
The Ig light chain restricted B6.\u0026#x003BA; - \u0026#x003BB; SEG mouse strain suggests that the...
14
Abstract In laboratory mice, the different Ig λ light chain subtypes (λ1, λ2, λx and λ3) are expressed on 60, 16, 16 and 8%, respectively, of the λ-positive peripheral B cells. Eighteen years ago, our laboratory characterized a λ1 – wild mouse strain: SPE (Mus spretus). In this report, we describe the characterization of another wild- derived Mus spretus inbred strain, SEG, that presents the same characteristic, namely the absence of λ1 expression. An almost congenic strain, B6.λ SEG , was detected in a series of recombinant congenic strains carrying 2% of SEG/Pas genome in a C57BL/6J background. This B6.λ SEG strain was crossed to Igh a Cκ – mice in order to derive two different additional congenic strains: B6.κ – λ SEG Igh a and B6.κ – λ SEG Igh b . In this paper, we characterize the genomic organization and the expression of the SEG IGL locus. Altogether, our data show that the SEG IGL locus is constituted by a single functional IGLJ2SEG-IGLC2SEG, two pseudo IGLJ4SEG1/2- IGLC4SEG1/2 gene clusters and two V gene segments: IGLV2SEG and IGLVXSEG. In particular, we show the absence of IGLV1 and IGLVSD26 gene segments. IGLVSD26 was reported to be present in some Mus m. musculus mice and absent in BALB/c. Here, we confirm its presence not only in other Mus m. musculus mice but also in Mus spretus mice. Consequently, we propose that IGLVSD26-related gene segments define a new family that we name Vλ4. The study of the organization of different IGL loci, in addition to the Vλ4 + reported here, could elucidate questions concerning the evolution of the λ locus. Keywords Wild mice · Lambda · Immunoglobulin · Evolution Introduction Among mammals, two immunoglobulin (Ig) light chain isotypes have been described, κ and λ. The proportion between these two isotypes can vary significantly at the serum and B-cell levels (95/5 in mouse, 60/40 in human, 5/95 in horse, almost 100% λ for chicken) (Butler 1997; Eisen and Reilly 1985). In laboratory mice, the organi- zation of the IGL locus has been shown to be very different from that of the IGK locus. The IGL locus shows a clustered organization with four J-C (including one pseudo) clusters divided in two recombination units that can produce four λ chains, namely: λ1, λ2, λ3 and λx, respectively, encoded by IGLV1-LJ1-LC1, IGLV2- LJ2-LC2, IGLV1-LJ3-LC3 and IGLVX-LJ2-LC2 combi- nations (Blomberg and Tonegawa 1982; Sanchez and Cazenave 1987; Sanchez et al. 1996; Storb et al. 1989). This organization was considered to be of an archaic type, close to that of the shark (Hinds and Litman 1986; Litman et al. 1993). The IGK locus shows another type of organization with a single IGCK, five IGJK (one pseudo) and at least 140 IGVK, giving rise to a more diverse repertoire of κ chains (Kirschbaum et al. 1998; Thiebe et al. 1999). The question of the influence of the IGL locus on the generation of B cells, Ig diversity and responses to different antigens (Ag) has already been addressed by us and others in normal laboratory and κ-deficient mice (Bot et al. 1996; Chen et al. 1993; Pricop et al. 1994; Sanchez et al. 1994, 1996; Zou et al. 1993). The targeted disruption of the IGCK gene can introduce a tremendous bias in the mouse Ig light chain repertoire by precluding the production of κ light chains. Consequently, the proportion of peripheral B-cell repertoire in the spleen becomes 60, 16, 16 and 8% for λ1, λ2, λx and λ3 positive B cells, respectively (Sanchez et al. 1999). This situation leads to a strong reduction of the combinatorial diversity Y.M. Amrani · D. Voegtlé · P.-A. Cazenave · A. Six ( ✉ ) Unité d’Immunophysiopathologie infectieuse, Institut Pasteur, CNRS URA 1961, Université Pierre et Marie Curie, Paris 6, Institut Pasteur, 25 rue du Docteur Roux, 75015 Paris, France e-mail: [email protected] Tel.: +33-1-45688581, Fax: +33-1-40613066 X. Montagutelli Unité de Génétique des Mammifères, Institut Pasteur, CNRS URA 1960, Institut Pasteur, 25 rue du Docteur Roux, 75015 Paris, France Immunogenetics (2002) 54:106–119 DOI 10.1007/s00251-002-0448-6 ORIGINAL PAPER Yacine M. Amrani · Danielle Voegtlé Xavier Montagutelli · Pierre-André Cazenave Adrien Six The Ig light chain restricted B6.κ - λ SEG mouse strain suggests that the IGL locus genomic organization is subject to constant evolution Received: 7 January 2002 / Revised: 12 February 2002 / Published online: 5 April 2002 © Springer-Verlag 2002
-
Upload
sorbonne-fr -
Category
Documents
-
view
2 -
download
0
Transcript of The Ig light chain restricted B6.\u0026#x003BA; - \u0026#x003BB; SEG mouse strain suggests that the...

Abstract In laboratory mice, the different Ig λ lightchain subtypes (λ1, λ2, λx and λ3) are expressed on 60,16, 16 and 8%, respectively, of the λ-positive peripheralB cells. Eighteen years ago, our laboratory characterizeda λ1– wild mouse strain: SPE (Mus spretus). In thisreport, we describe the characterization of another wild-derived Mus spretus inbred strain, SEG, that presents thesame characteristic, namely the absence of λ1 expression.An almost congenic strain, B6.λSEG, was detected in aseries of recombinant congenic strains carrying 2% ofSEG/Pas genome in a C57BL/6J background. ThisB6.λSEG strain was crossed to Igha Cκ– mice in order toderive two different additional congenic strains:B6.κ–λSEG Igha and B6.κ–λSEG Ighb. In this paper, wecharacterize the genomic organization and the expressionof the SEG IGL locus. Altogether, our data show that theSEG IGL locus is constituted by a single functionalIGLJ2SEG-IGLC2SEG, two pseudo IGLJ4SEG1/2-IGLC4SEG1/2 gene clusters and two V gene segments:IGLV2SEG and IGLVXSEG. In particular, we showthe absence of IGLV1 and IGLVSD26 gene segments.IGLVSD26 was reported to be present in some Mus m.musculus mice and absent in BALB/c. Here, we confirmits presence not only in other Mus m. musculus mice butalso in Mus spretus mice. Consequently, we propose thatIGLVSD26-related gene segments define a new familythat we name Vλ4. The study of the organization ofdifferent IGL loci, in addition to the Vλ4+ reported here,could elucidate questions concerning the evolution of theλ locus.
Keywords Wild mice · Lambda · Immunoglobulin ·Evolution
Introduction
Among mammals, two immunoglobulin (Ig) light chainisotypes have been described, κ and λ. The proportionbetween these two isotypes can vary significantly at theserum and B-cell levels (95/5 in mouse, 60/40 in human,5/95 in horse, almost 100% λ for chicken) (Butler 1997;Eisen and Reilly 1985). In laboratory mice, the organi-zation of the IGL locus has been shown to be verydifferent from that of the IGK locus. The IGL locusshows a clustered organization with four J-C (includingone pseudo) clusters divided in two recombination unitsthat can produce four λ chains, namely: λ1, λ2, λ3 andλx, respectively, encoded by IGLV1-LJ1-LC1, IGLV2-LJ2-LC2, IGLV1-LJ3-LC3 and IGLVX-LJ2-LC2 combi-nations (Blomberg and Tonegawa 1982; Sanchez andCazenave 1987; Sanchez et al. 1996; Storb et al. 1989).This organization was considered to be of an archaictype, close to that of the shark (Hinds and Litman 1986;Litman et al. 1993). The IGK locus shows another typeof organization with a single IGCK, five IGJK (onepseudo) and at least 140 IGVK, giving rise to a morediverse repertoire of κ chains (Kirschbaum et al. 1998;Thiebe et al. 1999).
The question of the influence of the IGL locus on thegeneration of B cells, Ig diversity and responses todifferent antigens (Ag) has already been addressed by usand others in normal laboratory and κ-deficient mice(Bot et al. 1996; Chen et al. 1993; Pricop et al. 1994;Sanchez et al. 1994, 1996; Zou et al. 1993). The targeteddisruption of the IGCK gene can introduce a tremendousbias in the mouse Ig light chain repertoire by precludingthe production of κ light chains. Consequently, theproportion of peripheral B-cell repertoire in the spleenbecomes 60, 16, 16 and 8% for λ1, λ2, λx and λ3 positiveB cells, respectively (Sanchez et al. 1999). This situationleads to a strong reduction of the combinatorial diversity
Y.M. Amrani · D. Voegtlé · P.-A. Cazenave · A. Six (✉ )Unité d’Immunophysiopathologie infectieuse, Institut Pasteur,CNRS URA 1961, Université Pierre et Marie Curie, Paris 6,Institut Pasteur, 25 rue du Docteur Roux, 75015 Paris, Francee-mail: [email protected].: +33-1-45688581, Fax: +33-1-40613066
X. MontagutelliUnité de Génétique des Mammifères, Institut Pasteur,CNRS URA 1960, Institut Pasteur, 25 rue du Docteur Roux,75015 Paris, France
Immunogenetics (2002) 54:106–119DOI 10.1007/s00251-002-0448-6
O R I G I N A L PA P E R
Yacine M. Amrani · Danielle VoegtléXavier Montagutelli · Pierre-André CazenaveAdrien Six
The Ig light chain restricted B6.κ-λSEG mouse strain suggests thatthe IGL locus genomic organization is subject to constant evolution
Received: 7 January 2002 / Revised: 12 February 2002 / Published online: 5 April 2002© Springer-Verlag 2002

107
of the light chain repertoire and consequently of the Igrepertoire (Sanchez et al. 1996).
To see the influence of a more reduced Ig light chaingenetic repertoire on the generation of diversity andresponse capacities to Ag, we took advantage of wildmouse strains “naturally” presenting a restricted λ reper-toire when compared to BALB/c or C57BL/6 (B6) mice.The Mus spretus SPE mouse lacks λ1 and probably λ3serum Ig (Amor et al. 1983). In addition to IGK genes,this strain is considered to possess a single IGL recombi-nation unit comprising IGLV2SPE and two IGLJ-LCclusters (Kindt et al. 1985; Mami and Kindt 1987a, b).Unfortunately, this strain was lost. Therefore, we studiedanother M. spretus SEG/Pas strain, which presentscharacteristics close to those of SPE, namely the absenceof λ1 and λ3 chain expression. SEG/Pas was crossedwith C57BL/6J mice to produce a series of interspecificrecombinant congenic strains (IRCS) carrying 2% ofSEG/Pas genome in a C57BL/6J background (unpublisheddata). One of these strains was shown to carry theSEG/Pas allele at the IGL locus and was named B6.λSEG.It was later crossed to Igha Cκ– mice to obtain B6.κ–λSEG
mice of both Igha and Ighb haplotypes.Since we wanted to use this strain as being deficient
for some of the λ genes, we first characterized the SEGIGL locus and showed that it comprises a single IGLV2-LVX-LJ2-LC2-LJ41-LC41-LJ42-LC42 recombination unit.Sequence analysis of the genes revealed that only λ2(IGLV2-LJ2-LC2) and λx (IGLVX-LJ2-LC2) chains canbe produced, in agreement with our previous observa-tions.
During our characterization, we showed the absenceof IGLVSD26 from the SEG IGL locus. This additionalIGLV gene segment was first described in two Mus m.musculus mice from Skive (Denmark) and Sladeckovce(Czech Republic) in association with a related IGLC1gene (Reidl et al. 1992). Interestingly, we could identifyIGLVSD26-related genes in some mouse strains belongingto M. m. musculus and North African M. spretus species,but not in Mus m. domesticus mice from which laboratorymice are derived. A correlation between the presence ofIGLVSD26 and IGL gene numbers has been establishedafter RFLP data analysis and comparison to other speciesthat sheds some light on the evolution of the IGL locus.
Materials and methods
Mouse strains used in this study
C57BL/6N mice were purchased from Charles River Laboratory(Saint Aubin les Elbeuf, France) and then maintained in our ani-mal facility. B6.κ–, 129/Sv.κ– (Sanchez et al. 1994) and B10.D2.κ–
mice were produced and bred in the Unité d’Immunochimie Anal-ytique. The B6.λSEG mouse strain is one from the IRCS developedindependently from this study (unpublished data). These strainswere obtained by backcrossing the M. spretus SEG/Pas strain fourtimes onto C57BL/6J, followed by inbreeding for over 20 genera-tions, without any selection. One of the resulting strains (laterrenamed B6.λSEG) was found to carry the SEG allele at the IGLlocus and was used in the present study. This is not strictly a con-genic strain but genome-wide genotyping has shown that this
strain has inherited less than 2% of its genome from the SEG/Passtrain. B6.κ–λSEG mice were obtained by crossing the Igha Cκ– andB6.λSEG mice and selected for the absence of λ1 by MS40–13 Abstaining (Amor et al. 1983). Both Igha and Ighb strains wereobtained.
T15i mice were a generous gift from Dr. K. Rajewsky(Cologne, Germany) (Taki et al. 1993). They are maintained in ouranimal facility at the Pasteur Institute.
We also used a series of wild-derived mice including M. m.musculus species: MBT, MBB, MBK (extinct strain), MBS, MBVfrom Bulgaria; MAI, MPW and PWK from Austria, Poland andthe Czech Republic, respectively; M. m. domesticus species: ULA,WLA, WGQ from France; BIK/g, 38CH and WMP from Israel,Italy and Tunisia, respectively; M. spretus species: SMZ (a generousgift of Dr. F. Bonhomme, University of Montpellier, France) andSMA from Morocco; SEG and SPR/Ei from Spain, SFM fromFrance and STF from Tunisia.
DNA preparation and PCR
Genomic DNA was purified from the tails of all mice used in thisstudy (see above) after overnight proteinase K digestion (150 mMNaCl, 100 mM EDTA, 0.5% SDS and 160 µg/ml Proteinase K,pH 8), followed by phenol/chloroform extraction and ethanolprecipitation (Sambrook et al. 1989).
PCR was carried out with 1/50th of the DNA preparation, 1.5or 2.5 mM MgCl2, 1 unit Taq polymerase (Promega, Madison,Wis., USA), 10 mM dNTPs and 0.6 µM of each primer in 50 µl of1× buffer provided by the manufacturer. The PCR program was:94°C for 1 min, followed by 30 cycles of 94°C for 1 min, 60°C for1 min, 72°C for 2 min, and then 72°C for 10 min. In some experi-ments, the Hot Star Taq polymerase (Qiagen, Courtaboeuf,France) was used with the following program: 94°C for 15 min,followed by 30 cycles of 94°C for 1 min, 60°C for 1 min, 72°C for2 min, and then 72°C for 10 min. Primers (20 bp length) are pre-sented in Fig. 2 (lower case) or can be found in our GenBank sub-mitted sequences (see legends to Figs. 2 and 5 for accession num-bers). Amplified DNA fragments were detected using ethidiumbromide staining after electrophoretic migration on 1.5% UltraPure agarose gel (Gibco BRL, Gaithersburg, Md., USA) in 1×TBEbuffer. Fragment length was estimated comparatively to the100 bp ladder (Pharmacia Biotech, Uppsala, Sweden).
Cloning and sequencing
Amplified DNA of interest was cloned immediately after PCR asfollows, according to the purity of amplification products: PCRproducts were either quantified with φx174 RF DNA/HaeIII frag-ments (Gibco BRL) and cloned directly with the TA Cloning kit(Invitrogen, Carlsbad, Calif., USA) or purified on LMP agarosegels (Gibco BRL) and cloned with the TOPO TA cloning kit(Invitrogen).
After bacterial transformation, plasmid DNA from randomlypicked white colonies was purified with the QIAwell 8 PLUSPlasmid kit (Qiagen). Presence of the insert was verified afterEcoRI digestion (Gibco BRL) and electrophoresis on a 2% agarosegel.
For each primer combination, four clones of two independentPCRs were sequenced using Universal (-40) primer (GTTTTCC-CAGTCACGAC) and Reverse primer (CAGGAAACAGTAT-GAC) by PCR with ddNTPs following the recommendations forthe Prism Dye Terminator Cycle Sequencing Ready Reaction kit(Applied Biosystem, PE Biosystems, Warrington, England). Asecond pair of internal primers was used to sequence theIGLV2SEG gene segment. PCR products were then purified, dena-tured by heating and addition of an equal volume of formamide-EDTA in blue dextran. Samples were loaded on a 6% acrylamidegel to be separated by electrophoresis on an ABI 373 automaticsequencer (ABI). The fragments were analyzed with sequencinganalysis 3.0 (ABI). The sequences were compared to a library ofIGL genes and gene segments in order to determine the orientation

108
and identity of the cloned inserts. The sequences were thenaligned with the reference sequences, when available, with theDNA sequence analysis package (CALTECH). For phylogeneticanalysis, Clustal X (EMBL, Heidelberg, Germany) and Phylip3.5c (J. Felsenstein, University of Washington) packages wereused. Phylogenetic trees were visualized with TreeView 1.6.5(Page 1996).
Fluorescence activated cell sorting
Peripheral blood lymphocytes (PBLs) from B6.κ–λSEG Igha and129/Sv.κ– mice were obtained after eye bleeding and ficoll separa-tion. PBLs were washed twice, 50 µl of 2–4.G2 hybridoma culturesupernatant was added for 30 min at 4°C to block Fc receptors.PBLs were stained first with 189-B3 (anti-λ2) (Sanchez et al.1991) for 15 min at 4°C conjugated with biotin, washed twice andthen stained with DS-1 (anti-Igh6a) conjugated with FITC, plusstreptavidin-PE (Pharmingen, San Diego, Calif., USA). PBLswere washed twice prior to analysis on a Becton-DickinsonFACScan.
RFLP analysis
High-molecular-weight germline DNA (12–15 µg) from B6,B6.λSEG Ighb, MAI, MBK, PWK, STF and SMZ mouse strainswas digested with either EcoRI, HindIII or BamHI overnight in afinal volume of 80 µl at 37°C. Digestion samples and λ/HindIIImarker were loaded on 0.7% agarose gels in 1×TAE buffer andrun for 36 h at 24 V after 5 min at 80 V. DNA in gels was depuri-nated, denatured and neutralized. DNA was transferred undervacuum onto nylon membranes (Qiabra, Nylon Plus, Qiagen), andfixed under UV light. Membranes were prehybridized overnight at65°C. Hybridizations were carried out with the appropriate32[P]dCTP radiolabeled probe in hybridization buffer overnight at65°C. Final washes of membranes were carried out with 3×SSC,0.05% SDS buffer, 10 min, 1×SSC, 0.05% SDS, 10 min, and0.2×SSC, 0.05% SDS at 65°C (high stringency) or 50°C (lowstringency) for 30–45 min (Southern 1975).
Probes used for hybridization
Vλ2SEG, VλxSEG, VλSD26, Cλ2SEG or Cλ4SEGB probes werederived from plasmids used to sequence the respective genesegments and genes following EcoRI digestion, migration on 1.2%agarose gel in modified 1×TAE buffer, band excision and purifica-tion on Ultrafree-DA columns (Millipore, Bedford, Mass., USA).Probes were radiolabeled using the Ready-to-go DNA LabelingBeads kit (dCTP) (Amersham Pharmacia, Piscataway, N.J., USA)overnight at 65°C.
Results
Nomenclature of λ genes and gene segments analyzedin this study
The reference sequences for laboratory mouse λ lightchain genes and gene segments are those from theBALB/c strain and we refer to them as IGLV1 (Vλ1),IGLV2 (Vλ2), IGLVX (Vλx), IGLJ1 (Jλ1), IGLJ2 (Jλ2),IGLJ3 (Jλ3), IGLJ4 (Jλ4), IGLC1 (Cλ1), IGLC2 (Cλ2),IGLC3 (Cλ3) and IGLC4 (Cλ4) (Arp et al. 1982;Blomberg and Tonegawa 1982; Sanchez et al. 1990;Selsing et al. 1982; Tonegawa et al. 1978). In addition,genomic sequences from the M. spretus SPE mousestrain are referred to as IGLV2SPE (Vλ2SPE), IGLJ2SPE
(Jλ2SPE), IGLJ4SPE (Jλ4SPE), IGLC2SPE (Cλ2SPE), andIGLC4SPE (Cλ4SPE) (Mami and Kindt 1987a, b).
The sequences to which we refer as IGLVSD26(VλSD26) and IGLVCZ81 (VλCZ81) are derived fromthe variable regions of SD26 and CZ81 hybridoma Iglight chain cDNAs, obtained from M. m. musculus micefrom Skive, Denmark and Sladeckovce, Czech Republic,respectively. An IGLV2SPE and an IGLC1 homologuehave also been described in the Czech and Danish miceand are derived from the Ig light chain variable region ofthe CZ72 hybridoma and the constant region of the Iglight chain hybridoma SD26 cDNAs, respectively (Reidlet al. 1992).
SEG IGL locus characterization
PCR and sequence analysis
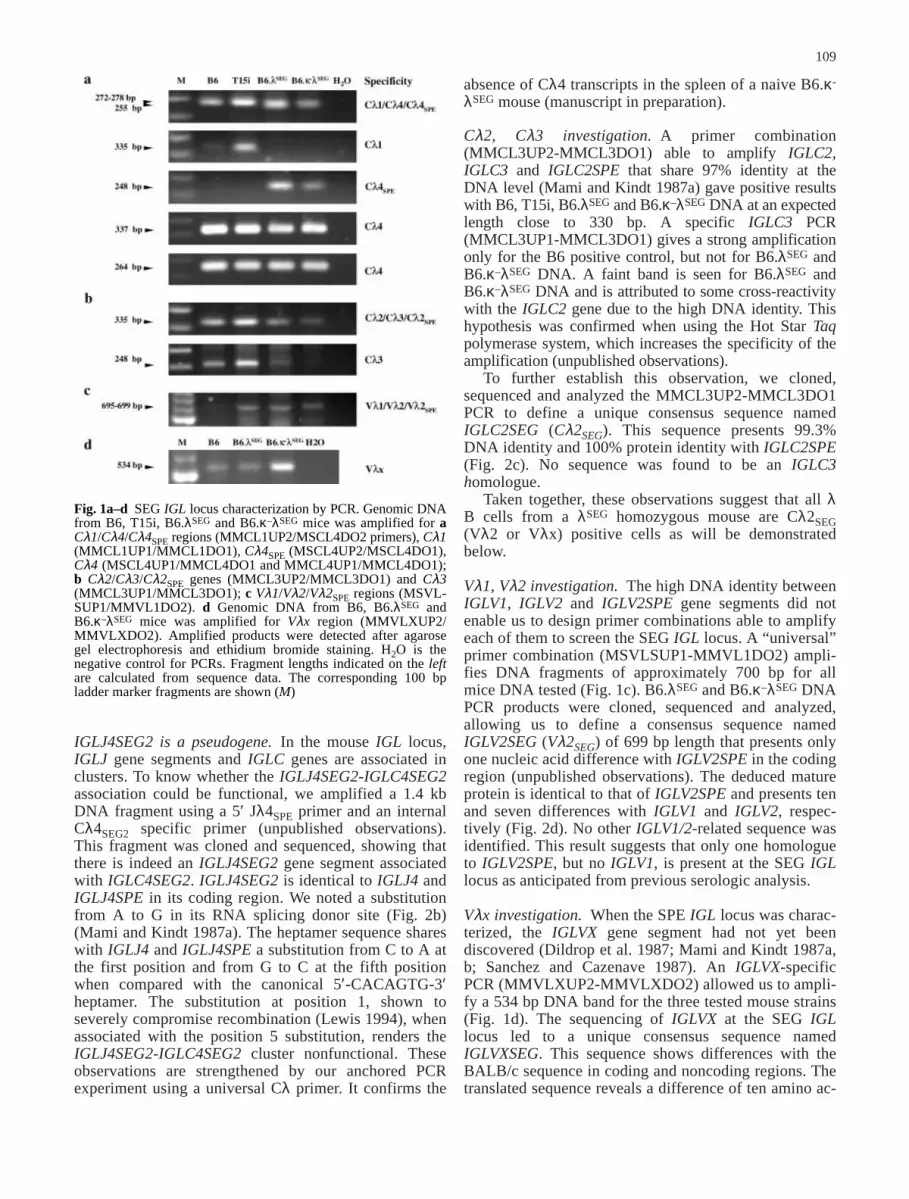
Cλ1, Cλ4 investigation. On the basis of availablesequence alignments, we have designed and used primercombinations able to amplify the IGLC1, IGLC4 andIGLC4SPE genes (MMCL1UP2-MSCL4DO2). B6, T15i,B6.λSEG and B6.κ–λSEG genomic DNAs show intenseamplification fragments at 250–270 bp, the expectedlengths being at 278, 272 and 255 bp, respectively, forIGLC1, IGLC4 and IGLC4SPE. The absence of IGLC1from the SEG IGL locus is suggested by the IGLC1specific PCR (MMCL1UP1-MMCL1DO1), which isnegative for B6.λSEG and B6.κ-λSEG DNA and positivefor both B6 and T15i control DNAs. A specificIGLC4SPE PCR (MSCL4UP2-MSCL4DO1) was positiveonly for B6.λSEG and B6.κ-λSEG. When using two combi-nations (MSCL4UP1-MMCL4DO1 and MMCL4UP1-MMCL4DO1) able to amplify IGLC4 but not IGLC4SPEor IGLC1, a positive result was observed for all theDNAs (Fig. 1a). The above data suggest that the SEG IGLlocus contains at least two IGLC4 gene homologues. Thecloning and sequencing of the MMCL1UP2-MSCL4DO2,MSCL4UP1-MMCL4DO1 and MMCL4UP1-MMCL4DO1PCR experiments confirmed this hypothesis and identi-fied two consensus sequences, IGLC4SEG1 (Cλ4SEG1)and IGLC4SEG1 (Cλ4SEG2). IGLC4SEG1 shows morethan 99% DNA identity with IGLC4SPE and is also apseudogene due to a 14 bp gap and stop codons in allthree reading frames; it presents 92% DNA identity withIGLC4 (Mami and Kindt 1987a). IGLC4SEG1 is associ-ated with a IGLJ4SEG1 (Jλ4SEG1) gene segment asdemonstrated by the amplification of a 1.3 kb or 1.5 kbfragment when using an internal Cλ4SEG1 specific primerwith an internal or an upstream Jλ4 primer, respectively(unpublished observations). IGLC4SEG2 presents 97.5%and 91% DNA identity with IGLC4 and IGLC4SPE,respectively. IGLC4SEG2 shares a 6 bp gap in the codingregion with IGLC4 and is associated with an IGLJ4SEG2(Jλ4SEG2) pseudogene segment (Blomberg and Tonegawa1982; Mami and Kindt 1987a; Miller et al. 1982). Its fullsequence has been obtained after the sequencing of theJλ4SEG2-Cλ4SEG2 fragment (see below).
absence of Cλ4 transcripts in the spleen of a naive B6.κ-
λSEG mouse (manuscript in preparation).
Cλ2, Cλ3 investigation. A primer combination(MMCL3UP2-MMCL3DO1) able to amplify IGLC2,IGLC3 and IGLC2SPE that share 97% identity at theDNA level (Mami and Kindt 1987a) gave positive resultswith B6, T15i, B6.λSEG and B6.κ–λSEG DNA at an expectedlength close to 330 bp. A specific IGLC3 PCR(MMCL3UP1-MMCL3DO1) gives a strong amplificationonly for the B6 positive control, but not for B6.λSEG andB6.κ–λSEG DNA. A faint band is seen for B6.λSEG andB6.κ–λSEG DNA and is attributed to some cross-reactivitywith the IGLC2 gene due to the high DNA identity. Thishypothesis was confirmed when using the Hot Star Taqpolymerase system, which increases the specificity of theamplification (unpublished observations).
To further establish this observation, we cloned,sequenced and analyzed the MMCL3UP2-MMCL3DO1PCR to define a unique consensus sequence namedIGLC2SEG (Cλ2SEG). This sequence presents 99.3%DNA identity and 100% protein identity with IGLC2SPE(Fig. 2c). No sequence was found to be an IGLC3homologue.
Taken together, these observations suggest that all λB cells from a λSEG homozygous mouse are Cλ2SEG(Vλ2 or Vλx) positive cells as will be demonstratedbelow.
Vλ1, Vλ2 investigation. The high DNA identity betweenIGLV1, IGLV2 and IGLV2SPE gene segments did notenable us to design primer combinations able to amplifyeach of them to screen the SEG IGL locus. A “universal”primer combination (MSVLSUP1-MMVL1DO2) ampli-fies DNA fragments of approximately 700 bp for allmice DNA tested (Fig. 1c). B6.λSEG and B6.κ–λSEG DNAPCR products were cloned, sequenced and analyzed,allowing us to define a consensus sequence namedIGLV2SEG (Vλ2SEG) of 699 bp length that presents onlyone nucleic acid difference with IGLV2SPE in the codingregion (unpublished observations). The deduced matureprotein is identical to that of IGLV2SPE and presents tenand seven differences with IGLV1 and IGLV2, respec-tively (Fig. 2d). No other IGLV1/2-related sequence wasidentified. This result suggests that only one homologueto IGLV2SPE, but no IGLV1, is present at the SEG IGLlocus as anticipated from previous serologic analysis.
Vλx investigation. When the SPE IGL locus was charac-terized, the IGLVX gene segment had not yet beendiscovered (Dildrop et al. 1987; Mami and Kindt 1987a,b; Sanchez and Cazenave 1987). An IGLVX-specificPCR (MMVLXUP2-MMVLXDO2) allowed us to ampli-fy a 534 bp DNA band for the three tested mouse strains(Fig. 1d). The sequencing of IGLVX at the SEG IGLlocus led to a unique consensus sequence namedIGLVXSEG. This sequence shows differences with theBALB/c sequence in coding and noncoding regions. Thetranslated sequence reveals a difference of ten amino ac-
109
IGLJ4SEG2 is a pseudogene. In the mouse IGL locus,IGLJ gene segments and IGLC genes are associated inclusters. To know whether the IGLJ4SEG2-IGLC4SEG2association could be functional, we amplified a 1.4 kbDNA fragment using a 5′ Jλ4SPE primer and an internalCλ4SEG2 specific primer (unpublished observations).This fragment was cloned and sequenced, showing thatthere is indeed an IGLJ4SEG2 gene segment associatedwith IGLC4SEG2. IGLJ4SEG2 is identical to IGLJ4 andIGLJ4SPE in its coding region. We noted a substitutionfrom A to G in its RNA splicing donor site (Fig. 2b)(Mami and Kindt 1987a). The heptamer sequence shareswith IGLJ4 and IGLJ4SPE a substitution from C to A atthe first position and from G to C at the fifth positionwhen compared with the canonical 5′-CACAGTG-3′heptamer. The substitution at position 1, shown toseverely compromise recombination (Lewis 1994), whenassociated with the position 5 substitution, renders theIGLJ4SEG2-IGLC4SEG2 cluster nonfunctional. Theseobservations are strengthened by our anchored PCRexperiment using a universal Cλ primer. It confirms the
Fig. 1a–d SEG IGL locus characterization by PCR. Genomic DNAfrom B6, T15i, B6.λSEG and B6.κ–λSEG mice was amplified for aCλ1/Cλ4/Cλ4SPE regions (MMCL1UP2/MSCL4DO2 primers), Cλ1(MMCL1UP1/MMCL1DO1), Cλ4SPE (MSCL4UP2/MSCL4DO1),Cλ4 (MSCL4UP1/MMCL4DO1 and MMCL4UP1/MMCL4DO1);b Cλ2/Cλ3/Cλ2SPE genes (MMCL3UP2/MMCL3DO1) and Cλ3(MMCL3UP1/MMCL3DO1); c Vλ1/Vλ2/Vλ2SPE regions (MSVL-SUP1/MMVL1DO2). d Genomic DNA from B6, B6.λSEG andB6.κ–λSEG mice was amplified for Vλx region (MMVLXUP2/MMVLXDO2). Amplified products were detected after agarosegel electrophoresis and ethidium bromide staining. H2O is thenegative control for PCRs. Fragment lengths indicated on the leftare calculated from sequence data. The corresponding 100 bpladder marker fragments are shown (M)

110
ids, four differences being located in the CDR3 (Fig. 2e).However, IGLVX still exhibits Vλx family characteris-tics, namely, a four-amino-acid longer CDR3 and areduced junctional diversity potential due to the presence
of a stop codon at the end of its coding sequence(Sanchez et al. 1990).
Taken together, the above results lead us to concludethat the SEG IGL locus comprises a single functional
Fig. 2a–e Gene and gene segment sequence comparisons.a Cλ4SEG1, Cλ4SEG2, Cλ4SPE, Cλ4 and Cλ1; Cλ4SEG1 and Cλ4SEG2gene sequences are compared to Cλ4SPE of Mus spretus SPEmouse, Cλ4 and Cλ1 of BALB/c mouse gene sequences (Gen-Bank accession nos.: AF357972, AF357974, M16628, J00598 andJ00587). Cλ4SEG2 sequence is deduced from sequence data con-secutive to Cλ4 and Jλ4-Cλ4 PCR experiments. b Jλ4SEG2,Jλ4SPE, and Jλ4; Jλ4SEG1 segment gene sequence is compared toJλ4SPE of Mus spretus SPE mouse, Jλ4 and Jλ1 of BALB/c mousegene segment sequences (GenBank accession nos.: AF357974,M16555, J00596 and X58419). c Cλ2SEG, Cλ2SPE, Cλ2 and Cλ3;Cλ2SEG gene sequence is compared to Cλ2SPE of M. spretus SPEmouse, Cλ2 and Cλ3 of BALB/c mouse gene sequences(GenBank accession nos.: AF357973, M16554, J00595 andJ00585). d Protein sequence comparisons of Vλ2SEG, Vλ2SPE, Vλ2
and Vλ1; Vλ2SEG protein sequence is compared to Vλ2SPE fromM. spretus SPE mouse, Vλ2 and Vλ1 from BALB/c mouse proteinsequences (GenBank accession nos.: AF357989, M17529, J00599and J00590). e VλxSEG and Vλx; VλxSEG gene sequence iscompared to Vλx from BALB/c mouse gene sequences (GenBankaccession nos.: AF357988 and M34597). All protein sequencesare translated from the respective GenBank nucleotide sequences.Primer regions are specified in lower case, dashes indicatesequence identity, slashes indicate deletions/insertions. Boldcharacters indicate the J nonamer and heptamer whereas the RNAsplicing site is underlined. Stars indicate stop codons and strokes,the beginning of the coding sequence or the boundaries of thedifferent domains. The locations of the leader, mature peptidesegments, FR and CDR are indicated according to Kabat andco-workers (1991)
111
IGLC2SEG, two pseudo IGLC4SEG1 and IGLC4SEG2genes, and only two functional IGLV gene segments,IGLV2SEG and IGLVXSEG. In order to confirm thisorganization, we performed FACS, SDS-PAGE of IgGsand RFLP studies for both B6 and B6.κ-λSEG. The resultsare presented below.
B6.κ–λSEG B cells express onlyCλ2SEG
+(Vλ2SEG/VλxSEG–Cλ2SEG) B-cell receptorsin the periphery
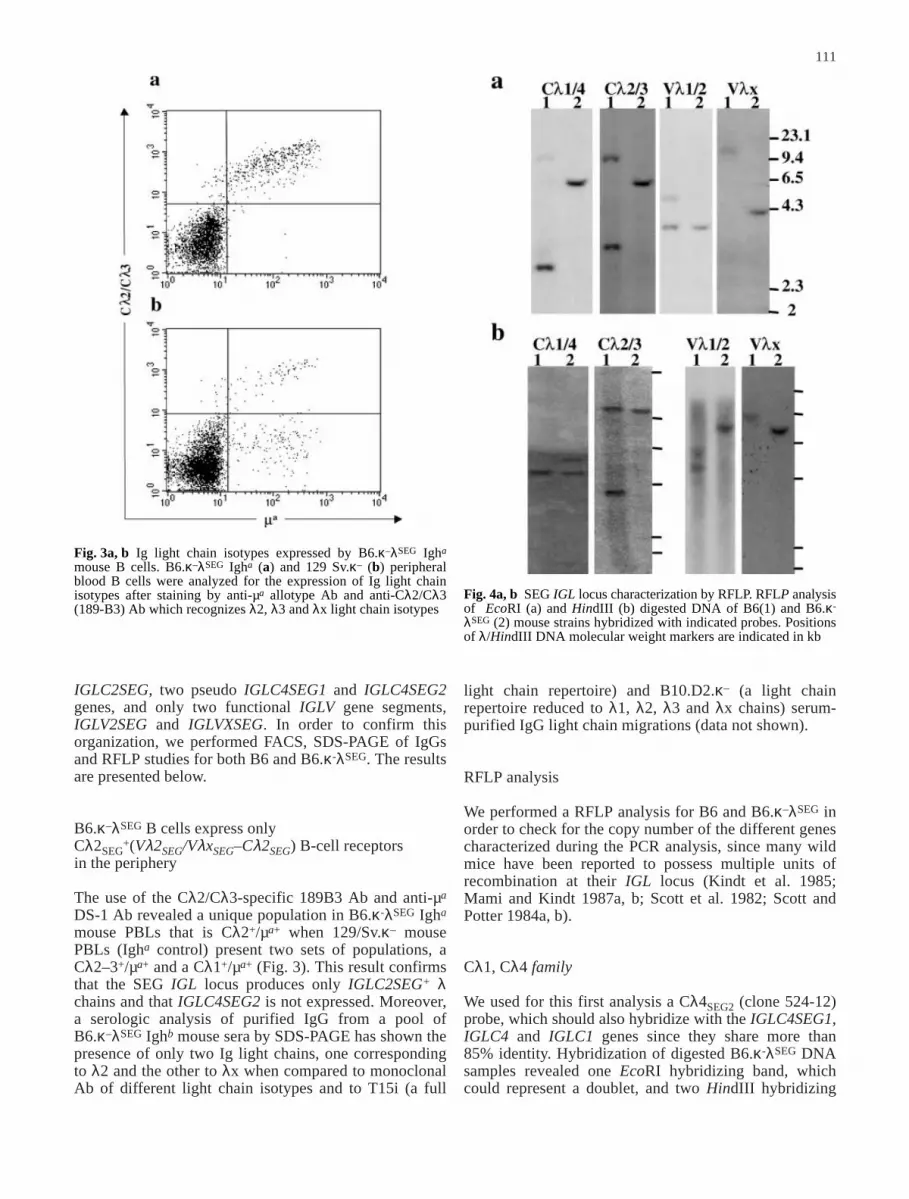
The use of the Cλ2/Cλ3-specific 189B3 Ab and anti-µa
DS-1 Ab revealed a unique population in B6.κ-λSEG Igha
mouse PBLs that is Cλ2+/µa+ when 129/Sv.κ– mousePBLs (Igha control) present two sets of populations, aCλ2–3+/µa+ and a Cλ1+/µa+ (Fig. 3). This result confirmsthat the SEG IGL locus produces only IGLC2SEG+ λchains and that IGLC4SEG2 is not expressed. Moreover,a serologic analysis of purified IgG from a pool ofB6.κ–λSEG Ighb mouse sera by SDS-PAGE has shown thepresence of only two Ig light chains, one correspondingto λ2 and the other to λx when compared to monoclonalAb of different light chain isotypes and to T15i (a full
light chain repertoire) and B10.D2.κ– (a light chainrepertoire reduced to λ1, λ2, λ3 and λx chains) serum-purified IgG light chain migrations (data not shown).
RFLP analysis
We performed a RFLP analysis for B6 and B6.κ–λSEG inorder to check for the copy number of the different genescharacterized during the PCR analysis, since many wildmice have been reported to possess multiple units ofrecombination at their IGL locus (Kindt et al. 1985;Mami and Kindt 1987a, b; Scott et al. 1982; Scott andPotter 1984a, b).
Cλ1, Cλ4 family
We used for this first analysis a Cλ4SEG2 (clone 524-12)probe, which should also hybridize with the IGLC4SEG1,IGLC4 and IGLC1 genes since they share more than85% identity. Hybridization of digested B6.κ-λSEG DNAsamples revealed one EcoRI hybridizing band, whichcould represent a doublet, and two HindIII hybridizing
Fig. 3a, b Ig light chain isotypes expressed by B6.κ–λSEG Igha
mouse B cells. B6.κ–λSEG Igha (a) and 129 Sv.κ– (b) peripheralblood B cells were analyzed for the expression of Ig light chainisotypes after staining by anti-µa allotype Ab and anti-Cλ2/Cλ3(189-B3) Ab which recognizes λ2, λ3 and λx light chain isotypes
Fig. 4a, b SEG IGL locus characterization by RFLP. RFLP analysisof EcoRI (a) and HindIII (b) digested DNA of B6(1) and B6.κ-
λSEG (2) mouse strains hybridized with indicated probes. Positionsof λ/HindIII DNA molecular weight markers are indicated in kb

112
bands under high (Fig. 4a, b) or low stringency condi-tions (unpublished observations) whereas BamHI diges-tion revealed two bands (Table 1). In the case of B6DNA, a faint hybridizing fragment attributable to IGLC1is detected with EcoRI (Fig. 4a) and BamHI (Table 1)digestions in addition to a single IGLC4 fragment. Notethat the IGLC1-containing restriction fragment was notdetected for B6 HindIII-digested DNA under high strin-gency conditions but it can be readily identified underless stringent conditions (unpublished observations).Hybridization data are summarized in Table 1. Theseresults are compatible with PCR and sequence data,confirming the presence of the two related IGLC4 genesand the absence of IGLC1 in the B6.κ–λSEG genomesince the Cλ4 probe does not contain HindIII or BamHIrestriction sites.
Note that under low stringency conditions an additionalband could be seen with EcoRI and HindIII digestscommon to all strains tested, which is attributed toIGLC5 (Mami et al. 1988).
Cλ2, Cλ3 family
These two genes share 97% nucleotide sequence identitywhile IGLC2SEG shares 99.3% with IGLC2. For thisreason, a Cλ2SEG probe (clone 183-7) should hybridizeto all three related genes. For the three restrictionenzymes used, B6.κ–λSEG presented only one band, even
if a slighter second band was present after BamHI diges-tion (Fig. 4a, b, Table 1). These two bands correspond tothe two IGLC4-hybridizing BamHI fragments. This fainthybridizing band is thus probably due to a limited crossreactivity with IGLC4SEG1/2. These results confirm thepresence of only one IGLC2/IGLC3 related gene at theSEG IGL locus.
When comparing Cλ4SEG and Cλ2SEG RFLP profilesfor B6.κ–λSEG, it appears that at least one IGLC4SEG geneis present on the same 6.3 kb EcoRI fragment withIGLC2SEG. The second IGLC4SEG gene is thus locatedeither on the same EcoRI fragment or on a comigratingfragment in the case of a doublet (Fig. 4a). Mami andKindt (1987a) showed that IGLC4SPE is also found on asimilar 6.3 kb EcoRI fragment with IGLC2SPE (Fig. 2 inMami and Kindt 1987a). After sequencing the wholeEcoRI fragment, but not the central 1,200 bp, they did notdetect any other IGLC gene. Our sequence analysis indi-cates that IGLC4SEG1 and IGLC4SPE are orthologues.Altogether, these observations place IGLC2SEG andIGLC4SEG1 on the same EcoRI fragment, IGLC4SEG2being situated outside this fragment, probably down-stream. The HindIII restriction profiles indicate that thethree SEG IGLC are on different HindIII fragments. FromBamHI restriction profiles, it appears that IGLC4SEG1and IGLC4SEG2 are on two different fragments and thatIGLC4SEG1 is on the same BamHI fragment withIGLC2SEG (unpublished observations). This result isagain in accordance with those from SPE since no BamHIrestriction site is found on the 6.3 kb EcoRI fragment.
Vλ1, Vλ2 family
IGLV1 and IGLV2 share 97% and 95.5%, respectively,nucleotide sequence identity with IGLV2SEG, althoughIGLV2 is more identical at the amino acid level toIGLV2SEG than to IGLV1. Consequently, a Vλ2SEGprobe (clone 186-2) should hybridize all IGLV1/2-relatedgene segments in the Mus genus. The presence of onlyone band after EcoRI and HindIII digestion (Fig. 4a andb), with one strongly hybridizing band together with twoslight bands after BamHI digestion for B6.κ-λSEG
(Table 1) suggests, along with our sequencing datapresented above, that the SEG IGL locus comprises onlythe IGLV2SEG gene segment.
Vλx family
For this experiment we used a VλxSEG probe (clone246-17) that hybridized to all tested DNAs. The VλxSEGprobe contains a BamHI restriction site. This explainsthe two BamHI hybridizing bands present for B6.κ–λSEG
mouse (Table 1). Since this mouse shows a single IGLVXhybridizing restriction fragment after EcoRI or HindIIIdigestions (Fig. 4a, b), we conclude the presence of asingle copy of IGLVXSEG at the SEG IGL locus assuggested by PCR and sequence analysis
Table 1 Summary of data obtained after RFLP analysis of B6,B6.κ–λSEG and Vλ4+ wild-derived inbred mouse strain DNAs withVλ and Cλ probes. C57BL/6, B6.κ-λSEG and Vλ4+ wild-derivedinbred mouse strains DNA RFLP profiles were obtained withprobes able to hybridize to the indicated gene families. The estimatednumber of genes and gene segments is deduced from the numberof hybridizing bands, presented in parentheses, after digestion byEcoRI, HindIII and BamHI restriction enzymes, respectively. ND Not determined
Strain Gene and gene segment families
Cλ1/4 Cλ2/3 Vλ1/2 Vλx Vλ4
C57BL/6 2 2 2 1 0(2, 2, 2) (2, 2, 2) (2, 2, 2) (1, 1, 1) (0, 0, 0)
B6.κ-λSEG 2 1 1 1 0(1a, 2, 2) (1, 1, 2b) (1, 1, 3c) (1, 1, 2d) (0, 0, 0)
MAI 3 7 3 2 5(4, 2, 4) (7, 7, 6) (3, 3, 3) (2, 1, 2) (5, 5, 6)
MBK 3 6 3 2 5(4, 2, 2) (7, 6, ND) (4, 3, 3) (2, 2, 1) (4, 5, 5)e
PWK 2 6 2 2 5(2, ND, ND) (7, ND, 5) (2, 3, 2) (1, 2, 2) (5, 5, 5)
STF 2 4 3 1 2(2, 1, 2) (3, 4, 4) (3, 2, 3) (1, 1, 2) (2, 2, 2)
SMZ 3 5 2 1 3(3, 3, 3) (6, 5, 5) (2, 2, 2) (1, 1, 2) (ND, 3, 3)
a Probably represents a doubletb One faint band corresponding to a probable cross hybridizationwith one of the Cλ4SEGc Two faint bands among the threed Presence of a BamHI restriction enzyme site in the probee Determined at low stringency

Taken together, the results above lead us to conclude thatmice with a SEG IGL locus can produce only two IGLlight chains arising from the rearrangement of eitherIGLV2SEG or IGLVXSEG to IGLJ2SEG and associationwith IGLC2SEG. The sequence analysis combined withFACS and SDS-PAGE pointed to the presence of onlyone functional IGLC, namely IGLC2SEG. The RFLPanalysis confirmed the sequence data by identifying thesame number of genes and gene segments: two IGLV,three IGLC and showing the absence of IGLV1, IGLC1and IGLC3. The case of a fourth IGLV gene segment,IGLVSD26, has been investigated and we confirmed itsabsence at the SEG IGL locus (see below).
Characterization of the Vλ4 family
VλSD26 refers to the Vλ region of an Ig produced by anhybridoma derived from a Danish (Skive region) Musmusculus musculus wild mouse strain. It is differentfrom the rest of the Vλ regions described for laboratorymouse strains with which it shares only 40–45% aminoacid identity and approximately 50% DNA identity.IGLVSD26 has a homologue present in a Czech(Sladeckovce region) mouse of the same species and isprobably a representative of a multiple gene segmentfamily as suggested by Southern blot analysis (Reidl etal. 1992). As no information is available for other mousestrains, except its absence from BALB/c, we decided toinvestigate its presence in B6.κ-λSEG along with otherwild inbred mouse strains of Mus musculus domesticus,Mus musculus musculus, Mus spretus and C57BL/6 byPCR and RFLP analysis.
Polymerase chain reaction analysis
We performed an IGLVSD26 and IGLVCZ81-specificPCR (MSD26UP1-MSD26DO1) on a large panel ofdifferent mouse strain DNAs: MBT, MBV, MAI, MPW,MBK, MBB, MBS and PWK of the Mus m. musculussubspecies; ULA, WLA, BIK/g, 38CH, WMP and WGQof the Mus m. domesticus subspecies; SFM, STF, SMZ,SMA, SEG and SPR/Ei of the Mus spretus species, inaddition to C57BL/6 and B6.κ-λSEG. None of the micebelonging to Mus m. domesticus including B6 and B6.κ-
λSEG showed positive amplification. On the other hand,three Mus m. musculus (MAI, MBK and PWK) and onlytwo Mus spretus (STF and SMZ strains, both from NorthAfrica: Tunisia and Morocco, respectively) DNAs wereamplified by our primers, indicating the presence ofIGLVSD26 homologues in their genomes. The negativeresult for B6.κ-λSEG DNA is consistent with the SEGmouse DNA result, the B6.κ-λSEG ancestor strain(Fig. 5a). The cloning of positive PCRs permitted thesequencing of two MAI clones presenting enough differ-ences to be considered as different gene copies: MAI1and MAI2. Similarly, we identified four differentVλSD26 family members in MBK, three in PWK, two in
113
STF and three in SMZ mouse strains (see Fig. 5 fordetails). These sequencing results indicative of a polymor-phic multigene family were confirmed by RFLP analysis(see below). Based on our sequencing information, wepropose to call this new gene family, Vλ4. Individualgene segments are referred to as IGLV4x, where x standsfor the mouse strain and the gene number in the family.The protein sequence of these different gene segments ispresented in Fig. 5c and is deduced from the respectivetranslated nucleotide sequences. Differences are concen-trated in CDR1 and CDR2, reflecting a probable environ-mental pressure, even if differences are also present inframework regions. It is noteworthy that very closesequences are found in both European (Mus m. musculus)and North African (Mus spretus) mice, as illustrated bythe phylogenetic tree in Fig. 5d. This tree shows a cleardifference between the three different Vλ mouse families,each family clustering with its rat and human homo-logues. Within the Vλ4 family, a first divergence betweenthe human and the rodent sequences indicates that miceand rats share a common ancestor. Secondly, all mouseIGLV4 sequences form a monophyletic group, whichdoes not include the rat sequence. This indicates that themouse Vλ4 family has diversified after the mouse (Mus)speciation. Thirdly, Vλ4 family members from differentmouse species cluster in the same branch. For example,Mus m. musculus IGLV4SD26 and IGLV4MBK2 are morerelated to Mus spretus IGLV4SMZ1 than to other Mus m.musculus Vλ4 family members.
Restriction fragment length polymorphism analysis
A deeper analysis by RFLP was carried out for the PCRVλ4+ MAI, MBK, PWK, STF and SMZ mouse strainstogether with B6 and B6.κ-λSEG. We used for this analysisa Vλ4SMZ probe (clone 585-4 from SMZ mouse, corre-sponding to IGLV4SMZ1) that is 99.6% homologous toIGLV4SD26. While B6 and B6.κ-λSEG strains DNA didnot show any hybridization pattern, the other DNAstrains showed multiple hybridizing bands with the threerestriction enzymes used. At high stringency, MAI andPWK DNA showed very similar profiles comprising fivebands with only one band difference after EcoRI andHindIII digestions. After BamHI digestion, the profilewas still similar except that MAI showed one more band,probably because of the presence of a BamHI restrictionsite in one of the five IGLV4 gene segments. For MBK,we have been unable to precisely determine the numberof bands, but a preliminary analysis showed the presenceof four to five restriction fragments after EcoRI orHindIII digestions and hybridizations at low stringency.Interestingly, RFLP profiles for MAI, MBK, STF andSMZ strains did not show any difference in the numberof bands at high and low stringency. STF and SMZshowed a different profile; the first presenting two bandsin each case and the second three bands after HindIII andBamHI digestions (Fig. 5b). These data, when combinedwith the sequencing results, suggest the presence of at
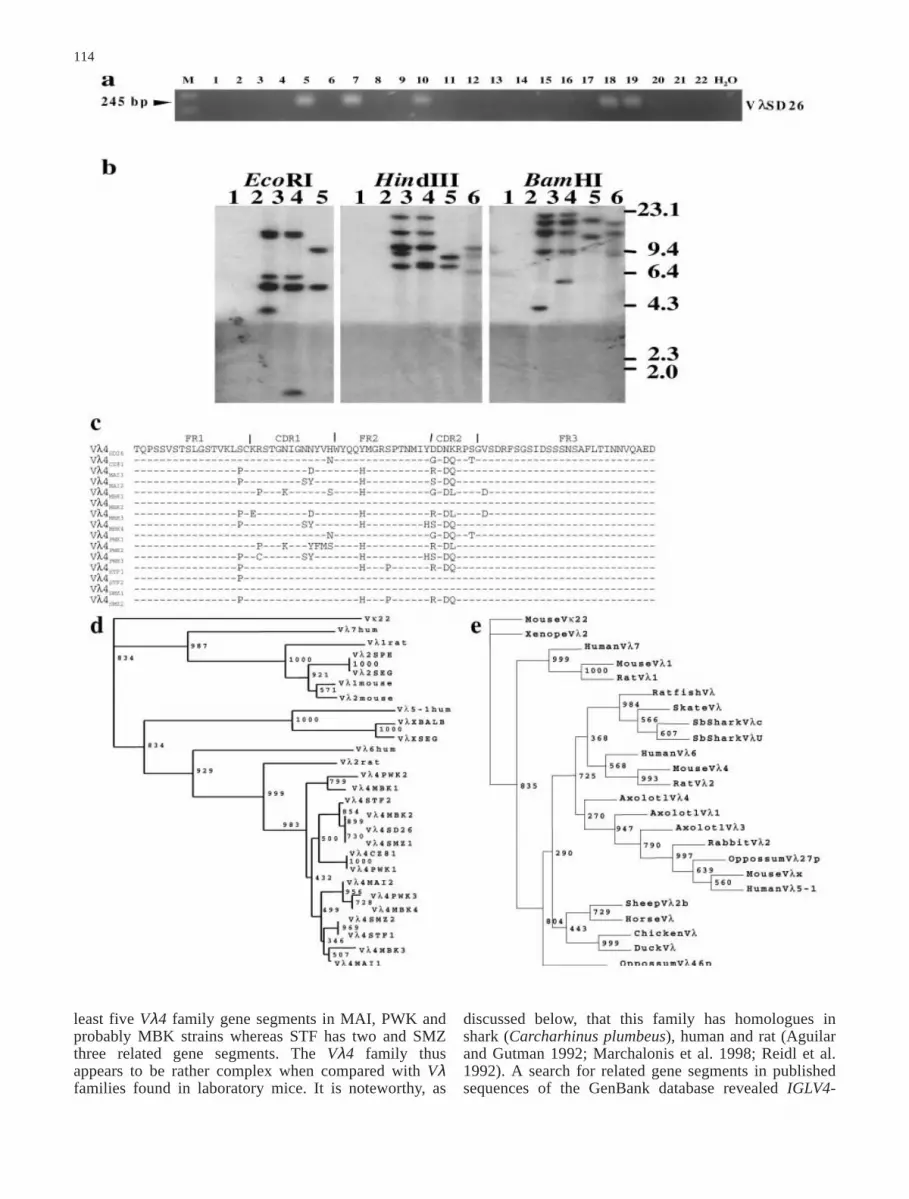
114
least five Vλ4 family gene segments in MAI, PWK andprobably MBK strains whereas STF has two and SMZthree related gene segments. The Vλ4 family thusappears to be rather complex when compared with Vλfamilies found in laboratory mice. It is noteworthy, as
discussed below, that this family has homologues inshark (Carcharhinus plumbeus), human and rat (Aguilarand Gutman 1992; Marchalonis et al. 1998; Reidl et al.1992). A search for related gene segments in publishedsequences of the GenBank database revealed IGLV4-

related sequences in the ratfish (Hydrolagus colliei),skate (Raja erinacea) and the mexican axolotl (Ambystomamexicanum) (Andre et al. 2000; Rast et al. 1994).Together with IGLV4, these sequences share an insertionof two more amino acids in the third framework (datanot shown). The Phylip parsimonious phylogenetic treein Fig. 5e illustrates this and suggests an early divergencebetween the three mouse Vλ families, together with theirknown homologues from other vertebrate species. Inaddition, the tree shows a close clustering between themouse, rat and human IGLV4 homologues and theratfish, skate and sandbar sequences. To a lesser extent,the axolotl IGLV4 sequence appears to be also related.
Finally, we extended the RFLP analysis in MAI,MBK, PWK, STF and SMZ mouse strains to all knownmouse IGL gene and gene segment families: Cλ1/Cλ4,Cλ2/Cλ3, Vλ1/Vλ2 and Vλx. Whereas the number ofIGLC1/IGLC4 genes seems to be very homogenous (twoto three), some variability is found for the IGLC2/IGLC3family members: (1) MAI, MBK and PWK have six toseven genes and (2) STF and SMZ have four to fivegenes. For IGLV1/IGLV2 and IGLVX gene segments, inboth cases, no significant difference is seen, with thenumber of gene segments varying from two to three andone to two copies, respectively (Table 1). As will bediscussed below, these observations strengthen the ideathat the IGL locus evolution has been very complex andis hard to determine, as no simple route can be drawnregarding the absence or presence of Vλ4 family members.
Discussion
The original aim of this study was to give a comprehen-sive description of the IGL locus of a new mouse modelcharacterized by a large reduction of the Ig light chainrepertoire. A wild-derived inbred strain of Mus spretusspecies, the SEG strain (originating in Grenada, Spain)was characterized by a deficiency in serum λ1 Ig expres-sion. This original observation, consistent with mole-cular data obtained in other geographically related Musspretus mice (Amor et al. 1983; Kindt et al. 1985; Mamiand Kindt 1987a, b; Scott and Potter 1984a, b), pointedout the possibility of developing a mouse strain produc-ing only two Ig light chains, namely λ2 and λx, insteadof the four found in conventional laboratory strains. Tothis end, we took advantage of the development of aseries of IRCS, one of which turned out to carry theSEG/Pas allele at the IGL locus in a C57BL/6J back-ground. This B6.λSEG was crossed to a κ- mouse in orderto obtain a B6.κ-λSEG strain that expresses no κ, λ1 andλ3 light chains. This strain expresses an extremelyreduced genetic repertoire of Ig light chains and shouldrepresent an interesting model to better understand themechanisms governing the diversification of Ig lightchains. It is completely different from strains known aslow λ1 B-cell producers. Those SJL and related strainsare able to rearrange and produce λ1 light chains whichdo not associate efficiently with heavy chains, due to aglycine to valine substitution in the IGLC1 region (Arpet al. 1982; Kim et al. 1994).
Our characterization of the SEG IGL locus wasinitially conducted by PCR analysis using primers ableto amplify all the known IGLV and IGLC gene segments.This first approach confirmed the presence of only twoIGLV gene segments and three IGLJ-IGLC clustersamong which only one is functional. The two V genesegments are IGLV2SEG and IGLVXSEG. IGLV2SEG isidentical to IGLV2SPE in its coding sequence at theDNA and protein levels. IGLVXSEG presents ten aminoacid differences with BALB/c and B6 IGLVX sequences.It also shows a stop codon at its end, which considerably
115
Fig. 5 Characterization of the Vλ4 family in wild mouse strains.a PCR genotyping. B6 (1), B6.κ–λSEG (2), MBT (3), MBV (4),MAI (5), MPW (6), MBK (7), MBB (8), MBS (9), PWK (10),ULA (11), WLA (12), BIK/g (13), 38CH (14), WMP (15), WGQ(16), SFM (17), STF (18), SMZ (19), SMA (20), SEG (21) andSPR/Ei (22) genomic DNAs were amplified with Vλ4 primers(MSD26UP1-MSD26DO1). Amplified fragments were detectedafter agarose gel electrophoresis and ethidium bromide staining.H2O is the PCR negative control. Fragment lengths were estimatedby comparison to the 100 bp ladder marker (M). b RFLP analysis.B6 (1), B6.κ–λSEG (2), MAI (3), PWK (4), STF (5) and SMZ (6)mouse strain DNAs were digested with indicated enzymes andhybridized with a Vλ4 probe (clone 585-4 corresponding toVλ4SMZ1 nucleotide sequence). Positions of λ/HindIII DNAmolecular weight markers are indicated on the right. c Vλ4 familyprotein sequence comparison. Vλ4 protein sequences are com-pared to Vλ4SD26 and Vλ4CZ81 protein sequences from M. m.musculus mice, Skive (Denmark) and Sladeckovce (Czech Repub-lic), respectively (GenBank accession nos.: M94349, M94351,AF357982, AF357983, AF357984, AF357985, AF357986,AF357987, AF357981, AF357980, AF357979, AF357976,AF357975, AF357978, AF357977). All protein sequences aretranslated from the respective GenBank nucleotide sequences. FRand CDR are indicated according to Kabat and co-workers (1991).End sequences are not available since the primers designed fromcDNA sequences used for the PCR do not cover the full sequenceof the gene segments. d Phylogenetic relationships of mouse, ratand human IGLV4 homologues. A bootstrap (repetition 1,000;seed=619) neighbor-joining phylogenetic tree of mouse, rat andhuman Vλ1, Vλx and Vλ4 families was constructed with theClustal X package. Bootstrap values are indicated at the nodes.The mouse Vκ22 (AJ235965) gene was used as an outgroup.e Parsimonious phylogenetic analysis of vertebrate Vλ families. APhylip parsimonious tree was constructed as follows: a multiplealignment of vertebrate Vλ protein sequences was produced withClustal X followed by manual correction to minimize gaps; 1,000bootstrapped alignments using SEQBOOT (seed=197) were ana-lyzed with PROTPARS. A consensus parsimonious tree was thendetermined using CONSENSUS and visualized with TreeView.Bootstrap values are indicated at the nodes. The mouse Vκ22(AJ235965) was used as an outgroup. See Figs. 3 and 6c formouse GenBank accession numbers. Other sequences are: Homosapiens human Vλ7 (X01015), human Vλ6 (X02422) and humanVλ5–1 (D87024); Rattus Norvegicus rat Vλ1 (M77356) and ratVλ2 (M77357); Monodelphis domestica oppossum Vλ27p(AAC98643) and oppossum Vl46p (AAC98659); Oryctolaguscunigulus rabbit Vλ2 (M27840); Ambystoma mexicanum axolotlVλ1 (AF317321), axolotl Vλ3 (AF317323) and axolotl Vλ4(AF317324); Gallus gallus chicken Vλ (M96962); Cairionamoscata duck Vλ (S49449); Equus caballus horse Vλ (AAA50981);Ovis aries sheep Vλ2b (AAB94909); Hydrolagus colliei ratfish Vλ(AAA59368); Raja erinacea skate Vλ (AAA59380); Carcharhinusplumbeus Sb shark Clx (M81314) and Sb shark U31 (U35007),Xenopus laevis Xenope Vλ (M94393)
▲

reduces the potential junctional diversity of λx chainsand a four-amino-acid longer CDR3 that is characteristicof this Vλ family, as it is conserved in the related rabbitand human IGLV gene segments (Sanchez et al. 1987,1990). Due to the relatedness between SEG and SPEstrains (see below), it is likely that IGLVX was alsopresent in the extinct SPE strain, although it has not beencharacterized.
Only one of the three IGLJ-IGLC clusters is functionaland formed by IGLJ2SEG associated with IGLC2SEG.IGLJ2SEG presents only one amino acid difference withBALB/c IGLJ2 (unpublished observations and manu-script in preparation); IGLC2SEG is 100% homologousto IGLC2 and IGLC2SPE in its protein sequence. Con-cerning the two other nonfunctional clusters, it appearsfrom sequence data that IGLJ4SEG1-IGLC4SEG1 isorthologous to the published IGLJ4SPE-IGLC4SPEcluster as the two IGLC4 genes share a 14 bp deletionthat distinguishes them from IGLC1, IGLC4 andIGLC4SEG2 (Mami and Kindt 1987a). On the otherhand, IGLJ4SEG2-IGLC4SEG2 is homologous to theBALB/c IGLJ4-IGLC4 cluster: it exhibits the same non-functional heptamer in the J sequence (Blomberg andTonegawa 1982; Miller et al. 1982). It is likely that theextinct SPE strain also possessed two IGLJ4-IGLC4clusters homologous to those identified in the SEG IGLlocus. This hypothesis is supported by the earlierobservation of two Cλ4 HindIII hybridizing bands onSouthern blots (see Fig. 4 in Kindt et al. 1985), althoughthe authors did not comment on the presence of thesetwo bands.
Many wild mice present multiple duplications of theirλ recombination units. In order to test for the possiblepresence of multiple gene copies of the PCR-identifiedgenes, we performed RFLP analysis. This last experi-ment confirmed the presence of a single recombinationunit comprising one IGLV2SEG and one IGLVXSEGgene segment associated with one functional IGLJ2SEG-IGLC2SEG and two nonfunctional IGLJ4-IGLC4 clusters.The SEG and SPE RFLP patterns were identical afterEcoRI and HindIII digestions, confirming the relatednessof SEG and SPE mice.
This genomic organization is consistent with FACSanalysis of B6.κ-λSEG mice showing the presence ofIgM+ B cells bearing only Cλ2, and SDS-PAGE analysiswhich identified only two λ chains, the heaviest corre-sponding to λx and the other to λ2, excluding thepresence of λ1 and λ3 chains.
In order to fulfill the description of the SEG IGLlocus, we have finally investigated the presence ofIGLVSD26-related gene segments. IGLVSD26 wasdescribed in a wild inbred M. m. musculus mouse strainfrom Skive (Denmark) together with a homologue in amouse strain of the same species from Sladeckovce(Czech Republic) (Reidl et al. 1992). PCR, as well asRFLP analysis, did not identify any IGLVSD26-relatedgene segment at the SEG IGL locus. Anchored-PCRanalysis of all Vλ-Cλ transcripts supports this observa-tion since only Vλ2SEG or VλxSEG-containing transcripts
116
were identified (manuscript in preparation). Taken together,these data offer strong support for the predicted organi-zation of the SEG IGL locus (Fig. 6).
Interestingly, after extending our PCR and RFLPanalyses to different wild inbred mouse strains, we iden-tified VλSD26 homologues in MAI, MBK and PWK M.m. musculus mouse strains from Austria, Bulgaria andthe Czech Republic, respectively, but also in North AfricanSTF and SMZ M. spretus mouse strains from Tunisiaand Morocco, respectively. No IGLVSD26-related genesegment was present in any of the wild M. m. domesticusinbred mouse strains tested, the species from whichlaboratory mouse strains are derived. The RFLP analysisconducted for PCR-positive strains confirmed thesequence data by showing the presence of multiple DNAfragments, establishing VλSD26 as a polymorphic multi-gene family that, therefore, we propose to call Vλ4.Surprisingly, the Vλ4 family is present in different Musspecies from various geographic origins. With regard tothe number of genes and gene segments present in theVλ4+ mouse strains tested, it appears that (1) all strainshave more IGLV gene segments and IGLJ-IGLC clustersthan the prototypic laboratory mouse, possibly due to anamplification of recombination units by duplicationevents; (2) IGLV4 gene segments are more numerous orequal to the sum of IGLV1/2 and IGLVX gene segmentsexcept for the STF strain; (3) all strains have IGLV1/2and IGLVX homologues, and (4) all strains have moreIGLC2/3 than IGLC1/4-related genes which suggests ahigher linkage of IGLV4 gene segments to IGLC2/3-related genes. This last point is apparently not in accor-dance with the reported association of IGLV4SD26 withan IGLC1-related gene in the Skive mouse strain (Reidlet al. 1992). However, a closer examination of DNAsequence alignments for all known IGLC1/4 indicatesthat this IGLC1-related gene presents mixed characteristicswith IGLC1 and IGLC4/4SEG1/4SEG2 genes (unpub-lished observations), and could, in fact, be part of a unitof recombination including IGLC2. These observationssuggest a large fluidity of the IGL locus genomic organi-zation, characterized by numerous independent duplica-tion/deletion events of partial or whole recombination
Fig. 6 The predicted SEG IGL locus organization. It has beendeduced according to our results and to the organization of theSPE (Mami and Kindt 1987a) and the laboratory mice loci (Storbet al. 1989). The hatched boxes represent the IGLJ4SEG1-IGLC4SEG1 cluster that is not present in the laboratory IGL micelocus. The absence of the second unit of recombination at the SEGIGL locus is indicated by the large cross. Note that this represen-tation does not mean that we consider that the SEG IGL locusorganization derives from that of laboratory mice following theloss of one unit of recombination

propose that the ancestral Mus IGL locus contained thethree IGLV gene segments related to IGLV1/2, IGLVXand IGLV4 with at least one IGLJ2-IGLC2-like and oneIGLJ1/4-IGLC1/4-like cluster. It is, however, difficult toreconcile our data and that of others concerning thenumber and structure of the ancestral IGLJ-IGLCclusters due to the high heterogeneity of RFLP profilesin wild mice. Again, this probably relates to the fluidityof the IGL locus organization. In particular, it is difficultto speculate whether the ancestral Mus IGL locus com-prised a single recombination unit, as in SPE and SEGmice, or two recombination units, which have beensubject to intensive duplication/deletion events duringevolution and speciation of the Mus genus. For example,our analysis suggests that the Vλ4 family, which isabsent in M. m. domesticus (at least in the series ofstrains studied), absent/present in M. m. musculus andabsent/present in M. spretus, has been lost independentlyin the three subspecies and amplified in M. spretus andM. m. musculus. In conclusion, the IGL locus genomicorganization appears to be subject to constant evolutiondue to numerous duplication/deletion events of bothIGLC and IGLV genes.
Since the IGLV4 gene segment is not present at theSEG IGL locus, our B6.κ–λSEG strain represents a newmouse model able to produce only two Ig light chains,λ2SEG (IGLV2SEG-IGLJ2SEG-IGLC2SEG) and λxSEG(IGLVXSEG-IGLJ2SEG-IGLC2SEG). This mouse pro-vides a unique model to study, under physiological con-ditions, whether gene conversion can be detected inmouse between IGLV2SEG and IGLVXSEG gene seg-ments (Weill and Reynaud 1996; Xu and Selsing 1994).The effect of drastic genetic reduction of Ig light chainloci will be tested in light of alternative diversificationmechanisms such as deletion/ insertion events (Wilson etal. 1998) and extended junctional diversity. All of thesequestions are currently under investigation in the labora-tory using our B6.κ–λSEG model both under physiologicalconditions and following immunization.
Acknowledgements We thank Olivier Gorgette, Carine Gris-liebeand Eric Deveaud for technical advice, Drs. Dominique Rueff-Juyand Pierre Boudinot for critical reading of the manuscript andhelpful discussion. Y.M.A. is supported by CROUS de Paris,France and Ministère de l’Enseignement Supérieur et de laRecherche Scientifique, Algeria.
References
Aguilar BA, Gutman GA (1992) Transcription and diversity ofimmunoglobulin lambda chain variable genes in the rat.Immunogenetics 37:39–48
Amor M, Guenet JL, Bonhomme F, Cazenave PA (1983) Geneticpolymorphism of lambda 1 and lambda 3 immunoglobulinlight chains in the Mus subgenus. Eur J Immunol 13:312–317
Andre S, Guillet F, Charlemagne J, Fellah JS (2000) Structure anddiversity of Mexican axolotl lambda light chains. Immuno-genetics 52:137–144
Arp B, McMullen MD, Storb U (1982) Sequences of immuno-globulin lambda 1 genes in a lambda 1 defective mouse strain.Nature 298:184–187
117
units. This mechanism has already been suggested as themain mechanism for the evolution of Ig/TCR loci andreferred to as a birth-and-death evolutionary process(Nei et al. 1997; Ohno 1970; Ota and Nei 1994). Thecomplex evolutionary process of the IGL locus renders itdifficult to propose an evolutionary scheme and to estab-lish a probable organization of the ancestral IGL mouselocus without referring to other vertebrates.
As mentioned above, IGLV4SD26 has been reportedto be related to the human IGLV6 (82% DNA identity)(Reidl et al. 1992). IGLV4SD26 orthologue gene segmentshave been described in rats (Aguilar and Gutman 1992).They present 90% DNA identity with IGLV4SD26 andconstitute a multigene segment family. More striking isthe sandbar shark (Carcharhinus plumbeus), a cartilagi-nous fish, that expresses a predominant IGLV genesegment presenting around 60% protein identity, concen-trated in frameworks and CDR1, with mouse and humanIGLV4 orthologues (Marchalonis et al. 1998). A searchfor related gene segments in published sequences ofthe GenBank database revealed similarities with otherspecies, namely the ratfish (Hydrolagus colliei), skate(Raja erinacea) and the mexican axolotl (Ambystomamexicanum) (Andre et al. 2000; Rast et al. 1994). Thoserelated sequences present a common feature, which isthe presence of two additional amino acids in the thirdframework. However, the skate and ratfish have oneamino acid less when considering the full V segmentlength. Based on these observations, it is noteworthy thatall of the above-mentioned species present a clusteredorganization of the IGL locus. With the exception ofshark, ratfish and skate, in which IGLV-J genes arefound joined in the germline as a derived characteristic(Litman et al. 1999; Rast et al. 1994), these species usejunctional and combinatorial diversity throughout theirlife for Ig repertoire diversification. In these species,IGHV, IGLV and IGKV gene segment families belong todifferent groups as defined by Sitnikova and Su (1998).Conversely, species using gene conversion or/and exten-sive somatic mutations as a source of diversification(chicken, rabbit, cattle, swine, sheep, horse) have notbeen reported as possessing a IGLV4 homologue; thesespecies generate their primary Ig repertoire in gut-associ-ated lymphoid tissues, generally from a single IGHV,IGLV or IGKV group. Lastly, these species tend to pro-duce more λ than κ Ig, except for the rabbit (10:90)(Butler 1997; Sitnikova and Su 1998). These observa-tions confirm our hypothesis that the Vλ4 family waspresent on the ancestral Mus IGL locus. Furthermore,IGLV4 appears to have been present on the ancestralvertebrate IGL-like locus, and conserved throughoutevolution except in those species that selected an alterna-tive Ig diversification mechanism such as gene conver-sion or hypermutation.
The presence of IGLV1/2 homologues in humans, ratsand other species, and IGLVX homologues in human,rats and rabbits, suggests their presence in the ancestralMus genus IGL locus (Hayzer 1990; Sanchez et al.1990). Taken together, these observations lead us to

Blomberg B, Tonegawa S (1982) DNA sequences of the joiningregions of mouse lambda light chain immunoglobulin genes.Proc Natl Acad Sci USA 79:530–533
Bot A, Nangpal A, Pricop L, Bogen B, Kaushik A, Bona CA(1996) V lambda-light chain genes reconstitute immuneresponses to defined carbohydrate antigens or haptens byutilizing different VH genes. Mol Immunol 33:1359–1368
Butler JE (1997) Immunoglobulin gene organization and themechanism of repertoire development. Scand J Immunol 45:455–462
Chen J, Trounstine M, Kurahara C, Young F, Kuo CC, Xu Y,Loring JF, Alt FW, Huszar D (1993) B cell development inmice that lack one or both immunoglobulin kappa light chaingenes. EMBO J 12:821–830
Dildrop R, Gause A, Muller W, Rajewsky K (1987) A new V geneexpressed in lambda-2 light chains of the mouse. Eur J Immunol17:731–734
Eisen HN, Reilly EB (1985) Lambda chains and genes in inbredmice. Annu Rev Immunol 3:337–365
Hayzer DJ (1990) Immunoglobulin lambda light chain evolution:Igl and Igl-like sequences form three major groups. Immuno-genetics 32:157–174
Hinds KR, Litman GW (1986) Major reorganization of immuno-globulin VH segmental elements during vertebrate evolution.Nature 320:546–549
Kabat EA, Wu TT, Perry HM, Gottesman KS, Foeller C (1991)Sequences of proteins of immunological interest. Public HealthService, NIH, Washington, DC
Kim JY, Kurtz B, Huszar D, Storb U (1994) Crossing the SJLlambda locus into kappa-knockout mice reveals a dysfunctionof the lambda 1-containing immunoglobulin receptor in B celldifferentiation. EMBO J 13:827–834
Kindt TJ, Gris C, Guenet JL, Bonhomme F, Cazenave PA (1985)Lambda light chain constant and variable gene complementsin wild-derived inbred mouse strains. Eur J Immunol 15:535–540
Kirschbaum T, Pourrajabi S, Zocher I, Schwendinger J, Heim V,Roschenthaler F, Kirschbaum V, Zachau HG (1998) The 3’part of the immunoglobulin kappa locus of the mouse. Eur JImmunol 28:1458–1466
Lewis SM (1994) The mechanism of V(D)J joining: lessons frommolecular, immunological, and comparative analyses. AdvImmunol 56:27–150
Litman GW, Rast JP, Shamblott MJ, Haire RN, Hulst M, Roess W,Litman RT, Hinds-Frey KR, Zilch A, Amemiya CT (1993)Phylogenetic diversification of immunoglobulin genes and theantibody repertoire. Mol Biol Evol 10:60–72
Litman GW, Anderson MK, Rast JP (1999) Evolution of antigenbinding receptors. Annu Rev Immunol 17:109–147
Mami F, Kindt TJ (1987a) C lambda 2 and C lambda 4 immuno-globulin light chain genes in a wild-derived inbred mousestrain. J Immunol 138:3980–3985
Mami F, Kindt TJ (1987b) The SPE wild mouse strain has a singleIg V lambda gene related to V lambda 1 and V lambda 2 ofBALB/c. Immunogenetics 26:291–295
Mami F, Cazenave PA, Kindt TJ (1988) Conservation of theimmunoglobulin C lambda 5 gene in the Mus gene. EMBO J7:117–122
Marchalonis JJ, Schluter SF, Bernstein RM, Hohman VS (1998)Antibodies of sharks: revolution and evolution. Immunol Rev166:103–122
Miller J, Selsing E, Storb U (1982) Structural alterations in Jregions of mouse immunoglobulin lambda genes are associatedwith differential gene expression. Nature 295:428–430
Nei M, Gu X, Sitnikova T (1997) Evolution by the birth-and-deathprocess in multigene families of the vertebrate immune system.Proc Natl Acad Sci USA 94:7799–7806
Ohno S (ed) (1970) Evolution by gene duplication. Springer,New York Berlin Heidelberg
Ota T, Nei M (1994) Divergent evolution and evolution by thebirth-and-death process in the immunoglobulin VH genefamily. Mol Biol Evol 11:469–482
118
Page RD (1996) TreeView: an application to display phylo-genetic trees on personal computers. Comp Appl Biosci 12:357–358
Pricop L, Brumeanu T, Elahi E, Moran T, Wang BS, Troustine M,Huszar D, Alt F, Bona C (1994) Antibody response elicited byT-dependent and T-independent antigens in gene targetedkappa-deficient mice. Int Immunol 6:1839–1847
Rast JP, Anderson MK, Ota T, Litman RT, Margittai M, ShamblottMJ, Litman GW (1994) Immunoglobulin light chain classmultiplicity and alternative organizational forms in early verte-brate phylogeny. Immunogenetics 40:83–99
Reidl LS, Kinoshita CM, Steiner LA (1992) Wild mice express anIg V lambda gene that differs from any V lambda in BALB/cbut resembles a human V lambda subgroup. J Immunol 149:471–480
Sambrook J, Fritsch EF, Maniatis T (1989) Molecular cloning:a laboratory manual. Cold Spring Harbor Laboratory Press,Cold Spring Harbor, NY
Sanchez P, Cazenave PA (1987) A new variable region inmouse immunoglobulin lambda light chains. J Exp Med 166:265–270
Sanchez P, Marche PN, Le Guern C, Cazenave PA (1987) Structureof a third murine immunoglobulin lambda light chain variableregion that is expressed in laboratory mice. Proc Natl Acad SciUSA 84:9185–9188
Sanchez P, Marche PN, Rueff-Juy D, Cazenave PA (1990) MouseV lambda x gene sequence generates no junctional diversityand is conserved in mammalian species. J Immunol 144:2816–2820
Sanchez P, Nadel B, Cazenave PA (1991) V lambda-J lambdarearrangements are restricted within a V-J-C recombinationunit in the mouse. Eur J Immunol 21:907–911
Sanchez P, Drapier AM, Cohen-Tannoudji M, Colucci E, BabinetC, Cazenave PA (1994) Compartmentalization of lambdasubtype expression in the B cell repertoire of mice with adisrupted or normal C kappa gene segment. Int Immunol 6:711–719
Sanchez P, Rueff-Juy D, Boudinot P, Hachemi-Rachedi S,Cazenave PA (1996) The lambda B cell repertoire of kappa-deficient mice. Int Rev Immunol 13:357–368
Sanchez P, Hachemi-Rachedi S, Boudinot P, Cazenave PA (1999)Expression of the immunoglobulin lambda locus in themouse. In: Zanetti M, Capra JD (eds) Harvard Academic,pp 151–174
Scott CL, Potter M (1984a) Polymorphism of C lambda genesand units of duplication in the genus Mus. J Immunol 132:2630–2637
Scott CL, Potter M (1984b) Variation in V lambda genes in thegenus Mus. J Immunol 132:2638–2643
Scott CL, Mushinski JF, Huppi K, Weigert M, Potter M (1982)Amplification of immunoglobulin lambda constant genes inpopulations of wild mice. Nature 300:757–760
Selsing E, Miller J, Wilson R, Storb U (1982) Evolution of mouseimmunoglobulin lambda genes. Proc Natl Acad Sci USA 79:4681–4685
Sitnikova T, Su C (1998) Coevolution of immunoglobulinheavy- and light-chain variable-region gene families. Mol BiolEvol 15:617–625
Southern EM (1975) Long range periodicities in mouse satelliteDNA. J Mol Biol 94:51–69
Storb U, Haasch D, Arp B, Sanchez P, Cazenave PA, Miller J(1989) Physical linkage of mouse lambda genes by pulsed-field gel electrophoresis suggests that the rearrangementprocess favors proximate target sequences. Mol Cell Biol 9:711–718
Taki S, Meiering M, Rajewsky K (1993) Targeted insertion of avariable region gene into the immunoglobulin heavy chainlocus. Science 262:1268–1271
Thiebe R, Schable KF, Bensch A, Brensing-Kuppers J, Heim V,Kirschbaum T, Mitlohner H, Ohnrich M, Pourrajabi S,Roschenthaler F, Schwendinger J, Wichelhaus D, Zocher I,Zachau HG (1999) The variable genes and gene families of the

Wilson P, Liu YJ, Banchereau J, Capra JD, Pascual V (1998)Amino acids and deletions contribute to diversify the humanIg repertoire. Immunol Rev 162:143–151
Xu B, Selsing E (1994) Analysis of sequence transfers resemblinggene conversion in a mouse antibody transgene. Science 265:1590–1593
Zou YR, Takeda S, Rajewsky K (1993) Gene targeting in the Igkappa locus: efficient generation of lambda chain-expressingB cells, independent of gene rearrangements in Ig kappa.EMBO J 12:811–820
119
mouse immunoglobulin kappa locus. Eur J Immunol 29:2072–2081
Tonegawa S, Maxam AM, Tizard R, Bernard O, Gilbert W (1978)Sequence of a mouse germ-line gene for a variable region ofan immunoglobulin light chain. Proc Natl Acad Sci USA 75:1485–1489
Weill JC, Reynaud CA (1996) Rearrangement/hypermutation/gene conversion: when, where and why? Immunol Today 17:92–97




















