
Expressing Arbitrary Reward Functions as Potential-Based Advice

Human endothelial cells expressing polyoma middle T induce tumors
Luca Primo1,2, Cristina Roca1,2, Chiara Ferrandi1,2, Luisa Lanfrancone3 and Federico Bussolino*,1,2
1Institute for Cancer Research and Treatment (I.R.C.C.), 10060 Candiolo (Torino), Italy; 2Department of Genetics, Biology andBiochemistry, School of Medicine, University of Torino, 10100 Torino, Italy; 3Department of Experimental Oncology, EuropeanInstitute of Oncology, 20100 Milano, Italy
The middle T oncogene of murine polyomavirus (PymT)rapidly transforms and immortalizes murine embryonicendothelial cells (EC), leading to the formation ofvascular tumors in newborn mice, by recruitment ofhost, non-transformed EC. These tumors are reminiscentof human vascular tumors like cavernous hemangioma,Kaposi's sarcoma or those characterizing Kasabach-Merrit syndrome. Here we investigate the in vitro and invivo behavior of human primary umbilical cord vein ECexpressing PymT. While PymT has been unable totransform human ®broblasts in earlier experiments orcontrols done here, mT expressing EC (PymT-EC)derived by infection with pLX-PymT retrovirus inducehemangiomas in nu/nu mice. These tumors contain notonly human cells but also recruited mouse EC as shownby the presence of human and murine CD31 positive EC.In vitro analysis shows that PymT-EC retain endothelialspeci®c markers like CD31, Von Willebrand factor, andVE-cadherin, and reach the con¯uence without signs ofovergrowth. They are also responsive to vascularendothelial growth factor-A. However, their proliferationrate is increased. The balance between urokinase-typeplasminogen activator and plasminogen activator inhi-bitor-1 is modi®ed; RNA and catalytic activity for theformer are elevated while PAI-1 RNA is reduced. Incontrast with murine model, where the PymT EC cellsbecome immortal, the e�ects induced by PymT in humanEC are transient. After 12 ± 15 passages, human PymTEC stop proliferating, assume a senescent phenotype,and lose the ability to induce hemangiomas. At the sametime both the amount of middle T protein and the level ofactivation of pp60c-src lower. Oncogene (2000) 19,3632 ± 3641.
Keywords: hemangioma; middle T; endothelial cells;polyomavirus; human
Introduction
Oncogenesis is a multi-step process that requires a longlatency period between the initiating event and theappearance of the tumor. However, in a few instances,expression of a single oncogene is su�cient to inducethe rapid development of tumors. In rodents, poly-omavirus induces a large variety of tumors, includingcarcinomas, lymphomas, hemangiomas and sarcomas(Gross, 1983). The expression of the middle Toncogene (PymT), one of the polyomavirus early
region genes, is su�cient to transform established®broblast cell lines, but cooperation with a secondoncogene such as polyomavirus large T, is required totransform primary cell cultures (Rassoulzadegan et al.,1982).
PymT has long been known to be su�cient toinduce hemangiomas in chicken (Kornbluth et al.,1986) or murine systems (Williams et al., 1988;Aguzzi et al., 1991). The murine system has beenstudied in the greatest detail. Transgenic miceexpressing PymT showed vascular tumors (Bautch etal., 1987), and a retroviral construct transducingPymT transformed murine endothelial cells (EC)(Williams et al., 1988; Bussolino et al., 1991;Garlanda et al., 1994; Giraudo et al., 1996). Wheninjected into mice these transformed cells causedhemangiomas, mostly composed of normal host ECand few cells with PymT (Williams et al., 1988;Garlanda et al., 1994). Though the mechanismsunderlying this rare and extreme type of oncogenesisare not understood, an imbalance of urokinase-typeplasminogen activator and its inhibitor is supposed tofavor abnormal degradation of extracellular matrix,permitting the migration of host cells (Montesano etal., 1990; Sabapathy et al., 1997).
The recruitment process is an example of positivetumor-host interactions where host cells acquire atumor phenotype or participate in the formation oftumor parenchyma. Transplantation of human tumorsin nu/nu mice resulted in the induction of stabletransplantable tumors of murine origin (Goldembergand Pavia, 1981, 1982; Beattie et al., 1982). In humans,Kaposi's sarcoma, a tumor generally considered tohave endothelial/mesenchymal origin, is characterizedby spindle-shaped cells representing the core of thelesion and eliciting an unbalanced network of solublemediators which is pivotal for mesenchymal cellrecruitment and lesion growth (Ensoli and Sturzl,1998). In this context a point of interest is that thesespindle cells cause vascular tumors by recruiting hostelements when injected into nu/nu mice (Salahuddin etal., 1988).
The molecular and cellular basis of cell recruitmentcontributing to human tumors, as well as the ontogenyof human hemangiomas and the possible involvementof a putative oncogene in their development areunknown (Drolet et al., 1999).
In this study, we demonstrate that PymT expressedin human EC by retroviral infection caused analteration of their phenotype. Furthermore, humanEC carrying PymT induced hemangiomas by host cellrecruitment in nu/nu mice and may be a useful modelto study the mechanisms of cell recruitment associatedto cancer and the pathogenesis of vascular tumors inhumans.
Oncogene (2000) 19, 3632 ± 3641ã 2000 Macmillan Publishers Ltd All rights reserved 0950 ± 9232/00 $15.00
www.nature.com/onc
*Correspondence: F Bussolino, I.R.C.C., Strada Provinciale 142,Km 3.95, 10060 Candiolo (Torino), ItalyReceived 22 February 2000; revised 11 April 2000; accepted25 May 2000

Results
EC-PymT expressed active mT oncogene
Human umbilical vein EC at ®rst passage were infectedwith the retroviral vector pLXSN carrying PymT orwith the vector alone. After selection with G418,PymT-EC and EC infected with vector alone (pLXSN-EC) were established. Figure 1 shows the PymToncogene expression in EC-PymT at 5th and 15thpassage. The Western blot analysis indicated thatPymT oncogene was expressed in EC-PymT at bothearly and late passages. However, the amount of thetransgene declined in older cultures (Figure 1a),whereas VE-cadherin expression, an endothelial cellspeci®c marker (Lampugnani et al., 1992), was notin¯uenced by culture passages (Figure 1b). These dataindicate that culture passages speci®cally a�ect middleT expression.
PymT has been shown to complex with, and elevatethe tyrosine kinase activity of pp60c-src (Bolen et al.,1984; Courtneidge and Smith, 1984). To verify theactivity of PymT oncogene, cell lysate of PymT-EC at5th and at 15th passage was immunoprecipitated by aspeci®c polyclonal antibody (pAb) anti-PymT (Mullaneet al., 1998) and the immunoprecipitates wereincubated with [g-32P]ATP and MG++ to visualize theassociated protein kinase activity. As expected, theimmunoprecipitates contained a phosphorylated bandof 56 kDa corresponding to PymT (Scha�hausen andBenjamin, 1981) (Figure 1c). Some experiments havebeen performed by adding enolase to the immunopre-
cipitates, in order to quantify the kinase activityassociated with PymT (Courtneidge and Smith, 1984;Kypta et al., 1990). Figure 1c shows that this substrateis more labeled in immunoprecipitates coming fromPymT-EC at early passages than in PymT-EC at latepassages. The labeling of enolase in PymT-EC at 5thpassage was 33-fold that in pLXSN-EC at the samepassage (Pym-T-EC: 4056+345 c.p.m.; pLXSN-EC:123+34; n=3). However, the enolase phosphorylationin PymT-immunoprecipitates from PymT-EC at 15thpassage was only 22-fold as compared to pLXSN-EC(PymT-EC: 2806+217 c.p.m.; pLXSN-EC: 131+41;n=3). The 50 kDa protein is far less phosphorylated inpLXSN-EC than PymT-EC and most likely representthe heavy chain of the immuno-precipitating antibody,which is an excellent substrate for the associated pp60src
kinase activity (Sartor and Robbins, 1993).
Characterization of PymT-EC
PymT-EC cell lines showed slight changes in morphol-ogy, but no sign of overgrowth (Figure 2b). Theyexpressed von Willebrand Factor (vWF) (Figure 2a),CD31 (Figure 2c) and VE-cadherin (not shown), threewell-known markers of vascular EC (Garlanda andDejana, 1997) with a similar pattern observed inpLXSN-EC (not shown). The expression of thesemarkers persisted up to 12 ± 15 passages when theexpression of PymT was lowered and the cells showeda senescent phenotype.
When grown in a tridimensional gel of extracellularmatrix proteins, EC develop networks of cellular cords
Figure 1 Human EC carrying PymT gene express the functional form of middle T oncogene. (a) Western blot analysis of PymToncogene in pLXSN-EC, PymT-EC at 5th passage (PymT-EC) and at 15th passage (PymT-EC*). (b) Analysis of VE-cadherinexpression performed on the same blot as in panel a. (c) In vitro phosphorylation of PymT oncogene and exogenous added enolasein pLXSN-EC (5th passage), PymT-EC (5th passage), and PymT-EC* (15th passage). The cells were lysed and subjected toimmunoprecipitation with anti-PymT antibody. After labeling in vitro with g-32P-ATP in the presence of 2.5 mg enolase, washedsamples were electrophoresed on a 8% SDS±PAGE and labeled bands visualized by autoradiography. In this experiment [32P]incorporation in enolase was as follows: pLXSN-EC 132 c.p.m.; pLXSN-EC: 3936 c.p.m.; pLXSN-EC*: 2863 c.p.m. Theseexperiments have been performed with similar results on six batches of PymT-EC and pLXSN-EC infected at di�erent times over aperiod of 20 months
Oncogene
Human endothelial cells carrying PymT induce hemangiomasL Primo et al
3633

that contain a central lumen as con®rmed bytransmission electron microscopy, a phenomenonnamed in vitro angiogenesis (Montesano et al., 1983,1993; Ingber and Folkman, 1989). In our experimentalconditions, control pLXSN-EC plated on a tridimen-sional gel of Matrigel formed branching, anastomozingcords with multicentric junctions within 18 h (Figure3a). In contrast, the e�ect of PymT expression in ECresulted in the impairment of in vitro angiogenic e�ect.PymT-EC organized in large cellular nodes withcobblestone-like morphology connected by a fewcord-like structures (Figure 3b). In order to excludethat the observed behavior of PymT-EC on Matrigel
was caused by a defect in adhesion machinery, wetested the adhesion of these cells on Matrigel or®bronectin coated plates. After 30 min of incubation,the number of adherent pLXSN-EC and PymT-ECwas similar (PymT-EC plated on Matrigel and on®bronectin: 1.1+0.36104 and 1.3+0.46104; pLXSN-EC plated on Matrigel and on ®bronectin: 1.2+0.16104 and 1.2+0.56104; n=3).
Expression of plasminogen activator urokinase-type andplasminogen activator inhibitor-1
When PymT-EC were suspended within ®brin gel, theydegraded it rapidly, suggesting that these cells had highlevels of ®brinolytic activity (data not shown). In fact,the plasminogen activator urokinase-type (uPA) activ-ity associated to cell membrane was increased in PymTtransduced cells (Table 1). To investigate the balancebetween uPA and plasminogen activator inhibitor-1(PAI-1), mRNA analysis was carried out. The level ofuPA transcript in PymT-EC was higher than inpLXSN-EC. When the same mRNA samples wereanalysed for PAI-1 expression, a marked decrease wasobserved as compared to control cells (Figure 4). Thesedata con®rm that PymT oncogene modi®es ®brinolytic
Figure 2 Expression of EC markers in PymT-EC. PymT-EC at5th passage seeded on collagen-coated coverslips were ®xed,permeabilized and stained with mAb anti-vWF (a) and anti-CD31(c) (magni®cation: 4006). (b) shows the same ®eld of aphotographed by phase-contrast microscope. These experimentshave been performed with similar results on four batches ofPymT-EC and pLXSN-EC infected at di�erent times over aperiod of 20 months
Figure 3 Phenotype of PymT-EC growth in a tridimensional gel.pLXSN-EC and PymT-EC at 6th passage were cultured onMatrigel for 18 h and observed by phase-contrast microscope.pLXSN-EC form branching, anastomozing cords (a) whereasPymT-EC organize in few cords which connect large cellularcobblestone-like areas (b) (magni®cation: 1006). These experi-ments have been performed with similar results on six batches ofPymT-EC and pLXSN-EC infected at di�erent times over aperiod of 16 months
Human endothelial cells carrying PymT induce hemangiomasL Primo et al
3634
Oncogene

potential of human EC as already reported in murineEC (Montesano et al., 1990).
PymT was unable to immortalize human EC
Expression of PymT in embryonic murine EC results inimmortal, established cell lines which retain thebiological features of EC and grow without additionof any speci®c endothelial growth factor (Williams etal., 1988; Bussolino et al., 1991; Garlanda et al., 1994;Giraudo et al., 1996).
Long-term culture of PymT-EC never gave rise topermanent cell lines. This suggested that PymTexpression was unable to immortalize human EC. Toexamine this in more detail, the growth properties ofPymT-EC were evaluated. PymT-EC at early passages(3 ± 10) grew more rapidly than pLXSN-EC inmedium containing 20% fetal calf serum (FCS).Moreover PymT-EC maintained their physiologicalmitogenic response to VEGF-A165. Like normalhuman EC, the proliferation of PymT-EC in presenceor absence of growth factors began to decline fromthe 11th to the 15th passage (Figure 5). To explorethe potential relationship between growth arrest andsenescence we looked for the presence of senescence-
associated b-galactosidase activity (SA-b-gal) (Dimri etal., 1995; Serrano et al., 1997) which has an optimalenzymatic activity at pH 6.0. The percentage of SA-b-gal positive cells in PymT-EC at early passages (3 ± 10)was low (passage 3: 1.20+0.05; passage 10: 3.4+0.21;n=3), but increased from the 10th passage reaching60 ± 70% of the cells at the 15th passage (13thpassage: 32.9+3.2; 15th passage: 63.4+9.65; n=3)(Figure 6).
Hemangiomas are induced by PymT-EC
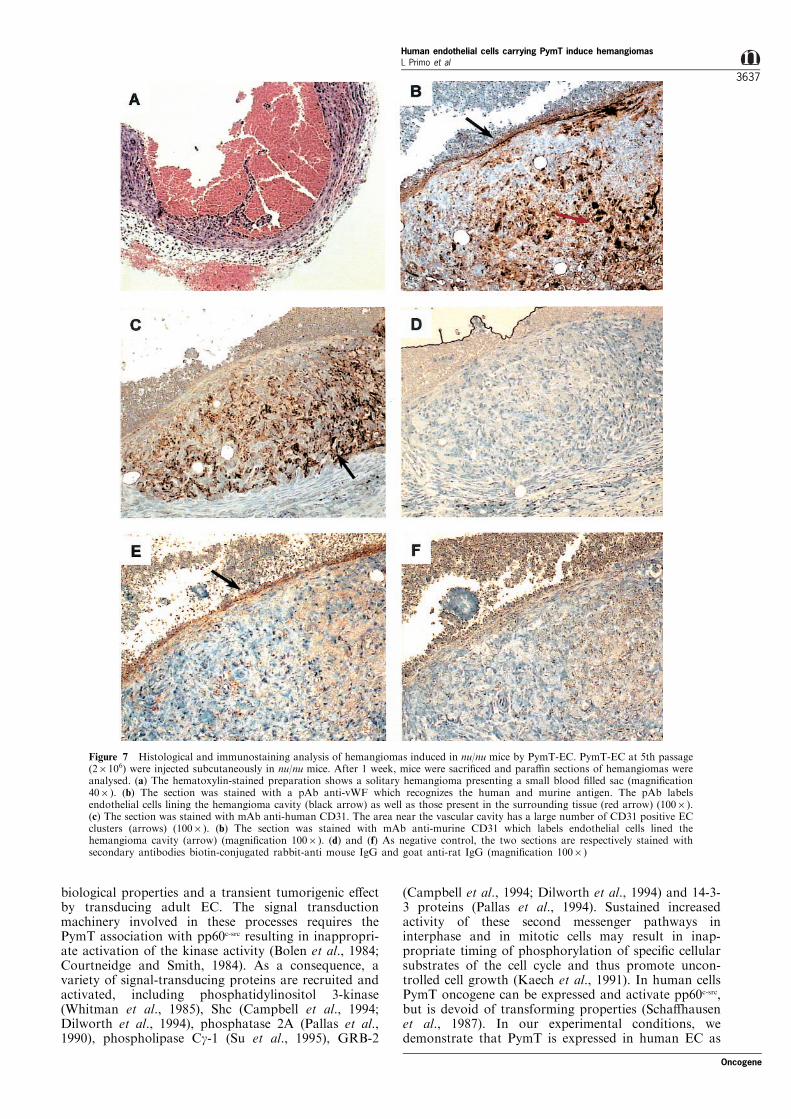
Subcutaneous injection of PymT-EC at early passagesinto nu/nu mice caused the appearance of hemangio-mas within 6 to 12 days at site of injection in allanimals injected. The ability of PymT-EC at laterpassages to induce hemangiomas was reduced (6/10)(P=0.0287 by Fisher's exact test) (Table 2). pLXSN-EC or primary human ®broblasts infected with pLX-PymT were unable to induce any lesions within 8weeks (Table 2). Hemangiomas were characterized bydiscrete blood-®lled cavities of variable size lined witha discontinuous layer of ¯attened cells (Figure 7).Histological sections were stained with speci®c mAbanti-vWF which recognizes human and murine cells,anti-human CD31 (Parums et al., 1990) and anti-mouse CD31 (Vecchi et al., 1994). Figure 7b showsthat hemangiomas were formed mainly by ECpositive for vWF. Moreover EC which lined thelarge cavities of hemangiomas were recognized by amAb anti-murine CD31 (Figure 7e), whereas thoseconstituting a heap near the cavities were recognizedby a mAb anti-human CD31 (Figure 7c). Theseresults show that the PymT-EC are recruiting hostEC to the tumor. With murine PymT-EC, hemangio-mas continue to grow. With human PymT-EC,regression of the hemangiomas was observed over aperiod of 3 weeks.
Table 1 uPA activity in human endothelial cells carrying mToncogenea
uPA released uPA cell associatedCells (IU/ml/24 h) (IU/16105 cells/24 h)
PymT-EC 0.04+0.002b 0.93+0.012*pLXSN-EC 0.05+0.009 0.32+0.010
auPa activity was measured by an indirect assay based on thecleavage of a chromogenic substrate by plasmin generated fromadded plasminogen. bMean+s.d. of ®ve determinations in one typicalexperiment out of three done. *P50.005 by t-test
Figure 4 Expression of uPA, PAI-1 and GAPDH mRNA in PymT-EC at 5th passage. Northern blot analysis of total RNA(15 mg) from PymT-EC and pLXSN-EC unstimulated (7) or stimulated with 20 ng/ml of VEGF-A165 (+) for 24 h for uPA mRNAlevels (a) and PAI-1 mRNA levels (b). These experiments have been performed with similar results on six batches of PymT-EC andpLXSN-EC infected at di�erent times over a period of 20 months from passage 5th to passage 10th
Oncogene
Human endothelial cells carrying PymT induce hemangiomasL Primo et al
3635

Discussion
In this study we demonstrate that human adult ECtransduced with a retroviral vector carrying PymTexhibit an altered phenotype in vitro and inducehemangioma formation in vivo. To our knowledge,this is the ®rst evidence that PymT is able to alterhuman cells and induce them to cause tumors inimmunode®cient mice. The e�ect of PymT seems to bespeci®c for EC, because human primary cultures of®broblasts infected by the same pLXSN-PymT vectorare not tumorigenic. The bulk of the tumor consists ofa cavity lined with a monolayer of murine EC and®lled with blood cells. Around it, there are capillariesand vessels constituted by human EC. This hemangio-ma has histological features similar to murine tumors(Williams et al., 1989), but there are many di�erencesbetween the two models. All nu/nu mice injected withPymT-EC survive and the hemangioma disappearsover a period of 3 weeks. In contrast, murine EC linescarrying PymT oncogene have a di�erent behavior innu/nu or syngenic mice. Hemangiomas induced bymurine cells in immunode®cient mice progress and killthe animals (Garlanda et al., 1994) and they do notregress in syngenic mice (Williams et al., 1989). This
di�erence could be in part explained by the di�erentcontext where the oncogene acts in murine and inhuman cells. Murine EC carrying PymT result to beimmortalized and transformed, but the e�ects of thisoncogene are evident on embryonic cells, suggestingthat murine EC precursors are permissive for middle Te�ect (Williams et al., 1988, 1989; Garlanda et al.,1994). Here, we obtained a transient change in cell
Figure 5 Proliferation curves of PymT-EC at 5th passage (~)and 15th passage (~) and of pLXSN-EC and 5th (*) and 15thpassage (*). 56103 cells were plated into 12-well-plates in 20%FCS. After overnight incubation, medium was changed andreplaced with fresh medium containing 20% FCS alone (a) orsupplemented with VEGF-A165 (20 ng/ml) (b). The cells were®xed at di�erent intervals of time and stained with 0.1% crystalviolet. Cell-associated dye was measured in microtiter plates at590 nm. Values were normalized to the optical density withstandard curves obtained with increasing number of PymT-EC orpLXSN-EC. Results (mean+s.d.) of one experiment (performedin triplicate) out of three are shown
Figure 6 Senescence-associated b-galactosidase (pH 6.0) activityin PymT-EC after the 5th passage (b) or in the same cells after the12th passage (a). The percentage of senescence associated b-gal(pH 6.0) positive cells was initially low, but increased at 12thpassage and reached 60% by 15th passage. Photographs are atthe same magni®cation (2006)
Table 2 Tumorigenic activity in nu/nu mice of human endothelialcells carrying mT oncogenea
Number of nudeNumber of nude mice developing
Cells mice injected hemangiomas
PymT-EC (5th passage) 12 12PymT-EC* (15th passage) 10 6pLXSN-EC (5th passage) 12 0Human fibroblast PymT 6 0(5th passage)
Human fibroblast pLXSN 4 0(5th passage)
anu/nu mice were injected subcutaneously with 26106 PymT-EC,pLXSN-EC and human ®broblasts infected with pLXSN or pLX-PymT. Mice were followed for 2 months. Tumors developed after 1week and disappeared after 3 weeks. Over a period of 2 monthspLXSN-EC and human ®broblasts did not develop any tumors andmice injected with PymT-EC did not show any tumor re-growth. Thepresented data have been obtained with the same batch of infectedcells. A second, similar experiment has been performed with similarresults by using two other batches of transduced cells
Human endothelial cells carrying PymT induce hemangiomasL Primo et al
3636
Oncogene
biological properties and a transient tumorigenic e�ectby transducing adult EC. The signal transductionmachinery involved in these processes requires thePymT association with pp60c-src resulting in inappropri-ate activation of the kinase activity (Bolen et al., 1984;Courtneidge and Smith, 1984). As a consequence, avariety of signal-transducing proteins are recruited andactivated, including phosphatidylinositol 3-kinase(Whitman et al., 1985), Shc (Campbell et al., 1994;Dilworth et al., 1994), phosphatase 2A (Pallas et al.,1990), phospholipase Cg-1 (Su et al., 1995), GRB-2
(Campbell et al., 1994; Dilworth et al., 1994) and 14-3-3 proteins (Pallas et al., 1994). Sustained increasedactivity of these second messenger pathways ininterphase and in mitotic cells may result in inap-propriate timing of phosphorylation of speci®c cellularsubstrates of the cell cycle and thus promote uncon-trolled cell growth (Kaech et al., 1991). In human cellsPymT oncogene can be expressed and activate pp60c-src,but is devoid of transforming properties (Scha�hausenet al., 1987). In our experimental conditions, wedemonstrate that PymT is expressed in human EC as
Figure 7 Histological and immunostaining analysis of hemangiomas induced in nu/nu mice by PymT-EC. PymT-EC at 5th passage(26106) were injected subcutaneously in nu/nu mice. After 1 week, mice were sacri®ced and para�n sections of hemangiomas wereanalysed. (a) The hematoxylin-stained preparation shows a solitary hemangioma presenting a small blood ®lled sac (magni®cation406). (b) The section was stained with a pAb anti-vWF which recognizes the human and murine antigen. The pAb labelsendothelial cells lining the hemangioma cavity (black arrow) as well as those present in the surrounding tissue (red arrow) (1006).(c) The section was stained with mAb anti-human CD31. The area near the vascular cavity has a large number of CD31 positive ECclusters (arrows) (1006). (b) The section was stained with mAb anti-murine CD31 which labels endothelial cells lined thehemangioma cavity (arrow) (magni®cation 1006). (d) and (f) As negative control, the two sections are respectively stained withsecondary antibodies biotin-conjugated rabbit-anti mouse IgG and goat anti-rat IgG (magni®cation 1006)
Oncogene
Human endothelial cells carrying PymT induce hemangiomasL Primo et al
3637

functional protein able to activate pp60c-src. Theoncogene o�ers a proliferative advantage to infectedcells, which is only transient and is abolished when thecells which have been cultured for 15 passages becamesenescent and the amount of oncogene expressed andthe activity of associated pp60c-src lowered. Thesechanges suggest a possible relationship between thedi�erent levels of PymT and of pp60c-src activity and thedi�erences in the proliferative response of EC. Incontrast, murine EC lines carrying mT and establishedby embryonic tissues have an increased and permanentactivation of signaling pathways triggered by mT,including phosphatidylinositol 3-kinase (Dong et al.,1996).
The in vitro transient e�ects of PymT on human ECsupport our in vivo results characterized by thedisappearance of hemangioma in nu/nu mice and thereduced tumorigenic e�ect of PymT-EC at laterpassages with a reduced growth rate. Similarly,mitomycin-C treated murine endothelioma cells, whichare unable to proliferate and have a short lifespan, stillinduced the formation of hemangiomas that regressedafter approximately 3 weeks (Williams et al., 1989).Murine EC carrying mT oncogene are highly immuno-genic (Garlanda et al., 1994; Biancone et al., 1999).Therefore, we cannot exclude that the PymT-EC-induced hemangiomas can be rejected by the residualimmunological response of nu/nu mice rather than aspontaneous regression due to the intrinsic features ofthe injected cells. However, this hypothesis is notsupported by the murine model. Hemangiomas inducedby murine EC carrying PymT do not regress in nu/numice (Garlanda et al., 1994; Williams et al., 1989),suggesting that the di�erent in vivo behavior of humanand murine PymT-EC is not obligatorily the result of adi�erent immunological response in the two models,but re¯ects di�erent molecular mechanisms of PymToncogene in the two cell types. In the light of ourresults, it is intriguing to speculate that PymToncogene does not a�ect the cell cycle of human EC,but modi®es transiently their phenotype, which favorsthe appearance of vascular transient lesions by host cellrecruitment. This altered phenotype includes (1) theunbalance of ®brinolytic system; and (2) the reducedability to di�erentiate into mature capillary, assuggested by the observation that PymT-EC form fewcapillary-like structures when plated on Matrigel. Itshould be also possible for the induction of autocrineloops involving soluble mediators as already reportedin mice (Montesano et al., 1990; Taraboletti et al.,1993; Ghigo et al., 1995; Muhlner et al., 1999).
This phenomenon is reminiscent of Kaposi's sarco-ma, a multifocal proliferative disease of vascular origin.Early lesions are in®ltrated by leukocytes, producing avariety of in¯ammatory cytokines, (Ensoli and Sturzl,1998), that induce normal cells to acquire the Kaposi'ssarcoma phenotype associated with HIV infection(Fiorelli et al., 1995). These cytokines can alsostimulate Kaposi's sarcoma cells themselves to produceother soluble mediators, including angiogenic factors.These molecules dictate the progression of the lesion byautocrine and paracrine mechanisms, regulating cellrecruitment and growth, with consequent angiogenesisand lesion formation (Ensoli and Sturzl, 1998). In somecases, Kaposi's sarcoma can regress spontaneously(Levy and Ziegler, 1983; Trattner et al., 1993), but in
later stages of development often associated to HIV-1infection, a cell clone may assume neoplastic features,subsequent to genotypic alterations, causing thehyperplastic lesions of Kaposi's sarcoma to transforminto real sarcomas (Lunardi-Iskandar et al., 1995;Albini et al., 1997). A putative candidate for thistransformation is human herpesvirus-8 (Flore et al.,1998).
Furthermore, it is also relevant to underline that acommon feature of human hemangiomas is a rapidproliferative phase followed by spontaneous involution(Drolet et al., 1999). Indeed the described model couldbe a useful tool to study involuting hemangiomas.
It has been recently reported for the ®rst time, theminimum number of de®ned genetic alterations unableto transform primary cultures of human epithelial and®broblasts cells. They include the ectopic expression ofthe telomerase catalytic subunit in combination withH-Ras and simian virus 40 large T oncogene (Hahn etal., 1999). Therefore, in contrast to murine EC modelwhere PymT oncogene transform in a single step way(Aguzzi et al., 1991; Williams et al., 1988), we canhypothesize that an unidenti®ed human oncogenesharing similar features to PymT could play a role asco-factor in the human vascular oncogenesis. Thishypothesis could open new perspectives in thepathogenesis of human vascular tumors and urges toidentify the sequence of molecular alterations leadingto vascular tumors.
Materials and methods
Cells
Human EC were extracted from umbilical cord veins growthand characterized as previously described (Bussolino et al.,1992). Human ®broblasts were obtained from human skin ofpatients submitted to dermatological surgery, digested for 1 hat 378C in DMEM (Sigma) containing 0.1% trypsin (Gibco)and 0.1% collagenase (Boehringer). After two washes withDMEM containing 10% FCS (Gibco), cell suspension wasplated in 100 mm diameter plastic dishes and grown until thecon¯uence. Fibroblasts were characterized for the expressionof vimentin, TE7 antigen, collagen I and collagen II, and forthe absence of cytocheratin, desmin, vWF, CD45 and smoothmuscle actin.
Retroviral-mediated gene transfer
Polyoma mT oncogene was subcloned in XbaI-Xho site of theexpression vector pLXSN under control of LTR promoter.Amphotropic cell line PA317 was transfected with pLXSNand pLXSN-PymT to producing retroviral supernatant withhigh viral titer. The virus-containing medium was collected,®ltered (0.45 mm ®lter, Millipore) and supplemented with4 mg/ml polybrene (Sigma). EC or human ®broblasts, at Ipassage, were plated in 100 mm tissue culture dishes in M19920% FCS and, for infections, the culture medium wasreplaced by the appropriate viral supernatant. The target cellswere incubated for 5 h at 378C and 5% CO2 and then thesupernatant was replaced by 10 ml of M199 20% FCS. After48 h, infected cells were placed under selection in G418(Gibco) (500 ng/ml) for 1 week.
Immunofluorescence studies
PymT-EC and pLXSN-EC were grown on coverslips coatedwith collagen I (Sigma), incubated for 24 h in medium M199
Human endothelial cells carrying PymT induce hemangiomasL Primo et al
3638
Oncogene

containing 20% FCS, ®xed at room temperature for 5 minwith paraformaldehyde (3%) solution, pH 7.6, containing 2%sucrose and then permeabilized (3 min at 488C) by Triton X-100 bu�er (20 mM HEPES, 300 mM sucrose, 50 mM NaCl,3 mM MgCl2, 0.5% Triton X-100, pH 7.4) (Bussolino et al.,1987). The coverslips were washed twice with phosphatebu�ered saline (PBS), incubated with 10% goat serum for10 min and then incubated 1 h at 378C with di�erent mAbs:anti-vWF (diluted 1 : 10) (Boehringer), anti-human CD31(diluted 1 : 50) (Dako), anti-VE-cadherin (diluted 1 : 50)(Lampugnani et al., 1992). The coverslips were washed threetimes in PBS, incubated 1 h in ¯uorescent isothiocyanate(FITC) conjugated goat anti-mouse (Sigma), washed threetimes in PBS and mounted in PBS-glycerol (1 : 1, v/v).Coverslips were observed by an Olympus (BX-60) microscopeequipped for epi¯uorescence.
Western blot analysis and immunoprecipitation experiment
The presence of PymT and its functional activity wasdetected by Western blot and by an in vitro kinase assay,respectively. Total proteins (15 mg) from con¯uent cells wereseparated by sodiumdodecyl sulphate polyacrilammide gelelectrophoresis (SDS ±PAGE) (8%), immunoblotted withmouse mAb (PM116 (1 : 100) which recognizes PymT (Hol-man et al., 1994), and visualized by ECL system (Pharmacia-Amersham-Biotech). For immunoprecipitation and in vitrokinase assay, con¯uent cells were washed three times withPBS containing 1 mM Na ortho-vanadate and lysed at 48C ina 25 mM HEPES bu�er pH 7.6, containing 150 mM NaCl,1% NP-40, and protease and phosphates inhibitors (pep-statin, 50 mg/ml; leupeptin, 50 mg/ml; aprotinin, 10 mg/ml;2 mM PMSF; soybean trypsin inhibitor, 500 mg/ml; ZnCl2,100 mM; Na ortho-vanadate, 1 mM). After centrifugation(20 min, 10 000 g), supernatants were pre-cleared by incuba-tion for 1 h with Protein A-Sepharose (Sigma). Samples(700 mg of proteins) were incubated with 5 ml of rabbitantiserum 45-1 which recognizes PymT in the aminoacidresidues 280 ± 300 (provided by Dr B Scha�hausen) for 1 hand immune complexes were recovered on Protein A-Sepharose. Beads were washed four times with lysis bu�er,twice with the same bu�er without NP-40 and once with tris-bu�ered saline. The in vitro kinase assay was performed in20 mM Tris-HCl bu�er (pH 7.2) containing 10 mM MgCl2,10 mCi [g32P]ATP (4000 Ci/mmole, Pharmacia-Amersham-Biotech), at room temperature for 10 min. In someexperiments, acid inactivated rabbit enolase (Sigma) (2.5 mgper assay) was added as substrate (Kypta et al., 1990).Proteins were separated by SDS±PAGE (8%), and the gelwas exposed to X-ray ®lms for 24 h. The incorporation oflabel into enolase was assayed by counting the excised bands.
RNA extraction and Northern analysis
Total cellular RNA was isolated by RNAzol (Tel-Test Inc.)according to manufacturer's instruction by PymT-EC andpLXSN-EC. Equal amounts of total RNA (15 mg/lane) wereelectrophoresed in 1% agarose gels containing 6.3%formaldehyde in 3-morpholino-propanesulfonic acid bu�er(Sigma) and blotted on a Nylon Duralon-UV membrane(Stratagene). Hybridization was carried out overnight at 428Cwith a-32P-dCTP-labeled (3000 Ci/mmol, Amersham) humanuPA probe (a 1.5 Kb PstI-PstI fragment of uPA), humanPAI-1 probe (a 3.2 Kb BglI-EcoRI fragment of PAI-1), andglyceraldehyde phosphate dehydrogenase (GAPDH) probe (a1.2 Kb PstI-PstI fragment).
In vitro matrigel and fibrin gel assays
Matrigel (Collaborative Biomedical Products) (0.4 ml) wasadded to each well of a 24-well plate and incubated at 378Cto allow gel formation. PymT-EC and pLXSN-EC at 80 ±
90% con¯uence were trypsinized, washed twice with PBS,plated on Matrigel at 4 ± 56104 cell/well, incubated in 5%CO2 at 378C for 18 h and then photographed.Fibrin gel was prepared dissolving bovine ®brinogen
(Sigma) immediately before the use in calcium andmagnesium free DMEM (Sigma) to obtain a ®nal proteinconcentration of 2.5 mg/ml and ®ltered through a 0.45 mmMillipore ®lter. Bovine thrombin (Sigma), at the concentra-tion of 25 U/ml (Sigma), was added 1/10 (v/v) to the®brinogen. The mixture was then immediately transferredinto 48-well plates and allowed to gel for 2 min before adding56104 PymT-EC or pLXSN-EC. Fresh complete medium(M199 supplemented with 20% FCS) was added to the wells.The ®brin gel cultures were subsequently incubated at 378Cfor 1 week. The culture medium was renewed every 2 ± 3 days(Montesano et al., 1990).
Cell growth assay
Cells (56103/well) were seeded in 24-well plates and allowedto attach for 16 h in M199 containing 20% FCS. Then, freshmedium with or without 20 ng/ml of VEGF-A165 (R&D)supplemented with 20% FCS was added. Cells were ®xed in2.5% glutaraldehyde, stained with 0.1% crystal violet in 20%methanol, and solubilized in 10% acetic acid (Bussolino etal., 1992). Cell growth was evaluated by measuringabsorbance at 590 nm in a plate reader (Perkin-Elmer). Acalibration curve was set up with known numbers of PymT-EC or pLXSN-EC and a linear correlation betweenabsorbance and cell counts was established up to 36105 cells.
uPA activity assay
To measure uPA activity released in the medium andassociated to cell surface, PymT-EC or pLXSN-EC weregrown at con¯uence on 48-well plates, washed and incubatedfor 36 h in M199 without phenol red, supplemented with20% FCS. uPA activity present in the medium was measuredby an indirect assay based on the generation of plasmin fromplasminogen. Fifty ml of ®ltered conditionated medium in50 ml of assay bu�er [50 mM Tris with 0.1% polyethilengli-cole 8000, 6 mM 6-aminohexanoic acid, and 0.1 mg/mlbovine plasminogen (American Diagnostica Inc.) pH 7.4]were incubated for 15 min at room temperature with0.125 mmoles of H-D-norleucyl-hexahydrotyrosyl-lysine-p-ni-troanilide diacetate salt (American Diagnostica Inc.), aspeci®c chromogenic substrate of plasmin. The adsorbancewas read at 404 nm in a plate reader (Perkin Elmer). Tomeasure cell associated uPA activity, 50 ml of assay bu�erand 50 ml of chromogenic substrate were added. After 15 minat room temperature the supernatants was recovered andadsorbance was read as described before. A calibration curvewas set up using an urokinase activity standard (AmericanDiagnostica Inc.).
Adhesion assay
Following coating of 96-well plates with Matrigel or®bronectin (Sigma) (both at 10 mg/ml) and saturation withPBS containing 3% bovine serum albumin (Sigma) for 2 h at378C, 26104 cells in 100 ml of growth medium were plated intriplicate. After incubation for 60 min at 378C, wells werewashed in PBS, ®xed in 2.5% glutaraldehyde (100 ml/well) for30 min and stained with 0.1% crystal violet and processed asdescribed before.
Injection in nu/nu mice and immunohistochemistry
PymT-EC, pLXSN-EC and human ®broblasts infected withpLX-PymT were dispersed with trypsin-EDTA, washed twicein M199 supplemented with 20% FCS, and resuspended inPBS. Three week-old nu/nu mice (Charles River Italia) were
Oncogene
Human endothelial cells carrying PymT induce hemangiomasL Primo et al
3639

inoculated subcutaneously with 26106 cells. Mice weremonitored daily, and size of the lesion was measured withcalibre. At sacri®ce, lesions were removed, ®xed in 4%bu�ered formaline and included in para�n. Tissue sections(7 mm) were depara�nized, rehydrated and incubated for10 min with 3% H2O2 in water to block endogenousperoxidase. Then a preincubation (30 min at room tempera-ture) with normal non-immune serum (1 : 50) was followed byincubation overnight with primary mAbs anti-human CD31(1 : 30) (Dako) and anti-murine CD31 (Vecchi et al., 1994),and pAB anti-vWF (1 : 100) (Dako). After three washes thesamples were incubated respectively, with biotinylated rabbitanti-mouse immunoglobulins (Dako), goat anti-rat immuno-globulins (Sigma) and goat anti-rabbit immunoglobulins(Dako), and then with streptABC complex/horseradishperoxidase (Dako). Tissue sections were washed anddeveloped with 3,3-diaminobenzidine tetrahydrochloride,and stained with hematoxylin.
Senescence-associated b-galactosidase
PymT-EC and pLXSN-ECs were washed once with PBS (pH7.2), ®xed with 3% formaldehyde, washed in PBS andincubated at 378C in the dark with fresh senescence-associated b-Gal stain solution (1 mg of 5-bromo-4-chloro-3-indolyl b-D-galactosidase dissolved in 1 ml PBS pH 6.0,5 mM potassium ferricyanide, 5 mM potassium ferrocyanide,
2 mM MgCl2) (Dimri et al., 1995). Staining was evident in 2 ±4 h and reached the maximum after 12 ± 16 h.
AcknowledgmentsWe are particularly grateful to B Scha�hausen (TuftsUniversity, Boston, USA) for antibodies anti-PymT andfor critically reading and reviewing of the manuscript. Theauthors thank: Drs E Dejana, A Vecchi (Istituto `MarioNegri', Milano, Italy) for Ab anti-human VE-cadherin andanti-murine CD31; Prof F Blasi (Ospedale San Ra�aele-Dibit, Milano, Italy) for uPA and PAI-1 cDNAs; Dr MRisio (Institute for Cancer Research and Treatment,Ordine del Mauriziano, Candiolo, Italy) for his suggestionsand encouragement in histological analysis; Prof MFDiRenzo (Institute for Cancer Research and Treatment,Candiolo, Italy) and Prof A Mantovani (Istituto `MarioNegri', Milano, Italy) for their fruitful discussions. Thisstudy was supported by Italian Association for CancerResearch (A.I.R.C.), Istituto Superiore di SanitaÁ (Pro-gramma Nazionale di Ricerca sull'AIDS: Patogenesi,immunitaÁ e vaccino per l'AIDS (40B.19) e Patologia,clinica e terapia dell'AIDS (30B.9), and Program onTumor Therapy), Ministero dell'UniversitaÁ e della RicercaScienti®ca e Tecnologica (60% and Programma di Ricercadi Rilevante Interesse Nazionale-1998 and 1999), andRegione Piemonte.
References
Aguzzi A, Kleihues P, Heckl K and Wiestler O. (1991).Oncogene, 6, 113 ± 118.
Albini A, Paglieri I, Orengo G, Carlone S, Aluigi MG,DeMarchi R, Matteucci C, Mantovani A, Carozzi F,Donini S and Benelli R. (1997). AIDS, 11, 713 ± 721.
Bautch VL, Toda S, Hassel AJ and Hanahan D. (1987). Cell,51, 529 ± 538.
Beattie GM, Knowles AF, Jensen FC, Baird SM and KaplanNO. (1982). Proc. Natl. Acad. Sci. USA, 79, 3033 ± 3036.
Biancone L, Stamenkovic I, Cantaluppi V, Boccellino M, DeMartino A, Bussolino F and Camussi G. (1999). J.Immunol., 162, 5263 ± 5268.
Bolen JB, Thiele CJ, Israel MA, Yonemoto W, Lipsich LAand Brugge JS. (1984). Cell, 38, 767 ± 777.
Bussolino F, Camussi G, Aglietta M, Braquet P, Boisa A,Pescarmona G, Sanavio F, D'Urso N and Marchiso PC.(1987). J. Immunol., 139, 2439 ± 2446.
Bussolino F, De Rossi M, Sica A, Colotta F, Wang JM,Bocchietto E, Padura IM, Bosia A, Dejana E andMantovani A. (1991). J. Immunol., 147, 2122 ± 2129.
Bussolino F, Di Renzo MF, Ziche M, Bocchietto E, OliveroM, Naldini L, Gaudino G, Tamagnone L, Co�er A andComoglio PM. (1992). J. Cell Biol., 119, 629 ± 641.
Campbell KS, Ogris E, Burke B, Su W, Auger KR, DrukerBJ, Scha�hausen BS, Roberts TM and Pallas DC. (1994).Proc. Natl. Acad. Sci. USA, 91, 6344 ± 6348.
Courtneidge SA and Smith AE. (1984). EMBO J., 3, 585 ±681.
Dilworth SM, Brewster CE, Jones MD, Lanfrancone L,Pelicci G and Pelicci PG. (1994). Nature, 367, 87 ± 90.
Dimri GP, Lee X, Basile G, Acosta M, Scott G, Roskelley C,Medrano EE, Linskens M, Rubelj I, Pereira-Smith O,Peacocke M and Campisi J. (1995). Proc. Natl. Acad. Sci.USA, 92, 9363 ± 9367.
Dong QG, Graziani A, Garlanda C, Wainstok De Calma-novici R, Arese M, Soldi R, Vecchi A, Mantovani A andBussolino F. (1996). Int. J. Cancer, 65, 700 ± 708.
Drolet BA, Esterly NB and Frieden IJ. (1999). N. Engl. J.Med., 341, 173 ± 181.
Ensoli B and Sturzl M. (1998). Cytokine Growth Factor Rev.,9, 63 ± 83.
Fiorelli V, Gendelman R, Samaniego F, Markham PD andEnsoli B. (1995). J. Clin. Invest., 95, 1723 ± 1734.
Flore O, Ra®l S, Ely S, O'Leary JJ, Hyjek EM and CesermanE. (1998). Nature, 394, 588 ± 592.
Garlanda C and Dejana E. (1997). Arterioscler. Thromb.Vasc. Biol., 17, 1193 ± 1202.
Garlanda C, Parravicini C, Sironi M, De Rossi M, Wainstokde Calmanovici R, Carozzi F, Bussolino F, Colotta F,Mantovani A and Vecchi A. (1994). Proc. Natl. Acad. Sci.USA, 91, 7291 ± 7295.
Ghigo D, Arese M, Todde R, Vecchi A, Silvagno F,Costamagna C, Dong QD, Alessio M, Heller R, Soldi R,Trucco F, Garbarino G, Pescarmona G, Mantovani A,Bussolino F and Bosia A. (1995). J. Exp. Med., 181, 9 ± 19.
Giraudo E, Arese M, Toniatti C, Strasly M, Primo L,Mantovani A, Ciliberto G and Bussolino F. (1996). J.Immunol., 157, 2618 ± 2623.
Goldemberg DM and Pavia RA. (1981). Science, 212, 65 ±67.
Goldemberg DM and Pavia RA. (1982). Proc. Natl. Acad.Sci. USA, 79, 2389 ± 2392.
Gross L. (1983). Oncogenic viruses, Vol. 2. Press, P. (ed):Oxford, pp. 737 ± 828.
Hahn WC, Counter CM, Lundberg AS, Beijersbergen RL,Brooks MW and Weinberg RA. (1999). Nature, 400, 464 ±468.
Holman PS, Gjoerup OV, Davin T and Scha�hausen BS.(1994). J. Virol., 68, 668 ± 673.
Ingber DE and Folkman J. (1989). J. Cell. Biol., 109, 317 ±330.
Kaech S, Covic L, Wyss A and Ballmer-Hofer K. (1991).Nature, 350, 431 ± 433.
Kornbluth S, Cross FR, Harbison M and Hanafusa H.(1986). Mol. Cell. Biol., 5, 1545 ± 1555.
Kypta RM, Goldberg Y, Ulug ET and Courtneidge SA.(1990). Cell, 62, 481 ± 492.
Lampugnani MG, Resnati M, Raiteri M, Pigott R, PisacaneA, Houen G, Ruco LP and Dejana E. (1992). J. Cell. Biol.,118, 1511 ± 1522.
Levy JA and Ziegler JL. (1983). Lancet, 2, 78 ± 81.
Human endothelial cells carrying PymT induce hemangiomasL Primo et al
3640
Oncogene

Lunardi-Iskandar Y, Gill P, Lam VH, Zeman RA, MichaelsF, Mann DL, Reitz MS, Kaplan M, Berneman ZN andCarter D. (1995). J. Natl. Cancer, 87, 974 ± 981.
Montesano R, Pepper MS, Mohle-Steinlein U, Risau W,Wagner FE and Orci L. (1990). Cell, 62, 435 ± 445.
Montesano R, Pepper MS and Orci L. (1993). J. Cell. Sci.,105, 1013 ± 1024.
Montesano R, Orci L and Vassalli P. (1983). J. Cell. Biol., 97,1648 ± 1652.
Muhlner U, Mohle-Steinlein U, Wizigmann-Voos S, Chris-tofori G, Risau W and Wagner EF. (1999). Oncogene, 18,4200 ± 4210.
Mullane KP, Ratnofsky M, Cullere X and Scha�hausen B.(1998). Mol. Cell. Biol., 18, 7556 ± 7564.
Pallas DC, Fu H, Haehnel LC, Weller W, Collier RJ andRoberts TM. (1994). Nature, 265, 535 ± 537.
Pallas DC, Shahrik LK, Martin BL, Jaspers S, Miller TB,Brautigan DL and Robert TM. (1990). Cell, 60, 167 ± 176.
Parums DV, Cordell JI, Micklem K, Heryet AR, Gatter KCand Mason DC. (1990). J. Clin. Pathol., 43, 752 ± 757.
Rassoulzadegan M, Cowie A, Carr A, Glaichenhaus N,Kamen R and Cuzin F. (1982). Nature, 300, 713 ± 718.
Sabapathy KT, Pepper SM, Kiefer F, Mohle-Steinlein U,Tacchini-Cottier F, Fekta I, Breier G, Risau W, CarmelietP, Montesano R and Wagner EF. (1997). J. Cell. Biol.,137, 953 ± 963.
Salahuddin SZ, Nakamura S, Biberfeld P, Kaplan MH,Markham PD, Larsson L and Gallo RC. (1988). Science,242, 430 ± 433.
Sartor O and Robbins KC. (1993). J. Biol. Chem., 268,21014 ± 21020.
Scha�hausen B and Benjamin TL. (1981). J. Virol., 40, 184 ±196.
Scha�hausen BS, Bockus BJ, Berkner KL, Kaplan D andRoberts TM. (1987). J. Virol., 61, 1221 ± 1225.
Serrano M, Lin AW, McCurrach ME, Beach D and LoweSW. (1997). Cell, 88, 593 ± 602.
Su W, Liu W, Scha�hausen SB and Roberts MT. (1995). J.Biol. Chem., 270, 12331 ± 12334.
Taraboletti G, Belotti D, Dejana E, Mantovani A andGiavazzi R. (1993). Cancer Res., 53, 3812 ± 3816.
Trattner A, Hodac E, David M and Sandbank M. (1993).Cancer, 72, 1779 ± 1783.
Vecchi A, Garlanda C, Lampugnani MG, Resnati M,Matteucci C, Stoppacciaro A, Schnurch H, Risau W,Ruco L and Mantovani A. (1994). Eur. J. Cell. Biol., 63,247 ± 254.
Whitman M, Kaplan DR, Scha�hausen B, Cantley L andRoberts TM. (1985). Nature, 315, 239 ± 242.
Williams LR, Courtneidge SA and Wagner EF. (1988). Cell,52, 121 ± 131.
Williams RL, Risau W, Zerwes HG, Aguzzi A and WagnerFE. (1989). Cell, 57, 1053 ± 1063.
Oncogene
Human endothelial cells carrying PymT induce hemangiomasL Primo et al
3641
Copyright © 2022 FDOKUMEN




















