
Impacts of Argentine ants on mealybugs and their natural enemies in California’s coastal vineyards
14
Ecological Entomology (2007), 32, 583–596 DOI: 10.1111/j.1365-2311.2007.00910.x © 2007 The Authors Journal compilation © 2007 The Royal Entomological Society 583 Introduction The Argentine ant, Linepithema humile (Mayr) (Hymenoptera: Formicidae), is one of the world’s more damaging invasive in- sects (Suarez et al., 2001; Holway et al., 2002a). In California, it was first recorded in 1907 (Woodworth, 1908) and is now widespread in urban environments and in low-elevation areas, particularly those with disturbed habitats and some soil mois- ture (Human et al., 1998; Suarez et al., 2001). Familiar as a pest of the kitchen and picnic table, the Argentine ant’s more serious impacts are on the natural biota and in agricultural production. By acting as either predator or fierce competitor, Argentine ants can displace native ants and other invertebrates (Ward, 1987; Suarez et al., 1998, 2001; Holway, 1999; Sanders et al., 2001; Correspondence: Kent M. Daane, 137 Mulford Hall, Department of Environmental Science, Policy and Management, University of California, Berkeley, CA 94720-3114, U.S.A. E-mail: [email protected] Impacts of Argentine ants on mealybugs and their natural enemies in California’s coastal vineyards KENT M. DAANE 1 , KAREN R. SIME 1 , JULIE FALLON 2 and MONICA L. COOPER 1 1 Department of Environmental Science, Policy and Management, Center for Biological Control, University of California, Berkeley, California, U.S.A. and 2 University of California Cooperative Extension, San Luis Obispo County, California, U.S.A. Abstract. 1. The Argentine ant, Linepithema humile, tends honeydew-excreting homopterans and can disrupt the activity of their natural enemies. This mutualism is often cited for increases in homopteran densities; however, the ant’s impact on natural enemies may be only one of several effects of ant tending that alters insect densities. To test for the variable impacts of ants, mealybug and natural enemy densities were monitored on ant-tended and ant-excluded vines in two California vineyard regions. 2. Ant tending increased densities of the obscure mealybug, Pseudococcus viburni, and lowered densities of its encyrtid parasitoids Pseudaphycus flavidulus and Leptomastix epona. Differences in parasitoid recovery rates suggest that P. flavidulus was better able to forage on ant-tended vines than L. epona. 3. Densities of a coccinellid predator, Cryptolaemus montrouzieri, were higher on ant-tended vines, where there were more mealybugs. Together with behavioural observations, the results showed that this predator can forage in patches of ant-tended mealybugs, and that it effectively mimics mealybugs to avoid disturbance by ants. 4. Ant tending increased densities of the grape mealybug, Pseudococcus maritimus, by increasing the number of surviving first-instar mealybugs. Parasitoids were nearly absent from the vineyard infested with P. maritimus. Therefore, ants improved either mealybug habitat or fitness. 5. There was no difference in mealybug distribution or seasonal development patterns on ant-tended and ant-excluded vines, indicating that ants did not move mealybugs to better feeding locations or create a spatial refuge from natural enemies. 6. Results showed that while Argentine ants were clearly associated with increased mealybug densities, it is not a simple matter of disrupting natural enemies. Instead, ant tending includes benefits independent of the effect on natural enemies. Moreover, the effects on different natural enemy species varied, as some species thrive in the presence of ants. Key words. Ant–homopteran mutualism, biological control, invasive species, Leptomastix epona, Linepithema humile, Pseudaphycus flavidulus, Pseudococcus maritimus, Pseudococcus viburni, spatial refuge.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Impacts of Argentine ants on mealybugs and their natural enemies in California’s coastal vineyards

Ecological Entomology (2007), 32, 583–596 DOI: 10.1111/j.1365-2311.2007.00910.x
© 2007 The AuthorsJournal compilation © 2007 The Royal Entomological Society 583
Introduction
The Argentine ant, Linepithema humile (Mayr) (Hymenoptera: Formicidae), is one of the world’s more damaging invasive in-sects ( Suarez et al. , 2001; Holway et al. , 2002a ). In California,
it was first recorded in 1907 ( Woodworth, 1908 ) and is now widespread in urban environments and in low-elevation areas, particularly those with disturbed habitats and some soil mois-ture ( Human et al. , 1998; Suarez et al. , 2001 ). Familiar as a pest of the kitchen and picnic table, the Argentine ant’s more serious impacts are on the natural biota and in agricultural production. By acting as either predator or fierce competitor, Argentine ants can displace native ants and other invertebrates ( Ward, 1987; Suarez et al. , 1998 , 2001; Holway, 1999; Sanders et al. , 2001;
Correspondence: Kent M. Daane, 137 Mulford Hall, Department of Environmental Science, Policy and Manage ment, University of California, Berkeley, CA 94720-3114, U.S.A. E-mail: [email protected]
Impacts of Argentine ants on mealybugs and their natural enemies in California’s coastal vineyards
K E N T M . D A A N E 1 , K A R E N R . S I M E 1 , J U L I E FA L L O N 2 a n d M O N I C A L . C O O P E R 1 1 Department of Environmental Science, Policy and Management,
Center for Biological Control, University of California, Berkeley, California, U.S.A. and 2 University of
California Cooperative Extension, San Luis Obispo County, California, U.S.A .
Abstract . 1. The Argentine ant, Linepithema humile , tends honeydew-excreting homopterans and can disrupt the activity of their natural enemies. This mutualism is often cited for increases in homopteran densities; however, the ant’s impact on natural enemies may be only one of several effects of ant tending that alters insect densities. To test for the variable impacts of ants, mealybug and natural enemy densities were monitored on ant-tended and ant-excluded vines in two California vineyard regions.
2. Ant tending increased densities of the obscure mealybug, Pseudococcus viburni , and lowered densities of its encyrtid parasitoids Pseudaphycus flavidulus and Leptomastix epona . Differences in parasitoid recovery rates suggest that P. flavidulus was better able to forage on ant-tended vines than L. epona .
3. Densities of a coccinellid predator, Cryptolaemus montrouzieri , were higher on ant-tended vines, where there were more mealybugs. Together with behavioural observations, the results showed that this predator can forage in patches of ant-tended mealybugs, and that it effectively mimics mealybugs to avoid disturbance by ants.
4. Ant tending increased densities of the grape mealybug, Pseudococcus maritimus , by increasing the number of surviving first-instar mealybugs. Parasitoids were nearly absent from the vineyard infested with P. maritimus . Therefore, ants improved either mealybug habitat or fitness.
5. There was no difference in mealybug distribution or seasonal development patterns on ant-tended and ant-excluded vines, indicating that ants did not move mealybugs to better feeding locations or create a spatial refuge from natural enemies.
6. Results showed that while Argentine ants were clearly associated with increased mealybug densities, it is not a simple matter of disrupting natural enemies. Instead, ant tending includes benefits independent of the effect on natural enemies. Moreover, the effects on different natural enemy species varied, as some species thrive in the presence of ants.
Key words . Ant – homopteran mutualism , biological control , invasive species , Leptomastix epona , Linepithema humile , Pseudaphycus fl avidulus , Pseudococcus maritimus , Pseudococcus viburni , spatial refuge .

584 Kent M. Daane et al.
© 2007 The AuthorsJournal compilation © 2007 The Royal Entomological Society, Ecological Entomology, 32, 583–596
Holway et al. , 2002a ) and can disrupt vertebrate populations through changes to the food web ( Suarez et al. , 2000; Suarez & Case, 2002 ).
In agricultural and ornamental ecosystems, Argentine ants have long been associated with outbreaks of honeydew-producing homopterans ( Bartlett, 1961; Way, 1963; Buckley, 1987; Lach, 2003; Ness & Bronstein, 2004 ). The mutualistic association has clear benefits for the ants, which are provided with a carbohy-drate food source, and in return, ant tending has been credited with protecting homopterans from natural enemies ( Way, 1963 ). However, the disruption of natural enemies by ants may not solely account for observed changes in homopteran densities. For exam-ple, ant tending does not equally affect all natural enemy species, and some species flourish in ant-tended systems ( Völkl & Kroupa, 1997; Barzman & Daane, 2001 ). Ant tending also cleans the hab-itat of honeydew and tending ants have been observed moving insects to different feeding locations ( Flanders, 1951; Way, 1963 ), both of which may improve homopteran survival.
The invasion of Argentine ants into California vineyards has been implicated in outbreaks of mealybugs (Hemiptera: Pseudococcidae) in North Coast and Central Coast appellations ( Phillips & Sherk, 1991; Daane et al. , 2006 ). In this study, the first hypothesis tested was that Argentine ants increased mealy-bug population densities and fruit damage. Ant-tending and ant-exclusion treatments were established in a vineyard located in the North Coast region that was infested with the grape mealy-bug, Pseudococcus maritimus (Ehrhorn), and two vineyards in the Central Coast region that were infested with the obscure mealybug, Pseudococcus viburni (Signoret). These locations al-lowed the impacts of Argentine ants to be followed with differ-ent mealybug species. To separate treatment effects, ant and mealybug densities were measured, as well as fruit damage. The second hypothesis tested was that ant tending influences mealy-bug populations through multiple mechanisms, including va-rying effects on different natural enemy species. To separate treatment effects, mealybug percentage parasitism was meas-ured separately for each parasitoid species, as was the abun-dance of mealybug predator species. Additionally, a laboratory experiment was conducted to compare the effectiveness of two parasitoid species in the presence or absence of foraging ants. The third hypothesis tested concerned whether the foraging pat-terns of ants created a spatial refuge for the mealybugs. Reeve and Murdoch (1986) observed that Argentine ants foraged mainly on the inner scaffolding of citrus trees, and suggested that this helped create a spatial refuge for California red scale, Aonidiella aurantii (Maskell), from parasitoids. Argentine ant foraging patterns on vines are similar and it was hypothesised that there would be less parasitism on the vine trunk and spurs, where there is more ant activity, compared with the leaves and canes, where there is less. To test for such a spatial refuge, per-centage parasitism of mealybugs on different vine sections was measured. Finally, population-level effects of ant tending were also investigated by measuring treatment differences in mealy-bug seasonal age structure, to determine if ant tending influ-enced mealybug development, and by analysing mealybug distribution on the vines, to determine if ants moved mealybugs to better feeding locations. Brief descriptions of the mealybug species and their natural enemies studied are first presented.
Mealybug species and damage
The grape mealybug and obscure mealybug cause economic damage by accumulating in fruit clusters, secreting honeydew that fouls the fruit and promotes fungal pathogens, and transmit-ting closteroviruses ( Flaherty et al. , 1992; Golino et al. , 2002 ). Mealybug populations can be suppressed with insecticides; however, there has been increasing use of sustainable farming practices in Californian vineyards and many of the insecticides used for mealybug control are incompatible with these practices ( Daane et al. , 2005 ). Although closely related, the two mealy-bug species differ in their origin, seasonal biology, and natural enemy complex.
The grape mealybug is native to North America ( Ben-Dov, 1995 ). In California vineyards, it has two generations per year, with populations overwintering as eggs or small immature stages located under the bark of the trunk or spurs ( Geiger & Daane, 2001 ). For most of the season, a majority of the grape mealybug population remains under the bark, but near harvest-time, mealybugs from the second generation move into the fruit clusters ( Geiger et al. , 2001 ). Although it been reported in California since the early 1900s ( Essig, 1914 ), a complex of natural enemies has typically provided acceptable control ( Clausen, 1924; Flaherty et al. , 1992 ). In fact, Flaherty et al. (1982) at-tributed occasional grape mealybug outbreaks to the disruption of natural enemies by synthetic insecticides. In recent years, ants may have replaced insecticides as the primary disruptive mechanism ( Phillips & Sherk, 1991; Daane et al. , 2006 ).
The California history of the obscure mealybug is less clear, in part from earlier taxonomic confusion – there are at least seven synonyms ( Ben-Dov, 1995 ). The obscure mealybug is not native to North America ( Miller et al. , 1984 ) and was not con-sidered a primary vineyard pest until the 1980s, when the grape industry expanded in the Central Coast appellations. Most likely it first moved from ornamental plants onto vines in areas where vineyards abutted residential landscaping. There are no reported studies of obscure mealybug population dynamics in California vineyards. Observations suggest it has three generations per year, with overlapping generations near harvest-time (K. M. Daane, unpublished data). Unlike the grape mealybug, the ob-scure mealybug readily feeds on grape leaves, exposing a large portion of the population to natural enemies. Although it is closely related to the grape mealybug, the two species have few natural enemy species in common ( Noyes & Hayat, 1994 ).
Natural enemy species
Several natural enemy species of the grape mealybug were found in the North Coast region before the study commen-ced. These include Acerophagous notativentris (Girault) and Pseudaphycus angelicus (Howard) (Hymenoptera: Encyrtidae) as the dominant parasitoid species, and the green lacewing Chrysoperla carnea Stephens (Neuroptera: Chrysopidae) ( Daane et al. , 1996 ). Additionally, ≈ 2200 adult mealybug destroyers, Cryptolaemus montrouzieri Mulsant (Coleoptera: Coccinellidae), which originated from material collected in southern Australia (K. S. Hagen, unpublished data), were released from April 1996

Argentine ants and vineyard insects 585
© 2007 The AuthorsJournal compilation © 2007 The Royal Entomological Society, Ecological Entomology, 32, 583–596
to August 1997. These predators were observed in the study site prior to treatment establishment. Because an effective natural en-emy complex was resident in the North Coast region, it was as-sumed that their densities did not need to be manipulated, only that exclusion treatments need be implemented.
In the Central Coast region, predators found before the study commenced were an unidentified Hyperaspis Redtenbacher (Coleoptera: Coccinellidae), the green lacewings C. carnea , C. comanche (Banks), and an unidentified Chrysopa Leach, and the brown lacewing Hemerobius pacificus Banks (Neuroptera: Hemerobiidae) ( Daane et al. , 1996 ). Because these resident nat-ural enemies did not sufficiently suppress the exotic obscure mealybug, an importation programme was started in 1996. Ap-proximately 1500 mealybug destroyers of the aforementioned southern Australia strain were released before the experiment commenced, from May 1996 to September 1997. To establish obscure mealybug parasitoids, Pseudaphycus flavidulus (Brethes) and Leptomastix epona (Walker) (Hymenoptera: Encyrtidae) were imported from insectary colonies maintained at Centro National de Entomología, La Cruz, Chile. From June 1997 to May 2000, ≈ 194 000 P. flavidulus and ≈ 5500 L. epona were released, including ≈ 73 600 P. flavidulus and ≈ 5000 L. epona in the two vineyards used in the current study during the experimental period. No parasitoids were released directly on vines in the experimental plots.
Table 1 summarises the mealybug and natural enemy species that were either resident to the region or augmented in or near vineyards containing the experimental plots.
Methods and materials
Field sites
Ant exclusion trials were conducted at two vineyards in the Central Coast region (Edna Valley appellation, San Luis Obispo County) and one vineyard in the North Coast region (Carneros appellation, Sonoma County). The Central Coast vineyards were an 8- and 16-year-old Chardonnay cultivar, and the North Coast vineyard was a 12-year-old Pinot Noir cultivar. All vineyards used drip-line irrigation systems, and were clean-cultivated by running a disc over the row middles and applying a herbicide (glyphosate) to the berms. During the 2-year sam-pling period, no insecticides were used.
Ant-exclusion and ant-tending treatments were initiated in July 1998 at the North Coast site and April 1999 at the Central Coast sites. At each site, treatments were set in a randomised
block design, with six replicates. Each replicate consisted of five adjacent vines, with 5 – 10 vines between ant-tended and ant-excluded treatments in each block, and 1 – 3 rows between blocks. To exclude ants, the bark was stripped from the basal 10 – 15 cm of each trunk and the exposed wood was wrapped with duct tape, which was then coated with Tanglefoot Pest Barrier (Tanglefoot, Grand Rapids, Michigan). Vines adjacent to each replicate had the canes and leaves removed in a 30 – 60 cm section, and the exposed trellis posts, wires, and irrigation lines were coated with a 5 – 10 cm barrier of Tanglefoot. The bar-riers were checked every 1 – 2 weeks and Tanglefoot was reap-plied as needed.
Insect sampling
In July (North Coast) and September (Central Coast) 1998, a pre-treatment survey was conducted to determine levels of mealybug infestation. In each of 10 rows, 8 – 12 vines were selected randomly and one cluster per vine was searched for mealybugs ( n = 100). The clusters selected were in contact with the cordon because these have higher mealybug densities ( Geiger et al. , 2001 ). The presence or absence of mealybugs was recorded for each cluster, and mealybug species were iden-tified using field diagnostic characters ( Godfrey et al. , 2002 ).
After the ant-exclusion treatments were applied, the densities of ant, mealybug, and natural enemy populations were estimated every 2 weeks from May through September and every 3 – 5 weeks from October through April. On each sample date, one vine per replicate was selected randomly and the number of ants moving up or down a selected band of the trunk or cordon was recorded during a 2-min period. Mealybug and natural enemy densities were then determined on four different vine sections: (i) a 150 cm 2 section of the upper trunk or cordon, (ii) three spurs located at the first, third, and fifth positions from the basal cordon, (iii) the seven basal leaves on a cane originating from each sampled spur (21 leaves per vine), and (iv) the three basal clusters on each sampled spur (nine clusters per vine). This was an estimated 15 – 25% of each vine section. For each sample, mealybug density was recorded in four development stage cat-egories: crawlers and first instars, second and third instars, adult females, and ovisacs. The mealybug condition was recorded as live, mummified (no exit hole), or parasitised (mummified with an exit hole). All mummified mealybugs were collected, placed individually in gelatine capsules, and held for parasitoid emer-gence at 23 ± 2 °C. Predators (green lacewing eggs and larvae and beetle larvae and adults) were counted in the field.
Table 1. Mealybug species and resident and released natural enemies at study locations.
Study site Year Mealybug species Resident parasitoids Augmented parasitoids Resident predators
North Coast (Vineyard A) 1998, 1999 P. maritimus P. angelicus , A. notativentris
None C. montrouzieri , C. carnea
Central Coast (Vineyards A and B) 1999 P. viburni None P. fl avidulus , L. epona C. montrouzieri , C. carnea , C. comanche , H. pacifi cus

586 Kent M. Daane et al.
© 2007 The AuthorsJournal compilation © 2007 The Royal Entomological Society, Ecological Entomology, 32, 583–596
Parasitoid success in the presence of foraging ants
Adult parasitoids were often observed being attacked by for-aging ants, a phenomenon that has been described in other systems ( Heimpel et al. , 1997; Völkl & Kroupa, 1997 ). These observations of released parasitoids and different recovery rates of P. flavidulus and L. epona led us to suspect that these species differed in their ability to reproduce successfully when ants were present. To test this hypothesis, laboratory trials were con-ducted to measure the parasitoids’ oviposition success in the presence and absence of Argentine ants. Ant colonies were de-rived from broods collected in North Coast vineyards. The labo-ratory colonies were housed in 30.2 litre plastic tubs filled with loose dirt for nest substrate and ringed with Tanglefoot to pre-vent escape. The colony was provisioned with navel orange-worm pupae, Amyelois transitella (Walker) (Lepidoptera: Pyralidae), for protein, and a dilute honey solution (1:3 honey and water) for carbohydrates. To reach the food, the ants were trained to forage through a plastic tube (1 m long × 1.27 cm di-ameter). By repositioning the tube, ants were directed into the test arena, which was a wood-sided sleeve cage (45 × 45 × 45 cm). Inside the cage, potatoes infested with ≈ 100 second- and third-instar obscure mealybugs were set on raised platforms that were either covered with Tanglefoot (ant-excluded treatment) or clean (ant-tended treatment). After the ants began foraging, L. epona and P. flavidulus were added (40 females and 15 males for each species). The cages were checked periodically thereafter to record ant and parasitoid interactions. After all parasitoids were dead (2 – 3 weeks), potatoes were isolated for 4 weeks and the emerged parasitoids were recorded by species. There were four replicates.
Statistics
Results are presented as means per treatment ( ± SE). Season-long treatment effects were analysed using repeated measures analysis of variance (anova); the statistical output is provided in the figure captions. Average treatment effects are compared by t -tests, with sample date and block used to pair measured pa-rameters. To estimate the reproductive potential, the per capita increase, the number of surviving immature mealybugs was di-vided by the number of ovisacs on each vine, using sample dates that correspond to the period of egg hatch. Percentage parasit-ism was determined using all mummified mealybugs, which is less conservative than rearing parasitoids from live mealybugs. Fruit damage was assessed by comparing the number of mealy-bugs in clusters in August and September samples (harvest-time). Treatment effects on mealybug distribution on the vine (live or parasitised) were determined using a general linear model for each paired comparison. The model used the per-centage mealybugs per vine section of the total per vine per sample date, as a function of treatment, sample date, and treatment × sample date interaction. Sample date was set as a categorical variable to exclude its impact on treatment. If the treatment × sample date interaction term is not significant ( P > 0.05), this analysis is equivalent to an analysis of covari-ance with sample date as the covariate. In this case a significant treatment effect indicates that ant presence alters the percentage
mealybugs on that vine section. A significant ( P < 0.05) interac-tion term indicates that per cent mealybug distribution, as a function of sample date, is significantly different by treatment. That is, the ant treatment altered the seasonal per cent mealybug distribution on that vine section. Similar analyses were con-ducted to determine treatment impact on mealybug develop-ment stages. For all analyses, data were transformed [log( x + 1)] as needed to stabilise the variance.
Results
Ant exclusion fi eld studies – Central Coast vineyards
In the pre-treatment survey, 76.2 ± 4.3 and 59.4 ± 4.9% of the fruit clusters in Central Coast vineyards were infested with obscure mealybug. The between-vineyard difference was significant ( t 198 = – 2.590, P = 0.010). Similarly, after the exclusion treatments were imposed, the two vineyards had different season-long mealybug densities when pairing similar treatments (repeated measures anova: ant-tended: F 1,9 = 10.549, P = 0.010; ant-excluded: F 1,9 = 6.913, P = 0.027). Analyses of mealybug and parasitoid densities were, therefore, conducted separately for each vineyard.
Season-long ant densities were higher in the ant-tended than the ant-excluded treatment ( Figs 1a and 2a ). In both vineyards, there was a mid-season dip in ant activity (ant-tended vines). Average in-season (June – October) ant foraging activity was quite high, greater than 30 ants per vine per 2 min count on ant-tended vines, while there was either no measured ant activity (Vineyard A) or ant activity was extremely low (Vineyard B) on ant-excluded vines ( Table 2 ).
Season-long mealybug densities were higher in the ant-tended than the ant-excluded treatment ( Figs 1b and 2b ). In Vineyard A, mealybug density in the ant-tended treatment steadily rose to a peak in August, when crawlers from the second generation emerged from the ovisacs. In Vineyard B, the two peaks in mealybug density correspond to crawler emergence in the first and second generations. Average in-season mealybug densities (June – October) were greater on ant-tended than ant-excluded vines, and there were 82.2-fold and 3.7-fold more infested fruit clusters in the ant-tended treatment in Vineyards A and B respec-tively ( Table 2 ). The per capita population increase (surviving immatures per ovisac) did not differ between treatments for either vineyard ( Table 2 ).
Per cent parasitism was greater on vines in the ant-exclusion treatment ( Figs 1c and 2c ). In Vineyard A, where ant exclusion was complete, overall percentage parasitism in the ant-excluded treatment rose quickly in late July, with an average in-season (June – October) level of 67.7 ± 4.7% compared with only 10.3 ± 1.6% in the ant-tended vines ( Table 2 ). Pseudaphycus flavidulus comprised 59.2% of the parasitoids reared in the ant-exclusion treatment ( n = 113) and 100% in the ant-tended treatment ( n = 7). Parasitism levels rose more slowly in the ant-exclusion treatment in Vineyard B, where ant exclusion was incomplete, reaching a maximum of only 28.7 ± 7.1%. Still, average in-season parasitism was higher in the ant-excluded treatment. Pseudaphycus flavidulus comprised 82.3% of the parasitoids reared in the ant-exclusion treatment ( n = 34) and 100% of the

Argentine ants and vineyard insects 587
© 2007 The AuthorsJournal compilation © 2007 The Royal Entomological Society, Ecological Entomology, 32, 583–596
parasitoids reared in the ant-tended treatment ( n = 5). Data from both vineyards combined showed that P. flavidulus was more common on the ant-tended vines than L. epona , compared with their ratio in the ant-excluded treatment ( � 2 = 7.136, P = 0.005).
The average in-season density of the mealybug destroyer was higher in ant-tended than ant-excluded treatments in both vineyards ( Table 2 ). There was no between-treatment difference in the number of chrysopids. The larvae found were identified as C. carnea ( n = 9); data include 24 chrysopid eggs that were not identified to
Fig. 1. Season-long average ( ± SE) sample counts in ant-tended ( • ) and ant-excluded ( ○ ) treatments for (a) Argentine ants, (b) obscure mealybugs, and (c) percentage parasitism of obscure mealybug in Central Coast Vineyard A . Treatment impact was signifi cantly different for each parameter measured (repeated measures anova for ants: F 1,9 = 43.42, P < 0.001; mealybug: F 1,9 = 159.2, P < 0.001; percentage parasitism: F 1,9 = 34.84, P < 0.001).

588 Kent M. Daane et al.
© 2007 The AuthorsJournal compilation © 2007 The Royal Entomological Society, Ecological Entomology, 32, 583–596
species. Adult Chrysoperla sp. and Hemerobius sp. were also ob-served but were not assigned to a treatment because the sampling methods often resulted in their flight between treatment vines. All predator groups were at low densities (<0.4 per vine per sample).
Analyses of the live and parasitised mealybug distribution on the vine used data combined from both vineyards as distribution patterns were similar between vineyards. There was a signifi-cantly smaller proportion of the live mealybug population on
Fig. 2. Season-long average ( ± SE) sample counts in ant-tended ( • ) and ant-excluded ( ○ ) treatments for (a) Argentine ants, (b) obscure mealybugs, and (c) percentage parasitism of obscure mealybug in Central Coast Vineyard B . Treatment impact was signifi cantly different for each parameter measured (repeated mea-sures anova for ants: F 1,10 = 12.34, P = 0.006; mealybug: F 1,10 = 8.91, P = 0.017; per-centage parasitism: F 1,10 = 8.690, P = 0.015).

Argentine ants and vineyard insects 589
© 2007 The AuthorsJournal compilation © 2007 The Royal Entomological Society, Ecological Entomology, 32, 583–596
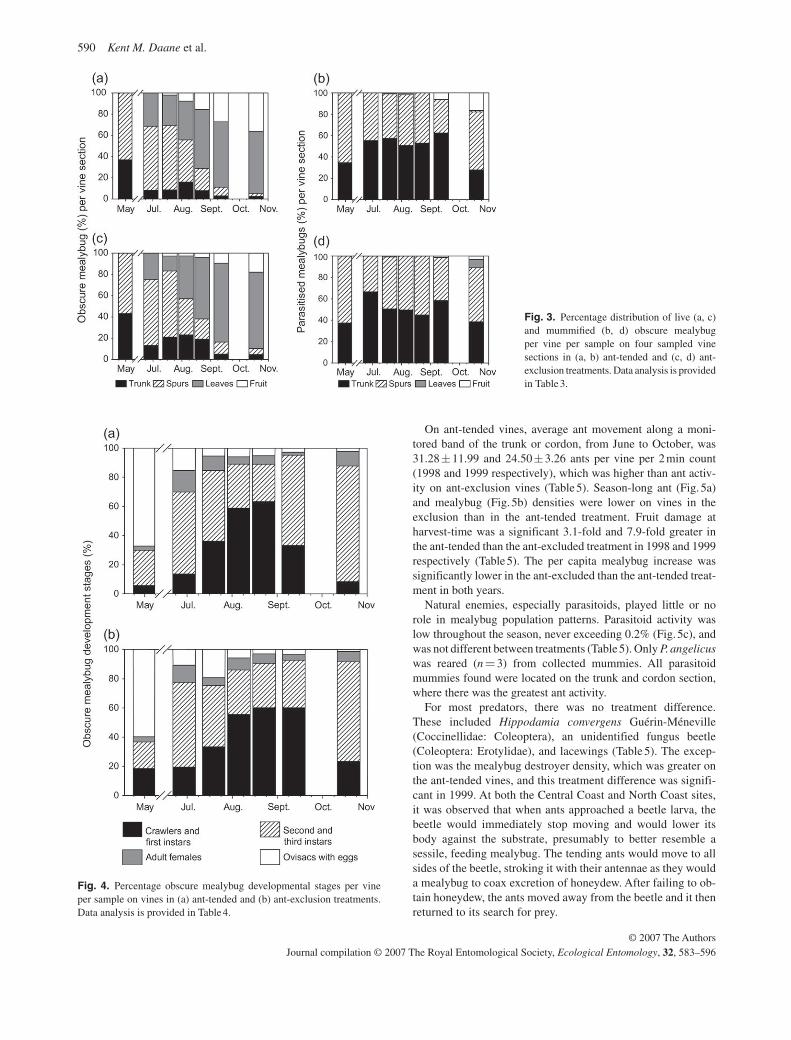
the trunk and more in the fruit clusters on the ant-tended vines ( Table 3 , Fig. 3a,c ). In all locations, sample date influenced mealybug distribution, reflecting mealybug movement from overwintering locations on the trunk and spurs to summer feed-ing locations on the leaves on fruit. Mummified mealybugs were primarily found on the trunk and spur sections ( Fig. 3b,d ). A greater proportion of the mummified mealybugs were found in fruit clusters in the ant-tended treatment ( Table 3 ), although this difference may be unreliable because of the low numbers of mealybugs found in the ant-tended treatment near harvest-time.
Obscure mealybug development, as estimated by the percent-age composition of different life stages in the population, was
relatively similar between treatments ( Fig. 4 ). There was a smaller percentage of crawlers and first instars ( P = 0.079) and more second and third instars ( P = 0.049) in the ant-tended than ant-excluded treatment ( Table 4 ).
Ant exclusion fi eld studies – North Coast vineyards
The pre-treatment survey showed that 81.7 ± 3.8% of the grape clusters were infested, all with the grape mealybug. Results of treatment impact are provided separately for 1998 and 1999.
Table 2. Average ( ± SE) density per vine per sample of Argentine ants, obscure mealybugs (MB), mealybug destroyer, green lacewings, and grape cluster infestation on vines in ant-exclusion and ant-tended treatments in two vineyards studied in the Central Coast wine grape region.
Vineyard Measured parameter Ant-tended Ant-excluded Paired t -test
Vineyard A Argentine ant * 33.8 ± 7.4 0 t 29 = 4.463, P < 0.001 Obscure MB † 303.2 ± 45.8 11.8 ± 4.1 t 33 = 6.430, P < 0.001 Parasitism (%) † 10.3 ± 1.6 67.7 ± 4.7 t 33 = – 12.41, P < 0.001 MB destroyer † 0.31 ± 0.09 0.07 ± 0.04 t 41 = 2.125, P = 0.040 Chrysopidae † 0.12 ± 0.07 0.21 ± 0.10 t 29 = – 0.783, P = 0.436 Cluster infestation ‡ 123.1 ± 28.8 1.5 ± 0.7 t 17 = 4.224, P = 0.001 Per capita change § 71.4 ± 18.2 62.2 ± 24.0 t 7 = 0.301, P = 0.772
Vineyard B Argentine ant * 38.6 ± 7.0 0.4 ± 0.3 t 29 = 5.476, P < 0.001 Obscure MB † 129.2 ± 20.5 59.2 ± 11.7 t 35 = 3.010, P = 0.005 Parasitism (%) † 11.0 ± 1.7 21.3 ± 3.3 t 35 = – 2.767, P = 0.009 MB destroyer † 0.29 ± 0.09 0.09 ± 0.05 t 41 = 2.033, P = 0.049 Chrysopidae † 0.14 ± 0.06 0.24 ± 0.09 t 29 = – 0.828, P = 0.410 Cluster infestation ‡ 53.6 ± 19.3 14.3 ± 8.7 t 17 = 1.799, P = 0.090
Per capita change § 19.9 ± 3.5 21.6 ± 8.3 t 10 = – 0.182, P = 0.859
* Ants moving up or down a selected band of the trunk or cordon during a 2-min period. † Insects found during a destructive subsample of each vine on four different plant parts: (i) a 150 cm 2 section of the upper trunk or cordon, (ii) three spurs, (iii) the seven basal leaves on a cane originating from each sampled spur, and (iv) the three basal clusters on each sampled spur. ‡ Mealybugs found in sampled clusters from August to September (harvest-time). § Number of surviving immature mealybugs per ovisac per vine, compared during periods of peak mealybug density when there was egg hatch and crawlers settling to feed.
Table 3. Relationship of treatment (ant-tending vs ant-exclusion) and sample date on the distribution of live and parasitised obscure mealybugs on four mea-sured vine sections, as a percentage of the total mealybug population per vine (see Fig. 3 ), for data collected in two Central Coast vineyards in 1999.
Vine section Source of variation
Live mealybugs Mummifi ed mealybugs
F -ratio P -value F -ratio P -value
Trunk/Cordon * Treatment 5.637 0.019 0.183 0.892 Sample date 11.885 <0.001 2.954 0.010 Treatment × Date 0.308 0.932 0.408 0.873
Spur * Treatment 0.029 0.866 0.066 0.798 Sample date 37.151 0.000 1.952 0.077 Treatment × Date 0.230 0.966 0.358 0.904
Leaves/Cane * Treatment 0.192 0.662 1.093 0.297 Sample date 31.261 <0.001 3.556 0.003 Treatment × Date 1.216 0.301 2.019 0.067
Grape clusters † Treatment 14.521 <0.001 4.948 0.029 Sample date 7.622 <0.001 4.859 0.004
Treatment × Date 0.761 0.519 2.147 0.101
* Degrees of freedom are 1,6,6,138. † Degrees of freedom are 1,3,3,82 as data were not collected before grape clusters formed.
590 Kent M. Daane et al.
© 2007 The AuthorsJournal compilation © 2007 The Royal Entomological Society, Ecological Entomology, 32, 583–596
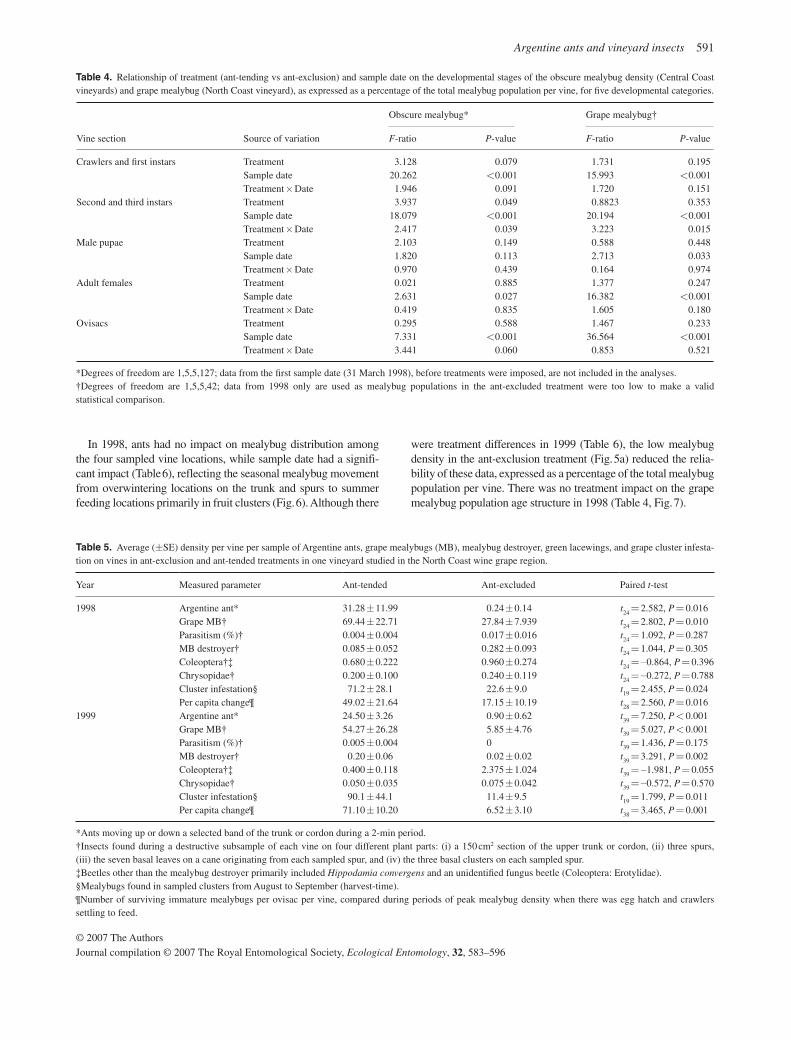
On ant-tended vines, average ant movement along a moni-tored band of the trunk or cordon, from June to October, was 31.28 ± 11.99 and 24.50 ± 3.26 ants per vine per 2 min count (1998 and 1999 respectively), which was higher than ant activ-ity on ant-exclusion vines ( Table 5 ). Season-long ant ( Fig. 5a ) and mealybug ( Fig. 5b ) densities were lower on vines in the exclusion than in the ant-tended treatment. Fruit damage at harvest-time was a significant 3.1-fold and 7.9-fold greater in the ant-tended than the ant-excluded treatment in 1998 and 1999 respectively ( Table 5 ). The per capita mealybug increase was significantly lower in the ant-excluded than the ant-tended treat-ment in both years.
Natural enemies, especially parasitoids, played little or no role in mealybug population patterns. Parasitoid activity was low throughout the season, never exceeding 0.2% ( Fig. 5c ), and was not different between treatments ( Table 5 ). Only P. angelicus was reared ( n = 3) from collected mummies. All parasitoid mummies found were located on the trunk and cordon section, where there was the greatest ant activity.
For most predators, there was no treatment difference. These included Hippodamia convergens Guérin-Méneville (Coccinellidae: Coleoptera), an unidentified fungus beetle (Coleoptera: Erotylidae), and lacewings ( Table 5 ). The excep-tion was the mealybug destroyer density, which was greater on the ant-tended vines, and this treatment difference was signifi-cant in 1999. At both the Central Coast and North Coast sites, it was observed that when ants approached a beetle larva, the beetle would immediately stop moving and would lower its body against the substrate, presumably to better resemble a sessile, feeding mealybug. The tending ants would move to all sides of the beetle, stroking it with their antennae as they would a mealybug to coax excretion of honeydew. After failing to ob-tain honeydew, the ants moved away from the beetle and it then returned to its search for prey.
Fig. 3. Percentage distribution of live (a, c) and mummifi ed (b, d) obscure mealybug per vine per sample on four sampled vine sections in (a, b) ant-tended and (c, d) ant-exclusion treatments. Data analysis is provided in Table 3 .
Fig. 4. Percentage obscure mealybug developmental stages per vine per sample on vines in (a) ant-tended and (b) ant-exclusion treatments. Data analysis is provided in Table 4 .
Argentine ants and vineyard insects 591
© 2007 The AuthorsJournal compilation © 2007 The Royal Entomological Society, Ecological Entomology, 32, 583–596
In 1998, ants had no impact on mealybug distribution among the four sampled vine locations, while sample date had a signifi-cant impact ( Table 6 ), reflecting the seasonal mealybug movement from overwintering locations on the trunk and spurs to summer feeding locations primarily in fruit clusters ( Fig. 6 ). Although there
were treatment differences in 1999 ( Table 6 ), the low mealybug density in the ant-exclusion treatment ( Fig. 5a ) reduced the relia-bility of these data, expressed as a percentage of the total mealybug population per vine. There was no treatment impact on the grape mealybug population age structure in 1998 ( Table 4 , Fig. 7 ).
Table 4. Relationship of treatment (ant-tending vs ant-exclusion) and sample date on the developmental stages of the obscure mealybug density (Central Coast vineyards) and grape mealybug (North Coast vineyard), as expressed as a percentage of the total mealybug population per vine, for fi ve developmental categories.
Vine section Source of variation
Obscure mealybug * Grape mealybug †
F -ratio P -value F -ratio P -value
Crawlers and fi rst instars Treatment 3.128 0.079 1.731 0.195 Sample date 20.262 <0.001 15.993 <0.001 Treatment × Date 1.946 0.091 1.720 0.151
Second and third instars Treatment 3.937 0.049 0.8823 0.353 Sample date 18.079 <0.001 20.194 <0.001 Treatment × Date 2.417 0.039 3.223 0.015
Male pupae Treatment 2.103 0.149 0.588 0.448 Sample date 1.820 0.113 2.713 0.033 Treatment × Date 0.970 0.439 0.164 0.974
Adult females Treatment 0.021 0.885 1.377 0.247 Sample date 2.631 0.027 16.382 <0.001 Treatment × Date 0.419 0.835 1.605 0.180
Ovisacs Treatment 0.295 0.588 1.467 0.233 Sample date 7.331 <0.001 36.564 <0.001
Treatment × Date 3.441 0.060 0.853 0.521
* Degrees of freedom are 1,5,5,127; data from the fi rst sample date (31 March 1998), before treatments were imposed, are not included in the analyses. † Degrees of freedom are 1,5,5,42; data from 1998 only are used as mealybug populations in the ant-excluded treatment were too low to make a valid statistical comparison.
Table 5. Average ( ± SE) density per vine per sample of Argentine ants, grape mealybugs (MB), mealybug destroyer, green lacewings, and grape cluster infesta-tion on vines in ant-exclusion and ant-tended treatments in one vineyard studied in the North Coast wine grape region.
Year Measured parameter Ant-tended Ant-excluded Paired t -test
1998 Argentine ant * 31.28 ± 11.99 0.24 ± 0.14 t 24 = 2.582, P = 0.016 Grape MB † 69.44 ± 22.71 27.84 ± 7.939 t 24 = 2.802, P = 0.010 Parasitism (%) † 0.004 ± 0.004 0.017 ± 0.016 t 24 = 1.092, P = 0.287 MB destroyer † 0.085 ± 0.052 0.282 ± 0.093 t 24 = 1.044, P = 0.305 Coleoptera † ‡ 0.680 ± 0.222 0.960 ± 0.274 t 24 = – 0.864, P = 0.396 Chrysopidae † 0.200 ± 0.100 0.240 ± 0.119 t 24 = – 0.272, P = 0.788 Cluster infestation § 71.2 ± 28.1 22.6 ± 9.0 t 19 = 2.455, P = 0.024 Per capita change ¶ 49.02 ± 21.64 17.15 ± 10.19 t 28 = 2.560, P = 0.016
1999 Argentine ant * 24.50 ± 3.26 0.90 ± 0.62 t 39 = 7.250, P < 0.001 Grape MB † 54.27 ± 26.28 5.85 ± 4.76 t 39 = 5.027, P < 0.001 Parasitism (%) † 0.005 ± 0.004 0 t 39 = 1.436, P = 0.175 MB destroyer † 0.20 ± 0.06 0.02 ± 0.02 t 39 = 3.291, P = 0.002 Coleoptera † ‡ 0.400 ± 0.118 2.375 ± 1.024 t 39 = – 1.981, P = 0.055 Chrysopidae † 0.050 ± 0.035 0.075 ± 0.042 t 39 = – 0.572, P = 0.570 Cluster infestation § 90.1 ± 44.1 11.4 ± 9.5 t 19 = 1.799, P = 0.011
Per capita change ¶ 71.10 ± 10.20 6.52 ± 3.10 t 38 = 3.465, P = 0.001
* Ants moving up or down a selected band of the trunk or cordon during a 2-min period. † Insects found during a destructive subsample of each vine on four different plant parts: (i) a 150 cm 2 section of the upper trunk or cordon, (ii) three spurs, (iii) the seven basal leaves on a cane originating from each sampled spur, and (iv) the three basal clusters on each sampled spur. ‡ Beetles other than the mealybug destroyer primarily included Hippodamia convergens and an unidentifi ed fungus beetle (Coleoptera: Erotylidae). § Mealybugs found in sampled clusters from August to September (harvest-time). ¶ Number of surviving immature mealybugs per ovisac per vine, compared during periods of peak mealybug density when there was egg hatch and crawlers settling to feed.

592 Kent M. Daane et al.
© 2007 The AuthorsJournal compilation © 2007 The Royal Entomological Society, Ecological Entomology, 32, 583–596
Parasitoid success in the presence of foraging ants
The ant-exclusion treatment produced both L. epona and P. flavidulus , while the ant-foraging treatment produced only P. flavidulus . There were no significant treatment differences, a result of the low percentage parasitism (<2% overall), which was a result of adult parasitoid mortality. The adults spent most of their time on the side and bottom of the cage, where many were killed by ants foraging inside the test arena. While the quantitative aspects of this experiment failed, the observations of ant – parasitoid interactions were more revealing. In the pres-ence of ants, L. epona females often appeared unaware of the
danger, moving slowly while handling hosts, and were often killed by ants. In contrast, P. flavidulus females moved quickly away from foraging ants and often oviposited successfully while ants tended mealybugs nearby.
Discussion
In this study, the relationship among Argentine ants, mealybugs, and various natural enemy species in California vineyards was investigated. Higher mealybug densities and fruit infestation levels were found on ant-tended than on ant-excluded vines. It was expected that the exclusion of ants and associated decrease in mealybug densities would be directly related to an increase in natural enemy numbers. Instead, the presence of ants aided mealybug populations through multiple mechanisms, and effects on natural enemy species varied.
The most often described scenario in ant – homopteran mu-tualisms is that ants tend phloem-feeders, collecting the ho-neydew and, in return, protecting them from natural enemies ( Bartlett, 1961; Way, 1963; Buckley, 1987; Buckley & Gullan, 1991; Stechmann et al. , 1996; Styrsky & Eubanks, 2007 ). In this study, there was clear evidence of this interaction at the Central Coast sites, where higher parasitism levels in the ant-excluded vines were associated with lower mealybug densities. Also observed was direct ant interference with parasitoid ovipo-sition, both in the laboratory and in the field. This was the ex-pected outcome as researchers have previously shown that ants can interfere with the biological control of various mealybug species ( Swirski et al. , 1980; Cudjoe et al. , 1993; González-Hernández et al. , 1999 ).
Most often, descriptions of ant interference with biological controls focus on key natural enemy species or do not differen-tiate the impacts of ants on specific natural enemy species (but see James et al. , 1999 ). Results from the Central Coast sites showed that P. flavidulus was more effective than L. epona when ants were present. This finding was corroborated by laboratory observations indicating that L. epona adults were more suscep-tible to attack by ants. Direct interference by ants with adult parasitoids was probably the primary cause of lowered parasit-ism rates, as has been reported for aphids ( Völkl & Kroupa, 1997 ). The biologies of P. flavidulus and L. epona have been compared in the laboratory ( Karamaouna & Copland, 2000 ), but not in the presence of ants. Both species tend to parasitise older mealybugs, but only L. epona host-feeds, which may pro-vide at least a partial explanation for the greater effectiveness of P. flavidulus . Many insect parasitoids require more time to host feed than to oviposit ( Heimpel & Collier, 1996 ), and species with shorter host-handling times tend to be more successful in the presence of foraging Argentine ants ( Barzman & Daane, 2001 ). Here, observations also suggest that P. flavidulus females respond more quickly than L. epona females to the approach of ants. This evasive response from P. flavidulus may be an evolved trait as adult parasitoid mortality inflicted by ants can result in significantly lowered lifetime reproductive potential ( Heimpel et al. , 1997 ). There are examples of parasitoids foraging suc-cessfully among ants, some species even seeking out ant-tended patches ( Völkl & MacKauer, 1993; Völkl, 1994 ).
Fig. 5. Season-long average ( ± SEM) sample counts in ant-tended ( • ) and ant-excluded ( ○ ) treatments for (a) Argentine ants, (b) grape mealy-bugs, and (c) percentage parasitism of grape mealybug in North Coast vineyard. Treatment impact was signifi cantly different for each para-meter measured (repeated measures anova for ants: F 1,8 = 50.39, P < 0.001; mealybug: F 1,8 = 32.67, P < 0.001; percentage parasitism: F 1,9 = 34.84, P < 0.001).

Argentine ants and vineyard insects 593
© 2007 The AuthorsJournal compilation © 2007 The Royal Entomological Society, Ecological Entomology, 32, 583–596
There are fewer documented examples of changes in predator effectiveness in the presence of foraging ants. Haney et al. (1987) reported an increase in the densities of citrus red mite, Panonychus citri (McGregor), on citrus trees with Argentine ants, and concluded that ants interfered with a key natural en-
emy, the spider mite destroyer, Stethorus picipes Casey. In the present study, in contrast, densities of mealybug destroyer lar-vae were higher on ant-tended vines, reflecting their ability to successfully forage in patches where the mealybug density is higher ( Garcia & O’Neil, 2000 ). Furthermore, the larvae of the mealybug destroyer closely resemble mealybugs in that they are covered with whitish strands of wax. It may be a common as-sumption that the beetle larvae mimic mealybugs to avoid detec-tion by ants, but no references could be found that documented the adaptive advantage of such mimicry by the beetle larvae, and moreover Bartlett (1961) reported ant interference with adult mealybug destroyers. In support of the mimicry hypothe-sis, it was found here that the mealybug destroyer populations fare better in the presence of ants than do the encyrtid parasi-toids, other lady beetles, and chrysopids. The beetle larvae’s behavioural changes when contacted by ants, which have not previously been described, further support the mimicry hypoth-esis. They also provide an excellent example, the first for coccinellid larvae, of a behavioural component in mimicry sig -nalling ( Edmunds, 1974; Mallet & Gilbert, 1995; Howarth et al. , 2004 ). The acceptance of the beetle larvae as mealybugs by the ants after antennal examination suggests that further study is warranted to determine if there is a chemical component to the mimicry display as well as visual or tactile cues.
Tending ants might also create a spatial refuge where the homopteran is protected from natural enemies ( Hawkins et al. , 1993 ). One of the more often cited examples concerns the California red scale on citrus in southern California. The scale population is under successful biological control and it has been suggested that the observed long-term stability of the popula-tion is due, in part, to a spatial refuge on the inner tree sections where parasitism by Aphytis melinus DeBach (Hymenoptera: Aphelinidae) was very low ( Reeve & Murdoch, 1986 ). The scale population on the inner tree scaffolding provided the ma-jority of reproductive females ( Murdoch et al. , 1989 ) and had lower parasitism rates even though acceptable host stages were abundant ( Walde et al. , 1989; Yu et al. , 1989 ). The researchers
Table 6. Relationship of treatment (ant-tending vs ant-exclusion) and sample date on the distribution of the grape mealybugs (percentage of the total mealybug population) on four sampled vine sections (trunk/cordon, spur, leaves/cane, and grape clusters) for data collected in a North Coast vineyard in 1998 and 1999.
Vine section Source of variation
1998 * 1999 *
F -ratio P -value F -ratio P -value
Trunk/Cordon Treatment 0.236 0.629 4.619 0.037 Sample date 0.485 0.022 2.618 0.019 Treatment × Date 0.342 0.884 0.389 0.920
Spur Treatment 0.162 0.689 7.188 0.011 Sample date 2.343 0.057 5.512 <0.001 Treatment × Date 0.488 0.784 0.981 0.488
Leaves/Cane Treatment 1.333 0.254 2.392 0.129 Sample date 4.148 0.037 1.181 0.332 Treatment × Date 0.729 0.606 1.512 0.181
Grape Treatment 1.125 0.295 4.322 0.043 Sample date 3.277 0.013 6.626 <0.001
Treatment × Date 0.481 0.788 0.732 0.663
* Degrees of freedom are 1,5,5,43 and 1,8,8,44 for 1998 and 1999 respectively. Data from collections between seasons (29 January 1999) or at the end of the trial (21 October 1999) are not used in the analysis.
Fig. 6. Percentage distribution of live grape mealybug per vine per sample on four sampled vine sections in (a) ant-tended and (b) ant-exclusion treatments. Data analysis is provided in Table 6 .

594 Kent M. Daane et al.
© 2007 The AuthorsJournal compilation © 2007 The Royal Entomological Society, Ecological Entomology, 32, 583–596
suggested that Argentine ants foraging mainly on the inner tree scaffolding created the spatial refuge ( Reeve & Murdoch, 1986 ), although ant exclusion experiments provided only weak support for this hypothesis ( Murdoch et al. , 1995 ). In fact, experimental manipulation of the refuge population indicated that Argen-tine ants did not create a spatial refuge from natural enemies ( Murdoch et al. , 1996 ) and that stability is a property of local interactions between California red scale and A. melinus ( Murdoch et al. , 2005 ).
In the vineyard systems studied, there was no evidence that a spatial refuge was created by ants that foraged more heavily on the inner vine sections (trunk and spurs) than the outer sections (leaves and canes). High parasitism levels were only found at the Central Coast sites, where parasitism by P. flavidulus and L. epona reduced obscure mealybug densities ( Figs 1 and 2 ). A refuge effect would have been indicated by a greater percentage of the mummified mealybugs found in the inner vine sections in the ant-excluded treatment, or a greater percentage of the mealybug population on the outer vine sections in the ant-tended treatment. Neither situation was observed ( Table 3 ). Rather, mum-mified mealybugs were found on the trunk and spurs, the woody sections of the vine, in both treatments Fig. 3b,d ), even though ant presence was higher on these vine sections. It is possible that mealybugs were parasitised in exposed locations and later moved. Some koinobiont parasitoids induce behavioural changes in their
Fig. 7. Percentage grape mealybug developmental stages per vine per sample on vines in (a) ant-tended and (b) ant-exclusion treatments. Data analysis is provided in Table 4 .
host ( Quicke, 1997 ), such as causing the host to move to more protected locations to avoid predation or hyperparasitism ( Brodeur & McNeil, 1992 ). However, the similar mealybug distribution patterns further support the null hypothesis that no spatial refuge was created, and instead simply indicate that parasitoids forage in locations where the hosts are most common.
In the North Coast site there was little natural enemy activity observed, especially parasitoids, and yet mealybug densities were reduced in ant-excluded treatments. These results suggest that ants provide benefits to the tended mealybugs other than protection from natural enemies. One possibility is that ant-tended mealybugs may also be larger, due to improved feeding habitat, and have higher fecundity ( Buckley, 1987 ). The per capita change in grape mealybug density, during the period when crawlers emerge from the ovisac, was higher in the ant-tended than ant-excluded treatment in both years ( Table 5 ). Both the removal of honeydew and the potential increase in either mealybug fecundity or survival of emerging crawlers would in-crease the mealybug population without the addition of protec-tion from natural enemies. In contrast, at the Central Coast vineyards there was no treatment impact on the per capita popu-lation increase in obscure mealybug density during the period of egg hatch ( Table 2 ). It was observed, however, that obscure mealybug population densities on some ant-tended vines were so high that the ants could not remove all the honeydew. Obscure mealybug crawlers were observed caught in the drying excre-ment, which impeded their movement to better feeding sites. Therefore, a likely benefit of ant tending is the removal of hon-eydew from the mealybug habitat as described by Buckley (1987 and references therein). Whether or not mealybug fecun-dity is improved by ant tending can not be directly assessed by this field measurement of the per capita population increase. Controlled laboratory experiments are needed.
Ants were observed carrying mealybugs, and it was hypoth-esised that this behaviour might help place the mealybugs on better feeding locations or sites better protected from natural enemies. The mealybug population distributions, however, were similar on different vine sections in ant-tended and ant-excluded vines, which indicates that the ants had no overall effect on mealybug location on the vine.
Previous studies on the ecological impacts of Argentine ant in-vasions have focused on their effects on ground-dwelling arthro-pods, using baits or pitfall traps to conduct comparative faunal surveys. These studies have amply documented the elimination of native ants by Argentine ants in California and elsewhere ( Ward, 1987; Human & Gordon, 1996; Holway, 1998 ). Effects on other ground-dwellers are mixed, however. In California, Holway (1998) found no differences in spider, carabid, or overall arthropod spe-cies abundance between sites infested with Argentine ants and sites that were not infested, but Cole et al. (1992) found significant changes in a Hawaiian ground arthropod community. Hawaii has no native ants, however, and its native fauna may thus be more susceptible to invading ants. As Holway (1998) points out, the use of pitfall traps does not allow investigation of other parts of the ecosystem where Argentine ants are expected to have an effect, including, most obviously, homopterans feeding on plants.
Here, the effects of the invasive Argentine ant were shown to include an increase in mealybug population density. These

Argentine ants and vineyard insects 595
© 2007 The AuthorsJournal compilation © 2007 The Royal Entomological Society, Ecological Entomology, 32, 583–596
effects carry across trophic levels, influencing the abundances of parasitoids and predators and the fruit quality of the host plant. It is also clear that the effects of Argentine ant invasions on natural enemy species are variable, even within guilds, and may depend on the extent to which individual species have ad-aptations that favour coexistence with ants. There are also obvi-ous ant effects on the grape crop. In general, impacts on plant fitness of ant – homopteran mutualisms are sparsely documented ( Styrsky & Eubanks, 2007 ). For Argentine ants, the only evi-dence comes from plant communities in which many species rely on seed-dispersing ants that are displaced by Argentine ants (which do not disperse seeds) ( Christian, 2001; Ness & Bronstein, 2004 ). Yet to be resolved are implications that the large Argentine ant populations residing in vineyards may be altering adjacent natural ecosystems. The three vineyards stud-ied abut natural areas of California chaparral and oak – grassland, where the soil is undisturbed and often too dry for the Argentine ant to become established ( Human et al. , 1998; Holway et al. , 2002b ). The cultivated vineyards, however, with their huge populations of Argentine ants, make a potential base camp for Argentine ant invasions into the surrounding natural ecosystems.
Acknowledgements
The America Vineyard Foundation, California Competitive Grants Program for Enology and Viticulture, Viticulture Con-sortium, and the California Table Grape Commission funded re-search on Argentine ant invasions in vineyards. We thank the Domaine Chandon, Paragon, and Wolff vineyards for use of their land; Chris Geiger, Andrea Jani, Kerry Weir, Eric Davidian, John Andrews, and Glenn Yokota for assistance in the laboratory, field, and insectary; Erik Nelson (University of California, Berkeley) and two anonymous reviewers for suggestions on an earlier version of this manuscript; and Serguei Triapitsyn (University of California, Riverside) and Renato Ripa (Instituto de Investigaciones Agropecurias, Centro National de Entomo logía, La Cruz, Chile) for help with importation of obscure mealybug parasitoids.
References
Bartlett , B.R . ( 1961 ) The infl uence of ants upon parasites, predators, and scale insects . Annals of the Entomological Society of America , 54 , 543 – 551 .
Barzman , M.S. & Daane , K.M . ( 2001 ) Host-handling behaviours in parasitoids of the black scale: a case for ant-mediated evolution . Journal of Animal Ecology , 70 , 237 – 247 .
Ben-Dov , Y . ( 1995 ) A Systematic Catalogue of the Mealybugs of the World (Insecta: Homoptera: Coccoidea: Pseudococcidae, and Puto-idae) with Data on Geographical Distribution, Host Plants, Biology, and Economic Importance . Intercept Ltd , Andover, U.K .
Brodeur , J. & McNeil , J.N . ( 1992 ) Host behaviour modifi ed by the en-doparasitoid Aphidius nigripes : a strategy to reduce hyperparasitism . Ecological Entomology , 17 , 97 – 104 .
Buckley , R.C . ( 1987 ) Interactions involving plants, Homoptera, and ants . Annual Review of Ecology and Systematics , 18 , 111 – 135 .
Buckley , R. & Gullan , P . ( 1991 ) More aggressive ant species (Hyme-noptera: Formicidae) provide better protection for soft scales and
mealybugs (Homoptera: Coccidae, Pseudococcidae) . Biotropica , 23 , 282 – 286 .
Christian , C.E . ( 2001 ) Consequences of a biological invasion reveal the importance of mutualism for plant communities . Nature , 413 , 635 – 639 .
Clausen , C.P . ( 1924 ) The parasites of Pseudococcus maritimus (Eh-rhorn) in California (Hymenoptera: Chalcidoidea). Part II. Biological studies and life history . University of California Publications in Entomology , 3 , 223 – 292 .
Cole , F.R. , Medeiros , A.C. , Loope , L.L. & Zuehlke , W.W . ( 1992 ) Effects of the Argentine ant on arthropod fauna of Hawaiian high-elevation shrubland . Ecology , 73 , 1313 – 1322 .
Cudjoe , A.R. , Neuenschwander , P. & Copland , M.J.W . ( 1993 ) Interfer-ence by ants in biological control of the cassava mealybug Phenacoc-cus manihoti (Hemiptera: Pseudococcidae) in Ghana . Bulletin of Entomological Research , 83 , 15 – 22 .
Daane , K.M. , Gonzalez , D. , Bianchi , M. , Bentley , W.J. , Godfrey , K.E. , Powell , D . et al . ( 1996 ) Mealybugs in grape vineyards . Grape Grower , 28 , 10 , 12 .
Daane , K.M. , Sime , K.R. , Hogg , B.N. , Cooper , M.L. , Bianchi , M.L. , Rust , M.K . et al . ( 2006 ) Effects of liquid insecticide baits on Ar-gentine ants in California’s coastal vineyards . Crop Protection , 25 , 592 – 603 .
Daane , K.M. , Smith , R.H. , Klonsky , K.M. & Bentley , W.J . ( 2005 ) Or-ganic vineyard management in California . Organic-Research.com , 5 , 37 – 55 .
Edmunds , M . ( 1974 ) Defence in Animals: A Survey of Anti-Predator Defences . Longman, Harlow , U.K .
Essig , E.O . ( 1914 ) The mealybugs of California . Monthly Bulletin of the California State Commission of Horticulture , 3 , 18 – 143 .
Flaherty , D.L. , Peacock , W.L. , Bettiga , L. & Leavitt , G.M . ( 1982 ) Chemicals losing effect against grape mealybug . California Agricul-ture , 36 , 15 – 16 .
Flaherty , D. L. , Phillips , P. A. , Legner , E. F. , Peacock , W. L. & Bentley , W. J . ( 1992 ) Mealybugs . Grape Pest Management ( ed . by D. L. Fla-herty , L. P. Christensen , W. T. Lanini , J. J. Marois , P. A. Phillips , and L. T. Wilson ), pp . 159 – 165 . Publication 3343, University of Cal-ifornia, Division of Agricultural and Natural Resources , Oakland, California .
Flanders , S.E . ( 1951 ) The role of the ant in the biological control of homopterous insects . Canadian Entomologist , 83 , 93 – 98 .
Garcia , J.F. & O’Neil , R.J . ( 2000 ) Effect of Coleus size and variegation on attack rates, searching strategy, and selected life history character-istics of Cryptolaemus montrouzieri (Coleoptera: Coccinellidae) . Biological Control , 18 , 225 – 234 .
Geiger , C.A. & Daane , K.M . ( 2001 ) Seasonal movement and sampling of the grape mealybug, Pseudococcus maritimus (Ehrhorn) (Homop-tera: Pseudococcidae) in San Joaquin Valley vineyards . Journal of Economic Entomology , 94 , 291 – 301 .
Geiger , C.A. , Daane , K.M. & Bentley , W.J . ( 2001 ) Development of a sampling program for improved management of the grape mealybug . California Agriculture 55 , 19 – 27 .
Godfrey , K.E. , Daane , K.M. , Bentley , W.J. , Gill , R.J. & Malakar-Kuenen , R . ( 2002 ) Mealybug in California Vineyards . Publication 21612, University of California, Division of Agriculture and Natural Resources , Oakland, California .
Golino , D.A. , Sim , S.T. , Gill , R. & Rowhani , A . ( 2002 ) California mealybugs can spread grapevine leafroll disease . California Agricul-ture , 56 , 196 – 201 .
González-Hernández , H. , Johnson , M.W. & Reimer , N.J . ( 1999 ) Impact of Pheidole megacephala (F.) (Hymenoptera: Formicidae) on the bi-ological control of Dysmicoccus brevipes (Cockerell) (Homoptera: Pseudococcidae) . Biological Control , 15 , 145 – 152 .

596 Kent M. Daane et al.
© 2007 The AuthorsJournal compilation © 2007 The Royal Entomological Society, Ecological Entomology, 32, 583–596
Haney , P.B. , Luck , R.F. & Moreno , D.S . ( 1987 ) Increases in densities of the citrus red mite, Panonychus citri (Acarina: Tetranychidae), in as-sociation with the Argentine ant, Iridomyrmex humilis (Hymenoptera: Formicidae), in Southern California (USA) citrus . Entomophaga , 32 , 49 – 58 .
Hawkins , B.A. , Thomas , M.B. & Hochberg , M.E . ( 1993 ) Refuge theory and biological control . Science , 262 , 1429 – 1432 .
Heimpel , G.E. & Collier , T.R . ( 1996 ) The evolution of host-feeding be-haviour in insect parasitoids . Biological Reviews , 71 , 373 – 400 .
Heimpel , G.E. , Rosenheim , J.A. & Mangel , M . ( 1997 ) Predation on adult Aphytis parasitoids in the fi eld . Oecologia , 110 , 346 – 352 .
Holway , D.A . ( 1998 ) Effect of Argentine ant invasions on ground-dwelling arthropods in northern California riparian woodlands . Oecologia , 116 , 252 – 258 .
Holway , D.A . ( 1999 ) Competitive mechanisms underlying the dis-placement of native ants by the invasive Argentine ant . Ecology , 80 , 238 – 251 .
Holway , D.A. , Lach , L. , Suarez , A.V. , Tsutsui , N.D. & Case , T.J . ( 2002a ) The causes and consequences of ant invasions . Annual Review of Ecology and Systematics , 33 , 181 – 233 .
Holway , D.A. , Suarez , A.V. & Case , T.J . ( 2002b ) Role of abiotic factors in governing susceptibility to invasion: a test with Argentine ants . Ecology , 83 , 1610 – 1619 .
Howarth , B. , Edmunds , M. & Gilbert , F . ( 2004 ) Does the abundance of hoverfl y (Syrphidae) mimics depend on the numbers of their hy-menopteran models? Evolution , 58 , 367 – 375 .
Human , K.G. & Gordon , D.M . ( 1996 ) Exploitation and interference competition between the invasive Argentine ant, Linepithema humile , and native ant species . Oecologia , 105 , 405 – 412 .
Human , K.G. , Weiss , S. , Weiss , A. , Sandlar , B. & Gordon , D.M . ( 1998 ) Effects of abiotic factors on the distribution and activity of the in-vasive Argentine ant (Hymenoptera: Formicidae) . Environmental Entomology , 27 , 822 – 833 .
James , D.G. , Stevens , M.M. , O’Malley , K.J. & Faulder , R.J . ( 1999 ) Ant foraging reduces the abundance of benefi cial and incidental arthro-pods in citrus canopies . Biological Control , 14 , 121 – 126 .
Karamaouna , F. & Copland , M.J.W . ( 2000 ) Oviposition behaviour, infl uence of experience on host size selection, and niche overlap of the solitary Leptomastix epona and the gregarious Pseudaphycus fl avidulus , two parasitoids of the mealybug Pseudococcus viburni . Entomologia Experimentalis et Applicata , 97 , 301 – 308 .
Lach , L . ( 2003 ) Invasive ants: unwanted partners in ant – plant interac-tions? Annals of the Missouri Botanical Garden , 90 , 91 – 108 .
Mallet , J. & Gilbert , L.E . ( 1995 ) Why are there so many mimicry rings? Correlations between habitat, behaviour and mimicry in Heliconius butterfl ies . Biological Journal of the Linnean Society , 55 , 159 – 180 .
Miller , D.R. , Gill , R.J. & Williams , D.J . ( 1984 ) Taxonomic analysis of Pseudococcus affi nis (Maskell), a senior synonym of Pseudococcus obscurus Essig, and a comparison with Pseudococcus maritimus (Ehrhorn) (Homoptera: Coccoidea: Pseudococcidae) . Proceedings of the Entomological Society of Washington , 86 , 703 – 713 .
Murdoch , W.W. , Briggs , C.J. & Swarbrick , S.J . ( 2005 ) Host suppression and stability in a parasitoid – host system: experimental demonstra-tion . Science , 309 , 610 – 613 .
Murdoch , W.W. , Luck , R.F. , Swarbrick , S.J. , Walde , S.J. , Yu , D.S. & Reeve , J.D . ( 1995 ) Regulation of an insect population under biologi-cal control . Ecology , 76 , 206 – 217 .
Murdoch , W.W. , Luck , R.F. , Walde , S.J. , Reeve , J.D. & Yu , D.S . ( 1989 ) A refuge for red scale: the role of size-selectivity by Aphytis : struc-tural aspects . Ecology , 70 , 1707 – 1714 .
Murdoch , W.W. , Swarbrick , S.L. , Luck , R.F. , Walde , S. & Yu , D.S . ( 1996 ) Refuge dynamics and metapopulation dynamics: an experi-mental test . American Naturalist , 147 , 424 – 444 .
Ness , J.H. & Bronstein , I.L . ( 2004 ) The effects of invasive ants on pro-spective ant mutualists . Biological Invasions , 6 , 445 – 461 .
Noyes , J.S. & Hayat , M . ( 1994 ) Oriental Mealybug Parasitoids of the Anagyrini (Hymenoptera: Encyrtidae) . London Natural History Mu-seum , London .
Phillips , P.A. & Sherk , C.J . ( 1991 ) To control mealybugs, stop honey-dew seeking ants . California Agriculture , 45 , 26 – 28 .
Quicke , D.L.J . ( 1997 ) Parasitic Wasps . Chapman & Hall , London . Reeve , J.D. & Murdoch , W.W . ( 1986 ) Biological control by the parasit-
oid Aphytis melinus , and population stability of the California red scale . Journal of Animal Ecology , 55 , 1069 – 1082 .
Sanders , N.J. , Barton , K.E. & Gordon , D.M . ( 2001 ) Long-term dynamics of the distribution of the invasive Argentine ant, Linepithe ma -humile , and native ant taxa in northern California . Oecologia , 127 , 123 – 130 .
Stechmann , D.H. , Völkl , W. & Stary , P . ( 1996 ) Ant-attendance as a critical factor in the biological control of the banana aphid Pentalonia nigronervosa Coq. (Hom. Aphididae) in Oceania . Journal of Applied Entomology , 120 , 119 – 123 .
Styrsky , J.D. & Eubanks , M.D . ( 2007 ) Ecological consequences of interactions between ants and honey-dew producing insects. Proceedings of the Royal Society. Series B . Biological Sciences , 274 , 151 – 164 .
Suarez , A.V. , Bolger , D.T. & Case , T.J . ( 1998 ) Effects of fragmentation and invasion on native ant communities in coastal southern Califor-nia . Ecology , 79 , 2041 – 2056 .
Suarez , A.V. & Case , T.J . ( 2002 ) Bottom-up effects on persistence of a specialist predator: ant invasions and horned lizards . Ecological Applications , 12 , 291 – 298 .
Suarez , A.V. , Holway , D.A. & Case , T.J . ( 2001 ) Patterns of spread in biological invasions dominated by long-distance jump dispersal: insights from Argentine ants . Proceedings of the National Academy of Sciences of the United States of America , 98 , 1095 – 1100 .
Suarez , A.V. , Richmond , J.Q. & Case , T.J . ( 2000 ) Prey selection in horned lizards following invasion of Argentine ant in Southern Cali-fornia . Ecological Applications , 10 , 711 – 725 .
Swirski , E. , Izhar , Y. , Wysoki , M. , Gurevitz , E. & Greenberg , S . ( 1980 ) Integrated control of the long-tailed mealybug, Pseudococcus longis-pinus (Homoptera: Pseudococcidae) in avocado plantations in Israel . Entomophaga , 25 , 415 – 426 .
Völkl , W . ( 1994 ) The effect of ant-attendance on the foraging behaviour of the aphid parasitoid Lysiphlebus cardui . Oikos , 70 , 149 – 155 .
Völkl , W. & Kroupa , A.S . ( 1997 ) Effects of adult mortality risks on parasitoid foraging tactics . Animal Behaviour , 54 , 349 – 359 .
Völkl , W. & MacKauer , M . ( 1993 ) Interactions between ants attending Aphis fabae ssp. cirsiiacanthoidis on thistles and foraging parasitoid wasps . Journal of Insect Behavior , 6 , 301 – 312 .
Walde , S.J. , Luck , R.F. , Yu , D.S. & Murdoch , W.W . ( 1989 ) A refuge for red scale: the role of size-selectivity by a parasitoid wasp . Ecology , 70 , 1700 – 1706 .
Ward , P.S . ( 1987 ) Distribution of the introduced Argentine ant ( Irido-myrmex humilis ) in natural habitats of the lower Sacramento Valley and its effects on the indigenous ant fauna . Hilgardia , 55 , 1 – 16 .
Way , M.J . ( 1963 ) Mutualism between ants and honeydew-producing homopterans . Annual Review of Entomology , 8 , 307 – 344 .
Woodworth , C.W . ( 1908 ) The Argentine ant in California . University of California Agricultural Experiment Station Circular , 38 , 1 – 11 .
Yu , D.S. , Luck , R.F. & Murdoch , W.W . ( 1989 ) Competition, resource partitioning and coexistence of an endoparasitoid Encarsia pernicio-si and an ectoparasitoid Aphytis melinus of the California red scale . Ecological Entomology , 15 , 469 – 480 .
Accepted 1 February 2007 First published online 1 October 2007




















