
In Rhizobium etli Symbiotic Plasmid Transfer, Nodulation Competitivity and Cellular Growth Require...
10
In Rhizobium etli Symbiotic Plasmid Transfer, Nodulation Competitivity and Cellular Growth Require Interaction among Different Replicons Susana Brom, 1 Alejandro Garcı ´a-de los Santos, Laura Cervantes, Rafael Palacios, and David Romero Centro de Investigacio ´n sobre Fijacio ´n de Nitro ´geno, Universidad Nacional Auto ´noma de Me ´xico, Apdo. Postal 565-A, C.P. 62210, Cuernavaca, Morelos, Me ´xico Received December 13, 1999; revised February 14, 2000 Bacteria belonging to the genus Rhizobium are able to develop two different lifestyles, in symbiotic association with plant roots or through saprophytic growth. The genome of Rhizobium strains is constituted by a chromosome and several large plasmids, one of them containing most of the genes involved in symbiosis (symbiotic plasmid or pSym). Our model strain Rhizobium etli CFN42 contains six plasmids. We have constructed multiple plasmid-cured derivatives of this strain and used them to analyze the contribution of these plasmids to free-living cellular viability, competitivity for nodula- tion, plasmid transfer, and utilization of diverse carbon sources. Our results show that the transfer of the pSym is strictly dependent on the presence of another plasmid; consequently under conditions where pSym transfer is required, nodulation relies on the presence of a plasmid devoid of nodulation genes. We also found a drastic decrease in competitivity for nodulation in multiple plasmid-cured derivatives when compared with single plasmid-cured strains. Cellular growth and viability were greatly diminished in some multiple plasmid-cured strains. The utilization of a number of carbon sources depends on the presence of specific plasmids. The results presented in this work indicate that functional interactions among sequences scattered in the different plasmids are required for success- ful completion of both lifestyles. © 2000 Academic Press For many years the genomic organization in bacteria was considered to be rather simple, con- sisting of a unique circular chromosome, and in some cases additional genetic material in the form of plasmids. The chromosome has been defined as the carrier of housekeeping genes. This view has been changing, with the description of linear and multiple chromosomes in some bacteria. It has been proposed that the genome of bacterial spe- cies is constituted by the endogenome (universal core of the species’ genes) and the exogenome (variable, with or without selective advantage) (Casjens, 1998); and we would further propose that both may have representation in chromo- somes and plasmids. In Rhizobium and related genera, the ge- nome is usually constituted of a chromosome and a variable number of large plasmids, which may constitute up to 40% of the total genome. Rhizobium strains have the capacity to establish nitrogen-fixing symbioses with leguminous plants. Most of the bacterial genes required for this function are normally present in one of these large plasmids, named as the symbiotic plasmid (pSym) (see Gar- cı ´a-de los Santos et al., 1996, for a review). The isolation and characterization of plas- mid-cured strains has allowed the description of different plasmid-associated functions in rhizo- bia in plasmids other than the pSym (Baldani et al., 1992; Brom et al., 1992; Hynes and McGre- gor, 1990). Among the plasmid-associated traits found are nitrogen fixation, competition for nodulation, genes required for lipopolysaccha- ride (LPS) 2 synthesis, and other catabolic genes. A group of methyl-accepting chemotaxis pro- teins, some of which seem to be involved also in competitiveness, have been found in R. legu- minosarum (Yost et al., 1998). Also mutations 1 To whom correspondence should be addressed. Fax: (73) 17 55 81. E-mail: [email protected]. 2 Abbreviations used: LPS, lipopolysaccharide; nal, nali- dixic acid; sp, spectinomycin; km, kanamycin; nm, neomy- cin; cm, chloramphenicol; rif, rifampin. 34 0147-619X/00 $35.00 Copyright © 2000 by Academic Press All rights of reproduction in any form reserved. Plasmid 44, 34 – 43 (2000) doi:10.1006/plas.2000.1469, available online at http://www.idealibrary.com on
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of In Rhizobium etli Symbiotic Plasmid Transfer, Nodulation Competitivity and Cellular Growth Require...

(
0
Plasmid44, 34–43 (2000)doi:10.1006/plas.2000.1469, available online at http://www.idealibrary.com on
In Rhizobium etli Symbiotic Plasmid Transfer, Nodulation Competitivityand Cellular Growth Require Interaction among Different Replicons
Susana Brom,1 Alejandro Garcı´a-de los Santos, Laura Cervantes,Rafael Palacios, and David Romero
Centro de Investigacio´n sobre Fijacion de Nitrogeno, Universidad Nacional Auto´noma de Me´xico,Apdo. Postal 565-A, C.P. 62210, Cuernavaca, Morelos, Me´xico
Received December 13, 1999; revised February 14, 2000
Bacteria belonging to the genusRhizobiumare able to develop two different lifestyles, in symbioticassociation with plant roots or through saprophytic growth. The genome ofRhizobiumstrains isconstituted by a chromosome and several large plasmids, one of them containing most of the genesinvolved in symbiosis (symbiotic plasmid or pSym). Our model strainRhizobium etliCFN42 containssix plasmids. We have constructed multiple plasmid-cured derivatives of this strain and used them toanalyze the contribution of these plasmids to free-living cellular viability, competitivity for nodula-tion, plasmid transfer, and utilization of diverse carbon sources. Our results show that the transfer ofthe pSym is strictly dependent on the presence of another plasmid; consequently under conditionswhere pSym transfer is required, nodulation relies on the presence of a plasmid devoid of nodulationgenes. We also found a drastic decrease in competitivity for nodulation in multiple plasmid-curedderivatives when compared with single plasmid-cured strains. Cellular growth and viability weregreatly diminished in some multiple plasmid-cured strains. The utilization of a number of carbonsources depends on the presence of specific plasmids. The results presented in this work indicate thatfunctional interactions among sequences scattered in the different plasmids are required for success-ful completion of both lifestyles. © 2000 Academic Press
inco
informedha
andhasprsomge)osmo
e-medstality
ithiallly
medar-.as-n ofzo-i
e-aitsorha-es.
ro-o in
s
Faali-
d my-c
For many years the genomic organizationbacteria was considered to be rather simple,sisting of a unique circular chromosome, andsome cases additional genetic material in theof plasmids. The chromosome has been definthe carrier of housekeeping genes. This viewbeen changing, with the description of linearmultiple chromosomes in some bacteria. Itbeen proposed that the genome of bacterialcies is constituted by the endogenome (univecore of the species’ genes) and the exogen(variable, with or without selective advanta(Casjens, 1998); and we would further propthat both may have representation in chrosomes and plasmids.
In Rhizobiumand related genera, the gnome is usually constituted of a chromosoand a variable number of large plasmiwhich may constitute up to 40% of the togenome.Rhizobiumstrains have the capac
1 To whom correspondence should be addressed.73) 17 55 81. E-mail: [email protected].
34147-619X/00 $35.00Copyright © 2000 by Academic PressAll rights of reproduction in any form reserved.
n-
ass
se-ale
e-
,
to establish nitrogen-fixing symbioses wleguminous plants. Most of the bactergenes required for this function are normapresent in one of these large plasmids, naas the symbiotic plasmid (pSym) (see Gcıa-de los Santoset al., 1996, for a review)
The isolation and characterization of plmid-cured strains has allowed the descriptiodifferent plasmid-associated functions in rhibia in plasmids other than the pSym (Baldanetal., 1992; Bromet al.,1992; Hynes and McGrgor, 1990). Among the plasmid-associated trfound are nitrogen fixation, competition fnodulation, genes required for lipopolysaccride (LPS)2 synthesis, and other catabolic genA group of methyl-accepting chemotaxis pteins, some of which seem to be involved alscompetitiveness, have been found inR. legu-minosarum(Yost et al., 1998). Also mutation
x:
2 Abbreviations used: LPS, lipopolysaccharide; nal, nixic acid; sp, spectinomycin; km, kanamycin; nm, neoin; cm, chloramphenicol; rif, rifampin.

elsi-
insinin
s
se
m
ex-rilelee i
a
c obg
tef a-t ds,a avbs1t f as ngp Thc lat l-g ncfs toc 00k sos vert wnA anR d in db edw rrei
av
o m-p dt s indC s toi idi
sw1 4),o am-m andct ).G e int ricc er.C yerfl iths ereg a-t ingp ro-n theirg oft
tyrly,ofticsna-
l
.
ndkers
sRG ingt o-s s.
35RHIZOBIUM ETLI SYMBIOTIC PLASMID TRANSFER
in the rhamnose utilization genes inR. trifoliiresult in an alteration in competition lev(Oresnik et al., 1998). A plasmid-encoded ctrate synthase gene has been describedR.tropici (Pardoet al., 1994), which affects itnodulation ability. Plasmid-encoded genesvolved in LPS synthesis have been founddifferent R. etli strains (Garcı´a-de los Santoand Brom, 1997; Vinuesaet al., 1999) and inAzospirillum brasilense(Katzy et al., 1998).
Some plasmids have been found to be estial for growth. p42f of theR. etlistrain CFN42is required for growth on minimal mediu(Brom et al., 1992). In theR. trifolii strainW14-2, single plasmid-cured derivatives (cept one) are not affected for growth in stesoil, but different combinations of multipplasmid-cured derivatives show a decreasgrowth that in some cases corresponds toabsolute inability for growth (Moe¨nne-Loccozand Weaver, 1995).Alcaligenes eutrophusH16ontains a plasmid that is essential for anaerrowth on nitrate (Siedowet al., 1999).Among the most relevant plasmid associa
unctions is their ability to transfer genetic merial. Self-transmissible symbiotic plasmissociated to production of bacteriocins, heen described in someR. leguminosarumtrains (Johnstonet al., 1982; Brewin et al.,980). Transfer of the pSym of a strain ofR.
rifolii was achieved through the formation otable cointegrate with the broad-host-ralasmid R68.45 (Scott and Ronson, 1982).omparison of geographically distant popuions ofRhizobiumisolated fromPhaseolus vuaris nodules presents circunstantial evide
or pSym transfer (Geniauxet al., 1993).Me-orhizobium lotihas recently been shownarry a symbiotic island of approximately 5b, which has been transferred to othertrains (Sullivan and Ronson, 1998). Howehe mechanism of its transmission is not knolso, broad-host-range plasmids pJP468.45 have been shown to be transferreonsterile soil (Kinkleet al., 1993), restrictey the fact that only some strains functionell as receptors and that the transfer occu
n soil or rhizosphere, but not in nodules.In this paper we present the results we h
-
n-
nn
ic
d
e
ee-
e
il,.dn
d
e
btained by analyzing transfer functions, coetition for nodulation, growth, viability, an
he capacity to use some carbon sourceifferent plasmid-cured derivatives ofR. etliFN42. The approach we used allowed u
dentify functions which involve interplasmnteractions.
MATERIALS AND METHODS
Bacterial strains and growth conditions.Thetrains used are listed in Table 1.R. etli strainsere grown at 30°C in PY medium (Noelet al.,984), minimal M9 medium (Beringer, 197r Fahraeus (Fahraeus, 1957) with 10 mMonium chloride and succinate as nitrogen
arbon sources.Escherichia coliandAgrobac-eriumstrains were grown on LB (Miller, 1972rowth rates were measured by the increas
urbidity in a Klett–Summerson photoelectolorimeter, with a 500- to 700-nm-range filtells were incubated in 250-ml Erlenmeasks with 100 ml of medium, at 30°C, whaking (100 rpm). Overnight cultures wrown on PY medium, washed with sterile w
er, and used for inoculation. Strains lack42b flocculate in liquid media; to avoid ereous measurements caused by this,rowth was determined by counting the cfu
he cultures, diluted in 10 mM MgSO4–0.01%Tween 40 (vol/vol) and plated on PY. Viabilimeasurements were performed similathrough determination of the cfu of dilutionsculture samples. When required, antibiowere added at the following concentrations:lidixic acid (nal) 20 mg ml21; spectinomycin(sp) 75mg ml21; kanamycin (km) 15mg ml21;neomycin (nm) 60mg ml21; cloramphenico(cm) 20mg ml21; rifampin (rif) 50 mg ml21 forRhizobiumor 100mg ml21 for Agrobacterium
Genetic manipulations.Transposon Tn5-GDYN-1 carries kanamycin–gentamicin aspectinomycin–streptomycin resistance marand sacRsacBgenes bordered by the IS50 in-ertion sequences of Tn5 (Floreset al., 1993).hizobium derivatives containing Tn5-DYN-1 insertions were obtained by mat
hem withE. coli S-17 containing the transpon and selecting nalR spR kmR transconjugant

er-5%
s
s esw 85(
id
sis-214dersenter-ghtnorble
R
s
36 BROM ET AL.
Plasmid curing was carried out by plating ovnight cultures on PY plates containing 12.sucrose. Sucrose-resistant colonies werelected and verified for spS and kmS. To con-truct the multiple plasmid-cured derivative obtained a derivative of strain CFNX1
containing peD) with a Tn5-GDYN-1 insertionin plasmid c. A derivative cured of this plasm
TAB
Bacterial Str
Strain Relevant featur
. etliCFN42 Wild type, contains plaCFN2001 CFN42 derivative (pa2 pCFNX182 CFN42 derivative (pa2)CFNX183 CFN42 derivative (pb2)CFNX184 CFN42 derivative (pc2)CFNX89 CFN42 derivative (pd2)CFNX185 CFN42 derivative (peD)CFNX186 CFN42 derivative (pf2)CFNX180 CFN42 with chromosoCFNX187 CFNX182 complementCFNX190 CFNX183 complementCFNX191 CFNX184 complementCFNX192 CFNX89 complementeCFNX193 CFNX185 complementCFNX194 CFNX186 complementCFN42recA2 recA2 derivative of CFNCFNX214 CFN42 derivative (pc2, pCFNX215 CFN42 derivative (pc2, pCFNX216 CFN42 derivative (pa2, pCFNX217 CFN42 derivative (pa2, pCFNX218 CFN42 derivative (pa2, pCFNX220 CFN42 derivative (pa2, pCFNX221 CFNX216 derivative (pCFNX223 CFNX217 derivative (pCFNX224 CFNX220 derivative (pCFNX195 CFNX182 derivative (pCFNX196 CFNX42recA2 derivativeCFNX197 CFN42 derivative withCFNX198 CFNX218 derivative w
Agrobacterium tumefaciensGMI9023 C-58 cured of its native
Escherichia coliS17/pSUP5011 Source of Tn5mobS17/pDR21 Source of Tn5 GDYN-1HB101/pRK2013 Conjugation helper
PlasmidspAGS10 region 3.6-kbEcoRI–BamHI fra
biosynthesis genes f
e-
,
was obtained through selection of sucrose-retant colonies, thus generating strain CFNX(pc2, peD). The procedure was repeated in orto sequentially lose the other plasmids prein CFN42. Conjugation experiments were pformed on PY plates at 30°C, using overnicultures grown to the stationary phase. Doand recipient were mixed in a 1:2 ratio. Suita
1
and Plasmids
lost or modified plasmids) Reference
ids a, b, c, d, e, and f Quintoet al., 1985Leemanset al., 1984Brom et al., 1992Brom et al., 1992Brom et al., 1992Brom et al., 1992Brom et al., 1992Brom et al., 1992
Tn5mobinsertion Bromet al., 1992with pa::Tn5mob Brom et al., 1992with pb::Tn5mob Brom et al., 1992with pc::Tn5mob Brom et al., 1992ith pd::Tn5mob Brom et al., 1992with pe::Tn5mob Brom et al., 1992with pf::Tn5mob Brom et al., 1992
J. Martınez, CIFNThis work
pf2, pa::Tn5) This workpeD, pf2) This workpc2, peD, pf2) This workpc2, pd2, peD, pf2) This workpc2, peD) This workn5 GDYN) This workn5 GDYN) This workn5 GDYN) This workn5mob) This work
ith pd::Tn5mobfrom CFNX195 This work:Tn5mobfrom CFNX195 This workpf::Tn5moband pAGS10 This work
asmids Rosenberg andHughet, 1984
Simon, 1984Floreset al., 1993
Figurski andHelinski, 1979
ent containinglpsb (LPSp42b)
Garcıa de los Santoand Brom, 1997
LE
ains
es (
smd2)
maledededd weded42eD)eD,c2,b2,b2,b2,d::Td::Td::Td::T
wpd:ith
pl
gmrom

tio17ve
as01/
iumthedein
redal
ei
is.ckntorinyberesnde
ofla
w ndd ra-t s
thellhe
T diaf ingc ntib n inF ont at2 det odu ndc anp
cy
t inion,-
mG ce-d ds.T iss sfero it ata oft 42f,w nep bet ds,s rp
T nt
oft ds.T thep a-
rain
37RHIZOBIUM ETLI SYMBIOTIC PLASMID TRANSFER
markers were used for transconjugant selecCFNX195 was obtained by crossing SpSUP5011 with CFNX185 (CFN42 derivatilacking p42a), and selecting for nalR and nmR.The Tn5mob-labeled derivatives were useddonors in triparental crosses, with HB1pRK2013 as a helper andAgrobacteriumGMI9023 as receptor. The selective medcontained rif and nm. Plasmid profiles oftransconjugants allowed us to identify therivatives containing the marker elementp42d. p42d of strain CFNX195 was transferto a rifR derivative of CFN42, in a triparentcross, selecting for nmR and rifR; and to arecA2
derivative of CFN42, selecting for cmR andnmR. Transconjugants were analyzed for thplasmid pattern and its hybridization withnifHand p42a.
Plasmid profiles and hybridization analysPlasmid patterns were visualized by the Ehardt procedure (Eckhardt, 1978), blotted onylon membranes, and hybridized under stgent conditions, using Amersham’s Rapid-hbuffer as specified by the manufacturer (Amsham Corp.). Probes were linearized by digtion with appropriate restriction enzymes alabeled with [32P]a-CTP using a RediprimDNA labeling system (Amersham Corp.).
Competition assays. Phaseolus vulgarisrootswere inoculated with a quantified mixturestrains (107 cells/plant). Bacteria for inocu
ere grown overnight on PY, centrifuged, ailuted in sterile water. Inoculum concent
ions were determined byA600. Cell numberwere adjusted in suspensions to formulatemixtures of inoculum strains. The final cnumbers were verified by serially diluting tinocula in 10 mM MgSO4, 0.01% (vol/vol)
ween 40, plating them on appropriate meor identification of each strain, and countfu. One of the strains always carried an aiotic resistance marker. Plants were growahraeus containing agar medium, in a c
rolled chamber with a 12-h photoperiod,8°C for 18 days. Nodule occupancy was
ermined by squashing surface-sterilized nles, isolating colonies on PY medium, ahecking their antibiotic resistance markerslasmid profile.
n./
-
r
-
-
--
e
-
-
--
d
RESULTS
p42a Is Self-Transmissible at High Frequen
The transference of the plasmids presenstrain CFN42 was evaluated by conjugatusing derivatives labeled with Tn5 in each plas
id as donors andA. tumefaciensstrainMI9023 as recipient, according to the proure described under Materials and Methohe results (Table 2) show that only p42aelf-transmissible at a high frequency. Tranf the pSym (p42d) was also detected, albelow frequency, while the transfer frequency
he other plasmids, p42b, p42c, p42e, and pas below our levels of detection. We defi42a as self-transmissible because it may
ransferred from other genomic backgrounuch asAgrobacterium,containing no othelasmids (data not shown).
he Transfer of the pSym Is Fully Dependeon the Presence of p42a and Partiallyon RecA
We determined the transfer frequencyhe pSym in different genomic backgrouno perform these experiments, we labeledSym of strain CFNX182 (pa-cured deriv
TABLE 2
Transfer Frequency ofR. etli CFN42 Plasmidsa
Donor Relevant genotypeTransfer
frequencyb
CFNX187 pa::Tn5mob 1 3 1022
CFNX190 pb::Tn5mob ndc
CFNX191 pc::Tn5mob ndCFNX192 pd::Tn5mob 2 3 1026
CFNX193 pe::Tn5mob ndCFNX194 pf::Tn5mob ndCFNX195 pa2, pd::Tn5mob ndCFNX197 pa1, pd::Tn5mob
(CFNX195)6 3 1025
CFNX196 recA2, pa1, pd::Tn5mob(CFNX195)
9 3 1027
a All crosses were repeated at least twice. StGMI9023 was always used as the receptor.
b Expressed as transconjugants per donor.c Not detectable (below 1029).

inoinhe
aonndonncy
icthain
C freqr al
esnotecthetheym
leca
ated
ym
g it edt er-m twc t oa
W
m( onP sl du-l ideb db andM 89a antwb ild-t mee ainC yma
ntser-chcir-
tiones-
/orc-s ofid
y athis
theless:gants 10,
38 BROM ET AL.
tive of CFN42) with Tn5mob, generatingstrain CFNX195. The pSym of straCFNX195 was subsequently transferred trecA2 derivative of CFN42 to construct straCFNX196. Strain CFNX197 resulted from ttransfer of the pSym of strain CFNX195 towild-type receptor (CFN42). The conjugatiand selection procedures are described uMaterials and Methods. These constructiallowed us to compare the transfer efficieof the same pSym from different genombackgrounds. The results (Table 2) showthe Tn5mob-labeled pSym from stra
FNX197 is transferred at the expecteduency from a wild-type donor (p42a1,ecA1); when the donor lacks a function
RecA, the transfer frequency diminishabout 60 times. In contrast, when the dolacks p42a, we are no longer able to detransfer of the pSym. Figure 1 exemplifiesplasmid patterns observed amongtransconjugants: Some show plasmids pSand p42a, and others contain a higher moular weight plasmid, which is shown to becointegrate of both plasmids, as demonstrby its hybridization with both of them, ansome (10% or less) present only the pSThis is similar in recA1 or recA2 back-
rounds. Our interpretation of these datahat the transfer of the pSym is achievhrough cointegration with p42a. Furthore, our results suggest that there are
ointegration mechanisms, one dependennd one independent of RecA.
FIG. 1. Plasmid pattern of transconjugants seleethidium-bromide-stained Eckhardt-type gels; lanewith a nifH probe; and lanes 3, 6, 9, and 12 showCFNX192 (donor): lanes 1, 2, and 3; transconjugawith pSym and p42a: lanes 7, 8, and 9; transconju11, and 12.
a
ers
t
-
rt
-
d
.
s
on
hen pSym Transfer Is Required, p42aBecomes Indispensable for Nodulation
Derivatives of CFN42 lacking the pSyCFNX89) are unable to form noduleshaseolus vulgarisroots. Also, derivative
acking p42b (CFNX183) are unable to noate, due to an impairment in lipopolysacchariosynthesis (Bromet al.,1992). We inoculateean roots (as described under Materialsethods) with a mixture of strains CFNXnd CFNX183. Approximately 30 nodules/plere obtained 15 days postinoculation.Rhizo-ium strains isolated from nodules had a w
ype plasmid profile (data not shown). The saxperiment was performed mixing strFN2001 (CFN42 derivative lacking the pSnd p42a) with a p42a2 derivative of strain
CFNX183. In this case no nodulated plawere obtained. Both experiments were pformed twice, inoculating six plants with eamixture. These data show that under somecumstances, a plasmid devoid of nodulagenes may be indispensable for symbiosistablishment.
Wild-Type Nodulation Competitivity LevelRequires Interaction of Different Plasmids
In order to approach the individual andjoint participation of plasmids in cellular funtions, we constructed a series of derivativestrain CFN42 which contain different plasmcombinations, ranging from one lacking onlsegment of p42e to one containing only
d for transfer of the pSym. Lanes 1, 4, 7, and 10 are, 5, 8, and 11 show the hybridization of the plasmid profi
hybridization of the plasmid profiles with p42a. Strainresenting only the pSym: lanes 4, 5, and 6; transconjut containing a cointegrate of the pSym and p42a: lane
ctes 2the
nt pgan
oesds
. 2insm
ionlas6
-micu-00in-in
on, aro-eirlum
esThigneab
inhasandym
inee-onin-forthatinus.
s
sedes toea-
hecel-se,no-and
ed
rep-(P)
3:vsach
■ 0t na-l
ingriv-
,C
,
C
39RHIZOBIUM ETLI SYMBIOTIC PLASMID TRANSFER
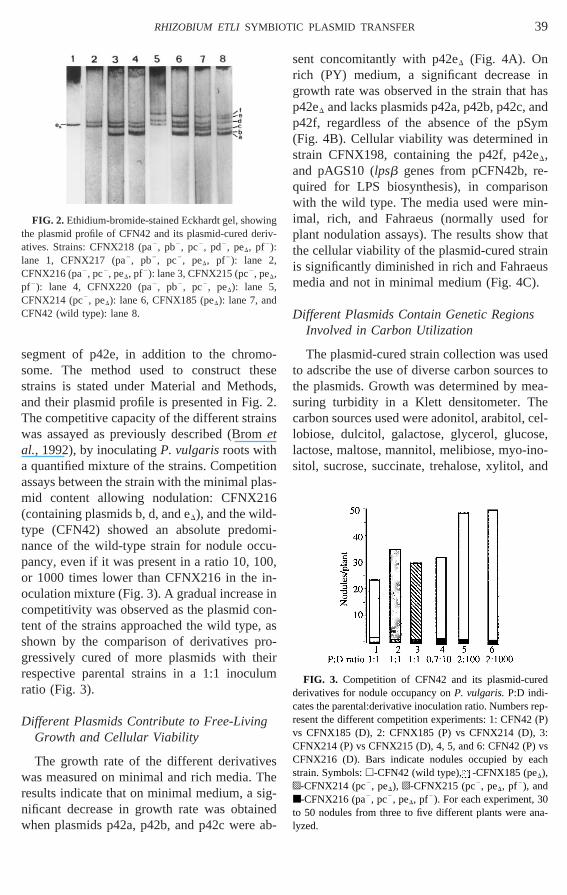
segment of p42e, in addition to the chromsome. The method used to construct thstrains is stated under Material and Methoand their plasmid profile is presented in FigThe competitive capacity of the different strawas assayed as previously described (Broetal., 1992), by inoculatingP. vulgarisroots witha quantified mixture of the strains. Competitassays between the strain with the minimal pmid content allowing nodulation: CFNX21(containing plasmids b, d, and eD), and the wildtype (CFN42) showed an absolute predonance of the wild-type strain for nodule ocpancy, even if it was present in a ratio 10, 1or 1000 times lower than CFNX216 in theoculation mixture (Fig. 3). A gradual increasecompetitivity was observed as the plasmid ctent of the strains approached the wild typeshown by the comparison of derivatives pgressively cured of more plasmids with threspective parental strains in a 1:1 inocuratio (Fig. 3).
Different Plasmids Contribute to Free-LivingGrowth and Cellular Viability
The growth rate of the different derivativwas measured on minimal and rich media.results indicate that on minimal medium, a snificant decrease in growth rate was obtaiwhen plasmids p42a, p42b, and p42c were
FIG. 2. Ethidium-bromide-stained Eckhardt gel, showthe plasmid profile of CFN42 and its plasmid-cured deatives. Strains: CFNX218 (pa2, pb2, pc2, pd2, peD, pf2):lane 1, CFNX217 (pa2, pb2, pc2, peD, pf2): lane 2
FNX216 (pa2, pc2, peD, pf2): lane 3, CFNX215 (pc2, peD,pf2): lane 4, CFNX220 (pa2, pb2, pc2, peD): lane 5CFNX214 (pc2, peD): lane 6, CFNX185 (peD): lane 7, and
FN42 (wild type): lane 8.
-e,.
-
-
,
-s
e-d-
sent concomitantly with p42eD (Fig. 4A). Onrich (PY) medium, a significant decreasegrowth rate was observed in the strain thatp42eD and lacks plasmids p42a, p42b, p42c,p42f, regardless of the absence of the pS(Fig. 4B). Cellular viability was determinedstrain CFNX198, containing the p42f, p42D,and pAGS10 (lpsb genes from pCFN42b, rquired for LPS biosynthesis), in compariswith the wild type. The media used were mimal, rich, and Fahraeus (normally usedplant nodulation assays). The results showthe cellular viability of the plasmid-cured strais significantly diminished in rich and Fahraemedia and not in minimal medium (Fig. 4C)
Different Plasmids Contain Genetic RegionInvolved in Carbon Utilization
The plasmid-cured strain collection was uto adscribe the use of diverse carbon sourcthe plasmids. Growth was determined by msuring turbidity in a Klett densitometer. Tcarbon sources used were adonitol, arabitol,lobiose, dulcitol, galactose, glycerol, glucolactose, maltose, mannitol, melibiose, myo-isitol, sucrose, succinate, trehalose, xylitol,
FIG. 3. Competition of CFN42 and its plasmid-curderivatives for nodule occupancy onP. vulgaris.P:D indi-cates the parental:derivative inoculation ratio. Numbersresent the different competition experiments: 1: CFN42vs CFNX185 (D), 2: CFNX185 (P) vs CFNX214 (D),CFNX214 (P) vs CFNX215 (D), 4, 5, and 6: CFN42 (P)CFNX216 (D). Bars indicate nodules occupied by estrain. Symbols:h-CFN42 (wild type), -CFNX185 (peD),p-CFNX214 (pc2, peD), o-CFNX215 (pc2, peD, pf2), and
-CFNX216 (pa2, pc2, peD, pf2). For each experiment, 3o 50 nodules from three to five different plants were ayzed.

outoildesilit
nde tobonsuc-the
ribe
entre-
las-as-be
est-in
erit-las-ymoth,n
is2a.eende-
for-re-
bems,t ofon-ese
herete-
rma-
im-toole
usof
hor-eent are9).ples
itsp ndm owtr N42(
pA inc foP eu-m leat ainw Y,c tye 0°CS
al
40 BROM ET AL.
trigonelline. We also checked arabinose, cmaric acid, hydroxybenzoic acid, malate, procatechuate, and sorbitol, but found that the wtype strain was unable to use any of thcarbon sources. The results show that the ab
FIG. 4. Growth rate and viability of CFN42 andlasmid-cured derivatives. A: Growth on rich (PY) ainimal media. Growth is expressed as the specific gr
ate compared to that of the wild type. Strains: 1: CFw.t.), 2: CFNX214 (pc2, peD), 3: CFNX220 (pa2, pb2, pc2,
peD), 4: CFNX215 (pc2, peD, pf2), 5: CFNX217 (pa2, pb2,c2, peD, pf2), 6: CFNX218 (pa2, pb2, pc2, pd2, peD, pf2).ll derivatives had a significantly different growth rateomparison to the wild-type strain, at 90% confidenceY and 95% for minimal medium, determined by a Nan–Keuls test. Each experiment was repeated at
wice. Absolute growth rate values for the wild-type strere 12.86 (PY, turbidity), 5.85 (MM, turbidity), 2.5 (Pfu), and 0.62 (MM, cfu). B, C, and D: Cellular viabilixpressed as cells/ml. Determined for 8 to 10 days at 3trains: CFN42 (w.t.) and CFNX198 (containing peD, pf,
and pAGS10) are indicated. B: PY medium; C: minimmedium; and D: Fahraeus medium.
---ey
to use glycerol is related to p42b, dulcitol atrigonelline are related to p42c, and melibiosp42e. The growth on a number of other carsources (cellobiose, galactose, xyitol, andcinate) was also seen to be diminished instrain CFNX198, but we were unable to ascthese phenotypes to a specific plasmid.
DISCUSSION
In this paper we address the study of differplasmid-associated features. The first onegards a characteristic commonly present in pmids: conjugative transfer. Only one of the plmids present in CFN42 (p42a) was found toself-transmissible at a high frequency. Interingly, the pSym could be transferred solelythe presence of p42a. Transconjugants inhing the pSym marker usually presented a pmid corresponding to a cointegrate of the pSand p42a, or both plasmids separately, in bwild-type, andrecA2 donors. Our interpretatioof these results is that transfer of the pSymachieved through its cointegration with p4The presence of genetic regions shared betwthese two plasmids has been previouslyscribed in our laboratory (Girardet al., 1991);these may be the substrates for cointegratemation. The fact that the pSym transfer fquency decreased about 60-fold inrecA2 do-nors suggests that cointegration mayaccomplished at least through two mechanisone dependent on and one independenRecA. The fact that some transconjugants ctaining only the pSym are also observed in thcrosses may correspond to derivatives wp42a was lost after the resolution of the coingrate, as no selective pressure for the penence of this plasmid was exercised.
Recent literature has acknowledged thepact of gene transfer in evolution, in additionadaptative functions. The analysis of the whsequence of theE. coli genome has allowedto determine that a very significant fractionthis genome (17.6%) was acquired throughizontal transfer. Also gene transfer has bshown to occur even between organisms thaphylogenetically very distant (Davidson, 199Comparative genomics has provided exam
h
r
st
.

insthere-or
ia,msndcoer
enon-verd
fheaat
lassd
tht thA)thechadeedain
herinsly,en
e-al
a tionc vio 2ac el( isa der m-p inge ain
Cwend-cy.onsr-the
togiesr-ky,d to
resnts,
s,der-of
wthted,
ywehe-.nte are--5),,omic
97
dly,tro-e of
.
-
o-ant
41RHIZOBIUM ETLI SYMBIOTIC PLASMID TRANSFER
where two very distant or not related protehave a similar function. This has led tointerpretation that original information has fquently been displaced by proteins of viralplasmidic origin (Forterre, 1999). In rhizobthe isolation of strains containing similar pSyin different chromosomal backgrounds (avice versa) suggests that pSym transfer hastributed to the generation of new strains (Hrera-Cerveraet al., 1999; Schofieldet al.,1987). Presently our knowledge about gtransfer and its regulation in natural envirments is still scarce (Davidson, 1999); howeplasmid transfer inR. melilotiseems to depenon the nitrogen source (Herrera-Cerveraet al.,1996); inPseudomonas,conjugative transfer ochromosomal genes in the rhizosphere of whas been reported, where plant roots grestimulate bacterial growth and conjugative pmid transfer (Troxleret al., 1997). Also, cellcompetent for transformation are developeinplanta under nutrient limitation inRalstoniasolanacearum(Bertolla et al., 1999).
Our results indicate that in the case ofR. etli,pSym transfer is absolutely dependent onpresence of another plasmid (p42a), so thapSym transfer will ultimately be subject to: (the presence of p42a, (B) the feasibility ofcointegration mechanisms, and (C) the menism that regulates transfer of p42a. The unstanding of these processes will be addressthe future. The presence of nonsymbiotic strin soil has been described (Segoviaet al.,1991).The pSym transfer mechanism describedmay be the link through which these straacquire their nodulation ability, not randombut subject to the regulatory mechanisms mtioned above.
The isolation of multiple plasmid-cured drivatives ofR. etli CFN42 allowed us to reve
drastic effect of plasmid loss on the nodulaompetitiveness of the strain. We had preusly shown that all plasmids, except p4ontribute to the wild-type competitive levBrom et al., 1992); here we show that there
drastic decrease in competitivity when aivative lacking an additional plasmid is coared with its parental strain. The most strikffect was exposed by the comparison of str
n--
e
,
tly-
ee
-r-ins
e
-
-,
-
s
FNX216 (containing only p42b, p42eD, andthe pSym) and CFN42 (wild type), whereshowed that CFNX216 is at least a thousafold less competitive for nodule occupanThese results imply that the genetic regiinvolved in competitivity present in the diffeent plasmids depend on each other to attainwild-type competition levels. This led uspropose that, in addition to the usual stratefor manipulating competitivity, pointed at paticular features (see Triplett and Sadows1992 for a review), other approaches, steerethe modification of global genomic featuthrough the use of genomic rearrangemesuch as amplifications (Mavinguiet al., 1997;Romero et al., 1995; Romero and Palacio1997), may open other perspectives to unstand and manipulate the competitive abilityRhizobium.
It is possible that plasmid-associated groability and competition phenotypes are relaas shown forR. trifolii (Oresniket al., 1998).Also, it is logical to think that cellular viabilitmay affect competitivity; neverthelessshould not discard the prospect of finding pnotypes that are independent of each other
Also for growth and viability under differeconditions plasmid loss was found to havdecremental effect, somewhat similar to thesults found withR. trifolii plasmid-cured derivatives (Moenne-Loccoz and Weaver, 199further supporting the notion that inRhizobiumplasmids and chromosomes share the gencore that defines a particular species.
ACKNOWLEDGMENTS
This work was partially supported by Grant IN2032from DGAPA, UNAM.
REFERENCES
Baldani, J. I., Weaver, R. W., Hynes, M. F., and EarD. B. (1992). Utilization of carbon substrates, elecphoretic enzyme patterns, and symbiotic performancplasmid-cured clover rhizobia.Appl. Environ. Microbiol58, 2308–2314.
Beringer, J. E. (1974). R factor transfer inRhizobium leguminosarum. J. Gen. Microbiol.84, 188–198.
Bertolla, F., Frostgard, A., Brito, B., Nesme, X., and Simnet, P. (1999). During infection of its host, the pl

a.
B V.,r o
B res,2).
nce
C re o
D ria
E of
F byiqu
F ofen-
Ftio-
i.
F byexal
G cterliima
96)ns.
om
.
R.m-i.
, G-t fiv-
996).gal
idsfor-
-
andin
.
ndDa-
ki-n benon-
,andlie-10.
nds.,
om
s.”ar-
ids
N nd
O st,ata-
lved-
P .
Q ,it-
R S.,.
42 BROM ET AL.
pathogenRalstonia solanacearumnaturally developsstate of competence and exchanges genetic materialMol.Plant–Microbe Interact.11, 467–472.
rewin, N. J., Beringer, J. E., Buchanan-Wollaston, A.Johnston, A. W. B., and Hirsch, P. R. (1980). Transfesymbiotic genes with bacteriocinogenic plasmids inRhi-zobium leguminosarum. J. Gen. Microbiol.116, 261–270.
rom, S., Garcı´a-de los Santos, A., Stepkowsky, T., FloM., Davila, G., Romero, D., and Palacios, R. (199Different plasmids ofRhizobium leguminosarumbv.phaseoli are required for optimal symbiotic performaJ. Bacteriol.174,5183–5189.asjens, S. (1998). The diverse and dynamic structubacterial genomes.Annu. Rev. Genet.32, 339–378.avidson, J. (1999). Genetic exchange between bactethe environment.Plasmid42, 73–91.
ckhardt, T. (1978). A rapid method for the identificationplasmid deoxyribonucleic acid in bacteria.Plasmid 1,584–588.
ahraeus, G. (1957). The infection of clover root hairnodule bacteria studied by a single glass slide technJ. Gen. Microbiol.16, 374–381.
igurski, D. H., and Helinski, D. R. (1979). Replicationan origin containing derivative of plasmid RK2 depdent on a plasmid function provided in trans.Proc. Natl.Acad. Sci. USA76, 1648–1652.
lores, M., Brom, S., Stepkowski, T., Girard, L., Da´vila, G.,Romero, D., and Palacios, R. (1993). Gene amplificain Rhizobium:Identification and in vivo cloning of discrete amplifiable DNA regions (amplicons) fromRhizo-bium leguminosarumbv. phaseoli.Proc. Natl. Acad. ScUSA90, 4932–4936.
orterre, P. (1999). Displacement of cellular proteinsfunctional analogues from plasmids or viruses couldplain puzzling phylogenies of many DNA informationproteins.Mol. Microbiol. 33, 4457–4465.arcıa-de los Santos, A., and Brom, S. (1997). Charaization of two plasmid-bornelpsb loci of Rhizobium etrequired for lipopolysaccharide synthesis and for optinteraction with plants.Mol. Plant–Microbe Interact.10,891–902.
Garcıa-de los Santos, A., Brom, S., and Romero, D. (19Rhizobium plasmids in bacteria–legume interactioWorld J. Microbiol. Biotechnol.12, 119–125.
Geniaux, E., Laguerre, G., and Amarger, N. (1993). Cparison of geographically distant populations ofRhizo-bium isolated from root nodules ofPhaseolus vulgarisMol. Ecol. 2, 1–8.
Girard, L., Flores, M., Brom, S., Romero, D., Palacios,and Davila, G. (1991). Structural complexity of the sybiotic plasmid ofRhizobium leguminosarumbv. phaseolJ. Bacteriol.173,2411–2419.
Herrera-Cervera, J. A., Caballero-Mellado, J., LaguerreTichy, H. V., Requena, N., Amarger, N., Martı´nez-Romero, E., Olivares, J., and Sanjuan, J. (1999). At leasrhizobial species nodulatePhaseolus vulgarisin a spanish soil.FEMS Microbiol. Ecol.30, 87–97.
f
.
f
in
e.
n
-
-
l
.
-
,
.,
e
Herrera-Cervera, J. A., Olivares, J., and Sanjuan, J. (1Ammonia inhibition of plasmid pRmeGR4a conjutransfer betweenRhizobium melilotistrains.Appl. Envi-ron. Microbiol. 62, 1145–1150.
Hynes, M. F., and McGregor, N. F. (1990). Two plasmother than the nodulation plasmid are necessary formation of nitrogen-fixing nodules byRhizobium leguminosarum. Mol. Microbiol.4, 567–574.
Johnston, A. W. B., Hombrecher, G., Brewin, N. J.,Cooper, M. C. (1982). Two transmissible plasmidsRhizobium leguminosarumstrain 300.J. Gen. Microbiol128,85–93.
Katzy, E. I., Matora, L. Y., Serebrennikova, O. B., aScheludko, A. V. (1998). Involvement of a 120-Mplasmid ofAzospirillum brasilenseSp245 in the production of lipopolysaccharides.Plasmid40, 73–83.
Kinkle, B. K., Sadowsky, M. J., Schmidt, E. L., and Kosnen, W. C. (1993). Plasmids pJP4 and R68.45 catransferred between populations of bradyrhizobia insterile soil.Appl. Environ. Microbiol.59, 1762–1766.
Leemans, J., Sobero´n, G., Cevallos, M. A., Ferna´ndez, L.Pardo, M. A., de la Vega, H., Flores, M., Quinto, C.,Palacios, R. (1984). General organization ofR. phaseonif plasmids. In: “Advances in Nitrogen Fixation Rsearch” (C. Veeger and W. E. Newton, Eds.), p. 7Nijhoff, Junk, and Pudoc, The Hague, The Netherla
Mavingui, P., Flores, M., Romero, D., Martı´nez-RomeroE., and Palacios, R. (1997). Generation ofRhizobiumstrains with improved symbiotic properties by randDNA amplification (RDA). Nat. Biotechnol.15, 564–569.
Miller, J. H. (1972). “Experiments in Molecular GeneticCold Spring Harbor Laboratory Press, Cold Spring Hbor, NY.
Moenne-Loccoz, Y., and Weaver, R. W. (1995). Plasmand saprophytic growth ofRhizobium leguminosarumbv.trifolii W14-2 in soil.FEMS Microbiol. Ecol.18, 139–144.oel, K. D., Sanchez, A., Ferna´ndez, L., Leemans, J., aCevallos, M. A. (1984).Rhizobium phaseolisymbioticmutants with transposon Tn5 insertions.J. Bacteriol.158,148–155.resnik, I. J., Pacarynuk, L. A., O’Brien, S. A. P., YoC. K., and Hynes, M. F. (1998). Plasmid-encoded cbolic genes inRhizobium leguminosarumbv. trifolii:Evidence for a plant-inducible rhamnose locus invoin competition for nodulation.Mol. Plant–Microbe Interact. 11, 1175–1185.
ardo, M. A., Lagu´nez, J., Miranda, J., and Martı´nez, E(1994). Nodulation ability ofRhizobium tropiciis condi-tioned by a plasmid-encoded citrate synthase.Mol. Mi-crobiol. 11, 315–321.uinto, C., de la Vega, H., Flores, M., Ferna´ndez, L.Ballado, T., Sobero´n, G., and Palacios, R. (1982). Reeration of nitrogen fixation gene sequences inRhizobiumphaseoli. Nature (London)299,724–728.omero, D., Martı´nez-Salazar, J., Girard, L., Brom,Davila, G., Palacios, R., Flores, M., and Rodrı´guez, C

the.
tiot.
lasr
andanil
andion
,on-i-
B.tide
.
m-
d
ofthati.
of-
s,nesre of
, D.
inJ.
Y 98).at
eins.
C
43RHIZOBIUM ETLI SYMBIOTIC PLASMID TRANSFER
(1995). Discrete amplifiable regions (amplicons) insymbiotic plasmid ofRhizobium etliCFN42.J. Bacteriol177,973–980.
Romero, D., and Palacios, R. (1997). Gene amplificaand genomic plasticity in prokaryotes.Annu. Rev. Gene31, 91–111.
Rosenberg, C., and Hughet, T. (1984). The pATC58 pmid of Agrobacterium tumefaciensis not essential fotumor induction.Mol. Gen. Genet.196,533–536.
Schofield, P. R., Gibson, A. H., Dudman, W. F.,Watson, J. M. (1987). Evidence for genetic exchangerecombination ofRhizobiumsymbiotic plasmids in a sopopulation.Appl. Environ. Microbiol.53, 2942–2947.
Scott, D. B., and Ronson, C. W. (1982). Identificationmobilization by cointegrate formation of a nodulatplasmid inRhizobium trifolii. J. Bacteriol.151,36–43.
Segovia, L., Pin˜ero, D., Palacios, R., and Martı´nez-RomeroE. (1991). Genetic structure of a soil population of nsymbioticRhizobium leguminosarum. Appl. Environ. Mcrobiol. 57, 426–433.
Siedow, A., Cramm, R., Siddiqui, R. A., and Friedrich,(1999). A megaplasmid-borne anaerobic ribonucleoreductase inAlcaligenes eutrophusH 16. J. Bacteriol181,4919–4928.
Simon, R. (1984). High frequency mobilization of gra
n
-
d
negative bacterial replicons by thein vitro constructeTn5-mob transposon.Mol. Gen. Genet.196,413–420.
Sullivan, J. T., and Ronson, C. W. (1998). Evolutionrhizobia by acquisition of a 500-kb symbiosis islandintegrates into a phe-tRNA gene.Proc. Natl. Acad. ScUSA95, 5145–5149.
Triplett, E. W., and Sadowski, M. J. (1992). Geneticscompetition for nodulation of legumes.Annu. Rev. Microbiol. 46, 399–428.
Troxler, J., Azelvandre, P., Zala, M., De´fago, G., and HaaD. (1997). Conjugative transfer of chromosomal gebetween fluorescent pseudomonads in the rhizosphewheat.Appl. Environ. Microbiol.63, 213–219.
Vinuesa, P., Reuhs, B. L., Breton, C., and Werner(1999). Identification of a plasmid-borne locus inRhizo-bium etliKIM5s involved in lipopolysaccharide O-chabiosynthesis and nodulation ofPhaseolus vulgaris.Bacteriol.181,5606–5614.
ost, C. K., Rochepeau, P., and Hynes, M. F. (19Rhizobium legumionsarumcontains a group of genes thappear to code for methyl-accepting chemotaxis protMicrobiology 144,1945–1956.
ommunicated by Donald Helinski




















