
Immunogold localization of a transglutaminase related to grana development in different maize cell...
9
Protoplasma (2001) 216:155-163 PROTOPLASMA Springer-Verlag 2001 Printed in Austria Immunogold localization of a transglutaminase related to grana development in different maize cell types E. Villalobos, J. M. Torn~, J. Rigau, I. OIl~s, I. Claparols, and M. Santos* Departament de Genetica Molecular, Institut de Biologia Molecular de Barcelona, Consejo Superior de Investigaciones Cientificas, Barcelona Received October 12, 2000 Accepted December 27, 2000 Summary. A comparative study of the subcellular localization of a plant transglutaminase (TGase; EC 2.3.2.13) in various in vivo and in vitro maize cell types was carried out with a polyclonal antibody raised against a 58 kDa TGase purified from Helianthus tubero- sus leaves. Immunocytochemical staining, followed by electron microscopy, showed that this enzyme was markedly present in the grana-appressed thylakoids of mature chloroplasts of the light- exposed cells. Moreover, during embryogenic callus chloroplast differentiation, the abundance of TGase in the grana-appressed thy- lakoids depended on the degree of grana development and was greater than in mature leaf chloroplasts. In addition to the 58 kDa form, two other forms of the protein (of 77 and 34 kDa) were obtained by Western blot. The 77 kDa form might correspond to the inactive form and was immunodetected in dense vesicles observed in dark-grown embryogenic callus cells. In adult leaves, the enzyme was also markedly present in the grana-appressed thylakoids of the mesophyll cell chloroplasts, though very scarce and dispersed in the bundle-sheath cell chloroplasts (which do not contain grana). The concordance of these localizations with those described for the light-harvesting antenna proteins of the photosystem II suggests that it is possible that this TGase has a functional role in photosyn- thesis, perhaps modulating the photosynthetic efficiency and the absorption of excess light by means of polyamine conjugation to the antenna proteins. Keywords: Chloroplast; Embryogenic callus; Grana; Maize; Photo- system II; Transglutaminase. Abbreviations: 2,4-D 2,4-dichlorophenoxyacetic acid; LHCPII photosystem II light-harvesting complex; PSI photosystem I; PSII photosystemII; TGase R-glutaminyl-peptide:amine-y-glu- tamyltransferase. * Correspondence and reprints: Departament de Genetica Molecu- lar, Institut de Biologia Molecular de Barcelona, Consejo Superior de Investigaciones Cientificas, Jordi Girona 18-26, 08034 Barcelona, Spain. Introduction Chloroplast differentiation is critical in determining the photosynthetic capacity of a plant. In green-plant chloroplasts, the inner membrane system consists of numerous flattened sacs, the thylakoids, differentiated into two structural domains: stacked grana thyla- koids (appressed) and connecting stroma-exposed thy- lakoids (nonappressed). Regular stacking of the thylakoids in the grana is characteristic of mature chloroplasts in angiosperms and gymnosperms (Mura- kami 1991). The primary photoreaction and electron transport are built into the thylakoid membrane system. Their constituent components are organized into four complexes: photosystem I (PSI), photosys- tem I1 (PSII), light-harvesting chlorophyll-protein complex (LHCP), and cytochrome complex. It has been established that, in chloroplasts with grana, PS II is located preferentially in appressed thylakoids and PSI in the stroma-exposed nonappressed thylakoids (Anderson 1987). In C4 plants (such as maize) there is a structural tissue differentiation between the photosynthetic cells: the chloroplasts of mesophyll cells contain grana, while those in bundle-sheath cells do not. Bundle- sheat cell chloroplasts are poor in PS II in relation to PSI (Ku et al. 1974) and they contain fewer LHCPII polypeptides than those of mesophyll cells (Schuster etal. 1985, Bassi etal. 1987). Sheen and Bogorad (1986) have partially characterized cDNA clones spe- cific to six LHCPII genes of maize. Three of these genes are preferentially expressed in mesophyll cells and their mRNAs constitute about 54% of the total LHCPII transcripts in greening maize leaves.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Immunogold localization of a transglutaminase related to grana development in different maize cell...

Protoplasma (2001) 216:155-163 PROTOPLASMA �9 Springer-Verlag 2001 Printed in Austria
Immunogold localization of a transglutaminase related to grana development in different maize cell types
E. Villalobos, J. M. Torn~, J. Rigau, I. OIl~s, I. Claparols, and M. Santos*
Departament de Genetica Molecular, Institut de Biologia Molecular de Barcelona, Consejo Superior de Investigaciones Cientificas, Barcelona
Received October 12, 2000 Accepted December 27, 2000
Summary. A comparative study of the subcellular localization of a plant transglutaminase (TGase; EC 2.3.2.13) in various in vivo and in vitro maize cell types was carried out with a polyclonal antibody raised against a 58 kDa TGase purified from Helianthus tubero-
sus leaves. Immunocytochemical staining, followed by electron microscopy, showed that this enzyme was markedly present in the grana-appressed thylakoids of mature chloroplasts of the light- exposed cells. Moreover, during embryogenic callus chloroplast differentiation, the abundance of TGase in the grana-appressed thy- lakoids depended on the degree of grana development and was greater than in mature leaf chloroplasts. In addition to the 58 kDa form, two other forms of the protein (of 77 and 34 kDa) were obtained by Western blot. The 77 kDa form might correspond to the inactive form and was immunodetected in dense vesicles observed in dark-grown embryogenic callus cells. In adult leaves, the enzyme was also markedly present in the grana-appressed thylakoids of the mesophyll cell chloroplasts, though very scarce and dispersed in the bundle-sheath cell chloroplasts (which do not contain grana). The concordance of these localizations with those described for the light-harvesting antenna proteins of the photosystem II suggests that it is possible that this TGase has a functional role in photosyn- thesis, perhaps modulating the photosynthetic efficiency and the absorption of excess light by means of polyamine conjugation to the antenna proteins.
Keywords: Chloroplast; Embryogenic callus; Grana; Maize; Photo- system II; Transglutaminase.
Abbreviations: 2,4-D 2,4-dichlorophenoxyacetic acid; LHCPII photosystem II light-harvesting complex; PSI photosystem I; PSII photosystemII; TGase R-glutaminyl-peptide:amine-y-glu- tamyltransferase.
* Correspondence and reprints: Departament de Genetica Molecu- lar, Institut de Biologia Molecular de Barcelona, Consejo Superior de Investigaciones Cientificas, Jordi Girona 18-26, 08034 Barcelona, Spain.
Introduction
Chloroplast differentiation is critical in determining the photosynthetic capacity of a plant. In green-plant chloroplasts, the inner membrane system consists of numerous flattened sacs, the thylakoids, differentiated into two structural domains: stacked grana thyla- koids (appressed) and connecting stroma-exposed thy- lakoids (nonappressed). Regular stacking of the thylakoids in the grana is characteristic of mature chloroplasts in angiosperms and gymnosperms (Mura- kami 1991). The primary photoreaction and electron transport are built into the thylakoid membrane system. Their constituent components are organized into four complexes: photosystem I (PSI), photosys- tem I1 (PSII), light-harvesting chlorophyll-protein complex (LHCP), and cytochrome complex. It has been established that, in chloroplasts with grana, PS II is located preferentially in appressed thylakoids and PSI in the stroma-exposed nonappressed thylakoids (Anderson 1987).
In C4 plants (such as maize) there is a structural tissue differentiation between the photosynthetic cells: the chloroplasts of mesophyll cells contain grana, while those in bundle-sheath cells do not. Bundle- sheat cell chloroplasts are poor in PS II in relation to PSI (Ku et al. 1974) and they contain fewer LHCPII polypeptides than those of mesophyll cells (Schuster etal. 1985, Bassi etal. 1987). Sheen and Bogorad (1986) have partially characterized cDNA clones spe- cific to six LHCPII genes of maize. Three of these genes are preferentially expressed in mesophyll cells and their mRNAs constitute about 54% of the total LHCPII transcripts in greening maize leaves.

156 E. Villalobos et al.: Immunolocalization of transglutaminase in maize
Transglutaminases (TGases, R-glutaminyl-peptide: amine-7-glutamyltransferases; EC 2.3.2.13) were de- tected for the first time in animals. These enzymes catalyze the covalent linkage of substrates with primary amino groups, such as polyamines, to a 7- carboxamide group of protein endo-glutamine res- idues. In animals, these enzymes posttranslationally modify structural proteins (Folk 1980).
The first observation of TGase-like activity in plants was in cycling cells and etiolated apices of Helianthus tuberosus (Serafini-Fracassini et al. 1988, 1989). In unopened Medicago sativa flower buds, ribulose-l,5- biphosphate carboxylase/oxygenase (Rubisco) has been found to constitute a major substrate (Margosiak et al. 1990). Del Duca et al. (1994) identified endoge- nous substrates of TGase in chloroplasts of Helianthus tuberosus leaves. The thylakoid proteins incorporated into polyamines by the action of TGase activity were recognized by polyclonal antibodies as apoproteins of the antenna complex (LHCPII, CP24, CP26, and CP29 proteins). Moreover, these authors suggested that a TGase should be located in the chloroplast membranes on the basis of a cross-reaction obtained with animal TGase antibodies and thylakoid proteins, where they obtained a unique TGase band of 58 kDa. However, the specific localization of the enzyme within the thylakoid membrane system has not, until now, been studied.
Our group has also detected a TGase with the same molecular mass (58 kDa) in maize green meristematic calluses and their chloroplasts with an animal TGase antibody. This TGase was light sensitive and affected by hormone deprivation (Bernet et al. 1999).
In the light of these previous investigations, and taking into account that the relationship between this enzyme and the chloroplast differentiation process should be crucial in elucidating the possible function of TGase in photosynthesis, the localization of this TGase by immunoelectron microscopy in various in vivo and in vitro maize cell types, including different stages of chloroplast development, was examined. Moreover, the ultrastructural data were comple- mented by the analysis of the possible enzyme iso- forms which are recognized by Western blot in the same cell types.
Material and methods
Plant material
Root and young-leaf samples were taken from germinated maize plantlets grown for 7 days under cool-white fluorescent lights
(90-100 I.tmol/m 2. s) with a 16 h light and 8 h dark photoperiod (roots and young leaves) or under dark conditions (roots). Mature- leaf samples were taken from adult plants grown for two months in a greenhouse (sunlight plus alternate Gro-lux and sodium vapor lamps).
Green-callus samples were obtained from long-time cultured meristematic calluses growing on proliferation medium in the pres- ence of 9 I.tM 2,4-dichloropfienoxyacetic acid (2,4-D) as growth regulator, in the previously reported conditions (Torn6 et al. 1984) and in the same photoperiod and light intensity conditions as the plantlets.
Embryogenic-tissue samples were obtained from embryogenic calluses (proceeding from immature embryos) cultured on prolifer- ation medium (Santos et al. 1993) in the presence of 9 gM 2,4-D, under cool-white fluorescent lights for 16 h light and 8 h dark or under dark conditions.
Differentiated-embryo samples were taken from somatic embry- oids obtained from 30-day-old embryogenic calluses cultured in pro- liferation medium. Samples were taken after 12 h, 24 h, and 8 days of culture on differentiation medium (half-concentrated Murashige and Skoog [1962] basal salts and organic components and no growth regulators), under a light intensity of 90-100 p.mol/m 2. s (strong light) and the same photoperiod conditions as above.
Western analyses were realized with samples of these embryo- genic calluses, exposed to different conditions: (I) calluses cultured always in dark conditions; (2) callus submitted for 4 h to strong light (90-100 I.tmol/m 2 . s); (3) calluses cultured for 30 days under weak light (5-10 p.mol/m 2. s); (4) calluses cultured for 200 days under weak light; (5) calluses proceeding from condition 4 and cultured for 40 days in dark conditions (etiolated callus); (6) etiolated calluses cultured for 4 h under strong-light conditions; (7) callus proceeding from dark conditions subcultured on differentiation medium for 12 h, 24 h, and 8 days under strong light; (8) etiolated calluses subcultured on differentiation medium and the same con- ditions as 7.
Within the embryogenic-callus samples used in Western blot analyses, only those most representative were processed for immu- noelectron microscopy observations. These samples resulted from treatments 1, 3, 4, 7, and 8.
Antibody production and specificity determination
A polyclonal antibody obtained in collaboration with the group of Serafini-Fracassini (Dondini 1998) from chicken with a purified chloroplast TGase of Helianthus tuberosus leaves as antigen was used to immunolocalize the enzyme. The antibody was obtained by inoculating diluted antigen into the chicken, extracting it from the yolk egg, and purifying it with 100 mM phosphate-buffered saline (PBS) (100 mM phosphate buffer and 0.1 M NaC1, pH7.5) plus 5.25 % polyethylene glycol 8000.
The antibody specificity was determined by Western blot with purified TGase protein (Dondini 1998). It was also titrated by Western blot as described below.
Moreover, the possible inhibition of TGase activity by addition of diluted antibody to the maize leaf protein extract in the reaction mixture was measured as described below.
Transglutaminase assay and enzymatic-activity inhibition
The enzymatic assays were performed under the same light condi- tions as those prevailing at the time of the sample collection. Leaf samples were extracted as indicated in Del Duca et al. (1994). Chloroplasts, thylakoids, and grana membranes were isolated as indicated in Berthold et al. (1981), using 50 mM HEPES instead of morpholineethanesulfonic acid in the thylakoid and grana fractions. The pH of the incubation mixture was 8.0. The enzymatic mixture

E. Villalobos et al.: Immunolocalization of transglutaminase in maize
included: 100 gl of supernatant (containing about 600-700 gg of total proteins) and 100 gl of 20 mM Tris buffer, pH 8, containing 0.6 mM putrescine, 9 mM 2-J3-mercaptoethanol, and 185 kBq [1,4- 3H]putrescine (0.85 TBq/mmol) (Amersham, Little Chalfont, U.K.) or 100 gl of 20 mM Tris buffer containing 3 mM CaC12 and 660 mM sorbitol when chloroplasts were used. After 30 vain of incubation at 30~ the reaction was blocked by adding 300gl of 10% trichloroacetic acid containing 2mM putrescine. Samples were repeatedly precipitated and the radioactivity was measured as described by Del Duca et al. (1994). To study the inhibition of the enzymatic activity by the inhibition of the PS II, 6.5 gM Diurdn (3-(3,4-dichlorophenyl)-l,l-dimethylurea; Sigma-Aldrich, Madrid, Spain) was applied to each cell fraction.
The inhibition of TGase activity by the plant antibody was mea- sured by adding different concentrations of diluted antibody to the maize leaf protein extracts in the enzymatic mixture. In the experi- ments concerning the influence of light, auxin (2,4-D) and culture times on embryogenic callus TGase activity, samples were extracted as indicated in Bernet et al. (1999).
Electrophoresis and Western immunoblotting
Western immunoblotting was performed according to the method of Towbin et al. (1979). Protein transfer was realized with a Trans Blot SD (Bio-Rad). Proteins adsorbed onto a nitrocellulose filter were blocked overnight in 5% nonfat milk diluted in PBS. After blocking, the sheet was incubated for 1 h with the chicken anti- TGase in different dilutions in PBS. The nonspecifically bound prod- ucts were removed by three 5 min washings in PBS, after which the nitrocellulose sheet was incubated for 1 h with a peroxidase- conjugated donkey anti-chicken immunoglobulin (IgY) as sec- ondary antibody (Jackson Immunoresearch Laboratories Inc., West Grove Pa., U.S.A.) diluted 1 : 6700 in PBS. The antigen-antibody complex was identified with the electrochemiluminescence Western blotting detection reagents (Amersham Pharmacia Biotech) accord- ing to the manufacturer's instructions.
Immunoelectron microscopy
Root and leaf slices, with a thickness less than 0.5 mm, as well as callus and embryo slices, were fixed by vacuum infiltration with 4% paraformaldehyde and 0.5% glutaraldehyde in 0.1M phosphate buffer (pH 7.4) for 2 h at room temperature. After washing with 0.1M phosphate buffer, samples were dehydrated through an ethanol series and embedded in Lowicryl K4M resin. Blocks were polymerized under a UV lamp at -20 ~ for 24 h. Ultrathin sections were mounted on gold grids (200 meshes).
The sections were treated with blocking solution (1% bovine serum albumin in PBS) for 15 min and were then incubated for 2 h at room temperature with different dilutions of the TG{tse antibody in blocking solution (1 : 4000, 1 : 8000, 1 : 16000). Antibody solution proceeded from an original stock containing 20 mg of antibody per ml of distilled water. After washing with blocking solution, sections were incubated for 1 h at room temperature in a solution of 12nm diameter colloidal gold-affinipure donkey anti-chicken IgY (Jackson Immunoresearch) diluted at a ratio 1 : 30 in the blocking solution. The sections were then washed with saline buffer and distilled water and subsequently stained with 2% uranyl acetate and lead citrate solution.
Moreover, as controls, some samples were treated only with blocking solution or with preimmune serum, instead of the antibody, following the same protocol as above.
After staining, all sections were examined with a transmission electron microscope (Hitachi H 600) operated at 80 kV.
To quantify the observed differences, the number of gold parti- cles per an area of 1/.tm 2 of appressed and nonappressed thylakoids
157
of a certain number of samples in the different chloroplast cell types was calculated.
Results
Antibody specificity and Western blot analyses
T h e spec i f ic i ty o f t h e a n t i b o d y u s e d was t e s t e d by
W e s t e r n i m m u n o b l o t t i n g a n d in an e n z y m a t i c T G a s e
assay, w i t h m a i z e m a t u r e - l e a f p r o t e i n ext rac ts . T h e
i m m u n o l o g i c a l p r o b e s r e c o g n i z e d a b a n d at a m o -
l e c u l a r mass o f 58 k D a , c o r r e s p o n d i n g to T G a s e
(Fig. 1 A ) , at a d i l u t i o n o f a n t i b o d y in b l o c k i n g
s o l u t i o n o f 1 : 10000 w i t h t h e o r ig ina l a n t i b o d y s tock
(20 m g / m l ) . In t h e e n z y m a t i c p r o t e i n assay, t h e add i -
t i o n o f 1 m g o f a n t i b o d y p e r ml o f t h e m a i z e l e a f
p r o t e i n ex t r ac t s in t h e r e a c t i o n m i x t u r e g ive a 6 8 %
i n h i b i t i o n o f T G a s e ac t iv i ty w i t h r e s p e c t t o t h e c o n t r o l
n o n i n h i b i t e d ext rac ts .
F u r t h e r m o r e , p r o t e i n ex t r ac t s o f t h e d i f f e r e n t
o r g a n s and e m b r y o g e n i c - c a l l u s cel l t ypes i n d i c a t e d
in M a t e r i a l a n d m e t h o d s w e r e a n a l y z e d by W e s t e r n
blot . T h r e e bands , at a p p r o x i m a t e l y 77, 58, a n d 34 k D a ,
A
B
Fig. 1 A, B. Western blot analysis of proteins from different tissues separated by 14% acrylamide gel electrophoresis (Laemmli 1970), using 1 : 10000 antibody dilution. The antibody cross-reaction was performed utilizing the plant anti-TGase antibody cited in the text as primary antibody. A Green-callus and organ protein extracts; green callus (gc), root (r), mature leaf (mll and ml2), young leaf (y/). B Embryogenic-callus protein extracts (1-8) from samples cultured under conditions 1-8

158 E. Villalobos et al.: Immunolocalization of transglutaminase in maize
were detected depending on the analyzed tissue. Figure 1A shows the presence of the isoforms in the different organs. Root samples did not show any band in dark- or in light-grown conditions. Young-plant leaves showed only a 77 kDa band; finally, mature leaves always presented a 58 kDa band, except in some cases where a 34 kDa band appeared instead.
Figure 1 B shows the influence of light intensity and t reatment length on the embryogenic-callus samples. Embryogenic calluses grown in dark conditions pre- sented a predominant 77 kDa band (Fig. 1 B, lane 1). The same calluses subcultured 4 h under strong-light conditions showed the same 77 kDa band, accom- panied by a feeble 34 kDa band (Fig. 1B, lane 2). Embryogenic calluses grown for 30 days in weak light presented only the 77 kDa band (Fig. 1B, lane 3). After 200 days of culture in weak light, calluses pre- sented a predominant 58 kDa band (Fig. l B, lane 4), as in the case of adult leaf (Fig. 1 A). When these cal- luses were submitted to dark conditions, the 58 kDa band disappeared and the 77 kDa and the 34 kDa bands appeared (Fig. 1 B, lane 5). When these etiolated calluses were submitted for 4 h to strong-light condi- tions, the 58 kDa band was detected (Fig. IB, lane 6). A similar and repetitive band pattern was obtained with embryogenic calluses grown in differentiation medium for 12 h to 8 days under strong-light condi- tions (Fig. 1 B, lanes 7 and 8). Green calluses showed a unique band of 34 kDa (Fig. 1A).
Effect of PSII protein inhibition on TGase activity
In order to corroborate the results described below, the effect of the inhibition of TGase activity on the different membrane fractions treated with Diur6n was measured. This compound specifically inhibits the activity of the PSII proteins present in the grana membranes. The inhibition of TGase activity was 37 %, 50%, and 70% in the purified chloroplast, thylakoid, and noninhibited grana fractions, respectively.
Effect of light on the TGase activity of embryogenic calluses
The TGase activity (pmol of putrescine per mg protein per h) of embryogenic calluses after 0, 6, and 9 h of culture under strong- or weak-light conditions, as well as the effect of hormone deprivation are presented in Table 1. As can be seen, after 6 h of culture under light conditions, all the samples showed a significantly
Table 1. TGase activity expressed as putrescine incorporation in embryogenic calluses
Treatment of calluses a Amount of putrescine (pmol/h. mg protein) incorporated after the following culture period: b
Oh 6h 9h
wl + 9 gM 2,4-D 16+1.7 60.2+10.2b 81.7+8.3b sl + 9 gM 2,4-D 16+1.7 41.1+_9.5 a 132.4+5.7 a sl + 0 gM 2,4-D 16+1.7 34.9+10.5 a 119.5+4.6a
wl, weak light culture; sl, strong-light culture b Different letters in the same column indicate significant differences at 0.01 > P > 0.001 level
higher TGase activity with respect to the 0 h control samples, independent of light intensity. However, after 9 h of exposure to light, significant differences in enzy- matic activity between samples cultured under strong light and those cultured under weak light were obtained, independent of the presence of auxin.
Ultrastructural localization of TGase in maize tissues
Cells lacking chloroplasts
Ultrathin sections of light- and dark-grown roots indi- cated that there was no labelling with gold particles in both types of cells (not shown). Ultrathin sections of embryogenic calluses grown in maintenance medium and dark conditions showed that, in these cells, only a few proplastids were present. In these proplastids, there was no labelling with gold particles (Fig. 2A). However, in the same cell types, significant gold labelling was detected in certain types of spherical vesicles which were dispersed in the cytoplasm (Fig. 2A, inset). These dense vesicles, with a diameter of 200-700 nm, contained an electron-dense core in which gold particles are deposed and which is sur- rounded by an electron-translucent layer. The same type of vesicles were present in abundance near weak- light-cultured callus chloroplasts (data not shown). These vesicles were not detected in the other cell types
under examination.
Green calluses
All the chloroplasts of the green meristematic callus cells (cultured in proliferation medium under strong- light conditions) presented granal structures. In these chloroplasts, gold labelling was scarce (1 : 4000 anti-

E. Villalobos et al.: Immunolocalization of transglutaminase in maize 159
Fig. 2A-F. Electron microscopy of im- munolabelled sections of maize cal- luses cultured on different media and light intensities. A Proplastid and vesicle (inset; bar, 100 nm) of an embryogenic callus cell from treatment 1; B chloroplast of a green-callus cell; C general aspect of an immature chloroplast observed in the embryogenic-callus cells from treatments 3 or 4; D random distribution of gold labelling in the same chloroplast as in C; E grana-concentrated gold labelling of a mature chloroplast proceeding from the same tissue as C; F grana-concentrated gold labelling in a chloroplast of the embryoid-callus cells from treatments 7 or 8. chl Chloroplast, cw cell wall, cy cytosol, e Stroma, g grana, m mitochondria, p proplastid, s starch grain, t thylakoid, v vesicle. Arrows indicate gold particle deposition. Bars: A and C, 200 nm; B and DF, 100 nm
b o d y dilution) and restr icted to grana-appressed thylakoids (Fig. 2B).
Different ia t ing embryogen ic calluses and embryos
A t the end of the subcul ture per iod of embryogen ic cal-
luses cul tured in the prol i fera t ion m e d i u m under weak
light (condit ions 3 and 4), m a n y immatu re chloroplasts
wi thout normal grana were observed (Fig. 2 C). In these
chloroplasts, gold labelling was distr ibuted r andomly
and de tec ted in the thy lakoid- forming grana (Fig. 2 D).
Moreover , some chloroplasts with normal ly deve loped
grana structures were also observed. In these chloro-
plasts, gold labelling was specifically concen t ra ted in the thy lakoid-appressed grana (Fig. 2 E).
Af te r 12 h of embryo culture in the different iat ion
m e d i u m under strong-l ight condit ions (7 and 8),
chloroplasts deve loped mature grana and an abun-
dance of starch grains, as expected for a differentiat-
ing embryo. In the grana-appressed thylakoids of
these chloroplasts, abundan t label (using 1 : 4000 and
1 : 8000 an t ibody dilutions) was observed (not shown).

160 E. Villalobos et al.: Immunolocalization of transglutaminase in maize
Fig. 3A-F. Electron microscopy of im- munolabelled sections of young and adult maize plant leaves cultured in the condi- tions described in Material and methods. A Chloroplast of a young leaf; B randomly immunolabelled thylakoids of the same chloroplast; C general aspect of mesophyll and bundle-sheat cell chloroplasts of mature leaf; D scarce immunolabelling of thylakoids in a bundle-sheat cell chloro- plast of a mature leaf; E absence of gold labelling in the chloroplast of a section of a mature leaf treated with preimmune serum; F grana-concentrated immunola- belling in a mesophyll cell chloroplast of a mature leaf. BSC Bundle sheat cell, M C
mesophyll cell, chl chloroplast, cw cell wall, cy cytosol, e stroma, g grana, s starch grain, t thylakoid, Arrows indicate gold particle deposition. Bars: A, 0.5 gm; B, D, and F, 50 nm; C, 1 gm; E, 100 nm
The same chloroplast morphology and gold labelling specificity was observed in embryos cultured 24 h and 7 days in the differentiation medium under strong- light conditions (Fig. 2F). As expected, C4 morphol- ogy was not present in 8-day-differentiating embryos.
Leafs
The chloroplast morphology of the 7-day-germinated plantlet leaves is presented in Fig. 3 A. In these chloro- plasts, thylakoids were not appressed to form grana
structures and gold labelling was observed only in a few cases (Fig. 3 B) with the highest antibody concen- tration (1 : 4000).
In the leaves of mature plants (60-day-old plants), the morphology of mesophyll and bundle-sheat cell chloroplasts was perfectly differentiated (see Fig. 3 C). In grana-less bundle-sheat cell chloroplasts, label was scarce (1:4000 antibody dilution) (Fig. 3D). However, abundant gold particles were detected specifically in grana-appressed thylakoids of the mesophyll cell chloroplasts at 1:4000, 1:8000, and
E. Villalobos et al.: Immunolocalization of transglutaminase in maize 161
21 5 0 -
,~ 40 e ~ 3oj
20
10
yl mlm mlb gc ecn ecdl ecd2
cell type
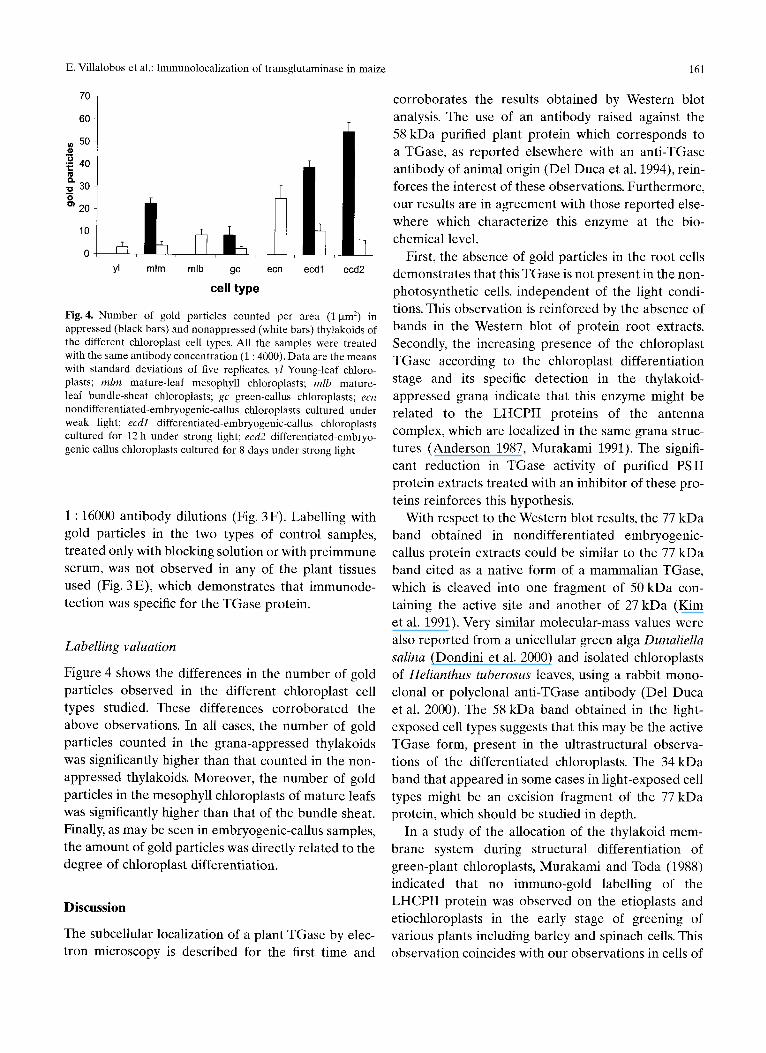
Fig. 4. Number of gold particles counted per area (1 gm 2) in appressed (black bars) and nonappressed (white bars) thylakoids of the different chloroplast cell types. All the samples were treated with the same antibody concentration (1 : 4000). Data are the means with standard deviations of five replicates, yl Young-leaf chloro- plasts; mira mature-leaf mesophyll chloroplasts; mlb mature- leaf bundle-sheat chloroplasts; gc green-callus chloroplasts; ecn nondifferentiated-embryogenic-callus chloroplasts cultured under weak light; ecdl differentiated-embryogenic-callus chloroplasts cultured for 12 h under strong light; ecd2 differentiated-embryo- genic-callus chloroplasts cultured for 8 days under strong light
1:16000 antibody dilutions (Fig. 3F). Labelling with gold particles in the two types of control samples, treated only with blocking solution or with preimmune serum, was not observed in any of the plant tissues used (Fig. 3 E), which demonstrates that immunode- tection was specific for the TGase protein.
Labelling valuation
Figure 4 shows the differences in the number of gold particles observed in the different chloroplast cell types studied. These differences corroborated the above observations. In all cases, the number of gold particles counted in the grana-appressed thylakoids was significantly higher than that counted in the non- appressed thylakoids. Moreover, the number of gold particles in the mesophyll chloroplasts of mature leafs was significantly higher than that of the bundle sheat. Finally, as may be seen in embryogenic-callus samples, the amount of gold particles was directly related to the degree of chloroplast differentiation.
D i s c u s s i o n
The subcellular localization of a plant TGase by elec- tron microscopy is described for the first time and
corroborates the results obtained by Western blot analysis. The use of an antibody raised against the 58 kDa purified plant protein which corresponds to a TGase, as reported elsewhere with an anti-TGase antibody of animal origin (Del Duca et al. 1994), rein- forces the interest of these observations. Furthermore, our results are in agreement with those reported else- where which characterize this enzyme at the bio- chemical level.
First, the absence of gold particles in the root cells demonstrates that this TGase is not present in the non- photosynthetic cells, independent of the light condi- tions. This observation is reinforced by the absence of bands in the Western blot of protein root extracts. Secondly, the increasing presence of the chloroplast TGase according to the chloroplast differentiation stage and its specific detection in the thylakoid- appressed grana indicate that this enzyme might be related to the LHCPII proteins of the antenna complex, which are localized in the same grana struc- tures (Anderson 1987, Murakami 1991). The signifi- cant reduction in TGase activity of purified PSII protein extracts treated with an inhibitor of these pro- teins reinforces this hypothesis.
With respect to the Western blot results, the 77 kDa band obtained in nondifferentiated embryogenic- callus protein extracts could be similar to the 77 kDa band cited as a native form of a mammalian TGase, which is cleaved into one fragment of 50 kDa con- taining the active site and another of 27 kDa (Kim et al. 1991). Very similar molecular-mass values were also reported from a unicellular green alga Dunaliella salina (Dondini et al. 2000) and isolated chloroplasts of Helianthus tuberosus leaves, using a rabbit mono- clonal or polyclonal anti-TGase antibody (Del Duca et al. 2000). The 58 kDa band obtained in the light- exposed cell types suggests that this may be the active TGase form, present in the ultrastructural observa- tions of the differentiated chloroplasts. The 34 kDa band that appeared in some cases in light-exposed cell types might be an excision fragment of the 77 kDa protein, which should be studied in depth.
In a study of the allocation of the thylakoid mem- brane system during structural differentiation of green-plant chloroplasts, Murakami and Toda (1988) indicated that no immuno-gold labelling of the LHCPII protein was observed on the etioplasts and etiochloroplasts in the early stage of greening of various plants including barley and spinach cells. This observation coincides with our observations in cells of

162 E. Villalobos et al.: immunolocalization of transglutaminase in maize
dark-grown maize tissues immunolabelled with the plant TGase antibody. In the cited experiment, after 4 h of illumination, small amounts of gold particles were found randomly in the thylakoid membrane system, in which the differentiation of grana and stroma thylakoids was initiated. This also coincides with our observations in the immature chloroplasts of maize embryogenic calluses cultured under weak light intensity and in young maize leaves (Figs. 2-4). Finally, as in LHCII immuno-labelled spinach chloroplasts, as the grana formation in our maize cells proceeds (dif- ferentiated embryos and adult leaves), the number of gold particles on the thylakoids increases significantly and they are concentrated on grana. Moreover, the increase in TGase presence in relation to chloro- plast differentiation, together with its significant presence in mesophyll chloroplasts of the maize C4- photosynthesis cell system, reinforces the relation between this TGase and the presence of grana. From these observations, it may be stated that the presence of the chloroplast TGase is concomitant with that of the PS II in the greening process and grana formation.
It has been shown that when the thylakoid mem- brane system commences (after 4-6 h of etioplast illumination), electron transport from H20 to NADP + becomes measurable and PS II becomes connected to PSI (Welburn 1980). The maturation of PSI and PS II complexes seems to be a prerequisite to the structural and functional events leading to grana differentiation in the thylakoid membrane system. Furthermore, phosphorylation of the major LHCPII complex is responsible for the transition from state I to state II (Bennett 1983), while another phosphorylation mech- anism induces the protection of the PSII reaction center from photoinhibition (Dainese et al. 1992). It has been demonstrated that TGase activity can be induced during the greening of an initially non- photosynthetically explant (Del Duca et al. 1993). Moreover, this activity increased at the beginning of the light period in maize green organogenic calluses (Bernet et al. 1999) and in embryogenic calluses (pre- sented here).
With respect to the gold particles present in the spherical dense vesicles observed only in the embryo- genic nondifferentiated calluses, their size and struc- ture indicate that these vesicles may correspond to the same type of transport vesicles observed in storage cells of various species such as wheat (Levanony et al. 1992), castor bean (Hara-Nishimura et al. 1998), and
pumpkin (Hayasi et al. 1999). In all these cases, vesi- cles are Golgi mediated and endoplasmic reticulum derived and contain the precursor form of a storage protein which is transported in these vesicles to protein storage vacuoles, where it is processed prote- olytically by a vacuolar processing enzyme to generate the mature form (Hara-Nishimura and Nishimura 1987). In our case, the presence of TGase (probably in its inactive form) in these vesicles which are observed only in the callus cells that do not have differentiated chloroplasts should be studied in order to relate this observations with the chloroplast differentiation process.
Although the physiological role of polyamine- binding chloroplast proteins is still unknown, the spe- cific localization of the enzyme which is responsible for this binding in the grana apparatus, where the an- tenna complex is also located, reinforces the hypothe- sis that this enzyme is related to the photosynthetic process.
These results also reinforced the biochemical iden- tification of the thylakoid chlorophyll proteins (in particular the LHCPII protein antenna complex) as substrates of endogenous TGase in isolated chloro- plasts of Helianthus tuberosus leaves with antibodies against TGases of animal origin (Del Duca et al. 1994). As has been reported of various tissues of other organ- isms, i.e., keratinocyte TGase (Rice et al. 1992), our results demonstrate at the electron-microscopic level that the chloroplast TGase is specifically located in the thylakoid-appressed grana, perhaps with the active site exposed to the stroma, as suggested by its pH optimum (Del Duca et al. 1994).
The specific subcellular localization of chloroplast TGase in the thylakoid-appressed grana of the light- exposed maize tissues presented here emphasizes the possible implication of this enzyme in photosynthesis, as has been hypothesized by other authors, and complements the previously published results. The increased presence of the enzyme throughout chloro- plast maturation adds new data to its implication in these processes. This implication could be related to the modulation of the photosynthetic efficiency as a protective system which regulates the absorption of excess light by polyamine conjugation to the LHCPII proteins or other antenna proteins.
Molecular studies of the chloroplast TGase to elu- cidate the function of the enzyme-mediated post- translational modification of the LHCPI1 proteins are currently underway in our laboratory.

E. Villalobos et al.: Immunolocalization of transglutaminase in maize 163
Acknowledgments
We particularly thank Dr. Donatella Serafini-Fracassini (Universitfi di Bologna, Italy) for her help in obtaining the plant TGase anti- body. We thank Dr. J. Casas for his work as head of the animal ser- vices, from which the antibody was obtained (CID of Barcelona, CSIC, Spain). We thank Servei de Microscopia (Facultat de Biolo- gia, Universitat de Barcelona, Spain) for their assistance in sample preparation. We thank Angel Sanchez (IBMB, Servei d'Imatges) for the processing of the figures. We thank Dr. Robin Rycroft for cor- recting the English text. This work was supported by a Spanish grant DGICYT PB-97 1138.
References
Anderson J-M (1987) Molecular organisation of thylakoid mem- branes. In: Amesz J (ed) Photosynthesis. Elsevier, Amsterdam, pp 273-297
Bassi R, Hoyer-Hansen G, Barbato R, Giacometti G-M, Simpson D-J (1987) Chlorophyll-proteins of the photosystem II antenna system. J Biol Chem 262:13333-13341
Bennett J (1983) Regulation of photosynthesis by reversible phos- phorylation of the light-harvesting chlorophyll a/b protein. Biochem J 212:1-13
Bernet E, Claparols I, Dondini L, Santos M-A, Serafini-Fracassini D, Torn6 J-M (1999) Changes in polyaminc content, arginine and ornithine decarboxylases and transglutaminase activities during light/dark phases (of initial differentiation) in maize calluses and their chloroplasts. Plant Physiol Biochem 37:899-909
Berthold D-A, Babcock G-T, Yokum C-F (1981) A highly resolved oxygen evolving PSII preparation from spinach thylakoid mem- branes. FEBS Lett 134:231-234
Dainese R Santini C, Ghiretti-Magaldi A, Marquardt J, Tidu V, Mauro S, Bergantino E, Bassi R (1992) The organisation of pigment proteins within photosystem II. In: Murata N (ed) Research in plant photosynthesis, vol 2. Kluwer, Dordrecht, pp 13-20
Del Duca S, Favali A, Serafini Fracassini D, Pedrazzini R (1993) Transglutaminase activity during greening and growth of Helianthus tuberosus explants in vitro. Protoplasma 174:1-9
- Tidu V, Bassi R, Esposito C, Serafini-Fracassini D (1994) Identifi- cation of chlorophyll-a/b proteins as substrates of transglutami- nase activity in isolated chloroplasts of Helianthus tuberosus L. Planta 193:283-289
- Dondini L, Della Mea M, Mufioz de Rueda R Serafini-Fracassini D (2000) Factors affecting transglutaminase activity catalysing polyamine conjugation to endogenous substrates in the entire chloroplast. Plant Physiol Biochem 38:429-439
Dondini L (1998) Poliammine legate e transglutaminasi nelle plante. PhD thesis, University of Bologna, Bologna, Italy
- Bonazzi S, Serafini-Fracassini (D) (2000) Recovery of growth capacity and of chloroplast transglutaminase activity induced by polyamines in a polyamine-deficient variant strain of Dunaliella salina. J Plant Physiol 157:473-480
Folk JE (1980) Transglutaminases. Annu Rev Biochem 49: 517- 531
Hara-Nishimura I, Nishimura M (1987) Proglobulin processing enzyme in vacuoles isolated from developing pumpkin cotyle- dons. Plant Physiol 85:440-445
-Sh imada T, Hatano K, Takeuchi Y, Nishimura M (1998) Transport of storage proteins to protein-storage vacuoles is medi- ated by large precursor-accumulating vesicles. Plant Cell. 10: 825- 836
Hayasi M, Toriyama K, Kondo M, Hara-Nishimura I, Nishimura M (1999) Accumulation of a fusion protein containing 2S albumin induces novel vesicles in vegetative cells of Arabidopsis. Plant Cell Physiol 40:263-272
Kim H-C, Idler WW, Kim G-, Han H J, Chung S-I, Steiner P-M (1991) The complete amino acid sequence of human transgluta- minase K enzyme reduced from the nucleic acid sequences of cDNA clones. J Biol Chem 266:536-539
Ku S-B, Gutierrez M, Kanai R, Edwards G-E (1974) Photosynthe- sis in mesophyll protoplasts and bundle sheat cells of various types of C4 plants II: chorophyll and Hill reaction studies. Z Pflanzenphysiol 72:320-337
Laemmli UK (1970) Cleavage of structural protein during the assembly of the head of bacteriophage T4. Nature 227:680-683
Levanony H, Rubin R, Altschuler Y, Galili G (1992) Evidence for a novel route of wheat storage proteins to vacuoles. J Cell Biol 119: 1117-1128
Margosiak S-A, Dharma A, Bruce-Carver M-R, Gonzalez A-R Louie D, Kuehn G-D (1990) Identification of the large subunit of ribulose 1,5-biphosphate carboxylase/oxygenase as a substrate for transglutaminase in Medicago sativa L. (alfalfa). Plant Physiol 92: 88-96
Murakami S (1991) Structural and functional organisation of the thylakoid membrane system in photosynthetic apparatus. J Elec- tron Microsc 41:424-433
- Toda T (1988) Cytochemical detection and immuno-go M local- ization of photosystem I and photosystem II in the thylakoid membrane system during its differentiation. In: Recent develop- ment of electron microscopy 1987. Business Center for Academic Societies, Tokyo, pp 257-262
Murashige T, Skoog F (1962) A revised medium for rapid growth and bioassays with tobacco tissue cultures. Plant Physiol 15: 473-497
Rice R-H, Qin Q, Phillips M-A (1992) Membrane anchorage of ker- atinocyte transglutaminase: analysis of mutants. In: Birckbichler P (ed) Proceedings of the Third International Conference of Trans- glutaminase and Protein Crosslinking Reactions. Noble Founda- tion, Ardmore, Okla, p 2
Santos M, Claparols I, Torn6 J-M (1993) Effect of exogenous argi- nine, ornithine, methionine and 7-aminobutyric acid on maize (Zea mays L.) embryogenesis and polyamine content. J Plant Physiol 142:74-80
Schuster G, Ohad I, Martineau B, Taylor W-C (1985) Differentiation and development of bundle-sheat and mesophyll thylakoids in maize: thylakoid polypeptide composition, phosphorylation and organisation of photosystem II. J Biol Chem 260:11866-11873
Serafini-Fracassini D, Del Duca S, D'Orazi D (1988) First evidence for polyamine conjugation mediated by an enzyme activity in plants. Plant Physiol 87:757-761
- -Torrigiani P (1989) Polyamine conjugation during the cell cycle of Helianthus tuberosus: non enzymatic and transglutaminase-like binding activity. Plant Physiol Biochem 27:659-668
Sheen, JY. Bogorad L (1986) Differential expression of six light- harvesting chlorophyll a/b binding protein genes in maize leaf cell types. Proc Natl Acad Sci 83:7811-7815
Torn6 JM, Santos MA, Blanco JL (1984) Methods of obtaining maize totipotent tissues II: atrophic tissues culture. Plant Sci Lett 33:317-325
Towbin H, Staehelin T, Gordon J (1979) Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc Natl Acad Sci USA 76: 4350.4354
Welburn AR (1980) Bioenergetic and ultrastructural changes asso- ciated with chloroplast development. Int Rev Cytol 80:133-191














![Grana acalculia 2006[1]NPS](https://static.fdokumen.com/doc/165x107/6313b81efc260b71020f3816/grana-acalculia-20061nps.jpg)





