
Disturbed Expression of Sox9 in Pre-Sertoli Cells Underlies Sex-Reversal in Mice B6.Ytir

0013-7227/91/1295-2345$03.00/0EndocrinologyCopyright © 1991 by The Endocrine Society
Vol. 129, No. 5Printed in U.S.A.
Identification, Characterization, and HormonalRegulation of 3',5'-Cyclic Adenosine MonophosphateDependent Protein Kinases in Rat Sertoli Cells*BRYNJAR F. LANDMARK, BEATHE FAUSKE, WINNIE ESKILD,BJ0RN SKALHEGG, SUZANNE M. LOHMANN, VIDAR HANSSON,TORE JAHNSEN, AND STEPHEN J. BEEBEt
Institute of Medical Biochemistry (B.F.L., W.E., V.H., T.J., S.J.B.), University of Oslo, N-0317 Oslo,Norway; Department of Anatomy (B.F.), University of Bergen, N-5009 Bergen, Norway; Institute ofPathology (B.S., T.J., S.J.B.), Rikshospitalet, N-0027 Oslo, Norway; and Medizin Universitdts Klinik, Laborfur Klinische Biochemie, 8700 Wurzburg, Germany
ABSTRACT. Recent studies have disclosed multiple isoformsof regulatory (R) and catalytic (C) subunits of cAMP-dependentprotein kinase (PKA) at the protein and messenger RNA(mRNA) levels. The purpose of the present study was to identify,characterize, and quantify individual R subunits in rat Sertolicells both at the mRNA and protein levels.
Unstimulated Sertoli cells contain high levels of R (~9.2 ±0.8 pmol/mg protein) and C (~7.3 ± 0.7 pmol/mg protein).Stimulation with (Bt)2cAMP (0.1 mMJ for 24 and 48 h revealeda time-dependent increase in [3H]cAMP-binding activity. Dur-ing the same time period the catalytic activity remained rela-tively constant, resulting in an increase in the R/C ratio fromapproximately 1.3 to 3.0.
Using diethylaminoethyl cellulose chromatography, 8-N3-[32P]cAMP photoaffinity labeling, autophosphorylation by y-[32P]ATP, and specific antibodies, we show that unstimulatedSertoli cells contain approximately 75% RIa, 25% RIIa, and very
low levels of RII3. Stimulation of Sertoli cells with (Bt)2cAMP(0.1 mM, 48 h) was associated with a 2.1-fold increase in RIO(6.6-14 pmol/mg) and a 10- to 20-fold increase in RII,, (<0.1-1.1 pmol/mg), with little or no change in RIIa (1.9-2.3 pmol/mg). Treatment with cAMP was associated with a slight increasein RI/RII ratio (3.3-4.1).
mRNA levels for RII,, increased 30- to 50-fold after(Bt)2cAMP stimulation, whereas only minor changes in mRNAlevels for RIa, RII«, and Co were observed (1.5- to 2.0-fold).mRNA levels for RIft C , and Cy were not detected in eitherunstimulated or in cAMP-stimulated Sertoli cells.
It is concluded that chronic treatment with cAMP changesthe relative proportion of R subunits of PKA in a mannerreflecting the changing levels in respective mRNAs. Further-more, such treatment; is associated with the appearance of a newPKA R subunit (Rllf), which is absent in untreated Sertoli cells.{Endocrinology 129: 2345-2354,1991)
SERTOLI cells provide a structural and functionalenvironment for the maintenance and progression
of mammalian spermatogenesis. FSH regulates a diver-sity of Sertoli cell functions through the activation ofadenylate cyclase and elevation of intracellular cAMP(1). Almost all known effects of cAMP in mammaliancells are believed to be mediated by cAMP-dependent
Received July 3, 1991.Address all correspondence and requests for reprints to: Dr. Brynjar
F. Landmark, Institute of Medical Biochemistry, University of Oslo,P.O. Box 1112 Blindern, N-0317 Oslo, Norway.
*This work was supported by grants from the Norwegian CancerSociety (to T.J. and V.H.), the Norwegian Research Council for Scienceand the Humanities (NAVF) (to T.J. and V.H.), Torsteds (to T.J. andV.H.), Anders Jahres Foundation for the Promotion of Science (to T.J.and V.H.), the Nordic Insulin Foundation (to T.J. and V.H.), the USEducation Foundation, Fulbright (to S.J.B.), and the Norwegian Mar-shall Fund (to S.J.B.).
fPresent address: Jones Institute for Reproductive Medicine, Dept.of Obstetrics/Gynecology, Eastern Virginia Medical School, Norfolk,VA 23507.
protein kinase (PKA) through subsequent phosphoryla-tion of substrate proteins. These include diverse proc-esses such as: regulation of carbohydrate and lipid me-tabolism, ion transport, enzyme induction, gene regula-tion (2), and secretion of androgen-binding protein(ABP) (3).
Two major isozymes of PKA, designated type I andtype II, are present in mammalian tissues (4). Thesediffer in the properties of the dimeric regulatory (R)subunit, which associates with the catalytic (C) subunitto inhibit its action in the absence of cAMP. Recentwork involving complementary DNA (cDNA) cloninghas revealed great heterogeneity of the PKA subunits.In mammals four closely related but distinct isoforms ofthe R subunits have been demonstrated, RIa (5, 6), RI^(7), RIIa, (8-10), and RII^ (11,12). Furthermore, cDNAsfor three isoforms of C subunits have been isolated andcharacterized, Co: (13, 14), C (15-17), and Cy (18).
Messenger RNAs (mRNAs) for various subunits of
2345

2346 PKA IN RAT SERTOLI CELLS Endo • 1991Vol 129* No 5
PKA in testis tissue reveal cell-specific distribution anddistinct regulation by hormones and cAMP (19). At theprotein level, it has been difficult to separately study theisomeric forms of the different subunits of PKA. Theaim of the present study was to identify, characterize,and quantify at the protein level the individual subunitsof PKA in Sertoli cells and examine possible regulatoryeffects of cAMP. In some of the experiments the levelsof the corresponding mRNAs were determined.
Materials and Methods
Sertoli cell culture
Sertoli cells were isolated from testes of 19-day-old immaturerats (Sprague-Dawley, Moller, Copenhagen, Denmark), andtreated as previously described (20, 21). Experiments werestarted on day 5 after plating. Fresh medium was added 12 hbefore stimulation and every 24 h thereafter.
RNA extraction and Northern transfer
Cells from two culture dishes (10 cm in diameter) werewashed three times in 4 ml each of ice-cold Ca2+- and Mg2+-free Hanks balanced salt solution (HBSS) and then scrapedand collected in HBSS. The cells were sedimented at 1500 rpmfor 5 min in a Sorvall (DuPont Co., Newton, CT) GLC-1centrifuge at 4 C. Three milliliters of 5 M guanidium isothio-cyanate were added, and the cells were homogenized with 20strokes of a tight-fitting pestle in a Dounce (Kebo-Lab, Oslo,Norway) homogenizer. Final RNA extraction was performedusing phenol/chloroform (22). RNA quantification was basedon absorbance at 260 nm. Twenty micrograms of RNA weredenatured in 50% formamide, 6% formaldehyde by heating at50 C for 15 min, then RNA was chilled on ice for 15 min andresolved on a 1.5% agarose gel containing 6.7% formaldehyde/20 mM sodium phosphate, pH 7.0.
RNA was blotted onto Biotrans (ICN, Irvine, CA) nylonfilters with 20x SSC (3 M NaCl, 0.3 M sodium citrate), andfilters were subsequently baked for 1 h at 80 C. Prehybridiza-tion, hybridization with nick-translated cDNA probes (1 X 106
cpm/ml, SA, 1-5 X 108 cpm/Vg DNA), and wash steps werecarried out as previously described (21). Autoradiography wasperformed using Kodak XAR film (Eastman Kodak Co., Roch-ester, NY) or Amersham Hyperfilm (Amersham, ArlingtonHeights, IL). After autoradiography, filters were washed andreprobed with subsequent cDNAs.
Preparation of Sertoli cell extracts
Before homogenization, cells in culture dishes were washedwith 3 X 5 ml Eagle's modified Minimum Essential Medium,then reincubated in a large vol (30-50 ml) of media for 1 h toremove cAMP before cAMP-binding experiments. The culturedishes were cooled for 15 min on ice and washed with homog-enization buffer [3 x 5 ml PE buffer (10 mM potassium phos-phate, 1 mM EDTA, pH 6.8) with 250 mM sucrose (PES)], andcells were scraped with 0.5 ml PES containing protease inhib-itors (10 /zg/ml each of chymostatin, antipain, leupeptin, andpepstatin) per dish. The cells were homogenized using 25
strokes with a tight-fitting Dounce homogenizer and centri-fuged (10 min, 15,000 X g, 4 C). Assays were performed on thesupernatants immediately after homogenization. Samples forgel electrophoresis were frozen in liquid nitrogen and stored at-70 C.
Triton extracts were made by several different methods.After centrifugation as above, the pellets were resuspended ineither PES buffer or PE buffer containing 150 mM NaCl and1% Triton X-100. Triton extracts were prepared either bymixing them well and leaving them for 1 h at 4 C or by applying10 strokes of a tight-fitting Dounce homogenizer to the samples.The samples were centrifuged (15,000 X g, 10 min, 4 C), andresidual insoluble particles were discarded.
cAMP-binding assays, protein kinase assays, anddetermination of R/C ratio
These assays were performed as described by Cobb andCorbin (23). In brief, the kinase assay contained 130 nM Kemp-tide substrate peptide, 200 fiM [7-32P]ATP, 25 mM magnesiumacetate, and 5 MM CAMP in 50 mM Tris-HCl (pH 7.4). The R/C ratio was calculated after determining the concentration ofR by [3H]cAMP binding [based on two cAMP binding sites permonomer (24)] and the concentration of C by phosphorylationof Kemptide based on the specific activity (15 /miol/min • mg,40,000 g/mol) of homogeneous C purified from bovine heart. Insoluble extracts from Sertoli cells Kemptide phosphorylationin the absence of cAMP was less than 2% of that seen in thepresence of cAMP.
Protein concentration
Protein concentration was determined by the method ofBradford (25), using a 7-globulin standard. All samples wereassayed in parallel at three different dilutions within the linearrange of the standard curve.
Diethylaminoethyl (DEAE)-cellubse chromatography
Two identical Whatman (Clifton, NJ) DE 52 columns (0.9X 7.5 cm, 4.8 ml) were equilibrated in PE-buffer (pH 6.8) inpreparation for analysis of untreated and (Bt)2cAMP-treatedSertoli cells. Extracts from equal amounts of cells were appliedto each column, and gradients (0-0.4 M NaCl, total vol 50 ml),created with a Buchler Instruments (Fort Lee, NJ) 2 5070Agradient mixer, were applied. Each column was washed with 30ml (6 bed vol) PE buffer, and fractions of 1 ml (approximately1 ml/min) were collected. Kinase assays and cAMP bindingassays were performed immediately after elution. The NaClconcentrations were determined by conductivity measure-ments.
Purification of PKA regulatory subunits by affinitychromatography
Cellular extracts prepared in PE buffer containing 10 mM /3-mercaptoethanol and 50 Kallikrein-inhibiting units/ml Trasy-lol (PEMT buffer) with 250 mM sucrose were applied directlyto a Sepharose-NH-(CH2)6-NH-C8-cAMP [8-amino (hexylam-ino)-cAMP Sepharose] affinity column (0.5 ml resin) whichhad been equilibrated with PEMT buffer.
PKA regulatory subunits were eluted in 6 M urea, dialyzed

PKA IN RAT SERTOLI CELLS 2347
overnight in PE buffer at 4 C, frozen in aliquots, and stored at—70 C. Binding assays were performed, and equal amounts ofbinding activity were analyzed by sodium dodecyl sulfate-poly-acrylamide gel electrophoresis (SDS-PAGE). The subunitswere visualized by Coomassie blue staining or by 8-N3-[
32P]cAMP-photoaffmity labeling and autophosphorylation using7-[32P]ATP and purified C subunit followed by autoradiogra-phy (see below).
Gel electrophoresis
Denaturing PAGE was performed using slab gels containing7.5% polyacrylamide in the separating gel and 4.5% polyacryl-amide in the stacking gel (26). The samples were denatured byboiling for 2 min in a solution containing final concentrationsof 1.4% SDS, 20% 0-mercaptoethanol, and 0.005% Bromophe-nol Blue.
Western blotting technique
Protein transfer onto nitrocullulose filter (Ancos, 01stykke,Denmark, 0.45 /tim) was performed on a semi-dry electroblotter(Ancos) following a procedure modified from Towbin et al. (27).Protein transfer buffer contained 20% methanol, 0.4 M glycine,and 0.05 M Tris. Transfer was performed at 100 mA overnightor at 200 mA for 3 h. The extent of transfer was judged byCoomassie blue staining with a solution of 20% methanol and7% acetic acid containing 0.1% Coomassie blue for at least 30min, followed by destaining with 20% methanol.
Radioimmunolabeling
Radioimmunolabeling of Western blots employing specificantibodies for the different R subunits was performed as pre-viously described by Jahnsen et al. (28). The RIIa and RII^antisera were characterized by Lohmann et al. (29).
8-Ns-P2P]cAMP incorporation
8-N3-[32P]cAMP was concentrated for 5 min in a Speed Vac
(Savant Instruments, Hicksville, NY), and the original meth-anol was replaced by an equal vol of distilled water. Labelingwas performed on crude protein extracts (1 mg/ml) or onpurified R subunits (170 Mg/ml) (30) for 30 min on ice in thedark with 1 nM 8-N3-[
32P]cAMP ± 1 mMunlabeled cAMPfollowed by exposure to UV light (Mineralight Lamps, modelUVS-54, Ultra-Violet Products, San Gabriel, CA, 254 nm), for15 min (at 10 cm). Denaturing solution was added, and thesamples were boiled as above. Proteins were separated by SDS-PAGE, Coomasie blue stained, and dried between sheets ofcellophane on a slab dryer (BioRad, Richmond, CA, model 483,60 C, 2 h). Autoradiograms were prepared using Kodak double-sided intensifying screens and Amersham Hyperfilm or KodakXAR film.
Phosphorylation of purified R using y-P2P]ATP
Autophosphorylation of RII subunits by purified bovine Csubunit was performed in a reaction mixture containing 10 mMpotassium phosphate, 1 mM EDTA, 200 »M 7-[32P]ATP (2 /xCi/mmol), 10 mM Mg(acetate)2, and excess C subunit (~50 ng//il).The reaction was left for 30 min at 30 C, then placed on icebefore stopping by addition of denaturing solution and boilingfor 2 min.
Immunoquantification
RI and total RII were quantified by incubating samples with[3H]cAMP, then adding the respective antisera and protein A-Sepharose and retaining bound [3H]cAMP on polypropylenecolumns (BioRad 731 1550) followed by elution in 1 M aceticacid as previously described by Ekanger et al. (31).
Materials
[3H]cAMP 34 Ci/mmol and 7-[32P]ATP (-5000 Ci/mmol)were purchased from Amersham, goat antirabbit [125I]immu-noglobulin G (7.6 fiCi/ng) was obtained from NEN Research(Boston, MA) and 8-N3-[
32P]cAMP (SA, 60-75 Ci/mmol) fromICN Biochemicals (Cleveland, OH). FSH (NIDDK-oFSH-17)was a gift from the NIH (Bethesda, MD). cAMP, AMP, 8-(6-aminohexyl) amino cAMP, N6-2'-O-dibutyryl cAMP, N6-ben-zoyl cAMP, 8-(4-chlorophenylthio)-cAMP, ATP grade I,aprotinin, and histone (calf thymus) type II-AS were purchasedfrom Sigma (St. Louis, MO). Chymostatin, antipain, leupeptin,pepstatin, and Kemptide phosphate acceptor protein were pur-chased from Peninsula Laboratories (Belmont, CA). Penicillinwas obtained from Apothekernes Laboratorium (Oslo, Nor-way), streptomycin from Evans PL (Longhurst, Horsham, Eng-land), and fungizone from Novo Industries (E. R. Squibb &Sons, Princeton, NJ). Fetal calf serum, Eagle's Minimum Es-sential Medium, and HBSS without Ca2+ and Mg2+ were pur-chased from GIBCO (Grand Island, NY). The 8-(6-amino-hexyl)amino cAMP affinity column was from Pharmacia(Bromma, Sweden). SDS, acrylamide, bis, temed, ammoniumpersulfate, 2-mercaptoethanol, Tris, glycine, Bromophenol blueand Coomassie blue were obtained from BioRad. All otherchemicals were of analytical grade.
Results
Effects of Bt2cAMP on mRNA levels of PKA subunits
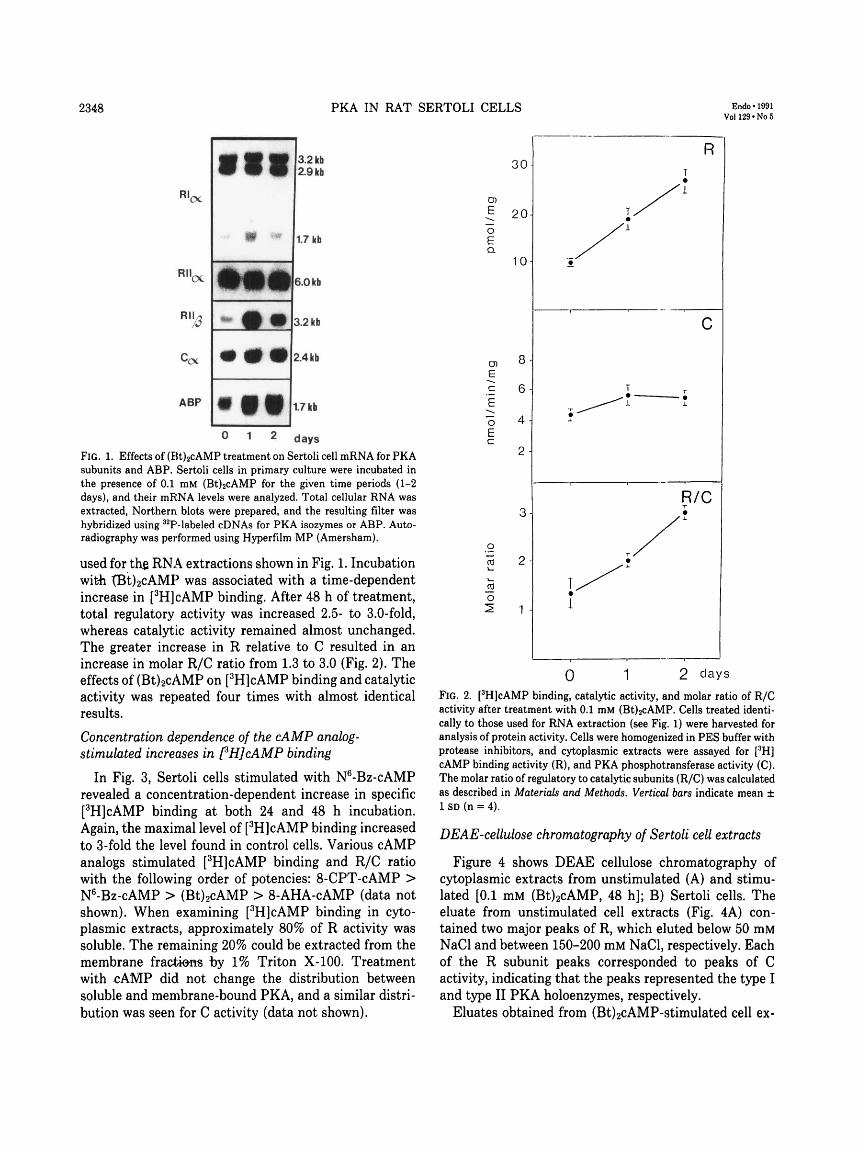
Figure 1 depicts autoradiographs of Northern filtersafter treatment of Sertoli cell cultures with (Bt)2cAMP(0.1 mM) for 24 and 48 h, respectively. As reported earlier(21), cAMP maximally induced all PKA mRNAs within24 h. RILj mRNA was most affected, with a 30- to 50-fold increase relative to basal levels. Treatment with(Bt)2cAMP had less effects on mRNAs for RIa, C«, andRIIa, although small but reproducible increases (1.5- to3-fold) were seen.
A cAMP-responsive gene, ABP, was used as a control,since its mRNA level is known to increase in Sertoli cellsafter (Bt)2cAMP stimulation. Treatment of Sertoli cellswith butyrate (2 mM) did not affect any of the PKAsubunit mRNA levels (data not shown).
Effects of (Bt)2cAMP on fHJcAMP binding and PKA-phosphotransferase activity
Figure 2 depicts the effects of (Bt)2cAMP (0.1 mM) on[3H]cAMP binding to R subunits and C activity of PKA.The R/C ratio was also calculated. The cells used in thisexperiment received treatment identical to that of cells
2348 PKA IN RAT SERTOLI CELLS Endo • 1991Voll29«No5
Rl( X
Rlloc
ABP
• ti
1
3.2 kb2.9 kb
1.7 kb
6.0 kb
3.2 kb
2.4 kb
1.7 kb
days
FIG. 1. Effects of (Bt)2cAMP treatment on Sertoli cell mRNA for PKAsubunits and ABP. Sertoli cells in primary culture were incubated inthe presence of 0.1 mM (Bt)2cAMP for the given time periods (1-2days), and their mRNA levels were analyzed. Total cellular RNA wasextracted, Northern blots were prepared, and the resulting filter washybridized using 32P-labeled cDNAs for PKA isozymes or ABP. Auto-radiography was performed using Hyperfilm MP (Amersham).
used for th$ RNA extractions shown in Fig. 1. Incubationwith tBt)2cAMP was associated with a time-dependentincrease in [3H]cAMP binding. After 48 h of treatment,total regulatory activity was increased 2.5- to 3.0-fold,whereas catalytic activity remained almost unchanged.The greater increase in R relative to C resulted in anincrease in molar R/C ratio from 1.3 to 3.0 (Fig. 2). Theeffects of (Bt)2cAMP on [3H]cAMP binding and catalyticactivity was repeated four times with almost identicalresults.
Concentration dependence of the cAMP analog-stimulated increases in pHJcAMP binding
In Fig. 3, Sertoli cells stimulated with N6-Bz-cAMPrevealed a concentration-dependent increase in specific[3H]cAMP binding at both 24 and 48 h incubation.Again, the maximal level of [3H]cAMP binding increasedto 3-fold the level found in control cells. Various cAMPanalogs stimulated [3H]cAMP binding and R/C ratiowith the following order of potencies: 8-CPT-cAMP >N6-Bz-cAMP > (Bt)2cAMP > 8-AHA-cAMP (data notshown). When examining [3H]cAMP binding in cyto-plasmic extracts, approximately 80% of R activity wassoluble. The remaining 20% could be extracted from themembrane fractions by 1% Triton X-100. Treatmentwith <:AMP did not change the distribution betweensoluble and membrane-bound PKA, and a similar distri-bution was seen for C activity (data not shown).
O)
E
oEa
E
c
E
oEc
3-
2 -
1 -
0 1 2 days
FIG. 2. [3H]cAMP binding, catalytic activity, and molar ratio of R/Cactivity after treatment with 0.1 mM (Bt)2cAMP. Cells treated identi-cally to those used for RNA extraction (see Fig. 1) were harvested foranalysis of protein activity. Cells were homogenized in PES buffer withprotease inhibitors, and cytoplasmic extracts were assayed for [3H]cAMP binding activity (R), and PKA phosphotransferase activity (C).The molar ratio of regulatory to catalytic subunits (R/C) was calculatedas described in Materials and Methods. Vertical bars indicate mean ±1 SD (n = 4).
DEAE-cellubse chromatography of Sertoli cell extracts
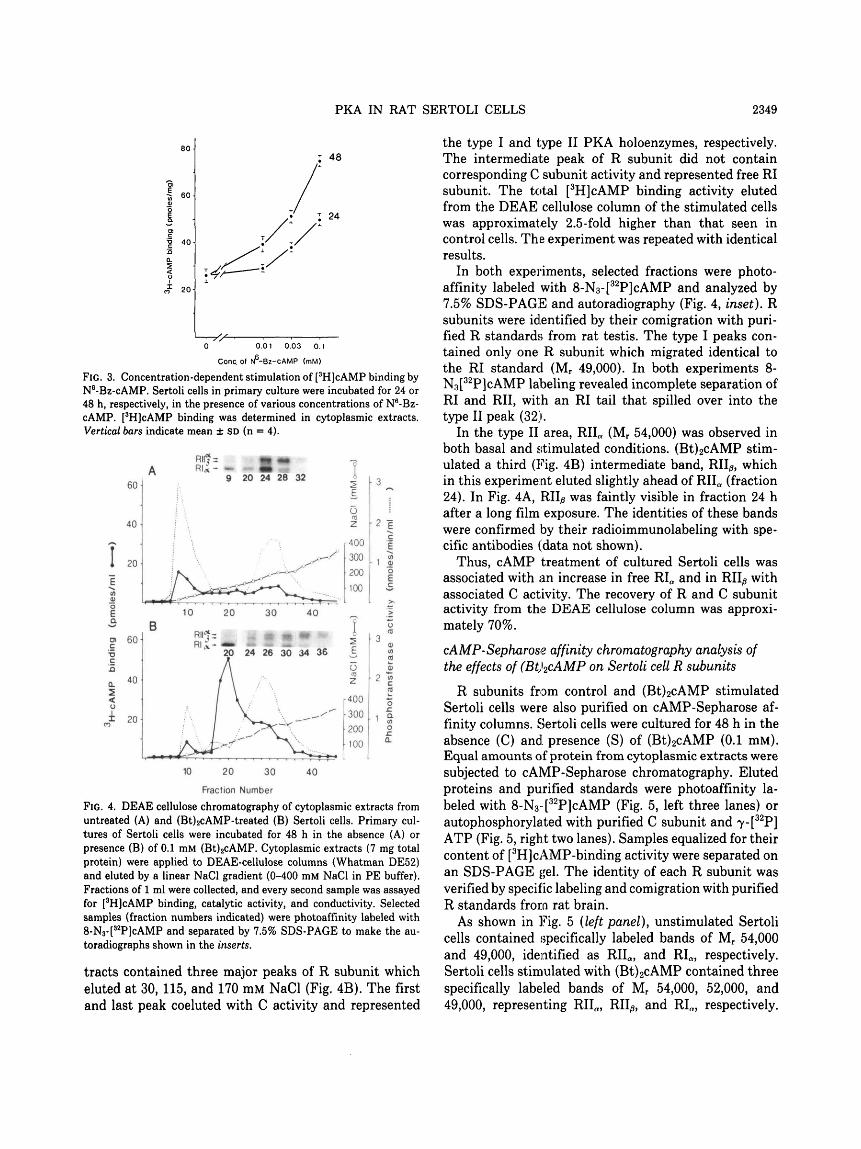
Figure 4 shows DEAE cellulose chromatography ofcytoplasmic extracts from unstimulated (A) and stimu-lated [0.1 mM (Bt)2cAMP, 48 h]; B) Sertoli cells. Theeluate from unstimulated cell extracts (Fig. 4A) con-tained two major peaks of R, which eluted below 50 mMNaCl and between 150-200 mM NaCl, respectively. Eachof the R subunit peaks corresponded to peaks of Cactivity, indicating that the peaks represented the type Iand type II PKA holoenzymes, respectively.
Eluates obtained from (Bt)2cAMP-stimulated cell ex-
PKA IN RAT SERTOLI CELLS 2349
: 48
24
0 0.01 0.03 0.1
Cone, of N^-Bz-cAMP (mM)
FIG. 3. Concentration-dependent stimulation of [3H]cAMP binding byN6-Bz-cAMP. Sertoli cells in primary culture were incubated for 24 or48 h, respectively, in the presence of various concentrations of N6-Bz-cAMP. [3H]cAMP binding was determined in cytoplasmic extracts.Vertical bars indicate mean ± SD (n = 4).
1 0)
o) 60-
10 20 30 40
RII'Js ill?* I20 24 26 30 34 36 £
0)(0
to
2 2
20 30 40
Fraction Number
FIG. 4. DEAE cellulose chromatography of cytoplasmic extracts fromuntreated (A) and (Bt)2cAMP-treated (B) Sertoli cells. Primary cul-tures of Sertoli cells were incubated for 48 h in the absence (A) orpresence (B) of 0.1 mM (Bt)2cAMP. Cytoplasmic extracts (7 mg totalprotein) were applied to DEAE-cellulose columns (Whatman DE52)and eluted by a linear NaCl gradient (0-400 mM NaCl in PE buffer).Fractions of 1 ml were collected, and every second sample was assayedfor [3H]cAMP binding, catalytic activity, and conductivity. Selectedsamples (fraction numbers indicated) were photoaffinity labeled with8-N3-[
32P]cAMP and separated by 7.5% SDS-PAGE to make the au-toradiographs shown in the inserts.
tracts contained three major peaks of R subunit whicheluted at 30, 115, and 170 mM NaCl (Fig. 4B). The firstand last peak coeluted with C activity and represented
the type I and type II PKA holoenzymes, respectively.The intermediate peak of R subunit did not containcorresponding C subunit activity and represented free RIsubunit. The total [3H]cAMP binding activity elutedfrom the DEAE cellulose column of the stimulated cellswas approximately 2.5-fold higher than that seen incontrol cells. The experiment was repeated with identicalresults.
In both experiments, selected fractions were photo-affinity labeled with 8-N3-[
32P]cAMP and analyzed by7.5% SDS-PAGE and autoradiography (Fig. 4, inset). Rsubunits were identified by their comigration with puri-fied R standards from rat testis. The type I peaks con-tained only one R subunit which migrated identical tothe RI standard (Mr 49,000). In both experiments 8-N3[
32P]cAMP labeling revealed incomplete separation ofRI and RII, with an RI tail that spilled over into thetype II peak (32).
In the type II area, RIIa (Mr 54,000) was observed inboth basal and stimulated conditions. (Bt)2cAMP stim-ulated a third (Fig. 4B) intermediate band, RII/3, whichin this experiment eluted slightly ahead of RIIa (fraction24). In Fig. 4A, RII^ was faintly visible in fraction 24 hafter a long film exposure. The identities of these bandswere confirmed by their radioimmunolabeling with spe-cific antibodies (data not shown).
Thus, cAMP treatment of cultured Sertoli cells wasassociated with an increase in free RIa and in RII^ withassociated C activity. The recovery of R and C subunitactivity from the DEAE cellulose column was approxi-mately 70%.
cAMP-Sepharose affinity chromatography analysis ofthe effects of (Bt)2cAMP on Sertoli cell R subunits
R subunits from control and (Bt)2cAMP stimulatedSertoli cells were also purified on cAMP-Sepharose af-finity columns. Sertoli cells were cultured for 48 h in theabsence (C) and. presence (S) of (Bt)2cAMP (0.1 mM).Equal amounts of protein from cytoplasmic extracts weresubjected to cAMP-Sepharose chromatography. Elutedproteins and purified standards were photoaffinity la-beled with 8-N3- [
32P]cAMP (Fig. 5, left three lanes) orautophosphorylated with purified C subunit and Y-[3 2P]
ATP (Fig. 5, right two lanes). Samples equalized for theircontent of [3H]cAMP-binding activity were separated onan SDS-PAGE gel. The identity of each R subunit wasverified by specific labeling and comigration with purifiedR standards from rat brain.
As shown in Fig. 5 (left panel), unstimulated Sertolicells contained specifically labeled bands of Mr 54,000and 49,000, identified as Rlla, and Rla, respectively.Sertoli cells stimulated with (Bt)2cAMP contained threespecifically labeled bands of Mr 54,000, 52,000, and49,000, representing RII«, RII^, and RIa, respectively.

2350 PKA IN RAT SERTOLI CELLS Endo • 1991Voll29«No5
origin-8-Ng p P] c AM P [T-32- P] ATP
dyefront-
FIG. 5. 8-N3-[32]PcAMP labeling (left) and autophosphorylation
(right) of affinity-purified R subunits from control and (Bt)2cAMP-stimulated Sertoli cells. Primary Sertoli cell cultures were incubatedfor 48 h in the absence (C) and presence (S) of 0.1 mM (Bt2)cAMP.Cellular extracts were applied to 8-(6-aminohexyl)-amino-cAMP affin-ity columns, and R subunits were eluted with 8 M urea. The R subunitswere dialyzed and photoaffinity labeled with 8-N3-[
32P]cAMP (left) orautophosphorylated with [T-3 2P]ATP in the presence of purified PKAC subunit (right). Equal amounts of [3H]cAMP binding activity weresubjected to SDS-PAGE in 7.5% acrylamide gels. Purified regulatorysubunits (50 ng) from rat brain were used as a standard (Std).
Mr
103
52^:4 9 -
RI
i
mm
c
ms
RII
* ~
C
mm
s
R
c
ns
FlG. 6. Effects of (Bt)2cAMP on R subunit immunoactivity. Primarycultures of Sertoli cells were incubated for 48 h in the absence (A) orpresence (B) of 0.1 mM (Bt)2cAMP. Cellular extracts were subjected toSDS-PAGE in 7.5% gels, electroblotted onto nitrocellulose, and incu-bated with monospecific antibodies for RIO) RIIa) and RIL. Bands werevisualized after incubation with [125I]immunoglobulin G (goat antirab-bit) and autoradiography. Purified rat brain R subunits were used asstandards.
Both RII subunits, but not RI, were autophosphorylatedby C subunit (Fig. 5, right two lanes). A smaller mol wtband of approximately 35,000 Mr was autophosphoryl-ated and labeled with 8-N3-[
32P]cAMP, indicating thatit may be a proteolytic fragment of RII, retaining thehinge region (containing the autophosphorylation site)and the cAMP binding sites of RII.
Radioimmunolabeling analysis of the effects of(Bt)2cAMP on Sertoli cell R subunits
Extracts from Sertoli cell cultures were separated on7.5% SDS-PAGE, transferred to nitrocellulose filters,and incubated with antibodies monospecific for each ofthe R subunits (Fig. 6). As shown, treatment of culturedSertoli cells for 48 h (S) resulted in a distinct increase in
RI« immunostaining. Densitometric scanning of autora-diographs indicated that the increase in RI« intensitywas 2.2-fold. Treatment of cultured Sertoli cells for 48 hwith (Bt)2cAMP (0.1 mM) was not associated with anychange in RIIa immunoactivity (Fig. 6, middle panel),whereas immunostaining of RILj in the same cells (Fig.6, right panel) increased dramatically (>10-fold).
Quantification of individual R subunit forms by PH]cAMP binding and immunoimmobilization
To perform more quantitative estimates of the abso-lute levels of the various R subunits in control and(Bt)2cAMP stimulated Sertoli cells, [3H]cAMP-labeledRI and RII subunits were removed from cell extractsusing columns containing specific antibodies and proteinA Sepharose. Using this procedure the antibodies coulddistinguish between RI and RII subunits, but poorlybetween RIIa and RII^. Therefore, the results of Westernanalysis (Fig. 6) were used to estimate the changes inRIIa and RILj after (Bt)2cAMP stimulation. From thisdata we calculated the levels of RIIa and RII^ in controland stimulated cells (Table 1). As shown in the table,(Bt)2cAMP treatment was associated with a 2.1-foldincrease in RI subunits (RIa). A similar change in RIa
protein levels was observed by Western analysis followedby densitometric scanning. The total amounts of RIIwere determined by immunoimmobilization to be ap-proximately 25-30% of that of RIa both in control andstimulated cells. The calculated levels of RIIa from cellscultured in the absence and presence of (Bt)2cAMP were1.9 and 2.3 pmol/mg protein, respectively. In comparison,(Bt)2cAMP stimulated the levels of RIIp from less than0.1 to 1.1 pmol/mg protein (Table 1). Thus, in nonstim-ulated Sertoli cells RIIa is more than 20 times that ofRII/s, whereas in the stimulated Sertoli cells we findcomparable amounts of RIIa and RII^. The ratio of RI toRII increased slightly during stimulation with(Bt)2cAMP.
TABLE 1. Estimate of specific R subunit levels in Sertoli cells
RIa RII Ratio RIIO RII,(pmol/mg) (pmol/mg) (RI/RII) (pmol/mg) (pmol/mg)
Control 6.6 ±0.3 2.0 ± 0.1 3.3 1.9 0.1(Bt)2cAMP 14.0 ±2.1 3.4 ±0.6 4.1 2.3 1.1
(n = 4) (n = 4)
Sertoli cells were incubated for 48 h in the absence (Control) orpresence of 0.1 mM (Bt)2cAMP in the growth medium. R subunits wereassayed by [3H]cAMP binding and immunoimmobilization as describedin Materials and Methods. Levels of individual RII subspecies werecalculated/estimated from the total RII levels and their specific in-crease from densitometric scanning of the Western blot autoradiograms(shown in Fig. 5). Values for [3H]cAMP binding are means ± SD forfour separate Sertoli cell isolations.

PKA IN RAT SERTOLI CELLS 2351
Discussion
The role of cAMP in reproductive function is relativelywell characterized and involves the covalent modificationof protein substrates via phosphorylation by the catalyticsubunit of PKA. PKA regulates cell function via post-translational activation or inactivation of enzymes thatregulate metabolism or by activation or inactivation ofgene regulatory proteins (2, 33, 34). A number of proteinsare induced by cAMP in various cell systems, and someof these are also substrates for PKA (35). In addition,several PKA autoregulatory components have been dem-onstrated, including autophosphorylation of the regula-tory subunits (36) and modification of cAMP-dependentphosphodiesterase activities via PKA phosphorylation(37, 38). cAMP also has been shown to regulate compo-nents of its own signal transduction system in thatseveral of the PKA mRNAs are regulated by cAMP viatranscriptional and/or posttranscriptional mechanisms(21, 39). In the present study we have characterized thecAMP-stimulated induction of mRNA and protein forall the PKA subunits in rat Sertoli cells.
The multiplicity of PKA subunits has recently beenmore fully defined, primarily through cDNA cloning andsequencing (5, 7-8,12,14-16). In Sertoli cells, the PKAsare represented by RIa, RIIa, RUp, and Ca, whereas Cp,Rip, and Cy are absent or present at very low levels (21).
Because caveats are inherent in each method for themeasurement of PKA protein levels, we have used severalmethods in the present investigation in order to substan-tiate the conclusions made. These include Western blot-ting analysis and [3H]cAMP binding before and afterDEAE cellulose chromatography, 8-AHA-cAMP Sepha-rose, and immunoimmobilization chromatography. Ad-ditional data were obtained from 8-N3[
32P]cAMP photo-affinity labeling of R subunits and autophosphorylationof RII subunits in conjunction with SDS-PAGE. Func-tional C subunit levels were obtained by specific activitymeasurements of heptapeptide phosphorylation.
By using 8-N3-[32P]cAMP photoaffinity labeling and
DEAE cellulose chromatography, Spruill et al. (32) havedemonstrated the presence of two distinct regulatorysubunits of PKA in Sertoli cells, corresponding to theRIa and Rlla in our studies. By DEAE cellulose chro-matography, Fakunding and Means (40) also demon-strated the presence of two types of protein kinases inSertoli cell-enriched testes in vivo. However, none ofthese investigators obtained indications of heterogeneity(RIIn vs. RII/?) among the RII subunits.
The present measurements of Sertoli cell PKA sub-units proteins indicate that cAMP stimulation is asso-ciated with a 2- to 3-fold increase in the ratio of R/Csubunits. The C subunit level increased only slightly;therefore, increases in the R subunit account for the ratio
change observed. In other tissues the levels of PKAsubunits have been shown to be subject to regulationduring cell growth, after transformation, and during dif-ferentiation (for review see Ref. 41). In addition, free RIsubunit like that demonstrated in the present study haspreviously been reported in cells of neural origin (42-45), in Chinese hamster ovary cells (46), and in Frienderythroleukemic cells (47). In contrast, a cAMP-depend-ent increase in the R/C ratio in rat hepatocytes inprimary culture has been shown to be due to C down-regulation rather than due to an increase in R (48).
The most distinctive change in PKA during cAMPstimulation of Sertoli cells was the induction of RII/jmRNA and protein. Interestingly, ovarian granulosacells, the female counterpart to testicular Sertoli cells,reveal a similar differential regulation of RII^ by cAMP(12, 49). During cAMP stimulation, the increase in RILjmRNA was approximately 30- to 50-fold within the first24-48 h. Other PKA subunit mRNAs only showed minorincreases after cAMP stimulation.
After cAMP stimulation, the RI/RII subunit ratio wasincreased only modestly, as determined by specific im-munoimmobilization method. In cAMP-stimulated cellsRII0 still represented less than 10% of the total R sub-unit. This may explain why RII^ has not been identifiedin Sertoli cells in previous studies (32, 40). Approxi-mately 75% of the total R subunit was RI«, both underbasal conditions and after cAMP stimulation.
The estimated concentration of RILj in Sertoli cells islow compared to that of RIa but substantial when com-pared with total levels of regulatory subunits in someother tissues or cells, for example rat hepatocytes (50).
In our 8-N3-[32P]cAMP photoaffinity studies of whole
cytoplasmic extracts, we always observed labeled RIa andRIIa. However, RII^ could not readily be detected underbasal conditions. In this case our results are very similarto those obtained by Spruill et al. (32) who detected onlytwo regulatory PKA subunits in cytoplasmic extractsfrom Sertoli cells. However, after fractionation of thecytoplasmic extracts by DEAE cellulose chromatographyor cAMP affinity chromatography, three R subunitscould be photoaffinity labeled. In these studies we dem-onstrated a selective induction of the RII/s protein aftercAMP stimulation. Under basal conditions autophos-phorylation detected only RIIa; however, after stimula-tion with (Bt)2cAMP, similar amounts of autophosphor-ylated RIIa and of RII^ were observed. During the auto-phosphorylation experiment, a smaller proteolyticfragment of the RII subunits with mol wt of approxi-mately 35,000 M:r was observed. The fact that this proteinwas observed only in cells treated with cAMP indicatesthat this may represent a breakdown product of RII^.Another possibility is that cAMP induces a proteasedegrading both RIIa and RII^. These autophosphoryla-

2352 PKA IN RAT SERTOLI CELLS Endo•1991Voll29«No5
tion studies clearly show the selective induction of theRIIp protein, however it does not allow any estimate ofthe relative RIIa to RILj levels, since the endogenousphosphate content of the RII subunits was not con-trolled.
It has been suggested that R subunit induction mayserve to attenuate the cAMP transduction signal byfavoring the inactive PKA holoenzyme (44, 48). For sucha mechanism, an increase in RII^ would not seem verypractical, since this isoform represents only a minorfraction of the total R. Since RIa is the predominantsubunit and is increased about 2-fold by cAMP stimula-tion, such a function would be more appropriate for RIa.Consistent with this hypothesis, much of the RIa subunitappears to be dissociated from the C subunit duringcAMP stimulation.
cAMP stimulation results in an apparent decrease intype I holoenzyme and an apparent increase in type IIholoenzyme. Since quantitative methods are not pres-ently available to adequately separate RII holoenzymeisoforms, it is not possible to determine if the apparentincrease in type II holoenzyme is due to RIIa or RII^.Overexpression of RIIa in transfected NIH3T3 cells re-sults in preferential assembly of PKA type II, even inthe presence of excess free RI (51). Whether this pref-erential association of RII holoenzymes results from invivo differences in R subunit affinities for C subunit ofpreferential assembly of type II holoenzyme via com-partmentalization is not known. Furthermore, the orderof potency of four different analogs is the same as thatobserved for kinase-mediated mechanisms in other cells(52). The mechanism(s) of this autoregulation has notbeen specifically determined, but multiple mechanismsare most likely operative, including altered transcriptionand translation as well as mRNA and protein turnoverrates. The stimulatory effects of cAMP on mRNA forRILj in Sertoli cells appears to involve both transcrip-tional regulation (53) as well as cAMP-mediated mRNAstabilization (53a).
The functional significance of the different isomericforms of R and C in Sertoli cells (or any cell) has stillnot been established. Their discrete cellular and subcel-lular localization (41, 54-56), their specific interactionwith R binding proteins (57-59), and differential regu-lation seen in the present study suggest that the isoformsmay have a role in redistribution of intracellular PKAactivity. Given the various PKA gene products, a greatnumber of PKA holoenzymes could result. It remains tobe determined whether isoforms of PKA holoenzymescontribute to differential regulation of the great diversityof cAMP-mediated responses.
AcknowledgmentsWe gratefully acknowledge the technical assistance of Ms. Guri
Opsahl and Ms. Gladys Josefsen. We thank Ms. Liv St0ttum forassisting in typing the manuscript.
References1. Means AR, Dedman JR, Tash JS, Tindall DJ, van Side M, Welsh
MJ 1980 Regulation of the testis Sertoli cell by follicle-stimulatinghormone. Annu Rev Physiol 42:59-79
2. Beebe SJ, Corbin JD 1986 Cyclic nucleotide-dependent proteinkinases. In: Boyer PD, Krebs EG (eds) The enzymes. AcademicPress, Orlando, vol 17:43-111
3. Hansson V, Trygstad 0, French FS, McLean WS, Smith AA,Tindall DJ, Weddington SC, Petrusz P, Nayfeh SN, Ritzen EM1974 Androgen transport and receptor mechanisms in testis andepididymis. Nature 250:387-391
4. Corbin JD, Keely SL, Park CP 1975 The distribution and dissocia-tion of cyclic adenosine 3':5'-monophosphate-dependent proteinkinases in adipose, cardiac, and other tissues. J Biol Chem 250:218-221
5. Lee DC, Carmichael DF, Krebs EG, McKnight SM 1983 Isolationof a cDNA clone for the type I regulatory subunit of bovine cAMP-dependent protein kinase. Proc Natl Acad Sci USA 80:3608-3612
6. Titani K, Sasagawa T, Ericsson LH, Kumar S, Smith SB, KrebsEG, Walsh K A 1984 Amino acid sequence of the regulatory subunitof bovine type I adenosine cyclic 3'5'-phosphate dependent proteinkinase. Biochemistry 23:4193-4199
7. Clegg CH, Cadd GC, McKnight SM 1988 Genetic characterizationof a brain-specific form of the type I regulatory subunit of cAMP-dependent protein kinase. Proc Natl Acad Sci USA 85:3703-3707
8. Scott JD, Glaccum MB, Zoller MJ, Uhler MD, Helfman DM,McKnight SM, Krebs EG 1987 The molecular cloning of a type IIregulatory subunit of the cAMP-dependent protein kinase from ratskeletal muscle and mouse brain. Proc Natl Acad Sci USA 84:5192-5196
9. Takio K, Smith SB, Krebs EG, Walsh KA, Titani K 1982 Primarystructure of the regulatory subunit of type II cAMP-dependentprotein kinase from bovine cardiac muscle. Proc Natl Acad SciUSA 79:2544-2548
10. Takio K, Smith SB, Krebs EG, Walsh KA, Titani K 1984 Aminoacid sequence of the regulatory subunit of bovine type II adenosinecyclic 3',5'-phosphate dependent protein kinase. Biochemistry23:4200-4206
11. Weldon SL, Mumby MC, Taylor SS 1985 The regulatory subunitof neural cAMP-dependent protein kinase II represents a uniquegene product. J Biol Chem 260:6440-6448
12. Jahnsen T, Hedin L, Kidd VJ, Beattie WG, Lohmann SM, WalterU, Durica J, Schulz TZ, Schiltz E, Browner M, Lawrence CB,Goldman D, Ratoosh SL, Richards JS 1986 Molecular cloning,cDNA structure, and regulation of the regulatory subunit of typeII cAMP-dependent protein kinase from rat ovarian granulosacells. J Biol Chem 261:12352-12361
13. Shoji S, Ericsson LH, Fischer EN, Titani K 1983 Amino acidsequence of the catalytic subunit of bovine type II adenosine cyclic3',5'phosphate-dependent protein kinase. Biochemistry 22:3702-3709
14. Uhler MD, Carmichael DF, Lee DC, Chrivia JC, Krebs EG Mc-Knight, SM 1986 Isolation of cDNA clones coding for the catalyticsubunit of mouse cAMP-dependent protein kinase. Proc Natl AcadSci USA 83:1300-1304
15. Uhler MD, Chrivia JC, McKnight SM 1986 Evidence for a secondisoform of the catalytic subunit of cAMP-dependent protein kinase.J Biol Chem 261:15360-15363
16. Showers, MO Maurer RA 1986 A cloned bovine cDNA encodes analternate form of the catalytic subunit of cAMP-dependent proteinkinase. J Biol Chem 261:16288-16291
17. Chrivia JC, Uhler MD, McKnight SM 1988 Characterization ofgenomic clones coding for the Ca and Cb subunits of mouse cAMP-dependent protein kinase. J Biol Chem 263:5739-5744
18. Beebe SJ, 0yen O, Sandberg M, Fr0ysa A, Hansson V, Jahnsen T1990 Molecular cloning of a unique, tissue-specific protein kinase(Gy) from human testis—representing a third isoform for the

PKA IN RAT SERTOLI CELLS 2353
catalytic subunit of cAMP-dependent protein kinase. Mol Endo-crinol 4:465-475
19. 0yen O, Fr0ysa A, Sandberg M, Eskild W, Joseph D, Hansson V,Jahnsen T 1987 Cellular localization and age-dependent changesin mRNA for cyclic adenosine 3',5'-monophosphate-dependentprotein kinases in rat testis. Biol Reprod 37:947-956
20. Dorrington JH, Roller NF, Fritz IJ 1975 Effects of follicle-stimu-lating hormone on cultures of Sertoli cell preparations. Mol CellEndocrinol 3:57-70
21. 0yen 0, Sandberg M, Eskild W, Levy FO, Knutsen G, Beebe SJ,Hansson V, Jahnsen T 1988 Differential regulation of messengerribonucleic acids for specific subunits of cyclic adenosine3',5'monophosphate (cAMP)-dependent protein kinase in rat Ser-toli cells. Endocrinology 122:2658-2666
22. Maniatis T, Fritsch EF, Sambrook J 1983 Molecular Cloning, aLaboratory Manual. Cold Spring Harbor Laboratory, Cold SpringHarbor
23. Cobb CE, Corbin JD 1988 Purification of cAMP-free and cAMP-bound forms of bovine heart cAMP-dependent protein kinaseholoenzyme. In: Corbin JD, Johnson RA (eds) Methods in Enzy-mology. Academic Press, San Diego, vol 159:202-208
24. Corbin JD, Sugden PH, West L, Flockart DA, Lincoln TM, Mc-Carthy D 1978 Studies on the properties and mode of action of thepurified regulatory subunit of bovine heart adenosine 3',5'-mono-phosphate-dependent protein kinase. J Biol Chem 253:3997-4003
25. Bradford MM 1976 A rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principle ofprotein-dye binding. Anal Biochem 72:248-254
26. Laemmli UK 1970 Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature 227:680-685
27. Towbin H, Staehelin T, Gordon J 1988 Electrophoretic transfer ofproteins from polyacrylamide gels to nitrocellulose sheets: proce-dure and some applications. Proc Natl Acad Sci USA 76:4350-4354
28. Jahnsen T, Lohmann SL, Walter V, Hedin L, Richards JS 1985Purification and characterization of hormone-regulated isoformsof the regulatory subunit of type II cAMP-dependent protein kinasefrom rat ovaries. J Biol Chem 260:15980-15987
29. Lohmann SL, Schwoch G, Reiser G, Port R, Walter U 1983Dibuturyl cAMP treatment of neuroblastoma-glioma hybrid cellsresults in selective increase in cAMP-receptor protein (R-I) asmeasured by monospecific antibodies. EMBO J 2:153-159
30. Robinson-Steiner AM, Beebe SJ, Rannels SR, Corbin JD 1984Microheterogeneity of type II cAMP-dependent protein kinase invarious mammalian species and tissues. J Biol Chem 259:10596-10605
31. Ekanger R, Vintermyr OK, D0skeland SO 1988 The amounts ofrat liver cyclic AMP-dependent protein kinase I and II are differ-entially regulated by diet. Biochem J 256:447-452
32. Spruill WA, Steiner AL, Tres LL, Kierszenbaum AL 1984 Char-acterization of cyclic AMP-binding proteins in rat Sertoli cellsusing a photoaffinity ligand. Mol Cell Biochem 60:147-157
33. Kuo Jf, Greengard P 1969 Cyclic nucleotide-dependent proteinkinases IV. Widespread occurrence of adenosine 3',5' monophos-phate-dependent protein kinase in various tissues and phyla of theanimal kingdom Proc Natl Acad Sci USA 64:1349-1355
34. Montminy MR, Bilezikjian LM 1987 Binding of a nuclear proteinto the cyclic-AMP response element of the somatostatin gene.Nature 328:175-178
35. Robert M, De Philip Kierszenbaum AL 1982 Hormonal regulationof protein synthesis, secretion and phosphorylation in cultured ratSertoli cells. Proc Natl Acad Sci USA 79:6551-6555
36. Rangel-Aldao R, Rosen OM 1977 Effect of cAMP and ATP on thereassociation of phosphorylated and nonphosphorylated subunitsof the cAMP-dependent protein kinase from bovine cardiac muscle.J Biol Chem 252:7140-7145
37. Gettys TW, Vine AJ, Simonds MF, Corbin JD 1988 Activation ofthe particulate low Km phosphodiesterase of adipocytes by addition
of cAMP-dependetn protein kinase. J Biol Chem 263:10359-1036338. Macphee CH, Reifsnyder DH, Moore TA, Lerea KM, Beavo JA
1988 Phosphorylation results in activation of a cAMP phosphodi-esterase in human platelets. J Biol Chem 263:10353-10358
39. Hedin L, McKnight SM, Lifka J, Durica JM, Richards JS 1987Tissue distribution and hormonal regulation of messenger ribonu-cleic acid for regulatory and catalytic subunits of adenosine 3',5'-monophosphate-dependent protein kinases during ovarian follicu-lar development and luteinization in the rat. Endocrinology120:1928-1935
40. Fakunding JL, Means AR 1977 Characterization and follicle-stim-ulating hormone activation of Sertoli cell cyclic AMP-dependentprotein kinases. Endocrinology 101:1358-1368
41. Lohmann SM, Walter U 1984 Regulation of the cellular andsubcellular concentrations and distribution of cyclic nucleotide-dependent protein kinases. In: Greengard P, Robinson GA (eds)Advances in Cs'clic Nucleotide and Protein Phosphorylation Re-search. Raven Press, New York, vol 18:63-117
42. Walter U, Costa M, Breakefield XO, Greengard P 1979 Presenceof free cyclic AMP receptor protein and regulation of its level bycyclic AMP in neuroblastoma-glioma hybrid cells. Proc Natl AcadSci USA 76:3251-3255
43. Prashad N, Rosenberg RN, Wischmeyer B, Ulrich C, Sparkman D1979 Induction of adenosine 3',5'-monophosphate binding proteinsby N6,O2'-dibutyryladenosine 3',5'-monophosphate in mouseneuroblastoma cells. Analysis by two-dimensional gel electropho-resis. Biochemistry 38:2717-2725
44. Liu AY-C, Chan T, Chen KY 1981 Induction of the regulatorysubunit of type I adenosine cyclic 3',5'-monophosphate-dependentprotein kinase in differentiated N-18 mouse neuroblastoma cells.Cancer Res 41:4579-4587
45. Loffler F, Lohmann SM, Walckhoff B, Walter U, Hamprecht B1985 Selective increase of RI subunit of cyclic AMP-dependentprotein kinase iin glia-rich primary cultures upon treatment withdibutyryl cyclic AMP. Brain Res 344:322-328
46. Singh TJ, Roth C, Gottesmann MM, Pastan IH 1981 Character-ization of cyclic AMP-resistant Chinese hamster ovary cell mutantslacking type I protein kinase. J Biol Chem 256:926-932
47. Schwartz DA, Rubin CS 1983 Regulation of cAMP-dependentprotein kinase subunit levels in Friend erythroleukemic cells. JBiol Chem 258:777-784
48. Houge G, Vintermyr OK, D0skeland SO 1990 The expression ofcAMP-dependent protein kinase subunits in primary rat hepato-cyte cultures. Cyclic AMP down-regulates its own effector systemby decreasing the amount of catalytic subunit and increasing themRNAs for the inhibitory (R) subunits of cAMP-dependent pro-tein kinase. Mol Endocrinol 4:481-488
49. Richards JS, Rolfes AI 1980 Hormonal regulation of cyclic AMPbinding to specific receptor proteins in rat ovarian follicles. J BiolChem 255:5481-5489
50. Ekanger R, Vintermyr O, Houge G, Sand T-E, Scott JD, KrebsEG, Eikhom TS, Christoffersen T, 0greid D, D0skeland SO 1989The expression of cAMP-dependent protein kinase subunits isdifferentially regulated during liver regeneration. J Biol Chem264:4374-4382
51. Otten AD, McKnight SM 1989 Overexpression of the type IIregulatory subunit of the cAMP-dependent protein kinase elimi-nates the type I holoenzyme in mouse cells. J Biol Chem 264:20255-20260
52. Beebe SJ, Blackmore PF, Chrisman TD, Corbin JD 1988 Use ofsynergistic pair of site-selective cAMP analogs in intact cells. In:Corbin JD, Johnson RA (eds) Use of Synergistic Pairs of Site-Selective cAMP Analogs in Intact Cells. Academic Press, SanDiego, vol 159:118-139
53. Tasken KA, Knutsen HK, Attramadal H, Tasken K, Jahnsen T,Hansson V, Eskild W 1991 Different mechanisms are involved incAMP-mediated induction of mRNAs for subunits of cAMP-de-pendent protein kinases. Mol Endocrinol 5:21-28

2354 PKA IN RAT SERTOLI CELLS Endo • 1991Voll29-No5
53a.Knutsen HK, Tasken KA, Eskild W, Jahnsen T, Hansson V, CyclicAMP-dependent stabilization of mRNAs for protein kinase A(PKA) subunits in rat Sertoli cells: rapid degradation of mRNAsfor PKA subunits is dependent on ongoing RNA and proteinsynthesis. Endocrinology, in press
54. 0yen 0, Scott JD, McKnight SM, Krebs EG, Hansson V, JahnsenT 1988 A unique mRNA species for a regulatory subunit of cAMP-dependent protein kinase is specifically induced in haploid germcells. FEBS Lett 229:391-394
55. Cadd G, McKnight SM 1989 Distinct patterns of c AMP -dependentprotein kinase gene expression in mouse brain. Neuron 3:71-79
56. De Camilli P, Moretti M, Donini SD, Walter U, Lohmann SM1986 Heterogeneous distribution of the cAMP receptor protein RII
in the nervous system: evidence for its intracellular accumulationon mucrotubules, microtubule-organizing centers, and in the areaof the Golgi complex. J Cell Biol 103:189-203
57. Lohmann SL, De Camilli P, Walter U 1988 Type II cAMP-dependent protein kinase regulatory subunit-binding proteins. In:Corbin JD, Johnson PA (eds) Methods in Enzymology. AcademicPress, San Diego, vol 159:189-203
58. Sarkar D, Erlichman J, Rubin CS 1984 Identification of a Calmod-ulin-binding protein that co-purifies with the regulatory subunit ofbrain protein kinase II. J Biol Chem 259:9840-9846
59. Leiser M, Rubin CS, Erlichman J 1986 Differential binding of theregulatory subunits (RII) of cAMP-dependent protein kinase IIfrom bovine brain and muscle to RH-binding proteins. J Biol Chem261:1904-1908
Ninth International Congress of Endocrinology
The Ninth International Congress of Endocrinology will be held in Nice (Acropolis, Convention Center,Nice, France) from August 30-September 5, 1992. The program will cover the entire field of basic andclinical endocrinology and will include plenary lectures, symposia, meet-the-professors sessions, and posterpresentations. For information, contact:
Ninth International Congress of EndocrinologySOCFI14, Rue Mandar75002 Paris, FrancePhone: 33 (1) 42 33 89 94Fax: 33(1) 40 26 04 44.
Copyright © 2022 FDOKUMEN




















