
Transient coexpression of cytokeratin and vimentin in differentiating rat sertoli cells
10
DEVELOPMENTAL BIOLOGY 117.35-44 (1986) Transient Coexpression of Cytokeratin and Vimentin in Differentiating Rat Sertoli Cells JORMA PARANKO,* MARKKU KALLAJOKI,* LAURI J. PELLINIEMI,? VELI-PEKKA LEHTO,$ AND ISMO VIRTANENI: *Imtitute of’Bicrmedicine, Depa,rtment of Anutomy, and tlaboratory of Electron Microscopy, University of Turku, Kiinamyllynkatu 10, SF- 20520 Turku, Finland, and +Depwtment of Pathology, University of Helsinki, Haartmaninkatu 3, SF-00290 HelsCnki, Finland Received July 5, 1985; accepted in revised f&n January 23, 1986 The expression of cytokeratin and vimentin type intermediate filaments were studied in fetal, postnatal, and adult rat testes. Immunocytochemical observations were correlated with the light and electron microscopic analysis of the developing organs. The Sertoli cell precursors in 15-day-old fetal testes contained both cytokeratin and vimentin. A gradual reorganization of both filaments, accompanied by a decrease of cytokeratin-positivity, was observed toward the end of the fetal period. The simultaneous presence of cytokeratin and vimentin in the same cells was shown by double immunofluorescence of newborn testes and the primary culture of dissociated testicular cells. In postnatal Sertoli cells, cytokeratin-positivity continued to decrease and disappeared by the age of 14 days. The increase in vimentin content and the appearance of axially oriented vimentin filaments coincided with the acquisition of the columnar shape of the Sertoli cells. The presence of cytokeratin and vimentin in fetal and newborn testes, and only vimentin in the adult testes was confirmed by immunoblotting. The present results suggest that major qualitative changes in the expression of intermediate filament proteins can take place during the embryonic development. The expression of cytokeratin in developing Sertoli cells, although only transient, supports the epithelial origin of these cells and can be applied as a marker for embryonic and early postnatal Sertoli cells. o 1986 Academic P~CSS, 1,~. INTRODUCTION The embryonic testis differentiates from the indiffer- ent gonad by formation of epithelial testicular cords, the future seminiferous tubuli, from unorganized cells on the anterior surface of the mesonephros (Jost et al., 1973; Pelliniemi, 19’75; Pelliniemi et al., 1984). Different developmental origins have been suggested for the Ser- toli cells (Wartenberg, 1978), and the interpretations of the results in various mammalian species-e.g., mouse (Upadhyay et al., 1981), sheep (Zamboni et al., 1981), pig (Pelliniemi, 1975), rabbit and human (Wartenberg, 1981), rat (Jost et al., 1981), and horse (Merchant-Larios, 1979)-are still controversial. Many biochemical and immunocytochemical studies have demonstrated that the cell type-specific expression of intermediate filaments can be used to reveal the de- velopmental origin and differentational stage of cell lin- eages both in viva and in vitro (Virtanen et al., 1981; Franke et ab, 1982; Osborn et al., 1982). Cytokeratins compose a heterogeneous group of polypeptides, present in different combinations in various epithelial cells (Moll et al., 1982; Franke et ah, 1984). Mesenchymal cells, on the other hand, express only vimentin-type intermediate filaments (Franke, 1978). The simultaneous presence of cytokeratin and vimentin filaments is rare in normal adult tissues (Franke et al., 1982; Osborn et ah, 1982), but is often seen in cultured epithelial cells (Franke et al., 19’78;Virtanen et ab, 1981; Franke et al., 1982, 1984). Recent immunocytochemical studies have reported that Sertoli cells from the testes of the adult rat (Franke et ah, 1979a) and human (Ramaekers et al, 1983), as well as the developing and adult pig (van Vorstenbosch et al., 1984a) contain only vimentin, which would support their mesenchymal origin. In the present study we have analyzed immunocyto- chemically cytokeratin and vimentin in the fetal, post- natal, and adult rat testes. Our results with several dif- ferent antibodies indicate that in the rat testis the Ser- toli cells contain both cytokeratin and vimentin during the fetal and early postnatal periods, but later only vi- mentin. These results speak in favor of an epithelial or- igin of Sertoli cells. MATERIALS AND METHODS Tissues Fetal, newborn, 5-, 8-, 9-, lo-, 14-, and 20-day-old post- natal and adult testes were obtained from Sprague- Dawley rats. The fetal ages studied were 15, 16, 18, and 20 days of pregnancy. The day after copulation was des- ignated as Day 0 of pregnancy. Primary Cell Culture For primary culture of newborn rat testicular cells, the testes were removed and cleaned under the dissection microscope. Thereafter, the contents of the testes were 35 0012-1606/86 $3.00 Copyright G 19% by Academic Press, Inc. All rights of reproduction in any form reserved.
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Transient coexpression of cytokeratin and vimentin in differentiating rat sertoli cells

DEVELOPMENTAL BIOLOGY 117.35-44 (1986)
Transient Coexpression of Cytokeratin and Vimentin in Differentiating Rat Sertoli Cells
JORMA PARANKO,* MARKKU KALLAJOKI,* LAURI J. PELLINIEMI,? VELI-PEKKA LEHTO,$ AND ISMO VIRTANENI:
*Imtitute of’Bicrmedicine, Depa,rtment of Anutomy, and tlaboratory of Electron Microscopy, University of Turku, Kiinamyllynkatu 10, SF-
20520 Turku, Finland, and +Depwtment of Pathology, University of Helsinki, Haartmaninkatu 3, SF-00290 HelsCnki, Finland
Received July 5, 1985; accepted in revised f&n January 23, 1986
The expression of cytokeratin and vimentin type intermediate filaments were studied in fetal, postnatal, and adult rat testes. Immunocytochemical observations were correlated with the light and electron microscopic analysis of the
developing organs. The Sertoli cell precursors in 15-day-old fetal testes contained both cytokeratin and vimentin. A
gradual reorganization of both filaments, accompanied by a decrease of cytokeratin-positivity, was observed toward the end of the fetal period. The simultaneous presence of cytokeratin and vimentin in the same cells was shown by double
immunofluorescence of newborn testes and the primary culture of dissociated testicular cells. In postnatal Sertoli cells, cytokeratin-positivity continued to decrease and disappeared by the age of 14 days. The increase in vimentin content
and the appearance of axially oriented vimentin filaments coincided with the acquisition of the columnar shape of the Sertoli cells. The presence of cytokeratin and vimentin in fetal and newborn testes, and only vimentin in the adult
testes was confirmed by immunoblotting. The present results suggest that major qualitative changes in the expression of intermediate filament proteins can take place during the embryonic development. The expression of cytokeratin in
developing Sertoli cells, although only transient, supports the epithelial origin of these cells and can be applied as a
marker for embryonic and early postnatal Sertoli cells. o 1986 Academic P~CSS, 1,~.
INTRODUCTION
The embryonic testis differentiates from the indiffer- ent gonad by formation of epithelial testicular cords, the future seminiferous tubuli, from unorganized cells on the anterior surface of the mesonephros (Jost et al., 1973; Pelliniemi, 19’75; Pelliniemi et al., 1984). Different developmental origins have been suggested for the Ser- toli cells (Wartenberg, 1978), and the interpretations of the results in various mammalian species-e.g., mouse (Upadhyay et al., 1981), sheep (Zamboni et al., 1981), pig (Pelliniemi, 1975), rabbit and human (Wartenberg, 1981), rat (Jost et al., 1981), and horse (Merchant-Larios, 1979)-are still controversial.
Many biochemical and immunocytochemical studies have demonstrated that the cell type-specific expression of intermediate filaments can be used to reveal the de- velopmental origin and differentational stage of cell lin- eages both in viva and in vitro (Virtanen et al., 1981; Franke et ab, 1982; Osborn et al., 1982). Cytokeratins compose a heterogeneous group of polypeptides, present in different combinations in various epithelial cells (Moll et al., 1982; Franke et ah, 1984). Mesenchymal cells, on the other hand, express only vimentin-type intermediate filaments (Franke, 1978). The simultaneous presence of cytokeratin and vimentin filaments is rare in normal adult tissues (Franke et al., 1982; Osborn et ah, 1982), but is often seen in cultured epithelial cells (Franke et al., 19’78; Virtanen et ab, 1981; Franke et al., 1982, 1984).
Recent immunocytochemical studies have reported that Sertoli cells from the testes of the adult rat (Franke et ah, 1979a) and human (Ramaekers et al, 1983), as well as the developing and adult pig (van Vorstenbosch et al., 1984a) contain only vimentin, which would support their mesenchymal origin.
In the present study we have analyzed immunocyto- chemically cytokeratin and vimentin in the fetal, post- natal, and adult rat testes. Our results with several dif- ferent antibodies indicate that in the rat testis the Ser- toli cells contain both cytokeratin and vimentin during the fetal and early postnatal periods, but later only vi- mentin. These results speak in favor of an epithelial or- igin of Sertoli cells.
MATERIALS AND METHODS
Tissues
Fetal, newborn, 5-, 8-, 9-, lo-, 14-, and 20-day-old post- natal and adult testes were obtained from Sprague- Dawley rats. The fetal ages studied were 15, 16, 18, and 20 days of pregnancy. The day after copulation was des- ignated as Day 0 of pregnancy.
Primary Cell Culture
For primary culture of newborn rat testicular cells, the testes were removed and cleaned under the dissection microscope. Thereafter, the contents of the testes were
35 0012-1606/86 $3.00 Copyright G 19% by Academic Press, Inc. All rights of reproduction in any form reserved.

36 DEVELOPMENTAL BIOLOGY
briefly trypsinized (0.25%, 10 min at 3’7°C) and plated on glass coverslips. The cells were grown 2 days in RPM1 1640 medium supplemented with 10% fetal calf serum (Flow, Irvine, Scotland) and antibiotics. After the cul- ture the coverslips were fixed in methanol at -20°C for 10 min.
Antibodies
Monoclonal antibodies to vimentin. Vimentin was iso- lated from detergent-resistant cytoskeletons of human fibroblasts by preparative gel electrophoresis as de- scribed earlier (Virtanen et al., 1981). Standard tech- niques (Kohler et ah, 1976) were used to raise the mouse hybridoma antibodies as described earlier in detail (Holthiifer et al, 1983). Antibody-producing hybrids were screened by a solid phase assay with purified vimentin, as well as by indirect immunofluorescence microscopy of various human and rat cells and tissues. For the pres- ent study one of the antibody-producing clones was used (24 BA6). This antibody reacts exclusively with mes- enchymal cells in frozen sections of human and rat tis- sues. Indirect immunofluorescence of cultures of human fibroblasts (from a local source), mouse 3T3 cells, and normal rat kidney (NRK) fibroblasts (from American Type Culture Collection, Rockville, Md.), revealed a typ- ical fibrillar pattern in the cytoplasm (Figs. la, b). Im- munoblotting of detergent-resistant cytoskeletons of cultured NRK fibroblasts revealed only a polypeptide with lkf, 57,000 (Fig. lc).
Polyclmal antibody to vimentin. Guinea pig antibodies to vimentin were raised against vimentin purified from mouse 3T3 cells and characterized as described earlier (Franke et al., 19’79b). These antibodies were a kind gift from Dr. Werner W. Franke (German Cancer Research Center, Heidelberg, FRG).
Monoclonal cytokeratin antibodies. RGE 53 antibodies were raised against keratin polypeptides isolated from HeLa cells and human plantar callus (Ramaekers et al., 1983). PKK 1 and PKK 3 antibodies were raised against cytokeratins isolated from a pig kidney epithelial cell line (LLCK-PK 1) as described earlier (Holthofer et al., 1984; Virtanen et ah, 1985). RGE 53 (Ramaekers et al., 1983), PKK 1, and PKK 3 (Holthiifer et aZ., 1983, 1984; Miettinen et al., 1984; Virtanen et al., 1985) antibodies have given a positive reaction only in various simple epithelial cells and in carcinomas derived from them. Immunoblotting of various cultured human epithelial cells with PKK 1 antibodies have shown a strong reaction with M, 55,000, ikf, 45,000, and M, 40,000 cytokeratins corresponding to cytokeratins 8,18, and 19 according to Moll et al. (1982) (Virtanen et al., 1985; Miettinen et ab, 1985). Both PKK 3 and RGE 53 antibodies reveal in im-
1
1
I
VOLUME 117. 1986
1 2
681
431
C
FIG. 1. (a) Normal rat kidney (NRK) tibroblasts reacted with the
nonoclonal vimentin antibody 24 BA6. Note the fibrillar cytoplasmic ocalization by immunofluorescence in all cells. (b) The same field in
chase-contrast. (c) Electrophoretically separated NRK fibroblast cy- .oskeletal polypeptides transferred onto nitrocellulose sheet and
stained for proteins (lane 1). Immunoreaction with monoclonal vi- nentin antibody (lane 2) reveals only a polypeptide with M, 57,000.
Molecular weight standards are indicated on the left side (M, X lOma). a, b) X700.
nunoblotting of human epithelial cells a single cyto- teratin polypeptide with M, 45,000 corresponding to cy- ;okeratin 18 (Moll et al., 1982; Ramaekers et al., 1983; Virtanen et ah, 1985). In rat hepatoma cell line (HTC, Thompson et al., 1966) PKK 1, PKK 3, and RGE 53 cy- ;okeratin antibodies revealed a cytoplasmic, wavy fi- n-illar system different from that obtained with vimen- in antibodies. In immunoblotting PKK 1 antibody re- Tealed polypeptides of M, 55,000 and 45,000, whereas both XGE 53 and PKK 3 antibodies only revealed an M, 45,000 lolypeptide (Fig. 2). Monoclonal vimentin antibodies, on ;he other hand, revealed a different prominent M, 57,000 lolypeptide (Fig. 2).
Polyclonal antibody to laminin. Polyclonal antibody *aised against laminin purified from EHS sarcoma Timpl et ah, 1979) was a kind gift from Dr. Jean-Michel ?oidart (University of Liege, Belgium).

PARANK~ ET AL. Cytokerati72 and Vimentirc in Sertoli Cells 37
1 2 3 4
FIG. 2. Electrophoretically separated polypeptides of rat hepatoma
cell line (HTC) cells transferred onto nitrocellulose and stained for proteins (lane l), or immunoreacted with cytokeratin PKK 1 (lane 2),
PKK 3 (lane 3), and monoclonal vimentin (lane 4) antihodies. PKK 1 antibody reveals two polypeptides (M, 55,000 and 45,000), PKK 3 an-
tihodg one ,I+‘, 45,000 polypeptide, and vimentin antihody a distinct M, 57,000 polypeptide.
17nnLu7lcj~uorescence Microscopy
Testes were frozen in liquid nitrogen immediately af- ter removal. Cryostat sections (8 pm) were air dried for a few minutes, and fixed in acetone at -20°C for 10 min, and allowed to air dry. The sections were washed in NaCl-P buffer (140 mM NaCl, 10 mM sodium phosphate, pH 7.2). The first antibody and a fluorescein isothiocy- anate (FITC)-conjugated goat anti-mouse IgG (Hybri- tech, Inc., San Diego, Calif.) were applied in sequence on the sections with standard washes. For double im- munoreactions, the sections were first reacted with guinea pig vimentin antibody (1:80), and FITC-rabbit anti-guinea pig antiserum (Cappel Laboratories, Coch- ranville, Pa.). For the laminin detection FITC-coupled goat anti-rabbit IgG (Cappel) was used. The second re- action was PKK 1 cytokeratin or monoclonal vimentin antibody followed by tetramethylrhodamine isothiocy- anate (TRITC)-coupled sheep anti-mouse IgG antiserum (Cappel). The sections were washed thoroughly in NaCl- P buffer, embedded in glycerol-Verona1 buffer (l:l, pH 8.4), and examined under a Leitz Dialux or Zeiss Uni-
versa1 microscope equipped with an epi-illuminator and filters for FITC and TRITC fluorescence.
Polyacrylamide Gel Electrophoresis and Immunoblotting
Samples from 16-day-old fetal and newborn rats were isolated for gel electrophoresis from the mesonephric tissue and the adjoining rete testis under a transillu- minated stereo microscope. Comparable amount of iso- lated seminiferous tubuli from adult rats were also used. These specimens, as well as cultures of HTC and NRK cells for control purposes, were first extracted in 0.5% Triton X-100 (BDH Chemicals, Poole, England) in 50 mM Tris-HCl, pH 7.4 supplemented with protease in- hibitors (1 mM phenymethyl sulfonylfluoride, 1 mM N- p-tosyl-L-lysine chlormethyl ketone; Sigma Chemical Co., St. Louis, MO.) at 0°C for 30 min, and then boiled in electrophoresis sample buffer (Laemmli, 1970). Poly- acrylamide gel electrophoresis in the presence of sodium dodecyl sulphate (SDS) was performed according to Laemmli (1970) using 8% slab gels. The transfer of pro- teins onto nitrocellulose sheets was performed accor- ding to Towbin et al. (1979). The proteins were stained with 0.1% Amido black. To identify the antigens on the nitrocellulose sheets, they were incubated overnight at 4°C in NaCl-P buffer containing 2% bovine serum al- bumin (BSA) and 0.2% sodium azide. The sheets were exposed to monoclonal antibodies (culture supernatant diluted 1:lO) in NaCl-P buffer supplemented with 2% BSA and 0.2% Triton X-100 for 1 hr. The sheets were then washed in NaCl-P, and reacted with peroxidase- conjugated rabbit anti-mouse immunoglobulins (Dako- patts, Copenhagen, Denmark; 1:lOO dilution) in the same buffer as above. After washes, the peroxidase re- action was made in 0.25 mg/ml 3,3’-diamino-benzidine- 4 HCl, 0.006% H202 in 0.025 M Tris-HCl, pH 7.4. The reaction was terminated with 0.2% sodium azide in NaCl-P.
Light and Electron Microscopy
For light microscopy, lo-day-old postnatal and adult rat testes were fixed by perfusion through the thoracic aorta with 5% glutaraldehyde (Merck, Darmstadt, FRG) in 0.16 mole/liter s-collidine-HCl buffer, pH 7.4, and postfixed with potassium ferrocyanide-osmium fixative (Karnovsky, 1971). Testes of 15-day-old fetuses were fixed by immersion.
For electron microscopy, 15-day-old fetal testes were separated from the mesonephric tissue and cut into thin slices. Tissues were incubated in 0.5% Triton X-100 sup- plemented with protease inhibitors as was described in the previous section. After washes in NaCl-P buffer, the

38 DEVELOPMENTAL BIOLOGY VOLUME 117, 1986
tissues were fixed with 5% glutaraldehyde, postfixed with osmium tetroxide, and processed for electron microscopy as previously described (Paranko et al., 1983).
RESULTS
The fetal testicular cords appeared at the age of 15 days, consisting of developing Sertoli cells and sper- matogonia organized as stratified cuboidal epithelium surrounded by a basement membrane (Fig. 3a). Most of
the spermatogonia with dark cytoplasm, prominent spherical nucleus, and globular nucleoli were found in the central region of the cords. The Sertoli cells with pale cytoplasm and ovoid or pleomorphic nucleus were located mainly along the basement membrane (Fig. 3a). The postnatal Sertoli cells became polarized and differ- entiated into tall columnar cells. The first meiotic sper- matocytes were seen by the age of 10 days. The adult testis was characterized by complete spermatogenesis
FIG. 3. Morphological changes from fetal to adult rat Sertoli cells (S). (a) Sertoli cells of 15-day-old fetal rat testicular cords (C) are found as pale cells mainly along the basement membrane. Spermatogonia (G) are characterized by dark cytoplasm. (b) In adults, Sertoli cells have acquired a tall columnar (arrow) shape. (c and inset) Electron micrographs of a detergent-extracted 15-day-old fetal rat testis reveals an
accumulation of intermediate filaments (F) in the basal cytoplasm of a Sertoli cell. D, lamina densa; I, interstitium. (a, b) X300; (c) X18,000; inset X48,000.

FIG. 4. Changes in the cytokeratins from fetal (a, c) to newborn (d), and postnatal (e-h) rat Scrtoli cells revealed by immunofluorescene monoclonal antibodies. (a) PKK 1 cytokeratin antibody gives an intense reaction in the basal (arrowheads) and an occasional lateral rea
(arrows) in the cytoplasm of the Sertoli cells in 15day-old fetuses. (b) Double immunofluorescence with laminin antibody reveals the base membrane (B) of the testicular cords and mesonephric tubuli (T) which were cytokeratin-positive in panel a. (c) An intense basal (arrowh’ and in some cells, a weak lateral (arrows) cytokeratin-positivity (PKK 1) is seen in a 16-day-old fetus. (d) Rete cells (R) of a newborn rat t
show an intense cytokeratin-positivity (PKK 1). Positive basal reaction in the Sertoli cells is indicated by arrowheads. (e-h) A series of pit of postnatal rat testes reveals a gradual decrease in cytokeratin-positivity (arrowheads) with PKK 1 (e) and RGE 53 (f) antibodies in a 5- old testis, and with PKK l-antibody (g) in a lo-day-old testis. (h) Cytokeratin is no longer present in a 14-day-old postnatal testis. C, testi
cords; I, interstitium, a, b X250; c X300; d-h X220.
with
ction ment eads)
;estis
tures -day- cular

40 DEVELOPMENTAL BIOLOGY VOLUME 117, 1986
and the presence of Sertoli cells with their typical large nucleolus (Fig. 3b). Electron microscopy of detergent extracted 15-day-old fetal gonads revealed an irregular meshwork of intermediate filaments at the basal aspect of the Sertoli cells (Fig. 3~).
Cytokeratin in Fetal and Postnatal Testis
All three cytokeratin antibodies used in this study (PKK 1, PKK 3, and RGE 53) gave a distinct basal im-
munoreaction in the fetal Sertoli cells (Figs. 4a and c). This reaction continued in some cells as a weak lateral extension, which was positive with PKK 1 (Figs. 4a and c), but negative with PKK 3 and RGE 53 antibodies. A decrease in the lateral immunoreaction was observed during the fetal development (Fig. 4d). In each case the cytokeratin-positivity was identified in the Sertoli cells by the antilaminin immunofluorescence (Fig. 4b) and phase contrast microscopy. The cells of the mesonephric
FIG. 5. Changes in the distribution of vimentin intermediate filaments from fetal (a), to postnatal (b), and adult (c) rat Sertoli cells (S) as revealed by monoclonal vimentin antibody. (a) Sertoli cells of 15.day-old fetal rat testicular cords (C) contain mainly basal accumulations of vimentin. (b) In a g-day-old postnatal testis baso-apically oriented accumulations of vimentin (arrows) are visible, and in the adult testis (c)
bundles of vimentin filaments run from the basal cytoplasm towards the lumen (L) of the seminiferous tubule. A series of a phase contrast (d) and double immunofluorescence (e, f) micrographs of a newborn rat testis. Immunoreaction with guinea pig polyclonal vimentin antibody (e), and monoclonal cytokeratin PKK 1 antibody (f) are both localized in Sertoli cells (S) of the testicular cords (C). Interstitial tissue (I) is also positive for vimentin in panels a, b, c, and e. (a-c) X210; (d-f) X400.
PARANKO ET AL. Cutokeratin and Vimentin in Sertoli Cells 41
tubuli (Figs. 4a and b) and rete testis (Fig. 4d) were cytokeratin-positive throughout the study.
In a newborn rat testis mainly a basal immunoreaction was observed (Fig. 4d), and as shown in Figs. 4e-g, a gradual decrease in the cytokeratin-positivity took place in the developing postnatal rat Sertoli cells. In a lo-day- old testis a faint basal immunoreaction was still detected (Fig. 4g), but was no longer seen in a 14-day-old testis (Fig. 4h). Similarly, the Sertoli cells of adult testes were always negative with all antibodies used. Cytokeratin- positivity gradually increased in the surface epithelium of developing testis.
Vimentin in Fetal and Postnatal Testis
The surface epithelium and the interstitium (Figs. 5a- c) of the testis, as well as the endothelium of the blood vessels, gave a positive reaction for vimentin at all ages. The cells of the rete testis and mesonephric tubuli were devoid of vimentin. The Sertoli cells at the fetal age of 15 days contained vimentin (Fig. 5a) with similar basal distribution as cytokeratin at the same age. The vimen- tin-positivity of the apical and lateral aspects of Sertoli cells increased toward the end of the fetal period (Fig. 5b) until sexual maturity. In the adult rat Sertoli cells, vimentin was localized in bundles of cytoplasmic fila- ments extending from the aggregate in the basal cyto- plasm toward the lumen of the seminiferous tubuli (Fig. 5~). Double immunofluorescence of frozen sections of a newborn rat testis (Fig. 5d) with polyclonal guinea pig vimentin (Fig. 5e) and PKK 1 cytokeratin (Fig. 5f) an- tibodies gave a positive reaction with both antibodies in the Sertoli cells, as well as an additional strong inter-
stitial reaction with vimentin antibodies. In the im- munocytochemical controls the intensity of the reactions followed the dilution of the primary antibodies and the antibody omission controls were negative.
Primary Cell Culture of Newborn Rut Testicular ,Cells
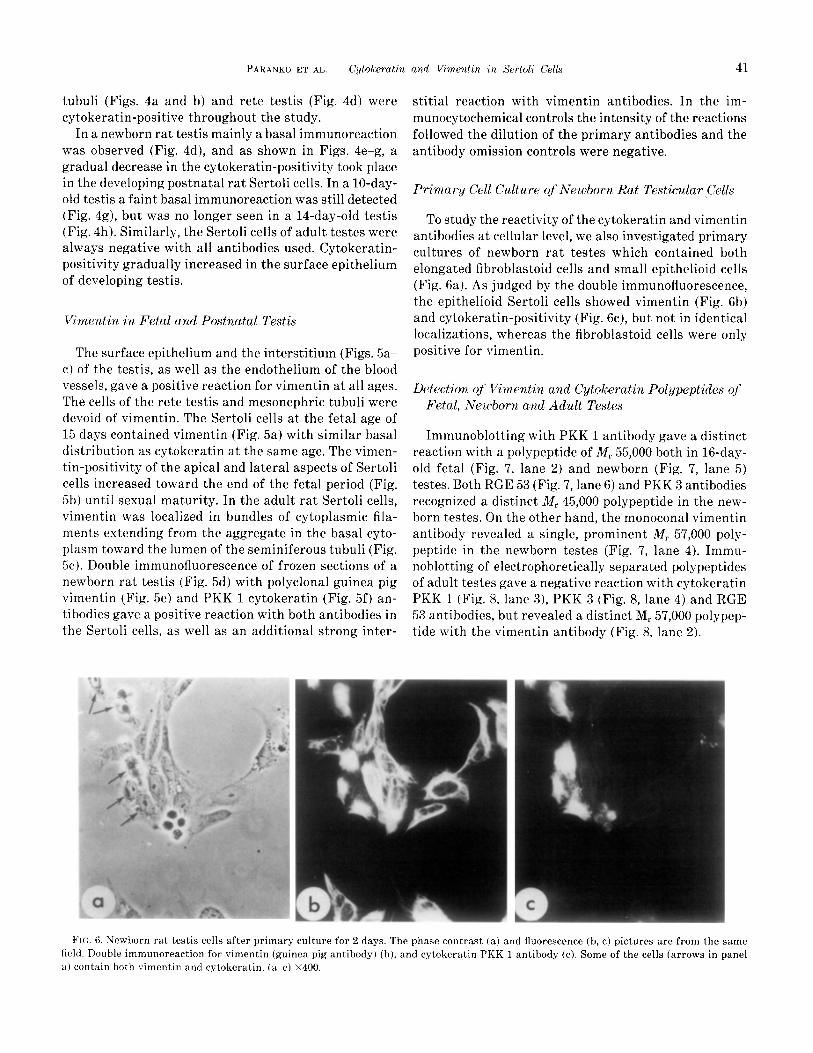
To study the reactivity of the cytokeratin and vimentin antibodies at cellular level, we also investigated primary cultures of newborn rat testes which contained both elongated fibroblastoid cells and small epithelioid cells (Fig. 6a). As judged by the double immunofluorescence, the epithelioid Sertoli cells showed vimentin (Fig. 6b) and cytokeratin-positivity (Fig. 6c), but not in identical localizations, whereas the fibroblastoid cells were only positive for vimentin.
Detection of Vimentin and Cytokeratin Polypeptides sf Fetal, Newborn and Adult Testes
Immunoblotting with PKK 1 antibody gave a distinct reaction with a polypeptide of M,. 55,000 both in 16-day- old fetal (Fig. 7, lane 2) and newborn (Fig. 7, lane 5) testes. Both RGE 53 (Fig. 7, lane 6) and PKK 3 antibodies recognized a distinct h!, 45,000 polypeptide in the new- born testes. On the other hand, the monoconal vimentin antibody revealed a single, prominent M, 57,000 poly- peptide in the newborn testes (Fig. 7, lane 4). Immu- noblotting of electrophoretically separated polypeptides of adult testes gave a negative reaction with cytokeratin PKK 1 (Fig. 8, lane 3), PKK 3 (Fig. 8, lane 4) and RGE 53 antibodies, but revealed a distinct M, 57,000 polypep- tide with the vimentin antibody (Fig. 8, lane 2).
FIG. 6. Newborn rat testis cells after primary culture for 2 days. The phase contrast (a) and fluorescence (b, c) pictures are from the same field. Double immunoreaction for vimentin (guinea pig antibody) (b), and cytokeratin PKK 1 antibody (c). Some of the cells (arrows in panel
a) contain both vimentin and cytokeratin. (a-c) x400.

42 DEVELOPMENTAL BIOLOGY VOLUME 117, 1986
941
68=
43-
1 2 3 4 5 6
k
FIG. 7. Electrophoretically separated polypeptides of 16-day-old fetal (lanes 1 and 2) and newborn (lanes 3-6) rat testis transferred onto
nitrocellulose and stained for proteins (lanes 1 and 3), and immuno-
reacted with cytokeratin PKK 1 (lanes 2 and 5), monoclonal vimentin (lane 4), or cytokeratin RGE 53 (lane 6) antibodies. Similar polypeptides
(M, 55,000) are recognized by PKK 1 cytokeratin antibody in the fetal (lane 2) and newborn (lane 5) testis, whereas RGE 53 cytokeratin an-
tibody recognizes a distinct (M, 45,000) poiypeptide (lane 6). In the newborn testis a prominent (M, 57,000) polypeptide is recognized by
the vimentin antibody (lane 4).
DISCUSSION
The primitive gonadal cords in male rats appear on Day 13 of fetal development (Jost et al., 1981; Paranko et al., 1983). The lack of fibronectin and the synthesis of basement membrane components are the first signs of their epithelial nature (Paranko et ah, 1983: Pelliniemi et al., 1984, 1985). Earlier immunocytochemical studies of mammalian testes have indicated that adult Sertoli cells of various species contain only vimentin-type in- termediate filaments (Franke et ab, 19’79a; Ramaekers et al., 1983; van Vorstenbosch et al., 1984a). This could be in agreement with their mesenchymal origin as sug- gested by Gropp and Ohno (1966). In the present study we were, however, able to show cytokeratin and vimen- tin-positivity in the Sertoli cells of fetal and early post- natal rat testes. We conclude that the early presence of cytokeratin suggests the epithelial origin of the Sertoli cells. Whether they originate from the coelomic surface
epithelium or the mesonephric epithelial structures, or both (Jost et al, 1973; Pelliniemi, 1975; Upadhyay et ah, 1981; Zamboni et ab, 1981, Wartenberg, 1983), remains to be confirmed. The observed basal accumulation of in- termediate filaments and microfilaments (van Vorsten- bosch et ab, 198413; Pelliniemi et ah, 1984) in the fetal Sertoli cells may have a role in the formation of testic- ular cords and polarization of the Sertoli cells.
The postnatal disappearance of cytokeratin is in agreement with the modified epithelial nature of Sertoli cells; they lack desmosomes (Franke et ah, 1979a) and possess specialized Sertoli-Sertoli cell junctions (Dym, 1977). The continuous cytokeratin-positivity and vimen- tin-negativity of the mesonephric tubuli and the rete testis cords, on the other hand, suggest a common epi- thelial origin for their cells. Cytokeratin-positivity of the rete cells seems to be a common feature of several mammalian species (Ramaekers et ab, 1983; van Vor- stenbosch et al,, 984a) and is in agreement with the true epithelial nature of these cells. Functionally it has been suggested that the rete cells would secrete a meiosis- inducing factor, whereas the Sertoli cells would secrete
1 2 34
FIG. 8. Electrophoretically separated polypeptides of isolated adult rat seminiferous tubuli transferred onto nitrocellulose sheet and stained for proteins (lane l), or immunoreacted with monoclonal vi- mentin (lane 2), cytokeratin PKK 1 (lane 3), and PKK 3 (lane 4) an-
tibodies. Vimentin (lane 2), but not cytokeratin (lanes 3 and 4), is present in the cells of seminiferous tubuli.

PARANKO ET AL. Cytokeratin and Vimentin in Sertoli Cells 43
a meiosis-inhibiting factor (Grinsted et al., 1979; War- tenberg, 1983).
The disappearance of cytokeratin and the spatial re- organization of vimentin were observed in the Sertoli cells during the first two postnatal weeks, at the time when the major maturational changes take place in the developing testis. At the age of 15-21 days, the Sertoli cells of the rat testis stop proliferation (Clermont and Perey, 1957; Orth, 1982), the blood-testis barrier be- comes functional (Gilula et al, 1976), and the first meiotic spermatocytes appear (Clermont and Perey, 1957; Berg- mann and Dierichs, 1983). Another temporally matching event with the postnatal disappearance of cytokeratin is the pubertal rise in the tissue concentration of an- drogens after a minimum at the age of two weeks (Ta- panainen, et al, 1984). The hormonal control of the in- termediate filaments is, however, still poorly understood. In culture the organization of intermediate filaments can be affected by growth factors (Keski-Oja et ab, 1981), FSH (Spruill et al., 1983a), and calcium (Spruill et ab, 198313). The observed changes in the intermediate fila- ments of Sertoli cells suggest that there may be impor- tant relationships between the maturation of the testis and the regulation of the cytoplasmic filament systems.
The simultaneous presence of cytokeratins and vi- mentin, as shown in the present study in fetal and early postnatal rat Sertoli cells, is a rare phenomenon in nor- mal epithelial cells, but is found in cultured epi- thelial cells (Franke et al., 197913; Virtanen et al., 1981). Both cytokeratin and vimentin have also been detected in the parietal endodermal cells of early mouse embryos (Lane et ah, 1983; Lehtonen et al, 1983), and in the col- lecting ducts of the developing human kidney (HolthGfer et al., 1984). After malignant change, some carcinomas derived from simple epithelia may also start to express vimentin in addition to cytokeratin (Holthiifer et al.,
1983; Miettinen et al., 1984). Our findings of the qualitative and organizational
changes in the intermediate filaments of developing Sertoli cells suggest that during the embryonic cytodif- ferentiation the cells may modulate the expression of their intermediate filaments. The cytokeratin-positivity of the fetal rat Sertoli cells, although transient, is suggestive for the epithelial origin of these cells. Fur- thermore, the present results indicate that the cytoker- atin- and vimentin-positivity is a useful marker for fetal and early postnatal rat Sertoli cells both in situ and in cell culture.
We thank Mrs. Raija Anderskn, Mrs. Sirpa From, Ms. Pipsa Kai-
painen, Mrs. Raili Taavela, and Mr. Lars-Henrik Wikgren for expert technical assistance, and Mrs. Marja Huhtinen for secretarial assis- tance. This study was supported finacially by the Sigrid Ju&lius Foun- dation, the Medical Research Council of the Academy of Finland, and
the University of Helsinki, Finland.
REFERENCES
BERGMANN, M., and DIERICHS, R. (1983). Postnatal formation of the
blood-testis barrier in the rat with special reference to the initiation
of meiosis. Anat. Embryol. 168, 269-275. CLERMONT, Y., and PEREY, B. (1957). Quantitative study of the cell
population of the seminiferous tubules in immature rats. Am. J.
Anat. 100; 241-268. DYM, M. (1977). The role of the Sertoli cell in spermatogenesis. In
“Male Reproductive System” (R. D. YATES, and M. GORDON, eds.),
pp. 155-169. Masson, St. Paul, Minn.
FRANKE, W. W., SCHMID, E. OSBORN, M., and WEBER, K. (1978). Different intermediate-sized filaments distinguished by immunofluorescence
microscopy. Proc. Natl. Acad. Sci. USA 75,5034-5038. FRANKE, W. W., GRUND, C., and SCHMID, E. (1979a). Intermediate-
sized filaments present in Sertoli cells are of the vimentin type. Eur.
J. Cell Biol. 19,269-275.
FRANKE, W. W., SCHMID, E., WINTER, S., OSBORN, M., and WEBER, K. (1979b). Widespread occurrence of intermediate-sized filaments of the vimentin-type in cultured cells from diverse vertebrates. Exp
Cell Res. 123, 25-46.
FRANKE, W. W., SCHMID, E., SCHILLER, D. L., WINTER, S., JARASCH,
E. D., MOLL, R., DENK, H., JACKSON, B. W., and ILLMENSEE, K. (1982). Differentiation-related expression of proteins of intermediate-size
filaments in tissues and cultured cells. Cold .!!e?/g Harbor 5’ym.n C&ant. Biol. 46,431-453.
FRANKE, W. W., SHILLER, D. L., HATZFELD, M., MAGIN, T. M., JORCANO,
J. L., MITTNACHT, S., SCHMID, E.. COHLBERG, J. A., and QIJINLAN,
R. A. (1984). Cytokeratins: Complex formation, biosynthesis, and interactions with desmosomes. In “Cancer Cells. The Transformed
Phenotype” (A. J. Levine, G. F. V. Woude, W. C. Topp, and J. D. Watson, eds,), pp. 177-190. Cold Spring Harbor Laboratory, Cold
Spring Harbor, N.Y.
GILULA, N. B., FAWCETT, D. W., and AOKI, A. (1976). The Sertoli cell occluding junctions and gap junctions in mature and developing mammalian testis. Dew. Biol. 50, 142-168.
GRINSTED, J., BYSKOV, A. G., and ANDREASEN, M. P. (1979). Induction
of meiosis in fetal mouse testis in vitro by rete testis tissue from pubertal mice and bulls. J. Reprod. Fert. 56, 653-656.
GROPP, A., and OHNO, S. (1966). The presence of a common embryonic blastema for ovarian and testicular parenchymal (follicular, inter-
stitial and tubular) cells in cattle, Bos taurus. 2. Zel(forsch. 74,505- 528.
HOLTHOFER, H., MIETTINEN, A., LEHTO, V-P., LEHTONEN, E., and VIR- TANEN, I. (1984). Expression of vimentin and cytokeratin types of
intermediate filament proteins in developing and adult human kid-
neys. Lab. Invest. 50, 552-559. HOLTH~FER, H., MIETTINEN, A., PAASIVUO, R., LEHTO, V.-P., LINI)ER,
E., ALFTHAN, O., and VIRTANEN, I. (1983). Cellular origin and dif- ferentiation of renal carcinomas. Lab. Invest. 49, 317-326.
JOST, A., VIGIER, B., PR~PIN, J., and PERCHELLET, J. P. (1973). Studies
on sex differentiation in mammals. Recent. Prop. Horm. Res. 29, l- 41.
JOST, A., MAGRE, S., and AGELOPOULOU, R. (1981). Early stages of tes- ticular differentiation in the rat. Hum. Genet. 58,59-63.
KARNOVSKY, M. J. (1971). Use of ferrocyanide-reduced osmium tetroxide in electron microscopy. J. Cell. Biol. Abstr. 284, 146.
KESKI-OJA, J., LEHTO, V.-P., and VIRTANEN, I. (1981). Keratin filaments of mouse epidermal cells are rapidly affected by epidermal growth factor. J. CelZ. Biol. 90, 537-541.
KOHLER, G., HOWE, S. C., and MILSTEIN, C. (1976). Fusion between immunoglobulin-secreting and nonsecreting myeloma cell lines. Eur. J Immunol. 6, 292-295.
LAEMMLI, U. K. (1970). Cleavage of structural proteins during the as-

44 DEVELOPMENTAL BIOLOGY VOLUME 117, 1986
sembly of the head of bacteriophage T4. Nature (Londm) 227, 680- 683.
LANE, E. B., HOGAN, B. L. M., KURKINEN, M., and GARRELS, J. I. (1983). Co-expression of vimentin and cytokeratins in parietal endoderm cells of early mouse embryo. Nature (London) 303, W-704.
LEHTONEN, E., LEHTO, V.-P., PAASIVUO, R., and VIRTANEN, I. (1983). Parietal and visceral endoderm differ in their expression of inter- mediate filaments. EMBO J. 2,1023-1028.
MERCHANT-LARIOS, H. (1979). Ultrastructural events in horse gonadal morphogenesis. J. Reprod. Pert. 27, S479-485.
MIETTINEN, M., FRANSSILA, K., LEHTO, V.-P., PAASIVUO, R., and VIR- TANEN, I. (1984). Expression of intermediate filament proteins in thyroid gland and thyroid tumors. Lab. Invest. 50,262-270.
MIETTINEN, M., VIRTANEN, I., and TALERMAN, A. (1985). Intermediate filament proteins in human testis and testicular germ cell tumors. Am. J. Pathol. 120,402-410.
MOLL, R., FRANKE, W. W., SCHILLER, D. L., GEIGER, B., and KREPLER, R. (1982). The catalog of human cytokeratins: patterns of expression in normal epithelia, tumors and cultured cells. Cell. 31, 11-24.
ORTH, J. M. (1982). Proliferation of Sertoli cells in fetal and postnatal rats: A quantitative autoradiographic study. Anat. Rec. 203, 485-
492. OSBORN, M., GEISLER, N., SHAW, G., SHARP, G., and WEBER, K. (1982).
Intermediate filaments. Cold Spring Hurbor Symp. @ant. Biol. 46,
413-429. PARANKO, J., PELLINIEMI, L. J., VAHERI, A., FOIDART, J-M., and LAK-
KALA-PARANKO, T. (1983). Morphogenesis and fibronectin in sexual differentiation of rat embryonic gonads. Diferentiation 23, S72-S81.
PELLINIEMI, L. J. (1975). Ultrastructure of the early ovary and testis in pig embryos. Am. J. Anat. 144,89-112.
PELLINIEMI, L. J., PARANKO, J., GRUND, S. K., FR~JDMAN, K., FOIDART, J-M., and LAKKALA-PARANKO, T. (1984). Morphological differentia- tion of Sertoli cells. INSERMSymp. Ser. 123,121-140.
PELLINIEMI, L. J., PARANKO, J., GRUND, S. K., FR~JDMAN, K., FOIDART, J-M., and LAKKALA-PARANKO, T. (1985). Extracellular matrix in tes- ticular differentiation. Ann. NYAcad. Sci. 438,405-416.
RAMAEKERS, F., HUYSMANS, A., MOESKER, O., KANT, A., JAP, P., HER-
MAN, C., and VOOIJS, P. (1983). Monoclonal antibody to keratin fil- aments, specific for glandular epithelia and their tumors. Lab. Invest 49,353-361.
SPRUILL, W. A., STEINER, A. L., TRES, L. L., and KIERSZENBAUM, A. L. (1983a). Follicle-stimulating hormone-dependent phosphorylation of vimentin in cultures of rat Sertoli cells, Proc. N&l. Acad. Sci. USA
80,993-997.
SPRUILL, W. A., ZYSK, J. R., TRES, L. L., and KIERSZENBAUM, A. L. (1983b). Calcium/calmodulin-dependent phosphorylation of vimentin in rat Sertoli cells. Proc. Natl. Acad Sci. USA 80, 760-764.
TAPANAINEN, J., KUOPIO, T., PELLINIEMI, L. J., and HUHTANIEMI, I.
(1984). Rat testicular endogenous steroids and number of Leydig cells between the fetal period and sexual maturity. Biol. Reprod. 31,
1027-1035.
THOMPSON, E. B., TOMKINS, G. M., and CURRAN, J. F. (1966). Induction of tyrosine oc-ketoglutarate transaminase by steroid hormones in a newly established tissue culture cell line. Proc. N&l. Acad. Sci.
USA 56,296-303.
TIMPL, R., RHODE, H., ROBEY, P. G., RENNARD, S. I., FOIDART, J. M., and MARTIN, G. R. (1979). Laminin-A glycoprotein from basement membranes. J. BioL Chem. 254,9933-9937.
TOWBIN, H., STAEHELIN, T., and GORDON, J. (1979). Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc. NatL Acud Sci. USA 76,4350- 4354.
UPADHYAY, S., LUCIANI, J. M., and ZAMBONI, L. (1981). The role of the mesonephros in the development of the mouse testis and its excurrent pathways. In “Development and Function of Reproductive Organs” (A. G. Byskov, and H. Peters, eds.), pp. 18-27. International Congress Series 559. Excerpta Medica, Amsterdam.
WARTENBERG, H. (1978). Human testicular development and the role of the mesonephros in the origin of a dual Sertoli cell system. An-
drologia 10, 1-21.
WARTENBERG, H. (1981). The influence of the mesonephric blastema on gonadal development and sexual differentiation. In “Development and Function of Reproductive Organs” (A. G. Byskov, and H. Peters, eds.), pp. 3-12. International Congress Series 559. Excerpta Medica, Amsterdam.
WARTENBERG, H. (1983). Structural aspects of gonadal differentiation in mammals and birds. Diflerentiation 23, S64-S71.
VAN VORSTENBOSCH, C. J. A. H. V., COLENBRANDER, B., and WENSING, C. J. G. (1984a). Cytoplasmic filaments in fetal and neonatal pig testis. Eur. .I Cell Biol. 34, 292-299.
VAN VORSTENBOSCH, C. J. A. H. V., SPEK, E., COLENBRANDER, B., and WENSING, C. J. G. (198413). Sertoli cell development of pig testis in the fetal and neonatal period. Biol. Reprod. 31, 565-577.
VIRTANEN, I., LEHTO, V.-P., LEHTONEN, E., VARTIO, T., STENMAN, S., KURKI, P., WAGER, O., SMALL, J. V., DAHL, D., and BADLEY, R. A. (1981). Expression of intermediate filaments in cultured cells. J. Cell Sci. 50, 45-63.
VIRTANEN, I., MIETTINEN, M., LEHTO, V.-P., KARINIEMI, A.-L., and PAA-
SIVUO, R. (1985). Diagnostic application of monoclonal antibodies to intermediate filaments. Ann, N. Y. Acad. Sci. 455,635-648.
ZAMBONI, L., UPADHYAY, S., BEZARD, J., and MAULOON, P. (1981). The role of the mesonephros in the development of the sheep testis and its current pathways. In “Development and Function of Reproductive Organs” (A. G. Byskov, and H. Peters, eds.), pp. 31-40. International Congress Series 559. Excerpta Medica, Amsterdam.




















