
Tsc1 (hamartin) confers neuroprotection against ischemia by inducing autophagy
Upload
independentCategory
view
0download
0
www.elsevier.com/locate/ynbdi
Neurobiology of Disease 19 (2005) 419–426
Synaptophysin enhances the neuroprotection of VMAT2 in
MPP+-induced toxicity in MN9D cells
Carol X.-Q. Chen, Steven Y. Huang,1 Limei Zhang, and Yong-Jian LiuT
Departments of Neurology and Neurobiology, University of Pittsburgh School of Medicine, W958 Biomedical Science Tower, 200 Lothrop Street, Pittsburgh,
PA 15213, USA
Received 4 August 2004; revised 10 January 2005; accepted 20 January 2005
Available online 5 March 2005
The use of the potent neurotoxin MPTP in producing a model for
Parkinson’s disease (PD) has allowed us to dissect the cellular processes
responsible for both selective neuronal vulnerability and neuroprotec-
tion in idiopathic PD. It has been suggested that vesicular monoamine
transporters (VMATs) play a critical neuroprotective role in MPP+
toxicity. However, little is known about how this detoxificative
sequestration in dopaminergic (DAergic) neurons is regulated at the
molecular and cellular levels. Using the DAergic cell line MN9D as an
in vitro model, we found that overexpression of VMAT2 (a neuronal
isoform of VMATs) protects the transformants from MPP+-induced
toxicity, consistent with the previous work on fibroblastic CHO cells.
We further found that the MN9D cells displayed lower expression levels
of secretory vesicle proteins such as synaptophysin. Overexpression of
synaptophysin in MN9D cells can significantly increase the resistance
of the transformants to MPP+ toxicity. The co-expression of VMAT2
and synaptophysin has shown synergistic protection for the trans-
formants, suggesting a role of synaptophysin in the biogenesis of
secretory vesicles and in influencing the targeting of VMAT2 to these
vesicles. Our work indicates that both the expression level of VMAT2
and capacity of vesicular packaging of DA are important in protecting
DAergic cells from MPP+ toxicity.
D 2005 Elsevier Inc. All rights reserved.
Keywords: Parkinson’s disease; Neuroprotection; MPP+ toxicity; MN9D;
VMAT2; Synaptophysin
0969-9961/$ - see front matter D 2005 Elsevier Inc. All rights reserved.
doi:10.1016/j.nbd.2005.01.014
Abbreviations: DAT, dopamine transporter; DA, dopamine; DAergic,
dopaminergic; EGFP, enhanced green fluorescent protein; AMPT, a-
methyl-p-tyrosine; PNS, postnuclear supernatant; Syn, synaptophysin;
TH, tyrosine hydroxylase; TBZ, tetrabenazine; VMAT2, vesicular mono-
amine transporter 2.
* Corresponding author. Fax: +1 412 624 9914.
E-mail address: [email protected] (Y.-J. Liu).1 Present address: AEMTEK, Inc., 46309 Warm Springs Blvd., Suite A,
Fremont, CA, USA.
Available online on ScienceDirect (www.sciencedirect.com).
Introduction
In several animal models, the neurotoxin MPTP induces a
selective loss of dopamine (DA) neurons in the substantia nigra of
the midbrain, and this loss resembles a clinical symptom
occurring in idiopathic Parkinson’s disease (PD) (Kopin and
Markey, 1988; Langston, 1987; Przedborski et al., 2001). The in
vitro neurotoxicity study on cell lines or primary cultures using
MPP+, an active form of MPTP, has allowed dissection of the
cellular processes responsible for both selective neuronal vulner-
ability and neuroprotection in idiopathic PD. The vesicular
sequestration of MPP+ mediated by overexpressed vesicular
monoamine transporters (VMATs) has provided a mechanism
for protecting fibroblastic CHO transformants from MPP+-induced
toxicity (Liu et al., 1992a) (for a review, see Ahnert-Hilger et al.,
2003; Gainetdinov and Caron, 2003; Liu and Edwards, 1997b;
Uhl et al., 2000). However, little has been done to directly
confirm whether similar neuroprotection of VMATs occurs in
dopaminergic (DAergic) cell lines or DAergic neurons. More
importantly, little is known about how this protection is regulated
at the molecular and cellular levels. As a neuronal isoform of the
vesicular transporters, VMAT2 normally transports monoamines
from both de novo synthesis and reuptake from the synapse into
secretory vesicles for regulated release (Kanner and Schuldiner,
1987). VMAT2 in DAergic neurons thus appears to play a dual
role: neuroprotection by detoxification and signaling in synaptic
transmission. In support of their potential role in detoxification,
the transporters share sequence similarity with a group of bacteria
multidrug resistant genes (Liu et al., 1992b; Schuldiner et al.,
1995). Nonetheless, the loss of DAergic neurons in the MPTP
model in the presence of VMAT2 has raised a reasonable doubt
concerning whether VMAT2 is sufficient to protect DAergic
neurons in vivo.
Over the past decade, there has been limited clinical evidence in
support of the neuroprotective function of VMAT2 although the
transporter has in fact served as a marker for assessment of PD
progression (Brooks et al., 2003). However, several recent studies
on VMAT2 knockout mice strongly suggest that impaired DA
packaging may induce DAergic neuron susceptibility to neurotoxins

C.X.-Q. Chen et al. / Neurobiology of Disease 19 (2005) 419–426420
(Fon et al., 1997; Gainetdinov et al., 1998; Takahashi et al., 1997;
Wang et al., 1997). These independent studies have consistently
shown that heterozygote VMAT2 knockout mice display increased
MPTP neurotoxicity and increased methamphetamine neurotoxic-
ity, suggesting that VMAT2-mediated protection correlates with the
expression level of the transporter. The VMAT2 knockdown model
has also shown consistent results with increased vulnerability to
MPTP toxicity (Mooslehner et al., 2001). More recently, Staal and
his colleagues suggested that higher expression of VMAT2 and
greater ability to sequester MPP+ in rat than in mouse may
contribute to the reduced vulnerability of rat DAergic neurons to
MPP+ neurotoxicity (Staal et al., 2000). The recent report on the
correlation between lower VMAT2 expression and higher neuro-
melanin pigment accumulation in human ventral substantia nigra
neurons also indicates a protective role for the transporter (Liang et
al., 2004). Interestingly, the significant resistance of the mutant
mouse Tottering to MPTP partially results from hyperinnervation of
noradrenergic terminals in the brain, which presumably offers
higher vesicular storage capacity in MPP+ sequestration (Kilbourn
et al., 1998). Furthermore, the neuroprotection of the transporter has
also been observed in CHO cells in dopamine toxicity (Weingarten
and Zhou, 2001). However, lack of an in vitro model for VMAT2
neuroprotection prevents further study of molecular mechanisms
involved in the protection and its regulation.
Secretory vesicles provide a reducing environment for the
function of VMAT2 during the storage of monoamines (Liu and
Edwards, 1997b). Thus, the availability and the function of
secretory vesicles might determine the capacity by which VMAT2
mediates sequestration of MPP+ and other neurotoxins. It has been
suggested that the impaired function of secretory vesicles at nerve
terminals might be associated with the pathogenesis caused by
overexpression of mutant proteins such as a-synuclein and parkin
in PD (Lotharius and Brundin, 2002; Zhang et al., 2000). However,
little is known about how vesicular proteins such as VMAT2 and
synaptophysin target to these specified vesicles and permit them to
perform normal function in synaptic transmission. On the other
hand, the size and contents of secretory vesicles may also regulate
the function. One such example is that the secretory granules (120
nm in diameter, counterparts of large dense core vesicles in brain)
in PC12 cells can concentrate monoamines more than 10,000 times
within the granules (~5 mM in lumen of vesicles) over the
cytoplasmic levels (Winkler, 1976). In contrast, synaptic vesicles
(40 nm in diameter) display less than 1000 times concentration.
This might contribute to the higher capacity for MPP+ detoxifica-
tion in PC12 cells than in other neuronal cell lines.
In this study, we used a DAergic cell line, MN9D, to establish
an in vitro model for MPP+ toxicity study. We aimed to determine
whether VMAT2 expression can indeed protect DAergic cells, as
suggested by the study with fibroblast CHO cells. Furthermore, our
study explored some unknown features of MN9D cells such as the
transport activity for dopamine transporter (DAT) and VMAT2,
and secretory vesicle properties.
Materials and methods
Chemicals
General chemicals used in this work were purchased from
Sigma (St. Louis, MO) unless otherwise noted. The following
antibodies were used in the experiments: polyclonal anti-synapto-
physin (1:6000 for blotting and 1:200 for immunofluorescent
staining, Zymed, South San Francisco, CA); monoclonal anti-
synaptophysin (1:4000 for blotting and 1:200 for immunofluor-
escent staining, Sigma); polyclonal anti-secretogranin (1:6000 for
blotting, Biodesign, Saco, ME); monoclonal anti-SV2 (1:3000 for
blotting, Hybridoma Bank, Iowa City, IA); monoclonal anti-
synapsin I (1:3000 for blotting, Chemicon, Temecula, CA), and
monoclonal anti-h-actin (1:5000 for blotting, Sigma).
Cell culture and transfection
All cells were maintained in 5% CO2 at 378C in medium
containing penicillin and streptomycin. PC12 cells were grown in
high-glucose Dulbecco’s modified Eagle’s-H21 medium (Invitro-
gen, San Diego, CA) supplemented with 5% Cosmic calf serum
and 10% equine serum (Hyclone, Logan, UT) as described
previously (Liu et al., 1994). MN9D and SY5Y cells were grown
in high-glucose DMEM supplemented with 10% fetal bovine
serum. CHO cells were grown in F12 supplemented with 10%
Cosmic calf serum. For making stable MN9D transformants, the
cells were transfected with VMAT2-EGFP-N2, synaptophysin/
pcDNA3.1/hygro, or pcDNA3.1/hygro vector alone using a
modified calcium phosphate transfection method. The transfected
cells were then treated for selection by either G418 (500 Ag/ml) or
hygromycin B (300 Ag/ml, Invitrogen, San Diego, CA). Positive
clones for EGFP-VMAT2 were selected by FACS (MoFlo high-
performance cell sorter, Cytomation, Inc., Fort Collins, CO). Other
positive transformants were screened by both Western blot analysis
and immunofluorescence staining. Several synaptophysin trans-
formants were further transfected with HA-VMAT2 and selected
by G418 to obtain double transformants.
cDNA plasmid constructs
An HA-tagged VMAT2 cDNA (Tan et al., 1998) was recovered
from a pCDNA3 vector by HindIII/XbaI digestion and was
subcloned into pEGFP-N2 vector (BD Clontech, CA). Synaptophy-
sin was amplified from a PC12 cell library using PCR with a pair of
primers (5V-ACTAAGCTTCATTGCTGCTGCTGCTGCTGCT-3Vand 5V-ATCTCGAGAGGCCTTCTC TTGAGCTGTT-3V) contain-ing restriction sites for HindIII and XhoI, respectively. Amplified
PCR fragment was digested and subcloned into pcDNA3.1/hygro
expression vector (Invitrogen). cDNA plasmids were prepared using
Qiagen Endo-free Midiprep kits (USA-Qiagen, Valencia, CA).
Cytotoxicity assay
About 3 � 104 cells to be tested were plated in each well of 24-
well plates for 24 h before adding the neurotoxin MPP+ at
concentrations between 10 and 1000 AM. After 3 days of
treatment, the cells were harvested with trypsin–EDTA (0.05%)
and stained with trypan blue. Live cells were then counted using a
hemacytometer. The percent viability of cells was calculated based
on the number of live cells divided by cells from the non-treated
control.
Western blotting analysis
Cells were harvested in buffer A (150 mM NaCl, 10 mM Hepes
(pH 7.4), 1 mM EGTA, and 0.1 mM MgCl2) with protease
inhibitors (0.4 nM PMSF, 2 ng/ml leupeptin, and 1 Ag/ml pepstatin).

Fig. 1. MN9D cell line is more sensitive to MPP+ toxicity than other
monoaminergic cell lines. MPP+ toxicity assay was performed on MN9D,
PC12, SY5Y, and CHO cells. Cells were subsequently treated with different
concentrations of MPP+ (0–1 mM) for 3 days. Cells were harvested with
0.025% trypsin–EDTA and stained with trypan blue. The cell viability (%)
is expressed as percentage of survival cells compared with that in non-
MPP+-treated control. Data are mean F SD (bars) values from four
experiments, each one of which was performed with duplicate incubations.
C.X.-Q. Chen et al. / Neurobiology of Disease 19 (2005) 419–426 421
The cells were then homogenized by sonication and centrifuged at
3000 rpm for 5 min at 48C to obtain postnuclear supernatant (PNS).
The PNS was further sedimented at 100,000 � g for 30 min at 48Cto obtain membrane protein fraction. The membrane pellet was
further resuspended in 3� SDS sample buffer (New England
Biolab, Beverly, MA) and incubated at room temperature for 5 min.
A 10–100 Ag protein sample was separated by electrophoresis
through 10% SDS–PAGE. After electrophoresis, the proteins were
transferred to nitrocellulose (BA-85, Schleicher-Schuell Bio-
science, Keene, NH), and HA-VMAT2 and synaptophysin were
visualized by enhanced chemiluminescence (Supersignal West
Pico, Pierce, Rockford, IL) using monoclonal anti-HA.11 (Babco,
Berkeley, CA) at a 1:3000 dilution and anti-synaptophysin at
1:6000 dilution, respectively. The secondary HRP-conjugated anti-
mouse antibody was used at a dilution of 1:5000 (Amersham,
Arlington Heights, IL). Protein concentration was determined by
protein assay kit (Bio-Rad, Hercules, CA). To quantify the intensity
of the detected bands, a densitometer was used (Personal
Densitometer S1, Molecular Dynamics, Amersham Biosciences,
Piscataway, NJ).
Subcellular fractionation
For cell fractionation by equilibrium sedimentation, the PNS
was layered onto a linear 0.65 to 1.55M sucrose gradient for
sedimentation in an SW41 rotor (Beckman Instruments, Palo Alto,
CA) at 30,000 rpm for 6 h at 48C. Fractions were collected from
the bottom of the tube and analyzed by Western blot.
Uptake assays
For the DAT uptake assay, intact cells cultured in 24-well
plates were used. 1 � 104 cells per well were plated on poly-d-
lysine pre-coated 24-well plates 1 day before the assay. Cells
were pretreated with or without 1 AM mazindol or 2 AMGBR12909 for 20 min, followed by a brief rinse with 0.5 ml of
ice-cold PBS solution supplemented with 1 mM MgCl2 and 0.1
mM CaCl2. Cells were then incubated in 300 Al of 10 nM [3H]-
MPP+ with or without mazindol at room temperature at intervals
of 0, 5, 10, 30, 60, and 90 min. The uptake was terminated by
rapid rinse with ice-cold PBS w/Ca2+, Mg2+ three times. Cell
extracts were prepared by solubilizing cells by 1% SDS, and the
radioactivity was determined by liquid scintillation counting. A
VMAT2 uptake assay was performed as described previously (Liu
and Edwards, 1997a). Briefly, PNS of cells were prepared as
described above. Since VMAT2 showed similar affinity to
monoamine substrates (Peter et al., 1994), for measuring the
transport activity of VMAT2 we chose [1,2-3H]-serotonin (NEN,
Boston, MA) over other monoamines because of its availability
and stability (substrates such as dopamine have a higher tendency
toward auto-oxidation; Stokes et al., 1999). In this transport
activity assay, 10 Al (1 Ag protein/Al) of PNS were incubated with
20 nM of [1,2-3H]-serotonin at 308C for the indicated times. The
uptake was terminated by filtration separation through a 0.45-AmMillipore filter (Millipore, Billerica, MA). The dried filters were
then used for scintillation counting.
Tetrabenazine binding assay
The binding assay was performed essentially as previously
described (Scherman and Henry, 1981). Membrane proteins
(50–100 Ag) from PNS preparation were incubated in 300 Al ofbinding buffer (0.3 M sucrose, 10 mM HEPES, pH 7.2) with 3
nM of [2-3H] a-dihydrotetrabenazine (3H-TBZ, ART-496, Amer-
ican Radiolabeled Chemicals, Inc., St. Louis, MO) at room
temperature. After 15–30 min incubation, the reaction was
stopped by the addition of 4 ml of ice-cold binding buffer
containing 10 AM TBZ and filtered through 0.45 Am HAWP
Millipore filters pretreated with 0.3% polyethylenimine. The
filters were then washed once more with 4 ml of the same buffer,
and the radioactivity was measured by liquid scintillation counter
(Beckman 9000, Beckman Coulter, Inc., Fullerton, CA). Non-
specific binding was determined by the addition of 2 AM cold
TBZ to the binding buffer.
Results
The MN9D cell line is more sensitive to MPP+ toxicity than are
other monoaminergic cell lines
To identify a cell line that may resemble the DAergic neurons
sensitive to MPP+, four cell lines, CHO, MN9D, PC12, and SH-
SY5Y, were chosen for the MPP+-induced toxicity assay. PC12
cells were more resistant to MPP+ than other cell lines with
EC50 of MPP+ about 500 AM (Fig. 1). SH-SY5Y cells also
showed responses to MPP+ similar to that of PC12 cells. As a
dopaminergic cell line, MN9D cells were more susceptible to
MPP+ than other cell lines with EC50 of ~60 AM. The
fibroblastic CHO cells showed a level of sensitivity between
those of the MN9D and the other two cell lines (Fig. 1). To
examine if accumulation of MPP+ was mediated by dopamine
transporter (DAT) and responsible for the selectivity, a [3H]-
MPP+ uptake assay was performed. All cell lines except the CHO
cell line showed similar time-dependent uptake of MPP+ in intact
cells (Fig. 2a). The uptake was inhibited by the DAT-specific
antagonists Mazindol (Fig. 2a) and GBR12909 (data not shown).
The results indicate that selective DAT-mediated uptake of MPP+
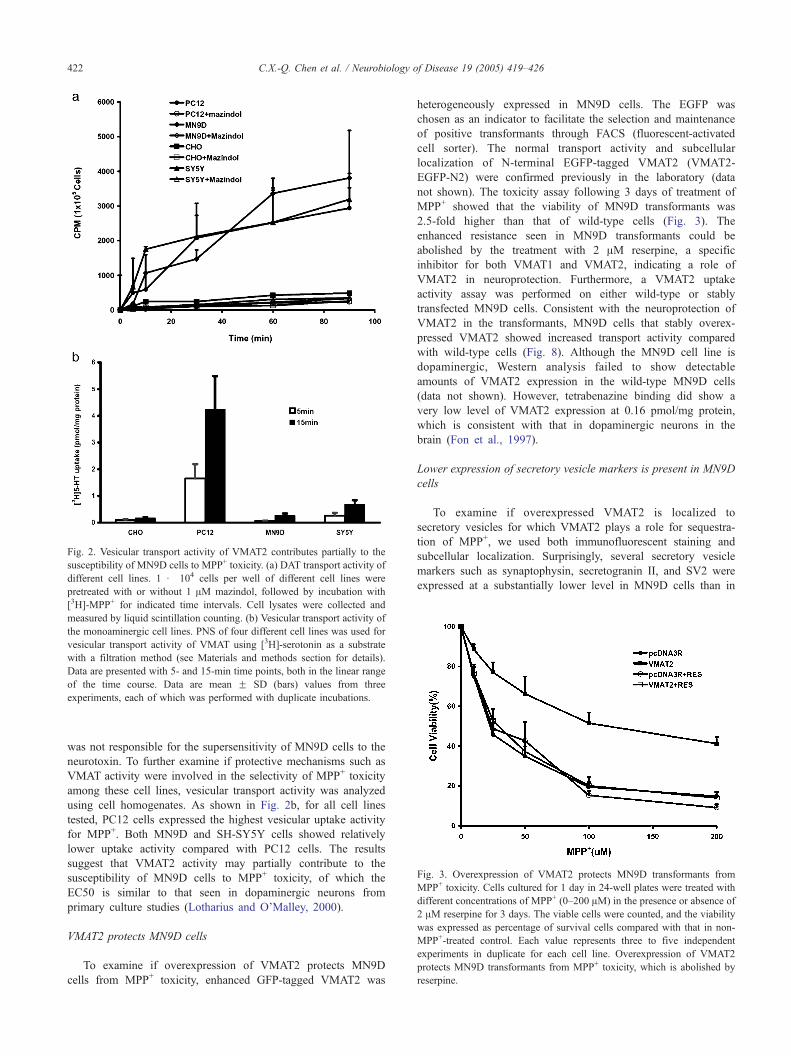
Fig. 3. Overexpression of VMAT2 protects MN9D transformants from
MPP+ toxicity. Cells cultured for 1 day in 24-well plates were treated with
different concentrations of MPP+ (0–200 AM) in the presence or absence of
2 AM reserpine for 3 days. The viable cells were counted, and the viability
was expressed as percentage of survival cells compared with that in non-
MPP+-treated control. Each value represents three to five independent
experiments in duplicate for each cell line. Overexpression of VMAT2
protects MN9D transformants from MPP+ toxicity, which is abolished by
reserpine.
Fig. 2. Vesicular transport activity of VMAT2 contributes partially to the
susceptibility of MN9D cells to MPP+ toxicity. (a) DAT transport activity of
different cell lines. 1 � 104 cells per well of different cell lines were
pretreated with or without 1 AM mazindol, followed by incubation with
[3H]-MPP+ for indicated time intervals. Cell lysates were collected and
measured by liquid scintillation counting. (b) Vesicular transport activity of
the monoaminergic cell lines. PNS of four different cell lines was used for
vesicular transport activity of VMAT using [3H]-serotonin as a substrate
with a filtration method (see Materials and methods section for details).
Data are presented with 5- and 15-min time points, both in the linear range
of the time course. Data are mean F SD (bars) values from three
experiments, each of which was performed with duplicate incubations.
C.X.-Q. Chen et al. / Neurobiology of Disease 19 (2005) 419–426422
was not responsible for the supersensitivity of MN9D cells to the
neurotoxin. To further examine if protective mechanisms such as
VMAT activity were involved in the selectivity of MPP+ toxicity
among these cell lines, vesicular transport activity was analyzed
using cell homogenates. As shown in Fig. 2b, for all cell lines
tested, PC12 cells expressed the highest vesicular uptake activity
for MPP+. Both MN9D and SH-SY5Y cells showed relatively
lower uptake activity compared with PC12 cells. The results
suggest that VMAT2 activity may partially contribute to the
susceptibility of MN9D cells to MPP+ toxicity, of which the
EC50 is similar to that seen in dopaminergic neurons from
primary culture studies (Lotharius and O’Malley, 2000).
VMAT2 protects MN9D cells
To examine if overexpression of VMAT2 protects MN9D
cells from MPP+ toxicity, enhanced GFP-tagged VMAT2 was
heterogeneously expressed in MN9D cells. The EGFP was
chosen as an indicator to facilitate the selection and maintenance
of positive transformants through FACS (fluorescent-activated
cell sorter). The normal transport activity and subcellular
localization of N-terminal EGFP-tagged VMAT2 (VMAT2-
EGFP-N2) were confirmed previously in the laboratory (data
not shown). The toxicity assay following 3 days of treatment of
MPP+ showed that the viability of MN9D transformants was
2.5-fold higher than that of wild-type cells (Fig. 3). The
enhanced resistance seen in MN9D transformants could be
abolished by the treatment with 2 AM reserpine, a specific
inhibitor for both VMAT1 and VMAT2, indicating a role of
VMAT2 in neuroprotection. Furthermore, a VMAT2 uptake
activity assay was performed on either wild-type or stably
transfected MN9D cells. Consistent with the neuroprotection of
VMAT2 in the transformants, MN9D cells that stably overex-
pressed VMAT2 showed increased transport activity compared
with wild-type cells (Fig. 8). Although the MN9D cell line is
dopaminergic, Western analysis failed to show detectable
amounts of VMAT2 expression in the wild-type MN9D cells
(data not shown). However, tetrabenazine binding did show a
very low level of VMAT2 expression at 0.16 pmol/mg protein,
which is consistent with that in dopaminergic neurons in the
brain (Fon et al., 1997).
Lower expression of secretory vesicle markers is present in MN9D
cells
To examine if overexpressed VMAT2 is localized to
secretory vesicles for which VMAT2 plays a role for sequestra-
tion of MPP+, we used both immunofluorescent staining and
subcellular localization. Surprisingly, several secretory vesicle
markers such as synaptophysin, secretogranin II, and SV2 were
expressed at a substantially lower level in MN9D cells than in

Fig. 5. Overexpression of synaptophysin attenuates MPP+ toxicity in MN9D
cells. (a) Synaptophysin-stable transformants were used to perform the
toxicity assay. pcDNA3.1/hygro vector-transfected MN9D transformants
were used as control. (b) Synergistic neuroprotective effect by double
overexpression of VMAT2 and synaptophysin in MN9D cells. The experi-
ments were performed essentially the same way as in Fig. 1.
Fig. 6. VMAT2 colocalizes with synaptophysin on synaptic vesicles in
MN9D cells. Density sucrose gradient fractionation was performed by sedi-
mentation of PNS of MN9D transformants through an equilibrium gradient
composed of 0.6–1.6 M sucrose. Fractions were collected and analyzed by
Western blot. VMAT2 was co-migrated with synaptophysin enriched in fracQ
tions 8–10 for secretory vesicles. Fractions were labeled from top to bottom.
Fig. 4. Low expression of secretory vesicle marker proteins in MN9D cells.
Membrane proteins from PNS (10 Ag for PC12 cells and 100 Ag for MN9D
cells) were loaded for PAGE and Western analyzed for secretory vesicle
marker proteins such as synaptophysin, secretogranin II, and SV2. To detect
synaptophysin in MN9D cell PNS, 100 Ag protein was used. Protein
samples were further normalized by h-actin.
C.X.-Q. Chen et al. / Neurobiology of Disease 19 (2005) 419–426 423
PC12 cells (Fig. 4). More specifically, the expression level of
synaptophysin was about 30- to 50-fold less than that in PC12
cells (Fig. 4, quantitative data not shown), suggesting that the
relatively small number of secretory vesicles in MN9D cells
might be responsible for their supersensitivity to the neurotoxin
MPP+.
Overexpressed synaptophysin attenuates MPP+ toxicity
Previous evidence has suggested that synaptophysin may play
a role in the biogenesis of secretory vesicles (Thiele et al., 2000).
We hypothesized that a low expression of synaptophysin
contributes to the reduced biogenesis of secretory vesicles in
MN9D cells. Therefore, we tested whether heterogeneous over-
expression of synaptophysin would increase the generation of
secretory vesicles and then the capacity of detoxification by
sequestration of MPP+ in MN9D cells. Determined by toxicity
assay, the synaptophysin-overexpressed cells showed increased
viability, with EC50 of ~92 AM in MPP+ treatment (Fig. 5a). This
result is comparable to that of VMAT2 transformants in the
MPP+ toxicity study (Fig. 3, EC50 = ~115 AM). The transport
activity assay also suggested an increased endogenous expression
of VMAT2 in secretory vesicles (refer to Discussion). Further-
more, the tetrabenazine binding assay on the synaptophysin
transformants has shown a consistent increase in endogenous
VMAT2 expression, from 0.27 pmol/mg protein to 0.41 pmol/mg
protein. Similar results on decreased sensitivity to MPP+ were
obtained from transient synaptophysin-transfected MN9D cells
(data not shown).
Co-expression of VMAT2 and synaptophysin provides synergistic
protection of MN9D transformants
To examine if overexpression of synaptophysin can facilitate
the neuroprotection of VMAT2 in MPP+ toxicity, we co-
expressed two proteins in MN9D cells. The transformants that
overexpress both proteins showed a ~2-fold increase in EC50 of
MPP+ compared with the overexpression of either protein alone
in the MPP+ toxicity assay (Fig. 5b). Both proteins also co-
localized to secretory vesicles as shown by double immuno-
fluorescent staining (data not shown) and subcellular fractionation
(Fig. 6). The results suggest a role for vesicular sequestration of

Fig. 9. Inhibition of dopamine synthesis attenuates MPP+ toxicity. MN9D
transformants transfected by either pcDNA3/RSV or VMAT2-EGFP-N2
were pretreated with or without 100 AM MPT for 24 h followed by the
addition of 50 or 200 AM MPP+. The viability of cells was measured using
essentially the same procedure as that in Fig. 1. MPT protects MN9D/
pcDNA3 from MPP+ toxicity but does not enhance VMAT2 protection.
Data are presented as meanF SD (bars) values from four experiments, each
of which was performed with quadruplicate incubations.
Fig. 7. Increased expression of secretory vesicle proteins in synaptophysin
and VMAT2 double-transfected MN9D cells. Lane 1, PC12/WT; Lane 2,
MN9D/WT; Lane 3, MN9D/Syn+VMAT2; Lane 4, MN9D/Syn. Membrane
proteins from PNS (10 Ag for PC12 cells and 100 Ag for MN9D cells) were
loaded for PAGE and Western blotting. The protein concentration was
further normalized by h-actin.
C.X.-Q. Chen et al. / Neurobiology of Disease 19 (2005) 419–426424
the neurotoxin MPP+ by secretory vesicles in which VMAT2 is
localized. Furthermore, several secretory vesicle markers also
showed increased expression levels. The increase was about
twofold for secretogranin II and fourfold for SV2 (Fig. 7),
indicating an increased biogenesis of secretory vesicles in the
transformants. In addition, the transport activity also suggests an
increased uptake of [3H]-serotonin, an indication of VMAT2
function (Fig. 8).
Inhibition of dopamine synthesis attenuates MPP+ toxicity
To test whether dopamine toxicity is involved inMPP+-mediated
toxicity and VMAT2 is sufficient for protection, we used a-methyl-
p-tyrosine (AMPT) as a TH-specific blocker to inhibit the synthesis
of dopamine in MN9D cells. As shown in Fig. 9, the inhibition of
dopamine synthesis can significantly reduce MPP+ toxicity by 30%.
Fig. 8. Neuroprotection of MN9D transformants correlates with the
transport activity of VMAT2. PNS of MN9D transformants was used for
transport activity assay with a filtration method (see Materials and methods
section for detail). Wild-type MN9D cells were used as control. Data are
presented as mean F SD (bars) values from at least three experiments, each
of which was performed with duplicate incubations. *P b 0.05, vs. control
(MN9D/WT), **P b 0.01 vs. control (MN9D/WT), #P b 0.05 vs. MN9D/
VMAT2.
This neuroprotection does not synergize with that by VMAT2
overexpression.
Discussion
We have shown that VMAT2 protects a DAergic cell line,
MN9D, from MPP+-induced toxicity. Our results also indicate that
the sensitivity of MN9D cells to MPP+ toxicity is relatively higher
than that of other dopamine-producing cell lines. This feature of
MN9D cells is consistent with that observed in primary dopamine
neurons. We have reported DAT-like activity in MN9D cells, which
is believed to be important for MPP+ toxicity. We also found that
the MN9D cells displayed lower expression levels of secretory
vesicle markers, suggesting a low amount of secretory vesicles in
this cell line. In our studies, the overexpression of either VMAT2
or synaptophysin in MN9D cells has shown significant increases in
resistance to MPP+ toxicity. Furthermore, the co-expression of two
proteins that may promote the vesicular targeting of VMAT2 and
the biogenesis of secretory vesicles has also shown synergistic
protection for the transformants.
Dopaminergic cell lines and MPP+ toxicity
Our results suggest that dopaminergic cell lines vary dramati-
cally in their response to MPP+ toxicity. This variation may be
determined by differences in DAT expression, expression level of
VMATs, and vesicle type (secretory granules versus synaptic
vesicles) and amount. Previously, we used the CHO cell line for
molecular cloning of VMAT2 (Liu et al., 1992b). Our present data
are consistent with previous unpublished work in which no
catecholamine transport activity was detected in wild-type CHO
cells. The MPP+-induced cell death might result from low
cytoplasmic accumulation of MPP+ upon exposure to high
concentrations of MPP+ (EC50 of 150 AM). In contrast, higher
cytoplasmic MPP+ (shown in Fig. 2) mediated by the amine
transporter system may occur in DAergic cell lines when they are

C.X.-Q. Chen et al. / Neurobiology of Disease 19 (2005) 419–426 425
exposed to much lower concentrations of MPP+ (e.g., EC50 of 60
AM for MN9D cells). However, DAT expression is not the only
deciding factor given that SH-SY5Y cells express high levels of
DAT but are resistant to MPP+ toxicity. Our results confirm the
notion that the capacity of detoxification can also be determined by
the expression of VMATs and the amount of secretory vesicles
where VMATs are localized. Early studies on PC12 cells have
demonstrated a relative resistance to the toxicity of MPP+ (Denton
and Howard, 1987; Greene and Rein, 1978). The vesicular
accumulation of [3H]-MPP+ in chromaffin cell homogenates
indicated a specific mechanism in this neuroprotection, which
further led to identification of the VMAT gene family (Liu et al.,
1992a; Reinhard et al., 1987). Additional in vitro pharmacological
studies have clearly supported a role of VMATs in the vesicular
sequestration of neurotoxin (Liu et al., 1992b; Peter et al., 1994).
Finally, differences in type and amount of secretory vesicles may
play a role. The lower expression level of VMAT2 in SH-SY5Y
determined by our uptake assay may indicate that VMAT2
expression does not explain resistance in this cell line. One possible
explanation is that this resistance may be attributed to the higher
detoxification capacity mediated by secretory granules which are
abundant in SY5Y cells (unpublished data). This secretory vesicle
profile represents a neuroendocrine phenotype similar to that of
PC12 cells, which are also relatively resistant. Thus, all these factors
must be considered in modeling MPP+ toxicity.
MN9D cells have demonstrated their potential as an in vitro
system for studying MPP+ toxicity because of their dopamine
neuron origin (Choi et al., 1991, 1992). Their sensitivity to MPP+
toxicity from our study is within the range of that seen in primary
dopamine neuronal cultures (Lotharius et al., 1999). Our character-
ization of this cell line has also indicated that both DAT-like
activity and low levels of VMAT2-like activity were detected. Our
RT-PCR analysis also indicates the expression of VMAT2 but not
VMAT1 in this cell line (data not shown). On the other hand, our
pharmacological analysis using both mazindol and GBR12909
inhibitors may not exclusively rule out whether other amine
transporters such as serotonin transporter are expressed. As MPP+
is a substrate for all monoamine transporters with similar affinity,
MPP+ toxicity may be also mediated by transporters other than
DAT. Analysis of the neurotransmitter transporter profile for
MN9D cells might provide some important information about
molecular mechanisms underlying MPP+ toxicity. However, it
seems inevitable that for any of the cell lines used in PD studies,
certain limitations may exist. In the case of MN9D cells, low
expression of secretory vesicle marker proteins may suggest a
relatively low amount of secretory vesicles as well as lower
capacity for detoxification. Overexpression of synaptophysin
increases the resistance of the transformants, which may result
from increased vesicular capacity for detoxification, an important
aspect in understanding the cellular mechanism underlying MPP+
toxicity.
VMAT2 detoxification and dopamine toxicity
As indicated by previous studies, vesicular sequestration of the
neurotoxin MPP+ may play a critical role in the neuroprotection of
fibroblast CHO transformants by VMAT2 (Liu et al., 1992a).
However, normal function of VMAT2 in the brain is mainly
involved in the packaging of monoamines, including dopamine,
into secretory vesicles (e.g., synaptic vesicles and large dense-core
vesicles). This raises the possibility that VMAT2 may be essential
in DAergic neurons to prevent cytoplasmic DA oxidation by
sequestering DA into vesicles. In contrast, under pathological
situations such as MPP+ insult, endogenous expression of the
transporter might not be sufficient to protect the cells. Interestingly,
a recent report indicated that DA may mediate MPP+ toxicity
through its intracellular oxidation and through production of non-
mitochondrial reactive oxygen species (ROS) (Lotharius and
O’Malley, 2000). Our result also supports this observation in that
the inhibition of DA synthesis attenuated MPP+ toxicity. More
importantly, depletion of DA did not enhance VMAT2 protection,
suggesting that vesicular sequestration of neurotoxins such as
MPP+ or DA may be sufficient for executing VMAT’s neuro-
protection. It would be interesting to examine whether vesicular
DA efflux occurs during MPP+-induced toxicity and how this
process is related to the pathogenesis of PD.
Synaptophysin and biogenesis of secretory vesicles
Although the function of synaptophysin remains unknown,
evidence has suggested that this abundant synaptic vesicle protein
may be involved in the biogenesis of secretory vesicles (Daly et al.,
2000; Thiele et al., 2000). Furthermore, it has also been speculated
that synaptophysin may play a role in stabilizing and modifying the
function of other synaptic proteins (Becher et al., 1999; Daly et al.,
2000; Edelmann et al., 1995; Galli et al., 1996). Our studies have
shown that VMAT2 may require dimerization in order to function
(unpublished data). It would be interesting to test whether
synaptophysin increases VMAT2 function through its interaction
with the transporter or other mechanisms. On the other hand, the
overexpression of synaptophysin may promote the formation of
secretory vesicles as suggested by the increased expression level of
other membrane proteins such as secretogranin II and SV2.
Therefore, the increased amount of secretory vesicle may enhance
the function of VMAT2 by providing membrane surface for protein
targeting and vesicular capacity for sequestering the neurotoxin
molecules.
Acknowledgments
We gratefully acknowledge Dr. Alfred Heller for providing
MN9D cells. We thank Ms. Carol Culver for reviewing the
manuscript. This work was supported by grants from the National
Institutes of Health, the Scaife Charitable Foundations and CMRF
of the University of Pittsburgh Medical Center (to Y. L.).
References
Ahnert-Hilger, G., Holtje, M., Pahner, I., Winter, S., Brunk, I., 2003.
Regulation of vesicular neurotransmitter transporters. Rev. Physiol.,
Biochem. Pharmacol. 150, 140–160.
Becher, A., Drenckhahn, A., Pahner, I., Margittai, M., Jahn, R., Ahnert-
Hilger, G., 1999. The synaptophysin–synaptobrevin complex: a hall-
mark of synaptic vesicle maturation. J. Neurosci. 19, 1922–1931.
Brooks, D.J., Frey, K.A., Marek, K.L., Oakes, D., Paty, D., Prentice, R.,
Shults, C.W., Stoessl, A.J., 2003. Assessment of neuroimaging
techniques as biomarkers of the progression of Parkinson’s disease.
Exp. Neurol. 184 (Suppl. 1), S68–S79.
Choi, H.K., Won, L.A., Kontur, P.J., Hammond, D.N., Fox, A.P., Wainer,
B.H., Hoffmann, P.C., Heller, A., 1991. Immortalization of embryonic

C.X.-Q. Chen et al. / Neurobiology of Disease 19 (2005) 419–426426
mesencephalic dopaminergic neurons by somatic cell fusion. Brain Res.
552, 67–76.
Choi, H.K., Won, L., Roback, J.D., Wainer, B.H., Heller, A., 1992. Specific
modulation of dopamine expression in neuronal hybrid cells by primary
cells from different brain regions. Proc. Natl. Acad. Sci. U. S. A. 89,
8943–8947.
Daly, C., Sugimori, M., Moreira, J.E., Ziff, E.B., Llinas, R., 2000.
Synaptophysin regulates clathrin-independent endocytosis of synaptic
vesicles. Proc. Natl. Acad. Sci. U. S. A. 97, 6120–6125.
Denton, T., Howard, B.D., 1987. A dopaminergic cell line variant
resistant to the neurotoxin 1-methyl-4-phenyl-1,2,3,6-tetrahydropyri-
dine. J. Neurochem. 49, 622–630.
Edelmann, L., Hanson, P.I., Chapman, E.R., Jahn, R., 1995. Synaptobrevin
binding to synaptophysin: a potential mechanism for controlling the
exocytotic fusion machine. EMBO J. 14, 224–231.
Fon, E.A., Pothos, E.N., Sun, B.C., Killeen, N., Sulzer, D., Edwards, R.H.,
1997. Vesicular transport regulates monoamine storage and release but
is not essential for amphetamine action. Neuron 19, 1271–1283.
Gainetdinov, R.R., Caron, M.G., 2003. Monoamine transporters: from
genes to behavior. Annu. Rev. Pharmacol. Toxicol. 43, 261–284.
Gainetdinov, R.R., Fumagalli, F., Wang, Y.M., Jones, S.R., Levey, A.I.,
Miller, G.W., Caron, M.G., 1998. Increased MPTP neurotoxicity
in vesicular monoamine transporter 2 heterozygote knockout mice.
J. Neurochem. 70, 1973–1978.
Galli, T., McPherson, P.S., De Camilli, P., 1996. The V0 sector of the
V-ATPase, synaptobrevin, and synaptophysin are associated on
synaptic vesicles in a Triton X-100-resistant, freeze-thawing sensi-
tive, complex. J. Biol. Chem. 271, 2193–2198.
Greene, L.A., Rein, G., 1978. Release, storage and uptake of catechol-
amines by a clonal cell line of nerve growth factor (NGF) responsive
pheochromocytoma cells. Brain Res. 129, 247–263.
Kanner, B.I., Schuldiner, S., 1987. Mechanisms of storage and transport of
neurotransmitters. CRC Crit. Rev. Biochem. 22, 1–38.
Kilbourn, M.R., Sherman, P., Abbott, L.C., 1998. Reduced MPTP
neurotoxicity in striatum of the mutant mouse tottering. Synapse 30,
205–210.
Kopin, I.J., Markey, S.P., 1988. MPTP toxicity: implications for research in
Parkinson’s disease. Annu. Rev. Neurosci. 11, 81–96.
Langston, J.W., 1987. Parkinson’s disease: current view. Am. Fam.
Physician 35, 201–206.
Liang, C.L., Nelson, O., Yazdani, U., Pasbakhsh, P., German, D.C., 2004.
Inverse relationship between the contents of neuromelanin pigment and
the vesicular monoamine transporter-2: human midbrain dopamine
neurons. J. Comp. Neurol. 473, 97–106.
Liu, Y., Edwards, R.H., 1997a. Differential localization of vesicular
acetylcholine and monoamine transporters in PC12 cells but not CHO
cells. J. Cell Biol. 139, 907–916.
Liu, Y., Edwards, R.H., 1997b. The role of vesicular transport proteins in
synaptic transmission and neural degeneration. Annu. Rev. Neurosci.
20, 125–156.
Liu, Y., Roghani, A., Edwards, R.H., 1992a. Gene transfer of a reser-
pine-sensitive mechanism of resistance to MPP+. Proc. Natl. Acad.
Sci. U. S. A. 89, 9074–9078.
Liu, Y., Peter, D., Roghani, A., Schuldiner, S., Prive, G.G., Eisenberg, D.,
Brecha, N., Edwards, R.H., 1992b. A cDNA that supresses MPP+
toxicity encodes a vesicular amine transporter. Cell 70, 539–551.
Liu, Y., Schweitzer, E.S., Nirenberg, M.J., Pickel, V.M., Evans, C.J.,
Edwards, R.H., 1994. Preferential localization of a vesicular mono-
amine transporter to dense core vesicles in PC12 cells. J. Cell Biol. 127,
1419–1433.
Lotharius, J., Brundin, P., 2002. Pathogenesis of Parkinson’s disease:
dopamine, vesicles and alpha-synuclein. Nat. Rev., Neurosci. 3,
932–942.
Lotharius, J., O’Malley, K.L., 2000. The parkinsonism-inducing drug
1-methyl-4-phenylpyridinium triggers intracellular dopamine oxidation.
A novel mechanism of toxicity. J. Biol. Chem. 275, 38581–38588.
Lotharius, J., Dugan, L.L., O’Malley, K.L., 1999. Distinct mechanisms
underlie neurotoxin-mediated cell death in cultured dopaminergic
neurons. J. Neurosci. 19, 1284–1293.
Mooslehner, K.A., Chan, P.M., Xu, W., Liu, L., Smadja, C., Humby, T.,
Allen, N.D., Wilkinson, L.S., Emson, P.C., 2001. Mice with very low
expression of the vesicular monoamine transporter 2 gene survive into
adulthood: potential mouse model for parkinsonism. Mol. Cell. Biol.
21, 5321–5331.
Peter, D., Jimenez, J., Liu, Y., Kim, J., Edwards, R.H., 1994. The
chromaffin granule and synaptic vesicle amine transporters differ in
substrate recognition and sensitivity to inhibitors. J. Biol. Chem. 269,
7231–7237.
Przedborski, S., Jackson-Lewis, V., Naini, A.B., Jakowec, M., Petzinger,
G., Miller, R., Akram, M., 2001. The parkinsonian toxin 1-methyl-4-
phenyl-1,2,3,6-tetrahydropyridine (MPTP): a technical review of its
utility and safety. J. Neurochem. 76, 1265–1274.
Reinhard Jr., J.F., Diliberto Jr., E.J., Viveros, O.H., Daniels, A.J., 1987.
Subcellular compartmentalization of 1-methyl-4-phenylpyridinium
with catecholamines in adrenal medullary chromaffin vesicles may
explain the lack of toxicity to adrenal chromaffin cells. Proc. Natl.
Acad. Sci. U. S. A. 84, 8160–8164.
Scherman, D., Henry, J.P., 1981. The catecholamine carrier of Bovine
chromaffin granules. Form of the bound amine. Mol. Pharmacol. 23,
431–436.
Schuldiner, S., Shirvan, A., Linial, M., 1995. Vesicular neurotransmitter
transporters: from bacteria to humans. Physiol. Rev. 75, 369–392.
Staal, R.G., Hogan, K.A., Liang, C.L., German, D.C., Sonsalla, P.K., 2000.
In vitro studies of striatal vesicles containing the vesicular monoamine
transporter (VMAT2): rat versus mouse differences in sequestration of
1-methyl-4-phenylpyridinium. J. Pharmacol. Exp. Ther. 293, 329–335.
Stokes, A.H., Hastings, T.G., Vrana, K.E., 1999. Cytotoxic and genotoxic
potential of dopamine. J. Neurosci. Res. 55, 659–665.
Takahashi, N., Miner, L.L., Sora, I., Ujike, H., Revay, R.S., Kostic, V.,
Jackson-Lewis, V., Przedborski, S., Uhl, G.R., 1997. VMAT2 knockout
mice: heterozygous display reduced amphetamine-conditioned reward,
enhanced amphetamine locomotion and enhanced MPTP toxicity. Proc.
Natl. Acad. Sci. U. S. A. 94, 9938–9943.
Tan, P.K., Waites, C., Liu, Y., Krantz, D.E., Edwards, R.H., 1998. A
leucine-based motif mediates the endocytosis of vesicular monoamine
and acetylcholine transporters. J. Biol. Chem. 273, 17351–17360.
Thiele, C., Hannah, M.J., Fahrenholz, F., Huttner, W.B., 2000. Cholesterol
binds to synaptophysin and is required for biogenesis of synaptic
vesicles. Nat. Cell Biol. 2, 42–49.
Uhl, G.R., Li, S., Takahashi, N., Itokawa, K., Lin, Z., Hazama, M., Sora, I.,
2000. The VMAT2 gene in mice and humans: amphetamine responses,
locomotion, cardiac arrhythmias, aging, and vulnerability to dopami-
nergic toxins. FASEB J. 14, 2459–2465.
Wang, Y.-M., Gainetdinov, R.R., Fumagalli, F., Xu, F., Jones, S.R.,
Bock, C.B., Miller, G.W., Wightman, R.M., Caron, M.G., 1997.
Knockout of the vesicular monoamine transporter 2 gene results in
neonatal death and supersensitivity to cocaine and amphetamine.
Neuron 19, 1285–1296.
Weingarten, P., Zhou, Q.Y., 2001. Protection of intracellular dopamine
cytotoxicity by dopamine disposition and metabolism factors. J. Neuro-
chem. 77, 776–785.
Winkler, H., 1976. The composition of adrenal chromaffin granules: an
assessment of controversial results. Neuroscience 1, 65–80.
Zhang, Y., Dawson, V.L., Dawson, T.M., 2000. Oxidative stress and
genetics in the pathogenesis of Parkinson’s disease. Neurobiol. Dis. 7,
240–250.
Copyright © 2022 FDOKUMEN




















