
Disturbed Expression of Sox9 in Pre-Sertoli Cells Underlies Sex-Reversal in Mice B6.Ytir
9
114 BIOLOGY OF REPRODUCTION 70, 114–122 (2004) Published online before print 17 September 2003. DOI 10.1095/biolreprod.103.016824 Disturbed Expression of Sox9 in Pre-Sertoli Cells Underlies Sex-Reversal in Mice B6.Y tir1 Norma Moreno-Mendoza, 3 Leda Torres-Maldonado, 3 Jesu ´s Chimal-Monroy, 3 Vincent Harley, 4 and Horacio Merchant-Larios 2,3 Department of Cell Biology and Physiology, 3 Instituto de Investigaciones Biome ´dicas, UNAM. Me ´xico, D.F. Me ´xico 04510 Prince Henry’s Institute of Medical Research, 4 Monash Medical Centre, Clayton, Victoria 3168, Australia ABSTRACT Sry in some varieties of Mus musculus domesticus fails to form normal testis when introduced into the C57BL/6J (B6) strain. We studied the developmental pattern of pre-Sertoli cells that express Sox9 by immunofluorescence and the profile levels of Sox9 transcripts by semiquantitative reverse transcriptase polymerase chain reaction and in situ hybridization in devel- oping gonads of B6-Y tir mice. Sox9-positive cells (pre-Sertoli cells) appeared in all B6.Y tir genital ridges at 11.5 and 12.5 days postcoitum (dpc). However, at 13.5 dpc, Sox9-positive cells were not detected only in 50% of the B6.Y tir gonads compared with 100% of B6 gonads. Although pre-Sertoli cells formed the seminiferous cords after 14.5 dpc in the medial region of the B6.Y tir gonad, the cranial and caudal regions formed ovarian tis- sue. Further, B6.Y tir ovaries have lower levels of Sox9 than ovo- testes at all fetal stages. These results suggest that although the pre-Sertoli cell lineage forms in B6.Y tir genital ridges, its further differentiation into Sertoli cells is apparently prevented. The cause may be the low levels of Sox9 and down-regulation of its product. Results suggest that inhibitory signals of Sox9 acting along the whole genital ridge or only at its cranial and/or caudal regions underlie formation of B6.Y tir ovaries or ovotestes, re- spectively. Furthermore, our results suggest that infertility of B6.Y tir females may be due to the abnormal presence of Sox9 transcripts in their ovaries. developmental biology, gene regulation, ovary, Sertoli cells INTRODUCTION It has been well established that the expression of Sry in undifferentiated gonads of mouse embryos initiates the events leading to testicular differentiation [1, 2]. In some cases of male to female sex reversal, Sry mutations or dif- fering Y alleles can result in differentiation of an ovary instead of a testis [3, 4]. An example occurs in gonads of the strain C57BL/6J (B6) when its Y chromosome is re- placed by the Y chromosome (Y dom ) of some varieties of Mus musculus domesticus [5, 6]. Failure of normal testis formation is observed in all B6-Y dom offspring (B6.Y tir from Tirano, Italy, or B6-Y pos from Poschiavo, Switzer- 1 Supported by a Young Investigator grant (CONACYT-J36337-N) to N.M.M. 2 Correspondence: Horacio Merchant-Larios, Instituto de Investigaciones Biome ´dicas, UNAM. Apartado Postal 70228, Me ´xico, D.F. Me ´xico 04510. FAX: 52 55 5662 3897; e-mail: [email protected] Received: 3 March 2003. First decision: 10 April 2003. Accepted: 2 September 2003. Q 2004 by the Society for the Study of Reproduction, Inc. ISSN: 0006-3363. http://www.biolreprod.org land). About half of the B6-Y dom embryos develop bilateral fetal ovotestes and become fertile males after birth, whereas the other half form ovaries and develop as infertile females [5–8]. It has been proposed that B6-Y dom sex reversal results from an improper interaction between the Y dom testis-de- termining gene (Sry) and dominant autosomal genes in the genome of the C57BL/6J strain [6, 9, 10], but the identity of these genes remains unknown. On the other hand, it was found that transcription of Sry in B6-Y tir gonads starts at the same time as in normal XY gonads, but its expression persists longer in B6-Y tir gonads [11]. The finding that lev- els of Sry were lower in sex-reversed B6.Y tir gonads than in non-sex-reversed B6-Y dom strains suggests that low lev- els and/or mistiming of Sry may cause failure of testis dif- ferentiation [12]. However, Lee and Taketo [13] observed that levels of Sry in normal male gonads of the C57BL/6J strain are as low as in the B6-Y tir gonad, suggesting that low levels of Sry transcripts cannot be the sole cause of B6-Y tir sex reversal. Thus the molecular events underlying sex reversal in the B6-Y tir mouse still remain to be eluci- dated. SOX9, a SRY-related gene, was originally identified as responsible for campomelic dysplasia, a severe form of dwarfism often associated with XY male-to-female sex re- versal [14, 15]. Like SRY, dosage may be important for proper function of SOX9, since mutations in humans appear to be present in only one allele, suggesting that sex reversal and campomelic dysplasia could be due to haploinsuffi- ciency of this gene product [16]. Sox9 has been detected in the mouse urogenital complex of both sexes at 10.5 dpc. Beginning at 11.5 dpc, just after Sry is expressed in the male gonad, Sox9 is up-regulated in male gonads and down-regulated in female gonads. After seminiferous cord formation, Sox9 expression is specific to Sertoli cells [17], and Sox9 is probably one of the first genes induced by Sry in male gonads [18]. The DNA-binding domains of both Sry and Sox9 proteins are highly conserved and can func- tionally substitute for each other [19]. A mouse line with a transgene inserted upstream of Sox9 showed that the de- velopment of XX males is consistent with Sry being an indirect repressor downstream target of Sox9 [20, 21]. Re- cently, female to male sex-reversal induced by ectopically expressing Sox9 in XX gonads indicated that it can substi- tute for Sry in the events necessary to form a testis and is likely to act downstream [22]. It is assumed that Sry and Sox9 are expressed in pre- Sertoli cells from which an orchestrated series of intercel- lular signals leads the undifferentiated genital ridge to form a testis [23]. Since it is known that Sry is expressed in B6-
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Disturbed Expression of Sox9 in Pre-Sertoli Cells Underlies Sex-Reversal in Mice B6.Ytir

114
BIOLOGY OF REPRODUCTION 70, 114–122 (2004)Published online before print 17 September 2003.DOI 10.1095/biolreprod.103.016824
Disturbed Expression of Sox9 in Pre-Sertoli Cells Underlies Sex-Reversalin Mice B6.Ytir1
Norma Moreno-Mendoza,3 Leda Torres-Maldonado,3 Jesus Chimal-Monroy,3 Vincent Harley,4
and Horacio Merchant-Larios2,3
Department of Cell Biology and Physiology,3 Instituto de Investigaciones Biomedicas, UNAM. Mexico,D.F. Mexico 04510Prince Henry’s Institute of Medical Research,4 Monash Medical Centre, Clayton, Victoria 3168, Australia
ABSTRACT
Sry in some varieties of Mus musculus domesticus fails toform normal testis when introduced into the C57BL/6J (B6)strain. We studied the developmental pattern of pre-Sertoli cellsthat express Sox9 by immunofluorescence and the profile levelsof Sox9 transcripts by semiquantitative reverse transcriptasepolymerase chain reaction and in situ hybridization in devel-oping gonads of B6-Ytir mice. Sox9-positive cells (pre-Sertolicells) appeared in all B6.Ytir genital ridges at 11.5 and 12.5 dayspostcoitum (dpc). However, at 13.5 dpc, Sox9-positive cellswere not detected only in 50% of the B6.Ytir gonads comparedwith 100% of B6 gonads. Although pre-Sertoli cells formed theseminiferous cords after 14.5 dpc in the medial region of theB6.Ytir gonad, the cranial and caudal regions formed ovarian tis-sue. Further, B6.Ytir ovaries have lower levels of Sox9 than ovo-testes at all fetal stages. These results suggest that although thepre-Sertoli cell lineage forms in B6.Ytir genital ridges, its furtherdifferentiation into Sertoli cells is apparently prevented. Thecause may be the low levels of Sox9 and down-regulation of itsproduct. Results suggest that inhibitory signals of Sox9 actingalong the whole genital ridge or only at its cranial and/or caudalregions underlie formation of B6.Ytir ovaries or ovotestes, re-spectively. Furthermore, our results suggest that infertility ofB6.Ytir females may be due to the abnormal presence of Sox9transcripts in their ovaries.
developmental biology, gene regulation, ovary, Sertoli cells
INTRODUCTION
It has been well established that the expression of Sry inundifferentiated gonads of mouse embryos initiates theevents leading to testicular differentiation [1, 2]. In somecases of male to female sex reversal, Sry mutations or dif-fering Y alleles can result in differentiation of an ovaryinstead of a testis [3, 4]. An example occurs in gonads ofthe strain C57BL/6J (B6) when its Y chromosome is re-placed by the Y chromosome (Ydom) of some varieties ofMus musculus domesticus [5, 6]. Failure of normal testisformation is observed in all B6-Ydom offspring (B6.Ytir
from Tirano, Italy, or B6-Ypos from Poschiavo, Switzer-
1Supported by a Young Investigator grant (CONACYT-J36337-N) toN.M.M.2Correspondence: Horacio Merchant-Larios, Instituto de InvestigacionesBiomedicas, UNAM. Apartado Postal 70228, Mexico, D.F. Mexico 04510.FAX: 52 55 5662 3897; e-mail: [email protected]
Received: 3 March 2003.First decision: 10 April 2003.Accepted: 2 September 2003.Q 2004 by the Society for the Study of Reproduction, Inc.ISSN: 0006-3363. http://www.biolreprod.org
land). About half of the B6-Ydom embryos develop bilateralfetal ovotestes and become fertile males after birth, whereasthe other half form ovaries and develop as infertile females[5–8].
It has been proposed that B6-Ydom sex reversal resultsfrom an improper interaction between the Ydom testis-de-termining gene (Sry) and dominant autosomal genes in thegenome of the C57BL/6J strain [6, 9, 10], but the identityof these genes remains unknown. On the other hand, it wasfound that transcription of Sry in B6-Ytir gonads starts atthe same time as in normal XY gonads, but its expressionpersists longer in B6-Ytir gonads [11]. The finding that lev-els of Sry were lower in sex-reversed B6.Ytir gonads thanin non-sex-reversed B6-Ydom strains suggests that low lev-els and/or mistiming of Sry may cause failure of testis dif-ferentiation [12]. However, Lee and Taketo [13] observedthat levels of Sry in normal male gonads of the C57BL/6Jstrain are as low as in the B6-Ytir gonad, suggesting thatlow levels of Sry transcripts cannot be the sole cause ofB6-Ytir sex reversal. Thus the molecular events underlyingsex reversal in the B6-Ytir mouse still remain to be eluci-dated.
SOX9, a SRY-related gene, was originally identified asresponsible for campomelic dysplasia, a severe form ofdwarfism often associated with XY male-to-female sex re-versal [14, 15]. Like SRY, dosage may be important forproper function of SOX9, since mutations in humans appearto be present in only one allele, suggesting that sex reversaland campomelic dysplasia could be due to haploinsuffi-ciency of this gene product [16]. Sox9 has been detected inthe mouse urogenital complex of both sexes at 10.5 dpc.Beginning at 11.5 dpc, just after Sry is expressed in themale gonad, Sox9 is up-regulated in male gonads anddown-regulated in female gonads. After seminiferous cordformation, Sox9 expression is specific to Sertoli cells [17],and Sox9 is probably one of the first genes induced by Sryin male gonads [18]. The DNA-binding domains of bothSry and Sox9 proteins are highly conserved and can func-tionally substitute for each other [19]. A mouse line with atransgene inserted upstream of Sox9 showed that the de-velopment of XX males is consistent with Sry being anindirect repressor downstream target of Sox9 [20, 21]. Re-cently, female to male sex-reversal induced by ectopicallyexpressing Sox9 in XX gonads indicated that it can substi-tute for Sry in the events necessary to form a testis and islikely to act downstream [22].
It is assumed that Sry and Sox9 are expressed in pre-Sertoli cells from which an orchestrated series of intercel-lular signals leads the undifferentiated genital ridge to forma testis [23]. Since it is known that Sry is expressed in B6-
115Sox9 EXPRESSION IN B6.Ytir SEX-REVERSED
TABLE 1. SOX9 epression in serially sectioned gonads of B6.Ytir embry-os.*
Age (dpc)
11.5
R L
12.5
R L
13.5
R L
15.5
R L
17.5
R L
18.5
R L
111111
111111
111111
111111
221112
216111
111122
161622
212126
211622
112211
122221
111111
111111
111111
111111
112122
662162
212212
112222
121226
121222
122222
111222
* Right (R) and left (L) gonads. N 5 12 B6. Ytir embryos at each age. 1,6, or 2 5 abundant, few, or absent SOX9-expressing cells, respectively.
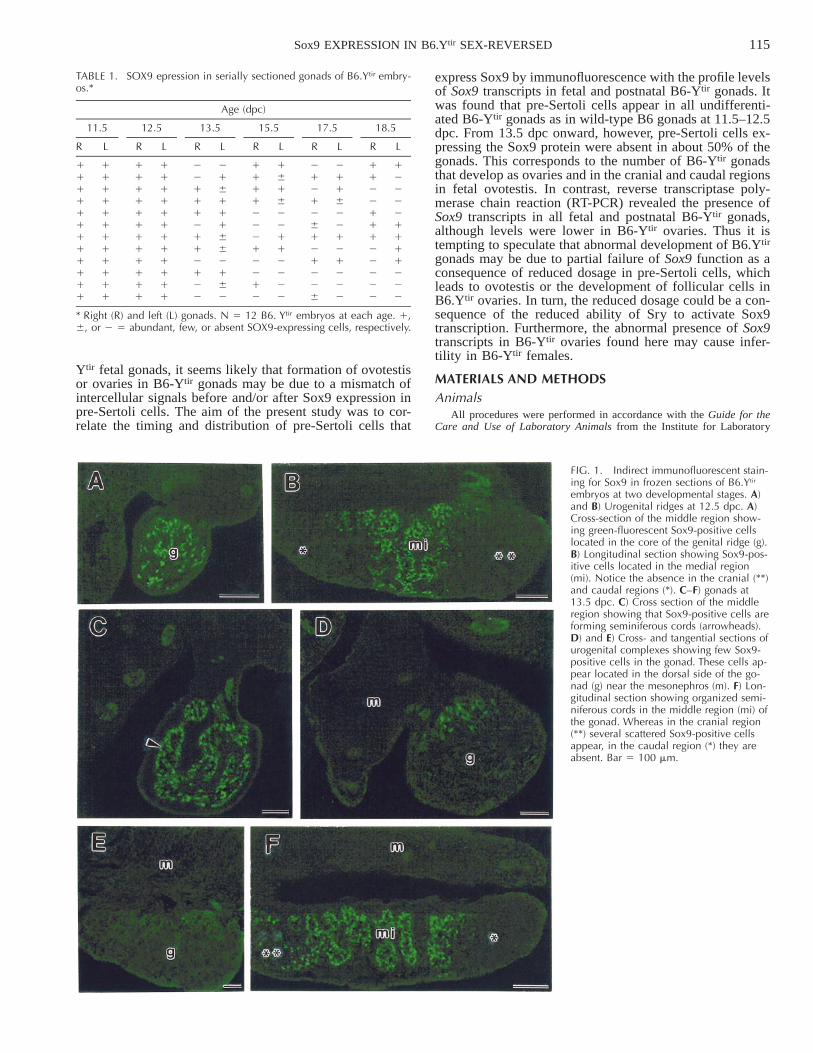
FIG. 1. Indirect immunofluorescent stain-ing for Sox9 in frozen sections of B6.Ytir
embryos at two developmental stages. A)and B) Urogenital ridges at 12.5 dpc. A)Cross-section of the middle region show-ing green-fluorescent Sox9-positive cellslocated in the core of the genital ridge (g).B) Longitudinal section showing Sox9-pos-itive cells located in the medial region(mi). Notice the absence in the cranial (**)and caudal regions (*). C–F) gonads at13.5 dpc. C) Cross section of the middleregion showing that Sox9-positive cells areforming seminiferous cords (arrowheads).D) and E) Cross- and tangential sections ofurogenital complexes showing few Sox9-positive cells in the gonad. These cells ap-pear located in the dorsal side of the go-nad (g) near the mesonephros (m). F) Lon-gitudinal section showing organized semi-niferous cords in the middle region (mi) ofthe gonad. Whereas in the cranial region(**) several scattered Sox9-positive cellsappear, in the caudal region (*) they areabsent. Bar 5 100 mm.
Ytir fetal gonads, it seems likely that formation of ovotestisor ovaries in B6-Ytir gonads may be due to a mismatch ofintercellular signals before and/or after Sox9 expression inpre-Sertoli cells. The aim of the present study was to cor-relate the timing and distribution of pre-Sertoli cells that
express Sox9 by immunofluorescence with the profile levelsof Sox9 transcripts in fetal and postnatal B6-Ytir gonads. Itwas found that pre-Sertoli cells appear in all undifferenti-ated B6-Ytir gonads as in wild-type B6 gonads at 11.5–12.5dpc. From 13.5 dpc onward, however, pre-Sertoli cells ex-pressing the Sox9 protein were absent in about 50% of thegonads. This corresponds to the number of B6-Ytir gonadsthat develop as ovaries and in the cranial and caudal regionsin fetal ovotestis. In contrast, reverse transcriptase poly-merase chain reaction (RT-PCR) revealed the presence ofSox9 transcripts in all fetal and postnatal B6-Ytir gonads,although levels were lower in B6-Ytir ovaries. Thus it istempting to speculate that abnormal development of B6.Ytir
gonads may be due to partial failure of Sox9 function as aconsequence of reduced dosage in pre-Sertoli cells, whichleads to ovotestis or the development of follicular cells inB6.Ytir ovaries. In turn, the reduced dosage could be a con-sequence of the reduced ability of Sry to activate Sox9transcription. Furthermore, the abnormal presence of Sox9transcripts in B6-Ytir ovaries found here may cause infer-tility in B6-Ytir females.
MATERIALS AND METHODSAnimals
All procedures were performed in accordance with the Guide for theCare and Use of Laboratory Animals from the Institute for Laboratory

116 MORENO-MENDOZA ET AL.
FIG. 2. Light microscopy of a semithinsection of a plastic-embedded B6.Ytir ovo-testis at 15.5 dpc. The medullary regiondisplays well-organized seminiferous cords(sc) formed by Sertoli (s) and germ cells(gc). In the cortical region, the transitionalovarian sex cords formed by epithelialcells (arrows), germ cells (gc), and darkpicnotic cells (arrowheads) are evident.Rectangles A and B correspond to the twoinsets of the electron-micrographs shownin Figure 3. Bar 5 50 mm.
Animal Research of the National Academy of Sciences [24]. B6.Ytir micekept in our colony (backcross generations N 30–35) that possess the B6genetic background and the Y chromosome from M. musculus domesticus(from Tirano, Italy) were produced as previously described [7]. B6.Ytir
males were mated with B6 females, and the day of vaginal plug was takento be 0.5 days postcoitum (dpc). Bilateral gonads were dissected fromembryos at 11.5 up to 18.5 dpc and 30 and 60 days postpartum (dpp).
Zfy PCR AnalysisThe chromosomal sex of the B6-Ytir embryos was determined by PCR
amplification of the 600 base pair (bp) fragment of the Y-encoded geneZfy. The PCR was performed using DNA isolated from the fetal tissuesor from the tails at 30 and 60 dpp. The tissue was homogenized andincubated in lysis buffer (100 mM Tris-HCl pH 8.0, 10 mM EDTA pH8.0, 0.5% Tween 20, 0.5% NP-40, and 0.4 mg/ml proteinase K) [13] at558C overnight, and proteinase K was inactivated by heating at 858C for10 minutes. Fifteen microliters of the lysate were diluted in 85 ml ofddH2O. Two microliters of each DNA sample was processed for PCRamplification in a total volume of 20 ml with 0.25 mM of each of thefollowing primers: Zfy1, 59-AAG ATA AGC TTA CAT AAT CAC ATGGA-39; Zfy2, 59-CCT ATG AAA TCC TTT GCT GCA CAT GT-39 forZfy; Actin 1, GGG TCA GAA GGA TTC CTA TG; and Actin 2, GGTCTC AAA CAT GAT CTG GG for b-actin used as an internal control[26]. The primers were in 10 mM Tris-HCl pH 8.3, 50 mM KCl, 0.01%gelatin, 2.0 mM MgCl2, 0.2 mM of each dNTP and 1.0 U Taq DNApolymerase.
The amplification program was 948C/10 min (once), 948C/15 sec,608C/15 sec, 728C/30 sec (35 cycles), and 728C/10 min (once) [26]. ThePCR reactions were electrophoresed on a 1% agarose gel in TAE 13buffer and visualized with ethidium bromide fluorescence in UV light.
Immunocytochemical Detection of SOX9Antigen-affinity antibody raised to the human SOX9 C-terminal 24-
amino-acid epitope (VPSIPQTHSPQHWEQPVYTQLTRP) was preparedaccording to the previously described protocol [17]. For immunocyto-chemical detection of Sox9, B6-Ytir gonads were dissected from embryosfrom 11.5 to 18.5 dpc and fixed in 4% paraformaldehyde in PBS for 10min, washed in PBS, and incubated in 30% sucrose overnight. The gonadswere embedded in OCT medium (Tissue-Tek, Sakura Finetek, Torrance,CA) and frozen at 2708C. Serial sections of 10-mm thickness were used.After air drying, sections were treated with 1% Triton X-100 in water,rinsed with PBS, and blocked using 1% bovine serum albumin. The sec-tions were then incubated overnight in Sox9 polyclonal antibody diluted1:500. After washing in PBS, the sections were treated with fluorescein-
goat anti-rabbit IgG (Sigma, St. Louis, MO) diluted 1:200 in blockingsolution.
RNA Isolation and RT-PCR
Gonads were dissected from B6 and B6-Ytir embryos of 18.5 dpc andfrom animals of 30 and 60 dpp. The tissues were immediately frozen indry ice and stored at 2708 C. Detection was carried out in the two fetalgonads or single gonads of postnatal animals. Unilateral fetal gonads fromhermaphrodite mice were pooled as two ovaries or two testes. A DNAprobe containing a 243 bp fragment of b-actin was used as the internalexpression control. Total RNA was isolated from gonads using Trizol re-agent (Life Technologies Invitrogen, Carlsbad, CA); based on the methodof Chomczynski and Sacchi [27], a few modifications were introduced toimprove the RNA isolation, the volume of chloroform was increased todouble, and the time of precipitation in isopropanol was increased from10 minutes to overnight. Total RNA was resuspended in 12 ml of DEPC-treated ddH2O. Reverse transcriptase polymerase chain reaction (RT-PCR)amplifications were performed using primers designed on the basis of themouse Sox9 as previously reported [18]. The sense primer was Sox9-1,59-GTG GCA AGT ATT GGT CAA-39; the antisense primer was Sox9-2, 59-GAA CAG ACT CAC ATC TCT-39. For expression control, ampli-fication of oligonucleotides used as internal controls were Actin 1 andActin 2 as previously described [25]. The amplification reactions werecarried out by means of the One Step RT-PCR kit (Life TechnologiesInvitrogen) using 3 ml of total RNA per reaction, 13 buffer reaction mixcontaining 0.2 mM of each dNTP, 1.2 mM MgSO4, and 200 ng of b-actinprimers or 300 ng of Sox9 primers in separate tubes. The enzyme usedwas a mixture of Superscript II RT and Taq DNA polymerase Platinum(Life Technologies Invitrogen) in a volume of 20ml. The amplificationprogram was 508C for 30 min (once); 948C/5 min (once), 948C/30 sec,568C/60 sec, 728C/60 sec (25 cycles), 728C/10 min (once) within the linearrange previously described [18]. In order to confirm that the amplifiedproducts were not genomic DNA in origin, we used an RT-free control(Taq DNA polymerase but not template DNA added) for each RT-PCR[28].
Ten microliters of each RT-PCR reaction were electrophoresed on a2.5% agarose gel with 0.1 mg/ml ethidium bromide in TAE 13 buffer[29]. Gels were visualized by UV transillumination and photographed witha DS-34 Polaroid camera (Polaroid, Waltham, MA). The size of the am-plified products was 310 bp for Sox9 and 243 for b-actin. The intensityof each band was quantified by densitometry using the Scion Image pro-gram. Results were expressed as arbitrary units of the ratio between Sox9and b-actin mRNA levels.
117Sox9 EXPRESSION IN B6.Ytir SEX-REVERSED
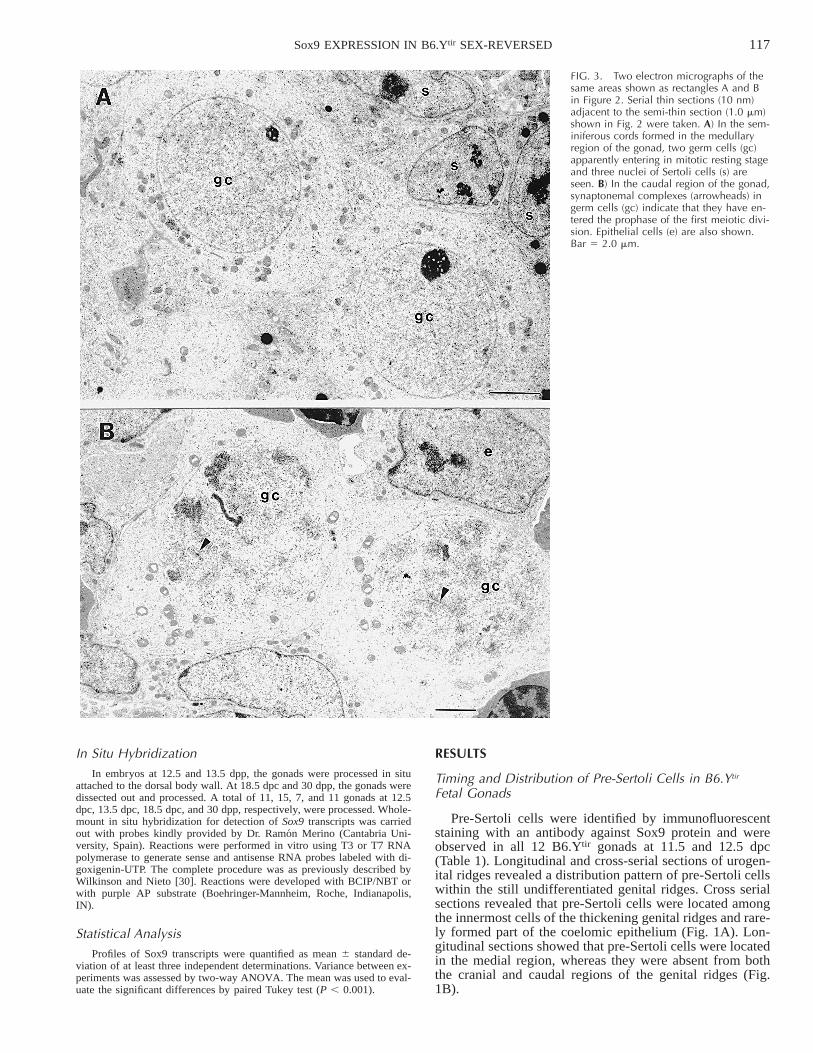
FIG. 3. Two electron micrographs of thesame areas shown as rectangles A and Bin Figure 2. Serial thin sections (10 nm)adjacent to the semi-thin section (1.0 mm)shown in Fig. 2 were taken. A) In the sem-iniferous cords formed in the medullaryregion of the gonad, two germ cells (gc)apparently entering in mitotic resting stageand three nuclei of Sertoli cells (s) areseen. B) In the caudal region of the gonad,synaptonemal complexes (arrowheads) ingerm cells (gc) indicate that they have en-tered the prophase of the first meiotic divi-sion. Epithelial cells (e) are also shown.Bar 5 2.0 mm.
In Situ HybridizationIn embryos at 12.5 and 13.5 dpp, the gonads were processed in situ
attached to the dorsal body wall. At 18.5 dpc and 30 dpp, the gonads weredissected out and processed. A total of 11, 15, 7, and 11 gonads at 12.5dpc, 13.5 dpc, 18.5 dpc, and 30 dpp, respectively, were processed. Whole-mount in situ hybridization for detection of Sox9 transcripts was carriedout with probes kindly provided by Dr. Ramon Merino (Cantabria Uni-versity, Spain). Reactions were performed in vitro using T3 or T7 RNApolymerase to generate sense and antisense RNA probes labeled with di-goxigenin-UTP. The complete procedure was as previously described byWilkinson and Nieto [30]. Reactions were developed with BCIP/NBT orwith purple AP substrate (Boehringer-Mannheim, Roche, Indianapolis,IN).
Statistical AnalysisProfiles of Sox9 transcripts were quantified as mean 6 standard de-
viation of at least three independent determinations. Variance between ex-periments was assessed by two-way ANOVA. The mean was used to eval-uate the significant differences by paired Tukey test (P , 0.001).
RESULTS
Timing and Distribution of Pre-Sertoli Cells in B6.Ytir
Fetal Gonads
Pre-Sertoli cells were identified by immunofluorescentstaining with an antibody against Sox9 protein and wereobserved in all 12 B6.Ytir gonads at 11.5 and 12.5 dpc(Table 1). Longitudinal and cross-serial sections of urogen-ital ridges revealed a distribution pattern of pre-Sertoli cellswithin the still undifferentiated genital ridges. Cross serialsections revealed that pre-Sertoli cells were located amongthe innermost cells of the thickening genital ridges and rare-ly formed part of the coelomic epithelium (Fig. 1A). Lon-gitudinal sections showed that pre-Sertoli cells were locatedin the medial region, whereas they were absent from boththe cranial and caudal regions of the genital ridges (Fig.1B).
118 MORENO-MENDOZA ET AL.
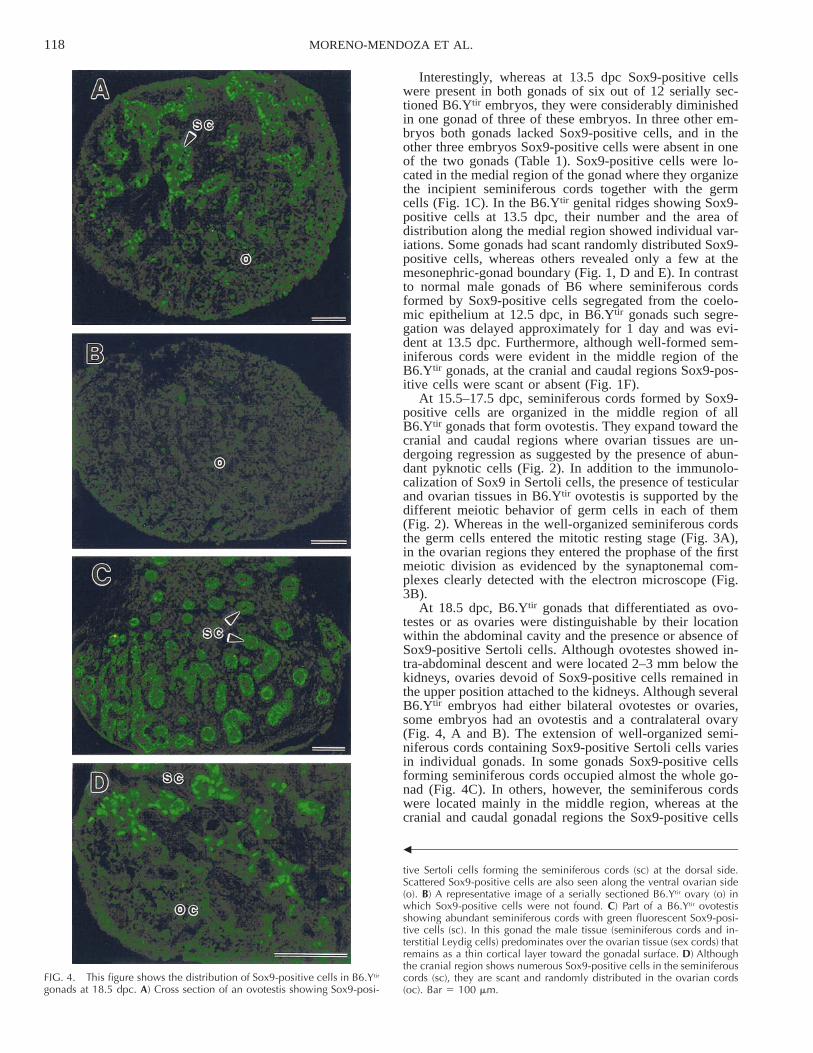
FIG. 4. This figure shows the distribution of Sox9-positive cells in B6.Ytir
gonads at 18.5 dpc. A) Cross section of an ovotestis showing Sox9-posi-
b
tive Sertoli cells forming the seminiferous cords (sc) at the dorsal side.Scattered Sox9-positive cells are also seen along the ventral ovarian side(o). B) A representative image of a serially sectioned B6.Ytir ovary (o) inwhich Sox9-positive cells were not found. C) Part of a B6.Ytir ovotestisshowing abundant seminiferous cords with green fluorescent Sox9-posi-tive cells (sc). In this gonad the male tissue (seminiferous cords and in-terstitial Leydig cells) predominates over the ovarian tissue (sex cords) thatremains as a thin cortical layer toward the gonadal surface. D) Althoughthe cranial region shows numerous Sox9-positive cells in the seminiferouscords (sc), they are scant and randomly distributed in the ovarian cords(oc). Bar 5 100 mm.
Interestingly, whereas at 13.5 dpc Sox9-positive cellswere present in both gonads of six out of 12 serially sec-tioned B6.Ytir embryos, they were considerably diminishedin one gonad of three of these embryos. In three other em-bryos both gonads lacked Sox9-positive cells, and in theother three embryos Sox9-positive cells were absent in oneof the two gonads (Table 1). Sox9-positive cells were lo-cated in the medial region of the gonad where they organizethe incipient seminiferous cords together with the germcells (Fig. 1C). In the B6.Ytir genital ridges showing Sox9-positive cells at 13.5 dpc, their number and the area ofdistribution along the medial region showed individual var-iations. Some gonads had scant randomly distributed Sox9-positive cells, whereas others revealed only a few at themesonephric-gonad boundary (Fig. 1, D and E). In contrastto normal male gonads of B6 where seminiferous cordsformed by Sox9-positive cells segregated from the coelo-mic epithelium at 12.5 dpc, in B6.Ytir gonads such segre-gation was delayed approximately for 1 day and was evi-dent at 13.5 dpc. Furthermore, although well-formed sem-iniferous cords were evident in the middle region of theB6.Ytir gonads, at the cranial and caudal regions Sox9-pos-itive cells were scant or absent (Fig. 1F).
At 15.5–17.5 dpc, seminiferous cords formed by Sox9-positive cells are organized in the middle region of allB6.Ytir gonads that form ovotestis. They expand toward thecranial and caudal regions where ovarian tissues are un-dergoing regression as suggested by the presence of abun-dant pyknotic cells (Fig. 2). In addition to the immunolo-calization of Sox9 in Sertoli cells, the presence of testicularand ovarian tissues in B6.Ytir ovotestis is supported by thedifferent meiotic behavior of germ cells in each of them(Fig. 2). Whereas in the well-organized seminiferous cordsthe germ cells entered the mitotic resting stage (Fig. 3A),in the ovarian regions they entered the prophase of the firstmeiotic division as evidenced by the synaptonemal com-plexes clearly detected with the electron microscope (Fig.3B).
At 18.5 dpc, B6.Ytir gonads that differentiated as ovo-testes or as ovaries were distinguishable by their locationwithin the abdominal cavity and the presence or absence ofSox9-positive Sertoli cells. Although ovotestes showed in-tra-abdominal descent and were located 2–3 mm below thekidneys, ovaries devoid of Sox9-positive cells remained inthe upper position attached to the kidneys. Although severalB6.Ytir embryos had either bilateral ovotestes or ovaries,some embryos had an ovotestis and a contralateral ovary(Fig. 4, A and B). The extension of well-organized semi-niferous cords containing Sox9-positive Sertoli cells variesin individual gonads. In some gonads Sox9-positive cellsforming seminiferous cords occupied almost the whole go-nad (Fig. 4C). In others, however, the seminiferous cordswere located mainly in the middle region, whereas at thecranial and caudal gonadal regions the Sox9-positive cells
119Sox9 EXPRESSION IN B6.Ytir SEX-REVERSED
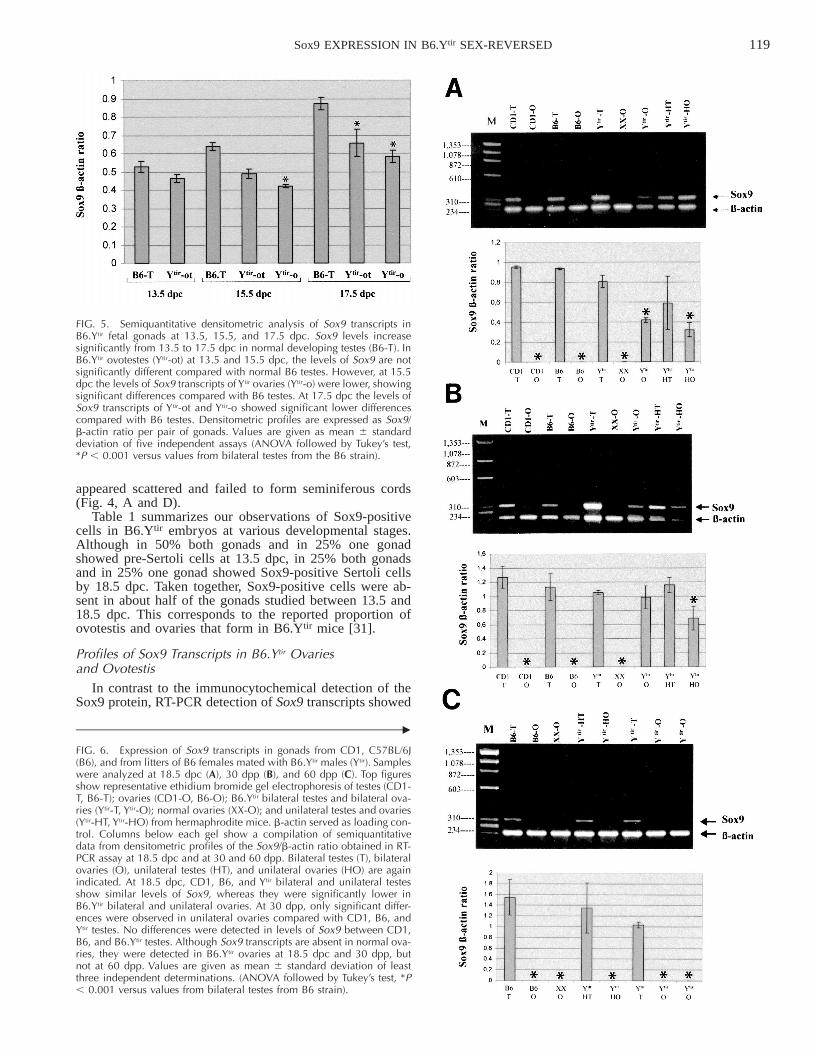
FIG. 5. Semiquantitative densitometric analysis of Sox9 transcripts inB6.Ytir fetal gonads at 13.5, 15.5, and 17.5 dpc. Sox9 levels increasesignificantly from 13.5 to 17.5 dpc in normal developing testes (B6-T). InB6.Ytir ovotestes (Ytir-ot) at 13.5 and 15.5 dpc, the levels of Sox9 are notsignificantly different compared with normal B6 testes. However, at 15.5dpc the levels of Sox9 transcripts of Ytir ovaries (Ytir-o) were lower, showingsignificant differences compared with B6 testes. At 17.5 dpc the levels ofSox9 transcripts of Ytir-ot and Ytir-o showed significant lower differencescompared with B6 testes. Densitometric profiles are expressed as Sox9/b-actin ratio per pair of gonads. Values are given as mean 6 standarddeviation of five independent assays (ANOVA followed by Tukey’s test,*P , 0.001 versus values from bilateral testes from the B6 strain).
c
FIG. 6. Expression of Sox9 transcripts in gonads from CD1, C57BL/6J(B6), and from litters of B6 females mated with B6.Ytir males (Ytir). Sampleswere analyzed at 18.5 dpc (A), 30 dpp (B), and 60 dpp (C). Top figuresshow representative ethidium bromide gel electrophoresis of testes (CD1-T, B6-T); ovaries (CD1-O, B6-O); B6.Ytir bilateral testes and bilateral ova-ries (Ytir-T, Ytir-O); normal ovaries (XX-O); and unilateral testes and ovaries(Ytir-HT, Ytir-HO) from hermaphrodite mice. b-actin served as loading con-trol. Columns below each gel show a compilation of semiquantitativedata from densitometric profiles of the Sox9/b-actin ratio obtained in RT-PCR assay at 18.5 dpc and at 30 and 60 dpp. Bilateral testes (T), bilateralovaries (O), unilateral testes (HT), and unilateral ovaries (HO) are againindicated. At 18.5 dpc, CD1, B6, and Ytir bilateral and unilateral testesshow similar levels of Sox9, whereas they were significantly lower inB6.Ytir bilateral and unilateral ovaries. At 30 dpp, only significant differ-ences were observed in unilateral ovaries compared with CD1, B6, andYtir testes. No differences were detected in levels of Sox9 between CD1,B6, and B6.Ytir testes. Although Sox9 transcripts are absent in normal ova-ries, they were detected in B6.Ytir ovaries at 18.5 dpc and 30 dpp, butnot at 60 dpp. Values are given as mean 6 standard deviation of leastthree independent determinations. (ANOVA followed by Tukey’s test, *P, 0.001 versus values from bilateral testes from B6 strain).
appeared scattered and failed to form seminiferous cords(Fig. 4, A and D).
Table 1 summarizes our observations of Sox9-positivecells in B6.Ytir embryos at various developmental stages.Although in 50% both gonads and in 25% one gonadshowed pre-Sertoli cells at 13.5 dpc, in 25% both gonadsand in 25% one gonad showed Sox9-positive Sertoli cellsby 18.5 dpc. Taken together, Sox9-positive cells were ab-sent in about half of the gonads studied between 13.5 and18.5 dpc. This corresponds to the reported proportion ofovotestis and ovaries that form in B6.Ytir mice [31].
Profiles of Sox9 Transcripts in B6.Ytir Ovariesand Ovotestis
In contrast to the immunocytochemical detection of theSox9 protein, RT-PCR detection of Sox9 transcripts showed
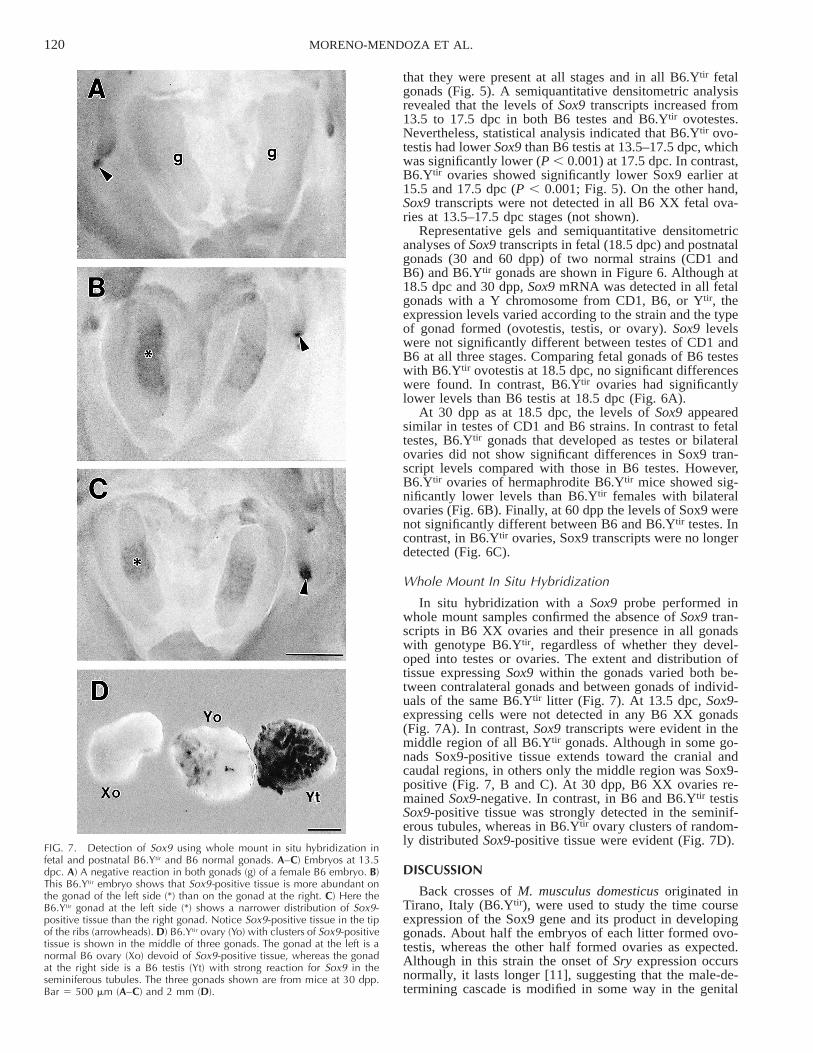
120 MORENO-MENDOZA ET AL.
FIG. 7. Detection of Sox9 using whole mount in situ hybridization infetal and postnatal B6.Ytir and B6 normal gonads. A–C) Embryos at 13.5dpc. A) A negative reaction in both gonads (g) of a female B6 embryo. B)This B6.Ytir embryo shows that Sox9-positive tissue is more abundant onthe gonad of the left side (*) than on the gonad at the right. C) Here theB6.Ytir gonad at the left side (*) shows a narrower distribution of Sox9-positive tissue than the right gonad. Notice Sox9-positive tissue in the tipof the ribs (arrowheads). D) B6.Ytir ovary (Yo) with clusters of Sox9-positivetissue is shown in the middle of three gonads. The gonad at the left is anormal B6 ovary (Xo) devoid of Sox9-positive tissue, whereas the gonadat the right side is a B6 testis (Yt) with strong reaction for Sox9 in theseminiferous tubules. The three gonads shown are from mice at 30 dpp.Bar 5 500 mm (A–C) and 2 mm (D).
that they were present at all stages and in all B6.Ytir fetalgonads (Fig. 5). A semiquantitative densitometric analysisrevealed that the levels of Sox9 transcripts increased from13.5 to 17.5 dpc in both B6 testes and B6.Ytir ovotestes.Nevertheless, statistical analysis indicated that B6.Ytir ovo-testis had lower Sox9 than B6 testis at 13.5–17.5 dpc, whichwas significantly lower (P , 0.001) at 17.5 dpc. In contrast,B6.Ytir ovaries showed significantly lower Sox9 earlier at15.5 and 17.5 dpc (P , 0.001; Fig. 5). On the other hand,Sox9 transcripts were not detected in all B6 XX fetal ova-ries at 13.5–17.5 dpc stages (not shown).
Representative gels and semiquantitative densitometricanalyses of Sox9 transcripts in fetal (18.5 dpc) and postnatalgonads (30 and 60 dpp) of two normal strains (CD1 andB6) and B6.Ytir gonads are shown in Figure 6. Although at18.5 dpc and 30 dpp, Sox9 mRNA was detected in all fetalgonads with a Y chromosome from CD1, B6, or Ytir, theexpression levels varied according to the strain and the typeof gonad formed (ovotestis, testis, or ovary). Sox9 levelswere not significantly different between testes of CD1 andB6 at all three stages. Comparing fetal gonads of B6 testeswith B6.Ytir ovotestis at 18.5 dpc, no significant differenceswere found. In contrast, B6.Ytir ovaries had significantlylower levels than B6 testis at 18.5 dpc (Fig. 6A).
At 30 dpp as at 18.5 dpc, the levels of Sox9 appearedsimilar in testes of CD1 and B6 strains. In contrast to fetaltestes, B6.Ytir gonads that developed as testes or bilateralovaries did not show significant differences in Sox9 tran-script levels compared with those in B6 testes. However,B6.Ytir ovaries of hermaphrodite B6.Ytir mice showed sig-nificantly lower levels than B6.Ytir females with bilateralovaries (Fig. 6B). Finally, at 60 dpp the levels of Sox9 werenot significantly different between B6 and B6.Ytir testes. Incontrast, in B6.Ytir ovaries, Sox9 transcripts were no longerdetected (Fig. 6C).
Whole Mount In Situ Hybridization
In situ hybridization with a Sox9 probe performed inwhole mount samples confirmed the absence of Sox9 tran-scripts in B6 XX ovaries and their presence in all gonadswith genotype B6.Ytir, regardless of whether they devel-oped into testes or ovaries. The extent and distribution oftissue expressing Sox9 within the gonads varied both be-tween contralateral gonads and between gonads of individ-uals of the same B6.Ytir litter (Fig. 7). At 13.5 dpc, Sox9-expressing cells were not detected in any B6 XX gonads(Fig. 7A). In contrast, Sox9 transcripts were evident in themiddle region of all B6.Ytir gonads. Although in some go-nads Sox9-positive tissue extends toward the cranial andcaudal regions, in others only the middle region was Sox9-positive (Fig. 7, B and C). At 30 dpp, B6 XX ovaries re-mained Sox9-negative. In contrast, in B6 and B6.Ytir testisSox9-positive tissue was strongly detected in the seminif-erous tubules, whereas in B6.Ytir ovary clusters of random-ly distributed Sox9-positive tissue were evident (Fig. 7D).
DISCUSSION
Back crosses of M. musculus domesticus originated inTirano, Italy (B6.Ytir), were used to study the time courseexpression of the Sox9 gene and its product in developinggonads. About half the embryos of each litter formed ovo-testis, whereas the other half formed ovaries as expected.Although in this strain the onset of Sry expression occursnormally, it lasts longer [11], suggesting that the male-de-termining cascade is modified in some way in the genital

121Sox9 EXPRESSION IN B6.Ytir SEX-REVERSED
ridges of B6.Ytir embryos. We asked if failure to developnormal testes is at the level of Sox9, a gene that closelyfollows downstream of Sry expression and can substitutethe role of Sry for complete testis differentiation [22].
The polyclonal antibody used here was raised against thehuman SOX9 C-terminal 24 amino-acid epitope. This re-gion forms part of the transcriptional activation domain andis highly conserved between species. The specificity of theantibody to Sox9 protein was previously shown to correlatewith Sox9 mRNA by in situ hybridization in developingmice gonads [17]. At 11.5 dpc, Sox9 was immunolocalizedin the cytoplasm of several cells, suggesting that functionalactivity of Sox9 may depend on its nuclear translocation inpre-Sertoli cells. In the present study, however, Sox9 wasalways located within the nuclei of pre-Sertoli cells at 11.5and 12.5 dpc, making it unlikely that the inhibition of dif-ferentiation of pre-Sertoli to Sertoli cells in B6.Ytir gonadsis due to failure of nuclear translocation of Sox9.
In B6.Ytir gonads that take the ovarian pathway, Sertolicells expressing Sox9 protein disappeared beyond 13.5 dpc;we did detect Sox9 transcripts in B6.Ytir ovaries, but atsignificantly lower levels than in gonads that form semi-niferous cords (at 17.5 dpc). Since Sox9 transcripts inB6.Ytir ovaries are lower than in B6 or CD1 testes, ourresults indicate that maintenance of Sox9 transcripts atthreshold levels may be necessary for pre-Sertoli cells toundergo further differentiation. Although Sry can weaklyinitiate Sox9 expression in the B6.Ytir gonads at 12.5 dpc,its low activity or amount may lead to failure of Sox9-positive cells to further differentiate into Sertoli cells. SinceSertoli cells are necessary for testis tissue differentiation, itis tempting to speculate that lower levels of Sox9 transcriptsin pre-Sertoli cells lead to B6.Ytir gonads developing asphenotypic ovaries.
As in normal males, Sox9-positive cells appear in thegenital ridges of B6.Ytir males in cells considered to be thepre-Sertoli cell lineage [17]. Although this lineage appearsin 100% of 11.5 and 12.5 dpc embryos, from 13.5 dpconward the number of Sox9-positive cells diminishes anddisappears in about half of B6.Ytir embryos. This observedloss of Sox9 protein suggests that unknown factors inB6.Ytir gonads prevent further differentiation of pre-Sertolicells and lead them down the female pathway. The factorscould be genetic modifiers that are manifested when Sryand/or Sox9 function is at threshold levels.
A regional distribution of cells that express Sox9 in un-differentiated B6.Ytir gonads was observed. Sox9-positivecells appeared in the middle region, whereas they were ab-sent from the cranial and caudal regions. Furthermore, crosssections of genital ridges at 11.5 and 12.5 dpc showedSox9-positive cells among the innermost cells and rarelyamong coelomic epithelial cells. This distribution was sim-ilar in all embryos at 11.5 and 12.5 dpc, suggesting thatregional location of pre-Sertoli cells in early genital ridgesdoes not preclude their fate to differentiate as ovotestes orovaries. The observed distribution of Sox9-positive pre-Sertoli cells in B6.Ytir genital ridges is similar to that ofSry-positive cells in morphologically undifferentiated gen-ital ridges of normal XY mice [32]. Thus the pattern of Sryexpression followed by the location of Sox9-positive cellsand the organization of the seminiferous cords [33, togetherwith current results] suggests that the presence of short-range extracellular cues acting along the genital ridge dur-ing normal testis differentiation may be disturbed in B6.Ytir
gonads. Alternatively, a cell autonomous effect due to weakSry expression causing weak Sox9 transcription may occur.
Thus normal entrance of cells from the mesonephros intothe normal developing testes [34, 35] may be partially ortotally prevented in B6.Ytir gonads [36].
Furthermore, in the present study Sox9 transcripts weredetected by RT-PCR in both ovotestis and ovaries of B6.Ytir
embryos at all developmental stages. This finding suggeststhat failure to up-regulate Sox9 in pre-Sertoli cells may beposttranscriptional of either Sry, Sox9, or both. Assumingthat cells immunopositive to Sox9 detected at 11.5 and 12.5dpc in B6.Ytir genital ridges are pre-Sertoli cells, currentresults indicate that 13.5 dpc represents a critical stage forthe developmental fate of these gonads.
In B6.Ytir ovotestis, seminiferous cords comprisingSox9-positive cells gradually develop toward the cranialand caudal regions of the gonad after 14.5 dpc. Althoughscattered Sox9-positive cells remain among the sex cordsin the ovarian regions, the great abundance of pyknoticcells indicates that in most fetal ovotestes the ovarian tissueregresses. Besides lacking clear epithelial cord organiza-tion, the ovarian regions of B6.Ytir ovotestis were identifiedby the onset of meiosis of the germ cells in these areas.Thus current results support previous observations sug-gesting that postnatal ovotestes in litters of B6.Ytir mice arerare because most or all ovarian tissue regresses beforebirth [9, 37].
The levels of Sox9 transcripts at 30 dpp are lower inunilateral (ovary and testis in each side) than bilateralB6.Ytir ovaries (two ovaries), but at 60 dpp Sox9 transcriptsare undetected in both. At 30 dpp, unilateral ovaries hadmore growing follicles than bilateral B6.Ytir ovaries. By 60dpp, most follicles disappeared in all B6. Ytir ovaries (un-published results). Sox9 transcripts are absent in normalovaries. Correlating the levels of Sox9 transcripts with theovarian morphology suggested that the lower levels of Sox9transcripts observed in unilateral ovaries at 30 dpp mayfavor follicular development. Considering that infertility inB6.Ytir females occurs at the ovarian level [31, 37, 38],current results suggest that the abnormal presence of Sox9transcripts may disturb oocyte development in follicles ofB6.Ytir ovaries.
ACKNOWLEDGMENTS
We are indebted to Dr. Teruko Taketo for the kind donation of theB6.Ytir strain. We thank Jose G. Baltazar and Alejandro Marmolejo fortechnical help, and Isabel Perez Montfort for English assistance.
REFERENCES
1. Koopman P, Gubbay J, Vivian N, Goodfellow P, Lovell-Badge R.Male development of chromosomally female mice transgenic for Sry.Nature 1991; 351:117–121.
2. Hacker A, Capel B, Goodfellow P, Lovell-Badge R. Expression ofSry, the mouse sex determining gene. Development 1995; 121:1603–1614.
3. Gubbay J, Collignon J, Koopman P, Capel B, Economou A, Mun-sterberg A, Vivian N, Goodfellow P, Lovell-Badge R. A gene mappingto sex-determining region of the mouse Y chromosome is a memberof a novel family of embryonically expressed genes. Nature 1990;346:245–250.
4. Berta P, Hawkins J, Sinclair A, Taylor A, Griffiths B, Goodfellow P,Fellous M. Genetic evidence equating SRY and the testis determiningfactor. Nature 1990; 348:448–450.
5. Eicher E, Washburn L. Inherited sex reversal in mice: identificationof a new primary sex-determining gene. J Exp Zool 1983; 228:297–304.
6. Eicher E, Washburn L. Genetic control of primary sex determinationin mice. Ann Rev Genet 1986; 20:327–360.
7. Nagamine C, Taketo T, Koo G. Morphological development of the

122 MORENO-MENDOZA ET AL.
mouse gonad in tda-1 XY sex reversal. Differentiation 1987; 33:214–222.
8. Biddle F, Nishioka Y. Assays of testis development in the mouse dis-tinguish three classes of domesticus-type Y chromosome. Genome1988; 30:870–878.
9. Eicher E, Washburn L. Does one gene determine whether a C57BL/6J-Ypos mouse will develop as a female or as an hermaphrodite? JExp Zool 2001; 290:322–326.
10. Carlisle C, Winking H, Weichenhan D, Nagamine C. Absence of cor-relation between Sry polymorphisms and XY sex reversal caused byM. m. domesticus Y chromosomes. Genomics 1996; 33:32–45.
11. Lee C, Taketo T. Normal onset, but prolonged expression, of Sry genein the B6.Ydom sex-reversed mouse gonad. Dev Biol 1994; 165:442–452.
12. Nagamine C, Morohashi K, Carlisle C, Chang D. Sex reversal causedby Mus musculus domesticus Y chromosomes linked to variant ex-pression of the testis-determining gene Sry. Dev Biol 1999; 216:182–194.
13. Lee C, Taketo T. Low levels of Sry transcripts cannot be the solecause of B6-Ytir sex reversal. Genesis 2001; 30:7–11.
14. Wagner T, Wirth J, Meyer J, Zabel B, Held M, Zimmer J, Pasantes J,Bricarelli F, Keutel J, Huster E, Wolf U, Tommerup N, Schempp W,Scherer G. Autosomal sex reversal and camptomelic dysplasia arecaused by mutations in and around the SRY-related gene SOX9. Cell1994; 79:1111–1120.
15. Foster J, Dominguez M, Guioli S, Kwok C, Weller P, Weissenbach J,Mansour S, Young I, Goodfellow P, Brook J, Schafer A. Campomelicdysplasia and autosomal sex reversal caused by mutations in an SRY-related gene. Nature 1994; 372:525–530.
16. Ryner L, Swain A. Sex in the ’90s. Cell 1995; 81:483–493.17. Morais da Silva S, Hacker A, Harley V, Goodfellow P, Swain A,
Lovell-Badge R. Sox9 expression during gonadal development im-plies a conserved role for the gene in testis differentiation in mammalsand birds. Nat Genet 1996; 14:62–68.
18. Kent J, Wheatley S, Andrews J, Sinclair A, Koopman P. A male spe-cific role for SOX9 in vertebrate sex determination. Development1996; 122:2813–2822.
19. Bergstrom D, Young M, Albrecht K, Eicher E. Related function ofmouse SOX3, SOX9 and SRY HMG domains assayed by male sexdetermination. Genesis 2000; 28:11–124.
20. Graves J. Interactions between SRY and SOX9 gene in mammals sexdetermination. Bioassays 1998; 20:264–269.
21. Bishop C, Whitworth D, Qin Y, Agoulnik A, Irina Agoulnik I, Har-rison W, Behringer R, Overbeek P. A transgenic insertion upstream ofSox9 is associated with dominant XX sex reversal in the mouse. NatGenet 2000; 26:490–494.
22. Vidal P, Chaboissier M, Rooij D, Schedl A. Sox9 induces testis de-velopment in XX transgenic mice. Nat Genet 2002; 28:216–217.
23. Koopman P. Sry and Sox9: mammalian testis-determining genes. CellMol Life Sci 1999; 55:839–856.
24. Institute for Laboratory Animal Research. Guide for the Care and Useof Laboratory Animals. Bethesda, MD: National Academy of Scienc-es; 1996.
25. Alonso S, Minty A, Bourlet Y, Buckingham M. Comparison of threeactin-coding sequences in the mouse: evolutionary relationships be-tween the actin genes of warm-blooded vertebrates. J Mol Evol 1986;23:11–22.
26. Nagamine C, Chan K, Kosak C, Lau Y. Chromosome mapping andexpression of a putative testis-determining gene in mouse. Science1989; 243:80–83.
27. Chomczynski P, Sacchi N. Single-step method of RNA isolation byacid guanidinium thiocyanate-phenol-chloroform extraction. AnnBiochem 1987; 162:156–159.
28. Melton D, Konecki D, Brennand J, Caskey T. Structure expressionand mutation of the hypoxanthine phosphoribosyltransferase gene.Proc Natl Sci U S A 1984; 81:2147–2151.
29. Sambrook J, Russell D. Molecular Cloning, 3rd ed. New York: ColdSpring Harbor Laboratory Press; 2001.
30. Wilkinson D, Nieto A. Detection of messenger RNA by in situ hy-bridization to tissue sections and whole mounts. Methods Enzymol1993; 225:361–373.
31. Eicher E, Washburn L, Whitney J, Morrow K. Mus poschiavinus Ychromosome in the C57Bl/6J murine genome causes sex reversal. Sci-ence 1982; 217:535–537.
32. Bullejos M, Koopman P. Spatially dynamic expression of Sry inmouse genital ridges. Dev Dynamics 2001; 221:201–205.
33. Kenneth H, Eicher E. Evidence that Sry is expressed in pre-Sertolicells and Sertoli and granulosa cells have a common precursor. DevBiol 2001; 240:92–107.
34. Buehr M, Gu S, McLaren A. Mesonephric contribution to testis dif-ferentiation in the fetal mouse. Development 1993; 117:273–281.
35. Merchant-Larios H, Moreno-Mendoza N. Mesonephric stromal cellsdifferentiate into Leydig cells in the mouse fetal testis. Exp Cell Res1998; 244:230–238.
36. Kenneth A, Capel B, Washburn L, Eicher E. Defective mesonephriccell migration is associated with abnormal testis cord development inC57BL/6J XY Mus domesticus mice. Dev Biol 2000; 225:26–36.
37. Taketo T, Nishioka Y, Nagamine C, Villalpando I, Merchant H. De-velopment and fertility of ovaries in the B6. Ydom sex-reversed femalemouse. Development 1989; 107:95–105.
38. Marmolejo A, Nishioka Y, Moreno N, Merchant H. Fertility of Ytir.B6sex-reversal females with XX orthotopic ovarian transplants. Biol Re-prod 1999; 61:1426–1430.




















