
CD1d structure and regulation on human thymocytes, peripheral blood T cells,B cells and monocytes
11
CD1d structure and regulation on human thymocytes, peripheral blood T cells, B cells and monocytes M. EXLEY,* J. GARCIA,* S. B. WILSON,{ F. SPADA,{1 D. GERDES,* S. M. A. TAHIR,* K. T. PATTON,* R. S. BLUMBERG", S. PORCELLI,{1 A. CHOTT** & S. P. BALK*{ *Cancer Biology Program, Hematology-Oncology Division, Department of Medicine, Beth Israel Deaconess Medical Center and Harvard Medical School, Boston, MA, USA, {Diabetes Unit, Massachusetts General Hospital and Harvard Medical School, Boston, MA, USA, {Lymphocyte Biology Section, Division of Rheumatology, Immunology, and Allergy, Brigham and Women’s Hospital and Harvard Medical School, Boston, MA, USA, "Gastroenterology Division, Brigham and Women’s Hospital and Harvard Medical School, Boston, MA, USA, **Department of Clinical Pathology, University of Vienna, Austria SUMMARY Human T cells expressing CD161 and an invariant T-cell receptor (TCR) a-chain (Va24 invt T cells) specifically recognize CD1d and appear to have immunoregulatory functions. However, the physiological target cells for this T-cell population, and whether alterations in CD1d expression contribute to the regulation of Va24 invt T-cell responses, remain to be determined. A series of antibodies were generated to assess CD1d expression, structure and regulation on human lymphoid and myeloid cells. CD1d was expressed at high levels by human cortical thymocytes and immunoprecipitation analyses showed it to be a 48 000-MW glycosylated protein. However, after solubilization, the majority of the thymocyte CD1d protein, but not CD1d expressed by transfected cells, lost reactivity with monoclonal antibodies (mAbs) against native CD1d, indicating that it was alternatively processed. Moreover, thymocytes were not recognized by CD1d-reactive Va24 invt T- cell clones. Medullary thymocytes and resting peripheral blood T cells were CD1d – , but low-level CD1d expression was induced on activated T cells. CD1d was expressed by B cells in peripheral blood and lymph node mantle zones, but germinal centres were CD1d – . Resting monocytes were CD1d + but, in contrast to CD1a, b and c, their surface expression of CD1d was not up-regulated by granulocyte–macrophage colony-stimulating factor (GM-CSF) and interleukin-4 (IL-4) activation. These results demonstrate constitutive CD1d expression by human professional antigen-presenting cells and that post-translational processing of CD1d may contribute to regulation of the activity of CD1d-specific T cells. INTRODUCTION The human CD1 locus encodes a family of proteins that are structurally related to major histocompatibility complex (MHC) class I proteins. 1–3 CD1a, b and c are most homologous to each other and are expressed by thymocytes, dendritic cells and activated monocytes, 4–9 while B cells express only CD1c. 10 T cells reactive with CD1a, b, or c have been isolated from peripheral blood, and lipids derived from mycobacteria have been identified as the antigens recognized by some of these clones. 8,11 CD1d is divergent in sequence from CD1a, b and c 3,12 and its tissue distribution is more widespread, including cells outside the lymphoid and myeloid lineages. 13–15 Possible CD1d-presented antigens include hydrophobic peptides, glycosylceramides and glycosylphosphatidylinositol. 16–18 Murine CD1d, even in the absence of an exogenous antigen, is recognized by a population of CD4 + or CD4 – CD8 – (double negative, DN), CD161 + (NK1) T cells that use an invariant Va14-Ja281 T-cell receptor (TCR) a-chain. 19–23 A population of human DN or CD4 + T cells that uses a homologous invariant Va24-JaQ TCR a-chain has been identified (Va24 invt T cells). 24,25 These human T cells also express CD161 (NKR- P1A), which acts as the major costimulatory molecule, and specifically recognize CD1d. 26,27 Functional studies suggest an Correspondence: Steven P. Balk, Hematology-Oncology Division, Beth Israel Deaconess Medical Center, HIM Building Room 1047, 330 Brookline Avenue, Boston, MA 02215, USA. Received 27 October 1999; revised 9 December 1999; accepted 9 December 1999. Abbreviations: GST, glutathione-S-transferase; b 2 m, b 2 -microglo- bulin; Va24 invt T cells, Va24 invariant T cells. Present address: 1Department of Microbiology and Immunology, Albert Einstein College of Medicine, 1300 Morris Park Avenue, Bronx, NY 1046, USA. Immunology 2000 100 37–47 # 2000 Blackwell Science Ltd 37
Transcript of CD1d structure and regulation on human thymocytes, peripheral blood T cells,B cells and monocytes

CD1d structure and regulation on human thymocytes, peripheral blood T cells,
B cells and monocytes
M. EXLEY,* J. GARCIA,* S. B. WILSON,{ F. SPADA,{1 D. GERDES,* S. M. A. TAHIR,* K. T. PATTON,*
R. S. BLUMBERG", S. PORCELLI,{1 A. CHOTT** & S. P. BALK*{ *Cancer Biology Program, Hematology-Oncology
Division, Department of Medicine, Beth Israel Deaconess Medical Center and Harvard Medical School, Boston, MA, USA,
{Diabetes Unit, Massachusetts General Hospital and Harvard Medical School, Boston, MA, USA, {Lymphocyte Biology Section,
Division of Rheumatology, Immunology, and Allergy, Brigham and Women's Hospital and Harvard Medical School, Boston, MA,
USA, "Gastroenterology Division, Brigham and Women's Hospital and Harvard Medical School, Boston, MA, USA,
**Department of Clinical Pathology, University of Vienna, Austria
SUMMARY
Human T cells expressing CD161 and an invariant T-cell receptor (TCR) a-chain (Va24invt T cells)
speci®cally recognize CD1d and appear to have immunoregulatory functions. However, the
physiological target cells for this T-cell population, and whether alterations in CD1d expression
contribute to the regulation of Va24invt T-cell responses, remain to be determined. A series of
antibodies were generated to assess CD1d expression, structure and regulation on human lymphoid
and myeloid cells. CD1d was expressed at high levels by human cortical thymocytes and
immunoprecipitation analyses showed it to be a 48 000-MW glycosylated protein. However, after
solubilization, the majority of the thymocyte CD1d protein, but not CD1d expressed by transfected
cells, lost reactivity with monoclonal antibodies (mAbs) against native CD1d, indicating that it was
alternatively processed. Moreover, thymocytes were not recognized by CD1d-reactive Va24invt T-
cell clones. Medullary thymocytes and resting peripheral blood T cells were CD1d±, but low-level
CD1d expression was induced on activated T cells. CD1d was expressed by B cells in peripheral
blood and lymph node mantle zones, but germinal centres were CD1d±. Resting monocytes were
CD1d+ but, in contrast to CD1a, b and c, their surface expression of CD1d was not up-regulated by
granulocyte±macrophage colony-stimulating factor (GM-CSF) and interleukin-4 (IL-4) activation.
These results demonstrate constitutive CD1d expression by human professional antigen-presenting
cells and that post-translational processing of CD1d may contribute to regulation of the activity of
CD1d-speci®c T cells.
INTRODUCTION
The human CD1 locus encodes a family of proteins that are
structurally related to major histocompatibility complex
(MHC) class I proteins.1±3 CD1a, b and c are most homologous
to each other and are expressed by thymocytes, dendritic cells
and activated monocytes,4±9 while B cells express only CD1c.10
T cells reactive with CD1a, b, or c have been isolated from
peripheral blood, and lipids derived from mycobacteria have
been identi®ed as the antigens recognized by some of these
clones.8,11 CD1d is divergent in sequence from CD1a, b and
c3,12 and its tissue distribution is more widespread, including
cells outside the lymphoid and myeloid lineages.13±15 Possible
CD1d-presented antigens include hydrophobic peptides,
glycosylceramides and glycosylphosphatidylinositol.16±18
Murine CD1d, even in the absence of an exogenous antigen,
is recognized by a population of CD4+ or CD4± CD8± (double
negative, DN), CD161+ (NK1) T cells that use an invariant
Va14-Ja281 T-cell receptor (TCR) a-chain.19±23 A population
of human DN or CD4+ T cells that uses a homologous
invariant Va24-JaQ TCR a-chain has been identi®ed (Va24invt
T cells).24,25 These human T cells also express CD161 (NKR-
P1A), which acts as the major costimulatory molecule, and
speci®cally recognize CD1d.26,27 Functional studies suggest an
Correspondence: Steven P. Balk, Hematology-Oncology Division,
Beth Israel Deaconess Medical Center, HIM Building Room 1047, 330
Brookline Avenue, Boston, MA 02215, USA.
Received 27 October 1999; revised 9 December 1999; accepted
9 December 1999.
Abbreviations: GST, glutathione-S-transferase; b2m, b2-microglo-
bulin; Va24invt T cells, Va24 invariant T cells.
Present address: 1Department of Microbiology and Immunology,
Albert Einstein College of Medicine, 1300 Morris Park Avenue, Bronx,
NY 1046, USA.
Immunology 2000 100 37±47
# 2000 Blackwell Science Ltd 37

immunoregulatory role for these Va24invt T cells,28±31 but the
precise nature of this role and that of CD1d remain to be
determined. This report describes the development and use of a
new series of CD1d antibodies to assess CD1d expression,
structure and regulation on human lymphoid and myeloid cells.
MATERIALS AND METHODS
Cells and culture
C1R cells, a human leucocyte antigen (HLA)-A and -B
de®cient B-lymphoblastoid cell line, transfected with CD1d
cDNA or a CD1d/CD1a chimeric cDNA, were as described
previously.26 Interleukin (IL)-2 used at 100 U/ml was provided
by the NIH Biological Response Modi®ers Program (Bethesda,
MD). Human thymocytes were obtained from paediatric
patients undergoing cardiac surgery. CD1d-reactive Va24invt
T-cell clones were derived from the peripheral blood of normal
donors, as described previously.26 Functional studies mon-
itored by proliferation and cytokine production were carried
out on resting Va24invt T-cell clones, as described pre-
viously.26,27 Brie¯y, Va24invt T-cell clones (105/well) were
incubated with irradiated stimulator cells in 96-well ¯at-
bottom plates and phorbol 12-myristate 13-acetate (PMA)
(1 ng/ml) was included, as indicated, to maximize costimula-
tion.26 Peripheral blood T-cell activation was carried out using
phytohaemagglutinin (PHA), as described previously.26 Mono-
cyte CD1a, b and c expression was induced by culture in
granulocyte±macrophage colony-stimulating factor (GM-CSF)
and IL-4 (100 units/ml of each),8 plus or minus lipopoly-
saccharide (LPS), for 24 hours.
Antibodies
Monoclonal and polyclonal antibodies were generated against
CD1d-glutathione-S-transferase (GST) fusion proteins and
(a)
C1R CD1a
C1R CD1a
C1R CD1b
C1R CD1c
C1R CD1d
C1R CD1b C1R CD1c
38 M. Exley et al.
# 2000 Blackwell Science Ltd, Immunology, 100, 37±47

against a CD1d-immunoglobulin fusion protein. A pool of
CD1d-GST fusion proteins containing the a1 domain alone,
the a1 and a2 domains, or the a1±a3 domains, were used to
immunize rabbits and mice. Rabbit antiserum was af®nity
puri®ed by passing multiple times through a GST column
followed by absorption to a column of agarose beads (Af®gel,
Bio-Rad, Hercules, CA) covalently conjugated with a mixture
of the three CD1d-GST fusion proteins. Monoclonal anti-
bodies (mAbs) against the pooled CD1d-GST fusion proteins
were generated by fusing hyperimmune BALB/c spleen cells to
murine myeloma (NS-1) cells. One antibody from this screen,
termed D5 (immunoglobulin G2b [IgG2b] isotype), reacted in
immunoblots against the a1 domain of CD1d (S. Balk,
unpublished).
A CD1d-immunoglobulin fusion protein was produced
by ®rst generating a BamHI site at the 3k end of the
CD1d a3 domain, using an oligonucleotide (antisense
CGGGATCCCCCCAGTAGAGGACGATG) and polymer-
ase chain reaction (PCR) ampli®cation. This BamHI site was
then used to fuse the CD1d leader sequence through the a3
domain (ending at the sequence Val±Leu±Tyr±Trp±Gly) to the
Fc portion of murine IgG2a, using an immunoglobulin Fc
expression vector (kindly provided by Dr Terry Strom, Beth
Israel Deaconess Medical Center, Boston, MA).32 The fusion
protein was secreted as a b2-microglobulin (b2m)-associated,
disulphide-linked dimer when expressed in hamster (CHO),
murine (NSO) or human (C1R) cells (S. Balk, unpublished).
CD1d knockout mice (M. Exley et al. submitted) were
immunized with the fusion protein and hybridomas were
subsequently screened by enzyme-linked immunosorbent assay
(ELISA) using the protein. The anti-CD1d mAbs raised against
the CD1d-immunoglobulin fusion protein used in this report
were 27.1, 42.1, and 51.1. Additional antibodies used in this
study included BBM.1, OKT6, 4A76, and M241, which
recognize b2m, CD1a, b and c, respectively.
Flow cytometry
Flow cytometry analyses were carried out using < 1r106 cells
in 50±100 ml of phosphate-buffered saline (PBS) containing
0.05% Na azide and 1% fetal calf serum (FCS). Peripheral
blood mononuclear cells (PBMC) were initially blocked by
incubation in 10% human serum. For indirect immuno¯uor-
escence, the primary antibodies were each used at 10±20 mg/ml
for 20±30 min at 4u. The secondary antibodies were ¯uorescein
(b) Thymocytes
Figure 1. Speci®city of anti-CD1d antibodies and thymocyte CD1d expression by indirect immuno¯uorescence. (a) Polyclonal rabbit anti-CD1d
and mouse monoclonal 27.1, 42.1 and 51.1 anti-CD1d antibodies were tested by indirect immuno¯uorescence against C1R cells stably transfected
with CD1a, b, c or d, as indicated. The top three panels (C1R CD1a, C1R CD1b and C1R CD1c) are the CD1a, b and c transfectants stained with
anti-CD1a (OKT6), anti-CD1b (4A76) and anti-CD1c (M241) monoclonal antibodies (mAbs), respectively. The thick lines represent speci®c
antibodies and the thin lines are control antibodies (normal rabbit or mouse immunoglobulin G [IgG]). (b) Freshly isolated human thymocytes were
stained by indirect immuno¯uorescence with the indicated speci®c (thick lines) or control (thin lines) antibodies.
39CD1d structure and regulation of expression
# 2000 Blackwell Science Ltd, Immunology, 100, 37±47

(a) (d)
(b)
(c)
IgG 27.1 anti-CD1d
IgG
42.1
ant
i-CD
1dIg
G42
.1 a
nti-C
D1d
IgG
42.1
ant
i-CD
1d
Anti-TCR
Anti-CD19
Anti-CD19
Anti-CD14
Anti-CD14
Donor 1 Donor 2 Donor 3 Donor 4
42.1 anti-CD1d 42.1 anti-CD1d
51.1 anti-CD1d 51.1 anti-CD1d
27.1 anti-CD1d
Figure 2. CD1d expression by peripheral blood lymphocytes and monocytes. Peripheral blood mononuclear cells from two
representative normal donors were analysed by two-colour immuno¯uorescence with the indicated directly ¯uorescein isothiocyanate
(FITC)- or phycoerythrin (PE)-conjugated speci®c and control antibodies. (a) Anti-T-cell receptor ab (TCR ab) versus control or 42.1
anti-CD1d analysis of cells in the resting lymphocyte gate. (b) Anti-CD19 versus control or 42.1 anti-CD1d analysis of cells in the
resting lymphocyte gate. (c) Anti-CD14 versus control or 42.1 anti-CD1d analysis of cells in the monocyte gate. (d) Histogram of
CD1d expression on B cells from two additional donors, examined as described above, gating of the B-cell (CD19+) population.
40 M. Exley et al.
# 2000 Blackwell Science Ltd, Immunology, 100, 37±47
isothiocyanate (FITC)-conjugated anti-mouse or anti-rabbit
F(abk)2 fragments (DAKO, Carpinteria, CA). Direct Ab
conjugates were from Pharmingen or DAKO. The 42.1 anti-
CD1d mAb was conjugated to FITC according to the
manufacturer's protocol (Molecular Probes, Eugene, OR).
Immunoprecipitation and immunoblotting
Cell-surface proteins were radiolabelled with 125I using
lactoperoxidase.33 Alternatively, cells (1±2r107/ml) were sur-
face labelled with 0.5 mg/ml sulphosuccinimidyl-6-(biotina-
mido) hexanoate (NHS-LC-Biotin; Pierce Chemicals,
Rockford, IL) in 25 mM HEPES, pH 8.0, 0.14 M NaCl, for
30 min at 4u. After washing, labelled cells were lysed in
immunoprecipitation buffer (0.15 M NaCl, 50 mM Tris, pH 7.8,
and 0.5% Nonidet P-40 [NP-40]) containing protease inhibi-
tors. For immunoblotting experiments, lysates from unlabelled
cells were prepared similarly.
Immunoprecipitations were performed using antibodies
coupled to protein G (IgG1 mAbs) or protein A±Sepharose
beads (Pierce). To minimize background from eluted IgG,
antibody was covalently coupled to the beads using dimethyl-
pimelimidate. Lysates were cleared by incubation with non-
immune serum bound to protein A± and/or protein G±
Sepharose beads. They were then incubated with speci®c
antibodies coupled to Sepharose beads, washed, eluted in non-
reducing sodium dodecyl sulphate±polyacrylamide gel electro-
phoresis (SDS±PAGE) sample buffer and analysed by SDS±
PAGE under reducing or non-reducing conditions.
After transfer to nitrocellulose, biotinylated proteins were
detected using streptavidin-horseradish peroxidase (HRP) and
enhanced chemiluminescence (ECL) (Amersham, Bucks, UK).
Non-labelled CD1d was detected by immunoblotting using the
af®nity-puri®ed rabbit anti-CD1d antibody, followed by anti-
rabbit immunoglobulin-HRP conjugates and ECL. b2m was
detected by immunoblotting of non-reducing gels with the
BBM.1 mAb. N-linked carbohydrates were removed by
digestion with N-glycanase (Genzyme, Cambridge, MA), as
described previously.33
Immunohistochemistry
Staining with anti-CD1 mAbs was performed on frozen
sections following a 10-min ®xation in acetone. Endogenous
peroxidase was ®rst blocked by incubation in 1% H2O2 in Tris-
buffered saline or in glucose (50 mg/ml) and glucose oxidase
(7 U; Sigma Chemical Co., St Louis, MO). Biotinylated horse
anti-mouse IgG was used as the secondary antibody followed
by Vectostain Elite ABC Reagent (Vector Labs, Burlinghame,
CA). The chromogens were 3-amino-9-ethyl-carbazole or
3,3k-diamino-benzidine.
RESULTS
Speci®city of anti-CD1d antibodies
A panel of human C1R B-cell clones transfected with CD1a, b,
c, or d were used to determine the speci®city of antibodies
generated against CD1d-GST and CD1d-immunoglobulin
fusion proteins. The af®nity-puri®ed rabbit anti-CD1d anti-
body, generated against CD1d-GST fusion proteins, speci®-
cally stained the CD1d transfectant (Fig. 1a, control versus
rabbit anti-CD1d). Similarly, the 27.1 mouse anti-human
(a)
Rabbit anti-CD1d 27.1 anti-CD1d
Fresh monocytes
(b)
Anti-CD1a Anti-CD1b
Anti-CD1c Anti-CD1d
Cultured monocytes
(c)
Anti-CD1a Anti-CD1b
Anti-CD1c Anti-CD1d
Monocytes with LPS
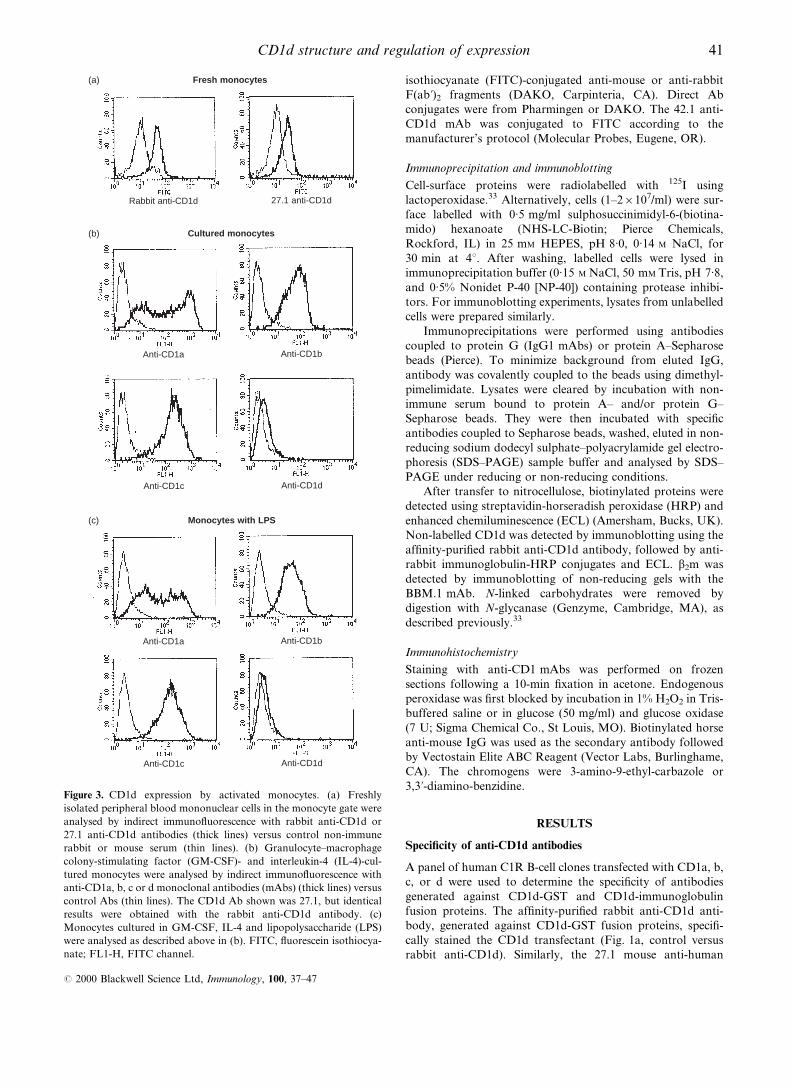
Figure 3. CD1d expression by activated monocytes. (a) Freshly
isolated peripheral blood mononuclear cells in the monocyte gate were
analysed by indirect immuno¯uorescence with rabbit anti-CD1d or
27.1 anti-CD1d antibodies (thick lines) versus control non-immune
rabbit or mouse serum (thin lines). (b) Granulocyte±macrophage
colony-stimulating factor (GM-CSF)- and interleukin-4 (IL-4)-cul-
tured monocytes were analysed by indirect immuno¯uorescence with
anti-CD1a, b, c or d monoclonal antibodies (mAbs) (thick lines) versus
control Abs (thin lines). The CD1d Ab shown was 27.1, but identical
results were obtained with the rabbit anti-CD1d antibody. (c)
Monocytes cultured in GM-CSF, IL-4 and lipopolysaccharide (LPS)
were analysed as described above in (b). FITC, ¯uorescein isothiocya-
nate; FL1-H, FITC channel.
41CD1d structure and regulation of expression
# 2000 Blackwell Science Ltd, Immunology, 100, 37±47

CD1d mAb, raised against a CD1d-IgG fusion protein,
speci®cally recognized CD1d (Fig. 1a, control versus 27.1
anti-CD1d). The 42.1 and 51.1 mouse anti-human CD1d mAbs
similarly reacted strongly with CD1d transfectants, although
they showed some cross-reactivity with CD1b (Fig. 1a). The
42.1 and 51.1 mAbs may also cross-react very weakly with
CD1a and c, as suggested by ¯ow cytometry results (Fig. 1a).
The D5 mouse anti-human CD1d mAb, raised against a CD1d-
GST fusion protein, did not bind to CD1d-transfected C1R
cells or other human CD1d transfectants, as determined by
¯ow cytometry (not shown), but was shown to bind human
CD1d on intact, transfected MDCK cells.34
CD1d expression by thymocytes
Northern blot analyses showed previously that CD1d tran-
scripts were expressed at very low levels by human thymocytes
compared with CD1a.3 Moreover, CD1d was not detected
previously on human thymocytes using a cross-reactive anti-
mouse CD1d mAb.14 Nonetheless, high-level cell-surface
expression of CD1d protein, comparable to that on CD1d
transfectants, was detected on the majority of thymocytes from
multiple donors using each of the anti-CD1d mAbs and the
CD1d-speci®c rabbit polyclonal Ab (Fig. 1b). Thymocyte
CD1d expression was also con®rmed by immunoprecipitations
(see below).
CD1d expression in peripheral blood
Figure 2 shows an analysis of CD1d expression on peripheral
blood cells from two representative normal donors (of < 40
analysed). CD1d expression was undetectable on the vast
majority of resting peripheral blood T cells, as determined by
using the 27.1, 42.1, 51.1 or rabbit anti-CD1d antibodies,
although very small numbers of weakly positive T cells were
sometimes observed (Fig. 2a and data not shown). In contrast,
CD1d staining of essentially all CD19+ B cells was observed
with the 42.1 mAb (Fig. 2B). Analyses of further donors and
the use of multiple antibodies con®rmed CD1d expression on B
cells (Fig. 2d and data not shown). This con®rmed the previous
®nding of CD1d on human B cells using a cross-reactive anti-
mouse CD1d mAb.14 CD1d expression by B cells was not
increased after activation in vitro with pokeweed mitogen, anti-
immunoglobulin M (anti-IgM), or LPS (results not shown).
The group 1 CD1 proteins (CD1a, b and c) are not
expressed on resting monocytes. However, CD1d expression
was clearly detected on freshly isolated CD14+ monocytes
(Figs 2c, 3a). It was shown previously that CD1a, b and c
expression could be strongly induced on monocytes by
stimulation with GM-CSF and IL-4.8,9 To determine whether
CD1d expression was regulated similarly, fresh monocytes
were cultured with GM-CSF and IL-4, plus or minus LPS
(Fig. 3b, 3c, respectively). In both cases there was induction of
CD1a, b and c expression, but surface CD1d expression was
not induced under these short-term culture conditions. These
results demonstrate that CD1d is expressed constitutively by
monocytes and its expression is regulated differently from the
group 1 CD1 proteins (CD1a, b and c).
Although peripheral blood T cells were CD1d±, expression
of CD1d could be induced by T-cell activation in vitro. CD1d
expression was observed within 24 hr of culture (Fig. 4a) and
persisted for at least 5 days (results not shown). Signi®cantly,
the analysis of peripheral blood from healthy donors revealed
some samples with similar low-level CD1d expression by the
majority of T cells (Fig 4b, 27.1 anti-CD1d, week 2). CD1d
expression by these donors did not appear to re¯ect
polymorphisms or other genetic differences in CD1d regula-
tion, as in all cases previous and/or subsequent samples from
the same donors were found to be CD1d± negative (Fig. 4b,
27.1 anti-CD1d, week 1). CD1d expression in each of these
samples correlated with modest levels of CD69 expression,
(a) Unstimulated Stimulated
100 101 102 103 104
Cou
nts
FL1-H
100
80
60
40
20
0100 101 102 103 104
100
80
60
40
20
0
Cou
nts
FL1-H
100 101 102 103 104
Cou
nts
Don
or 1
Don
or 2
FL1-H
100
80
60
40
20
0100 101 102 103 104
100
80
60
40
20
0
Cou
nts
FL1-H
(b) Week 1 Week 2
100 101 102 103 104
Cou
nts
200
160
120
80
40
0100 101 102 103 104
200
160
120
80
40
0
Cou
nts
Con
trol
vs27
.1 a
nti-C
D1d
100 101 102 103 104
Cou
nts
200
160
120
80
40
0100 101 102 103 104
200
160
120
80
40
0
Cou
nts
Con
trol
vsC
D 6
9
Figure 4. CD1d expression by activated T cells. (a) Freshly isolated
peripheral blood mononuclear cells (PBMC) from two donors were
cultured for 24 hr with or without stimulation and then analysed by
indirect immuno¯uorescence with 42.1 anti-CD1d monoclonal anti-
body (mAb) (42.1) (thick lines) or isotype-matched control Ab (thin
lines). Similar results were obtained with the other anti-CD1d mAbs
and the rabbit anti-CD1d Ab (not shown). The very weak staining in
the cells cultured without stimulation was not seen in the freshly
isolated resting T cells (results not shown). (b) Freshly isolated
peripheral blood mononuclear cells isolated 1 week apart from the
same donor were analysed by indirect immuno¯uorescence with the
indicated speci®c mAbs (thick lines) versus control antibodies (thin
lines). FL1-H, FITC channel.
42 M. Exley et al.
# 2000 Blackwell Science Ltd, Immunology, 100, 37±47

indicative of recent activation (Fig. 4b). Therefore, these
studies of CD1d expression in peripheral blood from healthy
donors indicated that CD1d expression by T cells was induced
by activation in vivo.
Immunohistochemical analysis of CD1d expression in thymus
and lymph node
The above ¯ow cytometric analysis showed strong CD1d
expression by most thymocytes. Consistent with this, immu-
nohistochemistry showed relatively uniform CD1d expression
by cortical thymocytes (Fig. 5a). However, there was a striking
down-regulation of CD1d expression by medullary thymo-
cytes, with CD1d+ thymocytes being very rare in the medulla.
CD1a expression was also diminished in the transition from
cortex to medulla, but many CD1a+ cells could nonetheless be
detected in the medulla (Fig. 5b). In lymph node, mantle zone
B cells were strongly CD1d+, while CD1d+ cells in germinal
centres were extremely rare (Fig. 5c). In the interfollicular T-
cell-rich zones, there were scattered, small-to-medium sized
mononuclear cells that were strongly CD1d+. Comparable
results were obtained with each of the anti-CD1d Abs.
Structure of CD1d expressed by human thymocytes
Although ¯ow cytometry with the 27.1, 42.1 and 51.1 mAbs
indicated a relatively high level of CD1d expression by
thymocytes, sequential immunoprecipitations with the
42.1 mAbs from biotinylated thymocyte lysates yielded rela-
tively small amounts of protein (Fig. 6a, lanes 2 and 3).
Comparable low levels of CD1d were immunoprecipitated by
the 27.1 and 51.1 mAbs (results not shown). In contrast,
substantially more CD1d was precipitated from the same
thymocyte lysates by the D5 anti-CD1d mAb (Fig. 6a, lane 4),
with the amount of biotin label being comparable to that
precipitated by the OKT6 anti-CD1a mAb (lane 5, T6). The
proteins precipitated by 42.1 and D5 migrated identically to the
major CD1d protein expressed by transfected C1R cells
(Fig. 6a, lane 6) (< 48 000 MW), and migrated at
37 000 MW after N-glycanase digestion (Fig. 6b), consistent
with thymocyte CD1d being fully glycosylated. These results
indicated that the epitopes recognized by the 27.1, 42.1, and
51.1 mAbs were lost after biotinylation and detergent solubi-
lization on the majority of the thymocyte cell-surface CD1d.
The D5 mAb was generated against a CD1d-GST fusion
protein and recognizes an epitope in the a1 domain (S. Balk,
unpublished). D5 binding to detergent-solubilized CD1d
appears to disrupt the association with b2m, as D5 immuno-
precipitates CD1d from transfected C1R cells without b2m.35
Although D5 immunoprecipitates CD1d, and not CD1a, b or c,
from the respective transfected C1R cells (reference 35 and data
not shown), the speci®city of this mAb was further assessed
directly on thymocyte lysates. To determine whether the
D5 mAb might be cross-reactive with CD1a expressed by
thymocytes (which migrates at the same position on SDS±
PAGE), thymocyte lysates were initially precleared three times
with an anti-CD1a mAb (Fig. 6c, CD1a, lanes 1±3). Immuno-
precipitations were then carried out with D5 on the CD1a
precleared versus fresh thymocyte lysates. The amount of
protein immunoprecipitated by D5 from the precleared lysate
(lane 4) versus fresh lysate (lane 5) was comparable, indicating
that D5 was not cross-reacting with thymocyte CD1a.
Although cross-reactivity of the D5 mAb with CD1b or
CD1c expressed by thymocytes was unlikely, as the apparent
molecular weight of these proteins on SDS±PAGE was lower
(not shown), this was nonetheless further addressed by
determining whether the D5 mAb could immunodeplete
CD1a, b, or c from thymocyte lysates. In these experiments,
CD1a, b and c were detected by immunoblotting with anti-b2m,
as immunoblotting antibodies for each of the group 1 CD1
proteins were not available. D5 was ®rst used to preclear half of
an unlabelled thymocyte lysate, followed by CD1a, b and c
immunoprecipitation and b2m immunoblotting. Figure 6(d)
shows that the amount of CD1a, b and c immunoprecipitated
was similar before (lanes 2±4) and after (lanes 5±7) two
successive preclears with the D5 mAb, which further con®rmed
that the D5 mAb was CD1d speci®c.
To address the possibility that epitopes on CD1d
recognized by the 42.1 and 51.1 mAbs were lost as a result
of biotin labelling, and to further con®rm the speci®city of the
mAbs, unlabelled thymocyte lysates were analysed by immu-
noblotting with the rabbit anti-CD1d Ab. Lysates were cleared
by two immunoprecipitations with the 42.1 mAb (Fig. 7a,
lanes 1 and 2) or 51.1 mAb (lanes 4 and 5), followed by
immunoprecipitation with the D5 mAb (lanes 3 and 6,
respectively). Immunoblotting with the rabbit anti-CD1d Ab
con®rmed that the 42.1 and 51.1 mAbs precipitated only a
small fraction of the total cellular CD1d relative to that
immunoprecipitated by the D5 mAb. In marked contrast to
these results with thymocyte CD1d, most of the CD1d in
lysates from CD1d-transfected C1R cells could be cleared by
immunoprecipitation with the 42.1 mAb (Fig. 7b, sequential
42.1 preclears in lanes 2±4 versus subsequent D5 immuno-
precipitations in lanes 5 and 6). Similar results were obtained
(a) (b) (c)
Figure 5. Immunoperoxidase analysis of CD1 expression in thymus and lymph node. (a) Thymus stained with the 51.1 anti-CD1d
monoclonal antibody (mAb). (b) Thymus stained with the OKT6 anti-CD1a mAb. (c) Lymph node stained with 51.1 anti-CD1d mAb.
Comparable results were obtained with the 27.1 and 42.1 anti-CD1d mAbs (not shown).
43CD1d structure and regulation of expression
# 2000 Blackwell Science Ltd, Immunology, 100, 37±47

with the 27.1 and 51.1 mAbs (results not shown). Taken
together, these results con®rmed biochemically CD1d expres-
sion by human thymocytes and demonstrated a structural
difference, detectable as the loss of one or more epitopes after
solubilization in non-ionic detergent, between a large fraction
of the CD1d protein expressed by thymocytes versus CD1d
expressed by transfected C1R cells.
The cytoplasmic tail of CD1d contains a tyrosine-based
endosomal-targeting motif that appears to be necessary for the
acquisition of speci®c lipid antigens.36 To determine whether
the structural difference between thymocyte and C1R CD1d
could re¯ect a post-translational modi®cation that occurs
during endosomal processing, a mutant CD1d, without this
endosomal targeting motif and containing instead the cyto-
plasmic tail from CD1a (CD1d/a), was examined. Figure 7(b)
shows that the vast majority of the CD1d/a protein could be
immunoprecipitated with the 42.1 mAb, indicating that the
structural difference between thymocyte and C1R CD1d does
not re¯ect a cytoplasmic tail-dependent processing step.
CD1d-speci®c Va24invt T cells do not respond to thymocyte
CD1d
Human Va24invt T cells can be stimulated by CD1d-transfected
cells expressing high levels of CD1d protein, comparable to the
levels on thymocytes, without the addition of an exogenous
antigen.26,27 Therefore, the ability of thymocytes to function as
stimulator cells for Va24invt T-cell clones was assessed. In
contrast to CD1d-transfected C1R and CHO cells, thymocytes
were unable to stimulate Va24invt T-cell clones (Fig. 8). Similar
results were obtained using four distinct Va24invt T-cell clones
(data not shown). Stimulation could not be reconstituted by
inclusion of phorbol ester or light glutaraldehyde ®xation
(Fig. 8), procedures that can circumvent the need for
costimulation in the case of some CD1d+ target cells.26
DISCUSSION
The expression of CD1d in the lymphoid and myeloid lineages
was assessed to identify potential physiological CD1d+ target
cells for CD1d-reactive T cells and to determine whether CD1d
expression was constitutive or regulated. CD1d expression
during T-cell development paralleled CD1a, b and c, which are
expressed at similar levels on immature cortical thymocytes and
down-regulated on mature thymocytes and peripheral blood
T cells.4±6 CD1d expression on peripheral blood B cells
paralleled that of CD1c, which is expressed on a large
proportion of mature B cells.10 However, these similarities in
the regulation of CD1a, b and c expression did not extend to
peripheral blood monocytes, as CD1a, b and c expression by
(a) (b)
(c) (d)
Thymus
42.1 D5 T6 42.1
1 2 3 4 5 6
1 2 3Streptavidin blot
MW (× 103)
66
46
30
MW (× 103)
46
MW (× 103)
14
MW (× 103)
46
30
C1R-d N-glc
Preclear
CD1a CD1d a b c a b c
1 2 3 4 5
Streptavidin blot
1 2 3 4 5 6 7
β2m blot
Figure 6. CD1d immunoprecipitations from thymocytes and speci®city of D5 monoclonal antibody (mAb). (a) Streptavidin blot of
immunoprecipitates from biotinylated thymocyte lysates: lane 1, normal mouse serum; lanes 2 and 3, sequential 42.1 anti-CD1d mAb
precipitations; lane 4, D5 anti-CD1d mAb; lane 5, OKT6 anti-CD1a precipitation; lane 6, 42.1 precipitation from CD1d-transfected
C1R cells. (b) Deglycosylation of CD1d from surface-iodinated thymocyte lysates: lane 1, normal mouse serum immunoprecipitation;
lanes 2 and 3, D5 anti-CD1d mAb immunoprecipitates untreated (lane 2) or treated lane 3 with N-glycanase (N-glc). (c) D5 immuno-
precipitation from CD1a precleared biotinylated thymocyte lysate: lanes 1±4, lysate immunoprecipitated sequentially with OKT6
(anti-CD1a) beads (lanes 1±3) followed by reimmunoprecipitation with D5 (anti-CD1d) beads (lane 4); lane 5, D5 precipitation from
an equal amount of lysate without a CD1a preclear. (d) Immunoprecipitates from unlabelled thymocyte lysates immunoblotted with
BBM.1 anti-b2m mAb: lanes 1±4, fresh thymocyte lysate; lanes 5±7, equivalent amounts of D5 anti-CD1d mAb-depleted thymocyte
lysate. Immunoprecipitations were as follows: lane 1, normal mouse serum; lanes 2 and 5, anti-CD1a (OKT6) mAb; lanes 3 and 6,
anti-CD1b (4A76) mAb; lanes 4 and 7, anti-CD1c (M241) mAb. Samples were analysed on a 15% gel under non-reducing conditions.
44 M. Exley et al.
# 2000 Blackwell Science Ltd, Immunology, 100, 37±47

monocytes is activation dependent.8,9 In contrast, cell-surface
CD1d expression by monocytes was constitutive and not up-
regulated by in vitro activation. Finally, CD1d expression by
mature peripheral blood T cells could be induced in vitro, and
appeared to be induced in vivo, by activation.
The lymph node immunohistochemistry was consistent
with the peripheral blood analysis, showing CD1d expression
primarily in the B-cell-rich mantle zones. Recent reports
showed higher level expression of murine CD1d by splenic
marginal zone B cells.37,38 Human lymph nodes do not have
well-de®ned marginal zones,39 but scattered CD1d+ cells
observed in the interfollicular T-cell zones could re¯ect this B-
cell population (or dendritic cells). Signi®cantly, CD1d+ cells
were not detected in germinal centres, indicating that B-cell
CD1d expression is down-regulated at this stage and that
CD1d may not play a role in the lymphocyte and dendritic cell
interactions that occur in germinal centres.
Immunoprecipitations con®rmed that thymocytes ex-
pressed relatively high levels of CD1d and showed that it
was fully glycosylated. They further showed that thymocyte
CD1d, but not CD1d expressed by transfected cells, lost
epitopes recognized by conformationally sensitive mAbs after
solubilization in non-ionic detergent. The structural differences
between CD1d expressed by thymocytes and CD1d transfec-
tants probably re¯ects post-translational processing at the level
of antigen binding or glycosylation. Correspondingly, CD1d
expressed by thymocytes also appeared to be functionally
distinct from CD1d on transfected cells, as thymocytes
expressing high levels of CD1d protein were unable to stimulate
CD1d-reactive Va24invt T cells. These results indicate that
while CD1d+ thymocytes may mediate the positive selection of
CD1d-reactive T cells,40 they are not the physiological target
cells of mature Va24invt T cells. Further studies are necessary to
determine the relationship between CD1d structure and
function on thymocytes. However, the data suggest that the
role of alternative CD1d processing by thymocytes may be to
promote the positive selection of Va24invt T cells while
preventing negative selection caused by high-af®nity binding
of CD1d by the Va24invt TCR.
Signi®cant human thymocyte CD1d expression was not
detected previously by ¯ow cytometry or immunohistochem-
istry using the cross-reactive rat anti-mouse CD1d mAbs, 1H1
and 3C11,14,15 although these mAbs detected human CD1d on
transfected cells, B cells and intestinal epithelial cells.14,15,33
The 1H1 and 3C11 mAbs similarly failed to immunoprecipitate
CD1d from human thymocytes (data not shown), consistent
with a distinct structure for thymocyte CD1d. We showed
previously, using the 1H1 and 3C11 mAbs, that CD1d was
alternatively processed in human intestinal epithelial cells,
yielding a non-glycosylated and non-b2m-associated protein,33
and these results have recently been con®rmed with additional
antibodies.41 These observations suggest that post-transla-
tional modi®cation may regulate CD1d function in a number
of cell types.
The rate of CD1d turnover in thymocytes appeared to be
extremely slow, based upon the relatively high level of CD1d
protein expression in conjunction with very low message
levels.3 However, immunohistochemistry demonstrated a
marked down-regulation of CD1d protein expression in the
transition from thymic cortex to the medulla. This suggests a
speci®c mechanism to clear CD1d protein from the cell surface
(a)
(b)
Thymus
42.1 D5 51.1 D5
1 2 3 4 5 6
1 2 3 4 5 6 7 8 9 10 11 12
42.1
Anti-CD1d immunoblot
Anti-CD1d immunoblot
D5 42.1 D5
C1R-CD1d C1R-CD1d/a
46 000 MW
46 000 MW
Figure 7. CD1d immunoprecipitations from thymocytes and trans-
fected cells immunoblotted with rabbit anti-CD1d. (a) Immuno-
precipitates from unlabelled thymocytes immunoblotted with
af®nity-puri®ed rabbit anti-CD1d: lanes 1 and 2, primary and
secondary 42.1 monoclonal antibody (mAb) precipitations, respec-
tively; lanes 4 and 5, primary and secondary 51.1 mAb precipitations,
respectively; lanes 3 and 6, D5 anti-CD1d precipitations from 42.1 and
51.1 precleared lysates, respectively. (b) Lysates from C1R cells
expressing wild-type CD1d (lanes 1±6) or a CD1d/a chimeric protein
(lanes 7±12) immunoprecipitated and immunoblotted with af®nity-
puri®ed rabbit anti-CD1d: lanes 1 and 7, normal mouse serum; lanes 2±
4 and 8±10, three sequential immunoprecipitations with 42.1 mAb;
lanes 5, 6 and 11, 12, sequential D5 mAb immunoprecipitations of the
42.1-cleared lysates. All samples were analysed on 12% reducing gels.
Stimulus T cells
No stimulus +
C1R mock +
C1R CD1d +
CHO CD1d (fix) +
Thymocytes (live) +
Thymocytes (fix) +
Thymocytes (live) −
0 20 40 60 80 100 120
% of maximum response
Figure 8. Lack of thymocyte CD1d recognition by Va24invt T cells. A
CD1d-reactive Va24invt clone, DN2.B9, was cultured with the indicated
stimulator cells in the presence of phorbol 12-myristate 13-acetate
(PMA). Proliferation was determined by 3H-thymidine incorporation.
The stimulator cells or DN2.B9 T cells were omitted from the top and
bottom experiments, respectively. The response to the C1R CD1d
transfectants was set at 100%. ®x, light glutaraldehyde ®xation; Mock,
transfected with empty expression vector.
45CD1d structure and regulation of expression
# 2000 Blackwell Science Ltd, Immunology, 100, 37±47

at this developmental stage. This mechanism could involve the
tyrosine-based endosomal targeting signal in the cytoplasmic
tail of CD1d (as well as CD1b and CD1c), but not CD1a.
In contrast to the results presented here in humans,
autoreactive recognition of murine thymocyte CD1d by
homologous murine Va14invt T-cell hybridomas has been
observed.23 This difference could re¯ect less stringent in vitro
requirements for TCR-mediated activation in the murine
hybridomas versus the human Va24invt T-cell clones. Another
signi®cant difference between CD1d in humans and mice is that
murine CD1d is expressed at low levels by both thymocytes and
mature T cells,13,42 and at slightly higher levels by B cells.37,38
We suggest that the expression of group 1 and group 2 CD1
proteins by human thymocytes and their absence in mature
human T cells may be co-ordinately regulated by CD1 locus-
speci®c enhancer and silencer elements, respectively, and that
these elements may have been lost with the deletion of the
group 1 (CD1a, b and c) genes from the murine CD1 locus. If
this is correct, then the regulated expression of CD1d in the
human T-lymphocyte lineage may re¯ect a physiological role in
immunoregulation. Speci®cally, CD1d expression by activated
human T cells may render them as direct in vivo stimulators or
targets of CD1d-reactive T cells. A role for Va24invt T cells in
modulating the function of activated T cells would be
consistent with the loss of Va24invt T cells in a number of
human autoimmune diseases.31,43
ACKNOWLEDGMENTS
We thank Dr T. Strom (Beth Israel Deaconess Medical Center, Boston)
for providing the IgG fusion vector. This work was supported by NIH
R01 grants AI42955 to S. P. B.; AI3319 and DK44319 to S. P. B. and R.
S. B.; DK51362 to R. S. B., AI45051 to S. B. W, AI40135 and an
Arthritis Foundation Investigator Award to S. P.; a Crohns and Colitis
Foundation of America grant to D. G.; and an American Hematology
Society award to K. T. P.
REFERENCES
1. MARTIN L.H., CALABI F., LEFEBVRE F.A., BILSLAND C.A. & MILSTEIN
C. (1987) Structure and expression of the human thymocyte
antigens CD1a, CD1b, and CD1c. Proc Natl Acad Sci USA 84,
9189.
2. ARUFFO A. & SEED B. (1989) Expression of cDNA clones encoding
the thymocyte antigens CD1a, b, c demonstrates a hierarchy of
exclusion in ®broblasts. J Immunol 143, 1723.
3. BALK S.P., BLEICHER P.A. & TERHORST C. (1989) Isolation and
characterization of a cDNA and gene coding for a fourth CD1
molecule. Proc Natl Acad Sci USA 86, 252.
4. MCMICHAEL A.J., PILCH J.R., GALFRE G., MASON D.Y., FABRE
J.W. & MILSTEIN C. (1979) A human thymocyte antigen de®ned
by a hybrid myeloma monoclonal antibody. Eur J Immunol 9,
205.
5. KAHN-PERLES B., WIETZERBIN J., CAILLOL D.H. & LEMONNIER F.
(1985) Delineation of three subsets of class I human T antigens
(HTA) on Molt-4 cells: serologic and regulatory relationship to
HLA class I antigens. J Immunol 134, 1759.
6. AMIOT M., BERNARD A., RAYNAL B., KNAPP W., DESCHILDRE C. &
BOUMSELL L. (1986) Heterogeneity of the ®rst cluster of differentia-
tion: characterization and epitopic mapping of three CD1
molecules on normal human thymus cells. J Immunol 136, 1752.
7. FITHIAN E., KUNG P., GOLDSTEIN G., RUBENFELD M., FENOGLIO C. &
EDELSON R. (1981) Reactivity of Langerhans' cells with hybridoma
antibody. Proc Natl Acad Sci USA 78, 2541.
8. PORCELLI S., MORITA C.T. & BRENNER M.B. (1992) CD1b restricts
the response of human CD4± T lymphocytes to a microbial antigen.
Nature 360, 593.
9. KASINRERK W., BAUMRUKER T., MAJDIC O., KNAPP W. & STOCKINGER
H. (1993) CD1 molecule expression on human monocytes induced
by granulocyte±macrophage colony-stimulating factor. J Immunol
150, 579.
10. SMALL T.N., KNOWLES R.W., KEEVER C. et al. (1987) M241 (CD1)
expression on B lymphocytes. J Immunol 138, 2864.
11. BECKMAN E.M., PORCELLI S.A., MORITA C.T., BEHAR S.M., FURLONG
S.T. & BRENNER M.B. (1994) Recognition of a lipid antigen by
CD1-restricted alpha beta+ T cells. Nature 372, 691.
12. CALABI F., JARVIS J.M., MARTIN L. & MILSTEIN C. (1989) Two classes
of CD1 genes. Eur J Immunol 19, 285.
13. BLEICHER P.A., BALK S.P., HAGEN S.J., BLUMBERG R.S., FLOTTE T.J.
& TERHORST C. (1990) Expression of murine CD1 on gastro-
intestinal epithelium. Science 250, 679.
14. BLUMBERG R.S., TERHORST C., BLEICHER P. et al. (1991) Expression
of a nonpolymorphic MHC class I-like molecule, CD1D, by
human intestinal epithelial cells. J Immunol 147, 2518.
15. CANCHIS P.W., BHAN A.K., LANDAU S.B., YANG L., BALK S.P. &
BLUMBERG R.S. (1993) Tissue distribution of the non-polymorphic
major histocompatibility complex class I-like molecule, CD1d.
Immunology 80, 561.
16. CASTANO A.R., TANGRI S., MILLER J.E. et al. (1995) Peptide binding
and presentation by mouse CD1. Science 269, 223.
17. KAWANO T., CUI J., KOEZUKA Y. et al. (1997) CD1d-restricted and
TCR-mediated activation of Va14 NKT cells by glycosylceramides.
Science 278, 1626.
18. JOYCE S., WOODS A.S., YEWDELL J.W. et al. (1998) Natural ligand of
mouse CD1d1: cellular glycosylphosphatidylinositol. Science 279,
1541.
19. COLES M.C. & RAULET D.H. (1994) Class I dependence of the
development of CD4+ CD8± NK1.1+ thymocytes. J Exp Med 180,
395.
20. OHTEKI T. & MACDONALD H.R. (1994) Major histocompatibility
complex class I related molecules control the development of
CD4+ 8± and CD4± subsets of natural killer 1.1+ T cell receptor-
alpha/beta+ cells in the liver of mice. J Exp Med 180, 699.
21. LANTZ O. & BENDELAC A. (1994) An invariant T cell receptor alpha
chain is used by a unique subset of major histocompatibility
complex class I-speci®c CD4+ and CD4± T cells in mice and
humans. J Exp Med 180, 1097.
22. ADACHI Y., KOSEKI H., ZIJLSTRA M. & TANIGUCHI M. (1995) Positive
selection of invariant V alpha 14+ T cells by non-major
histocompatibility complex-encoded class I-like molecules
expressed on bone marrow-derived cells. Proc Natl Acad Sci
USA 92, 1200.
23. BENDELAC A., LANTZ O., QUIMBY M.E., YEWDELL J.W., BENNINK J.R.
& BRUTKIEWICZ R.R. (1995) CD1 recognition by mouse NK1+ T
lymphocytes. Science 268, 863.
24. PORCELLI S., YOCKEY C.E., BRENNER M.B. & BALK S.P. (1993)
Analysis of T cell antigen receptor (TCR) expression by human
peripheral blood CD4- alpha/beta T cells demonstrates preferential
use of several V beta genes and an invariant TCR alpha chain.
J Exp Med 178, 1.
25. DELLABONA P., PADOVAN E., CASORATI G., BROCKHAUS M. &
LANZAVECCHIA A. (1994) An invariant V alpha 24-J alpha Q/V
beta 11 T cell receptor is expressed in all individuals by clonally
expanded CD4± T cells. J Exp Med 180, 1171.
26. EXLEY M., GARCIA J., BALK S.P. & PORCELLI S. (1997) Requirements
for CD1d recognition by human invariant Valpha24+CD4± CD8± T cells. J Exp Med 186, 109.
46 M. Exley et al.
# 2000 Blackwell Science Ltd, Immunology, 100, 37±47

27. EXLEY M., PORCELLI S., FURMAN M., GARCIA J. & BALK S.P. (1998)
CD161 (NKR-P1A) costimulation of CD1d-dependent activation
of human T cells expressing invariant V alpha 24 J alpha Q T cell
receptor alpha chains. J Exp Med 188, 867.
28. ARASE H., ARASE N., NAKAGAWA K., GOOD R.A. & ONOE K. (1993)
NK1.1+ CD4+ CD8± thymocytes with speci®c lymphokine
secretion. Eur J Immunol 23, 307.
29. YOSHIMOTO T. & PAUL W.E. (1994) CD4pos, NK1.1pos T cells
promptly produce interleukin 4 in response to in vivo challenge with
anti-CD3. J Exp Med 179, 1285.
30. YOSHIMOTO T., BENDELAC A., WATSON C., HU-LI J. & PAUL W.E.
(1995) Role of NK1.1+ T cells in a TH2 response and in
immunoglobulin E production. Science 270, 1845.
31. WILSON S.B., KENT S.C., PATTON K.T. et al. (1998) Extreme Th1
bias of invariant Valpha24JalphaQ T cells in type 1 diabetes.
Nature 391, 177.
32. ZHENG X.X., STEELE A.W., NICKERSON P.W., STEURER W., STEIGER J.
& STROM T.B. (1995) Administration of noncytolytic IL-10/Fc in
murine models of lipopolysaccharide-induced septic shock and
allogeneic islet transplantation. J Immunol 154, 5590.
33. BALK S.P., BURKE S., POLISCHUK J.E. et al. (1994) Beta 2-micro-
globulin-independent MHC class Ib molecule expressed by human
intestinal epithelium. Science 265, 259.
34. RODIONOV D.G., NORDENG T.W., PEDERSEN K., BALK S.P. & BAKKE
O. (1999) A critical tyrosine residue in the cytoplasmic tail is
important for CD1d internalization but not for its basolateral
sorting in MDCK cells. J Immunol 162, 1488.
35. KIM H.S., GARCIA J., EXLEY M., JOHNSON K.W., BALK S.P. &
BLUMBERG R.S. (1999) Biochemical characterization of CD1d
expression in the absence of beta2-microglobulin. J Biol Chem 274,
9289.
36. SPADA F.M., KOEZUKA Y. & PORCELLI S.A. (1998) CD1d-restricted
recognition of synthetic glycolipid antigens by human natural killer
T cells. J Exp Med 188, 1529.
37. AMANO M., BAUMGARTH N., DICK M.D. et al. (1998) CD1
expression de®nes subsets of follicular and marginal zone B cells
in the spleen: beta 2-microglobulin-dependent and independent
forms. J Immunol 161, 1710.
38. ROARK J.H., PARK S.H., JAYAWARDENA J., KAVITA U., SHANNON M. &
BENDELAC A. (1998) CD1.1 expression by mouse antigen-presenting
cells and marginal zone B cells. J Immunol 160, 3121.
39. SPENCER J., PERRY M.E. & DUNN-WALTERS D.K. (1998) Human
marginal-zone B cells. Immunol Today 19, 421.
40. BENDELAC A. (1995) Positive selection of mouse NK1+ T cells by
CD1-expressing cortical thymocytes. J Exp Med 182, 2091.
41. SOMNAY-WADGAONKAR K., NUSRAT A., KIM H.S. et al. (1999)
Immunolocalization of CD1d in human intestinal epithelial cells
and identi®cation of a beta2-microglobulin-associated form. Int
Immunol 11, 383.
42. BROSSAY L., JULLIEN D., CARDELL S. et al. (1997) Mouse CD1 is
mainly expressed on hemopoietic-derived cells. J Immunol 159,
1216.
43. SUMIDA T., SAKAMOTO A., MURATA H. et al. (1995). Selective
reduction of T cells bearing invariant V alpha 24J alpha Q antigen
receptor in patients with systemic sclerosis. J Exp Med 182, 1163.
47CD1d structure and regulation of expression
# 2000 Blackwell Science Ltd, Immunology, 100, 37±47




















