
Mortality Partitions and their Relevance to Research on Senescence
Upload
independentCategory
view
3download
0
IDENTIFICATION AND CHARACTERIZATION OF GENES INVOLVED IN THECARCINOGENESIS OF HUMAN SQUAMOUS CELL CERVICAL CARCINOMAQing CHENG
1, Wen Min LAU1, S.K. TAY
2, S.H. CHEW3, T.H. HO
4 and Kam M. HUI1*
1Laboratory of Gene Structure and Expression, Division of Cellular and Molecular Research, National Cancer Centre, Singapore2Department of Obstetrics and Gynaecology, Singapore General Hospital, Singapore3Department of Pathology and Gynaecological Oncology, KK Women’s and Children’s Hospital, Singapore4Department of Gynaecological Oncology, KK Women’s and Children’s Hospital, Singapore
We utilized RT-PCR differential display and cDNA mi-croarrays to identify cellular genes involved in the multi-stepcarcinogenesis of squamous cell cervical carcinoma. Thirty-eight cervical cancer patients in various stages of the diseaseand 5 non-cervical cancer patients were studied. Twenty-fivecDNA clones were identified and these were subsequentlydemonstrated to be consistently over-expressed in squa-mous cell cervical carcinoma biopsies of various FIGO stages.To further evaluate the possible role that these genes mayplay in the progression of disease, we performed Northernblot analysis and RNA-RNA in situ hybridization studies usingcervical cancer biopsies of various FIGO stages. Of particularinterest are the 2 clones G32C4B and G30CC that have beenidentified to be the NADH dehydrogenase 4 gene and thegene that encodes ribosomal protein S12 respectively whencompared to sequences available in the GenBank database.Increased expression of these 2 genes were detected in thematched normal tissues collected together with the lateFIGO stages of cervical cancer biopsies. In comparison, up-regulation of these 2 genes was not detected in cervicalsquamous epithelium collected from patients admitted forsurgery for non-malignant conditions, suggesting that ex-pression of these 2 genes may have altered in the adjacenthistopathologically “normal” cervical squamous epithelialtissue from cervical cancer patients. The ribosomal proteinS12 and the NADH dehydrogenase 4 genes may therefore bepotentially useful as early pre-transformation diagnosticmarkers for human cervical cancer.© 2002 Wiley-Liss, Inc.
Key words: human cervical cancer; differential display and cloning;cDNA microarrays; NADH subunit 4; ribosomal protein S12
Invasive carcinoma of the cervix develops from well-definedprecursor lesions referred to as cervical intraepithelial neoplasia(CIN) at the squamous-columnar junction. Longitudinal studieshave shown that untreated high grade CIN will develop intoinvasive carcinoma in 30–70% of cases over a period of 10–12years, though in about 10% of patients, lesions can progress fromin situ to invasive in less than a year.1,2 The prognosis for cervicalcancer is markedly affected by the extent of disease at the time ofdiagnosis.3 Among the major factors that influence prognosis arestage, volume and grade of tumor, histologic type, lymphaticspread and vascular invasion.4 Cytogenetic abnormalities andchromosome structural aberrations or allele loss have also beendetected in these lesions.5,6 The role of these genetic changes onthe pathogenesis of cervical cancer, however, has not been fullyevaluated. The molecular events that govern these changes areunclear at present.
Human papillomavirus (HPV) infection has long been impli-cated to play a significant role in the development of pre-invasiveor invasive carcinoma of the cervix.7,8 The HPV E6 and E7 genesand to a lesser extent E5, are involved in cellular transforma-tion.9,10 Although others appear to be at significant risk, however,it has been observed that some patients with HPV infection appearto be at minimal increased risk for the development of cervicalpre-invasive and invasive malignancies.11–13 These observationsindicate that other genetic factors, such as somatic genetic alter-ations, may play a decisive role in the development of malignantcancer.5,6 Intensive screening to search for candidate genes in
cervical cancer and CIN, with more accurate predictive value, isurgently needed, especially to identify genes that play a role in thevarious stages of the spectrum of the disease. This approach willeventually allow us to work out the hierarchy of their interactionstoward cervical carcinogenesis. In the present study, we haveemployed differential gene displays and cDNA microarrays assystematic methods to compare profiles of gene expression be-tween biopsies obtained from patients with squamous cell carci-noma of the cervix of various FIGO stages and normal squamousepithelium tissue.
MATERIAL AND METHODS
Tissue collection and RNA isolationBiopsies were collected from patients with squamous carcinoma
of the cervix. Punch biopsies with Tischler forceps were takenfrom the tumor lesion for histopathological assessment as well asfor RNA analysis. For histopathological analysis, the tissues werefixed in 10% neutral buffered formalin and processed into paraffinblocks. Multiple step sections were made and stained with haema-toxylin and eosin. Where indicated, PAS with and without diastasedigestion and mucicarmine stains were performed. Tissues col-lected for RNA analysis were snap frozen in liquid nitrogen.Wherever possible normal cervical tissues far away from the tumorareas were taken for controls. The staging of cancer was doneaccording to recommendation by the International Federation ofObstetrics and Gynaecology (FIGO).14 To avoid repetition inevaluating gene expression in the same patient, different patientswere included in different studies. In total, 38 cervical cancerpatients in various stages of the disease and 5 non-cervical cancerpatients were studied. Six of the cervical tissues and their adjacentnormal tissues were employed for the differential display tech-nique. The RNAs prepared from these same 6 individual tissueswere pooled and employed for the reverse Northern blot study.This allowed us to confirm clones that were overexpressed incervical cancer biopsies detected by differential display were also
Abbreviations: BCIP, X-phosphate/5-Bromo-4-chloro-3-indolyl-phas-phate; BL, basal-layer epithelial cells; BSA, bovine serum albumin; CA,carcinoma; CIN, cervical intraepithelial neoplasia; FIGO, InternationalFederation of Obstetrics and Gynaecology; H&E staining, hematoxylin andeosin staining; HPV, Human papillomavirus; NBT, 4-Nitro blue tetrazo-lium chloride; PL, para-layer epithelial cells; ST, stromal cells.
Grant sponsor: Singapore National Medical Research Council; Grantsponsor: Singapore National Science and Technology Board.
*Correspondence to: Laboratory of Gene Structure and Expression, Divi-sion of Cellular and Molecular Research, National Cancer Centre, Singapore.Fax: �65-226-3843 or �65-226-5694. E-mail: [email protected]
Received 30 July 2001; Revised 2 October 2001; Accepted 5 October2001
Published online 4 January 2002
Int. J. Cancer: 98, 419–426 (2002)© 2002 Wiley-Liss, Inc.DOI 10.1002/ijc.10177
Publication of the International Union Against Cancer

overexpressed when tested with another technique. Ten biopsies(obtained from another 10 independent cervical cancer patients),along with their adjacent normal tissues, were employed for ourDNA microarrays studies. For the Northern blot analyses, another8 biopsies from different patients with early stage cervical cancerand another 4 biopsies from patients with late stage cervical cancerwere studied. As stated in our original manuscript, 10 biopsiesobtained from patients with early stage cervical cancer and 4biopsies from patients with late stage cervical cancer were studiedby in situ hybridization. Of the 10 patients with early stage cervicalcancer that we employed for in situ hybridization, 4 patients werethe same as that employed for the Northern blot analyses. There-fore, there were altogether 6 new patients with early stage cervicalcancer and 4 new biopsies from patients with late stage cervicalcancer that were studied by in situ hybridization. For all experi-ments, total cellular RNA was isolated using TRIzol Reagent(Gibco BRL, Life Technologies, Grand Island, NY) according tothe manufacturer’s manual and stored at �80°C before beingemployed in experiments.
All cervical cancer biopsies were routinely screened for HPVand 85% of the cancer biopsies were found to be positive for HPV.For squamous cervical cancer they are of HPV types 16 and 18.The normal tissues were of squamous epithelial cells. The normalcervical tissues, however, were not routinely screened for thepresence of HPV. We also do not have a prevalence rate in ourpopulation as no survey has been done. Nevertheless, we con-firmed that the HPV cytopathic effects on cells of the normaltissues were not observed.
RT-PCR differential displayTotal RNA was purified from cervical cancer biopsies or the
adjacent histopathologically normal tissues. The RNA sampleswere treated with RNase-free DNase I (Promega, Madison, WI) toremove potential chromosomal DNA contamination. One �g eachof total RNA from normal and cancer biopsies was used forreverse transcription.
First-strand cDNAs were synthesized using a one-base anchoredoligo-dT primer (P21) in combination with 3 different primersP30, P31, P32 in individual pairs and PCR-amplified in parallel inthe presence of [alpha-33P]dATP. The parameters for PCR cyclingwere as follows: 40 cycles at 94°C for 30 sec, 40°C for 2 min and72°C for 30 sec and followed by 72°C for 10 min. The PCRproducts, together with size markers, were then separated on asequencing gel, Gel-Mix 6 (Gibco BRL, Life Technologies, GrandIsland, NY). Differentially expressed bands were cut out of the geland cDNA fragments eluted by boiling the gel pieces in 100�lH2O for 15 min before being used as templates for PCR re-amplification using the same conditions as described above. Aportion of the re-amplified PCR product was analyzed on a 2%agarose gel to check the efficiency of re-amplification and toconfirm the size of cDNA fragments. All PCR primers employedfor reverse transcription and PCR were obtained from Gibco BRL(Life Technologies, Grand Island, NY). Primers used included:P21 3�-GTTTTTTTTTTTCGAA-5�; P30 5�-AAGCTTGGT-GACA-3�; P31 5�-AAGCTTAGTC AG-3� and P32 5�-AAGCT-TCCACAGC-3�. AmpliTaq DNA polymerase was purchased fromPerkin-Elmer Corp. (Norwalk, CT). [alpha-33P]dATP (2,000 Ci/mmol) was obtained from New England Nuclear (Boston, MA.).
cDNA cloning and sequencingRe-amplified cDNA fragments were cloned into either the
pCR2.1 vector using the TA cloning system from Invitrogen (SanDiego, CA) or the pCR-TRAP Vector from GenHunter (Gen-Hunter Corp. Nashville, TN). DNA sequencing was performedusing either Sequenase Kit Version 2.0 (United States Biochemi-cal, Cleveland, OH), or by automated sequencing using ABI Prism377 DNA Sequencer (Perkin-Elmer Corp., Norwalk, CT). Thenucleotide sequences obtained were analyzed by BLAST search(National Centre for Biotechnology Information).
cDNA microarray fabrication and hybridizationcDNA clones were amplified in 100 �l PCR reactions, 5 �l PCR
product was analyzed on a 2% agarose gel after which the remain-ing PCR products were purified by isopropanol precipitation, re-suspended in 15 �l 3� SSC and printed onto poly-L-lysine (SigmaDiagnostics, St. Louis, MO) coated glass slides using a GMS 417arrayer (Genetic Microsystems Inc., Woburn, MA). Housekeepinggenes including G3PDH, �-actin, �-2-microglobulin, �-tubulin,cyclophilin and ubiquitin were also spotted as internal controls fornormalization. Each slide was hydrated for 2–3 sec over a steam-ing 100°C water-bath, snap-dried for 5 sec on a 100°C heatingblock and crosslinked with 550 mJ ultraviolet irradiation using aStratalinker (Stratagene, La Jolla, CA). The slide was then washedfor 10 min in 0.2% SDS and rinsed 5� in distilled water beforebeing denatured for 5 min in 100°C distilled water, desiccated for5 min in 95% ethanol and air-dried for 5 min in an 80°C oven.
Comparative hybridizations were carried out using cDNA re-verse-transcribed from total RNA isolated from tumor biopsies andmatched adjacent normal tissue of 10 individual cervical cancerpatients in varying stages of disease. Total RNA was reverse-transcribed into cDNA, labeled with Cy3 (for tumor cDNA) andCy5 (normal cDNA) fluorescent dyes and hybridized onto theslides according to the 3DNA™ Expression Array Detection kitprotocol recommendations (Genisphere Inc., Montvale, NJ). La-beled cDNA from cancer and normal tissues were hybridizedsimultaneously onto a single glass slide in 10 separate hybridiza-tions. Humidified microarray hybridization cassettes were used tocontain all slides during hybridization incubations (TeleChem In-ternational Inc, Sunnyvale, CA). Hybridized microarrays werescanned using a GMS 418 laser scanner (Genetic MicrosystemsInc, Woburn, MA). Separate images were acquired for Cy3 andCy5. The signal and background signals were then quantified withImagene 3.0 quantification software (BioDiscovery Inc., Los An-geles, CA). Generated signal values were analyzed and visualizedusing Cluster and Treeview programs (M. Eisen, University ofCalifornia, Berkeley; http://rana.lbl.gov/).
Signals were measured as the mean pixel intensity within eachcircumscribed spot and background was measured using the meanpixel intensity outside the circumscribed spot diameter within aspecific square location. Spots that were contaminated with fluo-rescent specks or other blemishes, or where background valueswere higher than the spot signal were considered as missing. Thesignal intensities from both Cy3 and Cy5 channels were normal-ized by applying a single scaling factor computed by taking theratio of the total signal intensities of each channel such that thesignal intensities of internal controls on each array would have aCy3/Cy5 ratio of 1.0. Spots that gave signals of 2.5-fold or moredifference in comparison to the internal controls would be consid-ered as differentially expressed.
Northern blot analysisTwenty �g of total RNA isolated from cancer biopsies or their
adjacent histopathologically normal tissues were employed forNorthern analysis. After electrophoresis, the RNA was transferredto Hybond-N� nylon transfer membrane (Amersham, Piscataway,NJ) and probed with random hexa-nucleotide-primed 32P-labeledcDNAs clones using high prime DNA labeling kit (Boehringer-Mannheim GmbH, Mannheim, Germany). Unincorporated nucle-otides were removed by NucTrap probe Purification Columns(Stratagene, La Jolla, CA). Hybridization and washes were per-formed as described previously.15 Hybridization to the housekeep-ing gene G3PDH cDNA probe (Clontech Laboratories, Inc., PaloAlto, CA) was used to quantitate RNA loading. After hybridizationand washing, the membranes were exposed . For analysis, theradioactivity associated with each band was quantitated by densi-tometric scanning using the BioRad FX PhosphorImager (BioRad,Richmond, CA).
420 CHENG ET AL.
RNA-RNA in situ hybridizationRNA-RNA in situ hybridization was performed using DIG
(digoxigenin)-labeled cRNA probe (DIG RNA Labeling Kit, Boe-hringer-Mannheim GmbH, Mannheim, Germany). Two sub-cloneswith opposite orientation of the gene fragment insert were furtherselected to generate sense and anti-sense probes. The labelingreaction was carried out according to the manufacturer’s protocolusing DIG RNA labeling Kit.
The cancer biopsies and the corresponding adjacent histopatho-logical normal biopsies were collected from patients of variouscancer stages. Normal epithelium collected from non-cervical can-cer patients were also employed in our study. Frozen biopsies werecut into sections of 10 �m and mounted directly on microscopeslides. The tissue sections were fixed by incubating at 50°C for 2min and subsequently at room temperature for 7 min in PBScontaining 4% paraformaldehyde. Pre-hybridization was per-formed by incubating at 37°C for 1 hr in blocking solution con-taining 5% skim milk powder, 4� SSPE, 50% deionized form-amide, 30 �g sheared salmon sperm DNA and 1% SDS. Tissuesections were then incubated with 200 ng/ml of DIG-labeledanti-sense or sense cRNA probe at 37°C overnight in hybridizationbuffer (16.6% dextran sulfate, 5% skim milk powder, 4� SSPE,50% deionized formamide and 1% SDS). The tissue sections werefurther incubated at room temperature for 2 hr with 100 mMTris-HCl pH 7.5 and 150 mM NaCl buffer containing 1% BSA and0.5% alpha-alkaline phosphatase-conjugated anti-DIG antibody(BM, MannLein, Germany). For color development, the tissue-section slides were soaked in 0.1 M Tris-HCl pH 9.5, 0.1 M NaCl,0.05 M MgCl2, 3.4% NBT, 1.8% BCIP, 2.4% levamisole over-night at room temperature in the dark. The color reaction wasstopped by incubating the slides in 10 mM Tris-HCl pH 8 and 1mM EDTA at room temperature for 5 min and counter-stainedwith Hematoxylin (BDH Laboratory Supplies, Dorset, England).
For H&E staining, tissue sections were soaked in hematoxylin(filtered haematoxylin contains 1 ml acetic acid v/v) for 4 min,washed in H2O, followed by dipping into 0.5% acid alcohol (0.5%HCl in 75% alcohol) for 2 sec and washed again in H2O. Tissuesections were then soaked in eosin solution (1% eosin stocksolution: 70% alcohol � 1:3) for 3 min and washed with runningH2O. Slides were viewed with the Olympus BX 60 microscope(Olympus Optical Co. Ltd, Tokyo, Japan).
RESULTS
Identification of differentially displayed cDNA fragments fromhuman cervical cancer biopsies
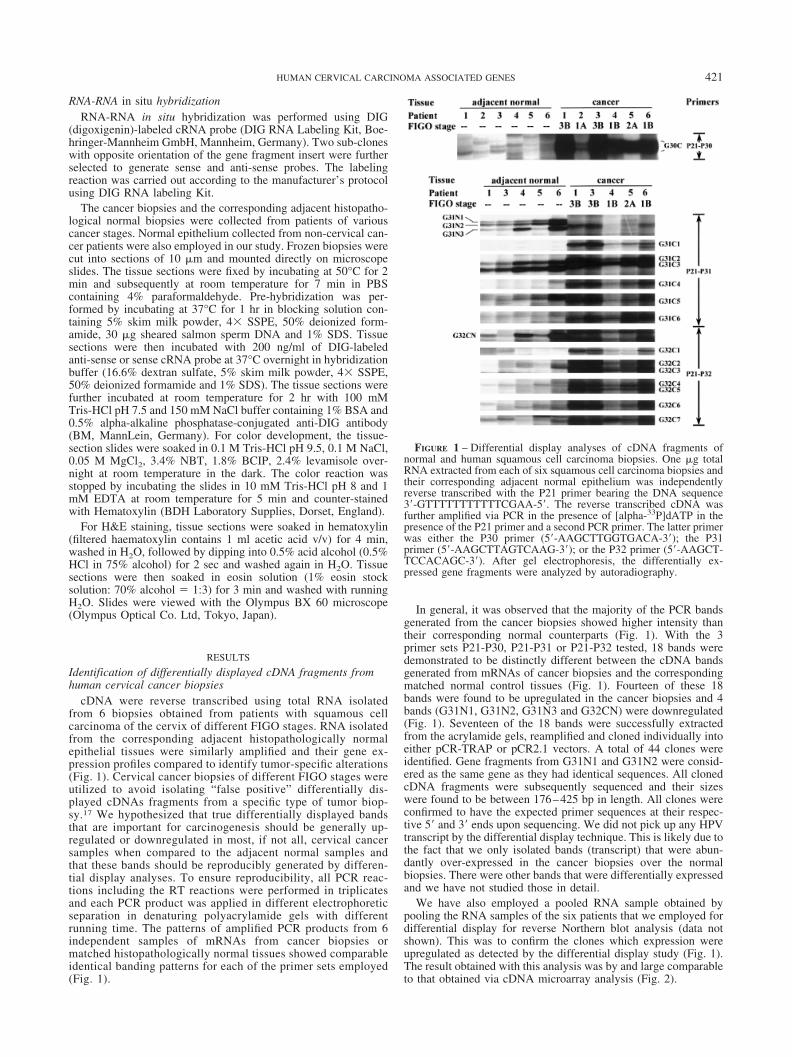
cDNA were reverse transcribed using total RNA isolatedfrom 6 biopsies obtained from patients with squamous cellcarcinoma of the cervix of different FIGO stages. RNA isolatedfrom the corresponding adjacent histopathologically normalepithelial tissues were similarly amplified and their gene ex-pression profiles compared to identify tumor-specific alterations(Fig. 1). Cervical cancer biopsies of different FIGO stages wereutilized to avoid isolating “false positive” differentially dis-played cDNAs fragments from a specific type of tumor biop-sy.17 We hypothesized that true differentially displayed bandsthat are important for carcinogenesis should be generally up-regulated or downregulated in most, if not all, cervical cancersamples when compared to the adjacent normal samples andthat these bands should be reproducibly generated by differen-tial display analyses. To ensure reproducibility, all PCR reac-tions including the RT reactions were performed in triplicatesand each PCR product was applied in different electrophoreticseparation in denaturing polyacrylamide gels with differentrunning time. The patterns of amplified PCR products from 6independent samples of mRNAs from cancer biopsies ormatched histopathologically normal tissues showed comparableidentical banding patterns for each of the primer sets employed(Fig. 1).
In general, it was observed that the majority of the PCR bandsgenerated from the cancer biopsies showed higher intensity thantheir corresponding normal counterparts (Fig. 1). With the 3primer sets P21-P30, P21-P31 or P21-P32 tested, 18 bands weredemonstrated to be distinctly different between the cDNA bandsgenerated from mRNAs of cancer biopsies and the correspondingmatched normal control tissues (Fig. 1). Fourteen of these 18bands were found to be upregulated in the cancer biopsies and 4bands (G31N1, G31N2, G31N3 and G32CN) were downregulated(Fig. 1). Seventeen of the 18 bands were successfully extractedfrom the acrylamide gels, reamplified and cloned individually intoeither pCR-TRAP or pCR2.1 vectors. A total of 44 clones wereidentified. Gene fragments from G31N1 and G31N2 were consid-ered as the same gene as they had identical sequences. All clonedcDNA fragments were subsequently sequenced and their sizeswere found to be between 176–425 bp in length. All clones wereconfirmed to have the expected primer sequences at their respec-tive 5� and 3� ends upon sequencing. We did not pick up any HPVtranscript by the differential display technique. This is likely due tothe fact that we only isolated bands (transcript) that were abun-dantly over-expressed in the cancer biopsies over the normalbiopsies. There were other bands that were differentially expressedand we have not studied those in detail.
We have also employed a pooled RNA sample obtained bypooling the RNA samples of the six patients that we employed fordifferential display for reverse Northern blot analysis (data notshown). This was to confirm the clones which expression wereupregulated as detected by the differential display study (Fig. 1).The result obtained with this analysis was by and large comparableto that obtained via cDNA microarray analysis (Fig. 2).
FIGURE 1 – Differential display analyses of cDNA fragments ofnormal and human squamous cell carcinoma biopsies. One �g totalRNA extracted from each of six squamous cell carcinoma biopsies andtheir corresponding adjacent normal epithelium was independentlyreverse transcribed with the P21 primer bearing the DNA sequence3�-GTTTTTTTTTTTCGAA-5�. The reverse transcribed cDNA wasfurther amplified via PCR in the presence of [alpha-33P]dATP in thepresence of the P21 primer and a second PCR primer. The latter primerwas either the P30 primer (5�-AAGCTTGGTGACA-3�); the P31primer (5�-AAGCTTAGTCAAG-3�); or the P32 primer (5�-AAGCT-TCCACAGC-3�). After gel electrophoresis, the differentially ex-pressed gene fragments were analyzed by autoradiography.
421HUMAN CERVICAL CARCINOMA ASSOCIATED GENES

Profile of gene expression of the cDNA clones in different FIGOstages
To study the potential role of the 44 cDNA clones in thepathogenesis of human cervical cancer, the gene expression profileof these 44 clones in 10 cervical carcinoma biopsies were com-pared to their corresponding adjacent normal tissues using cDNAmicroarrays. cDNA microarrays were generated by printing theDNA purified from the 44 cDNA clones in duplicates onto coatedglass slides. Among the 10 pairs of tissue biopsies studied, 3 pairswere obtained from Stage IB cancer patients, 2 from Stage IIA andStage IIB and 3 from Stage IIIB.
Results obtained from the cDNA analyses demonstrated upregu-lated expression of 25 of the 44 clones in all 10 cervical cancerpatients (Fig. 2, Table I). From our cDNA microarray analyses, nosignificant downregulation of gene expression could be detectedwith clones G31N1 and G32CN in the cancer biopsies (Fig. 2).These results were inconsistent with the results obtained fromdifferential display (Fig. 1) and these clones were not furtherstudied. Data from clones G31N3, G31C5D, G31C5F andG32C2A were technically inadequate and were not included in ouranalyses.
We reasoned that for a gene to be potentially employed as adiagnostic marker, its RNA should be in abundance. Twenty-twoof the 25 clones that have been confirmed to be upregulated bymicroarray analysis were employed as probes Northern blot anal-ysis with RNA isolated from the 4 cervical cancer cell lines, HeLa,CaSKi, SiHa and HT-3 (data not shown). Nine of these clonesdemonstrated strong hybridization signal. They were then chosenas probes for subsequent Northern Blot analyses of RNA purifiedfrom human cervical squamous cell carcinoma biopsies of variousFIGO stages and that of adjacent normal epithelium. It was ob-served that 2 of the clones, namely G30CC and G32C4B, showedelevated levels of expression in cervical cancer biopsies of early
FIGO stages (Fig. 3). In comparison to adjacent histopathologicalnormal epithelium, the expression of clone G30CC was upregu-lated by approximately 5-fold in Stage IIA cancer tissues whereasG32C4B was also upregulated by a similar magnitude in Stage IBcancer tissues. The expressions of these 2 clones were apparentlyalso elevated in the adjacent histopathologically normal epitheliumcollected from the late FIGO stage patients (Fig. 3). Late cervicalcancer tissues would include patients of FIGO Stages IIB and IIIBand in whom the carcinoma extends beyond the cervix withobvious parametrial involvement.14
To study the identity of clones G32C4B and G30CC, we em-ployed these clones as probes for the isolation of the correspondingfull-length cDNA genes by screening a cDNA library derived fromthe HeLa human cervical cancer cell line. Using ClonCapturecDNA Selection Kit, representative positive clones were isolatedfrom the HeLa cDNA library using G32C4B or G30CC as probes.The cDNA obtained were sequenced and they were found to behomologous to the reported NADH dehydrogenase subunit 4 geneand the ribosomal protein S12 mRNA respectively.
Level of gene expression of NADH dehydrogenase subunit 4gene and ribosomal protein S12 in adjacent normal tissues ofcervical cancer tissues
The results obtained from our Northern blot analyses indicatedthat the expression of NADH dehydrogenase 4 gene and theribosomal protein S12 gene was also elevated in the adjacentnormal epithelium collected together with the late clinical stages ofcervical cancer biopsies. To confirm the possibility that the NADHdehydrogenase 4 gene and the ribosomal protein S12 gene areprogressively upregulated in cancer adjacent biopsies as the cancerprogresses, as well as the cell types involved, we conductedRNA-RNA in situ hybridization studies. Squamous cell cervicalcarcinoma biopsies of various FIGO stages, along with their cor-
FIGURE 2 – cDNA microarray analyses ofthe expression of 40 of the 44 cDNA clonesisolated by differential display in differentFIGO stages of human cervical squamous cellcarcinoma biopsies. Data are presented in amatrix format: each row represents resultsobtained from a single cDNA clone and eachcolumn an experimental sample. Columns1–10 represent the 10 cervical squamous cellcarcinoma patients from which biopsies andmatched normal epithelium were taken. Thestages of disease are shown for each patient.For each sample, the ratio of the abundance oftranscripts of each clone in the cancer biopsyto the abundance of the same clone’s tran-script detected in adjacent histopathologicallynormal epithelium tissues is represented bythe color of the corresponding cell in thematrix. Green squares indicate transcript lev-els below the controls, red squares indicatetranscript levels greater than that of the con-trols, while black squares indicate transcriptlevels equal to the controls. The controls usedwere beta-2-microglobulin, alpha-tubulin, cy-clophilin, beta-actin, ubiquitin and G3PDH.Grey squares represent technically inadequateor missing data for that particular cell in thematrix.
422 CHENG ET AL.

responding adjacent normal tissues were employed for in situhybridization. A total of 14 patients with squamous cell carcinomaof the cervix and their corresponding adjacent normal tissues werestudied and representative results were shown in Figure 4.
The upregulated expression of the ribosomal protein S12 genewas detected within the basal immature epithelial cell layers (BL)
in 10 out of 10 adjacent histopathological normal epithelium ofearly stage cervical cancer patients (8 patients of Stage IB and 6patients of Stage IIA) (Fig. 4b). No hybridization signal could befound in the para-layers epithelial cells (PL) in all of these tissuesections. Overall, only weak hybridization signals could be de-tected within the basal immature epithelial cell layers for theNADH dehydrogenase 4 gene in adjacent histopathological normaltissues of early stage cervical cancer patients (2 out of 10 patients)(Fig. 4b). As expected, NADH dehydrogenase 4 gene and theribosomal protein S12 hybridized strongly to all stage cervicalcancer tissues (Fig. 4d). In comparison, when the adjacent his-topathologically normal tissues of late stage cervical cancer pa-tients were studied, strong hybridization signals for the NADHdehydrogenase 4 gene and the ribosomal protein S12 gene wereobtained in all the presumably histologically normal epithelial celllayers (4 out of 4 patients studied) (Fig. 4c). The hybridizationsignals were most intense in the basal immature epithelial cells(Fig. 4c). The negative control demonstrated that NADH dehydro-genase 4 gene and the ribosomal protein S12 gene did not hybrid-ize to normal cervical epithelium obtained from patients who wereadmitted for surgery for non-malignant conditions (5 out of 5patients) (Fig. 4a).
DISCUSSION
Current staging of cervical cancer (FIGO stage) is based on theextent of carcinoma spread. During carcinogenesis, the expression
TABLE I – GENES THAT ARE OVER-EXPRESSED IN SQUAMOUS CELL CARCINOMA
Differential display Reverse Northern Blot cDNA microarray Best match in GeneBank database (accession number)
G30CA G30CA G30CA Human 16S ribosomal RNA (6137796)G30CB G30CB Human EST yu04f09 (H72762)1
G30CC G30CC G30CC Human ribosomal protein S12 (RPS12) mRNA (NM_001016)G30CD G30CD G30CD Human mRNA for collagenase (X05231)G30CE G30CE G30CE No matchG30CH G30CH G30CH Human ZNF01 and HUMORFKG1B genes (AF205588)G30CI G30CI G30CI Human guanine nucleotide binding protein �5 (GNB5) (NM_006578)G30CJ G30CJ Human ribosomal protein L13a (RPL13A) mRNA (NM_012423)G30CK G30CK No matchG30CM Human chromosome X clone RP1-298J18 (AL096764)G31C1 G31C1 Human STS WI-15569 (G21217)G31C2 G31C2 No matchG31C3 G31C3 No matchG31C4A Human GAP-associated tyrosine phosphoprotein p62 mRNA (NM_006559)G31C4B G31C4B Homo sapiens chromosome 1 clone RP4-758N20 (AL03705)1
G31C4C G31C4C No matchG31C5A Human transmembrane protein BRI (BRI) mRNA (AF152462)G31C5B G31C5B No matchG31C5C G31C5C G31C5C No matchG31C5D G31C5D No matchG31C5E G31C5E G31C5E Human capping protein muscle Z-line, �1 mRNA (NM_003832)G31C5F No matchG31C5G G31C5G G31C5G No matchG31C5H G31C5H Human gene for casein kinase II subunit beta (X57152)G31C6A G31C6A G31C6A Human L-3-phosphoserine-phosphatase homo (CO9) mRNA (NM_003832)G31C6B G31C6B Human EST zh46b08.sl (W93382)1
G32C2A G32C2A No matchG32C2B G32C2B G32C2B Human general transcription factor IIB (GTF2B) mRNA (NM_001514)G32C2C G32C2C G32C2C Human clone 3A ETS-like protein mRNA (U30172)1
G32C3A G32C3A Human cDNA FLJ10841 fis, clone NT2RP4001339 (AK001703)G32C3B G32C3B Human NADH dehydrogenase subunit 6 (6137796)G32C3C G32C3C No matchG32C4A G32C4A Human mRNA for KIAA1311 protein (AB037732)G32C4B G32C4B G32C4B Human NADH dehydrogenase subunit 4 (6137796)G32C4C No matchG32C4D G32C4D G32C4D No matchG32C4E Human chromosome 17, clone HCIT48C15 (AC003104)G32C5A G32C5A G32C5A No matchG32C5B Human SIH003 mRNA (AF077040)G32C6 G32C6 G32C6 Human ubiquitin specific protease 3 (USP3) mRNA (NM_006537)G32C7 G32C7 G32C7 Human procollagen-proline, 2-oxoglutarate 4-dioxygenase (proline 4-hydroxylase),
alpha polypeptide 1 (P4HA1) mRNA (NM_000917)1The homologous sequence is in reverse orientation with respect to the cDNA fragment.
FIGURE 3 – Northern blot analyses of clones G30CC and G32C4B indifferent FIGO stages of human cervical squamous cell carcinoma.Twenty mg of total RNA obtained from cancer biopsies of differentFIGO stages and their corresponding adjacent normal epithelium wereemployed for Northern bolt hybridization. G3PDH was employed asan internal control to normalize the amount of RNA.
423HUMAN CERVICAL CARCINOMA ASSOCIATED GENES

patterns of a large number of genes are altered. Alterations in the geneexpression profiles between cervical cancer tissues of different FIGOstages have not been reported. Understanding the hierarchy of theirinteractions in cancer progression will help to elucidate the earlymolecular events and mechanisms associated with carcinogenesis andmay bring about development of better prognostic or predictive mark-ers. Our findings presented here show that differential display16–18
and cDNA microarrays can be employed as tools for rapid identifi-
cation of genes whose expression levels have altered during theprogression of human cervical neoplasia.
Of the 25 cDNA clones that are consistently upregulated inhuman cervical cancer, twelve clones are known genes with re-ported functions. The DNA sequence of clone G31C5H is homol-ogous to that of human casein kinase II subunit beta, an enzymethat is involved in transcription, signaling, proliferation and invarious stages of development.19 More significantly, Massimi et al.
FIGURE 4 – RNA-RNA in situ hybridization studies of clones G30CC and G32C4B with normal cervical squamous epithelium, cervicalsquamous cell carcinoma adjacent epithelium collected from early or late FIGO and squamous cell carcinoma biopsies. Consecutive tissuesections of 10 �m were hybridized to DIG-labeled anti-sense or sense probe derived from clone G30CC or G32C4B. Hematoxylin and Eosinstaining was employed for histology identification. Representative results obtained from clones G30CC and G32C4B to four different cervicaltissue biopsies are presented. (a) Normal cervical squamous epithelium collected from non-malignant patient. (b) Adjacent histopathologicalnormal tissues obtained from a early stage cervical squamous cell carcinoma patient. (c) Adjacent histopathological normal tissues obtained froma late stage cervical squamous cell carcinoma. (d) A squamous cell carcinoma biopsy section. BL, basal-layer epithelial cells; PL, para-layerepithelial cells; ST, stromal cells; SCC, squamous cell carcinoma. Scale bar � 80 �m.
424 CHENG ET AL.

had reported that the phosphorylation of HPV E7 by casein kinaseII enhances the disruption of the G1/S transition by HPV E7.20 Ittherefore appears that human casein kinase II subunit beta plays animportant role in human cervical neoplasia. Another 11 genes areinvolved in various molecular processes including energy con-sumption (G32C4B); signal transduction (G30CI and G31C6A);transcription regulation (G32C2B); translation (G30CA, G30CCand G30CJ); biosynthesis (G32C7 and G31C5E); and cellularmetabolism (G30CD and G32C6) (Fig. 2). Clone G30CA is ho-mologous to the human 16S ribosomal RNA that is required for thetranslation of mitochondrial subunits,21 whereas clone G32C4B ishomologous to the human NADH dehydrogenase subunit 4. Eightof the cDNA clones identified, G30CE, G31C2, G31C3, G31C5C,G31C5G, G32C3C, G32C4D and G32C5A, are novel DNA se-quences and do not match any of the reported gene sequences inthe GenBank or expressed sequence tags (EST) database (Fig. 2).In addition, the DNA sequences of clones G30CH, G31CI,G31C4B, G32C2C and G32C4A have been reported but as yet, nofunction has been assigned to the genes.
It has been suggested that gene fragments cloned from differ-ential display using arbitrary primer pairs may not represent theglobal view of the molecular events leading to tumor carcinogen-esis.16–18 Although the genes that are identified in this report mightnot represent the whole picture of cervical carcinogenesis, it wascomforting to confirm that over 70% of the genes identified byreverse Northern blot analysis to be upregulated in tissues withsquamous cell carcinoma of the cervix were similarly identifiedwhen cDNA microarrays were employed (Table I). These genesare therefore potentially most useful to be employed as tumor-specific molecular diagnostic markers.
Besides being highly expressed in tumor biopsies of early FIGOstage, our Northern blot results demonstrated that the ribosomalprotein S12 (clone G30CC) and the NADH dehydrogenase subunit4 gene (clone G32C4B) were also upregulated in adjacent his-topathologically normal tissues of stage 2B and 3B cervical cancerbiopsies (Fig. 3). These results were also confirmed using RNA-RNA in situ hybridization (Fig. 4). With Northern blot analysesand RNA-RNA in situ hybridization studies, a total of 8 cervicalcancer patients of late FIGO stages (4 patients each of Stages IIBand IIIB) have been investigated. The upregulated gene expressionof ribosomal protein S12 (clone G30CC) and NADH dehydroge-nase subunit 4 (clone G32C4B) could be consistently detected ineach of the adjacent histopathological normal epithelium (Figs. 3,4c). In comparison, upregulated gene expression of ribosomalprotein S12 (G30CC) was also detected in the adjacent histopatho-logically normal epithelium obtained from a total of 10 cervicalcancer patients of early FIGO stages (5 patients each of Stages IBand IIA) (Fig. 4b). The ribosomal protein S12 (G30CC) geneexpression was confined mainly within the basal immature epithe-lial cell layers (Fig. 4b). No upregulation in the gene expressionlevel of NADH dehydrogenase subunit 4 gene (G32C4B), how-ever, was detected in the adjacent histopathologically normal ep-ithelium tissues of early FIGO stages (Fig. 4b). Both the ribosomalprotein S12 (clone G30CC) and the NADH dehydrogenase subunit4 gene (clone G32C4B) were not detected in the squamous epi-thelial tissue sections of 5 non-cervical cancer patients (Fig. 4a).These results obtained from our in situ hybridization studiesclearly demonstrated that the gene expression profiles of the ribo-somal protein S12 (clone G30CC) and the NADH dehydrogenase4 genes (clone G32C4B) have been altered presumably in adjacenthistopathological normal epithelium.
Clone G32C4B, NADH dehydrogenase subunit 4, a mitochon-drial enzyme is involved in the metabolic pathway. It has beensuggested that metabolic transformation is likely to occur duringthe premalignant or early malignant stage of carcinogenesis.22
Yamamoto et al.23 had reported earlier that the level of NADHdehydrogenase subunits expression in pre-malignant colon adeno-mas was up regulated. Lu et al.24 and Mills et al.25 showed thatwhen tumor cells were induced to undergo terminal differentiation,the loss of tumorigenicity of the tumor cells was associated with
the downregulation of NADH dehydrogenase subunit 4. All theseevidence suggest NADH dehydrogenase subunit 4 (cloneG32C4B) play an important role in the carcinogenesis of cancer.There is also cumulative evidence to suggest that ribosomal pro-teins, in addition to participating in protein synthesis, are likely tobe involved in the malignant transformation of cells.26 For cloneG30CC, ribosomal protein S12, its overexpression has been re-ported in carcinomas of the colon and has been suggested to be anearly event in the neoplastic progression to malignancy.27
The presence of cancer cells in histopathological negative tissueassessment has previously been demonstrated within the surgicalmargin of head and neck cancer28 and in lymph nodes aroundcolorectal cancer29 by detection of K-ras and p53 mutations. Topromote invasion, tumor cells also require additional adaptationsbeyond simply an increase in cellular growth and these eventsinclude the increasing degrees of resistance to inhibitory homeo-static growth influences exerted by adjacent normal cells.30 Con-comitant changes in cells surrounding epithelial malignancy of aparacrine nature have therefore been suggested.31–36 Thereforechanges in stromal cells have been postulated to enhance thetumorigenic phenotypes of epithelial cells.37,38 Isolated carcinoma-associated fibroblast cells have been observed to exhibit an alter-ation in the expression of growth factors38–42 and the activatedstromal environment can promote tumorigenic conversion of non-tumorigenic epithelial cells.43,44 More recently, Tlsty et al.45 dem-onstrated that “pure” epithelial cell population isolated from thesetumors could produce a malignant tumor without further stimula-tion from carcinoma-associated fibroblast cells.
In cervical carcinogenesis, cancer cells often spread beyond thesurgical margin of resection.46 It is likely therefore that as thecancer cells spread, its adjacent tissues undergo molecular trans-formation before morphological and phenotypical changes. Ourpresent study utilized large biopsies from cervical cancer patientsand represented a vast mixture of cell types. Our results maytherefore likely reflect the importance of the cervical microenvi-ronment. In this report, we demonstrated that both ribosomalprotein S12 gene (clone G30CC) and the NADH dehydrogenase 4gene (clone G32C4B) peaked during early cervical carcinogenesisand maintained a high level throughout carcinogenesis. The up-regulation of gene expression of the ribosomal protein S12 geneand the NADH dehydrogenase subunit 4 gene in adjacent his-topathological normal epithelium could mark the initiation ofcellular transformation events that eventually will lead to cancerprogression. This agrees well with the clinical observation thatlocal recurrence will substantially decrease the survival rate ofcervical cancer patients47 and that most of the cervical cancerrecurs at or around the primary tumor sites. The ability to detecttranscripts of G30CC and G32C4B in otherwise phenotypicallynormal epithelium render them as useful diagnostic markers for theearly detection of human cervical cancer and as potential targetsfor early cancer therapy.
Invasive carcinoma of the cervix develops from well-definedprecursor lesions referred to as CIN.1,2 The identification of mo-lecular markers such as G32C4B and C30CC would allow one tocompare their expression profiles in progressing and non-progress-ing CIN lesions. The vast majority of low-grade CIN lesionsregress spontaneously and it has been suggested that approxi-mately 1% of CIN1 lesion and about 10% of CIN3 lesions actuallyprogress to cancer. Despite the observed low risk for cancerprogression, the current treatment of choice for all high-grade CINcervical lesions remains radical surgical resection. This is adopted,in the absence of adequate genetic markers to monitor diseaseprogression, to securely prevent the development of invasive car-cinomas. Thus, the lack of suitable cancer progression markerresults in a significant number of cervical cancer patients beingovertreated. Therefore, sensitive and objective diagnostic markers,such as those identified here, that could assess the potential of theinvasiveness of cervical neoplastic cells detected in cervical cancerpatients would be of tremendous clinical value.
425HUMAN CERVICAL CARCINOMA ASSOCIATED GENES

REFERENCES
1. Gustafsson L, Adami H-O. Natural history of cervical neoplasia;consistent results obtained by an identification technique. Br J Cancer1989;60:132–41.
2. Van Oortmarssen GJ, Habbema JD. Epidemiological evidence forage-dependent regression of pre-invasive cervical cancer. Br J Cancer1991;64:559–65.
3. Anton-Culver H, Bloss JD, Bringman D, et al. Comparison of ade-nocarcinoma and squamous cell carcinoma of the uterine cervix: apopulation-based epidemiologic study. Am J Obstet Gynecol 1992;166:1507–14.
4. Werner-Wasik M, Schmid CH, Bornstein L, et al. Prognostic factorsfor local and distant recurrence in Stage I and II cervical carcinoma.Int J Radiat Oncol Biol Phys 1995;32:1309–17.
5. Herrington CS. Human papillomaviruses (HPV) in gynecologicalcytology: from molecular biology to clinical testing. Cytopathology1995;6:176–89.
6. Lazo PA. The molecular genetics of cervical carcinoma. Br J Cancer1999;80:2008–18.
7. Brisson J, Morin C, Fortier M, et al. Risk factors for cervical intra-epithelial neoplasia: differences between low- and high-grade lesions.Am J Epidemiol 1994;140:700–10.
8. Koutsky LA, Holmes KK, Critchlow CW, et al. A cohort study of therisk of cervical intraepithelial neoplasia grade 2 or 3 in relation topapillomavirus infection. N Engl J Med 1992;27:1272–8.
9. zur Hausen H. Papillomaviruses causing cancer: evasion from host-cell control in early events in carcinogenesis. J Natl Cancer Inst2000;92:690–8.
10. Francis DA, Schmid SI, Howley PM. Repression of the integratedpapillomavirus E6/E7 promoter is required for growth suppression ofcervical cancer cells. J Virol 2000;74:2679–86.
11. De Villiers E-M. Heterogeneity of the human papillomavirus group.J Virol 1989;63:4898–903.
12. Ho GY, Bierman R, Beardsley L, et al. Natural history of cervico-vaginal papillomavirus infection in young women. N Engl J Med1998;338:423–8.
13. Kjaer SK, van den Brule AJ, Bock JE, et al. Determinants for genitalhuman papillomavirus (HPV) infection in 1000 randomly chosenyoung Danish women with normal Pap smear: are there different riskprofiles for oncogenic and nononcogenic HPV types? Cancer Epide-miol Biomark Prev 1997;6:799–805.
14. International Federation of Obstetrics and Gynecology. News. J Gy-necol Obstet 1987;25:87.
15. Soong TW, Hui KM. Locus-specific transcriptional control of HLAgenes. J Immunol 1992;149:2008–20.
16. Liang P, Pardee AB. Differential display of eukaryotic RNA by meansof the polymerase chain reaction. Science 1992;257:967–70.
17. Liang P, Averboukh L, Pardee AB. Distribution and cloning ofeukaryotic mRNAs by means of differential display: refinements andoptimization. Nucleic Acids Res 1993;21:3269–75.
18. Liang P, Zhu W, Zhang X, et al. Differential display using one-baseanchored oligo-dT primers. Nucleic Acids Res 1994;22:5763–4.
19. Guerra B, Issinger OG. Protein kinase CK2 and its role in cellularproliferation, development and pathology. Electrophoresis 1999;20:391–408.
20. Massimi P, Pim D, Storey A, et al. HPV-16 E7 and adenovirus E1acomplex formation with TATA box binding protein is enhanced bycasein kinase II phosphorylation. Oncogene 1996;12:2325–30.
21. Powers T, Noller HF. A functional pseudoknot in 16S ribosomalRNA. EMBO J 1991;10:2203–14.
22. Costello LC, Franklin RB. Bioenergetic theory of prostate malig-nancy. Prostate 1994;25:162–6.
23. Yamamoto A, Horai, S, Yuasa Y. Increased level of mitochondrialgene expression in polyps of famililial polyposis coli patients. Bio-chem Biophys Res Commun 1989;159:1100–6.
24. Lu X, Walker T, MacManus JP, et al. Differentiation of HT-29 humancolonic adenocarcinoma cells correlates with increased expression ofmitochondrial RNA: effects of trehalose on cell growth and matura-tion. Cancer Res 1992;52:3718–25.
25. Mills KI, Woodgate LJ, Gilkes AF, et al. Inhibition of mitochondrial
function in HL60 cells is associated with an increased apoptosis andexpression of CD14. Biochem Biophys Res Commun 1999;263:294–300.
26. Wool IG. Extraribosomal functions of ribosomal proteins. TrendsBiochem Sci 1996;21:164–5.
27. Pogue-Geile K, Geiser JR, Shu M, et al. Ribosomal protein genes areover expressed in colorectal cancer: isolation of a cDNA clone en-coding the human S3 ribosomal protein. Mol Cell Biol 1991;11:3842–9.
28. Brennan JA, Mao L, Hruban RH, et al. Molecular assessment ofhistopathological staging in squamous cell carcinoma of the head andneck. N Engl J Med 1995;332:429–35.
29. Hayashi N, Arakawa H, Nagase H, et al. Genetic diagnosis identifiesoccult lymph node metastases undetectable by the histopathologicalmethod. Cancer Res 1994;54:3853–6.
30. Kinzler KW, Vogelstein B. Landscaping the cancer terrain. Science1998;280:1036–7.
31. Chiquet-Ehrismann R, Mackie EJ, Pearson CA, et al. Tenascin: anextracellular matrix protein involved in tissue interactions during fetaldevelopment and oncogenesis. Cell 1986;47:131–9.
32. Basset P, Bellocq JP, Wolf C, et al. A novel metalloproteinase genespecifically expressed in stromal cells of breast carcinomas. Nature1990;348:699–704.
33. Wright JH, McDonnell S, Portella G, et al. A switch from stromal totumor cell expression of stromelysin-1 mRNA associated with theconversion of squamous to spindle carcinomas during mouse skintumor progression. Mol Carcinog 1994;10:207–15.
34. Singer C, Rasmussen A, Smith HS, et al. Malignant breast epitheliumselects for insulin-like growth factor II expression in breast stroma:evidence for paracrine function. Cancer Res 1995;55:2448–54.
35. Ronnov-Jessen L, Petersen OW, Bissell, MJ. Cellular changes in-volved in conversion of normal to malignant breast: importance of thestromal reaction. Physiol Rev 1996;76:69–125.
36. Hong WK, Sporn MB. Recent advances in chemoprevention of can-cer. Science 1997;278:1073–7.
37. Rinehar CA, Torti VR. Aging and cancer: the role of stromal inter-actions with epithelial cells. Mol Carcinog 1997;18:187–92.
38. Yee D, Rosen N, Favoni RE, et al. The insulin-like growth factors,their receptors and their binding proteins in human breast cancer.Cancer Treat Res 1991;53:93–106.
39. Yan G, Fukabori Y, McBride G, et al. Exon switching and activationof stromal and embryonic fibroblast growth factor (FGF)-FGF recep-tor genes in prostate epithelial cells accompany stromal independenceand malignancy. Mol Cell Biol 1993;3:4513–22.
40. Ellis MJ, Singer C, Hornby A, et al. Insulin-like growth factormediated stromal-epithelial interactions in human breast cancer.Breast Cancer Res Treat 1994;31:249–61.
41. Nakamura T, Matsumoto K, Kiritoshi A, et al. Induction of hepatocytegrowth factor in fibroblasts by tumor-derived factors affects invasivegrowth of tumor cells: in vitro analysis of tumor-stromal interactions.Cancer Res 1997;57:3305–13.
42. Frazier KS, Grotendorst GR. Expression of connective tissue growthfactor mRNA in the fibrous stroma of mammary tumors. Int. J Bio-chem Cell Biol 1997;29:153–61.
43. Olumi AF, Grossfeld GD, Hayward SW, et al. Carcinoma-associatedfibroblasts direct tumor progression of initiated human prostatic epi-thelium. Cancer Res 1999;59:5002–11.
44. Skobe M, Fusenig NE. Tumorigenic conversion of immortal humankeratinocytes through stromal cell activation. Proc Natl Acad Sci USA1998;95:1050–5.
45. Tlsty TD, Hein PW. Know thy neighbor: stromal cells can contributeoncogenic signals. Curr Opin Genet Dev 2001;11:54–9.
46. Denehy TR, Gregori CA, Breen JL. Endocervical curettage, conemargins and residual adenocarcinoma in situ of the cervix. ObstetGynecol 1997;90:1–6.
47. Burke TW, Hoskins WJ, Heller PB, et al. Clinical patterns of tumorrecurrence after radical hysterectomy in Stage IB cervical carcinoma.Obstet Gynecol 1987;69:382–5.
426 CHENG ET AL.
Copyright © 2022 FDOKUMEN




















