
IL25 causes Apoptosis of IL-25R-Expressing Breast Cancer Cells Without Toxicity to Nonmalignant...
35
DOI: 10.1126/scitranslmed.3001374 , 78ra31 (2011); 3 Sci Transl Med , et al. Saori Furuta Without Toxicity to Nonmalignant Cells Expressing Breast Cancer Cells - IL-25 Causes Apoptosis of IL-25R Editor's Summary Breast Tissue, Heal Thyself breast cancer drugs with broad therapeutic windows. al. strengthen the notion that targeting of the IL-25R signaling pathway forms the basis for the discovery of specific cell secretion of IL-17B might boost tumor growth if there is no IL-25 in the vicinity. Together, the findings of Furuta et surrounding epithelia might kill individual IL-25Rexpressing tumor cells as they form, but if one escapes, then cancer steals the show depends upon which one is on stage and where we are in the dramatic arc. IL-25 secreted by the IL-25Rexpressing tumors divide or die in vivo. Thus, in the context of mammary tumor development, which actor et al. hypothesize that it is the spatial and temporal expression of cytokines that determines whether this was not a surprising result, because IL-25 is known to bind IL-25R with a higher affinity than does IL-17B. Furuta express IL-25R. In contrast to IL-17B, addition of IL-25 to IL-17Bexpressing breast cancer cells induced apoptosis; deciphering whether IL-17B enhances, in an autocrine fashion, the tumorigenic potential of breast cancer cells that proliferate and invade; subsequent addition of the cytokine restored these functions. The authors are currently synthesis was blocked, with a small inhibitory RNA molecule, in cultured breast cancer cells, they lost their ability to another IL-25R ligand called IL-17B, which might compete with IL-25 for binding to the receptor. When IL-17B about one-third of the breast cancer biopsies tested, but not normal mammary cells, produce large amounts of It remains unknown how IL-25R imparts a growth advantage on breast cancer cells. The authors report that viable therapeutic approach for advanced breast cancer. had no toxic effect on normal tissues. These findings suggest that targeting of the IL-25R signaling pathway may be a had poor prognoses. Furthermore, IL-25 blocked the growth of IL-25Rexpressing mammary tumors in animals but tumor cells from patients and found that IL-25R was expressed in the greatest amounts on tumors from patients who potential in vivo role of IL-25R in breast cancer, the authors assessed whether the protein existed on the surface of no detrimental effect on nonmalignant MECs, which expressed only miniscule amounts of IL-25R. To investigate the assassin, IL-25, induced caspase-mediated apoptosis in cultured breast cancer cells that expressed IL-25R, but had et al. identified six factors secreted by MECs that killed breast cancer cells grown in the 3D culture. The most potent which cellular products are secreted. Using mass spectrometry analysis of the medium and functional assays, Furuta directed the formation of 3D aciniclusters of cells that form the glandular epithelium organized around a lumen into differentiation of other adjacent cells. The authors cultured nonmalignant MECs on a 3D laminin-rich matrix that Epithelial cells are known to function in tissue development and to produce factors that regulate the growth and therapeutic target for certain breast cancers. cytokine, interleukin-25 (IL-25), and show that signaling through the IL-25 receptor (IL-25R) may constitute a new midst of normal ones. Now, Furuta et al. find that normal mammary epithelial cells (MECs) deliver an anticancer conditions, normal cells might secrete substances that could stymie neighboring tumor cells that materialize in the cellular growth control is central to the tissue development process, researchers have suggested that under some development, the cradling microenvironment cooperates with cells in the creation of tissues and organs. Because actors play more complex roles and can display previously unappreciated intricacies of human biology. During the stage consists of a three-dimensional (3D) matrix designed to mimic the in vivo tumor microenvironment, cellular when limited good-guy, bad-guy personifications and thus reveal subtle nuances about the human condition. Similarly, Actors and their audiences alike prefer multidimensional characters because theyre more realistic than the is a registered trademark of AAAS. Science Translational Medicine rights reserved. The title NW, Washington, DC 20005. Copyright 2011 by the American Association for the Advancement of Science; all last week in December, by the American Association for the Advancement of Science, 1200 New York Avenue (print ISSN 1946-6234; online ISSN 1946-6242) is published weekly, except the Science Translational Medicine on April 13, 2011 stm.sciencemag.org Downloaded from
Transcript of IL25 causes Apoptosis of IL-25R-Expressing Breast Cancer Cells Without Toxicity to Nonmalignant...

DOI: 10.1126/scitranslmed.3001374, 78ra31 (2011);3 Sci Transl Med
, et al.Saori FurutaWithout Toxicity to Nonmalignant Cells
Expressing Breast Cancer Cells−IL-25 Causes Apoptosis of IL-25R
Editor's Summary
Breast Tissue, Heal Thyself
breast cancer drugs with broad therapeutic windows.al. strengthen the notion that targeting of the IL-25R signaling pathway forms the basis for the discovery of specific cell secretion of IL-17B might boost tumor growth if there is no IL-25 in the vicinity. Together, the findings of Furuta etsurrounding epithelia might kill individual IL-25R�expressing tumor cells as they form, but if one escapes, then cancer steals the show depends upon which one is on stage and where we are in the dramatic arc. IL-25 secreted by theIL-25R�expressing tumors divide or die in vivo. Thus, in the context of mammary tumor development, which actor et al. hypothesize that it is the spatial and temporal expression of cytokines that determines whetherthis was not a surprising result, because IL-25 is known to bind IL-25R with a higher affinity than does IL-17B. Furuta express IL-25R. In contrast to IL-17B, addition of IL-25 to IL-17B�expressing breast cancer cells induced apoptosis;deciphering whether IL-17B enhances, in an autocrine fashion, the tumorigenic potential of breast cancer cells that proliferate and invade; subsequent addition of the cytokine restored these functions. The authors are currentlysynthesis was blocked, with a small inhibitory RNA molecule, in cultured breast cancer cells, they lost their ability to another IL-25R ligand called IL-17B, which might compete with IL-25 for binding to the receptor. When IL-17Babout one-third of the breast cancer biopsies tested, but not normal mammary cells, produce large amounts of
It remains unknown how IL-25R imparts a growth advantage on breast cancer cells. The authors report thatviable therapeutic approach for advanced breast cancer.had no toxic effect on normal tissues. These findings suggest that targeting of the IL-25R signaling pathway may be ahad poor prognoses. Furthermore, IL-25 blocked the growth of IL-25R�expressing mammary tumors in animals but tumor cells from patients and found that IL-25R was expressed in the greatest amounts on tumors from patients whopotential in vivo role of IL-25R in breast cancer, the authors assessed whether the protein existed on the surface of no detrimental effect on nonmalignant MECs, which expressed only miniscule amounts of IL-25R. To investigate theassassin, IL-25, induced caspase-mediated apoptosis in cultured breast cancer cells that expressed IL-25R, but had et al. identified six factors secreted by MECs that killed breast cancer cells grown in the 3D culture. The most potentwhich cellular products are secreted. Using mass spectrometry analysis of the medium and functional assays, Furuta directed the formation of 3D acini�clusters of cells that form the glandular epithelium organized around a lumen intodifferentiation of other adjacent cells. The authors cultured nonmalignant MECs on a 3D laminin-rich matrix that
Epithelial cells are known to function in tissue development and to produce factors that regulate the growth andtherapeutic target for certain breast cancers.cytokine, interleukin-25 (IL-25), and show that signaling through the IL-25 receptor (IL-25R) may constitute a new midst of normal ones. Now, Furuta et al. find that normal mammary epithelial cells (MECs) deliver an anticancerconditions, normal cells might secrete substances that could stymie neighboring tumor cells that materialize in the cellular growth control is central to the tissue development process, researchers have suggested that under somedevelopment, the cradling microenvironment cooperates with cells in the creation of tissues and organs. Because actors play more complex roles and can display previously unappreciated intricacies of human biology. Duringthe stage consists of a three-dimensional (3D) matrix designed to mimic the in vivo tumor microenvironment, cellular
whenlimited good-guy, bad-guy personifications and thus reveal subtle nuances about the human condition. Similarly, Actors and their audiences alike prefer multidimensional characters because they�re more realistic than the
is a registered trademark of AAAS. Science Translational Medicinerights reserved. The title NW, Washington, DC 20005. Copyright 2011 by the American Association for the Advancement of Science; alllast week in December, by the American Association for the Advancement of Science, 1200 New York Avenue
(print ISSN 1946-6234; online ISSN 1946-6242) is published weekly, except theScience Translational Medicine
on
Apr
il 13
, 201
1st
m.s
cien
cem
ag.o
rgD
ownl
oade
d fr
om

http://stm.sciencemag.org/content/3/78/78ra31.full.htmlcan be found at:
and other services, including high-resolution figures,A complete electronic version of this article
http://stm.sciencemag.org/content/suppl/2011/04/11/3.78.78ra31.DC1.html can be found in the online version of this article at: Supplementary Material
http://www.sciencemag.org/about/permissions.dtl in whole or in part can be found at: article
permission to reproduce this of this article or about obtaining reprintsInformation about obtaining
is a registered trademark of AAAS. Science Translational Medicinerights reserved. The title NW, Washington, DC 20005. Copyright 2011 by the American Association for the Advancement of Science; alllast week in December, by the American Association for the Advancement of Science, 1200 New York Avenue
(print ISSN 1946-6234; online ISSN 1946-6242) is published weekly, except theScience Translational Medicine
on
Apr
il 13
, 201
1st
m.s
cien
cem
ag.o
rgD
ownl
oade
d fr
om

R E S EARCH ART I C L E
CANCER
IL-25 Causes Apoptosis of IL-25R–Expressing BreastCancer Cells Without Toxicity to Nonmalignant CellsSaori Furuta,1,2 Yung-Ming Jeng,1,3 Longen Zhou,1 Lan Huang,4,5 Irene Kuhn,2
Mina J. Bissell,2* Wen-Hwa Lee1*
011
As cells differentiate into tissues, the microenvironment that surrounds these cells must cooperate so that properlyorganized, growth-controlled tissues are developed and maintained. We asked whether substances produced fromthis collaboration might thwart malignant cells if they arise in the vicinity of normal tissues. Here, we identified sixfactors secreted by nonmalignant mammary epithelial cells (MECs) differentiating in three-dimensional laminin-richgels that exert cytotoxic activity on breast cancer cells. Among these, interleukin-25 (IL-25/IL-17E) had the highestanticancer activity without affecting nonmalignant MECs. Apoptotic activity of IL-25 was mediated by differentialexpression of its receptor, IL-25R, which was expressed in high amounts in tumors from patients with poor prog-noses but was low in nonmalignant breast tissue. In response to IL-25, the IL-25R on the surface of breast cancercells activated caspase-mediated apoptosis. Thus, the IL-25/IL-25R signaling pathway may serve as a new therapeutictarget for advanced breast cancer.
3, 2
on
Apr
il 1
stm
.sci
ence
mag
.org
Dow
nloa
ded
from
INTRODUCTION
Normal epithelial cells play active and critical roles in the maintenanceof tissue homeostasis. These cells regulate the secretion of autocrineand paracrine factors that promote the development of healthy or-gans and prevent aberrant growth of neighboring cells. We showedpreviously that in nonmalignant mammary epithelial cells (MECs),the breast cancer–associated gene 1 (BRCA1) protein forms a tran-scriptional repressor complex with the C-terminal binding proteininteracting protein (CtIP) and the zinc finger protein ZBRK1 that sup-presses angiopoetin-1 gene expression, which in turn inhibits angio-genesis in neighboring endothelial cells (1). During mammary glanddevelopment, MECs produce substances that inhibit epithelial growth,including mammary-derived growth inhibitor and transforminggrowth factor–b (2). In the mammary gland, myoepithelial cells con-tribute to basement membrane formation and tissue polarity and se-crete anti-invasive and antiangiogenic factors, including maspin andlaminin-111 (3, 4). We asked whether normal MECs also secrete fac-tors that directly and specifically inhibit malignant cell growth and/orsurvival and, if so, what are the underlying mechanisms for this intrin-sic antitumor activity.
We showed previously that conditioned medium (CM) from non-malignant MECs in the process of forming acini in three-dimensional(3D) laminin-rich extracellular matrix (lrECM) gels “reverts” the ma-lignant phenotype of breast cancer cells to basally polarized, growth-suppressed (dormant), acinar-like structures; this phenotypical reversionwas also shown with cell signaling inhibitors such as a b1 integrin–blocking antibody (5, 6). These observations suggest that acinus-formingnonmalignant MECs secrete factors that can suppress the phenotype
1Department of Biological Chemistry, College of Medicine, University of California, Irvine,CA 92697, USA. 2Department of Cancer and DNA Damage Responses, Life SciencesDivision, Lawrence Berkeley National Laboratory, Berkeley, CA 94720, USA. 3Departmentof Pathology, National Taiwan University Hospital, Taipei 100, Taiwan. 4Department ofPhysiology and Biophysics, College of Medicine, University of California, Irvine, CA92697, USA. 5Departments of Developmental and Cell Biology, College of Medicine,University of California, Irvine, CA 92697, USA.*To whom correspondence should be addressed. E-mail: [email protected] (M.J.B.);[email protected] (W.-H.L.)
www.S
of breast cancer cells growing in 3D cultures. We hypothesized thatsuch complex phenotypical reversion is likely a result of multiple sig-naling factors that in combination allow cancer cells to form quiescentacinar-like structures.
Here, we sought to identify and characterize these factors using sizefractionation of the CM from nonmalignant cells and functional as-says to identify the active molecules. However, fractionation revealedthat the CM could be divided into morphogenic activity in the in-soluble fraction and cytotoxic activity in the soluble fraction. Here,we characterized the molecule with the most potent tumor cell–killingactivity in the 10- to 50-kD fraction, which proved to be interleukin-25 (IL-25). We then showed that this cytokine is active against breastcancer cells that express high amounts of the cognate receptor, IL-25R,and that treatment of these cells with IL-25 caused the receptor to inter-act with death domain (DD)–associating proteins, activating caspase-mediated apoptosis.
To determine the relevance of the above findings to breast cancer,we measured the levels of IL-25R in two separate tissue cohorts de-rived from breast cancer patients and found that high levels of IL-25Rcorrelated with poor prognosis. These data suggest strongly that theIL-25/IL-25R signaling pathway may provide novel targets for treatingaggressive breast cancers.
RESULTS
Factors secreted from differentiating MECs kill breastcancer cellsTo identify and characterize the components secreted by the nonma-lignant cells, we fractionated their CM by solubility and molecular size.Each fraction of the CM was replenished with essential growth factors(7) and used instead of the usual media to culture MCF7 breast cancercells in 3D lrECM (Fig. 1A). The reverting/dormancy-inducing activ-ity of the total CM could be separated into two complementary activ-ities: morphogenic activity in the pellet fraction and cytotoxic activityin the soluble fraction. The pellet fraction caused breast cancer cellsto form spheroid structures reminiscent of differentiating MECs,
cienceTranslationalMedicine.org 13 April 2011 Vol 3 Issue 78 78ra31 1
R E S EARCH ART I C L E
although larger in size; this phenotype was distinct from those ob-served in breast cancer cells treated with total CM.
In contrast, the soluble fraction was cytotoxic for breast cancercells (Fig. 1A), and this activity was further enriched in the 10- to50-kD fraction. When treated with this fraction alone, there wereno viable cells after 1 week (Fig. 1C). None of the CM fractions ap-plied to the nonmalignant MCF10A cells in 3D cultures resulted incell-killing activity (Fig. 1, B and C). These observations indicate that,whereas differentiating MECs secrete factors in the 10- to 50-kD
www.S
range that exhibit tumor cell–specific cytotoxic activity, the non-malignant cells themselves are resistant to such activity. The highesttumor cell–killing activity was detected in CM from MECs that hadbeen differentiating for 4 days in the 3D cultures; this time scale cor-responds to the midpoint in the process of acini formation (Fig. 1D).The result suggests that secretion of the cytotoxic factors for cancercells is intimately linked to the progression of differentiation in 3Dcultures and that the activity is effective on all the cancer cell linestested.
cienceTranslationalMedicine.or
on
Apr
il 13
, 201
1st
m.s
cien
cem
ag.o
rgD
ownl
oade
d fr
om
Identification of factors secretedinto the 10- to 50-kD CMTo identify proteins specifically secretedby differentiating MECs, but not by breastcancer cells, we analyzed the 10- to 50-kDCM fraction (day 4) from differentiatingMCF10A cells and that fromMCF7 breastcancer cells also cultured in 3D lrECM bymass spectrometry using 2D liquid chro-matography (8). Through comparativeanalyses, we identified six differentially ex-pressed factors, including antiangiogenicproteins [antithrombin III (ATIII) and vi-tamin D–binding protein (VBP) (9, 10)],proinflammatory cytokines [IL-1F7 andIL-25 (also called IL-17E) (11, 12)], andgrowth and differentiation proteins [bonemorphogenetic protein 10 (BMP10) and fi-broblast growth factor 11 (FGF11) (13, 14)](Fig. 2A).
To verify the contribution of these fac-tors to tumor cell–killing activity, we im-munodepleted the 10- to 50-kD cytotoxicfraction from days 3 to 4 with antibodiesto the six factors individually and to p84control protein. The depletion efficacy wasassessed by immunoblots performed on theimmunoprecipitates (Fig. 2B). The immuno-depleted fraction was used to cultureMCF7breast cancer cells in 3D lrECM, and freshfractions were applied every 24 hours for1 week. Depletion of individual factors at-tenuated the cell-killing activity to a dif-ferent degree (Fig. 2C). However, protectionfrom cell-killing activity was especially pro-nounced for depletion of ATIII, IL-1F7,IL-25, and VBP, suggesting critical rolesfor each of these four factors in this process.
To determine which of the four factorscontained the most potent activity, we used293T cells to generate cell lines that stablyexpressed ATIII, IL-1F7, IL-25, and VBP atcomparable levels, determined by analysesof RNAs (Fig. 2D) and protein levels inthe media (15). Cells were seeded at thesame density, and the collected CM wasnormalized to total protein concentrationsand tested on MCF7 cells cultured in 3D
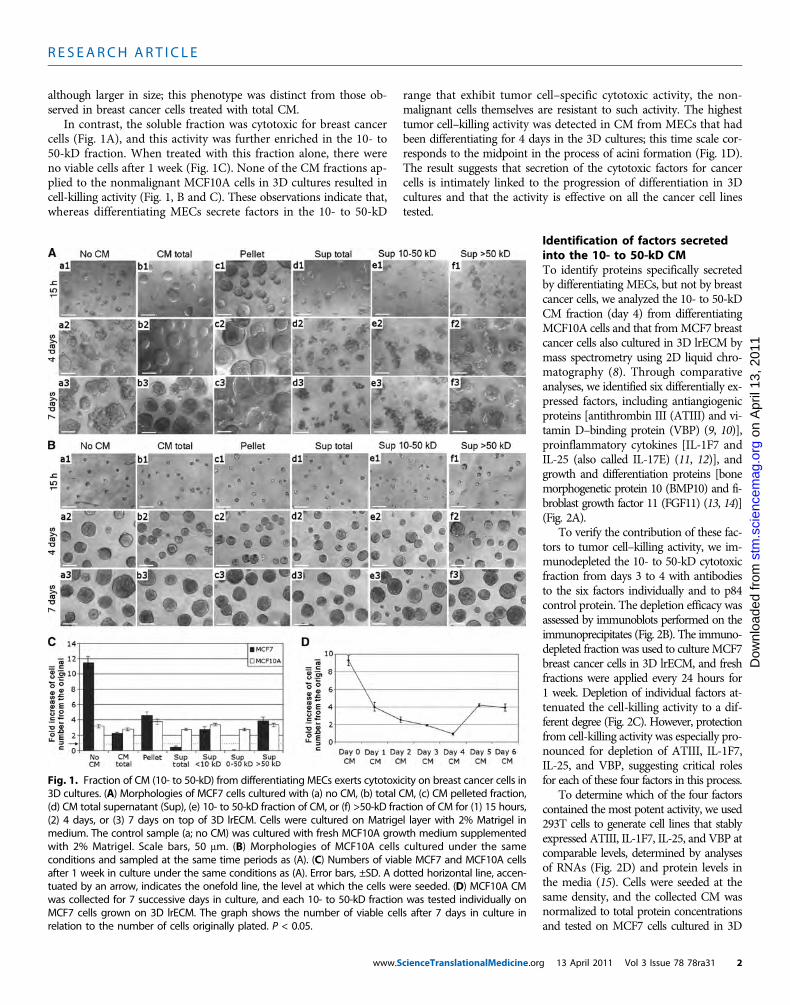
Fig. 1. Fraction of CM (10- to 50-kD) from differentiating MECs exerts cytotoxicity on breast cancer cells in3D cultures. (A) Morphologies of MCF7 cells cultured with (a) no CM, (b) total CM, (c) CM pelleted fraction,
(d) CM total supernatant (Sup), (e) 10- to 50-kD fraction of CM, or (f) >50-kD fraction of CM for (1) 15 hours,(2) 4 days, or (3) 7 days on top of 3D lrECM. Cells were cultured on Matrigel layer with 2% Matrigel inmedium. The control sample (a; no CM) was cultured with fresh MCF10A growth medium supplementedwith 2% Matrigel. Scale bars, 50 mm. (B) Morphologies of MCF10A cells cultured under the sameconditions and sampled at the same time periods as (A). (C) Numbers of viable MCF7 and MCF10A cellsafter 1 week in culture under the same conditions as (A). Error bars, ±SD. A dotted horizontal line, accen-tuated by an arrow, indicates the onefold line, the level at which the cells were seeded. (D) MCF10A CMwas collected for 7 successive days in culture, and each 10- to 50-kD fraction was tested individually onMCF7 cells grown on 3D lrECM. The graph shows the number of viable cells after 7 days in culture inrelation to the number of cells originally plated. P < 0.05.g 13 April 2011 Vol 3 Issue 78 78ra31 2
R E S EARCH ART I C L E
on
Apr
il 13
, 201
1st
m.s
cien
cem
ag.o
rgD
ownl
oade
d fr
om
lrECM. CM from cells that overexpressed ATIII, IL-1F7, and VBPshowed a cytostatic effect. However, CM from cells that overexpressedIL-25 showed a strong cytotoxic activity on MCF7 (Fig. 2E) as well ason two additional breast cancer lines, T47D and MDA-MB468 (Fig.2F). We therefore concentrated on the mechanism of the cytotoxic ac-tivity of IL-25 and its possible utility in treating malignant tumors. Asexpected, nonmalignant MCF10A cells were resistant to the cytotoxicactivity of IL-25–containing CM (Fig. 2F). IL-25 expression in 3D cul-
www.S
tures correlated with the cytotoxicity of the CM described above (Fig.1D) and peaked at day 4 in culture as well (Fig. 2G).
To determine the physiological significance of these modulations,we analyzed IL-25 expression in vivo in mice and on sections of nor-mal breast tissue. We compared human and mouse IL-25 genes andfound them to share similar regulatory elements in their promoter re-gions (Fig. 3A). Analysis of the mouse mammary glands as a functionof the pregnancy cycle showed that IL-25 expression gradually increases
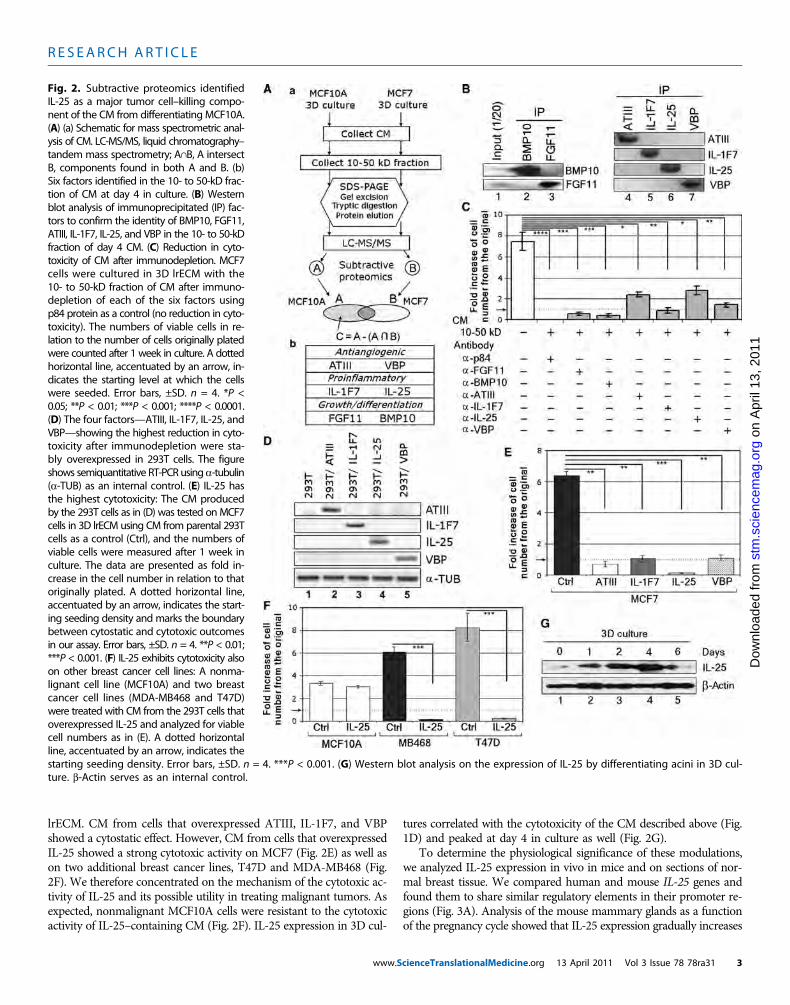
Fig. 2. Subtractive proteomics identifiedIL-25 as a major tumor cell–killing compo-nent of the CM from differentiating MCF10A.(A) (a) Schematic for mass spectrometric anal-ysis of CM. LC-MS/MS, liquid chromatography–tandem mass spectrometry; A∩B, A intersectB, components found in both A and B. (b)Six factors identified in the 10- to 50-kD frac-tion of CM at day 4 in culture. (B) Westernblot analysis of immunoprecipitated (IP) fac-tors to confirm the identity of BMP10, FGF11,ATIII, IL-1F7, IL-25, and VBP in the 10- to 50-kDfraction of day 4 CM. (C) Reduction in cyto-toxicity of CM after immunodepletion. MCF7cells were cultured in 3D lrECM with the10- to 50-kD fraction of CM after immuno-depletion of each of the six factors usingp84 protein as a control (no reduction in cyto-toxicity). The numbers of viable cells in re-lation to the number of cells originally platedwere counted after 1 week in culture. A dottedhorizontal line, accentuated by an arrow, in-dicates the starting level at which the cellswere seeded. Error bars, ±SD. n = 4. *P <0.05; **P < 0.01; ***P < 0.001; ****P < 0.0001.(D) The four factors—ATIII, IL-1F7, IL-25, andVBP—showing the highest reduction in cyto-toxicity after immunodepletion were sta-bly overexpressed in 293T cells. The figureshows semiquantitative RT-PCR using a-tubulin(a-TUB) as an internal control. (E) IL-25 hasthe highest cytotoxicity: The CM producedby the 293T cells as in (D) was tested onMCF7cells in 3D lrECM using CM from parental 293Tcells as a control (Ctrl), and the numbers ofviable cells were measured after 1 week inculture. The data are presented as fold in-crease in the cell number in relation to thatoriginally plated. A dotted horizontal line,accentuated by an arrow, indicates the start-ing seeding density and marks the boundarybetween cytostatic and cytotoxic outcomesin our assay. Error bars, ±SD. n = 4. **P < 0.01;***P < 0.001. (F) IL-25 exhibits cytotoxicity alsoon other breast cancer cell lines: A nonma-lignant cell line (MCF10A) and two breastcancer cell lines (MDA-MB468 and T47D)were treated with CM from the 293T cells thatoverexpressed IL-25 and analyzed for viablecell numbers as in (E). A dotted horizontalline, accentuated by an arrow, indicates the
starting seeding density. Error bars, ±SD. n = 4. ***P < 0.001. (G) Western blot analysis on the expression of IL-25 by differentiating acini in 3D cul-ture. b-Actin serves as an internal control.cienceTranslationalMedicine.org 13 April 2011 Vol 3 Issue 78 78ra31 3
R E S EARCH ART I C L E
on
Apr
il 13
, 201
1st
m.s
cien
cem
ag.o
rgD
ownl
oade
d fr
om
after puberty at about 5 weeks up to mid-pregnancy (Fig. 3, B to E) but declinesthereafter, suggesting that IL-25 is tempo-rally regulated in developing mammaryglands reminiscent of temporal regulationof IL-25 secretion during the differentia-tion process in 3D cultures (Fig. 2G). Onceacini are formed either by mouse mamma-ry glands at mid-pregnancy (Fig. 3, B to E)or by nonmalignant human breast cells in3D cultures (Fig. 2G), IL-25 expression wasdiminished. Indeed, when we analyzedIL-25 expression in normal breast tissuesections (n = 30) by immunohistochem-istry (IHC), IL-25 signal was almost un-detectable (Fig. 3F).
Purified IL-25 kills cancer cellsin culture and inhibits tumorgrowth in vivoTo ascertain whether the cytotoxic activ-ity of IL-25–containing CM did indeedresult from IL-25, we purified this cyto-kine from 293T cells that stably expressedIL-25. Because secreted IL-25 was expectedto be highly glycosylated, as is the case forother interleukin family members (16), thetotal glycoproteins were affinity-purifiedand then separated by gel filtration (size ex-clusion) chromatography. Pooled proteinfractions 21 to 23 contained similar amountsof IL-25 and bovine serum albumin (BSA)carrier protein. Otherwise, IL-25 was esti-mated to be 90 to 95% pure. Upon electro-phoresis in a denaturing gel, glycosylatedIL-25 migrated at a position that corre-sponded to ~48 kD (Fig. 4A).
To test the activity of this fraction, wetreated four breast cancer cell lines (MCF7,MDA-MB468, SKBR3, and T47D) withdifferent doses of IL-25. All four breastcancer cell lines were sensitive to the pu-rified IL-25 fraction in colony-formingassays, with IC50 (median inhibitory con-centration) values of ~10 ng/ml (500 pM;molecular size = 20 kD for the nonglyco-sylated protein) (Fig. 4B). In contrast, non-malignant MCF10A cells were relativelyresistant to IL-25 (Fig. 4B).
We tested the potency of IL-25 in vivousing xenografts of MDA-MB468 breastcancer cells grown in mammary fat padsof nude mice. The tumors were grown for10 days and then were injected with ve-hicle (phosphate-buffered saline, n = 7)or IL-25 (300 ng, n = 8) once a day for1 month. IL-25 significantly retarded growthof the tumors. After 1 month, the average
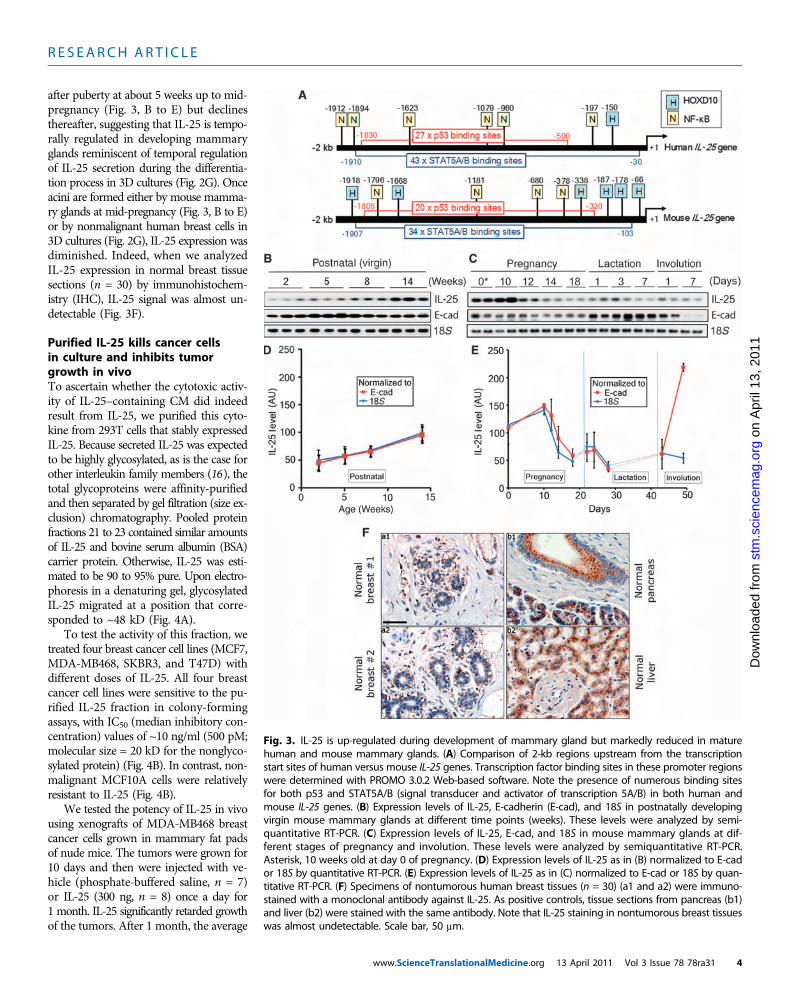
Fig. 3. IL-25 is up-regulated during development of mammary gland but markedly reduced in maturehuman and mouse mammary glands. (A) Comparison of 2-kb regions upstream from the transcriptionstart sites of human versus mouse IL-25 genes. Transcription factor binding sites in these promoter regionswere determined with PROMO 3.0.2 Web-based software. Note the presence of numerous binding sitesfor both p53 and STAT5A/B (signal transducer and activator of transcription 5A/B) in both human andmouse IL-25 genes. (B) Expression levels of IL-25, E-cadherin (E-cad), and 18S in postnatally developingvirgin mouse mammary glands at different time points (weeks). These levels were analyzed by semi-quantitative RT-PCR. (C) Expression levels of IL-25, E-cad, and 18S in mouse mammary glands at dif-ferent stages of pregnancy and involution. These levels were analyzed by semiquantitative RT-PCR.Asterisk, 10 weeks old at day 0 of pregnancy. (D) Expression levels of IL-25 as in (B) normalized to E-cador 18S by quantitative RT-PCR. (E) Expression levels of IL-25 as in (C) normalized to E-cad or 18S by quan-titative RT-PCR. (F) Specimens of nontumorous human breast tissues (n = 30) (a1 and a2) were immuno-stained with a monoclonal antibody against IL-25. As positive controls, tissue sections from pancreas (b1)and liver (b2) were stained with the same antibody. Note that IL-25 staining in nontumorous breast tissueswas almost undetectable. Scale bar, 50 mm.
www.ScienceTranslationalMedicine.org 13 April 2011 Vol 3 Issue 78 78ra31 4

R E S EARCH ART I C L E
www.ScienceTranslationalMedicine.or
on
Apr
il 13
, 201
1st
m.s
cien
cem
ag.o
rgD
ownl
oade
d fr
om
size of IL-25–treated tumors was three-fold smaller than that of control tumors(P = 0.0016) (Fig. 4C).
To assess the systemic stress/toxicitycaused by the treatment, we measured thebody weight of these mice and found nomeasurable difference between the con-trol and the treated groups (Fig. 4D). Inaddition, at the end of the experiment,we performed gross pathological exami-nations of various organs from the un-treated and IL-25–treated mice, includingintestine, kidney, heart, lungs, liver, andstomach based on size, color, presenceof cysts or any other gross lesions, aswell as any signs of inflammatory reac-tions or invasion. No apparent patholo-gies were found in any of these mice. Formore stringent safety tests, a fivefold high-er dose of IL-25 (1.5 mg) was injected intothe tail veins of C57 mice (vehicle: n = 3;IL-25: n = 5). Again, no signs of illness(lethargy or weight loss) were observed(15), indicating a lack of toxicity to non-malignant tissues.
When tumors from IL-25–treated anduntreated (control) MDA-MB468 xeno-grafts were excised and histologically exam-ined, the treated samples contained about50% fewer actively dividing tumor cells(Fig. 4E, b and c) that exhibited an averagemitotic index that was half that of untreatedtumors (Fig. 4E, a) (0.63 ± 0.055 versus1.1 ± 0.17, respectively, P = 0.053) (15). Insome of the IL-25–treated mice, the tu-mors had regressed completely, and thelesion contained only lymphocytic infil-trates (Fig. 4E, b and c). These findingssuggest a potential utility of IL-25 as anantitumor agent.
Breast tumor cells express highamounts of IL-25R comparedto nonmalignant cellsAs a mechanistic explanation for the selec-tive antitumor activity of IL-25 in mice,we speculated that IL-25–induced cyto-toxicity might be attributed to the differ-ential expression of IL-25R (also calledIL-17RB) on breast cancer cells. To test this,we screened for IL-25R expression in apanel of breast cancer cell lines that dis-played different characteristics. These in-cluded estrogen receptor (ER)–positivebreast cancer cells, such as the MCF7,MDA-MB361, T47D, and ZR75 cell lines,and ER-negative breast cancer cells, suchas the MCF10A, MDA-MB468, MB435-S,
Fig. 4. IL-25 exhibits cytotoxicity toward tumor cells both in colony assays and in vivo. (A) Secreted IL-25proteinwas purified from the CMof overexpressing 293T cells via affinity purification of glycosylated proteinsfollowed by gel filtration. Pooled gel-filtrated fractions containing glycosylated IL-25 (fractions 21 to 23) andother glycosylated proteins (fractions 17 to 20) were analyzed by Coomassie staining (CS, left) and Westernblot (WB, right). Note the abundant bovine serum albumin (BSA) eluted in fractions 17 to 20 and its carryoverin the IL-25 pool (fractions 21 to 23). MW,molecular weight. (B) Purified IL-25 protein described in (A) exhibitscytotoxicity on breast cancer cells. A nonmalignant cell line (MCF10A) and four breast cancer cell lines (MCF7,MDA-MB468, SKBR3, andT47D)were treatedwith varyingdoses of IL-25, and thenumbers of colonies formedwere counted after 10 days in cultures. Error bars, ±SD. n = 6. (C) Purified IL-25 inhibits the growth of xeno-grafted cancer cells. MDA-MB468 cells xenografted in themammary fat pads of nudemicewere treatedwithvehicle (control, n = 7) or IL-25 (300 ng, n = 8) for 31 days, and tumor sizes were measured twice weekly. P =0.0016. Error bars, ±SD. (D) The body weights of mice described in (C) showed no differences throughout theexperiment. Error bars, ±SD. n= 7 to 8. (E) Hematoxylin-eosin (H&E)–stained sections of (a) control and (b andc) IL-25–treated tumors as described in (C). The image (c) is a higher-magnification image of the boxed regionin (b). Control sample shows actively growing tumor cells [representative entities are indicated with arrows in(a)], whereas in the IL-25–treated sample, the tumorwas completely regressed and replacedwith lymphocyticinfiltrates [representative entities are shown in (c)]. Magnifications: ×400 (a and c); ×100 (b). Scale bars, 50 mm.
g 13 April 2011 Vol 3 Issue 78 78ra31 5

R E S EARCH ART I C L E
MB231, MB175-7, SKBR3, HS578T, HBL100, and HCC1937 cell lines(17). The analysis of transcripts by reverse transcription–polymerasechain reaction (RT-PCR) showed that IL-25R was expressed in higheramounts in all the breast cancer cell lines examined relative to nonma-lignant breast cell line MCF10A. Nevertheless, the IL-25R mRNA lev-els were surprisingly different among the cancer cell lines (Fig. 5A).On the other hand, the IL-25R protein concentrations in the breast can-cer cell lines were much more similar to each other than to IL-25R con-centrations in MCF10A cells and in another nonmalignant cell line,telomerase-immortalized human mammary epithelial (tHME) cells (Fig.5B). The low amounts of IL-25R measured in the nonmalignant MEC
www.S
cell lines are consistent with the IL-25R distribution reported in a pre-vious study for normal breast as well as other organs (11).
High amounts of IL-25R in breast cancer cells suggest that the re-ceptor may confer a growth advantage to these cells. To examine thispossibility, we depleted IL-25R from a panel of breast cancer cell linesusing an IL-25R–specific small interfering RNA (siRNA) (Fig. 5C) andmeasured the ability of the resulting cells to grow anchorage indepen-dently in soft agar (Fig. 5, D and E). Reduction of IL-25R markedlyinhibited (75 to 85%) the growth of all the cancer cell lines tested (Fig.5E), indicating that IL-25R is important for their anchorage-independentgrowth. To ascertain that the impaired ability of soft agar growth did
on
Apr
il 13
, 201
1st
m.s
cien
cem
ag.o
rgD
ownl
oade
d fr
om
Fig. 5. IL-25R is highly expressed in breasttumor cells, but not in nonmalignant MECs;the expression is involved in anchorage-independent growth ability of cells andinversely correlates with patients’ progno-sis. (A) RT-PCR analysis shows that breastcancer cell lines express higher amountsof IL-25R than nonmalignant breast cells.a-Tubulin (a-TUB) served as an internal con-trol. (B) Western blot analysis confirms thatIL-25R protein concentrations are higher inbreast cancer cell lines than in nonmalig-nant breast cell lines. b-Actin served as aninternal control. (C) Depletion of IL-25R invarious breast cell lines with a specific siRNA.The Western blot confirms a decrease inIL-25R amounts in a panel of breast cancercell lines (MCF7, MDA-MB468, MB435-S,MB231, SKBR3, and T47D) after treatmentwith IL-25R siRNA. b-Actin served as an inter-nal control. Luciferase (Luc) siRNA was usedas a nonspecific control. (D) Down-modulationof IL-25R impairs the ability of SKBR3 cancercells to grow on soft agar. Representativeimages of SKBR3 breast cancer cells grownfor 2 weeks on soft agar after treatment withLuc siRNA (a) or IL-25R siRNA (b) as in (C). n =6. (E) The number of colonies formed on softagar decreases in breast cancer cells aftertreatment with IL-25R siRNA as in (D). Errorbars, ±SD. KD, knockdown. (F) Expression ofsiRNA-resistant IL-25R restores IL-25R expres-sion after siRNA treatment. Western blot anal-ysis of IL-25R amounts in MDA-MB468 breastcancer cells expressing an IL-25R mutant(IL-25R RM) that is resistant to #1, but not#2, IL-25R siRNA after the cognate siRNAtreatment. pcDNA plasmid was used as a ve-hicle control; Luc siRNAwas used as a nonspe-cific siRNA control. (G) IL-25R RM expressionrescues the soft agar growth ability of MDA-MB468 cells treated with IL-25R siRNA. Cellswere transfected with vehicle or an IL-25RRM–expressing plasmid, treated with differ-
ent siRNA constructs as in (F), and then analyzed for the numbers of coloniesformed on soft agar. Error bars, ±SD. n = 6. (H) Specimens of nontumoroushuman breast tissue (a) versus human breast cancer (b) immunostained forIL-25R. Normal sample in upper left (a1) and tumor sample in upper right(b1) were both stained with an isotype-matched control IgG1 antibody,whereas the normal samples (a2) and (a3) and the tumor samples (b2)and (b3) were all stained with IL-25R antibody (GeneTex). Membranous stain-ing of IL-25R was seen in tumor cells and surrounding inflammatory cells, butnot in nonmalignant MECs. The images were captured at ×400 magnifica-tion. Scale bars, 25 mm. n = 69. (I) Survival analysis of patients with IL-25R(+)(n = 13) and IL-25R(−) (n = 56) tumors with XLSTAT-Life version 2007.4software. Statistical significance was assessed with the log-rank test.
cienceTranslationalMedicine.org 13 April 2011 Vol 3 Issue 78 78ra31 6

R E S EARCH ART I C L E
on
Apr
il 13
, 201
1st
m.s
cien
cem
ag.o
rgD
ownl
oade
d fr
om
not involve an off-target effect, we assessed whether coexpression ofan siRNA-resistant version of the IL-25R gene (IL-25R RM) could res-cue the growth properties of cancer cells. We generated an IL-25R RMconstruct that expressed an mRNA transcript harboring four silentmutations. In addition, we designed two siRNAs, only one of which tar-geted the region containing the mutations (siRNA #1). Western anal-ysis confirmed that the expression of IL-25R RM in MDA-MB468cancer cells rescued the IL-25R protein concentrations after treatmentwith siRNA #1, but not with siRNA #2 (Fig. 5F). Paralleling the re-stored IL-25R protein concentrations, expression of IL-25R RM rescuedthe ability of MDA-MB468 cells to grow in soft agar after treatmentwith IL-25R siRNA #1, but not with siRNA #2 (Fig. 5G). These resultsprovide clear evidence that IL-25R expression is directly responsiblefor anchorage-independent growth of breast cancer cells.
To determine the amount of IL-25R expression in human breastcancer specimens, we performed immunostaining using two differentIL-25R antibodies on two cohorts of resected ductal invasive carcinomasamples. The slides for analysis were selected to include both tumorand nontumorous epithelia. Of the 69 samples in the first cohort, whichconsisted of paraffin-embedded breast cancer tissue sections analyzedwith the first batch of antibody obtained from GeneTex (please referto Materials and Methods), there was intense membranous staining ofIL-25R in 18.8% of the tumor tissues analyzed (13 of 69) (Allred score:6 to 8) (Fig. 5H, b1 to b3) (18); in contrast, all nontumorous regions ex-amined (69 of 69) showed no or little staining (Allred score: 0 to 2) (Fig.5H, a1 to a3). Positive IL-25R staining correlated significantly with poorprognosis and high mortality rate in this cohort (P < 0.001) (Fig. 5I).Unfortunately, additional supplies of the GeneTex IL-25R antibody wereno longer available from the manufacturer, and of the other availableIL-25R antibodies, none could be used for IHC on paraffin sections(see Materials and Methods). On the basis of its accurate identificationof IL-25R in a number of assays, we chose a second IL-25R antibodyfrom R&D for our studies. We could use this antibody successfully forimmunofluorescence (IF) and for IHC on frozen tissue sections. By IF,the R&D antibody, similar to the GeneTex antibody, revealed accen-tuated localization of IL-25R at the cell membrane of cultured breastcancer cells, albeit with some additional staining in the Golgi (fig. S1A).IHC staining on a cohort of frozen sections of breast tissue samples (10nonmalignant and 20 malignant) revealed again tumor-specific staining(fig. S1B), as had the GeneTex antibody (Fig. 5H). Note that the stainingof frozen sections did not reveal the same degree of membrane speci-ficity or localization as the staining of paraffin sections (compare fig.S1B with Fig. 5H). Further specification of localization will need to awaitavailability of additional paraffin-compatible antibodies. Nevertheless,these two sets of IHC staining reveal high levels of IL-25R in tumor tis-sues, but none in normal tissues. The data thus provide a strongrationale for the observed tumor specificity of IL-25R cytotoxicityand its role as a marker of malignant progression and poor patientprognosis.
IL-25 stimulates formation of a cell death–inducingcomplex that activates caspase-mediated apoptosisIL-25/IL-25R signaling has been shown to induce a proinflammatoryresponse in certain tissues such as lung fibroblasts (19). In breast can-cer cells, on the other hand, we found that IL-25 induces cell death.We reasoned that if IL-25/IL-25R interaction can send a death signalin breast cancer cells, IL-25R may serve as the death receptor and con-tain a DD signature motif. We aligned the C-terminal region of IL-25R
www.S
(amino acids 362 to 467) with DDs of the apoptosis-stimulating frag-ment (FAS) receptor (FAS-R: amino acids 205 to 293) and tumor ne-crosis factor (TNF) receptor 1 (TNF-R1: amino acids 352 to 441) andfound that this region of IL-25R shares about 30% similarity with bothDDs (Fig. 6A). The residues highly conserved among DD-containingproteins are similar to those in the C-terminal region of IL-25R, exceptfor Trp72 (20) (see the numbering in Fig. 6A). Therefore, this regionof IL-25R appears to have a DD-like motif. Other members of theIL-17 receptor family, IL-17RA and IL-17RC, lack such a DD-likemotif (15).
To test whether IL-25 treatment induces receptor-mediated apoptosisof breast cancer cells, we treated MDA-MB468 and MCF10A cells,which express high and low amounts of IL-25R, respectively, with IL-25(Fig. 5, A and B). In MDA-MB468 breast cancer cells, but not in non-malignant MCF10A cells, IL-25 caused the cleavage of caspases 8 and 3within 30 min and measurable cleavage of poly(ADP-ribose) polymer-ase (PARP) after 24 hours (Fig. 6B), indicating the activation of apoptosis.Two alternative preparations of IL-25 protein—either immunoaffinity-purified from mammalian cells (fig. S2) or commercially availablebacterially produced recombinant protein—likewise exerted a potentproapoptotic effect on MDA-MB468 cells (fig. S3), corroborating themode of action of this protein.
To further confirm that IL-25R indeed mediates the cell death sig-naling of IL-25, we depleted IL-25R fromMDA-MB468 cells with siRNAs,which completely inhibited the protein expression after 60 hours (Fig. 6C).In cells treated with control luciferase siRNAs, IL-25 treatment causedcleavage of caspase 3 and PARP, whereas in cells depleted of IL-25R, suchphenomena were absent (Fig. 6D). These results substantiate the no-tion that the IL-25–dependent death signal is indeed mediated throughactivation of IL-25R.
We then explored how a death signal from IL-25 is transduced bythe receptor. If IL-25R were to have a DD-like motif, it would interactwith DD-associating proteins upon IL-25 binding. To test this hypoth-esis, we treated MDA-MB468 cells with IL-25 and analyzed for theinteractions between IL-25R and DD adaptor proteins: FAS-associatedprotein with death domain (FADD) and TNF-R1–associated death do-main protein (TRADD) (21). As a positive control, we also tested theinteraction of IL-25R with TNF receptor–associated factor 6 (TRAF6).TRAF6 was shown previously to be associated with IL-25R even in theabsence of the ligand through its binding to the motif around Glu341(Glu338 in mouse) of IL-25R (22), a region N-terminal to the putativeDD-like motif (amino acids 362 to 467).
We found that total amounts of IL-25R, FADD, and TRADD werehighly elevated upon IL-25 treatment (Fig. 6D; input level shown in Fig.6E). Moreover, IL-25R strongly interacted with FADD and TRADD onlyafter IL-25 addition, as detected by reciprocal immunoprecipitationwith IL-25R and FADD antibodies (Fig. 6E). The increases in FADD,TRADD, and IL-25R protein concentrations in response to IL-25 treat-ment may indicate their activation and contribute to their apparentincreased interactions. We confirmed that TRAF6 indeed remained as-sociated with IL-25R even in the absence of the ligand. Despite theunchanged total amount of TRAF6, the amount of TRAF6 that copre-cipitated with IL-25R increased in proportion to the elevated IL-25Rconcentrations upon IL-25 treatment (Fig. 6, D and E); these observa-tions suggest that IL-25R and TRAF6 are always bound together, aspreviously observed in cells in which the two proteins were overex-pressed ectopically (22). Moreover, reduction of FADD or TRADD inbreast cancer cells with specific siRNAs largely impaired the proapoptotic
cienceTranslationalMedicine.org 13 April 2011 Vol 3 Issue 78 78ra31 7
R E S EARCH ART I C L E
on
Apr
il 13
, 201
1st
m.s
cien
cem
ag.o
rgD
ownl
oade
d fr
om
activity of IL-25 (Fig. 6, F and G), suggesting that this function of IL-25depends on the FADD and TRADD proteins. These results indicatethat IL-25 binding to its receptor increases the stability of the IL-25R–associated complex, which contains FADD and TRADD. This complexthen triggers the activation of caspase-mediated apoptosis.
IL-25–mediated apoptotic signaling depends on theDD-like region of IL-25RTo dissect how IL-25 sends death signals through IL-25R, we ectopicallyexpressed inMCF10A cells either the wild-type IL-25R protein or a mu-tant from which either the TRAF6 binding domain (DTRAF6) or theDD-like region (DDD) was deleted (Fig. 7, A and B); MCF10A cellswere otherwise resistant to apoptosis mediated by IL-25 (Fig. 6B). Upon
www.S
IL-25 addition, MCF10A cells that overexpressed wild-type IL-25R dis-played the apoptotic response characterized by cleavage of caspase 3and PARP in a time-dependent manner (Fig. 7C), demonstrating thatIL-25R expression is sufficient to render these cells sensitive to apoptoticeffect of IL-25. In contrast, MCF10A cells that expressed the DDD IL-25Rmutant remained resistant to the IL-25 death signal. This finding in-dicates that the DD-like region of IL-25R is essential for mediating thedeath signal of IL-25, a finding that is consistent with the increasedassociation of IL-25R with DD adaptor proteins, FADD and TRADD,upon IL-25 treatment. Surprisingly, cells that expressed the DTRAF6IL-25R mutant showed an elevated level of apoptosis even in the ab-sence of IL-25 stimulation, suggesting that TRAF6 binding to IL-25Rconfers protection against apoptosis in MCF10A cells (Fig. 7C). These
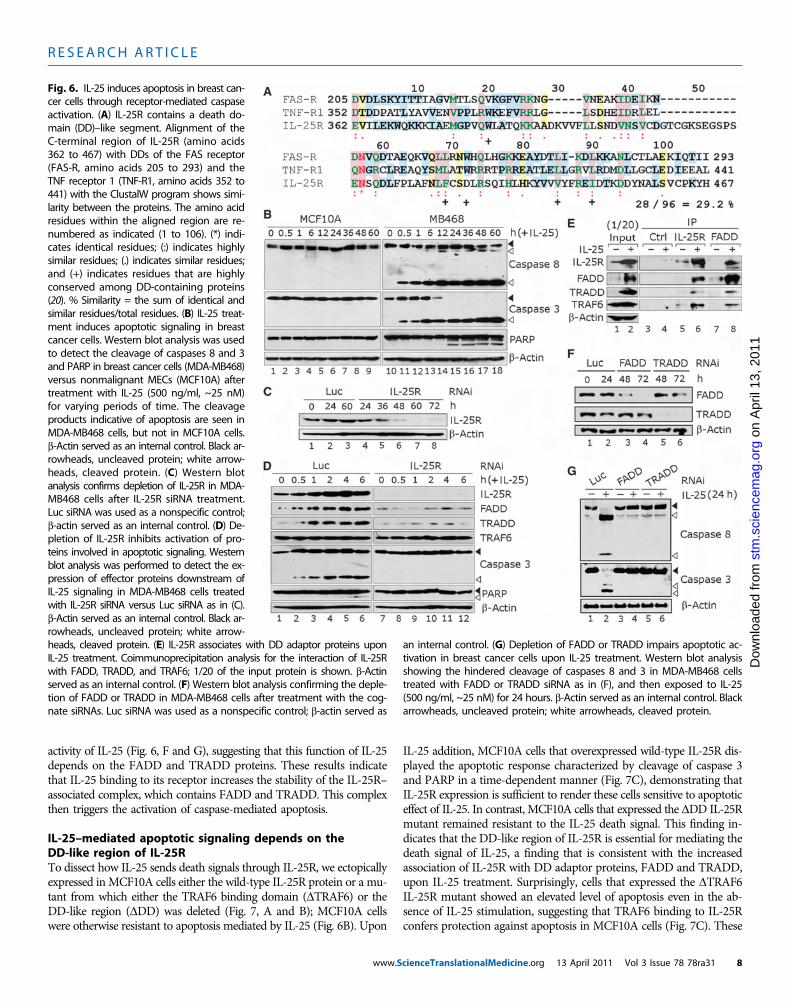
Fig. 6. IL-25 induces apoptosis in breast can-cer cells through receptor-mediated caspaseactivation. (A) IL-25R contains a death do-main (DD)–like segment. Alignment of theC-terminal region of IL-25R (amino acids362 to 467) with DDs of the FAS receptor(FAS-R, amino acids 205 to 293) and theTNF receptor 1 (TNF-R1, amino acids 352 to441) with the ClustalW program shows simi-larity between the proteins. The amino acidresidues within the aligned region are re-numbered as indicated (1 to 106). (*) indi-cates identical residues; (:) indicates highlysimilar residues; (.) indicates similar residues;and (+) indicates residues that are highlyconserved among DD-containing proteins(20). % Similarity = the sum of identical andsimilar residues/total residues. (B) IL-25 treat-ment induces apoptotic signaling in breastcancer cells. Western blot analysis was usedto detect the cleavage of caspases 8 and 3and PARP in breast cancer cells (MDA-MB468)versus nonmalignant MECs (MCF10A) aftertreatment with IL-25 (500 ng/ml, ~25 nM)for varying periods of time. The cleavageproducts indicative of apoptosis are seen inMDA-MB468 cells, but not in MCF10A cells.b-Actin served as an internal control. Black ar-rowheads, uncleaved protein; white arrow-heads, cleaved protein. (C) Western blotanalysis confirms depletion of IL-25R in MDA-MB468 cells after IL-25R siRNA treatment.Luc siRNA was used as a nonspecific control;b-actin served as an internal control. (D) De-pletion of IL-25R inhibits activation of pro-teins involved in apoptotic signaling. Westernblot analysis was performed to detect the ex-pression of effector proteins downstream ofIL-25 signaling in MDA-MB468 cells treatedwith IL-25R siRNA versus Luc siRNA as in (C).b-Actin served as an internal control. Black ar-rowheads, uncleaved protein; white arrow-
heads, cleaved protein. (E) IL-25R associates with DD adaptor proteins uponIL-25 treatment. Coimmunoprecipitation analysis for the interaction of IL-25Rwith FADD, TRADD, and TRAF6; 1/20 of the input protein is shown. b-Actinserved as an internal control. (F) Western blot analysis confirming the deple-tion of FADD or TRADD in MDA-MB468 cells after treatment with the cog-nate siRNAs. Luc siRNA was used as a nonspecific control; b-actin served asan internal control. (G) Depletion of FADD or TRADD impairs apoptotic ac-tivation in breast cancer cells upon IL-25 treatment. Western blot analysisshowing the hindered cleavage of caspases 8 and 3 in MDA-MB468 cellstreated with FADD or TRADD siRNA as in (F), and then exposed to IL-25(500 ng/ml, ~25 nM) for 24 hours. b-Actin served as an internal control. Blackarrowheads, uncleaved protein; white arrowheads, cleaved protein.
cienceTranslationalMedicine.org 13 April 2011 Vol 3 Issue 78 78ra31 8

R E S EARCH ART I C L E
on
Apr
il 13
, 201
1st
m.s
cien
cem
ag.o
rgd
from
results reveal the presence of two functionally distinct apoptotic reg-ulatory regions, a TRAF6-binding domain and a putative DD-like mo-tif, in the C terminus of IL-25R, demonstrating the intricacy of thesignaling cascades involved in ligand-activated IL-25R function.
Dow
nloa
de
DISCUSSION
It is long appreciated that the prevalence of cancer in human pop-ulations is far lower than one would predict based on DNA mu-tation rates (23). If oncogenic mutations are not sufficient toestablish tumors, how are tumors contained, suppressed, or elimi-nated before they become evident? A number of tumor surveillancemechanisms have been described, including the classic moleculartumor suppressors, immune surveillance, and suppression byECM and other microenvironmental factors. This study adds anew type of suppression to the list: factors secreted by normal dif-ferentiating cells that could kill or subdue their transformed coun-terparts.
We had shown previously that differentiating MECs secrete factorsthat allow phenotypic reversion of breast cancer cells, leading to for-mation of acinus-like structures and growth suppression (5). The se-creted factors could be partitioned into soluble and insoluble fractions;the former had tumor cell–killing activity and the latter had most ofthe “reverting” activity. Here, we identified six factors in the solublefraction that were secreted by differentiating MECs in 3D and shownto either kill or suppress the growth of tumor cells. This collection
www.ScienceTranslationalMedicine.or
of factors included antiangiogenic pro-teins (ATIII and VBP), proinflammatorycytokines (IL-1F7 and IL-25), and growthand differentiation proteins (FGF11 andBMP10). IL-25 exhibited the most potentcytotoxic activity toward breast cancer cells,whereas the other factors exhibited cytostat-ic activity. Here, we focused on the mech-anism of action of IL-25 and its potentialas a therapeutic agent in breast cancer.
IL-25 is a proinflammatory cytokinethat is expressed highly in certain organs,such as testis, prostate, and spleen, and isexpressed in low amounts in other organsincluding normal breast (11, 24). It is themost distant member of the IL-17 family ofproteins, sharing only 16 to 30% sequencehomology with the other family members(25). It plays a role in proinflammatoryresponses of lymphatic, kidney, and lungcells by inducing production of T helper2 (TH2)–type cytokines (11, 19, 24, 26).The function of IL-25 in other tissues re-mains to be elucidated.
We show here that IL-25 is temporallyup-regulated in developing normal mam-mary glands and induces caspase-mediatedapoptosis of breast cancer cells without af-fecting nonmalignant MECs either in cul-ture or in mice. The reason behind theresistance of nonmalignant cells to IL-25
is the differential expression of the receptor, IL-25R, high in breast can-cer cells but low or absent in nonmalignant MECs (Fig. 5, A and B).IL-25R overexpression contributes to tumorigenic potential, as shown bythe result that siRNA-induced reduction in the amounts of IL-25R im-paired breast cancer cells’ anchorage-independent growth in soft agar(Fig. 5, C to E). Examination of breast cancer specimens showed dis-tinct up-regulation of IL-25R in 19% of the samples, correlating stronglyin those with poor prognosis (Fig. 5I).
The exact mechanism by which IL-25R expression confers a growthadvantage to breast cancer cells remains to be determined. We postu-late that although IL-25R–expressing cancer cells do not express theapoptotic ligand IL-25 (Fig. 2A and fig. S4A), they may express anotherligand that contributes to their tumorigenic potential. Such a candidateligand appears to be IL-17B, which binds IL-25R with a markedly loweraffinity than that of IL-25 (11). We found that IL-17B was expressed inmost breast cancer cell lines that expressed high amounts of IL-25R,whereas IL-17B was absent from nonmalignant MCF10A cells (fig. S4A).Consistently, IL-17B was up-regulated in 30% of breast cancer speci-mens examined (12 of 40), but undetectable in normal tissues (0 of18) (fig. S4B). Small hairpin RNA (shRNA)–dependent reduction ofIL-17B amounts in MDA-MB468 breast cancer cells impaired theirgrowth and invasive potentials (fig. S4, C to G), whereas ectopic addi-tion of IL-17B protein enhanced both potentials (fig. S4, H to J). Theseresults in sum suggest that IL-17B may augment the tumorigenicity ofbreast cancer cells in an autocrine manner. This possibility is presentlyunder investigation. Whether IL-17B competes directly with IL-25 forreceptor binding is not known. However, it is known that both IL-25
Fig. 7. Death domain (DD)–like region of IL-25R renders cells sensitive to apoptotic signaling after treat-ment with IL-25. (A) Schematics for IL-25R protein expressed in mutation analyses. Wt, wild-type full-
length IL-25R protein; DTRAF6, mutated IL-25R with a deletion in TRAF6 binding domain (amino acidsD339–341); DDD, mutated IL-25R with a deletion in the DD-like region (amino acids D376–387). (B) West-ern blot shows that increased IL-25R amounts (Wt, DTRAF6, or DDD) after IL-25R were ectopicallyexpressed in MCF10A cells compared to the endogenous IL-25R amounts in parental cells (Ctrl). b-Actinserved as an internal control. (C) DD-like domain of IL-25R is essential for apoptotic signaling mediated byIL-25. Western blot analysis was used to detect cleavage of caspase 3 and PARP in MCF10A cells thatexpressed IL-25R as in (B) after treatment with IL-25 (500 ng/ml, ~25 nM) for varying time periods. b-Actinserved as an internal control. Black arrowheads, uncleaved protein; white arrowheads, cleaved protein. (D)Schematic for the cytotoxic activity of IL-25 specific to breast cancer cells that express IL-25R. Non-malignant MECs do not express IL-25R and are resistant to apoptosis induced by IL-25. Breast cancer cellsthat express IL-25R are susceptible to IL-25–induced apoptosis.g 13 April 2011 Vol 3 Issue 78 78ra31 9

R E S EARCH ART I C L E
on
Apr
il 13
, 201
1st
m.s
cien
cem
ag.o
rgD
ownl
oade
d fr
om
and IL-17B bind the extracellular domains of IL-25R in vitro, but thatthe IL-25 ligand shows a markedly higher affinity for the IL-25R thandoes IL-17B (11). Accordingly, it is speculated that IL-25 binding to thereceptor would outcompete IL-17B if both were present simulta-neously in the same cells. Some of our data, in fact, suggest that whenboth ligands are present, IL-25 binding to IL-25R is dominant overIL-17B binding to IL-25R. Figure S4A shows that IL-17B is expressedin most cancer cell lines tested. Yet, ectopic addition of IL-25 stillcauses the cells to apoptose. Nevertheless, the question could arise thatif IL-25 has such a strong affinity for IL-25R and this binding inducesapoptosis, why would tumors ever have a chance to form? We hypoth-esize that it is the localization and/or temporal availability of each ofthese ligands that accounts for the fact that IL-25R–expressing tumorsgrow rather than apoptose. That is, once a tumor is formed, if IL-17Bwere to be present in the tumor, it would act as a growth promoter.Because IL-25 is not generated by the tumor cells, IL-25–dependentapoptosis would be absent.
In lymphoid and renal cells, IL-25R activation by IL-25 induces aproinflammatory response mediated by TRAF6; in these two tissuecontexts, TRAF6 associates with its cognate binding domain in IL-25Rand activates nuclear factor kB (NF-kB), which in turn stimulates thetranscription of genes that encode inflammatory cytokines (11, 22). Incontrast, IL-25R activation by IL-25 in breast cancer cells causes the re-ceptor to interact with DD adaptor proteins FADD and TRADD, rap-idly activating caspases 8 and 3 sequentially for apoptotic signaling(Fig. 6, D to G). This action appears to be mediated by the DD-likeregion in the C terminus of IL-25R. Constitutive binding of TRAF6to IL-25R confers a protective effect on breast cancer cells, inhibitingapoptosis when IL-25 is not present (Fig. 7C). Such tissue-specific re-sponses to IL-25/IL-25R signaling may result from additional proteinsthat serve as switches between TRAF6/NF-kB signals and TRADD/FADD/caspase 8 signals. This type of molecular switching has beenshown previously for the related receptor, TNF-R1, which sharesabout 30% DD homology with IL-25R (Fig. 6A). TNF-R1 activationby TNF-a induces both NF-kB activation and apoptosis. However,NF-kB activation can be blocked by the brain- and reproductive organ–expressed (BRE) protein, which binds the juxtamembrane cytoplasmicregion of TNF-R1 and promotes apoptotic signaling (27). It is antici-pated that proteins with similar pathway-switching functions will bediscovered in the IL-25 pathway.
IL-25 thus holds promise as the basis for development of novel,effective breast cancer therapeutics with broad therapeutic windows.Our demonstration of the marked effect of IL-25 administration ininhibiting tumor growth with no apparent toxicity to the normal tis-sues in animals (Fig. 4, C and D) and the correlation of IL-25R ex-pression with poor prognosis in breast cancer patients support thiscontention. Unlike conventional immunotherapy in which a cytokine,such as IL-2, is administered by intravenous infusion (28) to provokeglobal immunologic responses (29), a therapy that targets IL-25/IL-25Rsignaling would induce apoptosis specifically in cancer cells that expressIL-25R. Expression of IL-25R in 19% of invasive ductal carcinomasplaces this receptor in a class of markers of interest for therapeutic tar-geting because of its prevalence, correlation with aggressiveness, andcell surface location. In these aspects, IL-25R could be compared toHer2/neu (30). Targeting Her2/neu has become an important strat-egy for treating HER2+ breast cancers by means of trastuzumab, amonoclonal antibody that binds and inhibits Her2/neu, resulting intumor regression. The findings reported here suggest that targeting
www.Sc
IL-25R in patients bearing tumors that overexpress this receptor shouldsimilarly result in strong clinical responses. We anticipate that our presentstudy will lead to the development of IL-25R–based diagnostics, facil-itating identification of the target population, and IL-25/IL-25R–basedtherapeutics, such as IL-25 peptidomimetics or IL-25R antagonistic anti-bodies, to efficiently and specifically induce target cell apoptosis in pa-tients with advanced breast cancer.
MATERIALS AND METHODS
See the Supplementary Material for the detailed Materials and Methods.
SUPPLEMENTARY MATERIAL
www.sciencetranslationalmedicine.org/cgi/content/full/3/78/78ra31/DC1Materials and MethodsTable S1. Oligonucleotide sequences used in this study.Fig. S1. Strong IL-25R staining in breast cancer cells as detected by a second IL-25R antibody.Fig. S2. Indirect immunoaffinity purification of IL-25.Fig. S3. Cell-killing activity of recombinant versus purified human IL-25 protein in breast cancercells in conventional cultures.Fig. S4. IL-17B is up-regulated in a subset of breast cancer cell lines and tissues and is impor-tant for their growth and invasive potentials.References
REFERENCES AND NOTES
1. S. Furuta, J. M. Wang, S. Wei, Y. M. Jeng, X. Jiang, B. Gu, P. L. Chen, E. Y. Lee, W. H. Lee, Removalof BRCA1/CtIP/ZBRK1 repressor complex on ANG1 promoter leads to accelerated mammarytumor growth contributed by prominent vasculature. Cancer Cell 10, 13–24 (2006).
2. X. L. Bourhis, R. A. Toillon, B. Boilly, H. Hondermarck, Autocrine and paracrine growth in-hibitors of breast cancer cells. Breast Cancer Res. Treat. 60, 251–258 (2000).
3. S. H. Barsky, N. J. Karlin, Myoepithelial cells: Autocrine and paracrine suppressors of breastcancer progression. J. Mammary Gland Biol. Neoplasia 10, 249–260 (2005).
4. T. Gudjonsson, L. Rønnov-Jessen, R. Villadsen, F. Rank, M. J. Bissell, O. W. Petersen, Normaland tumor-derived myoepithelial cells differ in their ability to interact with luminal breastepithelial cells for polarity and basement membrane deposition. J. Cell Sci. 115, 39–50(2002).
5. S. Furuta, X. Jiang, B. Gu, E. Cheng, P. L. Chen, W. H. Lee, Depletion of BRCA1 impairs differ-entiation but enhances proliferation of mammary epithelial cells. Proc. Natl. Acad. Sci. U.S.A.102, 9176–9181 (2005).
6. V. M. Weaver, O. W. Petersen, F. Wang, C. A. Larabell, P. Briand, C. Damsky, M. J. Bissell,Reversion of the malignant phenotype of human breast cells in three-dimensional cultureand in vivo by integrin blocking antibodies. J. Cell Biol. 137, 231–245 (1997).
7. S. K. Muthuswamy, D. Li, S. Lelievre, M. J. Bissell, J. S. Brugge, ErbB2, but not ErbB1, reinitiatesproliferation and induces luminal repopulation in epithelial acini. Nat. Cell Biol. 3, 785–792(2001).
8. X. Wang, C. F. Chen, P. R. Baker, P. L. Chen, P. Kaiser, L. Huang, Mass spectrometric char-acterization of the affinity-purified human 26S proteasome complex. Biochemistry 46,3553–3565 (2007).
9. Y. Cao, A. Lundwall, V. Gadaleanu, H. Lilja, A. Bjartell, Anti-thrombin is expressed in the be-nign prostatic epithelium and in prostate cancer and is capable of forming complexes withprostate-specific antigen and human glandular kallikrein 2. Am. J. Pathol. 161, 2053–2063(2002).
10. S. Kalkunte, L. Brard, C. O. Granai, N. Swamy, Inhibition of angiogenesis by vitamin D-bindingprotein: Characterization of anti-endothelial activity of DBP-maf. Angiogenesis 8, 349–360(2005).
11. J. Lee, W. H. Ho, M. Maruoka, R. T. Corpuz, D. T. Baldwin, J. S. Foster, A. D. Goddard, D. G. Yansura,R. L. Vandlen, W. I. Wood, A. L. Gurney, IL-17E, a novel proinflammatory ligand for the IL-17receptor homolog IL-17Rh1. J. Biol. Chem. 276, 1660–1664 (2001).
12. S. Kumar, C. R. Hanning, M. R. Brigham-Burke, D. J. Rieman, R. Lehr, S. Khandekar, R. B. Kirkpatrick,G. F. Scott, J. C. Lee, F. J. Lynch, W. Gao, A. Gambotto, M. T. Lotze, Interleukin-1F7B (IL-1H4/IL-1F7)is processed by caspase-1 and mature IL-1F7B binds to the IL-18 receptor but does not induceIFN-g production. Cytokine 18, 61–71 (2002).
ienceTranslationalMedicine.org 13 April 2011 Vol 3 Issue 78 78ra31 10

R E S EARCH ART I C L E
on
Apr
il 13
, 201
1tm
.sci
ence
mag
.org
13. P. M. Smallwood, I. Munoz-Sanjuan, P. Tong, J. P. Macke, S. H. Hendry, D. J. Gilbert, N. G. Copeland,N. A. Jenkins, J. Nathans, Fibroblast growth factor (FGF) homologous factors: New members ofthe FGF family implicated in nervous system development. Proc. Natl. Acad. Sci. U.S.A. 93,9850–9857 (1996).
14. H. Chen, S. Shi, L. Acosta, W. Li, J. Lu, S. Bao, Z. Chen, Z. Yang, M. D. Schneider, K. R. Chien,S. J. Conway, M. C. Yoder, L. S. Haneline, D. Franco, W. Shou, BMP10 is essential for maintainingcardiac growth during murine cardiogenesis. Development 131, 2219–2231 (2004).
15. S. Furuta, unpublished material.16. J. K. Kolls, A. Lindén, Interleukin-17 family members and inflammation. Immunity 21, 467–476
(2004).17. R. M. Neve, K. Chin, J. Fridlyand, J. Yeh, F. L. Baehner, T. Fevr, L. Clark, N. Bayani, J. P. Coppe,
F. Tong, T. Speed, P. T. Spellman, S. DeVries, A. Lapuk, N. J. Wang, W. L. Kuo, J. L. Stilwell,D. Pinkel, D. G. Albertson, F. M. Waldman, F. McCormick, R. B. Dickson, M. D. Johnson,M. Lippman, S. Ethier, A. Gazdar, J. W. Gray, A collection of breast cancer cell lines for the studyof functionally distinct cancer subtypes. Cancer Cell 10, 515–527 (2006).
18. D. C. Allred, J. M. Harvey, M. Berardo, G. M. Clark, Prognostic and predictive factors inbreast cancer by immunohistochemical analysis. Mod. Pathol. 11, 155–168 (1998).
19. S. Létuvé, S. Lajoie-Kadoch, S. Audusseau, M. E. Rothenberg, P. O. Fiset, M. S. Ludwig, Q. Hamid,IL-17E upregulates the expression of proinflammatory cytokines in lung fibroblasts. J. AllergyClin. Immunol. 117, 590–596 (2006).
20. K. Hofmann, J. Tschopp, The death domain motif found in Fas (Apo-1) and TNF receptor ispresent in proteins involved in apoptosis and axonal guidance. FEBS Lett. 371, 321–323 (1995).
21. S. J. Baker, E. P. Reddy, Modulation of life and death by the TNF receptor superfamily.Oncogene 17, 3261–3270 (1998).
22. Y. Maezawa, H. Nakajima, K. Suzuki, T. Tamachi, K. Ikeda, J. Inoue, Y. Saito, I. Iwamoto, Involve-ment of TNF receptor-associated factor 6 in IL-25 receptor signaling. J. Immunol. 176, 1013–1018(2006).
23. L. A. Loeb, Mutator phenotype may be required for multistage carcinogenesis. Cancer Res.51, 3075–3079 (1991).
24. M. R. Kim, R. Manoukian, R. Yeh, S. M. Silbiger, D. M. Danilenko, S. Scully, J. Sun, M. L. DeRose,M. Stolina, D. Chang, G. Y. Van, K. Clarkin, H. Q. Nguyen, Y. B. Yu, S. Jing, G. Senaldi, G. Elliott,E. S. Medlock, Transgenic overexpression of human IL-17E results in eosinophilia, B-lymphocytehyperplasia, and altered antibody production. Blood 100, 2330–2340 (2002).
25. T. A. Moseley, D. R. Haudenschild, L. Rose, A. H. Reddi, Interleukin-17 family and IL-17 recep-tors. Cytokine Growth Factor Rev. 14, 155–174 (2003).
26. M. M. Fort, J. Cheung, D. Yen, J. Li, S. M. Zurawski, S. Lo, S. Menon, T. Clifford, B. Hunte,R. Lesley, T. Muchamuel, S. D. Hurst, G. Zurawski, M. W. Leach, D. M. Gorman, D. M. Rennick,IL-25 induces IL-4, IL-5, and IL-13 and Th2-associated pathologies in vivo. Immunity 15, 985–995(2001).
www.Sc
27. C. Gu, A. Castellino, J. Y. Chan, M. V. Chao, BRE: A modulator of TNF-a action. FASEB J. 12,1101–1108 (1998).
28. M. B. Atkins, M. T. Lotze, J. P. Dutcher, R. I. Fisher, G. Weiss, K. Margolin, J. Abrams, M. Sznol,D. Parkinson, M. Hawkins, C. Paradise, L. Kunkel, S. A. Rosenberg, High-dose recombinant in-terleukin 2 therapy for patients with metastatic melanoma: Analysis of 270 patients treatedbetween 1985 and 1993. J. Clin. Oncol. 17, 2105–2116 (1999).
29. C. Jackaman, C. S. Bundell, B. F. Kinnear, A. M. Smith, P. Filion, D. van Hagen, B. W. Robinson,D. J. Nelson, IL-2 intratumoral immunotherapy enhances CD8+ T cells that mediate destruc-tion of tumor cells and tumor-associated vasculature: A novel mechanism for IL-2. J. Immunol.171, 5051–5063 (2003).
30. C. A. Hudis, Trastuzumab—mechanism of action and use in clinical practice. N. Engl. J. Med.357, 39–51 (2007).
31. Acknowledgments:We are grateful to C. Hines and R. Wei for providing reagents and J. Mott,A. Lo, E. Y. Lee, A. D. Borowsky, and D. Brownfield for helpful comments. We thank E. Lee andX. Tian for technical assistance and R. Baehner and L. Chan (UCSF Comprehensive CancerCenter) for pathological analyses of the second cohort. Funding: This work was supported bygrants from the NIH (RO1CA94170 to W.-H.L. and GM-74830 to L.H.), the National Cancer Institute(R37CA064786, U54CA126552, R01CA057621, U54CA112970, U01CA143233, and U54CA143836to M.J.B.), the U.S. Department of Energy (DE-AC02-05CH1123 to M.J.B.), the U.S. Departmentof Defense (W81XWH0810736 to M.J.B. and W81XWH-05-1-0322 to S.F.), and a physician scien-tist award from the National Health Research Institute in Taiwan (to Y.-M.J.). Author contribu-tions: W.-H.L. and M.J.B. were responsible for supervising the project and helping design theexperiments. I.K. contributed to designing some of the control experiments. S.F. was responsiblefor designing and executing most of the experiments. Y.-M.J. analyzed the first cohort of breastcancer tissue sections and patients’ clinical data. L.Z. performed mouse xenograft experiments.L.H. performed liquid chromatography–tandem mass spectrometry analysis. S.F., W.-H.L., M.J.B.,and I.K. contributed to writing and revising the manuscript. Competing interests: A patent(PCT/US2007/081395) has been filed related to the use of IL-25 for cancer treatment. All au-thors declare that they have no competing interests.
Submitted 11 June 2010Accepted 18 March 2011Published 13 April 201110.1126/scitranslmed.3001374
Citation: S. Furuta, Y.-M. Jeng, L. Zhou, L. Huang, I. Kuhn, M. J. Bissell, W.-H. Lee, IL-25 causesapoptosis of IL-25R–expressing breast cancer cells without toxicity to nonmalignant cells. Sci.Transl. Med. 3, 78ra31 (2011).
s
ienceTranslationalMedicine.org 13 April 2011 Vol 3 Issue 78 78ra31 11
Dow
nloa
ded
from

www.sciencetranslationalmedicine.org/cgi/content/full/3/78/78ra31/DC1
Supplementary Materials for
IL-25 Causes Apoptosis of IL-25R–Expressing Breast Cancer Cells Wit hout Toxicity to Nonmalignant Cells
Saori Furuta, Yung-Ming Jeng, Longen Zhou, Lan Huang, Irene Kuhn, Mina J. Bissell,*
Wen-Hwa Lee*
*To whom correspondence should be addressed. E-mail: [email protected] (M.J.B.); [email protected] (W.-H.L.)
Published 13 April 2011, Sci. Transl. Med. 3, 78ra31 (2011)
DOI: 10.1126/scitranslmed.3001374
The PDF file includes:
Materials and Methods Table S1. Oligonucleotide sequences used in this study. Fig. S1. Strong IL-25R staining in breast cancer cells as detected by a second IL-25R antibody. Fig. S2. Indirect immunoaffinity purification of IL-25. Fig. S3. Cell-killing activity of recombinant versus purified human IL-25 protein in breast cancer cells in conventional cultures. Fig. S4. IL-17B is up-regulated in a subset of breast cancer cell lines and tissues and is important for their growth and invasive potentials. References

1
Supplementary Information
MATERIAL AND METHODS
Cell cultures
Nonmalignant human mammary epithelial MCF10A cells were cultured as described (1); human
breast cancer cell lines (MCF7, MDA-MB361, T47D, ZR75, MDA-MB468, MDA-MB435-S,
MDA-MB231, MDA-MD175-7, SKBR3, HS578T, HBL100 and HCC1937) and human
embryonic kidney cells 293T were cultured as described (2) .
Microscopic imaging
Microscopic imaging of live cells was performed on a Zeiss Axiovert 200 M equipped with
Hamamatsu Photonics K.K. Deep Cooled Digital Camera using Axiovision 4.5 software (Carl
Zeiss) with phase I at 100X or phase II reflector at 200X magnification. Photomicrographs of
histology specimens were taken with Zeiss Axioplan 2 Imaging platform and AxioVision 4.4
Software at 100X or 400X magnification.
Assessment of the cytotoxicity of CM from MECs
MCF10A cells were plated at 4x104 cells in a 35mm-dish coated with 1ml Growth Factor
Reduced Matrigel (BD Biosciences) and covered with 3ml growth medium supplemented with
2% Matrigel as described (1). After 15hrs CM (2.5ml) was collected and separated into soluble
and pelleted fractions by centrifugation at 14,000 for 30min. The soluble fraction was size-
fractionated with Centricon-10 and -50 units (Millipore) following the manufacturer’s
instruction; the pellet was resuspended in 400µl of growth medium. 400µl each of the following

2
fractions were obtained: 1) total CM, 2) pellet, 3) total supernatant, soluble fractions of 4)
>50kDa, 5) 10-50kDa and 6) <10kDa. All the fractions were reconstituted with the essential
growth factors and 2% Matrigel and applied to MCF7 or MCF10A cells seeded at 5000
cells/well in Matrigel-coated 8-well chamber slides. The collection/application of CM was
performed every 12~15hrs for 1 week. To determine when differentiating MECs produce
cytotoxic factors, the 10-50kDa fraction of CM was harvested at each day (days 0-6) from
differentiating MCF10A in 3D lrECM using the above-mentioned condition. Different day
fractions were individually applied to MCF7 cells plated at 1000 cells/well in Matrigel-coated
96-well plate. Fresh CM was applied every 24hrs. For cell number counting, cells were
recovered from Matrigel after 1 week by digestion with dispase (BD Biosciences), and the viable
cell numbers were measured using trypan blue exclusion method.
Sample preparation for mass spectrometric analysis
The cytotoxic fraction (10-50kDa, day 4) of CM from differentiating MCF10A cells and the
soluble fraction of CM from MCF7 cells in 3-D laminin-rich ECM gels (day 4) were collected.
Proteins in each medium were concentrated by trichloroacetic acid (TCA) precipitation and
dissolved in boiling SDS sample buffer. Proteins were resolved by SDS-PAGE (10%) and
visualized with Coomassie Blue staining. Gel slices (2mm thickness) were excised, de-stained
with 25mM NH4HCO3 in 50% MeOH and digested with 50ng/ml trypsin in 50mM NH4HCO3
for 24 h at 37 °C. Peptides were extracted from gel slices with 3 volume of 50% acetonitrile,
vacuum-dried and resuspended in 0.1% formic acid. Following sample clean up in C18 ZipTips
(Mil lipore), peptides were eluted with 0.1% formic acid in 50% acetonitrile.

3
Liquid Chromatography and Tandem Mass Spectrometry (LC-MS/MS)
The cytotoxic fraction (10-50kDa, day 4) of CM from differentiating MCF10A cells and the
soluble fraction of CM from MCF7 cells in 3D matrix (day 4) were analyzed for LC-MS/MS
spectrometry as described (3, 4). For LC-MS/MS analysis, the digests were first separated by
cation exchange chromatography (polysulfoethyl A column, Nest Group) using a linear gradient
between solvents A (5mM KH2PO4, 30% acetonitrile, pH 3) and B (solvent A with 350mM KCl)
at a flow rate of 0.2 ml/min. Fractions were collected on the basis of UV absorbance (215 and
280nm) and desalted with C18 micro spin columns (Vivascience). LC-MS/MS was carried out
by nanoflow reverse phase liquid chromatography (RPLC) (Ultimate LC Packings) coupled on-
line to QSTAR XL tandem mass spectrometer (Applied Biosystems). RPLC was performed
using a capillary column (75µm x 150mm) packed with Polaris C18-A resin (Varian Inc.), and
the peptides were eluted using a linear gradient between solvents A (2% acetonitrile, 0.1%
formic acid) and B (98% acetonitrile, 0.1% formic acid) at a flow rate of 250 nl/min. Each full
MS scan was followed by three MS/MS scans where three most abundant peptide ions were
selected to generate tandem mass spectra. Two LC-MS/MS runs were performed on the same
sample to improve the dynamic range of mass spectrometric analysis.
Protein identification
For MS data analysis, monoisotopic masses (m/z) of peptide ions were obtained from the tandem
mass spectra using Mascot script in Analyst QS version 1.1 software (Applied Biosystems) with
the mass accuracy of ±200ppm. Certain chemical modifications (i.e., N-terminal acetylation or
pyroglutamine, methionine oxidation, asparagine deamination, carbamylation of the N-terminus
and lysine, phosphorylations of serine, threonine and tyrosine) were selected as variables during

4
the peptide search using Batch-Tag program in Protein Prospector version 4.25.0 software
(UCSF). Both Uniprot and NCBInr public databases were queried to identify the proteins. Search
Compare program in Protein Prospector was used to summarize the results including protein
scores and discriminating score among peptide fragments. The result obtained for CM from
differentiating MCF10A cells was compared to that for CM from MCF7 cells in 3-D cultures,
and proteins present only in the former sample were identified. Proteins with the best score (>20)
and discriminating score <6 were considered significant.
Immunodepletion
The cytotoxic fraction (10-50kDa, days 3-5) of CM was harvested from differentiating MCF10A
cells in 3-D culture. The medium was divided into six fractions (2ml each), and each fraction
was clarified with 100µl of protein G sepharose beads at 4°C for 2hrs. One µg of antibody
against BMP10, FGF11, ATIII, IL1F7, IL-25, VBP or p84 (Ctrl) was added to each fraction and
incubated at 4ºC overnight. Antibody-protein complex was precipitated by 100µl protein A/G
sepharose beads (1:1) at 4°C for 2hrs. The immunoprecipitates were washed in TEN buffer
(10mM Tris-HCl (pH8.0), 0.25mM EDTA, 50mM NaCl) supplemented with 0.1% NP-40 and
protease inhibitors, then analyzed by western blot. 1/20 of the immunodepleted fraction was also
examined by western analysis, and depletion was repeated 3 times to ensure complete loss of a
target protein. Depleted fractions were reconstituted with the essential growth factors and 2%
Matrigel, then used to plate MCF7 cells seeded at 5000 cells/well in Matrigel-coated 8-well
chamber slides. The fraction was applied every 24 hrs for 1 week. Cells were recovered from
Matrigel, and the viable cell numbers were measured.

5
Stable cell lines for ATIII, IL1F7, IL-25 and VBP
Full-length cDNA clones of ATIII, IL1F7, VBP (pDR-LIB) and IL-25 (pPCR-Script/Amp) were
obtained from ATCC. ATIII, IL1F7 and VBP cDNAs were excised at SmaI/XhoI sites and
subcloned into EcoRV/XhoI sites of pcDNA3.1/Hyg vector (Invitrogen), while IL-25 cDNA was
excised at HindIII/NotI and subcloned into pcDNA3.1/Hyg. 293T cells were transfected with the
respective plasmid and selected with 70µg/ml Hygromycin B (Roche). The comparable
expression levels of these proteins were confirmed by RT-PCR of the transcripts using primers
shown in Table.S1 and western analysis of secreted proteins in the conditioned media. For
determining the cytotoxic activity of each factor, 7ml of CM from 293T cells (4x106) was
harvested, concentrated by 2 fold with Centricon-10 and normalized based on the total protein
amount. The media were supplemented with growth factors plus 2% Matrigel and applied to
MCF7 cells seeded at 5000 cells/well in Matrigel-coated 8-well chamber slides. Fresh CM was
applied every 24 hrs for one week, and the viable cell numbers were counted. To generate a
stable cell line for IL-25 purification, IL-25 cDNA was subcloned into BamHI site of pQCXIH
retroviral vector (BD Biosciences). GP2-293 packaging cells were transfected with
pQCXIH/IL25 and pVSVG plasmids using Lipofectin, and retrovirus was harvested in the
conditioned medium. 293T cells were infected with retrovirus using 8µg/ml polybrene, and the
stable clones were selected with 70 µg/ml Hygromycin B (Roche). IL-25 retrovirus was
generated to establish a stable 293T cell line as described (5).
Purification of secreted IL-25 by concanavalin A column followed by gel filtration column
CM was collected from a stable 293T cell line expressing IL-25, supplemented with protease
inhibitors and loaded onto a column packed with concanavalin A-sepharose beads (CalBiochem)

6
pre-equilibrated with column buffer (10mM Tris (pH7.5), 150mM NaCl, 1mM CaCl2, 1mM
MnCl2). The column was washed with column buffer, and bound glycosylated proteins were
eluted with 0.5M α-methyl mannose in column buffer. The eluates were pooled, concentrated
with Centricon-10 and then separated by Superdex 200 gel filtration column (HR 10/30, 24mL;
Amersham Pharmacia) using elution buffer (50mM Na2HPO4 (pH7.5), 50mM NaCl) at a flow
rate of 0.4ml/min. Fractions were collected based on UV absorbance at 280nm and resolved on
10% SDS-PAGE for western analysis.
Purification of secreted IL-25 by indirect immunoaffinity chromatography
The IL-25 antibody was indirectly conjugated with agarose beads as follows. 1 mg of IL-25
antibody was dissolved in 2ml PBS and incubated with 1ml slurry of protein G- agarose (~50%)
for 1h at RT. Beads were washed in 5ml sodium borate (0.2M, pH 9.0), resuspended in 5ml
sodium borate supplemented with dimethylpimelimidate up to 20 mM, and incubated for 30 min
at RT. The reaction was stopped by 5ml 0.2M glycine (pH 8.0); the supernatant was drained and
replaced with a fresh 5ml 0.2M glycine. After 2h of incubation at RT, beads were suspended in
3ml PBS and transferred to a chromatography column. CM collected from a stable 293T cell line
expressing IL-25 was supplemented with protease inhibitors and pre-cleared through a column
pre-packed with 2 ml protein A/G (1:1) agarose. The flow-through was then loaded onto the
immunoaffinity column at a flow rate of 1 drop (~ 20 µl)/sec. The resin was washed with 10ml
each of PBS, wash buffer [10 mM Tris (pH 8.0), 140 mM NaCl, 0.025% NaN3, 0.5% Triton X-
100, 0.5% sodium deoxycholate], and then sodium phosphate (pH 6.3) buffer [50mM sodium
phosphate (pH 6.3), 0.1% Triton X-100, 0.5M NaCl]. The bound IL-25 protein was eluted in 20

7
ml glycine buffer [50 mM glycine (pH 2.5), 0.1% Triton X-100, 0.15M NaCl] , and neutralized
with 4 ml 1 M Tris (pH 9.0), and concentrated with Centricon-30 unit.
Colony formation assay
Breast cancer cells (MCF7, MDA-MB468, SKBR3 and T47D) at 1000/well and MCF10A cells
at 500/well were seeded in 6-well plates in triplicate and maintained for 24hrs. Designated
amounts of IL-25 were diluted in elution buffer to 100µl and then in 900µl growth medium to
culture cells. After 10 days cells were stained with 2% Methylene Blue in 50% EtOH, and the
numbers of colonies were counted.
Tumor inhibition assay in nude mice
Animal experiments were performed under federal guidelines and approved by Institutional
Animal Care and Use Committee at UCI. Exponentially growing MDA-MB-468 cells at 10x106
in 100µl of DMEM plus 5% Matrigel were injected into the mammary fat pad of 6-8 week old
athymic female BALB/c-nude mice (nu/nu) (Charles River Laboratories). After tumors grew to
80mm3 (day 10), mice were randomized into control (n=7) and experimental groups (n=8) to
receive on site injection of vehicle (elution buffer, 50µl) or IL-25 (300ng, 50µl) once every day
for 31 days. Mouse body weights and tumor volumes were measured twice weekly. Student’s t
test was used to determine p-value as indicated in the figure legend. At the end of experiments,
mice were sacrificed and subjected to pathological examinations of different organs, including
intestines, kidney, heart, lungs, liver and stomach, to test the presence of systemic inflammation.
Tumor xenografts were excised for histological examinations. For the safety test, IL-25 (1.5µg)

8
was injected into the tail veins of female C57 mice (vehicle: n=3; IL-25: n=5), and signs for
systemic stress (e.g., lethargy and weight loss) were monitored daily.
Breast tumor samples
Patient cohort 1: All cases were invasive ductal carcinomas, which account for more than 90% of
archived breast cancer samples in National Taiwan University Hospital. The grades of these
tumors were 17 grade 1, 40 grade 2, and 12 grade 3 according to Nottingham Histologic Score.
38 were stage 1, 20 stage 2, and 11 stage 3 according to AJCC staging system. Patient cohort 2:
All tumor cases were invasive ductal carcinomas between stages II and III according to Scarf-
Bloom-Richardson system. Normal tissues were obtained from reduction mammoplasty samples.
These tissues were provided by the Cooperative Human Tissue Network (NCI).
Histology and immunohistochemistry
Dissected tumors were fixed in 4% paraformaldehyde overnight and embedded in paraffin with a
tissue processor. 4-5µm sections were deparaffinized and hydrated. Tumor xenografts were
stained with hematoxylin and eosin, while human breast tumor specimens were processed for
immunohistochemistry. Antigen retrieval was performed in 0.01 M citric buffer at 100°C for 10
minutes. After being blocked in 3% H2O2 and nonimmune horse serum, the slides were reacted
with a monoclonal antibody against human IL-25R (GeneTex CAT # GTX13653; 1:100 dilution)
at 4°C overnight and incubated with link antibodies, followed by peroxidase conjugated
streptavidin complex (LSAB kit, DAKO Corp.) The peroxidase activity was visualized with
diaminobenzidine tetrahydroxychloride solution (DAB, DAKO) as the substrate. The sections
were lightly counterstained with hematoxylin. The positivity of staining was determined by

9
Allred Immunohistochemistry Scoring system (6), and more than 5% tumor cells stained in each
section (score >2+) was defined as positive by a trained pathologist. The survival curve of
patients was obtained by Kaplan-Meier analysis using XLSTAT-Life Version 2007.4 software.
Statistical significance was assessed by using the log-rank test. A second set of breast tissue
samples (10 normal and 20 malignant) were prepared as frozen sections (8-10 µm), fixed in 4%
paraformaldehyde for 20 min, stained with a different IL-25R antibody (R&D Systems CAT #
MAB1207; 1:50 dilution). NOTE: The manufacturer (GeneTex) of the IL-25R antibody clone
used for staining paraffin sections as described immediately above replaced it with another
clone; however, our testing showed that this replacement clone did not have the same
immunoreactivity as the first clone listed under the same catalog number and thus we chose not
to use this second clone from GeneTex. We found that by western blot, IP, detection of IL-25R
in knock-down and overexpression experiments the results obtained using the IL-25R antibody
available from R&D Systems were interchangeable with those obtained using the first clone of
GeneTex antibody. The only relevant difference being that the R&D antibody could not be used
for IHC on paraffin-embedded sections, and that is why the IHC analysis using the R&D
antibody was performed using frozen sections.
IL25R, FADD or TRADD siRNA treatment
Breast cancer cells or nonmalignant MCF10A cells were plated at 3.5x105/60mm dish and
maintained for 24 hrs. Cells were transfected with 400pmol of annealed siRNA for IL-25R,
FADD or TRADD (Table S1, Qiagen) using Oligofectamine (Invitrogen) according to
manufacturer’s instruction. Luciferase siRNA was used as a non-specific control.

10
Soft agar assay
One % agar was mixed with the equivalent volume of 2x DMEM plus 20% FBS and 2%
penicillin/streptomycin. One ml of the agar solution was poured into a 35 mm plate in triplicate
and solidified. 0.7% agar solution equilibrated to 40°C was mixed with 2x growth medium and
breast cancer cells at 4000 cells/ml and poured onto the base agar at 1 ml/plate. The solidified
agar was covered with 500µl growth medium and maintained in 37°C humidified incubator for 2
weeks. The plates were stained with 0.01% crystal violet for 30 min, and colonies were counted
under dissecting microscope.
Immunoprecipitation
Nine mg of whole cell lysates were collected in 3ml of Triton lysis buffer (25mM Tris (pH7.6),
150mM NaCl, 1% TritonX-100) supplemented with protease and phosphatase inhibitors. The
lysate was divided into three fractions (3mg protein/1ml each), and each fraction was clarified
with 50µl of protein G sepharose beads at 4°C for 2hrs. Two µg of antibody against p84 (Ctrl),
IL-25R or FADD (Cell Signaling) was added to each fraction and incubated at 4ºC overnight.
Antibody-protein complex was precipitated by 50µl protein A/G sepharose beads at 4°C for 2hrs
and washed in TEN buffer with 0.1% NP-40 and protease inhibitors. Immunoprecipitates were
resolved on 12% SDS-PAGE and detected by western analysis.
IL25R expression constructs
The expression construct of human IL-25R in pCMV6-XL5 vector was obtained from OriGene.
The wild-type IL-25R was subjected to mutagenesis to generate a siRNA-resistant mutant (RM),

11
a deletion mutant at TRAF6 binding domain (∆TRAF6) or death domain-like region (∆DD) by
using a corresponding primer pair (Table S1). Correct mutagenesis was validated by sequencing.
IL17B shRNA construction. Forward and reverse oligonucleotides (see supplementary Table
S1 for the sequences) were annealed to generate a double-stranded DNA oligonucleotide, which
in turn was ligated into BamH1/EcoR1 site of pGreen puro lentiviral vector (System
Biosciences). For control, luciferase shRNA vector was used (System Biosciences). Lentivirus
production and transduction of target cells were conducted following the guideline by the
manufacturer. Briefly, lentivirus vector and packaging plasmid mix (System Biosciences) were
transfected into 293FT cells (Invitrogen) using Lipofectamine® 2000. After 48 hours, medium
was harvested, filtered and used to infect target cells with the addition of polybrene (10 µg/ml).
After 24 hours medium was replaced. At 72 hours post-infection puromycin (0.5 µg/ml) was
added for selection and maintained throughout the culturing period.
Cell proliferation assay. Cells were plated at a density of 1 x 103 cells per well in 96-well
plates in DMEM plus 10% FBS and incubated at 37°C. Twenty-four hours before each time
point, the medium was replaced. At each time point, MTT (3-[4,5-dimethyl-2-thiazolyl]-2,5-
diphenyl-2H-tetrazolium bromide, Sigma-Aldrich) was added to cells to the final concentration
of 1.6 mg/ml, and the reaction was incubated at 37°C for 4 hours. Then, the medium was
removed and the precipitated reaction product was dissolved in MTT solvent (4 mM HCl, 0.1%
NP-40 in isopropanol). The absorbance was measured at 570 nm.

12
Invasion assays. Cells at 2.5 X 104 were placed on the top of a thin Matrigel layer in 24-well
invasion chamber (BD Biosciences) according to the manufacturer’s protocol. After 48 hours,
cells were stained with 0.2% methylene blue in 50% ethanol. Samples were prepared in triplicate
and invaded cells were counted on at least 3 different fields on the trans-well filters. Statistical
analysis was performed using an unpaired t-test. A p-value <0.05 was considered to be
significant.

13
Supplementary data
Table S1. Oligonucleotide sequences used in this study.
1Unique NdeI restriction site generated is shown in italics. 2Unique SacI restriction site generated is shown in italics. 3Unique NheI restriction site generated is shown in italics. 4Nucleotides substituted without changing the encoded amino acids are shown underlined.
Name F/R Sequence
RT-PCR Primers ATIII RT F 5’-GCT TTT GCT ATG ACC AAG CTG-3’ R 5’-TGC TTC ACT GCC TTC TTC ATT-3’ IL1F7 RT F 5’-AAA CCC GAA GAA ATT CAG CAT-3’ R 5’-CCC ACC TGA GCC CTA TAA AAG-3’ IL25 RT F 5’-TTC CTA CAG GTG GTT GCA TTC-3’ R 5’-CGC CTG TAG AAG ACA GTC TGG-3’ mIL25 RT F 5’-ATG TAC CAG GCT GTT GCA TTC TTG-3’ R 5’-CTA AGC CAT GAC CCG GGG CC-3’ VBP RT F 5’-AAT CAA GGC TCA GCA ATC TCA-3’ R 5’-CAT CTT TGT TTG TGG GCA ACT-3’ IL25R RT F 5’-AGA GGC CTT CCA GAC TCA GAC-3’ R 5’-AAA CCC GAT GAT AGT GCT GTG-3’ IL17B F 5’-CCT CAC AAC CTG CTG TTT CTT-3’ R 5’-ACA GCT GCA AGT TGA CCT CAC-3’ α-tubulin F 5’-TGA CCT GAC AGA ATT CCA GAC CA-3’ R 5’-GCA TTG ACA TCT TTG GGA ACC AC-3’ m18S F 5’-TCG GAA CTG AGG CCA TGA TT-3’ R 5’-CCT CCG ACT TTC GTT CTT GAT T-3’ mE-cadherin F 5’-CGA GAG AGT TAC CCT ACA TA-3’ R 5’-GTG TTG GGG GCA TCA TCA TC-3’ siRNA IL25R F 5’-r(CGC GAG CUU CAG UGG UGA U)dTdT-3’ R 5’-r(AUC ACC ACU GAA GCU CGC G)dTdT-3’ FADD F 5’- r(GAA GAC CUG UGU GCA GCA U)dTdT-3’ R 5’-r(AUG CUG CAC ACA GGU CUU C)dTdT-3’ TRADD F 5’- r(GGU CAG CCU GUA GUG AAU C)dTdT-3’ R 5’-r(GAU CA CUA CAG GCU GAC C)dTdT-3’ shRNA IL17B F 5’-GAT CCG TGG AGG AGT ATG AGA GGA ACT TCC TGT CAG ATT CCT
CTC ATA CTC CTC CAC TTT TTG-3’ R 5’-AAT TCA AAA AGT GGA GGA GTA TGA GAG GAA TCT GAC AGG AAG
TTC CTC TCA TAC TCC TCC ACG-3’ Mutagenesis IL25R ∆TRAF61 F 5’-ATT AAG GTT CTT GTG GTT TAC ATA TGT TTC CAT CAC ACA ATA-3’ R 5’-TAT TGT GTG ATG GAA ACA TAT GTA AAC CAC AAG AAC CTT AAT-3’ IL25R ∆DD2 F 5’-TGG CAG AAA AAG AAA ATA GCA GAG CTC GCA GCA GAC AAA GTC
GTC TTC CTT-3’ R 5’-AAG GAA GAC GAC TTT GTC TGC TGC GAG CTC TGC TAT TTT CTT TTT
CTG CCA-3’ IL25R RM3,4 F 5’-CAG AAG AAA CAA ACG CGG GCT AGC GTG GTG ATT CCA GTG ACT
GGG-3’ R 5’-CCC AGT CAC TGG AAT CAC CAC GCT AGC CCG CGT TTG TTT CTT
CTG-3’
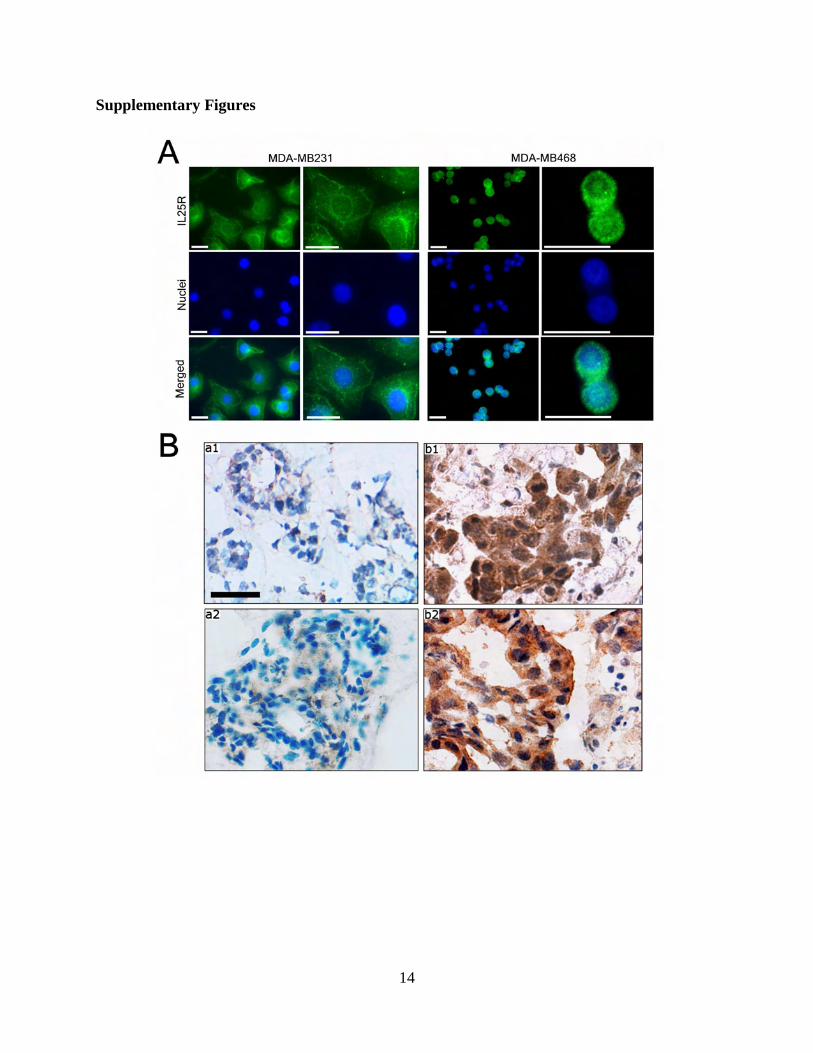
14
Supplementary Figures

15
Fig. S1. Strong IL-25R staining in breast cancer cells as detected by a second IL-25R antibody.
(A) Cultured MDA-MB231 and MDA-MB468 breast cancer cells were immunostained against
IL-25R with a monoclonal antibody from R&D Systems and subjected to immunofluorescent
imaging. Note the accentuated staining at cell membranes as well as Golgi. (B) Frozen sections
of normal human breast tissue (left, N=10) vs. breast cancer tissue (right, N=20) sections were
immunostained against IL-25R with the same antibody used in (A) and subjected to
immunohistochemical imaging. The staining specificity of this antibody is the same as that
observed by the antibody from GenTex; however, the membrane staining shown here is frozen
sections is not as accentuated or consistent as that shown using IHC on paraffin sections (Fig.
5H). Scale bar: 50µm.

16
Fig. S2. Indirect immunoaffinity purification of IL-25. (Left): Western blot of IL-25 purified by
acidic (low pH) elution. Conditioned medium from IL-25 expressing 293T cells was passed
through Anti-IL25 antibody-protein G-agarose column, and the column was washed with pH 8
and then pH 6.3 buffers. The bound protein(s) were eluted by a low pH buffer (pH 2.5)
containing glycine and then neutralized immediately. The eluate was size-fractionated by a series
of centrifugal filters (10, 50 and 100 kDa cut-offs.) Native Il-25 was enriched in the fraction
>100 kDa, possibly due to multimerization. (Right): The SDS-PAGE for samples corresponding
to those analyzed by western blot (left) after staining with Coomassie dye. The IL-25 protein
eluted in the >100 kDa fraction is fairly pure (>50%; 45~50 kDa as a monomer) with the
approximate concentration of ~80 ng/µl (lane 9).
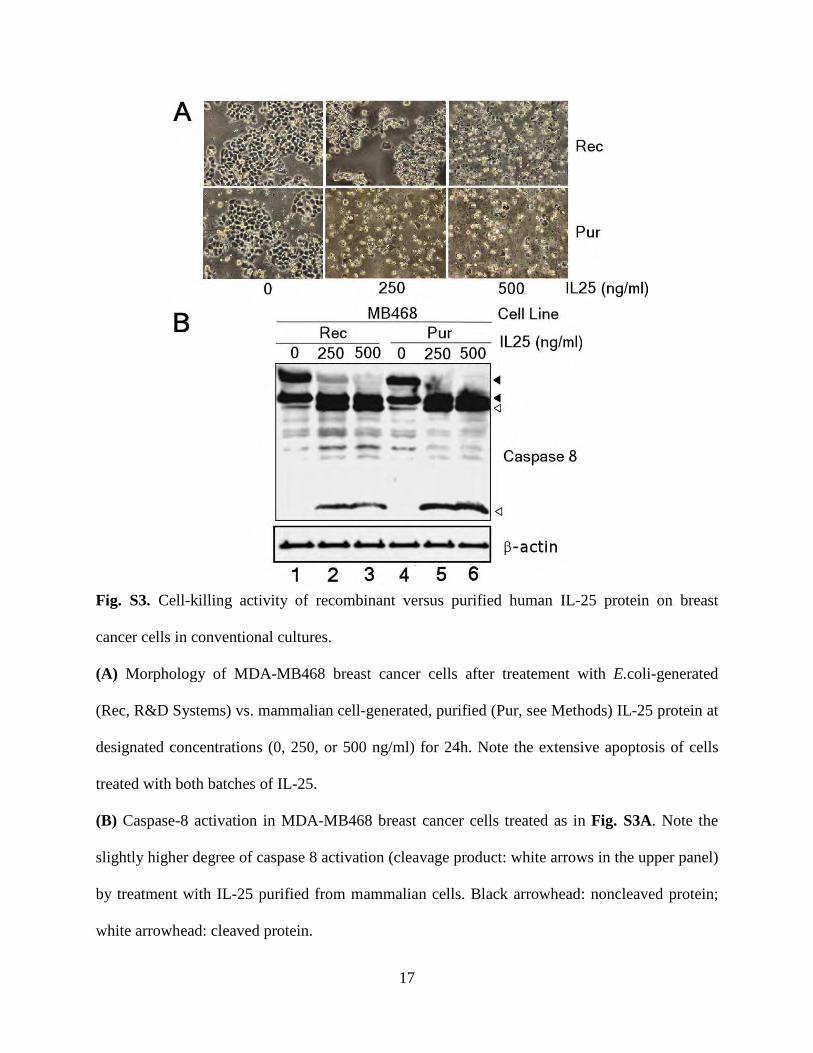
17
Fig. S3. Cell-killing activity of recombinant versus purified human IL-25 protein on breast
cancer cells in conventional cultures.
(A) Morphology of MDA-MB468 breast cancer cells after treatement with E.coli-generated
(Rec, R&D Systems) vs. mammalian cell-generated, purified (Pur, see Methods) IL-25 protein at
designated concentrations (0, 250, or 500 ng/ml) for 24h. Note the extensive apoptosis of cells
treated with both batches of IL-25.
(B) Caspase-8 activation in MDA-MB468 breast cancer cells treated as in Fig. S3A. Note the
slightly higher degree of caspase 8 activation (cleavage product: white arrows in the upper panel)
by treatment with IL-25 purified from mammalian cells. Black arrowhead: noncleaved protein;
white arrowhead: cleaved protein.

18

19
Fig. S4. IL-17B is up-regulated in a subset of breast cancer cell lines and tissues and is important
for their growth and invasive potentials.
(A) Opposing expression patterns of IL-25 and IL-17B in nonmalignant MECs (MCF10A) and
breast cancer cells detected by RT-PCR. α-tubulin (α-TUB) serves as an internal loading control.
(B) Specimens of nontumorous human breast tissue (a1-2) vs. human breast cancer (b1-2) were
immunostained with a monoclonal antibody against IL-17B. IL-17B was upregulated in 30% of
breast cancer specimens examined (12/40) while being undetectable in normal tissues (0/18).
Scale bar: 50 µm.
(C) Reduction of IL-17B in MDB-MB468 breast cancer cells with specific shRNA, in relation to
control. The result was determined by western blot.
(D) Reduction of IL-17B in MDA-MB468 cells impaired the proliferation rate, as determined by
MTT assay. *P-value < 0.05. N=12.
(E) Reduction of IL-17B in MDA-MB468 cells impaired the invasion rate, as determined by
Matrigel invasion assay. **P-value < 0.01. N=4.
(F) Reduction of IL-17B in MDA-MB468 cells impaired anchorage-independent growth in soft
agar. Scale bar: 50 µm.
(G) The number of colonies formed on in soft agar as in (F) were quantified and compared
between control and IL-17Bsh-treated cells. ***P-value < 0.001. N=6.
(H) Ectopic addition of recombinant IL-17B protein (50 ng/ml) to the culture medium promoted
the proliferation rate of MDA-MB468 cells, as determined by MTT assay. *P-value < 0.05.
N=12.
(I) Ectopic addition of recombinant IL-17B protein to the culture medium at different
concentrations promoted the clonogenic potential of MDA-MB468 cells. N=6.

20
(J) Ectopic addition of recombinant IL-17B protein to the culture medium at different
concentrations (50 or 100 ng/ml) promoted the invasiveness of MDA-MB468 cells. **P-value <
0.01. N=4.
Supplementary References
1. J. Debnath, S. K. Muthuswamy, J. S. Brugge, Morphogenesis and oncogenesis of MCF-
10A mammary epithelial acini grown in three-dimensional basement membrane cultures.
Methods 30, 256 (2003).
2. S. Furuta, X. Jiang, B. Gu, E. Cheng, P. L. Chen, W. H. Lee, Depletion of BRCA1
impairs differentiation but enhances proliferation of mammary epithelial cells. Proc.
Natl. Acad. Sci. U S A 102, 9176 (2005).
3. X. Wang, C. F. Chen, P. R. Baker, P. L. Chen, P. Kaiser, L. Huang, Mass spectrometric
characterization of the affinity-purified human 26S proteasome complex. Biochemistry
46, 3553 (2007).
4. R. J. Chalkley, P. R. Baker, L. Huang, K. C. Hansen, N. P. Allen, M. Rexach, A. L.
Burlingame, Comprehensive analysis of a multidimensional liquid chromatography mass
spectrometry dataset acquired on a QqTOF mass spectrometer: 2. New developments in
protein prospector allow for reliable and comprehensive automatic analysis of large
datasets. Mol. Cell Proteomics 4, 1194 (2005).
5. S. Furuta, J. M. Wang, S. Wei, Y. M. Jeng, X. Jiang, B. Gu, P. L. Chen, E. Y. H. P. Lee,
W. H. Lee, Removal of BRCA1/CtIP/ZBRK1 repressor complex on ANG1 promoter

21
leads to accelerated breast tumor growth contributed by prominent vasculature. Cancer
Cell 10, 13 (2006).
6. D. C. Allred, J. M. Harvey, M. Berardo, G. M. Clark, Prognostic and predictive factors in
breast cancer by immunohistochemical analysis. Mod. Pathol. 11, 155 (1998).




















