (Schmued et al., 2005). The brainections were immersed in 0.1% NaOH in 80% ethanol for 5 min,insed for 2 min in 70% ethanol and then for 2 min in distilledater. The slides were then transferred to a solution of 0.06%otassium permanganate for 10 min and were gently shaken on aotating platform. The slides were then rinsed for 2 min in distilled
ater and incubated with the FluoroJade staining solution a0.0001% FluoroJade-C, from Histo-Chem Inc., Jefferson, AR,SA), being gently shaken for 10 min. The sections were then
insed three times for 1 min with distilled water, dried and im-ersed in xylene and then coverslipped with DPX Mountant
Fluka Biochemika, Sigma-Aldrich) before being visualized underZeiss Axiovert 200 inverted fluorescence microscope (Hitec,
isbon, Portugal) equipped with a cooled CCD camera.The detection of nerve terminals was carried out using an
mmunohistochemical detection of synaptophysin, a protein lo-ated in synaptic vesicles (Masliah and Terry, 1993). The sectionsere first rinsed for 5 min in PBS and then three times for 5 minith Trizma buffer saline (TBS) (0.05 M Trizma base buffer con-
aining 150 mM of NaCl, pH 7.2) at room temperature. The sec-ions were blocked with TBS containing 0.2% Triton X-100 and0% goat serum during 45 min, incubated in the presence of theouse anti-synaptophysin antibody (1:500 dilution in TBS con-
aining 0.2% Triton X-100 and 10% normal goat serum) for 72 h at°C, rinsed three times for 10 min in TBS and subsequently
ncubated with goat anti-mouse secondary antibody conjugatedith a fluorophore (Alexa Fluor 488; from Molecular Probes, Lei-en, The Netherlands) (1:100 dilution in 0.1 M phosphate bufferontaining 0.2% Triton X-100 and 10% normal serum) for 2 h atoom temperature. After rinsing twice for 10 min in TBS and onceor 10 min in distilled water, the sections were dehydrated andassed through xylol before mounting on slides, using Vectashieldounting medium (Vector Laboratories, Batista Marques) andxamined under a fluorescence microscope or a confocal micro-cope. The intensity of synaptophysin labeling was estimated byetermining the total fluorescence readout in pre-defined areas20�20 �m) with at least three different areas in the CA3 stratumucidum being estimated per section. As an additional semi-quan-itative way of estimating synaptophysin immunoreactivity, we alsoounted the number of dots present in these pre-defined areasith the help of the MetaFluor 5.0 software (Universal Imagingorporation, West Chester, PA, USA).
tatistics
ata are mean�S.E.M. values or mean (95% confidence interval)f n experiments. Significance was considered at P�0.05 usingither a Student’s t-test or a one-way analysis of varianceANOVA) followed by post hoc Duncan or Newman-Keuls tests.
RESULTS
he blood levels of glucose (107.2�7.5 mg/dl) and thelasma corticosterone levels (117.8�12.3 �g/ml) mea-ured 24 h after the last restraint stress session were notignificantly (P�0.05) different than in control animals103.1�7.3 mg/dl and 98.6�1.2 �g/ml, respectively, n�6),lthough they were elevated (P�0.05) immediately at thend of the last immobilization period (glucose levels:27.7�7.6 mg/dl; corticosterone levels: 204.4�17.3 �g/l, respectively, n�3–4). However, 24 h after the lasteriod of immobilization, the sub-chronically stressed ratsisplayed a significant (P�0.05) lower gain of weight dur-
ng the 7 days of testing (15.8�6.9 g, n�6) compared withontrol rats (44.7�3.8 g), as well as a being the only grouphere gastric ulcers were observed (not shown). This
ndicates that a period of 24 h after the last period ofestraint stress was sufficient to wipe out the acute effectsf the last immobilization period and allows studying the
ccumulated effects of the repeated sub-chronic stress.
MA
Thw
1ibb((inwtiPrTasd5f(
SA
IchehjoapnFhcc
FdptcssbPlhriac*N
FepssmrueoA
G. M. A. Cunha et al. / Neuroscience 141 (2006) 1775–17811778
odification of the density of hippocampal
1 and A2A receptors
he selective A1 receptor antagonist, DPCPX, bound toippocampal nerve terminal membranes of control ratsith a Bmax of 961�26 fmol/mg protein and a KD of
ig. 1. Sub-chronic restraint stress (6 h daily for 7 days, sub-chronic)ecreases the density of adenosine A1 receptors in the rat hippocam-us. Panel A displays average saturation binding curves of the selec-ive A1 receptor antagonist 3H-DPCPX in hippocampal membranes ofontrol and sub-chronically stressed rats. The ordinates represent thepecific binding of 3H-DPCPX, obtained upon subtraction of the non-pecific binding, determined in the presence of 1 �M XAC, from totalinding. The results are mean�S.E.M. of four to five experiments.anel B displays a Western blot from one experiment (50 �g of protein
oaded in each lane) comparing the A1 receptor immunoreactivity inippocampal membranes from control and sub-chronically stressedats, as well as the average results (mean�S.E.M.) of A1 receptormmunoreactivity in four to five such experiments. In each Western blotnalysis, a re-probing for detection of �-tubulin was carried out toonfirm that similar amounts of protein were applied to the gel.
sP�0.05 compared with control (one-way ANOVA followed by
ewman-Keuls test).
.15 nM (95% confidence interval: 0.77–2.53 nM, n�5). Asllustrated in Fig. 1A, there was a lower (P�0.05) density ofinding of 3H-DPCPX in hippocampal nerve terminal mem-ranes derived from rats subjected to sub-chronic stressBmax of 773�27 fmol/mg protein) without change in KD
0.88 nM, 95% confidence interval: 0.62–1.20 nM, n�4),ndicating that hippocampal A1 receptors in nerve termi-als are down-regulated upon sub-chronic stress. Thisas confirmed by using Western blot analysis of A1 recep-
or immunoreactivity. As shown in Fig. 1B, the A1 receptormmunoreactivity was lower (15.2�3.6% lower, n�4,�0.05) in hippocampal nerve terminal membranes from
ats subject to sub-chronic stress compared with controls.he opposite was observed in relation to hippocampaldenosine A2A receptors, which density increased afterub-chronic stress. As illustrated in Fig. 2, the bindingensity of the selective A2A receptor antagonist, 3H-SCH8261 (10 nM) in hippocampal nerve terminal membranesrom rats subjected to sub-chronic stress was significantlyP�0.05) larger than that present in control rats (n�5).
ynapto-protective effects afforded by adenosine
2A receptor blockade
n accordance with previous studies indicating that sub-hronic restraint stress does not cause neurotoxicity in theippocampus (e.g. Magariños and McEwen, 1995; Sandit al., 2003; Sousa et al., 2000; Stewart et al., 2005), theistological analysis of hippocampal sections of rats sub-
ect to sub-chronic stress revealed a Cresyl Violet stainingf Nissl bodies in the pyramidal cell layer of the cornummonis qualitatively similar to that present in hippocam-al sections from control rats (data not shown). This lack ofeuronal damage was further confirmed by the lack ofluoroJade-C staining in either CA1 or CA3 regions inippocampal sections derived from rats subjected to sub-hronic stress (data not shown). As a positive control, weonfirmed the presence of an evident decrease of Nissl
ig. 2. Sub-chronic restraint stress (6 h daily for 7 days, sub-chronic)nhances the density of adenosine A2A receptors in the rat hippocam-us. The density of A2A receptors was estimated by comparing thepecific binding of a supra-maximal concentration (10 nM) of theelective A2A receptor antagonist 3H-SCH 58261 in hippocampalembranes of control and sub-chronically stressed rats. The ordinates
epresent the specific binding of 3H-SCH 58261 (10 nM), obtainedpon subtraction of the non-specific binding, determined in the pres-nce of 1 �M XAC, from total binding. The results are mean�S.E.M.f four to five experiments. * P�0.05 compared with control (one-wayNOVA followed by Newman-Keuls test).
taining and the appearance of FluoroJade-C stained py-
ras
sdTtii3flttcccamhs
hsp(rcwitscidstcwttw
Tthstmcu
uanfe
i
Ftirtmlfcdr(secnppirStreow*N
G. M. A. Cunha et al. / Neuroscience 141 (2006) 1775–1781 1779
amidal cells in sections derived from rats killed 7 daysfter an i.p. injection of 10 mg/kg of kainate (data nothown).
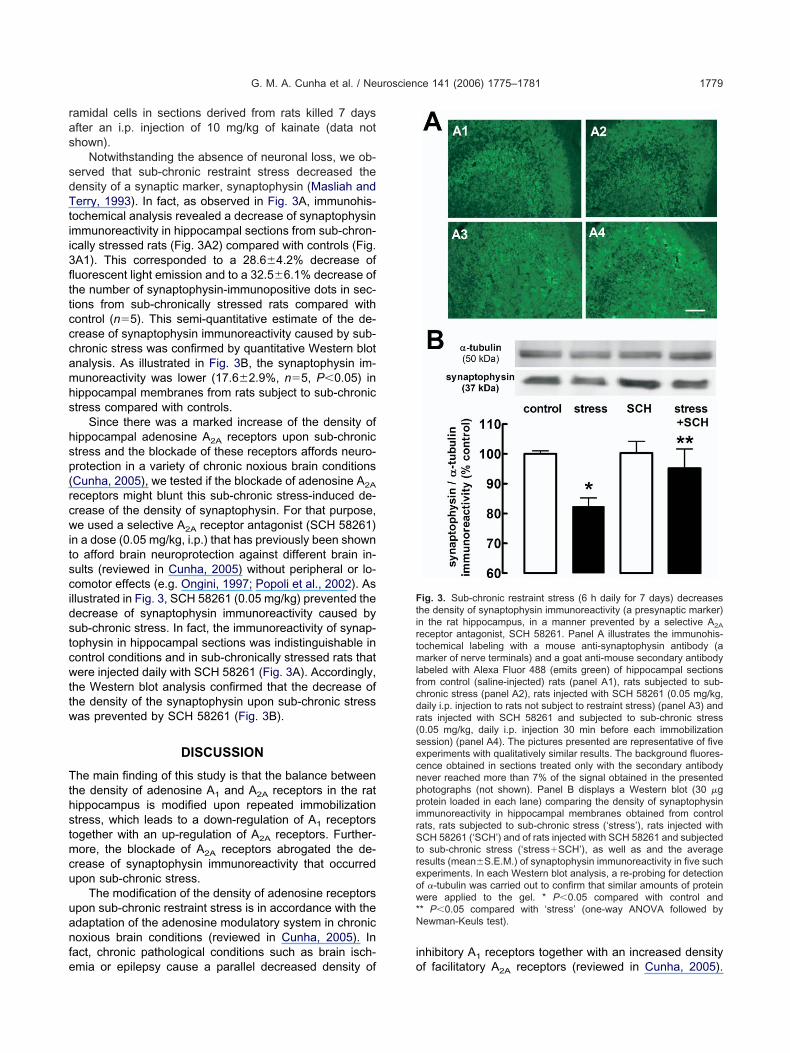
Notwithstanding the absence of neuronal loss, we ob-erved that sub-chronic restraint stress decreased theensity of a synaptic marker, synaptophysin (Masliah anderry, 1993). In fact, as observed in Fig. 3A, immunohis-
ochemical analysis revealed a decrease of synaptophysinmmunoreactivity in hippocampal sections from sub-chron-cally stressed rats (Fig. 3A2) compared with controls (Fig.A1). This corresponded to a 28.6�4.2% decrease ofuorescent light emission and to a 32.5�6.1% decrease ofhe number of synaptophysin-immunopositive dots in sec-ions from sub-chronically stressed rats compared withontrol (n�5). This semi-quantitative estimate of the de-rease of synaptophysin immunoreactivity caused by sub-hronic stress was confirmed by quantitative Western blotnalysis. As illustrated in Fig. 3B, the synaptophysin im-unoreactivity was lower (17.6�2.9%, n�5, P�0.05) inippocampal membranes from rats subject to sub-chronictress compared with controls.
Since there was a marked increase of the density ofippocampal adenosine A2A receptors upon sub-chronictress and the blockade of these receptors affords neuro-rotection in a variety of chronic noxious brain conditionsCunha, 2005), we tested if the blockade of adenosine A2A
eceptors might blunt this sub-chronic stress-induced de-rease of the density of synaptophysin. For that purpose,e used a selective A2A receptor antagonist (SCH 58261)
n a dose (0.05 mg/kg, i.p.) that has previously been showno afford brain neuroprotection against different brain in-ults (reviewed in Cunha, 2005) without peripheral or lo-omotor effects (e.g. Ongini, 1997; Popoli et al., 2002). Asllustrated in Fig. 3, SCH 58261 (0.05 mg/kg) prevented theecrease of synaptophysin immunoreactivity caused byub-chronic stress. In fact, the immunoreactivity of synap-ophysin in hippocampal sections was indistinguishable inontrol conditions and in sub-chronically stressed rats thatere injected daily with SCH 58261 (Fig. 3A). Accordingly,
he Western blot analysis confirmed that the decrease ofhe density of the synaptophysin upon sub-chronic stressas prevented by SCH 58261 (Fig. 3B).
DISCUSSION
he main finding of this study is that the balance betweenhe density of adenosine A1 and A2A receptors in the ratippocampus is modified upon repeated immobilizationtress, which leads to a down-regulation of A1 receptorsogether with an up-regulation of A2A receptors. Further-ore, the blockade of A2A receptors abrogated the de-
rease of synaptophysin immunoreactivity that occurredpon sub-chronic stress.
The modification of the density of adenosine receptorspon sub-chronic restraint stress is in accordance with thedaptation of the adenosine modulatory system in chronicoxious brain conditions (reviewed in Cunha, 2005). In
act, chronic pathological conditions such as brain isch-
mia or epilepsy cause a parallel decreased density of onhibitory A receptors together with an increased density
ig. 3. Sub-chronic restraint stress (6 h daily for 7 days) decreaseshe density of synaptophysin immunoreactivity (a presynaptic marker)n the rat hippocampus, in a manner prevented by a selective A2A
eceptor antagonist, SCH 58261. Panel A illustrates the immunohis-ochemical labeling with a mouse anti-synaptophysin antibody (aarker of nerve terminals) and a goat anti-mouse secondary antibody
abeled with Alexa Fluor 488 (emits green) of hippocampal sectionsrom control (saline-injected) rats (panel A1), rats subjected to sub-hronic stress (panel A2), rats injected with SCH 58261 (0.05 mg/kg,aily i.p. injection to rats not subject to restraint stress) (panel A3) andats injected with SCH 58261 and subjected to sub-chronic stress0.05 mg/kg, daily i.p. injection 30 min before each immobilizationession) (panel A4). The pictures presented are representative of fivexperiments with qualitatively similar results. The background fluores-ence obtained in sections treated only with the secondary antibodyever reached more than 7% of the signal obtained in the presentedhotographs (not shown). Panel B displays a Western blot (30 �grotein loaded in each lane) comparing the density of synaptophysin
mmunoreactivity in hippocampal membranes obtained from controlats, rats subjected to sub-chronic stress (‘stress’), rats injected withCH 58261 (‘SCH’) and of rats injected with SCH 58261 and subjected
o sub-chronic stress (‘stress�SCH’), as well as and the averageesults (mean�S.E.M.) of synaptophysin immunoreactivity in five suchxperiments. In each Western blot analysis, a re-probing for detectionf �-tubulin was carried out to confirm that similar amounts of proteinere applied to the gel. * P�0.05 compared with control and
* P�0.05 compared with ‘stress’ (one-way ANOVA followed byewman-Keuls test).
1
f facilitatory A2A receptors (reviewed in Cunha, 2005).

TcinatrtwacpabsAonqo(iaSwthe
tti(st(tspafepsreibiwr(nrbpcc
tisdds(AntishMfimp
IrtaFipAii
AaC
C
C
C
d
D
E
E
F
G. M. A. Cunha et al. / Neuroscience 141 (2006) 1775–17811780
his is in accordance with the down-regulation (i.e. de-reased function) of A1 receptors and up-regulation (i.e.
ncreased function) of A2A receptors in the limbic andeo-cortex upon chronic brain insults in the brain of adultnimals (reviewed in Cunha, 2005). It should be noted thathe modifications of the density of cortical A1 and A2A
eceptors after restraint stress were found to occur in nerveerminals, precisely the cortical sub-cellular compartmenthere these two receptors are most abundant (Rebola etl., 2003, 2005). However, it was not tested if thesehanges might also occur in other neuronal or glial com-artments that are known to be endowed with these twodenosine receptors (Fredholm et al., 2005). It should alsoe pointed out that the mechanisms underlying these re-traint stress-induced changes in the density of synaptic
1 and A2A receptors remain to be defined. This variationf the density of cortical adenosine receptors upon chronicoxious conditions has been proposed to be a conse-uence of the increased brain extracellular levels of aden-sine, which increase in a variety of noxious conditionsreviewed in Fredholm et al., 2005). Thus, it would benteresting to document whether the extracellular levels ofdenosine increase upon repetitive immobilization (seecaccianoce et al., 1989) in the hippocampus to confirmhether the sub-chronic restraint stress-induced modifica-
ion of the density of A1 and A2A adenosine receptors in theippocampus is related to increased levels of hippocampalxtracellular adenosine caused by this stressor.
The only clear histological and biochemical measurehat we found to be modified upon sub-chronic stress washe density of a pre-synaptic marker, synaptophysin, whichs an indirect index of the density of nerve terminalsMasliah and Terry, 1993). In fact, the reported sub-chronictress-induced decrease of the density of synaptophysin inhe hippocampus, which has also been reported by othersThome et al., 2001; but see Rosenbrock et al., 2005),entatively suggests that sub-chronic stress might causeynaptic loss, which has previously received a direct mor-hological confirmation (see Sandi et al., 2003; Sousa etl., 2000). In contrast, we found that sub-chronic stress
ailed to induce any measurable pattern of neurotoxicity, asvaluated by a lack of change of the pattern of hippocam-al Cresyl Violet or FluoroJade-C staining. This ability ofub-chronic stress to cause synaptotoxicity, but not neu-otoxicity, has previously been noted by others (Magariñost al., 1997; Sousa et al., 2000; Stewart et al., 2005) and is
n accordance with the reversibility of the stress-inducedehavioral impairments in rodents and humans. Interest-
ngly, limbic and neo-cortical adenosine A2A receptors,hich were found to be up-regulated upon sub-chronic
estraint stress, are mostly located in nerve terminalsLopes et al., 2004; Rebola et al., 2005), precisely the onlyeuronal compartment which was affected by sub-chronicestraint stress. This prompted the hypothesis that thelockade of adenosine A2A receptors might be a novelrotective strategy against the early hippocampal modifi-ations caused by stress, which was experimentally
onfirmed.One of the consequences of chronic stress is favoringhe implementation of a state of depression in susceptiblendividuals (Kloet et al., 2005). And recent data havehown that the pharmacological blockade or the geneticeletion of A2A receptors prevented the signs of behavioralespair in the tail suspension and forced swim tests, twocreening procedures predictive of antidepressant activityEl Yacoubi et al., 2001). This putative deleterious role of
2A receptors in depression (El Yacoubi et al., 2003) is inotable agreement with the presently raised hypothesishat the blockade of A2A receptors relieves the early stress-nduced hippocampal modifications. Finally, since depres-ion as well as the early stress-induced re-modeling ofippocampal circuits is under the control of serotonin (e.g.cEwen, 1999), it will be interesting to study if the bene-
cial effect of A2A receptor antagonists in these conditionsay be related to their known ability to control hippocam-al serotonin release (Okada et al., 2001).
CONCLUSION
n conclusion, the present study shows that sub-chronicestraint stress leads to a decreased density of A1 recep-ors together with a marked increase of the density ofdenosine A2A receptors in hippocampal nerve terminals.urthermore, it is shown that these A2A receptors may be
nvolved in the early stress-induced decrease of synapto-hysin immunoreactivity, suggesting that the blockade of
2A receptors may constitute a novel therapeutic possibil-ty to interfere with the early stress-induced modificationsn the hippocampus.
cknowledgments—This work was supported by Fundação paraCiência e para a Tecnologia (POCTI/FCB/44740/2002). G. M. A.unha was supported by CNPq-Brazil.
REFERENCES
hau A, Rose JC, Koos BJ (1999) Adenosine modulates corticotropinand cortisol release during hypoxia in fetal sheep. Am J ObstetGynecol 180:1272–1277.
oelho JE, Rebola N, Fragata I, Ribeiro JA, de Mendonça A, CunhaRA (2006) Hypoxia-induced desensitization and internalization ofadenosine A1 receptors in the rat hippocampus. Neuroscience138:1195–1203.
unha RA (2005) Neuroprotection by adenosine in the brain: from A1
receptor activation to A2A receptor blockade. Purinergic Signal1:111–134.
e Mendonça A, Sebastião AM, Ribeiro JA (2000) Adenosine: does ithave a neuroprotective role after all? Brain Res Rev 33:258–274.
uarte JMN, Oliveira CR, Ambrósio AF, Cunha RA (2006) Modificationof adenosine A1 and A2A receptor density in the hippocampus ofstreptozotocin-induced diabetic rats. Neurochem Int 48:144–150.
l Yacoubi M, Ledent C, Parmentier M, Bertorelli R, Ongini E, Costen-tin J, Vaugeois JM (2001) Adenosine A2A receptor antagonists arepotential antidepressants: evidence based on pharmacology andA2A receptor knockout mice. Br J Pharmacol 134:68–77.
l Yacoubi M, Costentin J, Vaugeois JM (2003) Adenosine A2A recep-tors and depression. Neurology 61(Suppl 6):S82–S87.
redholm BB, Chen JF, Cunha RA, Svenningsson P, Vaugeois JM(2005) Adenosine and brain function. Int Rev Neurobiol 63:
191–270.
G
G
H
J
K
K
L
L
L
M
M
M
M
M
O
O
P
R
R
R
R
S
S
S
S
S
S
S
S
T
U
G. M. A. Cunha et al. / Neuroscience 141 (2006) 1775–1781 1781
eiger JD, Glavin GB (1985) Adenosine receptor activation in brainreduces stress-induced ulcer formation. Eur J Pharmacol 115:185–190.
erwins P, Fredholm BB (1991) Glucocorticoid receptor activationleads to up-regulation of adenosine A1 receptors and down-regu-lation of adenosine A2 responses in DDT1 MF-2 smooth musclecells. Mol Pharmacol 40:149–155.
erman JP, Cullinan WE (1997) Neurocircuitry of stress: central con-trol of the hypothalamo-pituitary-adrenocortical axis. Trends Neu-rosci 20:78–84.
egou S, Yacoubi ME, Mounien L, Ledent C, Parmentier M, CostentinJ, Vaugeois JM, Vaudry H (2003) Adenosine A2A receptor genedisruption provokes marked changes in melanocortin contentand pro-opiomelanocortin gene expression. J Neuroendocrinol 15:1171–1177.
im JJ, Diamond DM (2002) The stressed hippocampus, synapticplasticity and lost memories. Nat Rev Neurosci 3:453–462.
loet ER, Joëls M, Holsboer F (2005) Stress and the brain: fromadaptation to disease. Nat Rev Neurosci 6:463–475.
opes LV, Cunha RA, Ribeiro JA (1999) ZM 241385, an adenosineA2A receptor antagonist, inhibits hippocampal A1 receptor re-sponses. Eur J Pharmacol 383:395–398.
opes LV, Rebola N, Pinheiro PC, Richardson PJ, Oliveira CR, CunhaRA (2003) Adenosine A3 receptors are located in neurons of the rathippocampus. Neuroreport 14:1645–1648.
opes LV, Halldner L, Rebola N, Johansson B, Ledent C, Chen JF,Fredholm BB, Cunha RA (2004) Binding of the prototypical aden-osine A2A receptor agonist CGS 21680 to the cerebral cortex ofadenosine A1 and A2A receptor knockout mice. Br J Pharmacol141:1006–1014.
agariños AM, McEwen BS (1995) Stress-induced atrophy of apicaldendrites of hippocampal CA3c neurons: comparison of stressors.Neuroscience 69:83–88.
agariños AM, Verdugo JM, McEwen BS (1997) Chronic stress alterssynaptic terminal structure in hippocampus. Proc Natl Acad SciU S A 94:14002–14008.
asliah E, Terry R (1993) The role of synaptic proteins in the patho-genesis of disorders of the central nervous system. Brain Pathol3:77–85.
attingly D (1962) A simple fluorimetric method for the estimation offree 11-hydroxycorticosteroids in human plasma. J Clin Pathol15:374–379.
cEwen BS (1999) Stress and hippocampal plasticity. Annu RevNeurosci 22:105–122.
kada M, Nutt DJ, Murakami T, Zhu G, Kamata A, Kawata Y, KanekoS (2001) Adenosine receptor subtypes modulate two major func-tional pathways for hippocampal serotonin release. J Neurosci21:628–640.
ngini E (1997) SCH 58261: a selective A2A adenosine receptorantagonist. Drug Dev Res 42:63–70.
opoli P, Pintor A, Domenici MR, Frank C, Tebano MT, Pezzola A,Scarchilli L, Quarta D, Reggio R, Malchiodi-Albedi F, Falchi M,Massotti M (2002) Blockade of striatal adenosine A receptor
2Areduces, through a presynaptic mechanism, quinolinic acid-in-
duced excitotoxicity: possible relevance to neuroprotectiveinterventions in neurodegenerative diseases of the striatum.J Neurosci 22:1967–1975.
ebola N, Pinheiro PC, Oliveira CR, Malva JO, Cunha RA (2003)Subcellular localization of adenosine A1 receptors in nerve termi-nals and synapses of the rat hippocampus. Brain Res 987:49–58.
ebola N, Canas PM, Oliveira CR, Cunha RA (2005) Different synap-tic and subsynaptic localization of adenosine A2A receptors in thehippocampus and striatum of the rat. Neuroscience 132:893–903.
en H, Stiles GL (1999) Dexamethasone stimulates human A1 aden-osine receptor (A1AR) gene expression through multiple regula-tory sites in promoter B. Mol Pharmacol 55:309–316.
osenbrock H, Koros E, Bloching A, Podhorna J, Borsini F (2005)Effect of chronic intermittent restraint stress on hippocampal ex-pression of marker proteins for synaptic plasticity and progenitorcell proliferation in rats. Brain Res 1040:55–63.
andi C, Davies HA, Cordero MI, Rodriguez JJ, Popov VI, Stewart MG(2003) Rapid reversal of stress induced loss of synapses in CA3 ofrat hippocampus following water maze training. Eur J Neurosci17:2447–2456.
caccianoce S, Navarra D, Di Sciullo A, Angelucci L, Endroczi E(1989) Adenosine and pituitary-adrenocortical axis activity in therat. Neuroendocrinology 50:464–468.
chmued LC, Stowers CC, Scallet AC, Xu L (2005) Fluoro-Jade Cresults in ultra high resolution and contrast labeling of degeneratingneurons. Brain Res 1035:24–31.
heline YI, Wang PW, Gado MH, Csernansky JG, Vannier MW (1996)Hippocampal atrophy in recurrent major depression. Proc NatlAcad Sci U S A 93:3908–3913.
ilva-Torres IL, Bonan CD, Crema L, Leon-Nunes M, Battastini AM,Sarkis JJ, Dalmaz C, Ferreira MB (2003) Effect of drugs active atadenosine receptors upon chronic stress-induced hyperalgesia inrats. Eur J Pharmacol 481:197–201.
ousa N, Lukoyanov NV, Madeira MD, Almeida OF, Paula-BarbosaMM (2000) Reorganization of the morphology of hippocampal neu-rites and synapses after stress-induced damage correlates withbehavioral improvement. Neuroscience 97:253–266.
tewart MG, Davies HA, Sandi C, Kraev IV, Rogachevsky VV, PeddieCJ, Rodriguez JJ, Cordero MI, Donohue HS, Gabbott PL, Popov VI(2005) Stress suppresses and learning induces plasticity in CA3 ofrat hippocampus: a three-dimensional ultrastructural study ofthorny excrescences and their postsynaptic densities. Neuro-science 131:43–54.
venningsson P, Fredholm BB (1997) Glucocorticoids regulate theexpression of adenosine A1 but not A2A receptors in rat brain.J Pharmacol Exp Ther 280:1094–1101.
home J, Pesold B, Baader M, Hu M, Gewirtz JC, Duman RS, HennFA (2001) Stress differentially regulates synaptophysin and syn-aptotagmin expression in hippocampus. Biol Psychiatry 50:809–812.
no H, Tarara R, Else JG, Suleman MA, Sapolsky RM (1989) Hip-pocampal damage associated with prolonged and fatal stress in
primates. J Neurosci 9:1705–1711.(Accepted 10 May 2006)(Available online 22 June 2006)




















