In vivo and in vitro modulation of MDR molecules in murine thymocytes
12
In vivo and in vitro modulation of MDR molecules in murine thymocytes Daniela F.P. Leite a,b , Juliana Echevarria-Lima a,b , Leonardo T. Salgado c , Marcia A.M. Capella b , Joa ˜o B. Calixto d , Vivian M. Rumjanek a, * a Instituto de Bioquı ´mica Me ´dica, Universidade Federal do Rio de Janeiro, Brazil b Instituto de Biofı ´sica Carlos Chagas Filho, Universidade Federal do Rio de Janeiro, Brazil c Departamento de Anatomia, Instituto de Cie ˆncias Biome ´dicas, Universidade Federal do Rio de Janeiro, Brazil d Departamento de Farmacologia, Universidade Federal de Santa Catarina, Brazil Received 8 March 2005; received in revised form 28 April 2005; accepted 8 August 2005 Abstract P-glycoprotein (Pgp/ABCB1) and multidrug resistance related protein 1 (MRP1/ABCC1) were first described in multidrug resistant tumor cells. It is presently known that both proteins are also expressed in a variety of normal cells, including lymphocytes. ABCB1 activity has already been detected in subpopulations of murine thymocytes, but there was little information on the expression or activity of ABCC1 in these cells. The present work studied in mice the expression of both proteins by RT-PCR and immunofluorescence. It was possible to identify the presence of ABCB1 and to detect the expression of ABCC1 in these cells. The functional activities of these proteins were also studied in vivo and in vitro measuring the extrusion of fluorescent dyes in association with MDR modulators. Cyclosporine A, verapamil and trifluoperazine inhibited the activity of thymic ABCB1. Indomethacin, probenecid and MK571 were effective in inhibiting ABCC1 activity by thymic cells. ABCB1 was only active in a small percentage of thymocytes being present in the immature double negative (not CD4 nor CD8) subpopulation and the mature single positive (CD4 or CD8) subpopulations. The functional activity of ABCC1, on the other hand, was more homogeneously distributed being found in all thymocyte subpopulations. Possible physiological roles for these transporters on thymocytes are discussed. D 2005 Elsevier B.V. All rights reserved. Keywords: Thymocytes; ABCC1; ABCB1; Multidrug resistance; Modulators 1. Introduction The multidrug resistance (MDR) phenomenon was first observed in tumor cells exposed to chemotherapy. These cells presented cross-resistance to a number of 1567-5769/$ - see front matter D 2005 Elsevier B.V. All rights reserved. doi:10.1016/j.intimp.2005.08.005 * Corresponding author. Laborato ´rio de Imunologia Tumoral, Instituto de Bioquı ´mica Me ´dica, ICB/Centro de Cie ˆncias da Sau ´ de, Universidade Federal do Rio de Janeiro, Cidade Universi- ta ´ria, Ilha do Funda ˜o, Rio de Janeiro, CEP 21941590, Brazil. Tel.: +55 21 25626780; fax: +55 21 22701635. E-mail address: [email protected] (V.M. Rumjanek). International Immunopharmacology 6 (2006) 204 – 215 www.elsevier.com/locate/intimp
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of In vivo and in vitro modulation of MDR molecules in murine thymocytes

www.elsevier.com/locate/intimp
International Immunopharmac
In vivo and in vitro modulation of MDR molecules
in murine thymocytes
Daniela F.P. Leite a,b, Juliana Echevarria-Lima a,b, Leonardo T. Salgado c,
Marcia A.M. Capella b, Joao B. Calixto d, Vivian M. Rumjanek a,*
a Instituto de Bioquımica Medica, Universidade Federal do Rio de Janeiro, Brazilb Instituto de Biofısica Carlos Chagas Filho, Universidade Federal do Rio de Janeiro, Brazil
c Departamento de Anatomia, Instituto de Ciencias Biomedicas, Universidade Federal do Rio de Janeiro, Brazild Departamento de Farmacologia, Universidade Federal de Santa Catarina, Brazil
Received 8 March 2005; received in revised form 28 April 2005; accepted 8 August 2005
Abstract
P-glycoprotein (Pgp/ABCB1) and multidrug resistance related protein 1 (MRP1/ABCC1) were first described in multidrug
resistant tumor cells. It is presently known that both proteins are also expressed in a variety of normal cells, including
lymphocytes. ABCB1 activity has already been detected in subpopulations of murine thymocytes, but there was little
information on the expression or activity of ABCC1 in these cells. The present work studied in mice the expression of both
proteins by RT-PCR and immunofluorescence. It was possible to identify the presence of ABCB1 and to detect the expression
of ABCC1 in these cells. The functional activities of these proteins were also studied in vivo and in vitro measuring the
extrusion of fluorescent dyes in association with MDR modulators. Cyclosporine A, verapamil and trifluoperazine inhibited the
activity of thymic ABCB1. Indomethacin, probenecid and MK571 were effective in inhibiting ABCC1 activity by thymic cells.
ABCB1 was only active in a small percentage of thymocytes being present in the immature double negative (not CD4 nor CD8)
subpopulation and the mature single positive (CD4 or CD8) subpopulations. The functional activity of ABCC1, on the other
hand, was more homogeneously distributed being found in all thymocyte subpopulations. Possible physiological roles for these
transporters on thymocytes are discussed.
D 2005 Elsevier B.V. All rights reserved.
Keywords: Thymocytes; ABCC1; ABCB1; Multidrug resistance; Modulators
1567-5769/$ - see front matter D 2005 Elsevier B.V. All rights reserved.
doi:10.1016/j.intimp.2005.08.005
* Corresponding author. Laboratorio de Imunologia Tumoral,
Instituto de Bioquımica Medica, ICB/Centro de Ciencias da
Saude, Universidade Federal do Rio de Janeiro, Cidade Universi-
taria, Ilha do Fundao, Rio de Janeiro, CEP 21941590, Brazil. Tel.:
+55 21 25626780; fax: +55 21 22701635.
E-mail address: [email protected] (V.M. Rumjanek).
1. Introduction
The multidrug resistance (MDR) phenomenon was
first observed in tumor cells exposed to chemotherapy.
These cells presented cross-resistance to a number of
ology 6 (2006) 204–215

D.F.P. Leite et al. / International Immunopharmacology 6 (2006) 204–215 205
unrelated agents. Different mechanisms were shown
to be involved in this phenomenon, including the
overexpression of two transporter proteins, members
of the ATP Binding Cassette family: P-glycoprotein
(Pgp/ABCB1) and/or multidrug resistance related pro-
tein (MRP1/ABCC1). These proteins act as ATP-
dependent pumps with multiple binding sites, allow-
ing a high affinity interaction with many different
substrates [1]. These transporters mediate the efflux
of drugs from the interior of cells against concentra-
tion gradients, thereby reducing the cytotoxicity of
chemotherapic agents [2]. The transporter molecules,
ABCB1 and ABCC1, may be also expressed in nor-
mal cells were they seem to have an important role in
cellular detoxification [3–5].
The family of ABCC transporter molecules is com-
posed of at least nine different proteins. ABCC1, the
first member of this family described, recognizes
neutral and anionic hydrophobic natural products
and transports glucoronides, gluthatione and its con-
jugates [6]. On the other hand, three proteins in mice
and two in humans compose the ABCB family. This
protein binds to hydrophobic drug substrates that are
either neutral or positively charged [7,8].
The expression of ABCB1 and ABCC1 has been
described in normal cells [9,10]. Both proteins are
present in bone marrow hematopoietic precursor
cells (CD34+) [11–13], immature and mature lympho-
cytes [11–16], dendritic cells [17,18], monocytes and
macrophages [19,20]. Despite their wide distribution,
the physiological roles of ABCB1 and ABCC1 in the
immune system are not well understood. ABCC1 is
differently expressed in Th1 and Th2 lymphocytes
and its expression is linked to the activation process
[21,22].
It has also been suggested that ABCB1 and
ABCC1 are important transporters of inflammatory
mediators. ABCB1 is important for the secretion of
platelet activation factor and ABCC1 for the secretion
of leukotriene C4 and prostaglandins [23–26]. Further-
more, steroids are also transported by both molecules
[27–29] and play a major physiological role on thy-
mocyte selection. The subpopulation most sensitive to
these hormones, the double positive (DP) thymocyte
[30], does not seem to have ABCB1 activity [14],
whereas single positive (SP) thymocytes (CD4+ or
CD8+) present ABCB1 activity and are resistant to
glucocorticoids. On the other hand, ABCC1 expres-
sion was identified in human thymus by real time
PCR [31], and in human Hassal corpuscle by antibo-
dies [10] and nothing is known in relation to its
presence or physiological function in mouse thymo-
cytes. Modulators and substrates of these two proteins
such as cyclosporine A [32], indomethacin [33], meth-
otrexate [33] and glucocorticoids [27–29] are very
often used clinically and may modify the immune
system.
The present work investigated ABCB1 and
ABCC1 expression in thymocytes, their activity in
vitro and in vivo and their sensitivity to different
MDR modulators, in an attempt to comprehend their
physiological and pathological roles in the thymus.
2. Materials and methods
2.1. Animals
The experiments were carried out in 2–4 months old or
seven days old C3H/HeJ male mice, obtained from the
Animal House of the Department of Medical Biochemistry.
Animals had free access to food and water, and were housed
and handled according to institutional guidelines complying
with Brazilian legislation. Animals were sacrificed and
thymi and kidneys immediately removed.
2.2. Materials
Fetal calf serum (FCS) was obtained from Gibco and
RPMI 1640 medium, l-glutamine, 25 mM Hepes, bovine
serum albumin (BSA), rhodamine 123 (Rho 123), probene-
cid, indomethacin, verapamil and trifluoperazine were pur-
chased from Sigma-Aldrich. MK571 was gently donated by
Merk; carboxy-2V-7V-dichlorofluorescein diacetate (CFDA)
was purchased from Molecular Probes. MK571 and Rho
123 were first dissolved in distilled water in a stock solution
of 10 mM and 1 mg/ml, respectively. Trifluoperazine was
first dissolved in fetal calf serum and RPMI (1 :5) and kept
as stock solution of 15 mM; verapamil was dissolved in
phosphate-buffered saline (pH 7.4) and kept in as stock
solution of 50 mM. Cyclosporine A (Sandimun—100 mg/
ml) was diluted in RPMI. Probenecid was dissolved in
alkaline water and then the pH was adjusted to 8.0 in a
stock solution of 50 mM. Indomethacin was first dissolved
in DMSO in a stock solution of 100 mM. All drugs were
divided into aliquots and stored in the freezer until the
moment of use when they were further diluted in RPMI
1640 medium supplemented with 1 mM l-glutamine and 25
mM Hepes. Primary rat monoclonal antibodies anti-MRP1

D.F.P. Leite et al. / International Immunopharmacology 6 (2006) 204–215206
(ABCC1) was purchased from Alexis Biochemicals and
the goat anti-rat secondary (fluorescein conjugated) anti-
body was purchased from Sigma. The antibodies anti-
CD3, anti-CD8 and anti-CD4 were purchased from
R&D System or Pharmingen. Primary antibody goat
anti-Pgp (ABCB1), as well as anti-goat secondary rhoda-
mine conjugated antibody, was purchased from Santa
Cruz Biotechnologies.
2.3. Thymocyte suspension
The thymus was washed in FCS and disposed in an
individual Petri dish containing 5 ml of RPMI 1640 medium
with 10% FCS. The organ was softly macerated with the
help of a sterile rubber. Cells were washed with phosphate-
buffered saline (pH 7.4) and the pellet was gently suspended
in RPMI 1640 medium with 10% FCS.
2.4. Immunofluorescence
For the detection of ABCC1 at the single-cell level, 106
cells were incubated with antibodies anti-CD4 (PerCP) and
anti-CD8 (PE) for 30 min on ice and cells were then washed
in PBS. Cells were permeabilized with FACS lysing solu-
tion (Becton & Dickinson) for 10 min at room temperature.
Cells were washed with PBS and incubated with the pri-
mary mouse monoclonal antibody: anti-ABCC1 diluted in
PBS with 1% BSA (1 :20) for 30 min at room temperature.
This was followed by incubation with goat anti-rat second-
ary antibody conjugated with fluorescein for 20 min at room
temperature. Cells were then washed in PBS and analyzed
by flow cytometry. For those experiments data were
acquired in a mode of 100000 events, and a gate based
on forward scatter (FSC) and side scatter (SSC) parameters
was made to analyze only viable cells, other gates were
made to determine the subpopulations. Analyses were per-
formed by flow cytometry using a FACScalibur (Becton and
Dickinson, USA). The data were analyzed by WinMID
software.
For the detection of ABCB1 at the single-cell level,
5�105 cells were cyto-centrifuged (800 rpm for 1 min),
fixed and permeabilized with 100% cold methanol for 5
min. Fixed cells were re-hydrated with 50 mM NH4Cl for 3
min. Glass slides containing fixed cells were washed with
PBS and incubated with 5% BSA for 10 min. This proce-
dure was repeated and cells were washed once again with
PBS before incubating them with 5% BSA for a further 30
min. After the incubation time, cells were washed and
incubated overnight in a cold humidified chamber at 4 8Cwith the primary antibody goat anti-ABCB1 diluted in PBS
with 1% BSA (1 :50). After washing, cells were incubated
with bovine anti-goat secondary antibody conjugated with
rhodamine for 45 min at room temperature. Cells were then
washed twice in PBS and once in water and the cover slips
were mounted on glass slides with 50% glycerol in PBS.
The fluorescence was visualized under an Axiovert 100
microscope (Zeiss, Germany).
2.5. RT-PCR
Total RNA from thymocyte suspensions and from
kidney preparations was extracted using the TRizol pro-
tocol. The RT-PCR was performed essentially as in the
one-step protocol of the Retrotools kit (Biotools, USA).
Briefly 0.5 or 1.0 Ag/Al of total RNA was incubated for 1
h with 2 Al of RT buffer, 2 Al of MnCl2 (5 mM), 5 Al of amix DNTP (10 mM), 0.75 Al of Retrotools polymerase
and 7.5 Al H2O DEPC and 1 Al 0.01% (v/v). Antisense
primer (abcc1—5V CGCAGGTTGTGCAGGCCGAT 3V orabcb1—5V CTGATGTTGCTTCGTCCAG 3V) at 60 8C in
a final volume of 10 Al (40 pM). The product was
immediately put on ice and 3 Al of 5�DNA free buffer,
1 Al of MgCl2 (50 mM) and 10.5 Al of H2O and 1 Al (10pM) of the sense primer (abcc1—5V GTAGAGTTCCGG-GATTAC 3V or abcb1—5V CACAGCTGGGCATTGTGT
3V) were added. The conditions for PCR were 2 min at
94 8C and then 35 cycles with 15 s at 94 8C, 30 s at 50
8C and 1 min at 72 8C, using a DNA thermal cycler
(Perklin Elmer GeneAmp PCR systems 2400). The PCR
products were size fractionated in 1.8% agarose gels
stained with 2.5 Al ethidium bromide (10 mg/ml). DNA
Ladder (1 kb plus) was purchased from Invitrogen.
2.6. Rhodamine 123 and CFDA accumulation by flow
cytometry
To measure ABCB1 activity the substrate Rho 123 [34]
was used. For this 1�106 cells were incubated in RPMI
10% FCS at 37 8C for 30 min in the presence of 150 ng/ml
Rho 123 with or without reversors (80 nM cyclosporine A; 5
AM verapamil; 300 AM indomethacin or 2.5 mM probene-
cid). The cells were then washed with PBS and left at 37 8Cfor a further 30 min with or without reversors, washed,
suspended in cold PBS and kept on ice. To measure
ABCC1 activity the same protocol was used, but in this
case 0.5 AM of CFDA was the substrate and the reversors
probenecid, indomethacin and MK571 were used at differ-
ent concentrations.
For triple-fluorescence analyses Rho 123 cells or CFDA
cells were incubated with antibodies anti-CD4 (PE) and anti-
CD8 (CY5) for 30 min on ice. Rho 123 fluorescence was
acquired through the FL1 (FITC) channel, CD4 on FL2 (PE)
channel and CD8 on FL3 (PI) channel. For those experi-
ments data were acquired in a mode of 100000 events, and a
gate based on forward scatter (FSC) and side scatter (SSC)
parameters was made to analyze only viable cells, other

D.F.P. Leite et al. / International Immunopharmacology 6 (2006) 204–215 207
gates were made to determine the subpopulations. Analyses
were performed by flow cytometry using a FACScalibur
(Becton and Dickinson, USA). The data were analyzed by
WinMID software.
2.7. ABCB1 and ABCC1 activity in vivo
To study ABCB1 activity in vivo Rho 123 was
injected in association or not with cyclosporine A as
described before [35] with small modifications. Briefly,
2–4 months old mice received a single intraperitoneal
(i.p.) injection of cyclosporine A (25 mg/kg), followed
1 h later by a single i.p. injection of Rho 123 (1 mg/kg).
The volume of administration was 10 Al/g. After 1 h, the
animals were sacrificed, thymi removed, and the thymo-
cytes isolated as described above. To analyze ABCC1
activity, 7 days old mice were injected with CFDA in
association or not with probenecid as described by Sun et
al. [36]. Briefly, mice received a single i.p. injection of
probenecid (400 mg/kg) in a final volume of 10 Al/gimmediately after an intravenous injection of CFDA (15
mg/kg) in a final volume of 5 Al/g. The control group
was injected with the vehicle alone. One hour after the
administration animals were sacrificed, thymi were imme-
diately removed and cell suspensions prepared as
described above.
3. Results
3.1. Expression of abcb1 and abcc1 in the thymus
The expression of abcb1 and abcc1 mRNA in the
thymus was analyzed. As a positive control kidney, an
organ known to express these molecules, was used (Fig.
1A). The mRNA for both proteins could be detected in
the thymus. This result shows that abcc1 is expressed in
murine thymus.
To analyze the expression of ABCC1 on thymocytes,
the cells were stained with anti-CD4, anti-CD8 and anti-
ABCC1 antibodies and studied by flow cytometry (Fig.
1B). Virtually the whole population was positive for
ABCC1, the only exception was the DN subpopulation
in which 8% of the cells did not express this protein. The
ABCB1 expression was determined by immunofluores-
cence, 5.8% (ranging from 2.5–9.2%) of total population
were positive for ABCB1 (data not shown).
3.2. ABCB1 activity detected by flow cytometry
Our results showed that thymic cells express mRNA
and the proteins ABCC1 and ABCB1. Thus the activity
of these proteins was evaluated using fluorescent sub-
strates. P-glycoprotein has a large spectrum of substrates
and Rho 123 was used in these series of experiments. It
has been shown that efflux of this fluorescent dye from
pre-labeled cells correlates with ABCB1 activity [37].
Fluorescence intensity between 100 and 102 was consid-
ered rhodamine negative or rhodamine low cells (Rho neg
and Rho low) and fluorescence intensity above 102—
rhodamine high cells (Rho high) (Fig. 2A). The analysis
of various experiments indicated that less than 5% of the
thymocytes were Rho low or Rho neg. To test the sensi-
tivity of ABCB1 activity to different modulators/reversors,
thymocytes were treated with verapamil (5 AM), trifluo-
perazine (5 AM) and cyclosporine A (80 nM). The effi-
cacy of the reversors was not equal; cyclosporine A was
more efficient and potent than verapamil or trifluoperazine
(Fig. 2B). Some authors suggested that Rho 123 is also a
substrate for ABCG2 and ABCC1 [38,39]. There is rela-
tively little or no evidence that ABCG2 is expressed in
thymus [38]. To test the possibility that the effect
observed was due to ABCC1 activity, thymocytes were
treated with indomethacin or probenecid, but none of
these classical reversors of ABCC1 were able to enhance
Rho 123 accumulation (Fig. 2B). This result suggests two
possibilities: ABCC1 might not be active in these cells or
Rho 123 is not a good substrate for ABCC1 expressed in
thymocytes.
3.3. Thymocytes subpopulations and ABCB1 activity
The staining with anti-ABCB1 antibodies and the
functional ABCB1 assay suggested that the expression
of ABCB1 in thymocytes was heterogeneous. To define
whether the heterogeneity represented a differential
expression among the various thymocyte subpopulations,
these cells were stained with Rho 123 followed by drug
extrusion, and the cells were, subsequently, stained with
anti-CD4 and anti-CD8 monoclonal antibodies (Fig. 3A–
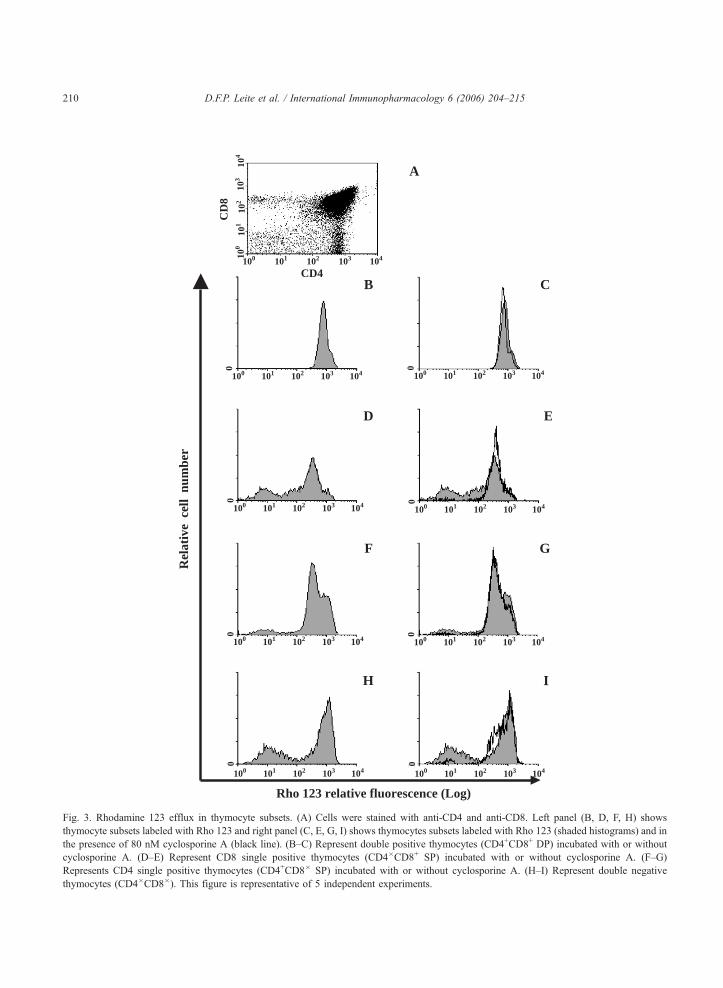
I). Our results are in agreement with those of MacDonald
et al. [14]. The double positive (DP) subpopulation
(CD4+CD8+) constituted more than 80% of the thymo-
cytes (Fig. 3A), but less than 1% of these cells lost their
fluorescence during the extrusion period (Fig. 3B) and
cyclosporine A did not modify the extrusion profile (Fig.
3C). Within the CD8+ single positive (SP) cells, the Rho
neg or Rho low cells constituted approximately 30% (Fig.
3D). Cyclosporine A treatment reduced the numbers of
CD8+ SP Rho 123 extruding cells more than 95% (Fig.
3E, black line), suggesting that almost 30% of CD8+ SP
had an active ABCB1. Within the CD4+ SP, approxi-
mately 10% of the cells were capable of reducing the
intracellular levels of Rho 123 (Fig. 3F) and cyclosporine

1 2 L 3 4
Rel
ativ
e ce
ll nu
mbe
r 92%
DN
MRP1 (Log)
MRP1 (Log)
Rel
ativ
e ce
ll nu
mbe
r
Total thymocytes
Rel
ativ
e ce
ll nu
mbe
r B 99.6%
MRP1 (Log)
MRP1 (Log)
Rel
ativ
e ce
ll nu
mbe
r 100%
DP
100%
CD8+ SP
99.4%
Rel
ativ
e ce
ll nu
mbe
r
CD4+ SP
MRP1 (Log)
A
250Kb
100
101
102
103
t104
100
101
102
103
104
100
101
102
103
104
100
101
102
103
104
100
101
102
103
104
012
8
010
00
20
010
05M1 M1
M1M1
M1
Fig. 1. Expression of ABCC1 and ABCB1 in thymocytes. (A) Expression of abcc1 (224 kb) and abcb1 (283 kb) in kidney and in thymus of
adult male mice. The expression was determined by RT-PCR, 0.5 Ag of total RNA was used to amplify abcc1 and 1 Ag of total mRNA to
amplify abcb1 in both organs. abcc1 expressed in the kidney (1) and in the thymus (2). abcb1 expressed in the thymus (3) and in the kidney (4).
(L) Ladder 250 kb. (B) ABCC1 detection in the various thymocytes subsets. Cells were permeabilized, and labeled with mAb anti-ABCC1.
Cells were labeled with anti-CD4 and anti-CD8 antibodies followed by permeabilization and incubation with anti-ABCC1 monoclonal antibody
and secondary antibody. Control was labeled only with the secondary antibody. DP—double positive (CD4+CD8+); DN—double negative
(CD4�CD8�); CD4 SP—single positive (CD4+CD8�); CD8 SP—single positive (CD4�CD8+).
D.F.P. Leite et al. / International Immunopharmacology 6 (2006) 204–215208

**
*
% I
nhib
itio
n
CSA VP TFP Prob Indo
B
-20
0
20
40
60
80
100
Fow
ard
scat
ter
Rho 123 fluorescence intensity
A10
230
100 101 102 103 104
Fig. 2. ABCB1 related activity in thymocytes. (A) Thymocyte
accumulation of rhodamine 123 (Rho 123). Thymocytes labeled
with Rho 123 for 30 min were allowed to extrude the dye for a
further 30 min. A gate based on rhodamine accumulation was
done. Fluorescence intensity between 100 and 102 was considered
Rho negative or Rho low and fluorescence intensity above 102
Rho high. Approximately, 4% of the total population of thymo-
cytes exported the dye. This figure is representative of 10 inde-
pendent experiments. (B) Inhibition of Rho 123 efflux by different
reversors: cyclosporine A—CSA (80 nM), verapamil—VP (5
AM), trifluoperazine—TFP (5 AM), probenecid—Prob (1 mM)
and indomentacin—Indo (300 AM). *p b0.001 different from
control. The results are the meanFSD of at least 4 independent
experiments.
D.F.P. Leite et al. / International Immunopharmacology 6 (2006) 204–215 209
A reversed around 85% of this population (Fig. 3G, black
line). The double negative thymocytes are the more
immature subpopulation and approximately 25% were
able to efflux Rho 123 (Fig. 3H) and cyclosporine A
reversed 95% of the ABCB1 activity (Fig. 3I, black
line). There was no difference between the sensitivities
of ABCB1 expressed on different subpopulations of thy-
mocytes to cyclosporine A, verapamil or trifluoperazine
(data not shown).
3.4. ABCB1 activity in vivo
To test if ABCB1 was active in vivo, mice were
injected (i.p.) with Rho 123 (1 mg/kg) following the
injection of cyclosporine A (25 mg/kg) or saline 1 h
previously, as described in Material and methods. The
mean of fluorescence intensity of the Rho low or negative
subpopulation of thymocytes triplicated in animals treated
with cyclosporine A, suggesting that this reversor was cap-
able of modulating ABCB1 activity in thymocytes in vivo
(data not shown).
3.5. ABCC1 activity by flow cytometry
As shown in Fig. 2B probenecid and indomethacin did
not reduce the number of Rho 123 extruding cells. To
verify if ABCC1 was active in thymocytes, these cells
were incubated with another fluorescent dye, CFDA,
which is transported by ABCC1 [40]. Other members of
ABCC family are expressed in much smaller amounts in
thymus [31] and therefore their activity is probably not
responsible for most CFDA extrusion. Although the accu-
mulation of CFDA in thymocytes was very small (Fig.
4A), treatment with the classical ABCC1 reversors:
MK571 100 AM (Fig. 4B), indomethacin 300 AM (Fig.
4C) or probenecid 2.5 mM (Fig. 4D), raised CFDA inten-
sity more than 10 times. Moreover, probenecid (0.5–2.5
mM), indomethacin (75–300 AM) and MK571 (25–100
AM) reversed ABCC1 activity in a dose dependent manner
(Fig. 5).
3.6. Thymocytes subpopulations and ABCC1 activity
To confirm that ABCC1 is active in all thymocyte
subpopulations, cells were incubated with CFDA with or
without 1 mM probenecid and after the extrusion period
cells were labeled with anti-CD4 and anti-CD8 mono-
clonal antibodies (Fig. 6). As expected, probenecid was
capable of increasing CFDA accumulation in all sub-
populations, showing that different from ABCB1,
ABCC1 activation does not seem to be developmentally
regulated.
3.7. ABCC1 activity in vivo
To test the activity of ABCC1 in vivo, young mice
were injected with CFDA (15 mg/kg) and probenecid
0
104 103102101100
A
B
D
F
H
C
E
G
I
CD
8
CD4
Rho 123 relative fluorescence (Log)
104
103
102
101
100
104 103102101100
0
104 103102101100
0
104 103102101100
0
104 103102101100 0
104 103102101100
0
104 103102101100
0
104 103102101100
0
104 103102101100
Rel
ativ
e c
ell
num
ber
0
Fig. 3. Rhodamine 123 efflux in thymocyte subsets. (A) Cells were stained with anti-CD4 and anti-CD8. Left panel (B, D, F, H) shows
thymocyte subsets labeled with Rho 123 and right panel (C, E, G, I) shows thymocytes subsets labeled with Rho 123 (shaded histograms) and in
the presence of 80 nM cyclosporine A (black line). (B–C) Represent double positive thymocytes (CD4+CD8+ DP) incubated with or without
cyclosporine A. (D–E) Represent CD8 single positive thymocytes (CD4�CD8+ SP) incubated with or without cyclosporine A. (F–G)
Represents CD4 single positive thymocytes (CD4+CD8� SP) incubated with or without cyclosporine A. (H–I) Represent double negative
thymocytes (CD4�CD8�). This figure is representative of 5 independent experiments.
D.F.P. Leite et al. / International Immunopharmacology 6 (2006) 204–215210

Indo + CFDA
CFDA
autofluorescence
CFDA
Prob + CFDA
CFDA
M
CFDA
C
A
CFDA
D
M
CFDA
B
MK571 + CFDA
145
0
100 101 102 103 104
CFDA fluorescence intensity
Rel
ativ
e ce
ll nu
mbe
r
145
0
100 101 102 103 104
145
0
100 101 102 103 104
145
0
100 101 102 103 104
Fig. 4. ABCC1 related activity in thymocytes. (A) Thymocytes were incubated with or without 0.5 AM CFDA. (B–D) Effect of reversors on
thymocytes incubated with CFDA. (B) 100 AM MK571. (C) 300 AM indomethacin (Indo). (D) 2.5 mM probenecid (Prob). Histogram analysis
is representative of six independent experiments.
D.F.P. Leite et al. / International Immunopharmacology 6 (2006) 204–215 211
(400 mg/kg) or saline (Fig. 7). As observed in vitro,
probenecid was capable of enhancing CFDA accumula-
tion homogeneously. This demonstrated that ABCC1 is
1
10
100
1000
Probenecid Indomethacin MK571
(mM) (µM) (µM)
0.5 1.0 2.5 75 150 300 25 50 100
CFDA
Mea
n o
f fl
uo
resc
en
ce i
nte
nsi
ty
Fig. 5. Dose dependent inhibition of ABCC1 activity. Mean fluor-
escence intensity of cells treated with different doses of ABCC1
reversors. Cells were labeled with 0.5 AM CFDA in the presence or
absence of probenecid (0.5–2.5 mM), indomethacin (75–300 AM),
or MK571 (25–100 AM) as described in Material and methods.
Representative of six independent experiments.
active in thymocytes and can be modulated in vivo
(Fig. 7).
4. Discussion
The present work describes the expression and
activity of ABCC1 in normal murine thymocytes,
results obtained are summarized in Table 1. Our
group has recently observed the expression of
ABCC1 in the murine thymona cell line EL4
[41]. In normal thymocytes ABCC1 was expressed
mainly in the plasma membrane (data not shown)
and similar to what has been reported in other cell
types it can be modulated by indomethacin, probe-
necid, and MK571 [33,36,41]. The accumulation of
CFDA, an ABCC1 fluorescent substrate, was homo-
geneously distributed among thymocytes, suggesting
that the expression of ABCC1 is similar among the
different subpopulations. This result was confirmed
when thymocytes were triple stained for anti-CD4,
anti-CD8 and CFDA in the presence or absence of
probenecid.

BA
DC
FE
HG
BA
DC
FE
HG HG
100 101 102 103 104 100 101 102 103 104
100 101 102 103 104 100 101 102 103 104
100 101 102 103 104100 101 102 103 104
100 101 102 103 104100 101 102 103 104
Rel
ativ
e ce
ll nu
mbe
r
CFDA flourescence intensity (Log)
Fig. 6. CFDA efflux from thymocytes subsets. Cells were stained with 0.5 AM of CFDA, anti-CD4 and anti-CD8. Left panel (A, C, E, G) shows
thymocyte subsets labeled with CFDA and right panel (B, D, F, H) shows thymocytes subsets labeled with CFDA (shaded histograms) in the
presence of 1 mM probenecid (black line). (A–B) CFDA accumulation in double positive (CD4+CD8+) thymocytes. (C–D) CFDA accumulation
in CD4+CD8� thymocytes. (E–F) CFDA accumulation in CD4�CD8+ thymocytes. (G–H) CFDA accumulation in double negative CD4�CD8�
thymocytes. This figure is representative of 3 independent experiments.
D.F.P. Leite et al. / International Immunopharmacology 6 (2006) 204–215212
ABCB1 was also observed in the thymus and this
observation is in agreement with the results of other
authors [14,15]. However, different from ABCC1, that
is active in all thymocyte subpopulations, ABCB1
seems to be developmentally regulated, as it was
only active in mature (single positive cells) and very
immature (duple negative cells) subpopulations.
The physiologic role of ABC transporters in the
thymus deserves further investigation. It is quite clear
from our results that both ABCB1 and ABCC1 activ-
ities may be modulated in vivo. These results suggest
that agents used quite often in the clinical setup, such as
cyclosporine A and indomethacin, may inhibit the
activity of ABCB1 and/or ABCC1 with unknown con-
sequences to the immune repertoire. Indeed, it has been
reported that cyclosporine A affects Tcell development
and repertoire selection in the thymus [42–44] and that
indomethacin induces T cell maturation in the EL4
thymoma cell line [45]. However, the possibility that
these effects might result from their action on ABC
transporters should be considered as probenecid and
indomethacin, both inhibitors of ABCC1, were shown
by us to induce differentiation of EL4 cells [41].
It has been described that leukotrienes, mainly
leukotriene C4, and prostaglandins, which may be
extruded via ABCC1, modulate immature thymocyte
proliferation, and alter the expression of surface mole-
cules, such as CD3, CD4 and CD8 [45–47]. ABCB1
has as natural substrates glucocorticoids [27,28] and
the subpopulation of thymocytes that is most sensitive

100
0
100 101 102 103 104
CFDA fluorescence intensity
Rel
ativ
e ce
ll nu
mbe
r
Fig. 7. ABCC1 related activity in vivo. Histogram depicting fluor-
escence intensity of thymocytes obtained from uninjected animals
(autofluorescence, shaded histogram), injected with 15 mg/kg
CFDA (second peak, - - -) and injected with CFDA and 400 mg/
kg of probenecid (third peak, —).
D.F.P. Leite et al. / International Immunopharmacology 6 (2006) 204–215 213
to these hormones, the double positive thymocytes
[30] do not present ABCB1 activity, suggesting a
role for this transporter on the control of intracellular
cortisol levels. In fact, when mice are treated with
Table 1
Summary of results
Pgp
mRNA detection +
Inhibitors (concentrations used) CSA (80 nM)
VP (5 AM)
TFP (5 AM)
Percent of the cells with transporter
expression
Total population—5.8
Within DN—nd
Within DP—nd
Within CD4 SP—nd
Within CD8 SP—nd
Percent of the total population with
transporter activity
Total population—4%
Determined with 80
Total population—2.5
Determined with 5 AMTotal population—2.8
Determined with 80
Percent of each thymocyte subset
with transporter activity
Within DN—23.7%
Within DPN1%
Within CD4 SP—8.5
Within CD8 SP—28.
Determined with 80
In vivo activity 5%
Determined with 25 m
Transporter activity was measured using rhodamine 123 as a substrate fo
substrate for MRP. CSA—cyclosporine A; VP—verapamil; TFP—trifl
negative (CD4�CD8�); DP—double positive (CD4+CD8+); CD4 SP—sin
nd—not determined.
hydrocortisone and the remaining thymocytes are
exposed to rhodamine in vitro, the percentage of
rhodamine extruding cells increase (data not shown).
There are however, evidences that ABCC1 is capable
of transporting corticosterone out of the cell [28]. Our
results indicate that all subpopulations of thymocytes
present an active ABCC1, making it unlikely that
physiologically the efflux of corticosterone by
ABCC1 is important for thymocyte survival.
Another possible role for ABCC1 in thymocytes
involves their migration within the thymus and to the
periphery. The expression of ABCC1 on dendritic
cells [17] and on mature T lymphocytes [48] was
shown to be important for their migration to lymph
nodes. Robbiani et al. showed that dendritic cell
migration involves leukotriene secretion, which is
mediated by ABCC1 [17]. Thymocyte emigration
involves a mechanism similar to the one described
in dendritic cells [49], suggesting that ABCC1 activ-
ity could participate in this process and might affect
the traffic of mature thymocytes. Furthermore,
MRP1
+
Indo (75–300 AM)
Prob (0.5–2.5 AM)
MK571 (25–100 AM)
% Total population—99.6%
Within DN—92%
Within DP—100%
Within CD4 SP—100%
Within CD8 SP—99.4%
Total population—90%
nM of CSA Determined with 1 mM of Prob
% Total population—97%
of VP Determined with 100 AM of MK571
% Total population—85%
nM of CSA Determined with 150 AM of Indo
Within DN—92%
Within DP—84%
% Within CD4 SP—97%
5% Within CD8 SP—84%
nM of CSA Determined with 1 mM of Prob
75%
g/kg of CSA Determined with 400 mg/kg of Prob
r Pgp and CFDA (carboxy-2V-7V-dichlorofluorescein diacetate) as a
uoperazine; Indo—indomethacin; Prob—probenecid; DN—double
gle positive (CD4+CD8�); CD8 SP—single positive (CD4-CD8+);

D.F.P. Leite et al. / International Immunopharmacology 6 (2006) 204–215214
ABCB1 was also reported to have a role on lympho-
cyte homing to lymph node [48] and may also play
role on thymocyte emigration.
In conclusion, the identification of the presence
and activity of ABCC1 and ABCB1 molecules in
thymocytes raises the question of whether these pro-
teins have a role in thymic differentiation and reper-
toire selection.
Acknowledgements
We would like to thank Dr. Franklin D. Rumjanek
and Daniel Rodrigues Furtado for technical support.
This work was supported by PRONEX, CNPq (Brazi-
lian National Research Council) and by Howard
Hughes Medical Institute (departmental sharing of
grant # 55003669). Daniela F.P. Leite was a recipient
of a PhD Fellowship from CNPq. Juliana Echevarria-
Lima and Leonardo T. Salgado were a recipient of a
PhD Fellowship from CAPES (Coordenacao de Aper-
feicoamento de Pessoal de Nıvel Superior).
References
[1] Neyfakh AA. Mystery of multidrug transporters: the answer
can be simple. Mol Microbiol 2002;44:1123–30.
[2] Gottesman MM, Pastan I. Biochemistry of multidrug resis-
tance mediated by the multidrug transporter. Annu Rev Bio-
chem 1993;62:385–427.
[3] Wijnholds J, Scheffer GL, Van Der Valk M, Van Der Valk P,
Beijnen JH, Scheper RJ, et al. Multidrug resistance protein 1
protects the oropharyngeal mucosal layer and the testicular
tubules against drug-induced damage. J Exp Med 1998;188:
797–808.
[4] Schinkel AH, Mayer U, Wagenaar E, Molca, Van Deemter L,
Smit JJ, et al. Normal viability and altered pharmacokinetics in
mice lacking mdr1-type (drug-transporting) P-glycoproteins.
Proc Natl Acad Sci U S A 1997;94:4028–33.
[5] Schinkel AH, Smit JJ, Van Tellingen O, Beijnen JH, Wagenaar
E, Van Deemter L, et al. Disruption of the mouse mdr1a P-
glycoprotein gene leads to a deficiency in the blood-brain
barrier and to increased sensitivity to drugs. Cell 1994;77:
491–502.
[6] Borst P, Elferink RO. Mammalian ABC transporters in health
and disease. Annu Rev Biochem 2002;71:537–92.
[7] Gottesman MM, Ambudkar SV. Overview: ABC transporters
and human disease. J Bioenerg Biomembr 2001;33:453–8.
[8] Borst P, Evers R, Kool M, Wijnholds J. A family of drug
transporters: the multidrug resistance-associated proteins.
J Natl Cancer Inst 2000;92:1295–302.
[9] Fojo AT, Ueda K, Slamon DJ, Poplack DG, Gottesman
MM, Pastan I. Expression of a multidrug-resistance gene
in human tumors and tissues. Proc Natl Acad Sci U S A
1987;84:265–9.
[10] Flens MJ, Zaman G.J, Van Der Valk P, Izquierdo MA, Schroei-
jers AB, Scheffer GL, et al. Tissue distribution of the multi-
drug resistance protein. Am J Pathol 1996;148:1237–47.
[11] Legrand O, Perrot JY, Tang RP, Simonin G, Gurbuxani S,
Zittoun R, et al. Expression of the multidrug resistance-asso-
ciated protein (MRP) mRNA and protein in normal peripheral
blood and bone marrow haemopoietic cells. Br J Haematol
1996;94:23–33.
[12] Laupeze B, Amiot L, Payen L, Drenou B, Grosset JM, Lehne
G, et al. Multidrug resistance protein (MRP) activity in normal
mature leukocytes and CD34-positive hematopoietic cells
from peripheral blood. Life Sci 2001;68:1323–31.
[13] Chaudhary PM, Roninson IB. Expression and activity of P-
glycoprotein, a multidrug efflux pump in human hematopoie-
tic stem cells. Cell 1991;66:85–94.
[14] MacDonald HPR, Bommhardt U, Cerottini JC. Developmen-
tally regulated expression of P-glycoprotein (multidrug resis-
tance) activity in mouse thymocytes. Eur J Immunol 1995;
25:1457–60.
[15] Pilarski LM, Paine D, Mcelhaaney JE, Cass CE, Belvh AR.
Multidrug transporter P-gycoprotein 170 as a differentiation
antigen on normal human lymphocytes and thymocytes: mod-
ulation with differentiation stage and during aging. Am J
Hematol 1995;49:332–5.
[16] Gupta S, Gollapudi SJ. The P-glycoprotein (MDR 1 gene
product) in cells of the immune system: its possible physiolo-
gic role and alteration in aging and human immunodeficiency
virus-1 (HIV-1) infection. J Clin Immunol 1993;13:289–301.
[17] Robbiani DF, Finch RA, Jager D, Muller WA, Sartorelli AC,
Randolph GJ. The leukotriene C(4) transporter MRP1 regu-
lates CCL19 (MIP-3beta, ELC)-dependent mobilization of
dendritic cells to lymph nodes. Cell 2000;103:757–68.
[18] Randolph GJ, Beaulieu S, Pope M, Sugawara I, Hoffman L,
Steinman RM, et al. A physiologic function for p-glycoprotein
(MDR-1) during the migration of dendritic cells from skin via
afferent lymphatic vessels. Proc Natl Acad Sci U S A 1998;
95:6924–9.
[19] Vellenga E, Tuyt L, Wierenga BJ, Muller M, Dokter W.
Interleukin-6 production by activated human monocytic cells
is enhanced by MK-571, a specific inhibitor of the multi-drug
resistance protein-1. Br J Pharmacol 1999;127:441–8.
[20] Puddu P, Fais S, Luciani F, Gherardi G, Dupuis ML, Romag-
noli G, et al. Interferon-gamma up-regulates expression and
activity of P-glycoprotein in human peripheral blood mono-
cyte-derived macrophages. Lab Invest 1999;79:1299–309.
[21] Lohoff M, Prechtl S, Sommer F, Roellinghoff M, Schmitt E,
Gradehandt G, et al. A multidrug-resistance protein (MRP)-
like transmembrane pump is highly expressed by resting mur-
ine T helper (Th) 2, but not Th1 cells, and is induced to equal
expression levels in Th1 and Th2 cells after antigenic stimula-
tion in vivo. J Clin Invest 1998;101:703–10.
[22] Prechtl S, Roellinghoff M, Scheper R, Cole SP, Deeley RG,
Lohoff M. The multidrug resistance protein 1: a functionally

D.F.P. Leite et al. / International Immunopharmacology 6 (2006) 204–215 215
important activation marker for murine Th1 cells. J Immunol
2000;164:754–61.
[23] Ernest S, Bello-Reuss E. Secretion of platelet-activating factor
is mediated by MDR1 P-glycoprotein in cultured human
mesangial cells. J Am Soc Nephrol 1999;10:2306–13.
[24] Leier I, Jedlitschky G, Buchholz U, Cole SPC, Deeley RG,
Kepper D. The MRP gene encodes an ATP-dependent export
pump for leukotriene C4 and structurally related conjugates.
J Biol Chem 1994;269:27807–10.
[25] Evers R, Cnubben NH, Wijnholds J, van Deemter L, van
Bladeren PJ, Borst P. Transport of glutathione prostaglandin
A conjugates by multidrug resistance protein 1. FEBS Lett
1997;419:112–6.
[26] Renes J, de Vries EG, Jansen PL, Muller M. The (patho)phy-
siological functions of the MRP family. Drug Resist Updat
2000;3:289–302.
[27] Farrell RJ, Menconi MJ, Keates AC, Kelly CP. P-glycopro-
tein-170 inhibition significantly reduces cortisol and cyclos-
porin efflux from human intestinal epithelial cells and T
lymphocytes. Aliment Pharmacol Ther 2002;16:1021–31.
[28] Webster JI, Carlstedt-Duke J. Involvement of multidrug resis-
tance proteins (MDR) in the modulation of glucocorticoid
response. J Steroid Biochem Mol Biol 2002;82:277–88.
[29] Gruol DJ, Bourgeois S. Expression of the mdr1 P-glycoprotein
gene: a mechanism of escape from glucocorticoid-induced
apoptosis. Biochem Cell Biol 1994;72:561–71.
[30] Cohen JJ. Glucocorticoid-induced apoptosis in the thymus.
Sem Immunol 1992;4:363–9.
[31] Langmann T, Mauerer R, Zahn A, Moehle C, Probst M,
Stremmel W, et al. Real-time reverse transcription-PCR
expression profiling of the complete human ATP-binding cas-
sette transporter superfamily in various tissues. Clin Chem
2003;49:230–8.
[32] Sorokin R, Kimura H, Schroder K, Wilson DH, Wilson DB.
Cyclosporine-induced autoimmunity. Conditions for expres-
sing disease, requirement for intact thymus, and potency esti-
mates of autoimmune lymphocytes in drug-treated rats. J Exp
Med 1986;164:1615–25.
[33] Duffy CP, Elliott CJ, O’Connor RA, Heenan MM, Coyle S,
Cleary IM, et al. Enhancement of chemotherapeutic drug
toxicity to human tumour cells in vitro by a subset of non-
steroidal anti-inflammatory drugs (NSAIDs). Eur J Cancer
1998;34:1250–9.
[34] Neyfakh AA. Use of fluorescent dyes as molecular probes for
the studies of multidrug resistance. Exp Cell Res 1988;
174:168–76.
[35] Marques-Santos L.F, Harab RC, de Paula EF, Rumjanek VM.
The in vivo effect of the administration of resistance-modulat-
ing agents on rhodamine 123 distribution in mice thymus and
lymph nodes. Cancer Lett 1999;137:99–106.
[36] Sun H, Johnson DR, Finch RA, Sartorelli AC, Miller DW,
Elmquist WF. Transport of fluorescein in MDCKII-MRP1
transfected cells and mrp1-knockout mice. Biochem Biophys
Res Commun 2001;284:863–9.
[37] Drach D, Zhao S, Drach J, Mahadevia R, Gattringer C, Huber
H, et al. Subpopulations of normal peripheral blood and bone
marrow cells express a functional multidrug resistant pheno-
type. Blood 1992;80:2729–34.
[38] Zhou S, Schuetz JD, Bunting KD, Colapietro AM, Sampath J,
Morris JJ, et al. The ABC transporter Bcrp1/ABCG2 is
expressed in a wide variety of stem cells and is a molecular
determinant of the side-population phenotype. Nat Med
2001;7:1028–34.
[39] Daoud R, Kast C, Gros P, Georges E. Rhodamine 123 binds to
multiple sites in the multidrug resistance protein (MRP1).
Biochemistry 2000;39:15344–52.
[40] Laupeze B, Amiot L, Courtois A, Vernhet L, Drenou B,
Fauchet R, et al. Use of the anionic dye carboxy-2V,7V-dichlor-ofluorescein for sensitive flow cytometric detection of multi-
drug resistance-associated protein activity. Int J Oncol
1999;15:571–6.
[41] Echevarria-Lima J, Kyle FC, Leite DFP, Capella LS, Capella
MAM, Rumjanek VM. Expression and activity of multidrug
resistance protein 1 in a murine thymoma cell line. Immunol-
ogy 2005;114:440–8.
[42] Urdahl KB, Pardoll DM, Jenkins MK. Cyclosporin A inhibits
positive selection and delays negative selection in alpha beta
TCR transgenic mice. J Immunol 1994;152:2853–9.
[43] Zadeh HH, Goldschneider I. Demonstration of large-scale
migration of cortical thymocytes to peripheral lymphoid tis-
sues in cyclosporin A-treated rats. J Exp Med 1993;178:
285–93.
[44] Cairns JS, Mainwaring MS, Cacchione RN, Walker JA,
McCarthy SA. Regulation of apoptosis in thymocytes. Thy-
mus 1993;21:177–93.
[45] Tomooka S, Serushago BA, Koga Y, Taniguchi K,
Nomoto K. Indomethacin-induced sialic acid-mediated
changes in surface markers from cortical type to medullary
type in murine thymoma line EL4. Immunobiol 1986;171:
345–56.
[46] Delebassee S, Gualde N. Effect of arachidonic acid metabo-
lites on thymocyte proliferation. Ann Inst Pasteur Immunol
1988;139:383–99.
[47] Daculsi R, Vaillier D, Carron JC, Gualde N. Effect of PGE2 on
the cell surface molecule expression in PMA treated thymo-
cytes. Immunol Lett 1998;60:81–8.
[48] Honig SM, Fu S, Mao X, Yopp A, Gunn MD, Randolph GJ, et
al. FTY720 stimulates multidrug transporter- and cysteinyl
leukotriene-dependent T cell chemotaxis to lymph nodes.
J Clin Invest 2003;111:627–37.
[49] Campbell JJ, Pan J, Butcher EC. Cutting edge: developmental
switches in chemokine responses during T cell maturation.
J Immunol 1999;163:2353–7.