
Effects of roasting on barley β-glucan, thermal, textural and pasting properties
Upload
independentCategory
view
2download
0
Fish & Shellfish Immunology (1998) 8, 409–424Article ID: fi980148
Immunomodulation by dietary vitamin C and glucan inrainbow trout (Oncorhynchus mykiss)
VIVIANE VERLHAC1*, ALEX OBACH
3, JACQUES GABAUDAN1, WILLY SCHU}EP
2AND
REID HOLE3
1CRNA, Société Chimique Roche, B.P. 170, 68305 Saint-Louis Cedex, France2Analytical Department, F. Hoffmann–La Roche Ltd, CH–4002, Basle,
Switzerland3Nutreco Aquaculture Research Centre (ARC), P.O. Box 353, N–4033 Forus,
Norway
(Received 11 November 1997, accepted after revision 23 March 1998)
The aim of this study was to determine the influence of dietary glucan andvitamin C, alone and in combination, on the cellular and humoral immuneresponses of rainbow trout. After 4 weeks of adaptation on a control dietcontaining glucan at 0 ppm and vitamin C at 150 ppm, four groups of rainbowtrout were fed the experimental diets containing vitamin C at 150 and1000 ppm plus a fixed amount of glucan, or no glucan for 2 weeks. The fishwere then switched back to the control diet. Non-specific immunity wasinvestigated at the end of the two-week experimental feeding period, and thenagain 4 weeks later. Macrophage activities such as oxidative burst andpinocytosis were determined along with complement activation and lysozymelevels. The specific immune response was evaluated as: (1) proliferation oflymphocytes induced by mitogen and (2) the antibody response after vacci-nation against enteric redmouth disease. Fish were vaccinated at the end ofthe 2-week experimental feeding period and antibody titres were determinedevery second week for the 16 following weeks. The results of this trial showeda significant e#ect of dietary vitamin C but not glucan on the non-specificimmune response of rainbow trout such as increased oxidative burst, pino-cytosis and lysozyme activity. Dietary treatments had no e#ect on alternativepathway of complement activation. A significant enhancing e#ect of dietarybeta-1,3/1,6 yeast glucan was observed on concanavalin A-induced prolifer-ation of lymphocytes and the antibody response after vaccination againstenteric redmouth disease. When considering the antibody titres at the peaktime, glucan or vitamin C significantly enhanced the response. This was alsoobserved on serum complement levels measured at the same time. Results fromthis trial helped to establish the beneficial e#ect of a dietary combination ofglucan and vitamin C on the immune response of rainbow trout, as well as toevaluate the specific e#ect of each substance. ? 1998 Academic Press
Key words: immunomodulation, nutrition, glucan, vitamin C, rainbow trout.
I. Introduction
During the last decade, increasing attention has been paid to the use ofimmunostimulants in disease control, especially to those which elevate
*Corresponding author: Email: [email protected]
4091050–4648/98/060409+16 $30.00/0 ? 1998 Academic Press

410 V. VERLHAC ET AL.
non-specific defence mechanisms (Anderson, 1992), activating early protectionagainst infections.
Beta-glucans are polysaccharides composed only of glucose moleculeslinked beta 1-3 and beta 1-6. They are normally found in the cell walls of yeastand mycelial fungi. Glucans function as potent immunostimulants in mam-mals (Di Luzio, 1985) and in fish (Robertsen et al., 1990). In fish, they canactivate macrophages, increasing their capacity to kill pathogens (Jørgensenet al., 1993a). They have also been shown to reinforce other non-specificimmune factors such as lysozyme and complement activities (Engstad et al.,1992). When administered orally, glucans have proven protective e#ect inseveral fish species and against a variety of bacterial pathogens includingAeromonas salmonicida (Nikl et al., 1993), Vibrio salmonicida (Raa et al., 1992),or Pasteurella piscicida (Noya et al., 1995).
The capacity of vitamin C to stimulate the immune response has beenestablished in a number of animal species including fish. The impact ofvitamin C on the immune system of fish has been reviewed by Lall and Olivier(1993) and Waagbø (1994). Elevated doses of dietary vitamin C have beenshown to enhance several non-specific immune parameters such as macro-phage activities (Blazer, 1982), lymphocyte proliferation (Verlhac et al., 1993),natural cytotoxicity (Verlhac & Gabaudan, 1994), complement activity(Hardie et al., 1991) and lysozyme activity (Waagbø et al., 1993). Elevatedantibody levels after vaccination have also been reported in fish fed highlevels of vitamin C (Navarre & Halver, 1989; Waagbø et al., 1993). In previousexperiments, Verlhac et al. (1996) studied the e#ect of combinations of glucansand doses of vitamin C varying from 150 to 4000 mg ascorbic acid equivalentper 1 kg of feed on the immune response of fish. The results showed that thecombinations of glucans and elevated doses of vitamin C had a positive impacton both the non-specific and specific immune parameters tested. The presentexperiment aims to further investigate the immunomodulatory role of eachsubstance in fish.
II. Materials and Methods
EXPERIMENTAL DESIGN
The experiment was designed to evaluate the immunomodulating propertiesof glucan and vitamin C after a short feeding period of 2 weeks, and theduration of any e#ect. After 4 weeks of adaptation on a control diet (no glucanand 150 ppm vitamin C), fish were fed the four experimental diets containingvitamin C at either 150 or 1000 mg ascorbic acid-equivalent per 1 kg of feedand with or without glucan (at a constant level) for 2 weeks. They were thenswitched back to the control diet during the 4 following weeks. The immuneresponse was investigated at the end of the experimental feeding period(week 0), and 4 weeks after switching back to the control diet.
ANIMALS AND REARING CONDITIONS
Rainbow trout (n=360) with an initial body weight of 146·2&4·2 g wererandomly divided between twelve 205 l tanks on a recirculating system

IMMUNOMODULATION BY DIETARY VITAMIN C 411
supplied with fresh ground water. Three tanks were randomly assigned toeach dietary treatment. Water temperature was recorded daily and variedfrom 14) C to 17) C during the experiment.
EXPERIMENTAL DIETS, FEEDING CONDITIONS AND GROWTH
The experimental diets (Table 1) were produced by Nutreco ARC(Stavanger, Norway). Vitamin C was added as ascorbate-2-polyphosphate(ROVIMIX> STAY-C>, 25 Ho#mann–La Roche, Basle, Switzerland) beforeextrusion. All vitamin C levels are given in mg ascorbic acid (AA)-equivalentper 1 kg of feed.
Beta-1,3/1,6 yeast glucan (MacroGard>, Biotec–Mackzymal, Tromsø,Norway) derived from cell wall of Saccharomyces cerevisiae, was included infeed before extrusion. The inclusion level was as recommended by themanufacturer. An approximate analysis of the diet is given in Table 1.
Fish were fed twice daily according to feeding ration tables, 7 days a weekthroughout the experiment.
Fish from each individual tank were bulk-weighed every 2 weeks starting atthe beginning of trial. Specific growth rate (% BW/day) was calculated as:
SGR=Ln (final mean body weight (g))"Ln (initial mean body weight (g))
#100time interval (days)
IMMUNE PARAMETERS
Two series of tanks were allocated to the study of cellular and humoralresponses (macrophage activities, mitogen-induced proliferation of peripheralblood lymphocytes, complement and lysozyme levels) and tissue vitamin Ccontents. In order to enable the testing of the four dietary groups for each ofthe parameters at exactly 4-week intervals, the start of the experimentalfeeding was delayed by 3 days in the two series of tanks and the sameparameter was evaluated on fish from the same series throughout the exper-iment. A third series of tanks were allocated to the study of the antibodyresponse after the fish had been vaccinated against enteric redmouth disease.
Table 1. Declared and measured vitamin C content of the experimental diets
Treatment Yeast glucan
Vitamin C(in mg ascorbic acid equivalent per kg of feed)
Declared Measured
No Glu+Vit C 150 " 150 161Glu+Vit C 150 + 150 170No Glu+Vit C 1000 " 1000 818Glu+Vit C 1000 + 1000 911
Approximate analysis of the diets in % dry matter: crude protein: 47·7; total lipid: 25·2;carbohydrates (nitrogen free extract and crude fibre): 11·0; ash: 16·2.

412 V. VERLHAC ET AL.
Phagocyte activities were evaluated for their oxidative burst and pinocyto-sis capacities on the same fish. The oxidative burst activity was measured bychemiluminescence assay. Neutrophils and macrophages were isolated fromthe head-kidney on a Percoll density gradient (Plytycz et al., 1989). Chemilu-minescence response was measured on an automated luminometer (Bioorbit1251, Wallac) after stimulation of phagocytes with chicken serum-opsonisedzymosan (Mathews et al., 1990). Substrates used to assess the chemilumines-cence response were either luminol (5-amino-2,3-dihydro-1,4-phthalazinedione)or lucigenin (bis-N-methylacridinium nitrate). Luminol allows the detection ofmajor reactive oxygen species: hydrogen peroxides (H2O2), hydroxyl radicals(OH") and singlet oxygen (1O2
·) while lucigenin allows the detection ofsuperoxide anions (O2
"). Phagocyte suspensions were prepared in Cortlandsaline (Wolf, 1963) at 107 cells per ml. Luminol (Sigma) was stored at 10"2
M indimethylsulphoxide (DMSO). The working solution was obtained by dilutingthe stock solution at 1:100. Lucigenin (Sigma) was prepared at 1 mg per ml ofCortland saline. Two hundred ìl luminol or lucigenin working solutions,100 ìl phagocyte suspension and 700 ìl opsonised zymosan working solutionswere mixed in polystyrene vials. The volume was adjusted to 1 ml withCortland saline. Zymosan was the last component added; subsequently thetubes were immediately put into the luminometer thermoregulated at 22) C.The time-course of chemiluminescence response was followed by counting therate of photons emitted at constant time intervals (166 sec) for 4 h 30 min.Results are expressed as the area under the curve of chemiluminescenceresponse for individual fish versus time (mV#sec). This method was adaptedfrom Scott et al. (1985) and Stave et al. (1983). The chemiluminescence responseof non-stimulated phagocytes was below 5 and 2·4#104 mV#sec with luminoland lucigenin, respectively.
Pinocytosis was also measured on the same individuals by the uptake ofneutral red droplets by headkidney phagocytes. The pinocytic activity wasdetermined by spectrophotometric measurement of ingested neutral red dyeaccording to Mathews et al. (1990).
Concerning the mitogen-induced proliferation assay, 5#105 lymphocytesisolated on Ficoll gradient from peripheral blood were cultured in 240 ìlof culture medium supplemented with 2% heat-inactivated rainbow troutserum (Verlhac & Gabaudan, 1994), in presence of Concanavalin A (Con A)at 10 ìg/ml or lipopolysaccharides (LPS) from Escherichia coli 0127:B8 at50 ìg/ml. Cultures were pulsed with 3H-thymidine after 72 h of incubation at22) C and the radioactivity was measured after an 18-h pulse. Values forunstimulated controls ranged from 100 to 600 cpm. Results are expressed asstimulation index (SI).
Classical and alternative pathways of complement activation weremeasured in plasma. Blood samples from the fish were collected by caudal veinpuncture in heparinised syringes and centrifuged at 3000 rpm for 5 min at 4) C.The 50%-haemolysis assays for classical and alternative pathways wereconducted in microplates with goldfish and rabbit red blood cells, respect-ively. The method for classical pathway determination was that of Sakai(1981) modified by Verlhac and Gabaudan (1994). The alternative pathway wasdetermined using the method of Yano (1992), but modified as follows: the tests

IMMUNOMODULATION BY DIETARY VITAMIN C 413
were performed using rabbit red blood cells as target cells in the presence ofethylene glycol tetra-acetic acid (EGTA) and Mg2+.
Lysozyme level in plasma was determined by the turbidimetric assay inmicroplates according to Ellis (1990). Results were expressed in units oflysozyme per ml of plasma. One unit is defined as the amount of samplecausing a decrease in absorbance of 0·001/min at 450 nm.
Antibody response was determined after vaccination of individually taggedfish against enteric redmouth disease by intraperitoneal injection of 109
formalin-killed Yersinia ruckeri per fish in 0·2 ml of phosphate-bu#ered saline(PBS, pH 7·5) (Yersivax, Rhône–Mérieux Laboratories, Lyon, France). Vacci-nation was applied at the end of the 2-week experimental feeding period.Every second week after immunisation, serum from all the fish was collected.The fish were monitored for 10 weeks post-treatment. Antibody titres weredetermined by indirect enzyme-linked immunosorbent assay (ELISA) (Vergnet& Dunier, 1993) using 96-well microplates coated with 100 ìl of the antigen,Yersinia ruckeri vaccine diluted at 1:50 in 0·05M carbonate–bicarbonate bu#er(pH 9·6), per well. Following overnight incubation at 4) C, unbound antigenwas removed by three successive washings with distilled water. The micro-plates were kept at 4) C until use. In order to saturate the plates, 200 ìl of a10% solution of skimmed milk powder dissolved in distilled water were addedto each well of coated plates.
After 1 h incubation at room temperature, the microplates were washedthree times with a 2% skimmed milk powder dissolved in distilled water, usinga microplate washer (Biorad). Serial dilutions from 1:200 to 1:3200 of the serato be tested were made in PBS–Tween (PBS containing 0·05% Tween 20,Sigma). Each well received 100 ìl of diluted serum. A standard antiserum wasalso used. After 1 h 30 min incubation at room temperature, the plates werewashed four times with 2% skimmed milk solution. Then, 100 ìl of I-14monoclonal antibody diluted at 1:50 in PBS–Tween were added to each well.The plates were incubated at room temperature for 1 h and washed 5 times asabove. One hundred ìl of horseradish peroxidase-labelled goat anti-mouse IgG (KPL) diluted at 1:1000 were added to each well. After 1 h incubation, themicroplates were washed 3 times with 2% skimmed milk solution and 3 timeswith PBS–Tween. One hundred ìl of TMB Microwell Peroxidase substrate(KPL) were distributed in each well. After 10 min incubation at roomtemperature, 100 ìl of 1M orthophosphoric acid were added to each well to stopthe reaction. Absorbance was measured at 450 nm with a 540-nm referencefilter on a Biorad spectrophotometer. The standard antiserum was used as areference in the calculation of antibody titres (Arkoosh & Kaattari, 1990). Oneunit of antibody activity was calculated as the number of ìl of standardantiserum giving 50% of the maximum absorbance. For each serum to betested, the number of ìl of serum corresponding to the 50% absorbance of thestandard is determined. The antibody titre of the test sera in units (U)/ml iscalculated as follows:
Activity of test serum =
(Activity of the standard, U/ìl) #(50% volume of standard)
50% volume of test serum

414 V. VERLHAC ET AL.
Areas under the curves (AUC) of antibody response were calculated forindividual fish. Statistical analysis was run on AUC. Complement levelsactivated through the classical pathway were also measured 2 and 8 weeksafter immunisation and compared to the antibody titres at the same time.
VITAMIN C ANALYSES
Ascorbic acid (AA) contents of liver, headkidney, plasma and leucocytes offive fish per treatment were measured by the fluorimetric assay (Vuilleumier &Keck, 1989) prior to the experimental feeding and at each sampling time. AAcontent of the experimental diets was also determined according to Wanget al. (1988).
STATISTICAL ANALYSIS
Two-way analysis of variance and Newman–Keul’s multiple range test wererun to compare the dietary treatment values at any one sampling time usingthe STATITCF> program. Statistical analysis was run in the same way for allthe parameters tested. Results of statistical analysis are presented below thecorresponding tables and figures.
III. Results
GROWTH PERFORMANCE
Specific growth rate (SGR) and feed conversion ratio (FCR) varied accord-ing to feeding rate. SGR ranged from 1·3 to 1·6 and FCR ranged from 0·9 to 1·0.
TISSUE VITAMIN C LEVELS
Liver, headkidney and plasma ascorbic acid levels (Table 2) were measuredbefore the beginning of the experimental feeding and at weeks 0 and 4, andreflected the dietary intake in vitamin C. No variations were observed overthe feeding period in the two groups fed vitamin C at 150 ppm, with or withoutglucan. An increase in tissue AA storage at week 0 was shown in the groupsfed the elevated dose of vitamin C. Four weeks after the fish had been switchedback to the control diet, plasma and liver AA levels in the groups fed theelevated dose of vitamin C had returned to the level of the groups fed vitaminC at 150 ppm. Leucocyte ascorbic acid content was measured at the end of the2-week experimental feeding period and 4 weeks after the fish had beenswitched back to the control diet (Table 2). The amount of vitamin C stored inthe leucocytes from the fish fed vitamin C at 1000 ppm was greater than that offish fed 150 ppm just after the end of the experimental feeding period. Thisdi#erence was still observed 4 weeks after the fish had been switched back tothe control diet. Headkidney ascorbic acid levels measured before the begin-ning of the experimental feeding and at weeks 0 and 4 reflected the dietaryintake in vitamin C (Table 2). With regard to AA levels in headkidney, novariations were observed over the feeding period in the groups fed vitamin Cat 150 ppm. An increase in tissue AA storage at the end of the 2-week
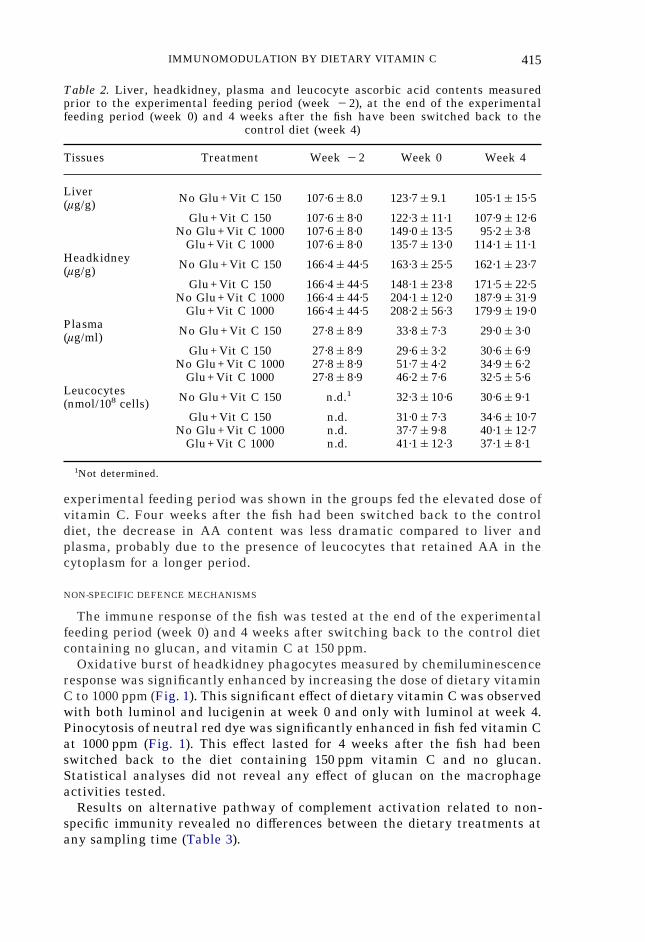
IMMUNOMODULATION BY DIETARY VITAMIN C 415
experimental feeding period was shown in the groups fed the elevated dose ofvitamin C. Four weeks after the fish had been switched back to the controldiet, the decrease in AA content was less dramatic compared to liver andplasma, probably due to the presence of leucocytes that retained AA in thecytoplasm for a longer period.
NON-SPECIFIC DEFENCE MECHANISMS
The immune response of the fish was tested at the end of the experimentalfeeding period (week 0) and 4 weeks after switching back to the control dietcontaining no glucan, and vitamin C at 150 ppm.
Oxidative burst of headkidney phagocytes measured by chemiluminescenceresponse was significantly enhanced by increasing the dose of dietary vitaminC to 1000 ppm (Fig. 1). This significant e#ect of dietary vitamin C was observedwith both luminol and lucigenin at week 0 and only with luminol at week 4.Pinocytosis of neutral red dye was significantly enhanced in fish fed vitamin Cat 1000 ppm (Fig. 1). This e#ect lasted for 4 weeks after the fish had beenswitched back to the diet containing 150 ppm vitamin C and no glucan.Statistical analyses did not reveal any e#ect of glucan on the macrophageactivities tested.
Results on alternative pathway of complement activation related to non-specific immunity revealed no di#erences between the dietary treatments atany sampling time (Table 3).
Table 2. Liver, headkidney, plasma and leucocyte ascorbic acid contents measuredprior to the experimental feeding period (week "2), at the end of the experimentalfeeding period (week 0) and 4 weeks after the fish have been switched back to the
control diet (week 4)
Tissues Treatment Week "2 Week 0 Week 4
Liver(ìg/g) No Glu+Vit C 150 107·6&8.0 123·7&9.1 105·1&15·5
Glu+Vit C 150 107·6&8·0 122·3&11·1 107·9&12·6No Glu+Vit C 1000 107·6&8·0 149·0&13·5 95·2&3·8
Glu+Vit C 1000 107·6&8·0 135·7&13·0 114·1&11·1Headkidney(ìg/g) No Glu+Vit C 150 166·4&44·5 163·3&25·5 162·1&23·7
Glu+Vit C 150 166·4&44·5 148·1&23·8 171·5&22·5No Glu+Vit C 1000 166·4&44·5 204·1&12·0 187·9&31·9
Glu+Vit C 1000 166·4&44·5 208·2&56·3 179·9&19·0Plasma(ìg/ml) No Glu+Vit C 150 27·8&8·9 33·8&7·3 29·0&3·0
Glu+Vit C 150 27·8&8·9 29·6&3·2 30·6&6·9No Glu+Vit C 1000 27·8&8·9 51·7&4·2 34·9&6·2
Glu+Vit C 1000 27·8&8·9 46·2&7·6 32·5&5·6Leucocytes(nmol/108 cells) No Glu+Vit C 150 n.d.1 32·3&10·6 30·6&9·1
Glu+Vit C 150 n.d. 31·0&7·3 34·6&10·7No Glu+Vit C 1000 n.d. 37·7&9·8 40·1&12·7
Glu+Vit C 1000 n.d. 41·1&12·3 37·1&8·1
1Not determined.

416 V. VERLHAC ET AL.
Plasma lysozyme levels determined just after the end of the experimentalfeeding (week 0) were enhanced in fish fed glucan and vitamin C at 1000 ppm.Statistical analysis showed a significant e#ect of vitamin C (Table 3). Fourweeks after the end of the experimental feeding, no significant e#ect ofvitamin C or glucan was observed.
SPECIFIC DEFENCE MECHANISMS
There was a significant enhancing e#ect of dietary glucan on the prolifer-ative response of peripheral blood lymphocytes induced by Concanavalin Ajust after the experimental feeding (week 0) and 4 weeks later (week 4) (Fig. 2).No significant variations were observed on the e#ect of dietary vitamin C onCon A-induced proliferation of peripheral blood lymphocytes and the diets didnot a#ect responses to lipopolysaccharides.
Individual kinetics of antibody responses were determined on individuallytagged fish from each dietary group. Results were expressed as the area under
6
0AU
C in
105 ×
mV
× s
ec4321
Week 0
5
Week 4S* NS
(a)
Vitamin C effectNS NSGlucan effectS* NSInteraction
20
0AU
C in
105 ×
mV
× s
ec
10
5
Week 0
15
Week 4S*** S**
(b)
Vitamin C effectNS NSGlucan effectNS NSInteraction
1.6
0
µg n
eutr
al r
ed/2
× 1
06 cel
ls
1.2
0.60.40.2
Week 0
1.4
Week 4
S** S***
(c)
Vitamin C effectNS NSGlucan effectNS NSInteraction
0.81.0
No Glu + Vit C 150Glu + Vit C 150
No Glu + Vit C 1000Glu + Vit C 1000
Fig. 1. E#ect of glucan and vitamin C on macrophage activities in rainbow trout:oxidative burst measured by chemiluminescence induced by zymosan usinglucigenin (a) and luminol (b) as substrates expressed as the area under the curveof chemiluminescence (AUC in 105#mV#sec) and pinocytosis (c) expressed inìg neutral red/2#106 cells. Results represent the mean of 10 fish per treatment.S: *P<0·1; **P<0·05; ***P<0·01. NS: P>0·1.

IMMUNOMODULATION BY DIETARY VITAMIN C 417
the curve (AUC) of antibody response for each fish and dietary treatmentgroups were statistically compared on this basis. A significant e#ect of dietaryglucan was observed in the areas under the curves of antibody response whileno significant e#ect of vitamin C was observed (Fig. 3).
Complement activated through the classical pathway was determined 2 and8 weeks post-immunisation. Results are presented in Table 4 as well as theantibody titres corresponding to the same sera. Two weeks after vaccination,antibody titres were still very low and no di#erences were observed amongtreatments. Complement activated through the classical pathway measured inthe same sera reached similar levels among the treatments. Eight weeks aftervaccination, the comparison of antibody titres showed a significant increasein antibodies related to both glucan and vitamin C. Complement activatedthrough the classical pathway was enhanced in the three groups fed glucanand/or vitamin C at 1000 ppm compared to the control group. Statisticalanalysis revealed a significant interaction between the two factors, glucanand vitamin C.
IV. Discussion
The results showed that vitamin C as ascorbate-2-polyphosphate and beta-1,3/1,6 glucan either given singly or in combination were able to enhancenon-specific and specific defence mechanisms of rainbow trout. Under theexperimental conditions, the results suggested that vitamin C enhancednon-specific immune parameters such as phagocyte activities and lysozymelevel while beta-1,3/1,6 glucan influenced the specific response to an antigen interms of antibody response to vaccination. Lymphocyte proliferation in thepresence of Concanavalin A which induces the proliferation of T cells was alsoenhanced by glucan. It is interesting that the classical pathway of complementactivation was modulated by both vitamin C and glucan when measured in the
Table 3. E#ect of glucan and vitamin C on alternative pathway of complementactivation and lysozyme level in rainbow trout at weeks 0 and 4 after the end of theexperimental feeding period. Results are expressed in AP50 units per ml of plasma forthe complement and in U/ml of plasma for lysozyme and represent the mean of 10 fish
per treatment
Treatment
Alternative pathway ofcomplement activation Lysozyme level
Week 0 Week 4 Week 0 Week 4
No Glu+Vit C 150 17·0&3·7 12·3&2·2 1562&378 1131&322Glu+Vit C 150 15·7&3·8 12·1&2·1 1568&494 1331&274No Glu+Vit C 1000 16·4&4·3 12·4&1·6 1584&236 1416&249Glu+Vit C 1000 16·9&2·9 12·7&1·6 1887&302 1122&206Vitamin C e#ect NS NS S* NSGlucan e#ect NS NS NS NSInteraction NS NS NS S*
S: *P<0·1. NS: P>0·1.

418 V. VERLHAC ET AL.
case of an in vivo activation such as the vaccination. This enhancementmeasured at the peak of the antibody response, 8 weeks post-vaccinationcorrelates with a similar significant increase in antibody titres due to bothglucan and vitamin C.
Verlhac et al. (1996) demonstrated that the oxidative burst of macrophagesmeasured by chemiluminescence in the presence of luminol which enables thedetection of several reactive oxygen species (ROS) derived from the super-oxide anion (O2
") such as hydrogen peroxide (H2O2), hydroxy radicals (OH")and singlet oxygen (1O2
·) was enhanced by vitamin C. This experimentconfirmed the e#ect as well as the duration of the response 4 weeks after thefish had been switched back to the control diet. Furthermore, a comparablee#ect of vitamin C was observed on the production of O2
" measured using
60
0S
tim
ula
tion
inde
x
40
30
20
10
Week 0
50
Week 4NS NS
(a)
Vitamin C effectS** S*Glucan effectNS NSInteraction
No Glu + Vit C 150Glu + Vit C 150
No Glu + Vit C 1000Glu + Vit C 1000
80
0
Sti
mu
lati
on in
dex
40
30
20
10
Week 0
50
Week 4NS NS
(b)
Vitamin C effectNS NSGlucan effectNS NSInteraction
60
70
Fig. 2. Influence of dietary glucan and vitamin C on proliferation of peripheral bloodlymphocytes induced by Concanavalin A (a) and lipopolysaccharide (b). Results areexpressed as the stimulation index (SI) and represent the mean of 8 fish pertreatment. Control values did not exceed 500 cpm. S: *P<0·1; **P<0·05. NS: P>0·1.

IMMUNOMODULATION BY DIETARY VITAMIN C 419
lucigenin as a substrate, despite the fact that the increase was not significant4 weeks later. In addition to the production of ROS, another aspect ofmacrophage activities such as pinocytosis was analysed and found to bemodulated in a similar manner by dietary vitamin C. The results thereforeshowed a correlation between the di#erent aspects of macrophage activitywhich have been investigated concerning the capacity of vitamin C given at ahigh dietary dose to modulate the activity of these cells. An e#ect of a highdietary dose of vitamin C has been demonstrated on the production of eitherO2
" or other ROS by rainbow trout macrophages (Verlhac & Gabaudan, 1992,1994) while Hardie et al. (1991) did not find any significant variation in O2
"
production measured via the reduction of nitroblue tetrazolium. Ingestion oflatex beads (Blazer, 1982; Verlhac et al., 1993) and phagocytosis of yeast cells(Roberts et al., 1995) were also enhanced by dietary vitamin C. The beneficiale#ect of beta-1,3/1,6 yeast glucan on macrophage activities, especially theirbactericidal and phagocytic properties has been clearly demonstrated whenthis substance was delivered by injection (Jørgensen et al., 1993a,b; Noyaet al., 1995) but the results on the capacity of this substance to enhance
No Glu + Vit C 150Glu + Vit C 150No Glu + Vit C 1000Glu + Vit C 1000
100
0
An
tibo
dy t
itre
s in
un
its/
µl o
f se
rum
40
20
14Days
60
80
28 42 56 70 84
Vitamin C effectGlucan effect
Interaction
NSS**NS
2690 + 10383574 + 17863302 + 16704304 + 2004
AUC values
––––
Fig. 3. E#ect of glucan and vitamin C on antibody response after vaccination againstenteric redmouth disease in rainbow trout. Results are presented as the kinetics ofantibody response per treatment group (n=15) and as the areas under the curves ofantibody response (AUC). Antibody titres are calculated in units per ml of serumusing a standard immune serum and AUC in (antibody units/ìl of serum)#week.S: **P<0·05. NS: P>0·1.

420 V. VERLHAC ET AL.
macrophage activities when administered orally are less clear. Noya et al.(1995) observed no variations in the erythrophagocytosis by peritoneal macro-phages of gilthead seabream fed beta-1,3/1,6 yeast glucan for 3 weeks. How-ever, Yoshida et al. (1995) demonstrated an enhancement of the non-specificprotection (NBT-positive cells and bacterial clearance) in African catfish fedbeta-1,3/1,6 yeast glucan for 4 weeks. After 5 weeks of feeding beta-1,3/1,6 yeastglucan to turbot, oxidative burst of headkidney macrophages was significantlyenhanced but the enhancement was no longer observed 2 weeks after the fishhad been switched back to the control diet (Ogier de Baulny et al., 1996).Rainbow trout fed beta-1,3/1,6 yeast glucan for 1 week presented higheroxidative burst, phagocytic activity, myeloperoxidase activity and potentialkilling activity, when compared to control fish fed no glucan (Siwicki et al.,1994). The e#ect of beta-1,3/1,6 yeast glucans by oral delivery should be furtherinvestigated in relation to the fish species, the feeding period, and theinclusion level.
The alternative pathway of complement activation was not influenced byglucan and vitamin C in this experiment while in the previous experiment runin our laboratory (Verlhac et al., 1996), an enhancement of this parameter wasobserved in the groups of fish fed vitamin C at elevated doses immediatelyafter the end of the 2-week experimental feeding period. Few data areavailable on the dietary immunomodulation of this parameter in fish.
The results of the e#ect of dietary vitamin C on enhancing lysozyme levelsin non-vaccinated fish showed an enhancement of this parameter in Atlanticsalmon (Waagbø et al., 1993), in turbot (Roberts et al., 1995) while nosignificant variations were observed in rainbow trout (Verlhac et al., 1995,1996). Results of Verlhac et al. (1996) on lysozyme level measured in rainbowtrout fed glucan and vitamin C in graded doses from 150 to 4000 ppm showedan increase in plasma lysozyme with increasing levels of vitamin C but thevariations were not considered to be significant due to high individualvariations. In this experiment, this parameter was enhanced by dietary
Table 4. Antibody titres (units per ìl of serum) and complement levels activated through theclassical pathway (CH50 units per ml) measured 2 and 8 weeks after vaccination against
enteric redmouth disease
Treatment
Classical pathway ofcomplement activation Antibody titres
Week 2post-vaccination
Week 8post-vaccination
Week 2post-vaccination
Week 8post-vaccination
No Glu+Vit C 150 17·3&4·8 19·6&4·0 24·5&16·5 46·3&23·2Glu+Vit C 150 20·0&5·7 26·9&5·5 19·4&10·3 74·1&44·3No Glu+Vit C 1000 19·7&5·5 27·0&3·8 24·6&17·4 64·8&29·4Glu+Vit C 1000 21·0&8·1 24·4&4·2 28·8&9·5 94·2&54·5Vitamin C e#ect NS S** NS S*Glucan e#ect NS S* NS S***Interaction NS S*** NS NS
S: *P<0·1; **P<0·05; ***P<0·01. NS: P>0·1.

IMMUNOMODULATION BY DIETARY VITAMIN C 421
vitamin C. Beta-1,3/1,6 yeast glucan fed to rainbow trout and turbot had noenhancing e#ect on lysozyme levels (Verlhac et al., 1996; Ogier de Baulnyet al., 1996).
Concerning the proliferation of peripheral blood lymphocytes, previoustrials have shown a significant enhancement of this parameter when fish hadbeen fed vitamin C at an elevated dose for longer periods (Verlhac &Gabaudan, 1994; Dunier et al., 1995; Wahli et al., 1997) while in this exper-iment, no e#ect was observed. This discrepancy in the e#ect of vitamin C onlymphocyte proliferation could be explained by a di#erence in the feedingperiod, showing that a 1000 ppm dose of vitamin C fed for 2 weeks was notsu$cient to ensure a stimulation of the proliferative response under ourexperimental conditions. Further investigations are required to determine thetime–dose response with dietary vitamin C on this parameter. No other dataare available on the e#ect of dietary glucan on the proliferative response offish but Hamuro and Chihara (1984) have defined lentinan, another beta-1,3/1,6 yeast glucan, as a T-cell oriented immunopotentiator functioning as amaturation factor of lymphoid cells in mammals.
Vaccinated fish fed glucan developed a higher antibody response whendetermined as the area under the kinetics of antibody titres, whatever thelevel of vitamin C in the diet. This observation confirmed results from Verlhacet al. (1996). Several studies have demonstrated the e$cacy of glucan as anadjuvant by increasing the antibody response by simultaneous injection withthe vaccine (Rørstad et al., 1993; Aakre et al., 1994). The comparison of thee#ect of each substance at the peak of the response in terms of antibody titresand classical pathway of complement activation revealed a significantenhancement of these parameters which was related to glucan and also tovitamin C. This finding shows that the classical pathway of complementactivation could perhaps be better modulated when the specific immuneresponse of the fish is activated. In view of these results, it should be notedthat measuring the e#ect of vitamin C on antibody production did not lead tothe same conclusions on the basis of areas under the curves or antibody titresat the peak time. On the other hand, results concerning the glucan e#ect werecomparable on both AUC and antibody titre peaks. Several studies haveshown an increase in the antibody response in fish fed an elevated dose ofvitamin C when measured at the maximum phase of the response (Erdal et al.,1991; Waagbø et al., 1993; Dunier et al., 1995; Li & Lovell, 1985; Navarre &Halver, 1989).
We are grateful to Christophe Zickler, Denis Constant and Didier Simon for theirtechnical assistance in this experiment.
References
Aakre, R., Wergeland, H. I., Aasjord, P. M. & Endresen, C. (1994). Enhanced antibodyresponse in Atlantic salmon (Salmo salar L.) to Aeromonas salmonicida cell wallantigens using a bacterin containing beta-1,3-M-glucan as adjuvant. Fish &Shellfish Immunology 4, 47–61.
Anderson, D. P. (1992). Immunostimulants, adjuvants, and vaccine carriers in fish:applications to aquaculture. Annual Review of Fish Diseases 2, 281–307.

422 V. VERLHAC ET AL.
Arkoosh, M. R. & Kaattari, S. L. (1990). Quantification of fish antibody to a specificantigen by an enzyme-linked immunosorbent assay (ELISA). In Techniques inFish Immunology (J. S. Stolen, T. C. Fletcher, D. P. Anderson, B. S. Roberson &W. B. van Muiswinkel, eds) pp. 15–24. U.S.A.: SOS Publications.
Blazer, V. (1982). The e#ects of marginal deficiency of ascorbic acid and alpha-tocopherol on the natural resistance and immune response of rainbow trout(Oncorhynchus mykiss). Ph.D. thesis, Univ. Microfilms Intern.–U.S.A., pp. 113.
Di Luzio, N. R. (1985). Update on the immunomodulating activities of glucans. SpringerSeminar Immunopathology 8, 387–400.
Dunier, M., Vergnet, C., Siwicki, A. K. & Verlhac, V. (1995). E#ect of lindane exposureon rainbow trout (Oncorhynchus mykiss) immunity. IV. Prevention of non-specific and specific immunosuppression by dietary vitamin C (ascorbate-2-polyphosphate). Ecotoxicology & Environmental Safety 30, 259–268.
Ellis, A. E. (1990). Lysozyme assays. In Techniques in Fish Immunology (J. S. Stolen,T. C. Fletcher, D. P. Anderson, B. S. Roberson & W. B. Muiswinkel, eds)pp. 101–103. U.S.A.: SOS Publications.
Engstad, R. E., Robertsen, B. & Frivold, E. (1992). Yeast glucan induces increase inlysozyme and complement-mediated haemolytic activity in Atlantic salmonblood. Fish & Shellfish Immunology 2, 287–297.
Erdal, J. L., Evensen, O., Kaurstad, O. K., Lillehaug, A., Solbakken, R. & Thorud, K.(1991). Relationship between diet and immune response in Atlantic salmon(Salmo salar L.) after feeding various levels of ascorbic acid and omega-3 fattyacids. Aquaculture 98, 363–379.
Hamuro, J. & Chihara, G. (1984). Lentinan, a T-cell oriented immunopotentiator: itsexperimental and clinical applications and possible mechanism of immunomodu-lation. In Immune Modulation Agents and their Mechanisms (R. L. Fenichel &M. A. Chirigos, eds) pp. 409–436. New York and Basel: Marcel Dekker, Inc.
Hardie, L. J., Fletcher, T. C. & Secombes, C. J. (1991). The e#ect of dietary vitamin C onthe immune response of the Atlantic salmon (Salmo salar). Aquaculture 95,201–214.
Jørgensen, J. B., Sharp, G. J. E., Secombes, C. J. & Robertsen, B. (1993a). E#ect of ayeast–cell-wall glucan on the bactericidal activity of rainbow trout macro-phages. Fish & Shellfish Immunology 3, 267–277.
Jørgensen, J. B., Lunde, H. & Robertsen, B. (1993b). Peritoneal and headkidney cellresponse to intraperitoneally injected yeast glucan in Atlantic salmon, Salmosalar L. Journal of Fish Diseases 16, 313–325.
Lall, S. P. & Olivier, G. (1993). Role of micronutrients in immune response and diseaseresistance in fish. In Fish Nutrition in Practice, Vol. 61 (INRA, ed.) pp. 101–118.Les Colloques.
Li, Y. & Lovell, R. T. (1985). Elevated levels of dietary ascorbic acid increase immuneresponses in channel catfish. Journal of Nutrition 115, 123–131.
Mathews, E. S., Warinner, J. E. & Weeks, B. A. (1990). Assays of immune functions infish macrophages: techniques used as indicators of environmental stress. InTechniques in Fish Immunology (J. S. Stolen, T. C. Fletcher, D. P. Anderson, B.S. Roberson & W. B. van Muiswinkel, eds) pp. 155–163. U.S.A.: SOS Publications.
Navarre, O. & Halver, J. E. (1989). Disease resistance and humoral antibody productionin rainbow trout fed high levels of vitamin C. Aquaculture 79, 207–221.
Nikl, L., Evelyn, T. P. T. & Albright, L. J. (1993). Trials with an orally andimmersion-administered beta-1,3 glucan as an immunoprophylactic againstAeromonas salmonicida in juvenile chinook salmon Oncorhynchus tshawytscha.Disease of Aquatic Organisms 17, 191–196.
Noya, M., Magarinos, B. & Lamas, J. (1995). La administracion intraperitoneal y oralde glucano afecta al sistema inmune no especifico y a la resistencia de la dorada,Sparus aurata, a la pasteurelosis. In Proceedings of the Fifth National Congressof Aquaculture (F. Castello & A. Caldere, eds) pp. 734–738. Sant Carles de laRapita, Spain, May 1995.

IMMUNOMODULATION BY DIETARY VITAMIN C 423
Ogier de Baulny, M., Quentel, C., Fournier, V., Lamour, F. & Le Gouvello, R. (1996).E#ect of long-term oral administration of beta-glucan as an immunostimulant oran adjuvant on some non-specific parameters of the immune response of turbotScophthalmus maximus. Diseases of Aquatic Organisms 26, 139–147.
Plytycz, B., Flory, C. M., Galvan, I. & Bayne, C. T. (1989). Leukocytes of rainbow trout(Oncorhynchus mykiss) pronephros: cell types producing superoxide anion.Developmental and Comparative Immunology 13, 217–224.
Raa, J., Rørstad, G., Engstad, R. & Robertsen, B. (1992). The use of immunostimulantsto increase resistance to aquatic organisms to microbial infections. In Diseasesin Asian Aquaculture (M. I. Shari#, R. P. Subasinghe & J. R. Arthur, eds)pp. 39–50. Fish health section, Asian Fisheries Society, Manila, Philippines.
Roberts, M. L., Davies, S. J. & Pulsford, A. L. (1995). The influence of ascorbic acid(vitamin C) on non-specific immunity of turbot (Scophthalmus maximus L.). Fish& Shellfish Immunology 5, 27–38.
Robertsen, B., Rørstad, G., Engstad, R. & Raa, J. (1990). Enhancement of non-specific disease resistance in Atlantic salmon, Salmo salar L. by a glucan fromSaccharomyces cerevisiae cell walls. Journal of Fish Diseases 13, 391–400.
Rørstad, G., Aasjord, P. M. & Robertsen, B. (1993). Adjuvant e#ect of a yeast glucan invaccines against furunculosis in Atlantic salmon (Salmo salar L.). Fish &Shellfish Immunology 3, 179–190.
Scott, A. L., Rogers, W. A. & Klesius, P. H. (1985). Chemiluminescence by peripheralblood phagocytes from channel catfish: function of opsonin and temperature.Developmental and Comparative Immunology 9, 241–250.
Siwicki, A. K., Anderson, D. P. & Rumsey, G. L. (1994). Dietary intake of immuno-stimulants by rainbow trout a#ects non-specific immunity and protects againstfurunculosis. Veterinary Immunology & Immunopathology 41, 125–139.
Stave, J. W., Roberson, B. S. & Hetrick, F. M. (1983). Chemiluminescence of phagocyticcells isolated from the pronephros of striped bass. Developmental and Compara-tive Immunology 7, 269–276.
Vergnet, C. & Dunier, M. (1993). Enzyme-linked immunosorbent assay (ELISA) toquantify rainbow trout (Oncorhynchus mykiss) antibodies specific to Yersiniaruckeri. Fish Disease Diagnostics Prev. Meth., 121–134.
Verlhac, V. & Gabaudan, J. (1992). E#ect of a high dietary dose of ascorbate-2-monophosphate on the immune response of rainbow trout (Oncorhynchusmykiss). In Ascorbic Acid in Domestic Animals (C. Wenk, R. Fenster & L. Völker,eds) pp. 456–461. Proceedings of the 2nd symposium, 9–12 October 1990, KartauseIttingen, Switzerland.
Verlhac, V., N’Doye, A., Gabaudan, J., Troutaud, D. & Deschaux, P. (1993). Vitaminnutrition and fish immunity: influence of antioxidant vitamins (C and E) onimmune response of rainbow trout (Oncorhynchus mykiss). In Fish Nutrition inPractice (INRA, ed.) pp. 167–177. Les Colloques.
Verlhac, V. & Gabaudan, J. (1994). Influence of vitamin C on the immune system ofsalmonids. Aquaculture Fisheries Management 25, 21–36.
Verlhac, V., Gabaudan, J. & Schüep, W. (1995). Immunomodulation in fish: II. E#ect ofdietary vitamin C. In Proceedings of the 2nd Roche Aquaculture Centre Confer-ence on Nutrition and Disease, 15 June 1995 (K. Kurmaly, ed.), Bangkok,Thailand.
Verlhac, V., Gabaudan, J., Obach, A., Schüep, W. & Hole, R. (1996). Influence of dietaryglucan and vitamin C on non-specific and specific immune response of rainbowtrout (Oncorhynchus mykiss). Aquaculture 143, 123–133.
Vuilleumier, J. P. & Keck, E. (1989). Fluorometric assay of vitamin C in biologicalmaterials using a centrifugal analyser with fluorescence attachment. Journal ofMicronutrient Analysis 5, 25–34.
Waagbø, R., Glette, J., Nilsen, E. R. & Sandnes, K. (1993). Dietary vitamin C, immunityand disease resistance in Atlantic salmon (Salmo salar). Journal of FishPhysiology & Biochemistry 12, 61–73.

424 V. VERLHAC ET AL.
Waagbø, R. (1994). The impact of nutritional factors on the immune system in Atlanticsalmon, Salmo salar L.: a review. Aquaculture and Fisheries Management 25,175–197.
Wahli, T., Verlhac, V., Gabaudan, J., Meier, W. & Schüep, W. (1998). Influence ofcombined vitamins C and E on immunity and disease resistance of rainbow troutOncorhynchus mykiss. Journal of Fish Diseases 21, 127–137.
Wang, X. Y., Liao, M. L., Hung, T. H. & Seib, P. A. (1988). Liquid chromatographydetermination of L-ascorbate 2-polyphosphate in fish feeds by enzymatic releaseof L-ascorbate. Journal of the Association of Official Analytical Chemists 71(6),1158–1161.
Yano, T. (1992). Assays of hemolytic complement activity. In Techniques in FishImmunology (J. S. Stolen, T. C. Fletcher, D. P. Anderson, S. L. Kaattari & A. F.Rowley, eds) pp. 131–141. U.S.A.: SOS Publications.
Yoshida, T., Kruger, R. & Inglis, V. (1995). Augmentation of non-specific protection inAfrican catfish, Claria gariepinus (Burchell), by the long-term oral admin-istration of immunostimulants. Journal of Fish Diseases 18, 195–198.
Sakai, D. K. (1981). Heat inactivation of complements and immune hemolysis reactionsin rainbow trout, masu salmon, coho salmon, goldfish and tilapia. Bull. Jap. Soc.Scient. Fish. 47, 565–571.
Wolf, K. (1963). Physiological salines for fresh-water teleosts. Progressive FishCulturist 25, 135–140.
Copyright © 2022 FDOKUMEN




















